
Submitted:
27 December 2024
Posted:
30 December 2024
You are already at the latest version
Abstract
This descriptive review summarizes the most recent findings on the induction and distribution of viable non-culturable (VBNC) Listeria monocytogenes in food production conditions and food. The aim was to obtain information on the factors that favor the transition to the VBNC state in L. monocytogenes, its resuscitation capacity, and, according to scientific articles published since 2020, how food contamination by the bacterium in VBNC state can be prevented. The methods used for VBNC L. monocytogenes detection were also reviewed. A few studies reported the presence of VBNC L. monocytogenes in food, in which fresh produce and chicken meat were considered. Different physicochemical stresses such as exposure to disinfectants with diverse actions and essential oils, desiccation, low temperatures, absence of nutrients, high NaCl and iron concentrations, and low pH adjusted with acetic acid were reported to induce the VBNC state in L. monocytogenes. The VBNC forms of L. monocytogenes were able to regain growth and virulence. This could pose a safety risk that cannot be revealed by the standard culture-dependent methods recommended for L. mono-cytogenes detection. Therefore, the presence in food and food production plants of VBNC L. mon-ocytogenes should be prevented by appropriate use of hurdles and cleaning/disinfection proce-dures. The opportunity to harmonize VBNC cell detection methods for regular use in food safety evaluation also emerged.
Keywords:
Listeria monocytogenes
; VBNC
; detection methods
; inducing stress factors
; occurrence in food
; resuscitation
1. Introduction
The genus Listeria comprises Gram-positive facultative anaerobic rod-shaped bacteria with a marked capacity to inhabit a variety of environmental niches and animal hosts, determined by their ability to withstand physicochemical stressors and activate persistence mechanisms. These characteristics make Listeria spp. prone to colonizing food production plants and contaminating foodstuff. The species Listeria monocytogenes in an intracellular pathogen with potential to cause invasive infections, such as septicemia, meningitis, and endocarditis, that are 99% foodborne in immunocompromised persons, the elderly, pregnant women, and newborns. This pathogen can also cause abortion in humans and animals [1,2,3]. Upon L. monocytogenes infection, healthy adults do not develop symptoms or manifest a gastrointestinal illness that is usually not diagnosed [4]. However, in pregnancy listeriosis, after a two-three weeks incubation period, can manifest as a self-limiting illness with flu-like symptoms but the bacterium can cross the placenta and infect the fetus with consequences such as spontaneous abortion, preterm birth, still birth and newborn infections. These complications were reported in 80% cases of listeriosis in pregnant women [5].
L. monocytogenes is classified in the four phylogenetic lineages I, II, III and IV in which 13 serotypes are distributed. Serotypes are further divided into sequence types (STs) and clonal complexes (CCs). About 95% isolates from food and infection cases belong to serotypes 1/2a, 1/2b, 1/2c, and 4b, among which 1/2b and 4b serotypes, belonging to lineage I, are more frequently associated with human clinical cases. Strains of Lineage II are more common in food and food processing plants while those belonging to Lineages III and IV are isolated mainly from animals. The internalin A, a cell well-anchored protein encoded by the inlA gene, is considered the main virulence factor in the hypervirulent strains of L. monocytogenes [6]. Most cases of listeriosis were caused by serotype 1/2a, while serotype 4b was associated with higher hospitalization and mortality rates [7].
In the European Union (EU), 2738 cases of listeriosis were confirmed in 2022 with a fatality rate of 18.1%, the highest compared to previously reported data, that increased to 2,952 confirmed cases, 96.5% hospitalizations and 19.7% fatality rate in the year 2023 according to the most recent reports on zoonotic diseases in the EU [8,9]. In 2022, 2770 confirmed listeriosis, including 17 strong-evidence and 18 weak-evidence foodborne outbreaks that affected 296 persons, with 242 hospitalizations and 28 deaths, were reported by 30 European countries through The European Surveillance System (TESSy). Whole genome sequence (WGS) reported to TESSy showed small but long-lasting multi-country epidemiological clusters that persisted for several years. The ECDC-EFSA One Health WGS System, already at the end of 2022, defined that more than one-third of the multi-country outbreak isolates matched non-human isolates, showing the potential of WGS application in identifying the origin of the outbreaks [4].
In the meat, fish/seafood, dairy, and fruit/vegetable sectors L. monocytogenes represents one of the main microbiological food contamination threats due to its capacity to persist in the food and feed processing environment (FFPE). Obstacles to the elimination of persistent L. monocytogenes in the FFPE are insufficient hygiene barriers and inadequate design of equipments, cleaning and disinfection procedures. To identify a possible contamination by L. monocytogenes a well-designed environmental sampling program (EMP) must be implemented, also involving the facility personnel, and, in case of persistent L. monocytogenes detection, a 'seek-and-destroy' approach must be applied, namely the application of cause analysis and corrective actions to eliminate the contamination at spefic sites. Persistence can be assessed by genetically characterizing the food plant isolates e.g. by sigB allelic typing (sigB AT) and single nucleotide polymorphism (SNP) analysis of whole genomes [10,11,12].
Beyond implementing the correct use of sanitizers, drainage systems, raw ingredient contamination control, and improvement of compartmentalization in the manufacturing plant, it should be elucidated to what extent the genotypes associated with persistence in the production environment are linked to listeriosis outbreaks and cases by making available genetic data of outbreak strains along with the epidemiological investigation results [10,11].
Stresses to which L. monocytogenes is exposed in food, such as suboptimal pH, temperatures and osmotic pressure, heating, and smoking, induce variations in the expression of genes related to pathogenesis and virulence, such as chemotaxis, flagella motility, transmembrane receptors, signal transduction and can also lead to the formation of sub-lethally injured cells, able to recover by cell repair mechanisms, and/or VBNC cells that do not grow in commonly used culture media in the recommended incubation conditions [9,13,14]. L. monocytogenes adopts different persistence strategies that determine its tolerance to food processing stressors. These are biofilm formation, development of “persisters”, and the transition to the viable but not culturable (VBNC) state [10,15,16,17,18,19,20,21,22,23].
Indeed, L. monocytogenes can form biofilms on the surfaces of food processing environments that withstand stressful conditions such as low temperature and desiccation [18]. Moreover, L. monocytogenes may give origin to “persisters”, i.e. cells that survive cleaning and disinfection thus reoccurring in the production plant and final products. This subpopulation of cells is slow replicating or non-dividing and transiently multidrug tolerant [15].
Differently, VBNC cells are in a deeper dormancy status with a minimal metabolism, active transcription and translation, intact DNA and membrane and capacity to resuscitate in particular conditions [16]. Sub-lethally injured cells that remain metabolically active but cannot be resuscitated in the standard culturing conditions may enter the VBNC state [22]. VBNC cells formation was observed in over one hundred microbial species and may occur following starvation, exposure to extreme temperatures, oxygen limitation, high salinity, pH changes, UV light, ultrasound, suboptimal osmotic pressure, and exposure to toxic agents, varying on microbial species and strains. In addition, VBNC formation may be induced even in nutrient-rich environments such as food. The mechanisms by which bacterial cells enter the VBNC state are still largely unknown, though some studies provided evidences on the involvement of stress response proteins, transcriptional regulators, membrane and cell division related proteins, and toxin/antitoxin systems [16].
Considering that the detection of culturable cells according to standard methods is carried out in specific culture conditions and defined time frames, nonculturability is defined as the lack of colony formation after the prescribed incubation time. This criterion can be used to distinguish VBNC cells from persister cells [16]. The detection of L. monocytogenes in foods is carried out by the culture-dependent standard methods EN ISO 11290–1 and EN ISO 11290–2 to accomplish detection and enumeration, respectively, as specified by Regulation 2073/2005 amended by Regulation 2024/2895 for the European Union [24,25]. These methods require some days for time to results and are unsuitable to detect VBNC cells [26].
Since VBNC L. monocytogenes may represent a threat to consumer’s health, its reliable and rapid detection would considerably improve food safety assessment. Therefore, this review summarizes the latest findings on the VBNC state of L. monocytogenes with special attention to the evidence of VBNC state induction in food and food production conditions and possible transmission of infectious VBNC through contaminated food. Scientific publications were retrieved from Google Scholar (https://scholar.google.com/schhp?hl=it, accessed on 29 November 2024) and Scopus (https://www-scopus-com.bibliosan.idm.oclc.org/search/form.uri?display=basic#basic, accessed on 29 November 2024) with the search string “viable AND Listeria monocytogenes AND food”. The aim was to obtain a complete overview of what currently is known regarding the conditions that lead to VBNC formation in L. monocytogenes, what is still to be known, and treatments that are effective in preventing the occurrence of this form of the pathogen. Results in the 2020-2024 years window were whole text screened for the occurrence of the words “viable non-culturable (or non-culturable)” or “VBNC” or “VNC”. Finally, the scientific articles pertinent to the scope of this review were synthesized.
2. Detection Methods Applied to Detect VBNC L. monocytogenes
An increase in number of publications regarding viable pathogen detection in food was reported from the year 2010 to 2022 and the methods used were based on fluorescence microscopy, fluorescence labeling coupled with flow cytometry (FC), viability PCR (vPCR), impedance-based methods, and surface Raman spectroscopy. Articles focused on molecular detection of VBNC L. monocytogenes used as targets the genes hly, iap, 16S rRNA gene, and prfA, and were aimed at method optimization and detection from various food matrices such as powdered infant formula, milk, and shrimp [26].
2.1. Optical Detection Methods
Early studies on VBNC cells compared bacterial culture with microscopic counting of cells stained with fluorescent dyes able to differentiate viable and dead cells [27]. Immunolabeling with antibodies bound to fluorophores such as fluorescein isothiocyanate (FITC) also allowed specific detection of bacteria in food. In this case, the antibody and the dye are first incubated in the dark with the target bacteria and then cells are detected by FC [28].
Among viability staining dyes, fluorescein diacetate (FDA) and the similar dye carboxyfluorescein diacetate (CFDA) are able to permeate the membrane of metabolically active cells and inside the cells are converted by unspecific esterases into fluorescent compounds that are retained by cells with intact cytoplasmic membranes [16,28,29]. Propidium iodide (PI) is an indicator of membrane integrity and cell death that enters cells with damaged membranes and binds to DNA at either major or minor grooves with red fluorescence emission. Differential staining with FDA or CFDA and PI allows distinguishing viable (green fluorescence), sub-injured (brown fluorescence), and dead (red fluorescence) cells [14,29]. A similar principle is exploited by the commercial staining system LIVE/DEADTM BacLightTM kit composed of two fluorescent dyes SYTO9 and PI. Other dyes used for viable cell differential staining are 5-cyano-2,3-ditolyl tetrazolium chloride (CTC), 4′,6-diamidino-2-phenylindole (DAPI), ethidium bromide, Bis-(1,3-Dibutylbarbituric Acid)Trimethine Oxonol (DiBAC4 (3)), calcein and acridine orange [28,30,31].
Direct epifluorescent filter technique (DEFT) enumerates microbes by filtering samples pre-treated with chemical reagents or enzymes on polycarbonate filters and staining with fluorescent viability staining such as CFDA or acridine orange dyes followed by microscopic analysis. When applied to determine VBNC L. monocytogenes cells in hard cheese during cold storage it showed limitations such as cell injury caused by the chemical reagents or missed detection of weak signals [28].
FC allows the detection of viable cells based on cell wall/membrane integrity, cell energy status, and enzyme activities such as substrate uptake. In FC, cells suspended in a liquid pass at high speed through a laser beam. Cell detection is carried out by exploiting light scattering or fluorescence emission. The signals are sent to detectors and converted into digital signals read by a computer. Before FC, food samples must be pre-treated to decrease background noise. As an example, dairy products need pre-treatments such as the dispersion of milk powder in a buffer and the extraction of cheese juice with a hydraulic press. Moreover, proteases and detergents such as Triton X-100 can be applied to degrade proteins and break somatic cells. Filtration and centrifugation are also carried out [28]. FC is not applicable to all food matrices and optical assay present common disadvantages such as susceptibility to cell membrane damage, background fluorescence and misidentification of dead cells with an intact membrane as viable [27,31].
2.2. Molecular Detection Methods for Viable Cell Detection
Molecular methods have been widely used in detecting bacteria in VBNC state in food matrixes and water for their high sensitivity, specificity, and detection speed. The most commonly adopted is quantitative polymerase chain reaction (qPCR) applied to cells treated with DNA intercalating dyes before DNA extraction [27,28]. This method is designated as “viability PCR” (vPCR) and allows an accurate quantification of only viable bacteria. It consists in applying a specific qPCR test to DNA extracted from cells treated with dyes which enter into the cells with a compromised membrane integrity and bind to their DNA blocking amplification after exposure to light. In theory, DNA intercalating dyes cannot penetrate intact cell membranes [18,28,31,32,33].
Different from standard culture detection methods applied to bacterial pathogen detection in food, vPCR provides results before products with a short shelf life are sold out [28]. If qPCR is performed with and without intercalating dye pre-treatment, viable cells, and total cells can be detected and quantified separately [34]. These methodologies must be validated for each type of matrix to avoid overestimation of the VBNC cells for the presence of dead cells with intact membranes [27].
Ethidium monoazide (EMA) was the first dye used among DNA intercalating dyes able to enter the membrane of dead cells [28,32] but presented limitations such as the extent of penetration influenced by the species of microorganisms and cytotoxic effects on viable cells [27,28]. Moreover, the usefulness of EMA is also limited by the ability of the dye to penetrate viable cells with consequent underestimation of their number [18].
Propidium monoazide (PMA) has a lower toxicity than EMA and a higher selectivity for cells with damaged membranes for its double positive charge, differently from EMA which has only one positive charge [28]. The azide group of PMA allows its crosslinking with the DNA of dead cells which is permanently modified while the DNA of viable cells remains intact and can be detected in the subsequent PCR reaction. The PMAxx derivative of PMA exerts better discrimination between viable and dead cells in complex matrices [35] and a combination of EMA and PMAxx improves selectivity for viable cells. Indeed, EMA is effective in distinguishing dormant cells that lack active efflux pumps to expel the dye. The mixture of these two dyes is commercially available under the name PEMAX [27].
In the vPCR, dye-treated samples are exposed to light, for example, from the blue light PMA-Lite LED or a 500-650 W halogen lamp, and dye crosslinking to DNA is usually allowed to occur for 5-15 min. Bacterial cells are concentrated by centrifugation and the pellet is used for DNA extraction. The pellet can also be stored at −20 °C until DNA extraction. The theoretical limits for L. monocytogenes quantification in water and fresh produce by vPCR with PMAxx were 2.4 and 1.0 Log cells/100 ml or gram, respectively, with primers targeting the Listeriolysin O hly gene. The levels of VBNC were determined as the difference between viable bacteria quantified by vPCR and the culturable bacteria counted on plates [20,27,28].
Before the vPCR tests, several sample pretreatments must be carried out. As an example, for solid dairy products, these include homogenization in phosphate buffer saline (PBS) or trisodium citrate solution, removal of fat and proteins by centrifugation, and pellet collection. For liquid samples such as milk and fermented milk, no homogenization is needed. The pellets are then re-suspended in a solution transparent to light mixed with the dye solution, incubated in the dark for a minimum of 5 min at room temperature, and exposed to light while held on ice for 2–5 min to induce dye crosslinking. Finally, cells are centrifuged and used for DNA extraction and vPCR [28].
It was reported that PMA treatment does not completely abolish amplification from dead cells possibly because cell debris hinders its penetration through the membrane. Therefore, sodium deoxycholate (SD), that is an anionic surfactant that disrupts the cell membranes of damaged or dead cells, was used to enhance L. monocytogenes dead cell permeability. A PMA concentration of 20 μM and 0.01% SD with an exposure time of 5 min were found to optimally protect the viable cells at the same time inhibiting amplification from non-viable cells. However, an SD concentration higher than 0.02% caused the inhibition of amplification also from viable cells [36]. In some studies, sodium dodecyl sulfate (SDS) was used to decrease amplification from dead cells before treatment with PMA by enhancing the permeability of the membranes but this practice is not easily applicable to bacteria at low concentrations and complex food matrices [33].
Immunomagnetic separation (IMS) allows to selectively bind and separate foodborne pathogens from a food suspension, thus eliminating the matrix interference effect. However, its efficiency can be reduced by minor changes in pH, monodispersity, and cross-linking time that influence the activity of the antibody adsorbed on the immunomagnetic beads. Therefore, an IMS method based on the streptavidin-biotin system was developed to enhance capture capability. However, antibodies were required in high amounts to enhance the capture efficiency. A novel biotin exposure–based IMS assay coupled with SDS and multiplex vPCR with PMA was developed for the rapid detection of viable bacterial pathogens including L. monocytogenes in milk. Biotinylated antibodies were used to bind to the cells, and then streptavidin-coated magnetic beads (SA-MB) were used to concentrate the pathogens that were treated with SDS and PMA. Finally, the genomic DNA was extracted and used in multiplex vPCR. The sensitivity of detection ranged between 10 to 103 CFU/ml for the target bacteria and the limit of detection (LOD) in artificially contaminated milk was 10 CFU/ml for all pathogens, showing an increased sensitivity compared to the test without IMS [33].
Nanomaterials AuNPs, AgNPs, silver nanowires, and PEI@Fe3O4, were tested as qPCR signal enhancers able to increase the normalized fluorescence (ΔRn) value. AuNPs with 20 nm particle size and 100-fold diluted displayed the highest signal enhancement with an increase of 20%. The sensitivity of the AuNPs-SD-PMA-qPCR assay was 5 × 102 CFU/ml for L. monocytogenes detected simultaneously with Salmonella enterica in enrichment cultures from dairy products. Thermal lysis for 15 min for DNA extraction reduced the detection time and the number of needed reagents by improving portability for on-site detection. The use of AuNPs can be easily implementable because it is non-toxic, highly biocompatible, not expensive, and easily produced and modified [36].
Isothermal amplification techniques coupled with intercalating dyes were introduced in recent years as an alternative to vPCR. One of these is loop-mediated isothermal amplification (LAMP), a cyclic strand displacement DNA synthesis process that allows the synthesis of 106-109 copies of target DNA in less than 1 h at a constant temperature of 60 °C - 65 °C. Amplification is visualized in real-time for the formation of a white magnesium pyro-phosphate precipitate that can be measured with a turbidimeter or by adding a fluorescent dye, such as Sybr Green I, to visualize amplification under UV light. LAMP test is less sensitive to inhibition by the food matrix compared to PCR and can be carried out in water baths or heaters, which makes it a portable method [26,28,31,37]. The specificity is high since up to 6 specific primers matching the target regions are used. These include forward inner primer (FIP) and backward inner primer (BIP), forward outer primer (F3) and backward outer primer (B3) and forward loop primer (FLP) and backward loop primer (BLP). The sensitivity of LAMP was found to be higher than that of qPCR with a LOD of 6 CFU/reaction for L. monocytogenes. LAMP coupled with PMA or EMA proved faster and 100-fold more sensitive than vPCR for food samples [28].
Most LAMP methods do not include an internal amplification control (IAC) due to the difficulty of multiplexing the reaction, and LAMP amplification products are differentiated based on their melting temperatures. However, an L. monocytogenes-specific test was developed to comprise an IAC constituted by a plasmid containing the original target fragment with inserted the sequence CCATCCGACGGCGGGCGGCC that increased the Tm of the IAC fragment allowing its distinction by melting curve analysis. This LAMP method effectively blocked amplification from up to 106 CFU/ml dead cells, and its LOD95 defined by plate counts was 2.7 CFU/25 ml of milk. The performance of the method was tested on a total of 59 spiked milk samples but not on VBNC cells [37].
Competitive annealing mediated isothermal amplification (CAMP) is another isothermal amplification reaction with incubation at 60-65 °C for 60 min with a simpler primer design compared to LAMP. It was tested for the detection of L. monocytogenes in milk and milk powder and was coupled with PMA cross-linking or to PMA and hydroxynaphthol (HNB) to visualize amplification by color change (violet to blue). Visual detection in this method was also achieved by using gold nanoparticle probes (AuNP) whose dispersion and aggregation can be visually detected based on color change. In negative samples, nanoparticles form aggregates conferring a red to blue-grey color to the solution, while in positive samples the AuNP probe hybridizes with the target and remains dispersed with no color change [28].
A novel technology was developed to detect viable L. monocytogenes based on PMA-LAMP with four primers combined with nanozyme strips, that are cost effective and equipment-free with the possibility of visual detection. This technology included the use of BIO- and FITC-modified primers targeting the hlyA gene and a magnetic nanozyme modified with a biotin-conjugated antibody specific for L. monocytogenes. The peroxidase-like activity of the magnetic nanozyme generated a visually readable color and quantitative detection could be achieved by measuring the optical intensity of the test line (T line) by ImageJ with a detection limit of 5.4 fg/μl [38].
Fe3O4 NPs S15 nanozyme was tested for viable L. monocytogenes detection after mixing with the amplification products of PMA-LAMP and allowed to react with nitrocellulose (NC) membranes on which anti-mouse IgG and anti-FITC antibodies were immobilized to form C and T lines, respectively. If L. monocytogenes was present in the sample, the PMA-LAMP products conjugated with the nanozyme probe by the combination of BIO and anti-biotin antibody. One side of positive products was labeled with Fe3O4 NPs while the other side was labeled with FITC so the amplification products could be captured by anti-FITC antibody on the T line while the free nanozyme probes stopped in the C line. The color reaction was enhanced by the enzymatic reaction between the nanozyme and 3,3-diaminobenzidine (DAB)/H2O2 enzymatic substrate. If viable L. monocytogenes was absent, only the C line appeared, while if no color appeared, the result was considered not valid. The method allowed to detect 1.164 × 10 CFU/ml and 1.021 × 10 CFU/ml L. monocytogenes, in biofilms developed on stainless steel and lettuce, respectively [38].
Other isothermal amplification techniques include polymerase spiral reaction (PSR) and crossing priming amplification (CPA). The CPA assay was applied to L. monocytogenes by using hylA gene as a target and calcein for product visualization. It involves use of several primers and probes, with one or more cross primers, and exploits the Bst DNA polymerase for strand displacement of the target DNA. The amplification products are detected by turbidimetry and electrophoresis or addition of fluorescent dyes such as calcein and SYBR Green [31].
A PMA-CPA assay with a sensitivity similar to a PCR-based assay was applied for the direct detection of VBNC L. monocytogenes cells in artificially contaminated rice flour. Dead bacterial cells killed by high temperature (121°C) and high pressure (100 kpa) were included as a negative control and genomic DNA was included to verify no amplification from extracellular DNA. PMA was used in a final concentration of 10 μg/ml, with incubation at room temperature in the dark for 5 min, and exposure to light from a 500 W halogen lamp for 15 min. After DNA extraction, the CPA assay could quantify bacterial cells in rice flour suspension in a concentration range of 104–108 CFU/ml [31].
A disadvantage of vPCR and other molecular viability tests based on DNA amplification is that cells in sublethal states with a damaged membrane cannot be detected. Colorimetric sensors can aid in vPCR to detect only viable pathogens by using streptavidin magnetic beads-dual aptamers (SMB-Apts) sandwich coupled with an AuNPs colorimetric sensor [28].
One limitation of PMA-based vPCR was the poor discrimination between viable and dead cells when applied to high levels of L. monocytogenes heat treated at 60 °C for 6 min. False positive results were attributed to the shadowing of non-viable cells in the photo-activation step. In addition, this method might overestimate the number of viable cells since dead cells with intact membranes could be counted as viable [39]. To increase vPCR accuracy, the use of a stable internal process control (ISPC), similar to the one developed for Campylobacter jejuni, was recommended to normalize and subtract the signal originating from dead cells [40].
An alternative approach to vPCR is the specific amplification/detection of mRNA by reverse transcription followed by qPCR (RT-qPCR) [41]. Since mRNAs have only a few minutes of half-life, they are only detectable when cells are viable through reverse transcription to cDNA and PCR amplification [28]. A comparison among commercial RNA extraction kits based on 260/280 and 260/230 optical density (OD) ratios, RNA integrity (RIN) values, and absence of residual gDNA after DNAse treatment was performed to select the most suitable for quantitative detection of L. monocytogenes. Subsequent multiplex RT-qPCR targeted on hly, actA, and a non-competitive internal amplification control (NC-IAC) allowed to obtaining LOD50 and LOD95 values of 1.2 CFU/25 g and 5.1 CFU/25 g, respectively, from enrichment cultures of artificially spiked smoked salmon [41]. Other studies also used hly and inlA-targeted RT-qPCR to analyze the viability of L. monocytogenes in food production conditions [39,42]. Limitations of mRNA usage for viable bacteria detection are its instability and the risk of contamination with DNA. Moreover, RNA extraction from complex food matrices still requires optimization [28].
Figure 1 reports a simplified scheme highlighting the experimental workflows that were applied for the detection of viable L. monocytogenes cells.
3. Distribution of VBNC L. monocytogenes in Food and Food Production Plants
A few recent studies investigating the presence of VBNC L. monocytogenes in food considered ready-to-eat (RTE) salads and chicken meat [17,34,43]. RTE salads require extensive handling and are exposed to recontamination from processing surfaces and equipment after listericidal treatments. The storage of these products in refrigerated conditions could allow the development of L. monocytogenes [17].
Bernardo et al. [17] reported that, based on hygiene indicators and L. monocytogenes determination by culture-based methods, an RTE salad considered in the study could have a shelf-life of eight days when stored at 4°C, according to Regulation (EC) No. 2073/2005 food safety criteria [25]. However, on day 8, the concentration of L. monocytogenes obtained by vPCR with PMAxx was at least 3 Log CFU/g higher than that obtained by colony counts. No amplification was detected in salad extracts submitted to boiling before vPCR. Therefore, the vPCR results exceeded the food safety criteria of 2 Log CFU/g and showed that the standard culture-based methods might underestimate the number of viable L. monocytogenes cells affected by processing treatments that hamper cultivability. However, the potential interference of dead cells was not evaluated [17].
A study on poultry meat investigated the presence of VBNC cells of L. monocytogenes in two slaughterhouses in 11 wings, 3 thighs, 12 drumsticks, and 4 breast samples. After homogenization of 25 g of sample in 225 ml of 0.1% peptone water, 1 ml was used to determine the total cell concentration, and 1 ml was used for vPCR after treatment of cells with PMAxx 25 µM. DNA extraction was performed on 750 µL of the two aliquots, treated and not treated with PMAxx, using a commercial Kit with proteinase K. The vPCR technique showed a sensitivity of 100%, a specificity of 66.7%, and an efficiency of 83.3%, with 0.67 kappa coefficient agreement when compared with culturing by enrichment, colony isolation on Oxoid Chromogenic Listeria agar (OCLA) followed by colony identification by species-specific PCR (OCLA-PCR). The rather low specificity and agreement were because 5 samples were positive on vPCR but negative on OCLA-PCR. These five samples most probably contained VBNC L. monocytogenes cells indicating the occurrence of these forms in poultry meat. Previous studies based on plate counts reported lower contamination levels for poultry, and this can be explained by the ability of vPCR to detect also VBNC cells. The high sensitivity score of vPCR (100%) against OCLA-PCR indicated the absence of false negative results for the method [34].
A second investigation found L. monocytogenes in 75.0% of 65 samples from two slaughterhouses by culturing methods and qPCR. The results were not fully concordant since some samples were positive only for one of the techniques. Viability PCR allowed to detect L. monocytogenes in half of the carcasses from one slaughterhouse but the microorganism was not detected by culture-dependent methods. In another slaughterhouse, L. monocytogenes was detected in 100% of samples by culture-dependent methods only (38.5%) or both by culturing and qPCR (61.5%). Comparison among culture-dependent detection, qPCR, and vPCR allowed to determine that the contamination levels in the positive samples were 4.01 Log CFU/g for total cells, 3.21 Log CFU/g for viable cells, and 1.00 Log CFU/g for culturable cells, 3.20 Log CFU/g for VBNC cells and 3.93 Log CFU/g for dead cells [43].
4. Factors Inducing the VBNC State in L. monocytogenes
According to a review published in 2021, studies carried out before 2020 reported VBNC induction in L. monocytogenes as determined by the detection of respiratory activity by CTC-DAPI staining in seawater and in “microcosm water” (i.e., sterile filtered water with pH 6.0), thus in the absence of nutrients. Those VBNC cells retained metabolic activity for up to ten weeks, and a high inoculum concentration decreased the induction of the VBNC state, probably due to higher nutrient availability within the population. The VBNC state was induced also in biofilms exposed to tap water. Viability PCR highlighted the presence of VBNC L. monocytogenes in pig manure, food environments, and on the surface of vegetables such as parsley and spinach [32].
Staining with CFDA showed that L. monocytogenes entered the VBNC state in long-ripened hard cheese left for up to 90 days at 6°C in meat exudate and less in smoked salmon juice. Direct viability count coupled with fluorescence in situ hybridization (FISH) with a 16S rRNA probe and vPCR showed the existence of VBNC L. monocytogenes in RTE vegetables [32].
4.4. Role of Disinfecting Agents in VBNC L. monocytogenes Induction
One of the concerns dealt with in studies regarding VBNC L. monocytogenes was the effect of RTE fresh produce process wash water (PWW) in the induction of VBNC cells. PWW is indeed a relevant vector of microbial cross-contamination of clean products from contaminated products that is prevented by adding sanitizers among which chlorine is the most commonly used. However, it was demonstrated that chlorine induces the VBNC state of the foodborne pathogens L. monocytogenes and S. enterica so suitable quantification methods are required to detect and control VBNC cells in PWW [27].
An optimized detection and quantification method of VBNC cells in PWW was developed after comparison of different combinations of dyes in FC, and of PMAxx, or EMA and PMAxx combination in vPCR. The method was validated in industrial settings by using six L. monocytogenes isolates from leafy vegetables in a mixture. A PMAxx concentration of 75 μM was tested since a 50 μM concentration added to PWW did not completely inhibit amplification from dead cells, possibly due to the presence of interfering organic matter, while 100 μM appeared toxic for culturable cells. Amplification from heat-treated cells was abolished after 30 min of incubation with the dye at 40°C but the PCR signal from chlorine-treated cells was not completely inactivated. PMAxx combined with EMA at concentrations of 75 μM and 10 μM, respectively, and incubation at 40°C for 40 min followed by 15 min exposure to light reduced the amplification from dead cells after chlorine treatment [27].
In the shredded lettuce PWW no significant changes in the concentration of free chlorine (10 mg/l) and pH were observed, and the culturable bacteria maintained a total count below 3 Log CFU/ml. However, a higher number of viable cells was detected by using EMA-PMAxx-vPCR, thus showing the induction of VBNC L. monocytogenes. FC with double-staining indicated a higher number of viable cells compared to vPCR, possibly for an overestimation caused by the presence of cells with intact membranes but damaged DNA, proteins, and lipids. Therefore, fluorescent markers able to highlight damages to these molecules should be used in FC for improved viable cell discrimination. The VBNC L. monocytogenes cells induced by 10 mg/l chlorine could be detected after 1 h and remained stable after 3 h [27].
Residual concentrations in PWW of 20 mg/l for chlorine, 80 mg/l for peroxyacetic acid (PAA), and 3 mg/l for chlorine dioxide (ClO2), the most commonly used sanitizers in vegetable washing, are recommended by scientific studies, guidelines, and regulations. The operational limits for these sanitizers must be set above these minimum residual levels, and their optimization for different types of fresh-cut produce was the object of many studies [35]. It has been reported that in industrial settings with free chlorine concentrations below the operational limit of 10 mg/l, there was an induction of VBNC bacteria, and, in one study, 100% of 106 CFU/ml L. monocytogenes cells exposed to 12 mg/l free chlorine entered the VBNC state. The induction of the VBNC state was higher at higher concentrations of organic matter, and the use of the recommended operational limits for chlorine of 20–25 mg/l was effectively inactivated L. monocytogenes in PWW without the induction of VBNC cells. Therefore, the efficacy of the recommended washing practices was evaluated in a pilot plant.
For chlorine, the levels of culturable and VBNC L. monocytogenes cells in PWW were undetectable after 1 min of contact, independently of the type of washed produce. Instead, PAA did not completely inactivate the pathogen in any type of PWW at 80 mg/l concentration [35]. For ClO2 at the 3 mg/l recommended concentration, significant differences were observed among different types of PWW. In particular, the presence of VBNC cells was detected in shredded lettuce PWW. Since the recommended concentrations of PAA and ClO2 induced the VBNC state, it was concluded that the current operational limits of these disinfecting agents should be revised by increasing the concentration or the contact time [35].
The optimized vPCR PMAxx method developed by Truchado et al. [27] was used to evaluate the ability of VBNC L. monocytogenes and E. coli O157:H7 cells to tolerate sanitizers, cause cross-contamination and resuscitate in leafy greens during storage [20]. To determine the free chlorine concentration needed in PWW to induce the VBNC state, the concentrations 2.5 and 5 mg/l, were tested on a mixture of six L. monocytogenes serotype 1/2a isolates from leafy vegetables. With 2.5 mg/l chlorine, VBNC and culturable L. monocytogenes cells were detected while with 5 mg/l chlorine, all cells were inactivated. The VBNC cells in the PWW cross-contaminated the product after 1 min wash with 3 mg/l of ClO2, and after 5, 10, and 15 days of storage at 7 °C at least one replicate originated colonies indicating the resuscitation of VBNC cells. On iceberg lettuce and baby spinach during the 15-day shelf life, the levels of inoculated VBNC cells did not significantly change, though these were not culturable [20].
Sterile distilled water (SDW), baby spinach rinse water (SRW), and romaine lettuce rinse water (LRW) were used as media to carry out sanitizer treatments against 8 Log CFU/ml of two foodborne strains of L. monocytogenes serogroup 1/2a. In SDW, L. monocytogenes became undetectable by selective plating after exposure to chlorine at a concentration of 0.5 mg/l for 30 s, while in spinach or lettuce rinse water, due to the presence of organic matter, inhibition of growth in plate was obtained with 10 mg/l of free chlorine. L. monocytogenes was undetectable in plate after treatment with 30 mg/l PAA, except for one colony in one of the four replicates for LRW. For PAA treatment, the effect of organic matter on pathogen inactivation was much less pronounced than for chlorine. After exposure to PAA 30 and 50 mg/l in different media, the pathogens were detected only by vPCR with a decrease of only 1–2 Log from the initial numbers [44].
For treatments with free chlorine in SRW and LRW, selective plating and vPCR were significantly correlated. In SDW, L. monocytogenes at an initial level of 7.88 Log CFU/ml became undetectable by selective plating, non-selective enrichment, and vPCR with PMAxx when exposed to 50 mg/l free chlorine for 30 s, whereas it was detected by vPCR, but not by selective plating, following treatment with 100 mg/l PAA. Cells treated with free chlorine stained red or brown with the LIVE/DEAD BacLight Bacterial viability kit, while in the PAA-treated samples, most cells stained green suggesting uncompromised membrane integrity and viability [44].
Negative stain transmission electron microscopy (TEM) showed that most cells were damaged with electron-dense internal aggregations, large electron-lucent areas, and shrunk cytoplasm separated from the cell wall. However, a small proportion of cells resembled untreated cells and were probably VBNC. These cells could not be resuscitated so the authors hypothesized that they could be damaged though the cell membrane was intact. Nevertheless, since chlorine-induced VBNC L. monocytogenes had been previously shown to remain infectious for Caenorhabditis elegans, the possibility that these cells pose a food safety risk was not excluded [44].
CFDA/PI differential staining coupled with FC revealed three different subpopulations of L. monocytogenes Scott A outbreak strain, serogroup 4b, when exposed to PAA or low pH adjusted with HCl or acetic acid (AA), namely viable and metabolically active cells with an intact membrane (CFDA+PI−), dead cells with a damaged membrane (CFDA−PI+), and metabolically active cells with a damaged membrane classified as sub-lethally injured (CFDA+PI+). The CFDA+PI− cells were considered to be in VBNC state if unable to restart growth in tryptic soy yeast extract broth or agar (TSBYE or TSAYE) [22].
After exposure to 20, 30, and 40 ppm of PAA at 20°C for 1, 2, and 3 h, CFDA+ metabolically active cells ranged between 40.00% and 47.27%. The percentages of injured cells (CFDA+/PI+) increased after 2h of incubation. After exposure for 3h to 40 ppm PAA at 20°C, the total culturable and non-injured populations significantly decreased, and the whole population was nonculturable and CFDA+, thus indicating the induction of the VBNC state [22]. Exposure to 20 ppm PAA at 20°C for 90 min and 180 min resulted in high percentages of CFDA+PI− cells, 16.17 and 13.33 folds compared to the control, while exposure to 30 and 40 ppm of PAA for 90 and 180 min increased the proportion of dead cells. However, exposure to higher PAA concentrations for 30 min led to the formation of numbers of 5.8 to 8.5 fold higher injured cells than the control [14].
Exposure to high acidity at pH 3.0 and 2.7 adjusted with HCl determined a small decline in metabolically active cells after 150 and 300 min. The decline was higher when AA was used as an acidifying agent, with a reduced formation of CFDA+PI+ cells and an increase of dead cells. The number of cells recovering on TSAYE compared to the non-agarized medium was significantly lower in all conditions, except for treatment with 20 ppm PAA for 180 min and 30 ppm PAA for 90 min, interpreted as the ability of PAA to induce the VBNC state in L. monocytogenes. After exposure to pH 3.0 adjusted with HCl a small percentage of cells became non culturable and the percentage increased to 33% at pH 2.7 after 300 min but only a small fraction of cells seemed to be in the VBNC state. When acidification to pH 2.7 was done with AA, a significant percentage of CFDA+PI− cells lost culturability, thus indicating VBNC induction. The lag phases for cells able to recover increased to a maximum of 120 h for cells exposed to PAA and to 21-24 h for exposure to pH 2.7 for 150-300 min [14].
The effects of two disinfectants commonly used in the seafood industry, one containing a quaternary ammonium compound and glutaraldehyde (QA) and one containing hydrogen peroxide (HP), were evaluated for the ability to induce VBNC cells of L. monocytogenes in single-species and mixed-species biofilms. L. monocytogenes Lm1 (serogroup 1/2a-3a), Carnobacterium maltoromaticum P1AV5, and C. divergens P9AV5, all isolated from food contact surfaces of seafood processing plants, were used to produce biofilms on stainless steel and polyvinyl chloride (PVC) coupons. The biofilms were treated for 20 min at room temperature with 2% (v/v) solutions of the disinfecting agents according to the recommendations for use [45].
The difference between the number of cells determined by qPCR and vPCR with PMA, expressed as genome equivalents (GE), represented the number of dead cells while the difference between the number of cells determined by vPCR and counts on agar plates corresponded to the number of VBNC cells. Listeria cells remained viable and culturable in the biofilm not treated with disinfectants. The difference between qPCR and vPCR measurements in biofilms of stainless steel were 2.06 Log GE/cm2 and 1.70 Log GE/cm2 after HP and QA treatment, respectively. However, the decreases in L. monocytogenes culturable cell numbers were 5.22 Log CFU/cm2 and 4.49 Log CFU/cm2, respectively, indicating the presence of VBNC cells in the biofilms with differences of 2.95 log GE/cm2 and 2.12 log GE/cm2 compared to the culturable cells. Similar results were obtained for the biofilms on PVC [45].
Sampling of food contact surfaces in four commercial smoked salmon processing plants was also carried out before and after cleaning and disinfection for 8 months to examine the prevalence of culturable and VBNC L. monocytogenes. Before cleaning and disinfection, the prevalence of L. monocytogenes cells was 3.17% for culturable cells, 10.32% for viable cells, and 15.87% for total cells, while after cleaning and disinfection, the prevalence of L. monocytogenes was 1.59% for culturable cells, 9.52% for viable cells, and 14.29% for total cells, thus indicating a high rate of VBNC formation though with differences in prevalence among single plants [45].
Essential oils are natural antimicrobials particularly attractive for use in food production. Those extracted from Origanum vulgare L. (oregano, OVEO) and Rosmarinus officinalis L. (rosemary, ROEO), alone or in combination, inhibited L. monocytogenes in food matrices. Therefore, their capacity to induce L. monocytogenes VBNC formation in culture media and conditions mimicking food matrices was analyzed for a mixture of L. monocytogenes strains ATCC 7644 and ATCC 19112 (serotype 1/2c of human origin), and ATCC 19117 (serotype 4d of sheep origin). Exposure to 5 and 2.5 μL/ml of OVEO determined culturability loss after 60 min in phosphate-buffered saline (PBS) and after 180 min in meat broth. Exposure to 10 and 5 μL/ml of ROEO in PBS determined culturability loss after 60 and 180 min, respectively, while in meat broth culturability loss occurred after 180 min at both concentrations thus appearing delayed in the presence of nutrients [29].
L. monocytogenes cells that lost culturability did not recover after 6 h, in agreement with previous observations with L. monocytogenes Scott A. With a recovery time of 24 h, L. monocytogenes cells exposed to OVEO did not regain culturability, while those exposed to 5 μL/ml of ROEO in PBS and 10 μL/ml in meat broth restored culturability, with average viable counts in the range of 3.0 – 4.8 Log CFU/ml from initial viable counts of about 7 Log CFU/ml. These results indicated that L. monocytogenes cells exposed to ROEO in PBS and meat broth entered the VBNC state [29].
The presence of VBNC cells was analyzed by FC and double staining with bis-1,3-dibutyl barbituric acid (BOX) and PI as indicators of membrane potential and membrane integrity, respectively. Exposure to OVEO and ROEO caused the formation of 76.4 to 97.2% of cells with depolarized and damaged cell membranes (BOX+ PI+) in PBS and 97 to 98.5% in meat broth. After 24 h of recovery treatment, the cells with a viability phenotype (BOX− PI−), especially for cells exposed to ROEO, increased reaching 69.1-83.5% in PBS and 44.9 - 75.5% in meat broth [29].
The subpopulations FDA+ PI+, i.e. with damaged cell membrane but with enzymatic activity ranged from 33.5 to 55.8% and 19.7-44.8% for cells exposed to OVEO and ROEO in PBS and meat broth, respectively. The number of dead cells (FDA− PI+) ranged from 10.8 to 22.7% and 8.6-29.5% for cells exposed to OVEO and ROEO in PBS and meat broth, respectively. However, the subpopulations with undamaged cell membranes increased after 6 h of recovery treatment and, for cells exposed to ROEO, reached percentages of 72.6 and 69.7% after exposure in PBS and 99.9 and 86.6% after exposure in meat broth. After 24 h of recovery, cells exposed to ROEO in PBS showed 79.5 and 91% subpopulations with undamaged cell membranes. Subpopulations with enzymatic activity after exposure to OVEO and ROEO in meat broth increased from 58.5 to 71.7% after 24 h of recovery [29].
Cell efflux and respiratory functions, beyond membrane damage, might influence culturability. Therefore, ethidium bromide (EB) staining was performed to investigate efflux activity. This dye can cross intact cell membranes and is pumped out by cells with active efflux systems so cells with impaired efflux activity are EB+. Cells exposed to OVEO and ROEO originated similar percentages of EB+ subpopulations, namely 57.2-84.1% and 55.6-81% in PBS and meat broth. Gradual increases in EB− subpopulations were observed after 24 h of recovery from exposure to ROEO in PBS (34.1 and 61.4% for the concentrations 10 and 5 μL/ml). After 24 h of recovery treatment cells exposed to OVEO increased the efflux activity whereas those exposed to ROEO increased the EB+ subpopulation. The results of viable counts after recovery diverged for the two essential oils but cells in which the culturability was not restored had improved physiological functions and appeared to be viable. Further investigations should elucidate whether OVEO and ROEO-induced VBNC cells can restore the ability to multiply in food matrices [29].
Lavender essential oil (LEO) has an attractive aroma and is widely used in the food industry because of its broad antibacterial, antifungal, antiviral, nematicidal, and anti-mite properties. It is also largely used in pharmaceutical products and cosmetics for its multiple health-promoting properties. The effects of LEO against L. monocytogenes ATCC19115 were defined toward biofilms of the pathogen by using plate counts and vPCR with PMA. Biofilm formation of L. monocytogenes on sterile coverslips in polystyrene microtiter plates was evaluated by the crystal violet staining method [46].
The effect of LEO on L. monoctogenes was compared with that of sodium hypochlorite. The minimum inhibitory concentration (MIC) for LEO and sodium hypochlorite against L. monocytogenes were 1·6 v/v and 0·219 mg/ml, respectively. When treated with 4 MIC of LEO, the numbers of viable cells and culturable cells in biofilm formed at 32°C (warm biofilm) decreased by 6·35 and 9·01 Log CFU/ml, and those in the biofilm formed at 10°C (cold biofilm) decreased by 3·16 and 1·93 Log CFU/ ml, respectively. When treated with 4 MIC of sodium hypochlorite, the proportion of viable cells and culturable cells decreased by 3·68 and 4·76 Log CFU/ml in the warm biofilm, and 2·88 and 2·91 Log CFU/ ml in the cold biofilm, respectively. Consequently, LEO was more effective in reducing the number of viable cells compared to sodium hypochlorite. However, both LEO and sodium hypochlorite-induced VBNC populations that for LEO treatment, ranged in number from 1·74 to 3·58 and from 2·10 to 3·26 Log CFU/ml in the warm biofilm and cold biofilm, respectively [46].
After 40 days of treatment with 1/2 MIC of LEO, the numbers of VBNC cells for planktonic cells and biofilm were 2·51 and 0·50 Log CFU/ml, respectively. The VBNC cell numbers in the 1/2 MIC sodium hypochlorite treatment were 4·93 Log CFU/ml for planktonic cells and 1·08 Log CFU/ml for the biofilm. When biofilms were treated with LEO or sodium hypochlorite at the MIC for 40 days, the numbers of VBNC cells for planktonic cells and biofilm were 2·61 and 0·64 Log CFU/ml and 4·93 and 1·08 Log CFU/ml, respectively. Therefore, a low concentration of LEO induced fewer VBNC cells in biofilm and planktonic cells compared to sodium hypochlorite [46].
Electrolyzed water (EW) is generated through the electrolysis of diluted NaCl or HCl solutions in a specialized chamber yielding different types of EW, including slightly acidic electrolyzed water (SAEW), with disinfecting properties. SAEW has a pH of 5–6.5, and is gentle on surfaces so suitable for use in the food industry. SAEW contains chlorine, which has a strong oxidizing capacity and covalently binds to proteins modifying their structure in the chlorination process. To determine the concentration of chlorine required for VBNC induction, L. monocytogenes, cells were treated with SAEW for 5 min at different concentrations. With SAEW containing 4 and 6 mg/l of available chlorine (ACC) only 0.31 and 0.72 Log CFU/ml reductions were achieved, respectively, while 8 mg/l chlorine completely abolished culturability. Staining with the LIVE/DEADTM BacLightTM kit showed that 23 to 34% of the cells were viable after treatment with 8 and 10 mg/l of ACC, while standard plating provided negative results, indicating induction of the VBNC state. Most cells appeared to be intact by optical microscopy but scanning electron microscopy (SEM) showed a shrinkage of the cell membrane and the formation of aggregates in the VBNC cells [30].
Iron is essential for energy metabolism, cell signaling, gene expression, regulation of cell growth and differentiation but when the redox-cycling reactions catalyzed by iron are poorly controlled, the process of ferroptosis may occur. This process is an iron-dependent nonapoptotic cell death, also exploited for cancer therapy, in which iron in excess catalyzes the Fenton reaction in which H2O2 and iron generate hydroxyl radicals that cause lipid peroxidation [19].
L. monocytogenes ATCC19114 showed different susceptibilities to iron and it was susceptible to inorganic ferrous and ferric iron at a final concentration of 200 µM for 3 h. Cells exposed to FeSO4 did not grow in culture media, and also the organic compound Cit-Fe (III) led to failed colony formation in more than 90% of cells. L. monocytogenes ATCC19114 cells exposed to FeSO4 did not stain with PI nor with SYTOX in FC analysis so it was deduced that FeSO4 did not cause cell death. To elucidate if FeSO4 induced DNA degradation, the cells exposed to FeSO4 for 3 h were incubated in a saline solution containing 400 µg/ml thymol for 30 min and it was observed that thymol led to PI-stained cells, indicating that after FeSO4 exposure cells were viable [19].
The induction of VBNC cells by FeSO4 occurred in a dose-dependent manner, but even at a low concentration of 25 µM, FeSO4 induced the VBNC state in approximately 90% of cells. The process occurred in less than 15 min at a range of temperatures from 5 to 42 °C and for strains belonging to different serotypes. Morphological changes from rod-shaped to spheroids, size reduction, and shrinkage were observed in the FeSO4-induced VBNC cells. These were able to resuscitate in fresh BHI broth after a prolonged lag phase and were virulent for C. elegans with survival rates similar to normal L. monocytogenes cells after 10 days from infection [19].
Notably, while normal L. monocytogenes cells were efficiently killed by 240 µg/ml ampicillin, 60 µg/ml tetracycline and 60 µg/ml chloramphenicol, in BHI broth, VBNC cells developed resistance, though still remaining sensitive to tetracycline. When resuscitated, the population of VBNC cells displayed again the sensitivity to ampicillin [19].
Common indicators of ferroptosis, namely reactive oxygen species (ROS) and lipid peroxidation products, did not significantly increase in L. monocytogenes cells exposed to FeSO4, showing that ferroptosis did not occur. Exposure to the intracellular Fe2+ chelator 2,2′-Bipyridine almost completely blocked the VBNC state formation by FeSO4. After the addition of 2,2′-Bipyridine, VBNC formed after exposure to Haz-Tab chlorine tablets almost recovered growth as the normal cells while those formed by exposure to benzalkonium chloride (BC) died. It was concluded that the chelation of intracellular Fe2+ hindered VBNC formation, supporting the universal involvement of Fe2+ in VBNC state formation in L. monocytogenes. On the other hand, the application of Lip-1 and ascorbate inhibitors of ferroptosis blocked the VBNC state induction and arrested VBNC formation induced by BC or Haz-Tab with similar effects as 2,2′-Bipyridine. This observation suggested that cascades related to ferroptosis were involved in the VBNC state formation in L. monocytogenes [19].
Table 1 summarizes the results obtained on VBNC state induction in L. monocytogenes by antimicrobial compounds, with indication of the effective concentrations.
3.2. Effect of Physicochemical Stresses in Food Production on the VBNC Induction in L. monocytogenes
Some stress factors such as presence of particular ingredients, sub-optimal temperature and presence of inhibitors are inherent to food and food production processes. In a study regarding the use of Raman–deuterium isotope probing (Raman-DIP) microspectroscopy to characterize VBNC cells of L. monocytogenes, this technique was evaluated for the analysis of the effects of different food-related stressors, namely storage temperature, presence of salts and exposure to disinfectants using strains L. monocytogenes Lm1 and EGD-e. Viability was also analyzed by using qPCR, vPCR with PMA, and culturing. An incubation temperature effect was observed with the presence of a VBNC population at 37°C, and at 4°C. For cells exposed to 2% and 20% NaCl, differences of 1.5 Log UFC/ml and 0,72 Log UFC/ml between viable and culturable cells were observed for L. monocytogenes Lm1, thus showing the presence of a VBNC population. Results obtained for L. monocytogenes EGD-e were similar [47].
The persistence of L. monocytogenes in food processing environments is the major source of contamination of RTE foods. Therefore, a study was conducted to devise the rapid enumeration of viable L. monocytogenes cells by vPCR in a food processing environment after heat treatment and desiccation on stainless steel with and without biofilm formation. PMA concentration, incubation time and temperature, light source for photoactivation and exposure time, target genes sequence, and amplicon length were optimized to suppress the signal from dead cells. A 50 μM concentration of PMA was used since it appeared non-toxic to cells. A linearity was observed between viable L. monocytogenes cells enumerated using vPCR and plate counts when the difference between sanitizer-killed cells and viable cells did not exceed 4 Log CFU. However, vPCR with a short amplicon overestimated the number of viable cells (false positives) in samples containing cells killed by heat or isopropanol [18].
A long-amplicon vPCR assay with an estimated limit of quantification (LOQ) of 1.32 Log CFU/reaction correctly indicated that heat treatment at 90 °C for 15 min reduced the L. monocytogenes cell number from 8.02 Log CFU/ml to below the detection limit of 1.22 log CFU/ml, whereas a short-amplicon PMA-qPCR assay used for comparison provided false positive results. Viability PCR with either amplicon size caused a slight but not significant underestimation of the viable cell count. The membrane destabilization pretreatment with SDS before PMA treatment did not significantly enhance the effect of PMA in removing the false-positive DNA signal from heat-killed cells and was toxic for viable L. monocytogenes cells. Similar vPCR results were obtained for two strains, L. monocytogenes 568 and 08-5578, throughout an experiment of biofilm desiccation, except for the overestimation of the viable cell concentration at day 10 for L. monocytogenes 568. At day 10 both strains exhibited a remarkable drop in culturable cells, which was also observed with vPCR but only for L. monocytogenes 08–5578 [18].
In an experiment of biofilm desiccation for 8 days at 15 °C and 33% RH, the plate count quantified a reduction of culturable cells of 1.38 Log CFU/cm2 for the two L. monocytogenes strains. The reduction of viable cells measured by long-amplicon vPCR was 1.40 ± 0.09 log CFU/cm2 for L. monocytogenes 08–5578 and 0.71 ± 0.06 log CFU/cm2 for L. monocytogenes 568. Therefore, the desiccation survival of L. monocytogenes 568 was higher for biofilm cells than non-biofilm cells. The DNA extraction procedure was optimized to obtain a reliable cell number determination by the long-amplicon PMA-qPCR [18].
Zolfaghari et al. [42] evaluated the fate of L. monocytogenes ATCC 19115 (serotype 4b) in water microcosm and rainbow trout fillets with 0% or 30% NaCl at refrigeration temperature (4 ± 2°C). The results showed that bacteria in a water microcosm lost their culturability after 13 days with 0% NaCl concentration and after 27 days of incubation in the presence of 30% NaCl. However, bacteria in rainbow trout fillets remained culturable with and without NaCl. RT-PCR targeted on the 16 S rRNA gene was positive for all treatments during the period of the study, indicating the entering of L. monocytogenes into the VBNC state in water microcosm with 0% and 30% NaCl. Moreover, VBNC L. monocytogenes retained the expression of hly and inlA virulence genes so the authors concluded that VBNC L. monocytogenes retains pathogenicity [42].
Another study evaluated the ability of L. monocytogenes to form VBNC cells in the presence of high NaCl concentrations at different temperatures and to resuscitate under different nutritional conditions. Viability PCR with PMA was used and compared with the Chinese national standard method for L. monocytogenes enumeration. Four strains of L. monocytogenes were examined including L. monocytogenes ATCC 19115 (serotype 4b), ATCC 19111 (serotype 1/2a). Single colonies were inoculated into TSBYE and incubated until reaching the exponential phase, then an induction suspension containing around 106 CFU/ml of cells from the exponential phase was prepared with pH adjusted to 5.0. The suspension was aliquoted and stored at -20°C to ensure comparability among experiments [7].
At 4°C, the number of culturable bacteria decreased significantly in the presence of 20% NaCl and 30% NaCl, dropping below the limit of detection after 70 days in the 30% NaCl solution. Culturable L. monocytogenes ATCC 19115 was below the limit of detection on day 70 in the presence of 20% NaCl, while L. monocytogenes ATCC 19111 and MRL300007 decreased until day 21 or 49 and stabilized later. At 25°C, a significant decrease in culturable bacteria was observed for all four strains. In particular, L. monocytogenes ATCC 19115 at 20% and 30% NaCl and ATCC 19111 at 30% NaCl dropped below the detection limit on day 14. In the presence of 20% NaCl, the loss of culturability occurred on day 21 for some strains, while L. monocytogenes MRL300007 lost culturability in the presence of 30% NaCl on day 35 [7].
The VBNC counts of L. monocytogenes ATCC 19115 were 4.31 Log CFU/ml at 4 °C and 3.23 Log CFU/ml at −20 °C after 70 days at high-concentration of NaCl and all cells were in VBNC state. L. monocytogenes ATCC 19111 at 20% and 30% NaCl concentration showed a higher number of VBNC cells, 5.08 and 5.38 Log CFU/ml, respectively, compared to the other strains [7].
In the resuscitation experiments, only the strain ATCC 19111 was resuscitated without dilution of the VBNC cell suspension, while all strains recovered in BHI medium from 1000-fold diluted VBNC cell suspensions. Tryptic soy broth (TSB) plus 5% Tween 80 allowed a better resuscitation, except for L. monocytogenes MRL300007. TSB added with 2 mg/ml sodium pyruvate also improved resuscitation. Recovery in pasteurized milk at 4°C was observed only for L. monocytogenes ATCC 19115 VBNC cells. The same strain appeared to be the only one entering the VBNC state at 4°C. When the high concentration of NaCl solution stress was removed and pasteurized milk was added as a nutrient solution for incubation, only L. monocytogenes MRL300007 did not recover, maybe for the too low number of cells in VBNC state formed during incubation in the stressful conditions [7].
The types of fresh produce implicated in listeriosis outbreaks in the US included cantaloupe, packaged lettuce, stone fruits, and celery [48]. A study regarding the survival of L. monocytogenes on fresh-cut products during storage at the temperature values prevailing during commercialization found that for three strains from cantaloupe outbreaks, L. monocytogenes FS 2025 1/2b, FS 2030 1/2a, FS 2061 1/2b, the culturable cells significantly decreased on fresh-cut carrots, fresh-cut pineapple and fresh-cut celery and broccoli at 4 °C by. The populations of L. monocytogenes slightly increased on fresh-cut cantaloupe and mango and did not change significantly on fresh-cut apple, cauliflower, romaine lettuce, pepper, and onion [49].
Temperature abuse at 15 °C higher value than the normal storage temperature did not change the overall outcomes of L. monocytogenes survival except for blueberry, on which L. monocytogenes showed a lower decline. The rates of L. monocytogenes population decline were accelerated on fresh-cut carrots, apples tomatoes, grapes, and avocados while the growth rate on cantaloupe and mango increased. For cauliflower, celery, onion, and romaine lettuce L. monocytogenes growth was observed in temperature abuse conditions, while for fresh-cut broccoli and green peppers, the L. monocytogenes populations did not change significantly [49].
Live/dead staining of L. monocytogenes cells recovered from carrot showed a ratio of green fluorescent cells with intact membranes comparable to the initial inoculum, indicating that the loss of cultivability of L. monocytogenes did not reflect viability. When L. monocytogenes recovered from fresh-cut carrot was examined by vPCR with PMAxx, a viable population of 3.84 Log higher than that quantified by selective plating on Day 0 was detected. Moreover, the population determined by plating decreased below the LOD after storage at 4, 10, and 15 °C for 6 and 14 days, whereas vPCR showed a reduction of only 1 Log, indicating the presence of VBNC L. monocytogenes still after 2 weeks [49].
RTE foods with a pH lower than 4.4 and water activity lower than 0.92 are considered not permissive for the growth of L. monocytogenes, but those lacking the above characteristics can support the growth of L. monocytogenes and must be subjected to time/temperature control measures to prevent L. monocytogenes development during storage [24]. Raw carrots could be considered a food that supports the growth of L. monocytogenes based on pH (6.43–6.7) and water activity (aw > 0.98) values but several studies showed that this food matrix does not support the growth of L. monocytogenes and can even suppress its survival under various storage conditions thus showing intrinsic antilisterial activity by as yet unidentified substances [48].
Strains of the Listeria species L. grayi, L. innocua, L. monocytogenes, L. seeligeri, and L. welshimeri exposed to fresh-cut carrots for 15 min in 10% buffered peptone water (BPW) from initial concentrations of about 9 Log CFU/ml showed a recovery range of 6.6 - 7.3 Log CFU/ml. In addition, L. monocytogenes FS2025, a strain associated with an outbreak from cantaloupe, showed a recovery from boiled carrots ranging from 7.1 to 7.6 Log CFU/g against an average 4.7 ± 0.3 Log CFU/g recovery from fresh-cut carrot samples, indicating inhibition of Listeria spp. in fresh carrots [48].
Quantitative PCR without PMAxx pre-treatment showed that rinsates from both fresh-cut and boiled carrots contained comparable levels of L. monocytogenes cells. Moreover, L. monocytogenes in the rinsate from boiled carrot showed comparable levels in culture-dependent or culture-independent enumeration, while in the fresh-cut carrot rinsate, more than 99% of L. monocytogenes cells lost cultivability. On the other hand, vPCR showed similar numbers of L. monocytogenes in fresh-cut carrots and boiled carrot rinsates, implying that exposure to fresh-cut carrots could induce the VBNC state in L. monocytogenes. Transmission electron microscopy (TEM) showed the formation of mesosome-like structures in cells exposed to the rinsate from fresh cut-carrots whose role in the formation of VBNC cells should be elucidated [48].
The physicochemical stress factors other than antimicrobials that favored the formation of VBNC cells of L. monocytogenes in food or mimicking food stressors are synthesized in Table 2.
5. Ultrastructural, Molecular and Transcriptomic Changes in L. monocytogenes VBNC Cells
A thorough description of the physiological characteristics of L. monocytogenes VBNC cells is available for those induced by starvation in water. In the study of Carvalho et al. [21], the L. monocytogenes EGDe, serogroup 1/2a, was incubated for 28 days in commercial mineral water with a low mineral content and quality-controlled composition at initial concentrations ranging from 109 to 106 cells/ml. Culturability decreased at a higher rate and extent at lower starting bacterial concentration to less than 1 CFU/ml at day 28, while with an initial concentration of 108 cells/ml a reduction of 2 log culturable cells/ml occurred. FC carried out by staining the viable cell population with CFDA showed that the total and viable cell numbers remained nearly unchanged. Moreover, the ATP cell content, which is a marker of cell viability, declined after 7 days and increased later reaching levels higher than those expected from the culturable cells from day 21, thus indicating a contribution from the VBNC forms [21].
The use of L. monocytogenes cells expressing the Green fluorescent protein (GFP) allowed to observe since day 7 the formation of coccoid cells, sometimes near rod-shaped structures resembling empty cell walls, that increased in number until day 28. The coccoid cells did not stain as Gram-positive because of peptidoglycan loss that was complete at day 28, as shown by Ultra-High Performance Liquid Chromatography (UHPLC) analysis [21].
Differential staining of the cell wall with a fluorescent lectin able to bind teichoic acids and of the cell membrane with an antibody specific for L. monocytogenes of the 1/2 serogroup showed that rod-shaped cells, observed by cryogenic electron tomography (cryo-ET), presented cell wall gaps from which spherical protoplast egressed. This phenomenon was also observed for three other L. monocytogenes strains and other Listeria sensu stricto species L. ivanovii, L. innocua, L. marthii, L. seeligeri and L. welshimeri, thus suggesting that this is a common mechanism in Listeria species [21].
Known and putative autolysins, namely lmo0394, p60, aut, lmo1215, lmo1521, lmo2522, ami, namA, were strongly downregulated after 7 days. Therefore, the cell wall degradation was probably carried out by a pre-existing pool of autolysins. However, 90% of NamA-deficient mutants of L. monocytogenes EGDe still maintained the cell wall at day 14, compared to 44% of the wild-type. A similar behavior was observed in deletion mutants of the secA2 component of the Sec system ATPase that mediates the secretion of the NamA autolysin [21].
Gas chromatography coupled with mass spectrometry highlighted a decrease in the relative abundance of anteiso- branched-chain fatty acid species (a-BFA) a-C15:0 and a-C17:0, key regulators of membrane fluidity, and an increase of linear saturated (SFA) and unsaturated (UFA) fatty acids between day 0 and day 28. The use of a ratiometric probe that detects local membrane fluidity changes suggested a decrease in membrane fluidity during the first 14 days. Diffusion of the fluorogenic dye Nile red detected by total internal reflection fluorescence correlation spectroscopy (TIR-FCS) showed a reduction of the diffusion coefficient in rod-shaped cells between 7 and 14 days, the period of most severe cell wall damage, while coccoid cells showed a similar diffusion coefficient after 7 and 14 days, indicating an adaptation to a wall-less status [21].
Transcriptome analysis showed the downregulation of genes associated with nucleotide and coenzyme biosynthesis, transcription regulation, uptake of phosphate and some carbohydrates, cell envelope biosynthesis, and maintenance, cell division, energy production, and protein secretion at day 7. Upregulated genes included those encoding amino acid metabolism, uptake of glucose, mannose and metal ions, protein translation and folding, transport of carnitine/glycine betaine compatible solutes, that protect from osmotic stress, and glutathione metabolism, that protect from oxidative stress. Moreover, nearly half of the regulon controlled by the stress-response sigma factor SigB was induced. However, a SigB-deficient L. monocytogenes EGDe mutant became VBNC faster than the wild-type, indicating for SigB a modulation rather than an essential role for survival during starvation. The genes encoding the synthases relA, relP, and relQ of the alarmone guanosine pentaphosphate ((p)ppGpp) activator of the stringent response were not upregulated but a ∆relAPQ strain switched to the VBNC status faster than the wild-type indicating that the stringent response plays a role in the early phase of VBNC formation [21].
In another study on VBNC cells generated by SAEW treatment, global proteome analysis with tandem mass tags (TMT) labeling showed that RpsZ, RpmI, Lmo1306, BetL, Lmo2564, RplX, RpmD, RplT, RplO, RnpA, and Lmo0208 associated with ribosomal functions were significantly upregulated in VBNC cells, while proteins Lmo1608, FloA, TrpD, GadB, and GlyS related to aminoacyl-tRNA biosynthesis were significantly downregulated. Therefore, it was suggested that protein synthesis was initiated but peptide chain elongation was impeded. A down-regulation of LuxS in VBNC L. monocytogenes was observed that might indicate the inactivation of autoinducer-2 (AI-2) mediated quorum sensing (QS) and consequent inhibition of processes such as biofilm formation, production of virulence factors and toxins, motility, and drug resistance [30].
Comparative transcriptome profile showed that upregulated genes in FeSO4-induced VBNC cells were significantly more numerous than the control group. Significantly enhanced processes included stress response and ribosome binding with upregulation of transcription factors rpoC, rpoB, and rpoD, genes associated with the VBNC state formation lmo0668, trxA, trxB, and Imo0593, and genes associated to stress response lisK, lmo0539, lmo1502, lmo0521, lmo0913, rsbX, spxA, metK, tig, and htrA [19].
6. Conditions for VBNC L. monocytogenes Resuscitation
The process of resuscitation is defined as the conversion of non-culturable cells into culturable cells also without any change in cell numbers [14]. The resuscitation of VBNC L. monocytogenes was observed when the relative humidity was close to 100%, but not in a dry environment and possibly on cheese rind, but this was not unequivocally demonstrated. Regain of virulence was excluded by an experiment with human colon cells (HT-29), and in a murine model but pre-incubation in embryonated chicken eggs restored pathogenicity. Models of VBNC L. monocytogenes formation in human epithelial cells suggested that subpopulations of unculturable cells regained active growth [32].
Some studies used conventional culture media to demonstrate resuscitation of VBNC cells, such as TSAYE and TSBYE, BHI broth, tryptic soy broth (TSB) added with pyruvate or pasteurized milk and half Listeria Fraser broth with long incubation times [7,14,19,20]. However, in one study, resuscitation of chlorine and PAA-treated L. monocytogenes did not occur in TSB, TSB supplemented with 0.3% of sodium pyruvate or 100 U/ml of catalase, and synthetic medium M9 salts (Becton Dickinson) supplemented with 0.4% glucose and 0.5 mM of each of the 20 amino acids previously reported to stimulate resuscitation. In addition, cell-free supernatants from log phase cultures were tested as resuscitation media without success [44].
Resuscitation of VBNC L. monocytogenes induced by disinfectants was obtained by incubating 1 ml of lettuce homogenate in BPW in 9 ml of half Listeria Fraser broth, supplemented with 0.1% of sodium pyruvate, incubating for 20 h at 37 °C and then transferring 100 μL of the culture to 9 ml of Fraser supplemented with 0.1% sodium pyruvate incubated for 24 h at 37 °C. Samples were considered positive when the medium turned black. To confirm resuscitation, positive samples were streaked onto agar Listeria Ottaviani Agosti (ALOA) and OCLA and at least two typical L. monocytogenes colonies were tested by hly PCR after DNA extraction by boiling [20].
It was also reported that nutrient supplementation in culture media and non-embryonated chicken eggs did not allow the resuscitation of the VBNC forms of L. monocytogenes EGDe generated by prolonged starvation in water. Instead, inoculation into embryonated chicken eggs of the isogenic GFP-expressing VBNC variant exhibited growth two days after inoculation. The resulting cells showed the presence of a cell wall, were rod-shaped, and able to infect human trophoblasts (JEG-3) and hepatocytes (HepG2) after 6 h of contact with polymerization of host actin tails as efficiently as vegetative cells. This was a demonstration that VBNC forms of L. monocytogenes with prolonged survival under nutritional limitation can revert to infectious cells [21]. Regaining of virulence in animal models was observed in some studies [19].
Resuscitation factors (Rpfs), initially identified in the bacterium Micrococcus luteus are involved in the process. In dormant cells, peptidoglycan has a low turnover and its degradation is necessary to reinitiate active growth. The presence of an Rpf factor at a pM concentration was shown to increase the number of previously dormant cells by at least 100-fold. Similar proteins are widely distributed among high G+C Gram-positive bacteria, including corynebacteria, mycobacteria, streptomycetes, and some firmicutes and have been extensively employed to promote bacterial growth or resuscitation to enhance microbial diversity and isolate species difficult to culture. However, their mechanism of action is not completely elucidated. L. monocytogenes encodes two Rpf orthologs, Lmo0186 and Lmo2522, that have a lytic transglycosylase domain and muralytic activity on crude preparations of L. monocytogenes cell walls. These enzymes stimulated growth in minimal media but were not yet tested for the ability to resuscitate VBNC cells [32].
7. Discussion and Future Perspectives
From the recent scientific literature it emerged that most of the studies carried out were aimed at identifying the stress factors that induce L. monocytogenes VBNC formation in conditions mimicking those existing in food and food production plants, especially analyzing the effects of the most common sanitizers [20,27,35,44]. The evidence that four of these, i.e. chlorine, PAA, BC, and ClO2 led to the formation of VBNC populations appeared particularly worrisome for the consequences that can arise from the presence of viable L. monocytogenes in RTE products that go undetected by the standard culturing procedures. Those findings indicated that the VBNC issue should not be ignored and that the highly sensitive molecular methods of VBNC cell detection, such as FC coupled with viability staining, vPCR, isothermal amplification methods coupled with DNA intercalating dyes, and mRNA detection should be optimized and validated to develop shared procedures for routine application, together with the standard methods, for the control of this pathogen.
In particular, the methods based on mRNA detection, that are those requiring a minimum number of phases compared to the DNA-based methods, were applied only in a few instances and were not yet compared to the DNA-based methods for the sensitivity and specificity parameters [39,41,42]. In addition, a highly sensitive detection method not prone to the presence of amplification inhibitors, namely Droplet digital PCR (ddPCR), though being widely used to detect foodborne pathogens with low concentration in samples, was not yet applied for the detection of VBNC L. monocytogenes. In this technique, the PCR mixture is divided into nanodroplets, and the initial target DNA copy number in the sample can be quantified according to the number of positive and negative droplets, thus allowing absolute quantification without the need for a standard curve [26]. The application of ddPCR coupled with PMA treatment was already tested with Salmonella Typhimurium with promising results in terms of sensitivity [50].
Beyond mRNA, rRNA precursors (Pre-rRNA), which account for more than 25% of total rRNA in the cells, are used as targets in a technique named molecular viability test (MVT). In this technique, Pre-rRNAs are detected when pathogens are growing actively. The quantification of viable cells is achieved by comparing quantification cycle values produced by RT-qPCR before and after nutritional stimulation. MVT successfully detected viable L. monocytogenes and other pathogens in milk [28]. Also for RNA detection an isothermal amplification method named nucleic acid sequence-based amplification (NASBA) is available. This method requires three enzymes i.e., T7 RNA polymerase, RNase H and the Avian myeloblastosis virus (AMV) reverse transcriptase, and two primers, i.e. one including the T7 promoter sequence and one complementary to the target sequence, and a constant temperature of 41 °C. The entire reaction can be completed in 2 h through cycles of reverse transcription, RNA degradation, double-stranded DNA synthesis, and target transcription [28].
Sequencing-based techniques are increasingly applied to detect foodborne pathogens in microbial communities also allowing source-tracking in the food industry. Direct metatranscriptome RNA-seq and multiplex RT-PCR were compared in detecting viable E. coli O157:H7, Salmonella Enteritidis, and L. monocytogenes in Romaine lettuce and juice extract with nanopore MinION sequencing. Metatranscriptome analysis, which includes the steps of RNA extraction, library preparation, and sequencing, allowed the defining of the physiological state of the cells by mapping the sequences to metabolic databases (e.g., Kyoto Encyclopedia for Genes and Genomes (KEGG) database), although the sensitivity was lower than for multiplex RT-PCR. Moreover, the technique presents limitations such as a complex sample pre-treatment, the need for enrichment steps, and advanced bioinformatics expertise needing improvements for application to the study of VBNC cell physiology [51].
Limitations of VBNC molecular detection methods are the loss of some bacteria in the pretreatment steps and final results can affected by background noise. Therefore, IMS with immunomagnetic beads (IMB) conjugated with antibodies to capture target microorganisms was introduced but not yet been applied to detect VBNC cells. The use of bentonite-coated activated carbon (BCAC) instead of IMB, to remove DNA inhibitors also proved successful in sensitive detection of bacteria in milk. Other tools used to target foodborne pathogens were streptavidin magnetic beads-dual aptamers with high binding affinity, structural stability, ease of storage, and ease of synthesis in vitro and vancomycin-modified poly-l-lysine magnetic beads [28]. All these strategies should be evaluated in the ability to detect VBNC cells.
Among the natural compounds that attract interest as substitutes for chemical preservatives in food OVEO, ROEO, and LEO were tested and shown to favor VBNC L. monocytogenes induction. Therefore, beyond attempting concentration and exposure time optimization of their usage in different foods to prevent this hazard, other natural compounds need to be tested for the capability to induce VBNC formation in L. monocytogenes. An example is luteolin, a flavonoid that showed efficacy in destroying the cell membrane integrity, with an increase in the number of non-viable cells, producing modifications in cell morphology, inhibitory effects on biofilm formation, and enhanced antibiotics diffusion within biofilms. This molecule was found to exert a detrimental effect on three tested strains of L. monocytogenes treated with the MIC (32 to 64 µg/µl), with a an increase of nonviable cells identified by confocal laser scanning microscope combined with the Live/Dead BacLight Kit. A 2 MIC concentration of luteolin induced remarkable cell membrane damage, as demonstrated by field emission scanning electron microscopy (FESEM), and an increase of dead cells based on fluorescence staining, thus showing to be suitable for preventing VBNC formation [52].
Green sanitizer formulation, such as one comprising the natural phenolic compound gallic acid, hydrogen peroxide and DL-lactic acid, which induced the VBNC state in L. innocua, should be evaluated for VBNC formation also in L. monocytogenes strains, as well as erythorbyl laurate (EL) that showed promising results in the inactivation of L. innocua when coupled with mild heating [53,54]. Other antimicrobials to be tested in the capacity to inhibit VBNC induction are bacteriocins, paricularly those inhibiting listerial growth such as nisin, enterocin CRL35, and lactocin [55]. To date, only nisin has been tested for death induction in two L. monocytogenes strains in stationary phase by the BD Cell Viability Kit coupled with FC. It was observed that after 4 h of treatment with 75 μg/ml nisin, 25% of the cells were viable for the L. monocytogenes A1 strain, a food isolate, but after 24 h, 91.7% of the cells were dead or VBNC/injured. For the collection strain L. monocytogenes M5, 16.2% of cells were viable after 4 h and 95.2% of cells were dead after 24 h. Therefore, the capacity of nisin to leave a residual number of VBNC cells should be better investigated [56].
In addition, the effects on VBNC formation of alternatives to heat exposure to improve food microbiological safety should be defined. As an example, high-pressure technology (HPP) or pascalization, a low-temperature treatment of food that improves microbiological safety with minimal effects on the nutritional, functional, and/or sensory properties, is largely applied to extend the shelf life of many liquid and solid foods at refrigeration temperature, should be tested for induction of the VBNC state in L. monocytogenes. Indeed, some studies reported that, immediately after HPP with different combinations of pressure values (200–600 MPa), times of exposure (3–30 min), temperatures (4–45°C), and storage duration and temperature (14–75 days; 4–37°C), culturable L. monocytogenes was not detected but grew during storage, suggesting a possible VBNC state induction. However, none of those studies utilized appropriate methods to demonstrate this hypothesis [57]. Other physical treatment methods, e.g. radiofrequency and microwave applied to control microorganisms in a variety of products, must still be evaluated for the ability to induce the VBNC status [55].
A few studies reviewed here regarded the detection of VBNC cells of L. monocytogenes directly in food and on contact surfaces in food production plants after cleaning and disinfection, highlighting the presence of VBNC cells of the pathogen [17,34,43,45]. The findings strongly suggested that many more food categories and processes still need to be evaluated for the induction of VBNC L. monocytogenes and for the levels of these forms of the pathogen. Indeed, it is important to note that in a study regarding RTE salad the number of VBNC cells was higher than the law limit of acceptability for the pathogen at the end of the shelf-life period fixed on the basis of the standard culture method [17]. This result, together with different confirmations that food matrices such as fresh-cut carrots can favor L. monocytogenes VBNC formation [48,49], clearly indicated the necessity to use methods alternative to the culture-dependent ones to determine the presence of viable L. monocytogenes. An example is given by apples with or without wax coating on which a decrease of count for three major disease-causing serotypes, 1/2a, 1/2b, and 4b of L. monocytogenes was observed at 1°C and room temperature but it was not elucidated if the reduction was due to death or to the transition to the VBNC state [58].
The strong capacity to form biofilms is a characteristic of some L. monocytogenes strains such as L. monocytogenes EGD-e and it is favored by endotoxin proteins for initial adhesion and the SecA2 protein export pathway for structure formation and cell–cell interactions. In food processing environments, L. monocytogenes can form mixed biofilms with E. coli, Pseudomonas aeruginosa, Staphylococcus aureus, Lactiplantibacillus plantarum, and Vibrio parahaemolyticus that cause food spoilage and cross-contamination. Biofilm formation is one of the main mechanisms for L. monocytogenes persistence in food production plants so applying effective cleaning and disinfection treatments to destroy and prevent biofilm formation in food manufacturing plants is a necessity. In addition, the presence of VBNC cells in the biofilm was reported [46]. These insidious persistent forms of L. monocytogenes can be efficiently contrasted only by adopting multi-hurdle strategies, since physical treatments provide insufficient sterilization effects on biofilms, biological techniques are expensive and poorly stable, and chemical sanitizers induce pathogen tolerance and presence of residues [31]. Baicalin, the 7-O-glucuronide flavonoid derived from the roots of Scutellaria baicalensis, an inhibitor of AI-2 in biofilm-forming bacteria, postbiotics, that are metabolites derived from probiotics and showed great potential against biofilm-forming bacteria [59], bacteriophages or their recombinant endolysins [55] should be tested also against L. monocytogenes biofilms and prevention of VBNC formation .
The studies analyzing resuscitation came to different conclusions on the regain of culturability by VBNC L. monocytogenes even when the same non-selective culture media were used [19,44]. The discrepancies could be due to the different stages of the VBNC state for the cells tested, the different lengths of incubation time, and differences in the medium composition that can depend on the preparation procedures and ingredient quality. Moreover, the resuscitation capacity of VBNC L. monocytogenes in foods was little investigated by methodologies unequivocally demonstrating the VBNC status of the inoculated cells, except for a study carried out earlier than the year 2020 [32] and the experiments with pasteurized milk and leafy green by Zhao et al. [7] and Truchado et al. [20], respectively, so further investigations are required on this aspect.
8. Conclusions
This review highlighted that the study of VBNC cell formation in response to environmental factors in L. monocytogenes was mainly devoted to evaluating the effect of common sanitizers in the washing process of RTE salads, while other food sectors have been little considered. Only three studies regarded the natural detection of VBNC L. monocytogenes cells in foods during storage, indicating that more food categories known to be possibly contaminated by this pathogen, especially those dedicated to fragile consumers, should be investigated in this respect.
It must be underlined that, to date, no coordinated efforts were carried out for finding consensus strategies to combat VBNC pathogen occurrence in food, and an improvement of the VBNC cell detection methods is felt to be necessary to improve sensitivity and reduce laborious sample pre-treatment steps. To this aim, methods based on mRNA detection and ddPCR deserve particular attention for evaluation and validation in food safety assessment.
Variability among L. monocytogenes different foodborne strains in the ability to form VBNC cells should also be better evaluated.
Finally, cultural resuscitation methods should be applied to L. monocytogenes VBNC cells induced by different stress factors and at different stages of dormancy to identify the appropriate conditions that allow their detection.
Author Contributions
Conceptualization, all authors; methodology, F.R.; investigation, all authors; resources, G.C.; writing—original draft preparation, F.R.; writing—review and editing, P.P. and F.P.; supervision, G.C.; project administration, G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Italian Ministry of Health, grant number IZS AM 04_23 RC, project title “Miglioramento delle performance dei metodi di analisi utilizzati per la ricerca di Listeria monocytogenes in alimenti e tamponi provenienti da ambienti di produzione degli alimenti”.
Institutional Review Board Statement
Not relevant.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Roberts, B. N.; Chakravarty, D.; Gardner, J. C., 3rd; Ricke, S. C.; Donaldson, J. R. Listeria Monocytogenes Response to Anaerobic Environments. Pathogens 2020, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.; Strawn, L.K.; Chapman, B.J.; Dunn, L.L. A Systematic Review of Listeria Species and Listeria monocytogenes Prevalence, Persistence, and Diversity throughout the Fresh Produce Supply Chain. Foods 2021, 10, 1427. [Google Scholar] [CrossRef]
- Orsi, R. H.; Liao, J.; Carlin, C. R.; Wiedmann, M. Taxonomy, Ecology, and Relevance to Food Safety of the Genus Listeria with a Particular Consideration of New Listeria Species Described between 2010 and 2022. MBio 2024, 15, e0093823. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Listeriosis - Annual Epidemiological Report for 2022. https://www.ecdc.europa.eu/en/publications-data/listeriosis-annual-epidemiological-report-2022. 8 Feb 2024. Accessed on 1 November 2024.
- Castaño Frías, L.; Tudela-Littleton Peralta, C.; Segura Oliva, N.; Suárez Arana, M.; Cuenca Marín, C.; Jiménez López, J.S. Case Series of Listeria monocytogenes in Pregnancy: Maternal–Foetal Complications and Clinical Management in Six Cases. Microorganisms 2024, 12, 2306. [Google Scholar] [CrossRef]
- Van de Merwe, C.; Simpson, D. J.; Qiao, N.; Otto, S. J. G.; Kovacevic, J.; Gänzle, M. G.; McMullen, L. M. Is the Persistence of Listeria Monocytogenes in Food Processing Facilities and Its Resistance to Pathogen Intervention Linked to Its Phylogeny? Appl. Environ. Microbiol. 2024, 90, e0086124. [Google Scholar] [CrossRef]
- Zhao, Q.; Xu, Z.; Liu, X.; Zhu, H.; Li, Z.; Liu, Y.; Yang, J.; Dong, Q. Formation and Recovery of Listeria Monocytogenes in Viable but Nonculturable State under Different Temperatures Combined with Low Nutrition and High NaCl Concentration. Food Res. Int. 2024, 192, 114774. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2023 Zoonoses Report. EFSA J. 2024, 22, e9106. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Koutsoumanis, K. ; Allende, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; Nauta, M.; Nonno, R.; Peixe, L.; Ru, G.; Simmons, M.; Skandamis, P.; Suffredini, E.; Fox, E.; Gosling, R. B.; Gil, B. M.; Møretrø, T.; Stessl, B.; da Silva Felício, M. T.; Messens, W.; Simon, A. C.; Alvarez-Ordóñez, A. Persistence of Microbiological Hazards in Food and Feed Production and Processing Environments. EFSA J. 2024, 22, e8521. [Google Scholar] [CrossRef]
- Centorotola, G.; Guidi, F.; D’Aurizio, G.; Salini, R.; Di Domenico, M.; Ottaviani, D.; Petruzzelli, A.; Fisichella, S.; Duranti, A.; Tonucci, F.; Acciari, V. A.; Torresi, M.; Pomilio, F.; Blasi, G. Intensive Environmental Surveillance Plan for Listeria Monocytogenes in Food Producing Plants and Retail Stores of Central Italy: Prevalence and Genetic Diversity. Foods 2021, 10, 1944. [Google Scholar] [CrossRef]
- Bolten, S.; Lott, T. T.; Ralyea, R. D.; Gianforte, A.; Trmcic, A.; Orsi, R. H.; Martin, N. H.; Wiedmann, M. Intensive Environmental Sampling and Whole Genome Sequence-Based Characterization of Listeria in Small- and Medium-Sized Dairy Facilities Reveal Opportunities for Simplified and Size-Appropriate Environmental Monitoring Strategies. J. Food Prot. 2024, 87, 100254. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, F.; Butler, F.; Krasteva, I.; Schirone, M.; Iannetti, L.; Torresi, M.; Di Pancrazio, C.; Perletta, F.; Maggetti, M.; Marcacci, M.; Ancora, M.; Di Domenico, M.; Di Lollo, V.; Cammà, C.; Tittarelli, M.; Sacchini, F.; Pomilio, F.; D’Alterio, N.; Luciani, M. Integrative Analysis of Transcriptomic and Immunoproteomic Data Reveals Stress Response Mechanisms in Listeria Monocytogenes. Heliyon 2024, 10, e39832. [Google Scholar] [CrossRef] [PubMed]
- Arvaniti, M.; Orologas-Stavrou, N.; Tsitsilonis, O. E.; Skandamis, P. Induction into Viable but Non Culturable State and Outgrowth Heterogeneity of Listeria Monocytogenes Is Affected by Stress History and Type of Growth. Int. J. Food Microbiol. 2024, 421, 110786. [Google Scholar] [CrossRef]
- Vidovic, S.; Paturi, G.; Gupta, S.; Fletcher, G. C. Lifestyle of Listeria Monocytogenes and Food Safety: Emerging Listericidal Technologies in the Food Industry. Crit. Rev. Food Sci. Nutr. 2024, 64, 1817–1835. [Google Scholar] [CrossRef]
- Liu, J.; Yang, L.; Kjellerup, B. V.; Xu, Z. Viable but Nonculturable (VBNC) State, an Underestimated and Controversial Microbial Survival Strategy. Trends Microbiol. 2023, 31, 1013–1023. [Google Scholar] [CrossRef]
- Bernardo, R.; Duarte, A.; Tavares, L.; Barreto, A. S.; Henriques, A. R. Listeria Monocytogenes Assessment in a Ready-to-Eat Salad Shelf-Life Study Using Conventional Culture-Based Methods, Genetic Profiling, and Propidium Monoazide Quantitative PCR. Foods 2021, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Kragh, M. L.; Thykier, M.; Truelstrup Hansen, L. A Long-Amplicon Quantitative PCR Assay with Propidium Monoazide to Enumerate Viable Listeria Monocytogenes after Heat and Desiccation Treatments. Food Microbiol. 2020, 86, 103310. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhu, X.; Zhang, N.; Zhang, H.; Li, H.; Qi, Y.; Mo, H.; Hu, L. Direct Ferrous Sulfate Exposure Facilitates the VBNC State Formation Rather than Ferroptosis in Listeria Monocytogenes. Microbiol. Res. 2023, 269, 127304. [Google Scholar] [CrossRef]
- Truchado, P.; Gómez-Galindo, M.; Gil, M. I.; Allende, A. Cross-Contamination of Escherichia Coli O157:H7 and Listeria Monocytogenes in the Viable but Non-Culturable (VBNC) State during Washing of Leafy Greens and the Revival during Shelf-Life. Food Microbiol. 2023, 109, 104155. [Google Scholar] [CrossRef]
- Carvalho, F.; Carreaux, A.; Sartori-Rupp, A.; Tachon, S.; Gazi, A. D.; Courtin, P.; Nicolas, P.; Dubois-Brissonnet, F.; Barbotin, A.; Desgranges, E.; Bertrand, M.; Gloux, K.; Schouler, C.; Carballido-López, R.; Chapot-Chartier, M.-P.; Milohanic, E.; Bierne, H.; Pagliuso, A. Aquatic Environment Drives the Emergence of Cell Wall-Deficient Dormant Forms in Listeria. Nat. Commun. 2024, 15, 8499. [Google Scholar] [CrossRef]
- Arvaniti, M.; Tsakanikas, P.; Papadopoulou, V.; Giannakopoulou, A.; Skandamis, P. Listeria Monocytogenes Sublethal Injury and Viable-but-Nonculturable State Induced by Acidic Conditions and Disinfectants. Microbiol. Spectr. 2021, 9, e0137721. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Wagner, E.; Møretrø, T.; Heir, E.; Moen, B.; Rychli, K.; Langsrud, S. Pervasive Listeria Monocytogenes Is Common in the Norwegian Food System and Is Associated with Increased Prevalence of Stress Survival and Resistance Determinants. Appl. Environ. Microbiol. 2022, 88, e0086122. [Google Scholar] [CrossRef] [PubMed]
- European Community. Consolidated text: Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. OJ 2005, L 338 22.12.2005, https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A02005R2073-20200308, accessed on 29 November 2024.
- European Union. Commission Regulation (EU) 2024/2895 of 20 November 2024 amending Regulation (EC) No 2073/2005 as regards Listeria monocytogenes. OJ 2024, L 21.11.2024, https://eur-lex.europa.eu/eli/reg/2024/2895/oj, accessed on 29 November 2024.
- Dos Reis Lemos, L. M.; Maisonnave Arisi A., C. Viability dyes quantitative PCR (vPCR) assays targeting foodborne pathogens - scientific prospecting (2010–2022). Microchemical J. 2024, 197, 109769. [Google Scholar] [CrossRef]
- Truchado, P.; Gil, M. I.; Larrosa, M.; Allende, A. Detection and Quantification Methods for Viable but Non-Culturable (VBNC) Cells in Process Wash Water of Fresh-Cut Produce: Industrial Validation. Front. Microbiol. 2020, 11, 673. [Google Scholar] [CrossRef]
- Qi, Y.; Li, S.; Zhang, Y.; You, C. Recent Advances in Viability Detection of Foodborne Pathogens in Milk and Dairy Products. Food Control 2024, 160, 110314. [Google Scholar] [CrossRef]
- de Medeiros Barbosa, I.; da Cruz Almeida, É. T.; Gomes, A. C. A.; de Souza, E. L. Evidence on the Induction of Viable but Non-Culturable State in Listeria Monocytogenes by Origanum Vulgare L. and Rosmarinus Officinalis L. Essential Oils in a Meat-Based Broth. Innov. Food Sci. Emerg. Technol. 2020, 62, 102351. [Google Scholar] [CrossRef]
- Chang, H.-Y.; Gui, C.-Y.; Huang, T.-C.; Hung, Y.-C.; Chen, T.-Y. Quantitative Proteomic Analysis on the Slightly Acidic Electrolyzed Water Triggered Viable but Non-Culturable Listeria Monocytogenes. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Liu, J.; Xiang, Z.; Huang, T.; Xu, Z.; Ma, Q.; Yuan, L.; Soteyome, T. Development and Verification of Crossing Priming Amplification on Rapid Detection of Virulent and Viable L. Monocytogenes: In-Depth Analysis on the Target and Further Application on Food Screening. Lebenson. Wiss. Technol. 2024, 204, 116422. [Google Scholar] [CrossRef]
- Lotoux, A.; Milohanic, E.; Bierne, H. The Viable but Non-Culturable State of Listeria Monocytogenes in the One-Health Continuum. Front. Cell. Infect. Microbiol. 2022, 12, 849915. [Google Scholar] [CrossRef]
- Shi, X.; Yu, L.; Lin, C.; Li, K.; Chen, J.; Qin, H. Biotin Exposure-Based Immunomagnetic Separation Coupled with Sodium Dodecyl Sulfate, Propidium Monoazide, and Multiplex Real-Time PCR for Rapid Detection of Viable Salmonella Typhimurium, Staphylococcus Aureus, and Listeria Monocytogenes in Milk. J. Dairy Sci. 2021, 104, 6588–6597. [Google Scholar] [CrossRef]
- Panera-Martínez, S.; Capita, R.; García-Fernández, C.; Alonso-Calleja, C. Viability and Virulence of Listeria Monocytogenes in Poultry. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Gil, M. I.; Allende, A. Peroxyacetic Acid and Chlorine Dioxide Unlike Chlorine Induce Viable but Non-Culturable (VBNC) Stage of Listeria Monocytogenes and Escherichia Coli O157:H7 in Wash Water. Food Microbiol. 2021, 100, 103866. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Pang, X.; Li, X.; Bie, X.; Sun, J.; Lu, Y. Rapid and Accurate AuNPs-Sodium Deoxycholate-Propidium Monoazide-qPCR Technique for Simultaneous Detection of Viable Listeria Monocytogenes and Salmonella. Food Control 2024, 166, 110711. [Google Scholar] [CrossRef]
- Roumani, F.; Azinheiro, S.; Carvalho, J.; Prado, M.; Garrido-Maestu, A. Loop-Mediated Isothermal Amplification Combined with Immunomagnetic Separation and Propidium Monoazide for the Specific Detection of Viable Listeria Monocytogenes in Milk Products, with an Internal Amplification Control. Food Control 2021, 125, 107975. [Google Scholar] [CrossRef]
- Shi, D.; Shi, H. Combining Loop-Mediated Isothermal Amplification and Nanozyme-Strip for Ultrasensitive and Rapid Detection of Viable Listeria Monocytogenes Cells and Biofilms. Lebenson. Wiss. Technol. 2022, 154, 112641. [Google Scholar] [CrossRef]
- Wideman, N. E.; Oliver, J. D.; Crandall, P. G.; Jarvis, N. A. Detection and Potential Virulence of Viable but Non-Culturable (VBNC) Listeria Monocytogenes: A Review. Microorganisms 2021, 9, 194. [Google Scholar] [CrossRef]
- Stingl, K.; Heise, J.; Thieck, M.; Wulsten, I. F.; Pacholewicz, E.; Iwobi, A. N.; Govindaswamy, J.; Zeller-Péronnet, V.; Scheuring, S.; Luu, H. Q.; Fridriksdottir, V.; Gölz, G.; Priller, F.; Gruntar, I.; Jorgensen, F.; Koene, M.; Kovac, J.; Lick, S.; Répérant, E.; Rohlfing, A.; Zawilak-Pawlik, A.; Rossow, M.; Schlierf, A.; Frost, K.; Simon, K.; Uhlig, S.; Huber, I. Challenging the “Gold Standard” of Colony-Forming Units - Validation of a Multiplex Real-Time PCR for Quantification of Viable Campylobacter Spp. in Meat Rinses. Int. J. Food Microbiol. 2021, 359, 109417. [Google Scholar] [CrossRef]
- Azinheiro, S.; Ghimire, D.; Carvalho, J.; Prado, M.; Garrido-Maestu, A. Next-Day Detection of Viable Listeria Monocytogenes by Multiplex Reverse Transcriptase Real-Time PCR. Food Control 2022, 133, 108593. [Google Scholar] [CrossRef]
- Zolfaghari, M.; Rezaei, M.; Mohabbati Mobarez, A.; Forozandeh Moghaddam, M.; Hosseini, H.; Khezri, M. Virulence Genes Expression in Viable but Non-Culturable State of Listeria Monocytogenes in Fish Meat. Food Sci. Technol. Int. 2020, 26, 205–212. [Google Scholar] [CrossRef]
- Panera-Martínez, S.; Rodríguez-Melcón, C.; Del Campo, C.; Alonso-Calleja, C.; Capita, R. Prevalence and Levels of Cells of Salmonella Spp. and Listeria Monocytogenes in Various Physiological States Naturally Present in Chicken Meat. Food Control 2025, 167, 110770. [Google Scholar] [CrossRef]
- Gu, G.; Bolten, S.; Mowery, J.; Luo, Y.; Gulbronson, C.; Nou, X. Susceptibility of Foodborne Pathogens to Sanitizers in Produce Rinse Water and Potential Induction of Viable but Non-Culturable State. Food Control 2020, 112, 107138. [Google Scholar] [CrossRef]
- Brauge, T.; Faille, C.; Leleu, G.; Denis, C.; Hanin, A.; Midelet, G. Treatment with Disinfectants May Induce an Increase in Viable but Non Culturable Populations of Listeria Monocytogenes in Biofilms Formed in Smoked Salmon Processing Environments. Food Microbiol. 2020, 92, 103548. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Chen, Q.; Zhang, X.; Peng, J.; Zhang, M.; Zhong, Q. The Elimination Effects of Lavender Essential Oil on Listeria Monocytogenes Biofilms Developed at Different Temperatures and the Induction of VBNC State. Lett. Appl. Microbiol. 2022, 74, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Trigueros, S. , 2022. Metabolism measurement by Raman microspectroscopy : Application to the detection of viable but non-culturable Listeria cells, 2: en ligne. France. Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail", 'STAR - Dépôt national des thèses électroniques', 'Archive ouverte en agrobiosciences. Laboratoire de Boulogne sur mer. Retrieved from https://coilink.org/20.500.12592/49kr0zh on 08 Nov 2024. COI, 1259. [Google Scholar]
- Bolten, S.; Mowery, J.; Gu, G.; Redding, M.; Kroft, B.; Luo, Y.; Nou, X. Listeria Monocytogenes Loss of Cultivability on Carrot Is Associated with the Formation of Mesosome-like Structures. Int. J. Food Microbiol. 2023, 390, 110121. [Google Scholar] [CrossRef]
- Kroft, B.; Gu, G.; Bolten, S.; Micallef, S. A.; Luo, Y.; Millner, P.; Nou, X. Effects of Temperature Abuse on the Growth and Survival of Listeria Monocytogenes on a Wide Variety of Whole and Fresh-Cut Fruits and Vegetables during Storage. Food Control 2022, 137, 108919. [Google Scholar] [CrossRef]
- Wu, S.; Flint, S.; Yu, P.-L. Changes of Cell Permeability during Listeria Monocytogenes Persistence under Nisin Treatment. J. Food Process. Preserv. 2020, 44. [Google Scholar] [CrossRef]
- Yang, M.; Cousineau, A.; Liu, X.; Luo, Y.; Sun, D.; Li, S.; Gu, T.; Sun, L.; Dillow, H.; Lepine, J.; Xu, M.; Zhang, B. Direct Metatranscriptome RNA-Seq and Multiplex RT-PCR Amplicon Sequencing on Nanopore MinION - Promising Strategies for Multiplex Identification of Viable Pathogens in Food. Front. Microbiol. 2020, 11, 514. [Google Scholar] [CrossRef]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial Mechanism of Luteolin against Staphylococcus Aureus and Listeria Monocytogenes and Its Antibiofilm Properties. Microb. Pathog. 2020, 142, 104056. [Google Scholar] [CrossRef]
- Chang, Y.; Bai, J.; Yu, H.; Yang, X.; Chang, P.-S.; Nitin, N. Synergistic Inactivation of Listeria and E. Coli Using a Combination of Erythorbyl Laurate and Mild Heating and Its Application in Decontamination of Peas as a Model Fresh Produce. Food Microbiol. 2022, 102, 103869. [Google Scholar] [CrossRef]
- Mwangi, E.W.; Shemesh, M.; Rodov, V. Investigating the Antibacterial Effect of a Novel Gallic Acid-Based Green Sanitizer Formulation. Foods 2024, 13, 3322. [Google Scholar] [CrossRef]
- Gupta, P.; Adhikari, A. Novel Approaches to Environmental Monitoring and Control of Listeria Monocytogenes in Food Production Facilities. Foods 2022, 11, 1760. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Flint, S.; Yu, P.-L. Changes of Cell Permeability during Listeria Monocytogenes Persistence under Nisin Treatment. J. Food Process. Preserv. 2020, 44. [Google Scholar] [CrossRef]
- Wiśniewski, P.; Chajęcka-Wierzchowska, W.; Zadernowska, A. Impact of High-Pressure Processing (HPP) on Listeria Monocytogenes-an Overview of Challenges and Responses. Foods 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Su, Y.; Hua, Z.; Chiu, T.; Wang, Y.; Mendoza, M.; Hanrahan, I.; Zhu, M.-J. Evaluating Serotype-Specific Survival of Listeria Monocytogenes and Listeria Innocua on Wax-Coated Granny Smith Apples during Storage. Int. J. Food Microbiol. 2024, No. 110964, 110964. [Google Scholar] [CrossRef]
- Ozma, M. A.; Fadaee, M.; Hosseini, H. M.; Ataee, M. H.; Mirhosseini, S. A. A Critical Review of Postbiotics as Promising Novel Therapeutic Agents for Clostridial Infections. Probiotics Antimicrob. Proteins 2024. [Google Scholar] [CrossRef]
Figure 1.
Experimental workflows applied for the detection of viable cells of L. monocytogenes [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42].
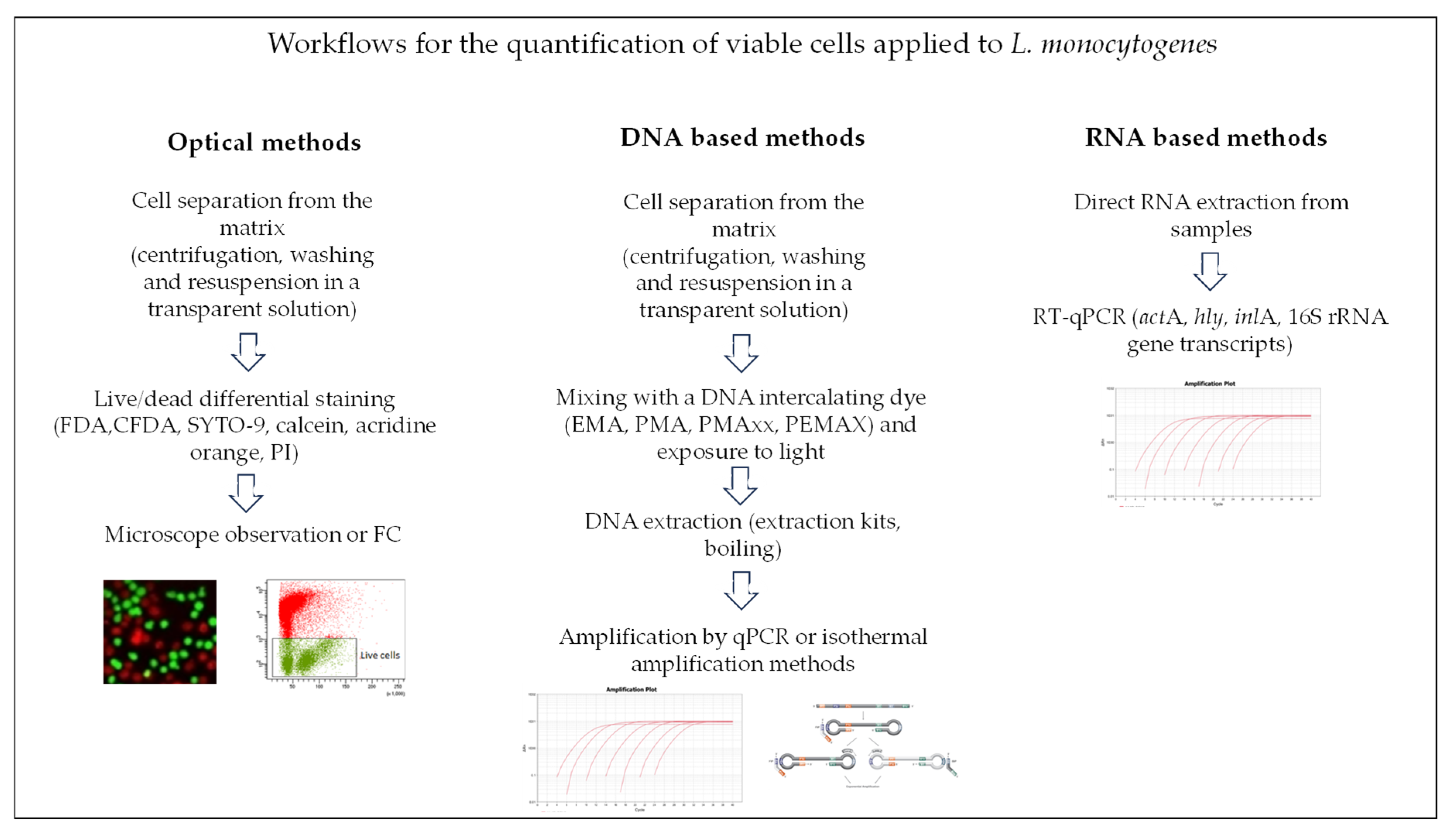

Table 1.
Antimicrobials that induced the VBNC state in L. monocytogenes, their concentration or intensity, and strains studied.
Table 1.
Antimicrobials that induced the VBNC state in L. monocytogenes, their concentration or intensity, and strains studied.
| Stress factor (s) | Intensity -concentration | L. monocytogenes strain | Reference |
|---|---|---|---|
| chlorine | 10 mg/l | cocktail of six vegetable isolates, serotype 1/2a | [27] |
| ClO2 | 3 mg/l | cocktail of six vegetable isolates, serotype 1/2a | [20,35] |
| PAA | 80 mg/l | cocktail of six vegetable isolates, serotype 1/2a | [35] |
| PAA | 100 mg/l | Two foodborne strains, serogroup 1/2a | [44] |
| PAA | 40 mg/l | Scott A, serotype 4b | [22] |
| AA | pH 2.7 | Scott A, serotype 4b | [14] |
| QA | 2% v/v | Lm1 (serogroup 1/2a-3a), seafood production plant isolate | [45] |
| HP | 2% v/v | Lm1 (serogroup 1/2a-3a), seafood production plant isolate | [45] |
| OVEO | 5 and 2.5 μL/ml in PBS and meat Broth | Mixture of ATCC 7644 and ATCC 19112 (serotype: 1/2c of human origin), and ATCC 19117 (serotype: 4d of sheep origin) | [29] |
| ROEO | 5 μL/ml in PBS and 10 μL/ml in meat broth | Mixture of ATCC 7644 and ATCC 19112 (serotype: 1/2c of human origin), and ATCC 19117 (serotype: 4d of sheep origin) | [29] |
| LEO | 4 MIC (MIC=1·6 v/v) | ATCC19115 | [46] |
| Sodium hypochlorite | 4 MIC (MIC=0·219 mg/ml) | ATCC19115 | [46] |
| SAEW | 8 and 10 mg/l of available chlorine (ACC) | BCRC14845 | [30] |
| FeSO4 | 200 μM | ATCC19114 | [19] |
PAA, peroxyacetic acid; AA, acetic acid; QA, disinfectant containing quaternary ammonium and glutaraldehyde; HP, hydrogen peroxide, OVEO, oreganon essential oil; ROEO, rosemary essential oil; LEO, lavender essential oil; SAEW, slightly acidic electrolyzed water.
Table 2.
Physicochemical stress factors that induced the VBNC state in L. monocytogenes during real or simulated food production conditions.
Table 2.
Physicochemical stress factors that induced the VBNC state in L. monocytogenes during real or simulated food production conditions.
| Stress factor (s) | Intensity - concentration | L. monocytogenesstrain | Reference |
|---|---|---|---|
| biofilm desiccation | 8 days at 15 °C and 33% RH | 568 | [18] |
| starvation | Water microcosm |
ATCC 19115 (serotype 4b) | [42] |
| NaCl | 30% | ATCC 19115 (serotype 4b) | [42] |
| NaCl | 2 – 20% | Lm1 (serogroup 1/2a-3a), EGD-e (serotype 1/2a) | [47] |
| NaCl | 20-30% at 4 and -20°C | Four strains including ATCC 19115 (serotype 4b), ATCC 19111 (serotype 1/2a) | [7] |
| fresh-cut carrots, | FS 2025 serotype 1/2b, FS 2030 serotype 1/2a, FS 2061 serotype 1/2b (cantaloupe outbreak) | [49] | |
| Raw carrots | FS2025 (serotype 1/2b cantaloupe outbreak) | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



