
Submitted:
25 December 2024
Posted:
26 December 2024
You are already at the latest version
Abstract
The potential of Streptomyces jeddahensis for lipid production was investigated, focusing on its ability to utilize various carbon sources under different temperature conditions. Glucose and fatty acid derivatives (sodium hexanoate, octanoate, and decanoate) were tested at 30°C, 37°C, and 45°C to optimize growth and lipid biosynthesis. The results revealed that 37°C was the optimal temperature for lipid accumulation, with sodium octanoate leading to the highest lipid content. While growth declined at 45°C, significant lipid production persisted, indicating an adaptive stress response. Gene expression analysis identified upregulation of key metabolic genes, including STSP_03080 (DGAT) and STSP_67970 (fabD), under thermal stress, highlighting enhanced fatty acid biosynthesis pathways. This study demonstrates the capacity of S. jeddahensis to efficiently convert medium-chain fatty acids into lipids, supporting its use as a sustainable and scalable platform for biodiesel production and other lipid-based industrial applications.
Keywords:
Streptomyces jeddahensis
; fatty acid derivatives
; sodium hexanoate
; sodium octanoate
; sodium decanoate
1. Introduction
The increasing global demand for sustainable and renewable energy sources has driven significant interest in microbial lipid production as a promising avenue for developing eco-friendly alternatives to fossil-derived oils. Among the oleaginous microorganisms, certain species of Streptomyces have gained attention due to their ability to accumulate lipids, particularly triacylglycerols (TAGs), which serve as precursors for biodiesel and various industrial bioproducts. These microorganisms can produce lipids constituting 20% to 80% of their biomass under nitrogen-limiting conditions[1,2,3,4]. making them a viable candidate for scalable lipid production. Oleaginous microbes, such as Rhodococcus opacus and Cryptococcus curvatus, have been extensively studied for their ability to accumulate TAGs when grown on both simple sugars, like glucose, and complex substrates, such as fatty acid derivatives[5], the process of TAG formation is illustrated in Figure 1.
This capability enables them to transform low-cost, renewable feedstocks into high-value lipids, highlighting their economic and environmental advantages[5,6]. Despite the substantial research focus on these organisms, Streptomyces species remain relatively underexplored. However, they demonstrate considerable potential for lipid production, particularly in synthesizing branched-chain fatty acids and high-valued lipids when cultivated with carbon sources such as glucose or cellobiose. One of the notable members of this genus, S. jeddahensis, stands out for its metabolic diversity and exceptional lipid accumulation capability, producing lipids up to 64 %(w/w) of its dry cell weight. This species can grow under a wide range of temperature conditions, up to 50°C, which is advantageous for industrial-scale production. These features make it a novel and promising host for the biotechnological development of lipid-based products[7].
The process by which Streptomyces accumulates lipids is influenced by multiple factors, with two of the most crucial being the carbon source[8] and growth temperature[9,10]. Carbon sources are essential because they provide energy for microbial growth and serve as precursors for lipid biosynthesis. While glucose is commonly used as a carbon source in many microbial cultures[11,12], certain fatty acids, have been shown to promote lipid accumulation in various microorganisms[13]. These fatty acids can directly impact metabolic pathways that lead to lipid formation, acting both as an energy source and as precursors for fatty acid biosynthesis. In particular, medium-chain fatty acids like sodium hexanoate are known to activate specific metabolic pathways, such as β-oxidation, which enhances lipid storage in many oleaginous microorganisms.
This study focuses on optimizing lipid production in S. jeddahensis using glucose and fatty acid derivatives, including sodium hexanoate, sodium octanoate, and sodium decanoate, as alternative carbon sources. Glucose is widely used due to its abundance, cost-effectiveness, and efficient metabolism through glycolysis to acetyl-CoA, a precursor for fatty acid synthesis[14]. Medium-chain fatty acids, such as hexanoate, octanoate and hexanoate, activate specific biosynthetic pathways, including β-oxidation, and enhance lipid accumulation[15,16,17,18,19]. By leveraging these substrates, this research aims to elucidate the metabolic potential of S. jeddahensis and its ability to serve as a sustainable platform for lipid production.
2. Materials and Methods
2.1. Medium, Bacterial Strains and Maintenance
The glucose and all other reagents were purchased from Sigma-Aldrich (Tokyo, Japan) and Nacalai Tesque, Inc. (Kyoto, Japan). Sodium hexanoate, sodium octanoate, and sodium decanoate used in this study were purchased from Tokyo Chemical Industry, co., Ltd (Tokyo). The oleaginous microorganism used in the experiments was Streptomyces jedahensis DSM 101878 sourced from the Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures. The strains were maintained at TSB agar medium containing (g/L): peptone from casein, 17; peptone from soymeal, 3; D(+)-glucose, 2.5; NaCl, 5; K2HPO4, 2.5; agar, 20.
2.2. Cultivation
In this study, the accumulation of lipids within S. jeddahensis cells was evaluated following a modified protocol adapted from Sass P et al[20], conducted in 250 mL Erlenmeyers flasks. The experimental setup began by introducing approximately 1 cm2 S. jeddahensis into 45 mL of Tryptic Soy broth (TSB) liquid medium. This initial culture was incubated at 30°C for 2 days at 160 rpm, serving as the pre-seed culture. For the seed culture, 5 mL of pre-seed culture was transferred into 50 mL of TSB medium and incubated under the same condition for 18 h. The fermentation phase seed culture with OD600=1 was transferred into 60 mL of Mineral Salt Medium (MSM) to facilitate the accumulation of triacylglycerols (TAGs). A range of carbon sources were examined, including glucose, sodium hexanoate, sodium octanoate, and sodium decanoate. MSM with reduced nitrogen content contains (g/L): urea, 0.25; yeast extract, 0.16; CaCl2, 0.02; KH2PO4, 1.52; MgSO4.7H2O, 0.52; Na2HPO4, 4, supplemented with different carbon sources: glucose, sodium hexanoate, sodium octanoate, and sodium decanoate at 2.5 g/L of each. Cultivation was performed on an orbital shaker at 160 rpm for 2 days. Cells were harvested by centrifugation at 10.000 x g for 5 minutes at 4°C and washed with MilliQ water. The cells were frozen at -20°C overnight and freeze-dried for 48 hours.
2.3. Dry Cell Weight Measurement
Cells from each culture were collected by centrifugation at 5000 x g for 5 minutes. The wet weight of the cell pellets was determined. Subsequently, the cell pellets were washed with sterile water. After washing, the cell pellets were reweighed. The collected cell pellets were then subjected to freeze-drying for 48 hours. The cell dry weight was recorded after freeze-drying before the lipid contents were extracted.
2.4. Lipid Contents
To prepare bacterial cell samples for lipid extraction, 30 mg of freeze-dried cells were combined with 0.6 mm zirconia beads in a 2 mL Eppendorf tube. The cell mixture was treated with lysozyme (5mg/mL) to lyse the bacterial cell walls and incubated for 30 minute. After incubation, the samples were freeze-dried again. Chloroform (1 mL) and methanol (0.5 mL) were added to the mixture. The samples were disrupted using a Shake Master Neo, operating at a speed of 1500 rpm for 5 minutes thrice. This disruption process was performed in three consecutive cycles to ensure thorough mixing. Following the disruption, the samples were centrifuged at 14,000 rpm for 5 minutes to separate the supernatant from the pellet. The supernatant was carefully removed, and 1.5 mL MilliQ water was added to the pellet. The samples underwent another round of disruption under the same conditions as previously described. After the second disruption, the supernatant was discarded, and the remaining sample was dried in a block incubator set at 60°C for 3 days. The final dry weight of the samples was recorded to calculate the lipid content by comparing it with the initial weight of the freeze-dried bacterial cells.
2.5. Fatty Acid Methyl Ester (FAME) Analysis
A gas chromatograph-mass spectrometer (GCMS-QP2020 NX, Shimadzu Corporation) was employed to analyze the cellular fatty acid composition. Chromatographic separation was achieved on a DB-23 column (60 m x 0.25 mm x 0.15 µm; Agilent Technologies, Palo Alto, CA). The temperature program for the analysis commenced at 50°C, which was maintained for 2 minutes, followed by a ramp of 25°C per minute to 175°C, and subsequently, a slower ramp of 4°C per minute to a final temperature of 230°C. Approximately 30 mg of freeze-dried bacterial cells were accurately measured and combined with 0.6 mm zirconia in a 2 mL microcentrifuge tube. The mixture was then treated with 500 µL of Reagent A and Reagent B, as specified by the fatty acid methylation kit’s instructions (Fatty acid Methylation Kit, Nacalai Tesque, Kyoto). Cell lysis was achieved by agitating the mixture with a Shake Master neo at 1500 rpm for three cycles of 5 minutes each. Following lysis, the sample was incubated at 37°C for 60 minutes on heat block to allow for the methylation of glycerides and sterol esters.
In subsequent step, 500 µL of Reagent C (from the same kit) was added to the mixture, which was then subjected to the same cell disruption protocol followed by a 20-minute incubation at 37°C to promote the methylation of free fatty acids. After the methylation process, 1 mL of the kit’s extraction solvent was added. The mixture was centrifuged at 14.000 rpm for 1 minute, and the clear supernatant was transferred into a new microcentrifuge tube. For Gas Chromatography-Mass Spectrometry (GC-MS) analysis, 90 µL of this supernatant was combined with 10 µL of an Internal Standard (IS) solution in a glass vial. The IS was prepared by diluting 10 µL of methyl benzoate in 990 µL of hexane, yielding a final volume of 1 mL.
2.6. Bodipy Staining for Qualifying Lipid Inside the Cells
To visualize intracellular triacylglycerol lipids, BodipyTN 493/503 (4,4-difluoro-5, 7-dimethyl-4-bora-3a, 4a-diaza-s-indacene) obtained from Invitrogen (Thermo Fisher Scientific) was utilized for staining. The dye was initially dissolved in ethanol to prepare a 5 mg/mL stock solution, which was subsequently stored at -20°C in the absence of light. Cell cultures (approximately 100 µL each) were centrifuged at 13,000 rpm for 30 seconds, and the supernatant was discarded. The cell pellets were washed with 100 µL phosphate-buffered saline (PBS) at pH 7. Following supernatant removal, the pellets were resuspended in 100 µL of the same buffer. To this suspension, 1 µL of Bodipy staining solution was mixed thoroughly. The stained cells were incubated at room temperature for 10 minutes. A 10 µL sample of the stained cell suspension was placed on a microscope slide for observation with a Keyence BZ-X810 fluorescence microscope.
2.7. qPCR Analysis of Gene Transcript Levels in S. jeddahensis Strain
qRT-PCR assays were used to investigate whether the gene was expressed. Firstly, S. jeddahensis strains were grown to OD600=1. cDNA from DNase-treated 0.5 µg of total RNA was synthesized in a 20 µl reverse transcription reaction containing random hexamers and SuperScript III reverse transcriptase (Invitrogen). All real-time PCR reactions were performed with a Bio-Rad iCycler (iQ5 system). All primer used are listed in Table 1. Changes in gene expression were calculated using the formula 2-∆∆Cq, as previously described[21]. Gene expression data for each run were organized into a matrix labeled as “countData,” where rows represented genes, and columns corresponded to samples. Metadata containing information about the samples, referred to as “colData,” included details on sample conditions and biological replicates, which were numbered (1, 2, or 3). Differentially expressed genes (DEGs) were identified by analyzing expression level variations between two groups of samples using the DESeq2 package in R, which applies a negative binomial distribution. DEGs were defined based on a threshold of |log2-fold change (LFC)| ≥ 2 and an adjusted p-value ≤ 0.05 (equivalent to -log10 adjusted p-value ≥ 1.30). A list of DEGs meeting these criteria was compiled. Furthermore, a protein-protein interaction (PPI) network analysis of the DEGs was performed using STRING (https://string-db.org/) version 11.5.
3. Results and Discussion
3.1. Effect of Temperature and Carbon Source on S. jeddahensis Growth
Microorganisms require the uptake of carbon sources and other nutrients from their environment to sustain cell growth and metabolic balance[22]. Additionally, temperature plays a crucial role in regulating bacterial growth [23]. This study evaluated the growth dynamics and metabolic efficiency of S. jeddahensis under varying carbon sources and incubation temperatures. These findings offer valuable insights into how temperature and carbon availability influence the bacterium’s growth and metabolism.
Glucose is commonly the preferred carbon source and is the primary energy source for many organisms[24]. Moreover, sodium hexanoate has been shown to support the growth of various bacterial strains, such as Cupriavidus necator[25]. Similarly, sodium octanoate is a potent carbon source for C. necator[26], supporting efficient biomass production without detrimental effects. Octanoate has been observed to promote effective biomass production, making it a favorable carbon source for bacterial growth. Several bacterial species can also utilize decanoate as a carbon and energy source[27]. The four carbon sources utilized in the study were glucose, sodium hexanoate, sodium octanoate, and sodium decanoate. The growth was monitored at three distinct temperatures 30°C, 37°C, and 45°C. Cell dry weight (CDW) measurements were recorded at 0, 12, 24, and 48 hours, with triplicate samples taken at each time point to ensure reproducibility and reliability of the data.
The results demonstrated that S. jeddahensis exhibited the highest CDW values at 30°C across all the tested carbon sources, indicating that this temperature is the most favorable for growth (Figure 2A, E, C, G). At this temperature, the bacterium showed robust growth, highlighting 30°C as the optimal biomass production condition. On the other hand, at 45°C, a significant decline in CDW was observed for all carbon sources, suggesting a critical threshold for thermal tolerance in this strain. This reduction in growth at higher temperatures may be attributed to cellular damage, protein denaturation, or impaired enzymatic activity, as previously noted by Schroda et al 2023[28]. Temperature sensitivity in S. jeddahensis could also be linked to the strain’s inability to maintain metabolic processes necessary for cell division and growth under thermal stress. Among the carbon sources tested, sodium octanoate emerged as the most efficient substrate for biomass production, with the highest variability in CDW observed at 30°C (Figure 2E). Interestingly, sodium hexanoate and sodium octanoate were consumed within 24 hours at 30°C and 37°C (Figure 2 D, F). However, at 45°C, the consumption rate slowed significantly, and these carbon sources were depleted much slower (Figure 2H). This further suggests that elevated temperatures inhibit key metabolic processes such as carbon source uptake and utilization and enzyme activities crucial for substrate metabolism[29]. Research has consistently shown that bacteria readily utilize medium-chain fatty acids like sodium hexanoate and sodium octanoate for growth. This was evident in the present study[25,26], where these substrates were efficiently consumed by S. jeddahensis. In contrast, glucose displayed the most significant reduction in residual levels at 45°C, with no measurable glucose remaining after 12 hours (Figure 2B). The rapid glucose consumption at 45°C suggests that glucose is rapidly metabolized under higher temperatures.
3.2. Lipid Accumulation in S. jeddahensis
Storage lipids play crucial roles in cells, including regulating membrane fatty acid composition, serving as a reservoir for reducing equivalents and metabolic intermediates, and acting as precursors for essential lipid biosynthesis, helping balance metabolism under fluctuating environmental conditions[30]. The experiment aimed to evaluate the lipid accumulation in S. jeddahensis cultured with different carbon sources (glucose, sodium hexanoate, sodium octanoate, and sodium decanoate) at three different temperatures (30°C, 37°C, and 45°C). The lipid content was measured as a percentage of cell dry weight (%, w/w), and Bodipy staining was used to visualize lipid accumulation within the cells. Figure 3 shows the observation of lipid accumulation in S. jeddahensis at 37°C using Bodipy staining, glucose (Figure 3A), sodium hexanoate (Figure 3B), sodium octanoate (Figure 3C), sodium decanoate (Figure 3D) indicating the presence of lipid accumulation within the cells. The other visualizations for 30°C and 45°C were shown in Figures S1, respectively.
The quantitative analysis of lipid content (%, w/w) across different temperatures and carbon sources is shown in Figure 4. The results indicate that temperature and carbon source significantly affect lipid accumulation in S. jeddahensis. At 30°C lipid contents were generally lower across all carbon sources. Glucose resulted in the higher lipid accumulation. At 37°C appeared to be optimal accumulation, with sodium octanoate leading to the highest lipid content, followed by glucose and sodium hexanoate. At 45°C lipid accumulation decreased at higher temperatures, with all carbon sources showing reduced lipid contents compared to 37°C. The result demonstrate that both the type of carbon source and the incubation temperature play crucial roles in lipid accumulation in S. jeddahensis. The highest lipid content was observed at 37°C with sodium octanoate as the carbon source. This suggests that medium-chain fatty acids (such as octanoate) are more efficiently converted into lipids by S. jeddahensis under optimal temperature conditions. Previous research has shown that oleaginous bacteria can accumulate significant amounts of lipids when provided with appropriate carbon sources and environmental conditions. For instance, a study by Yang et al [31] highlighted the potential of oleaginous microbes for lipid accumulation and renewable energy generation. They found that the choice of carbon source and culture conditions, such as temperature and nutrient availability, significantly influenced lipid production. Similarly, Koreti et al[32] reviewed the lipid accumulation capabilities of various oleaginous bacteria and emphasized the importance of optimizing culture conditions to enhance lipid yield. They noted that medium-chain fatty acids, like those used in this study, were particularly effective in promoting lipid accumulation. The enhanced lipid accumulation observed with sodium hexanoate and sodium octanoate can be attributed to the metabolic pathways involved in fatty acid synthesis. Oleaginous bacteria, including S. jeddahensis, can convert excess carbon into storage lipids, primarily triacylglycerols (TAGs). The efficiency of this conversion process is influenced by the chain length of the fatty acids provided as carbon sources. Bodipy staining, as used in this study, is a reliable method for visualizing lipid droplets within cells. Patel et al[33] discussed the advantages of using Bodipy for lipid detection, noting its high specificity and photostability.
3.3. Total Fatty Acid in S. jeddahensis
The presented stacked bar chart (Figure 5) reveals the total fatty acid (total FA) composition of S. jeddahensis under varying carbon sources and incubation temperatures. The carbon sources used were glucose, sodium hexanoate, sodium octanoate, and sodium decanoate, and the cultures were incubated at three different temperatures: 30°C, 37°C, and 45°C. This experiment highlights the interplay between carbon source utilization and environmental conditions in shaping fatty acid metabolism in S. jeddahensis. Each set of conditions induced specific shifts in the proportions of fatty acid types, underscoring the strain's metabolic flexibility. The stacked bar chart reveals the total (FA) composition of S. jeddahensis under varying carbon sources and incubation temperatures. The carbon sources used were glucose, sodium hexanoate, sodium octanoate, and sodium decanoate, and the cultures were incubated at three different temperatures: 30°C, 37°C, and 45°C. This experiment highlights the interplay between carbon source utilization and environmental conditions in shaping fatty acid metabolism in S. jeddahensis. Each set of conditions induced specific shifts in the proportions of fatty acid types, underscoring the strain's metabolic flexibility and potential applications in biotechnological industries. The type of carbon source strongly influenced the fatty acid composition of S. jeddahensis. Cultures grown on glucose, a simple carbon source, exhibited a balanced distribution of fatty acids, with C16:1 and C14:0 dominating the profiles across all temperatures. This indicates that glucose promotes a generalist metabolic pathway, producing both medium- and long-chain fatty acids in relatively stable proportions.
In contrast, cultures grown on sodium hexanoate displayed a marked increase in C16:1 and C16:0, particularly at 30°C. Hexanoate, a medium-chain fatty acid precursor, likely serves as a direct substrate for elongation and desaturation pathways, explaining the higher abundance of these medium-chain fatty acids. Similar trends were observed with sodium octanoate, where an elevated proportion of C16:0 and C16:1 was noted. Octanoate enhances the synthesis of longer-chain unsaturated fatty acids, reflecting its role in extending fatty acid chains through β-oxidation intermediates.
Sodium decanoate, a longer-chain fatty acid precursor, produced distinctive profiles characterized by increased C15:0 at 45°C. This indicates that decanoate promotes the biosynthesis of polyunsaturated fatty acids under high-temperature conditions, suggesting a potential stress adaptation mechanism by S. jeddahensis. These results demonstrate the strain’s ability to metabolize diverse carbon substrates into tailored fatty acid profiles, a feature that could be harnessed for industrial fatty acid production.
At 37°C, a more balanced distribution of saturated and unsaturated fatty acids emerged. This intermediate temperature likely optimizes enzymatic activities, enabling efficient chain elongation and desaturation. Cultures grown on glucose and sodium octanoate showed increased proportions of long-chain unsaturated fatty acids, such as C18:2, reflecting enhanced metabolic activity in fatty acid biosynthesis. At 45°C, significant shifts in fatty acid composition were observed, particularly in cultures grown on sodium decanoate. Polyunsaturated fatty acids, such as C18:2, became prominent, indicating an adaptive response to thermal stress. High temperatures likely induce the expression of desaturases, enzymes responsible for introducing double bonds into fatty acid chains. This adaptation enhances membrane fluidity, maintaining cellular integrity under heat stress. These findings align with previous studies that show elevated polyunsaturated fatty acid production in bacteria exposed to high temperatures.
The metabolic flexibility of S. jeddahensis revealed by this study highlights its potential for biotechnological applications. The ability to tailor fatty acid profiles by manipulating carbon sources and temperature conditions offers valuable insights into producing specific fatty acids for industrial purposes. For example, the elevated production of C16:1 and C18:1 under sodium hexanoate and octanoate supplementation could be exploited to synthesize biofuels, as these fatty acids are key components of biodiesel.
Moreover, the enhanced production of polyunsaturated fatty acids, such as C18:2 (Figure 5), The GC-MS results also showed higher peaks at 37°C and 45°C compared to 30°C. However, we were unable to identify the specific compounds responsible for these peaks, as they were not present in the BAME standard we used (Figure S3-S6), under high-temperature conditions and culture with sodium decanoate supplementation could interest the nutraceutical and pharmaceutical industries. This study's findings align with and extend previous research on bacterial fatty acid metabolism. For instance, Gorte et al. [34] demonstrated that carbon source and temperature significantly influence microbial lipid profiles, mainly glucose, which promotes the production of saturated fatty acids.
3.4. Gene Expression Analysis
As previously mentioned, the lipid content was highest when sodium octanoate was used as the carbon source at 37°C, even though the highest CDW was observed at 30°C. Another interesting finding from the lipid content result is that the lipid content was still significant at 45°C despite the cell dry weight not being very high (Figure 4). Bacterial responses to temperature changes highlight the induction of heat-shock proteins and other stress-related genes at higher temperatures[35]. We conducted a detailed gene expression analysis at 37°C and 45°C using sodium octanoate to understand the underlying mechanisms. We analyzed several genes that play critical roles in lipid metabolism, as detailed in Table 1, with rpoB as a housekeeping gene.
The STRING analysis emphasizes the functional importance of lipid biosynthetic genes in S. jeddahensis. The high connectivity of accD5_1 (acetyl-CoA carboxylase) underscores its role as a key regulator of fatty acid biosynthesis, catalyzing the first committed step in the pathway. The strong interactions of fabF and fabG, which are involved in elongation and reduction steps in fatty acid synthesis, further highlight the metabolic prioritization of lipid production[36]. Proteins such as plsC (1-acyl-sn-glycerol-3-phosphate acyltransferase) and gpsA (glycerol-3-phosphate dehydrogenase) were identified, underscoring their roles in glycerolipid biosynthesis. Environmental conditions, such as carbon sources and temperature, likely play a role in modulating the expression of these genes. Peripheral interactions involving plsC_1 suggest its role in glycerolipid and polyketide biosynthesis. Specifically, plsC_1, a bacterial 1-acyl-sn-glycerol-3-phosphate acyltransferase, catalyzes the acylation of lysophosphatidic acid to produce phosphatidic acid, a precursor for membrane glycerophospholipids.[37]. The bacterium's reliance on lipid biosynthesis under heat stress aligns with observations in other oleaginous microorganisms, where lipid accumulation is a protective mechanism. Regulatory proteins like pucR were also identified, indicating a potential role in modulating stress responses, including regulating heat shock proteins under thermal stress[38].
Regulatory proteins, pucR were also noted, indicating potential roles in modulating stress responses. Such regulatory elements mediate heat shock protein expression under thermal stress in other bacteria. These findings suggest that S. jeddahensis adapts to elevated temperatures by enhancing lipid biosynthesis pathways, with acyl carrier proteins playing a central role. Understanding these mechanisms lays a foundation for exploring thermally induced metabolic pathways and their industrial applications, such as biofuel production[31].
The RNA-Seq analysis compared the transcriptomes of S. jeddahensis at 37°C and 45°C using DESeq2. This analysis provided insights into temperature-dependent gene expression patterns, crucial for understanding the bacterium's metabolic adaptations under varying thermal conditions. The results were visualized using volcano plots, bar plots, and heatmaps, offering a comprehensive view of the differential gene expression data. The volcano plot (Figure 7A) highlights most genes with modest log2 fold changes (FC) and low statistical significance. However, a subset of genes displays substantial log2 FC values and high statistical significance (p-value < 0.05), suggesting their potential role as key metabolic regulators under specific conditions. Genes at the plot's extremes may encode enzymes critical for medium-chain fatty acid metabolism. Overall, only a few genes show significant differential expression. Features with high |log2 FC| (far left or right) and high -log10(p-value) are significantly upregulated or downregulated. The bar chart (Figure 7B) illustrates differential gene expression from the experiment, highlighting upregulated genes (red) such as STSP_67970 (fabD) and STSP_03080 (DGAT), alongside downregulated genes (blue) like STSP_12530. This suggests S. jeddahensis undergoes significant metabolic and regulatory shifts to adapt to varying temperatures. The upregulated genes are likely involved in carbon metabolism, heat response, or biosynthetic pathways, consistent with responses observed under stress conditions like the presence of different kinds of carbon source fatty acids and elevated temperatures. Conversely, downregulated genes may indicate suppression of specific pathways, such as glucose metabolism, in favor of fatty acid utilization [39].
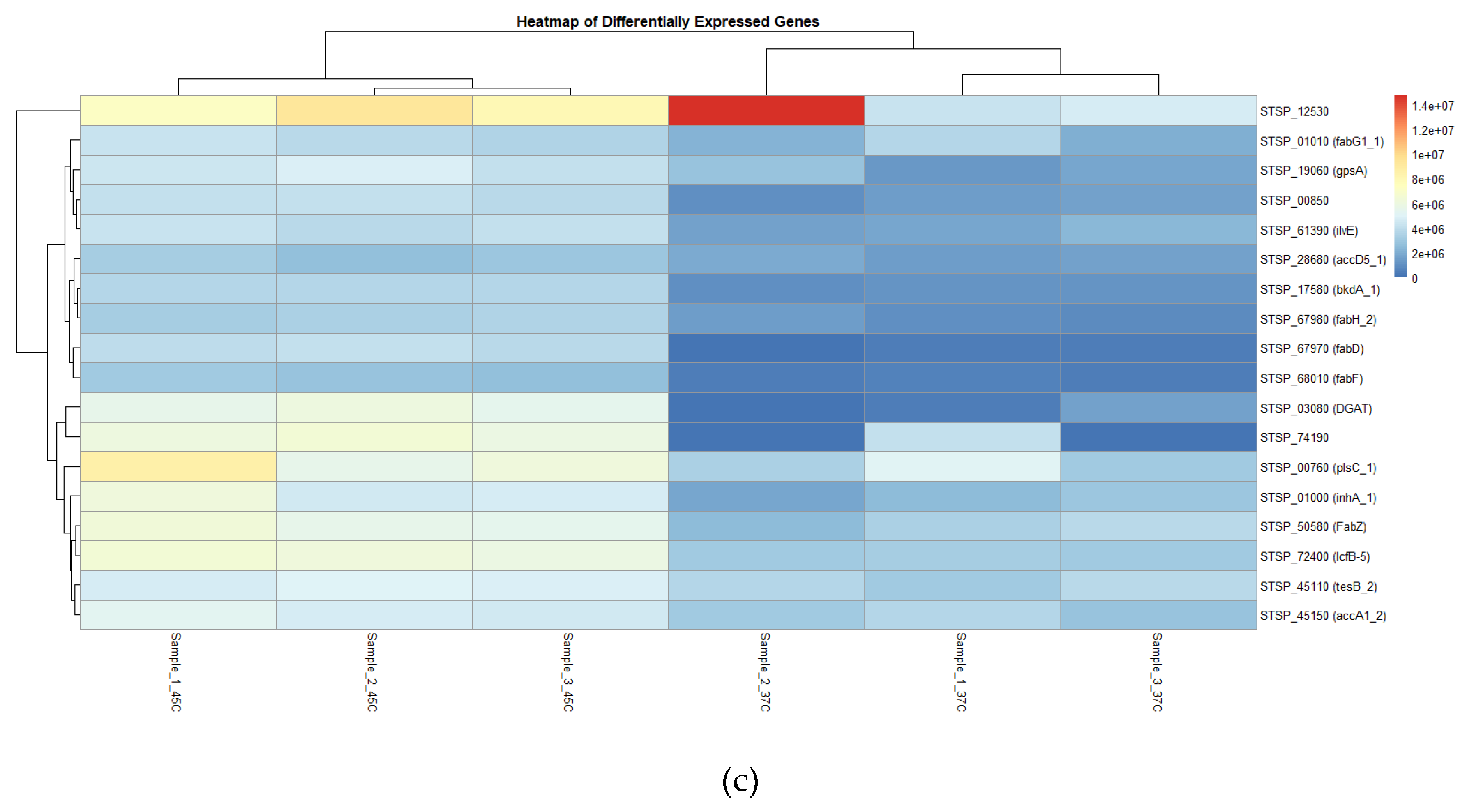
For instance, research by Bucca et al. [40] on S. coelicolor under thermal stress identified similar upregulation in heat shock proteins metabolism-related genes. This emphasizes conserved stress responses across Streptomyces species and provides insights into their adaptive mechanisms. The heatmap (Figure 7C) reveals distinct clusters of genes with higher expression (indicated in red) at 45°C compared to 37°C. For instance, genes such as STSP_12530, STSP_01010 (fabG1_1), and STSP_19060 (gpsA) exhibit elevated expression levels at 45°C. These genes likely involve critical metabolic pathways such as fatty acid biosynthesis and stress response. Notably, STSP_28660 (accD5_1), a gene associated with acetyl-CoA carboxylase activity, also showed a significant increase in expression, suggesting that higher temperatures may enhance lipid metabolism, potentially to cope with membrane fluidity changes at elevated temperatures.
Figure 6.
The STRING interaction network of the analyzed genes. Each filled node represents a gene, with edges between nodes indicating interactions between the protein products encoded by these genes. The edges are color-coded to represent different types of evidence supporting these interactions. Genes are color-coded based on their names and functional categories.
Figure 6.
The STRING interaction network of the analyzed genes. Each filled node represents a gene, with edges between nodes indicating interactions between the protein products encoded by these genes. The edges are color-coded to represent different types of evidence supporting these interactions. Genes are color-coded based on their names and functional categories.
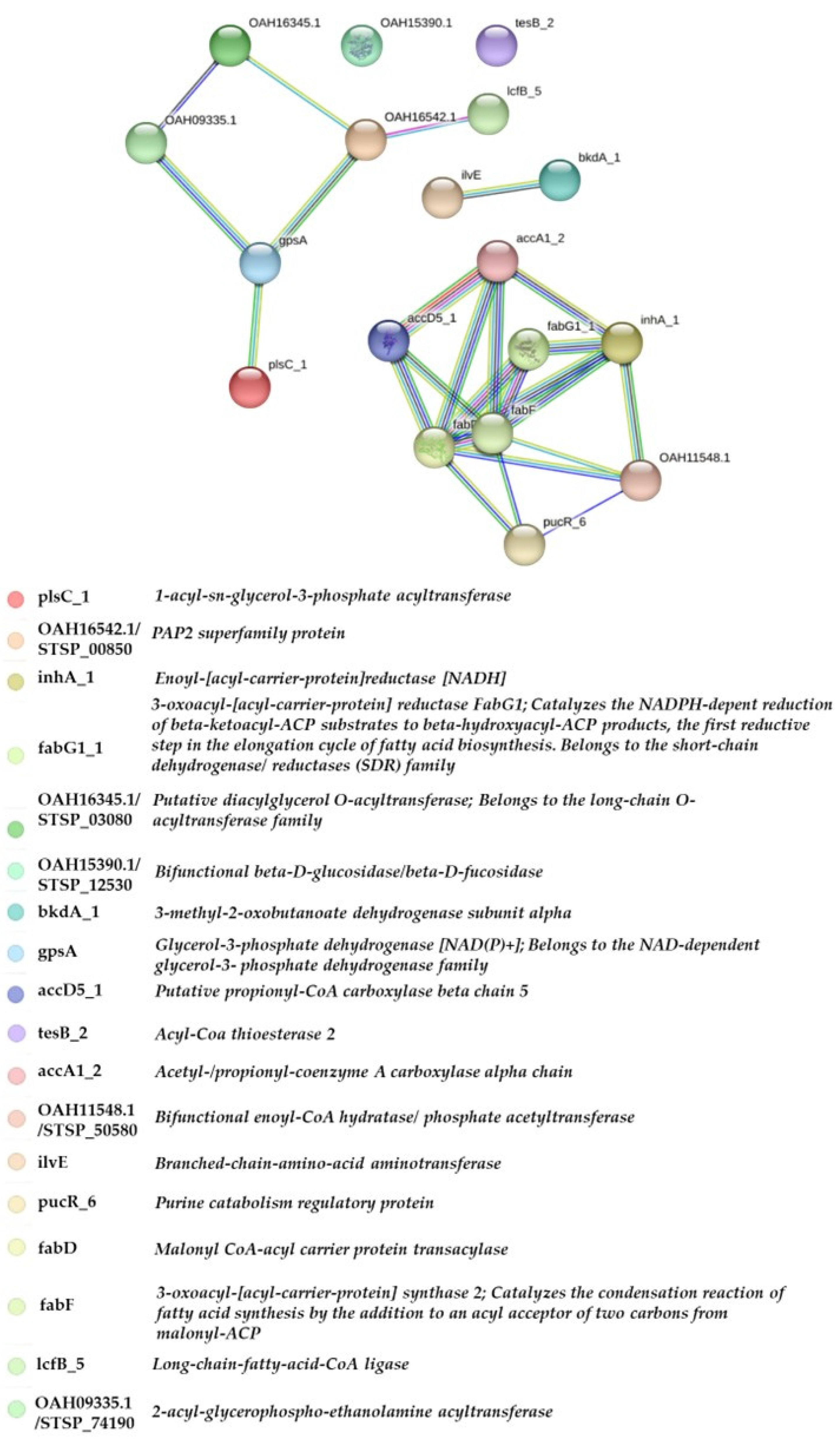
In contrast, specific genes demonstrated reduced expression at 45°C. For example, STSP_45150 (accA2_1) and STSP_45110 (tesB_2) showed lower expression levels at higher temperatures. These genes are likely involved in alternative metabolic pathways, which might be suppressed in favor of other pathways better suited for adaptation to heat stress. The clustering of gene expression profiles further highlights the functional partitioning of genes based on temperature, suggesting a shift in the metabolic network of S. jeddahensis to adapt to environmental conditions. The results align with the hypothesis that temperature significantly influences secondary metabolism in Streptomyces. Higher temperatures may induce stress-response pathways and enhance the biosynthesis of secondary metabolites, which are critical for the organism's survival under suboptimal conditions. Sodium octanoate, as a carbon source, appears to amplify these effects by promoting the activation of specific pathways related to lipid metabolism and energy production. This finding aligns with prior research that highlights the role of carbon source availability and temperature in regulating Streptomyces metabolic pathways.
The expression of genes such as STSP_03080 (DGAT), involved in triacylglycerol biosynthesis, indicates a potential accumulation of storage lipids at 45°C. This adaptive mechanism could serve as an energy reservoir to endure thermal stress. Similarly, the upregulation of STSP_61390 (ilvE), a gene associated with branched-chain amino acid metabolism, suggests an increased demand for amino acid precursors, which could play a role in maintaining protein stability and function under heat stress. Similar temperature-induced gene expression responses have been observed in other bacterial species. For instance, in Escherichia coli, the expression of heat shock proteins is regulated by RNA thermometers, which modulate gene expression in response to temperature changes[35]. These RNA thermometers are located in the 5’-untranslated region (5’-UTR) of genes encoding heat shock proteins and function as thermosensors to regulate gene expression at the post-transcriptional level[41]. The significantly upregulated genes at 45°C likely contribute to maintaining membrane fluidity and protecting cellular integrity under thermal stress.
In addition to acpP, other genes were identified as being temperature-responsive. Among the upregulated genes were STSP_68010 (fabF) encoding 3-Ketoacyl-ACP Synthase II, STSP_67970 (fabD) encoding a transcriptional activator of the fab operon, STSP_74190 encoding 1-Acylglycerol-3-Phosphate Acyltransferase (AGPAT), and STSP_03080 encoding Diacylglycerol Acyltransferase (DGAT). DGAT, a CoA-dependent acyltransferase enzyme, plays a key role in the synthesis of triacylglycerols (TAGs) in prokaryotes. Interestingly, DGAT exhibits dual activity, functioning both as an acyl-CoA:fatty alcohol acyltransferase (wax ester synthase, WS) and a diacylglycerol acyltransferase (DGAT), utilizing a wide range of acyl-CoA substrates for the synthesis of TAGs or wax esters (WEs)[42,43]. Conversely, some genes, such as STSP_12530, were downregulated in response to temperature changes. STSP_12530 encodes β-Glucosidase, an enzyme critical for bacterial metabolism, particularly in breaking down complex carbohydrates. β-Glucosidase hydrolyzes β-glycosidic bonds in compounds like cellobiose (a cellulose degradation product) and other oligosaccharides, converting them into glucose[44,45].
Lipid biosynthesis in bacteria depends on key metabolic intermediates such as pyruvate, acetyl-CoA, and glycerol-3-phosphate. Pyruvate, the end product of glycolysis, serves as a critical switch point for carbon flux distribution within central metabolism. It can act as a precursor for sugar phosphate synthesis via gluconeogenesis or be utilized to replenish tricarboxylic acid (TCA) cycle intermediates through anaplerotic reactions, thereby supporting anabolic processes. Together, these pathways highlight bacteria's metabolic versatility and adaptability to optimize lipid production under varying environmental conditions.
Figure 7.
Visual gene expression using Deseq2. (A) Volcano Plot of Differential Gene Expression. The volcano plot shows gene expression changes between 37°C and 45°C. Dots represent significant genes (log2FC > 1 or < -1, p < 0.05). (B) Bar chart of Log2 Fold Changes for Selected Genes. The bar plot highlights log2 fold changes between conditions, showing upregulation (Red) and downregulation (Blue). (C) The heatmap visualizes expression patterns across conditions, clustering genes like STSP_68000 and STSP_67970 based on significant temperature responsiveness. Red high and blue indicate low expression, respectively.
Figure 7.
Visual gene expression using Deseq2. (A) Volcano Plot of Differential Gene Expression. The volcano plot shows gene expression changes between 37°C and 45°C. Dots represent significant genes (log2FC > 1 or < -1, p < 0.05). (B) Bar chart of Log2 Fold Changes for Selected Genes. The bar plot highlights log2 fold changes between conditions, showing upregulation (Red) and downregulation (Blue). (C) The heatmap visualizes expression patterns across conditions, clustering genes like STSP_68000 and STSP_67970 based on significant temperature responsiveness. Red high and blue indicate low expression, respectively.
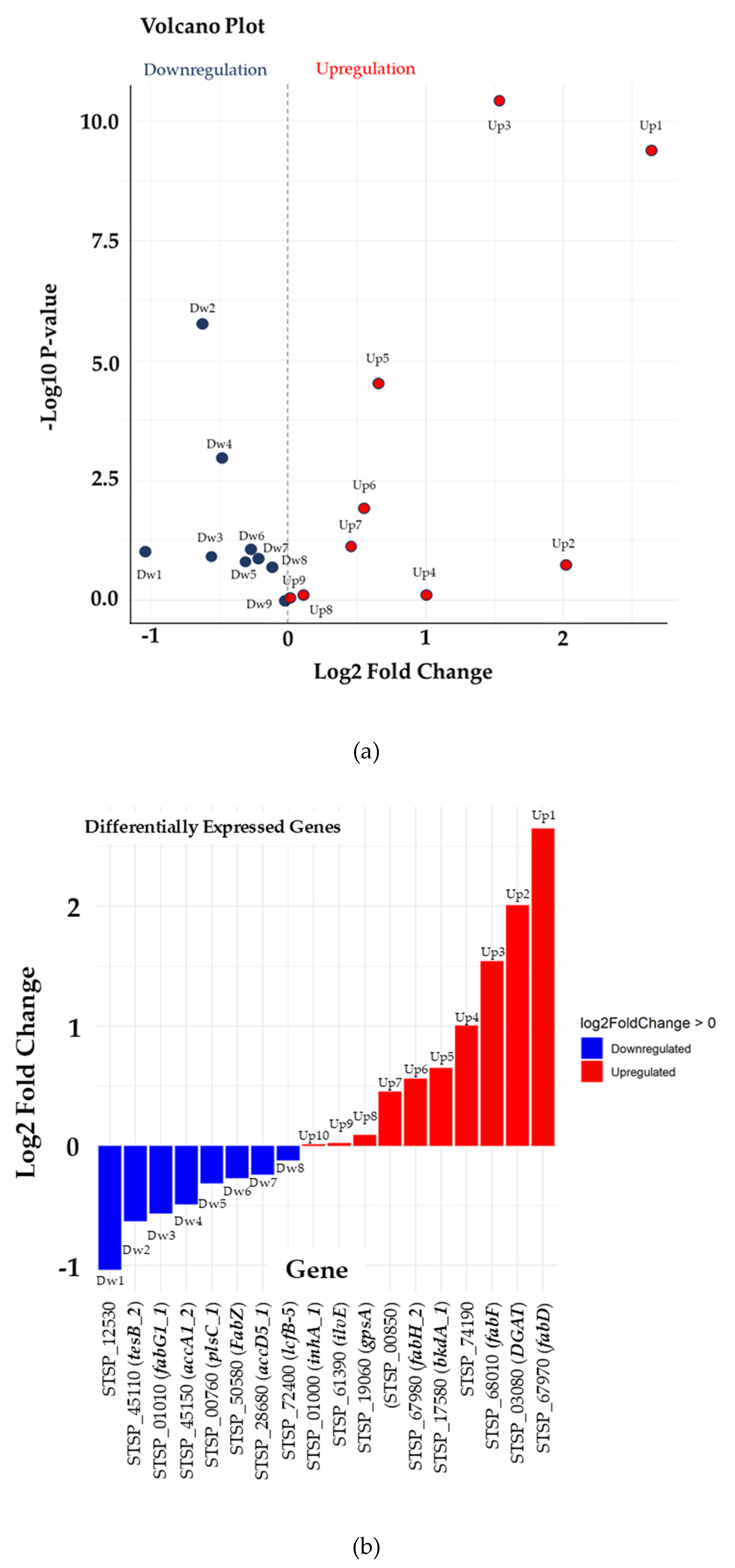
This study found that S. jeddahensis's ability to accumulate high levels of lipids, particularly when cultured with medium-chain fatty acids at optimal temperatures, suggests that this bacterium could be a valuable candidate for biodiesel production. Using renewable and cost-effective carbon sources, combined with optimized culture conditions, can enhance the feasibility and sustainability of microbial biodiesel production.
4. Conclusions
This study highlights Streptomyces jeddahensis as a robust and efficient microorganism for lipid production, particularly when cultured with medium-chain fatty acids under optimal conditions. Sodium octanoate emerged as the most effective substrate for lipid accumulation, with 37°C as the optimal temperature for growth and biosynthesis. Despite reduced biomass at higher temperatures, lipid production remained significant due to stress-induced metabolic adaptations. The upregulation of lipid biosynthesis genes such as STSP_03080 (DGAT) and STSP_67970 (fabD), emphasizes the bacterium’s potential for industrial biofuel applications. These findings establish S. jeddahensis as a promising candidate for renewable energy production, with opportunities for further optimization through genetic engineering and process development to enhance its industrial scalability.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Prihardi Kahar, Pamella Apriliana; Data curation, Pamella Apriliana, Prihardi Kahar; Formal analysis, Pamella Apriliana, Nova Rachmadona, Witta Kartika Restu; Funding acquisition, Chiaki Ogino; Investigation, Pamella Apriliana and Prihardi Kahar; Methodology, Prihardi Kahar, Pamella Apriliana; Project administration, Prihardi Kahar, Chiaki Ogino; Resources, Pamella Apriliana and Prihardi Kahar; Supervision, Prihardi Kahar, Chiaki Ogino and Akihiko Kondo; Validation, Pamella Apriliana and Prihardi Kahar; Writing – original draft, Pamella Apriliana and Prihardi Kahar.
Funding
This work was supported by the International Joint Program, Science and Technology Research Partnership for Sustainable Development (SATREPS) (Innovative Bio-production in Indonesia; iBioI) from the Japan Science and Technology Agency and the Japan International Cooperation Agency (JST and JICA), Japan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no potential conflicts of interest.
References
- Dávila Costa, J.S.; Leichert, L.; Alvarez, H.M.; Herrero, O.M. Label-Free and Redox Proteomic Analyses of the Triacylglycerol-Accumulating Rhodococcus Jostii RHA1. Microbiology (N Y) 2015, 161, 593–610. [Google Scholar] [CrossRef] [PubMed]
- Wältermann, M.; Luftmann, H.; Baumeister, D.; Kalscheuer, R.; Steinbüchel, A. Rhodococcus Opacus Strain PD630 as a New Source of High-Value Single-Cell Oil? Isolation and Characterization of Triacylglycerols and Other Storage Lipids. Microbiology (N Y) 2000, 146, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Röttig, A.; Atasayar, E.; Meier-Kolthoff, J.P.; Spröer, C.; Schumann, P.; Schauer, J.; Steinbüchel, A. Streptomyces jeddahensis Sp. Nov., an Oleaginous Bacterium Isolated from Desert Soil. Int J Syst Evol Microbiol 2017, 67, 1676–1682. [Google Scholar] [CrossRef]
- Amara, S.; Seghezzi, N.; Otani, H.; Diaz-Salazar, C.; Liu, J.; Eltis, L.D. Characterization of Key Triacylglycerol Biosynthesis Processes in Rhodococci. Sci Rep 2016, 6, 24985. [Google Scholar] [CrossRef]
- Kim, H.M.; Chae, T.U.; Choi, S.Y.; Kim, W.J.; Lee, S.Y. Engineering of an Oleaginous Bacterium for the Production of Fatty Acids and Fuels. Nat Chem Biol 2019, 15, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jin, G.; Wang, Y.; Shen, H.; Zhao, Z.K. Lipid Production on Free Fatty Acids by Oleaginous Yeasts under Non-Growth Conditions. Bioresour Technol 2015, 193, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Röttig, A.; Hauschild, P.; Madkour, M.H.; Al-Ansari, A.M.; Almakishah, N.H.; Steinbüchel, A. Analysis and Optimization of Triacylglycerol Synthesis in Novel Oleaginous Rhodococcus and Streptomyces Strains Isolated from Desert Soil. J Biotechnol 2016, 225, 48–56. [Google Scholar] [CrossRef]
- Peacock, L.; Ward, J.; Ratledge, C.; Dickinson, F.M.; Ison, A. How Streptomyces Lividans Uses Oils and Sugars as Mixed Substrates. Enzyme Microb Technol 2003, 32, 157–166. [Google Scholar] [CrossRef]
- Berry, E.D.; Foegeding, P.M. Cold Temperature Adaptation and Growth of Microorganisms. J Food Prot 1997, 60, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Beales, N. Adaptation of Microorganisms to Cold Temperatures, Weak Acid Preservatives, Low PH, and Osmotic Stress: A Review. Compr Rev Food Sci Food Saf 2004, 3, 1–20. [Google Scholar] [CrossRef]
- Sánchez, S.; Chávez, A.; Forero, A.; García-Huante, Y.; Romero, A.; Sánchez, M.; Rocha, D.; Sánchez, B.; Ávalos, M.; Guzmán-Trampe, S.; et al. Carbon Source Regulation of Antibiotic Production. J Antibiot (Tokyo) 2010, 63, 442–459. [Google Scholar] [CrossRef]
- L. Demain Arnold “Carbon Source Regulation of Idiolite Biosynthesis in Actinomycetes”. Regulation of Secondary Metabolism in Actinomycetes; CRC Press, 2020; ISBN 9781003068600.
- O-Thong, S.; Zhu, X.; Angelidaki, I.; Zhang, S.; Luo, G. Medium Chain Fatty Acids Production by Microbial Chain Elongation: Recent Advances. In; 2020; pp. 63–99.
- Lin, J.; Shen, H.; Tan, H.; Zhao, X.; Wu, S.; Hu, C.; Zhao, Z.K. Lipid Production by Lipomyces Starkeyi Cells in Glucose Solution without Auxiliary Nutrients. J Biotechnol 2011, 152, 184–188. [Google Scholar] [CrossRef]
- Kim, G.J.; Lee, I.Y.; Yoon, S.C.; Shin, Y.C.; Park, Y.H. Enhanced Yield and a High Production of Medium-Chain-Length Poly(3-Hydroxyalkanoates) in a Two-Step Fed-Batch Cultivation of Pseudomonas Putida by Combined Use of Glucose and Octanoate. Enzyme Microb Technol 1997, 20, 500–505. [Google Scholar] [CrossRef]
- Fei, Q.; Wewetzer, S.J.; Kurosawa, K.; Rha, C.; Sinskey, A.J. High-Cell-Density Cultivation of an Engineered Rhodococcus Opacus Strain for Lipid Production via Co-Fermentation of Glucose and Xylose. Process Biochemistry 2015, 50, 500–506. [Google Scholar] [CrossRef]
- Cabecas Segura, P.; Onderwater, R.; Deutschbauer, A.; Dewasme, L.; Wattiez, R.; Leroy, B. Study of the Production of Poly(Hydroxybutyrate-Co-Hydroxyhexanoate) and Poly(Hydroxybutyrate- Co -Hydroxyvalerate- Co- Hydroxyhexanoate) in Rhodospirillum Rubrum. Appl Environ Microbiol 2022, 88. [Google Scholar] [CrossRef]
- Hu, C.; Wu, S.; Wang, Q.; Jin, G.; Shen, H.; Zhao, Z.K. Simultaneous Utilization of Glucose and Xylose for Lipid Production by Trichosporon Cutaneum. Biotechnol Biofuels 2011, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, P.; Mosrati, R.; Corroler, D. Medium Chain Length Polyhydroxyalkanoates Biosynthesis in Pseudomonas Putida Mt-2 Is Enhanced by Co-Metabolism of Glycerol/Octanoate or Fatty Acids Mixtures. Int J Biol Macromol 2017, 98, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Sass, P. (ed. ) Antibiotics: Methods and Protocols, Methods in Molecular Biology 2nd Edition; New York, 2022; Antibiotics.
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bosdriesz, E.; Wortel, M.T.; Haanstra, J.R.; Wagner, M.J.; de la Torre Cortés, P.; Teusink, B. Low Affinity Uniporter Carrier Proteins Can Increase Net Substrate Uptake Rate by Reducing Efflux. Sci Rep 2018, 8, 5576. [Google Scholar] [CrossRef] [PubMed]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of Temperature and Nitrogen Concentration on the Growth and Lipid Content of Nannochloropsis Oculata and Chlorella Vulgaris for Biodiesel Production. Chemical Engineering and Processing: Process Intensification 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Schilling, O. A Protein-Dependent Riboswitch Controlling PtsGHI Operon Expression in Bacillus Subtilis: RNA Structure Rather than Sequence Provides Interaction Specificity. Nucleic Acids Res 2004, 32, 2853–2864. [Google Scholar] [CrossRef]
- Trakunjae, C.; Boondaeng, A.; Apiwatanapiwat, W.; Janchai, P.; Neoh, S.Z.; Sudesh, K.; Vaithanomsat, P. Statistical Optimization of P(3HB-Co-3HHx) Copolymers Production by Cupriavidus Necator PHB−4/PBBR_CnPro-PhaCRp and Its Properties Characterization. Sci Rep 2023, 13, 9005. [Google Scholar] [CrossRef] [PubMed]
- Lindenkamp, N.; Volodina, E.; Steinbüchel, A. Genetically Modified Strains of Ralstonia Eutropha H16 with β-Ketothiolase Gene Deletions for Production of Copolyesters with Defined 3-Hydroxyvaleric Acid Contents. Appl Environ Microbiol 2012, 78, 5375–5383. [Google Scholar] [CrossRef]
- Iram, S.H.; Cronan, J.E. The β-Oxidation Systems of Escherichia Coli and Salmonella Enterica Are Not Functionally Equivalent. J Bacteriol 2006, 188, 599–608. [Google Scholar] [CrossRef]
- Schroda, M.; deVitry, C. Molecular Chaperones, Proteases, and Unfolded Protein Responses. In The Chlamydomonas Sourcebook; Elsevier, 2023; pp. 647–689.
- Russell, A.D. Lethal Effects of Heat on Bacterial Physiology and Structure. Sci Prog 2003, 86, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, H.M.; Steinbüchel, A. Physiology, Biochemistry, and Molecular Biology of Triacylglycerol Accumulation by Rhodococcus. In; 2010; pp. 263–290.
- Yang, Y.; Jalalah, M.; Alsareii, S.A.; Harraz, F.A.; Thakur, N.; Zheng, Y.; Alalawy, A.I.; Koutb, M.; Salama, E.-S. Potential of Oleaginous Microbes for Lipid Accumulation and Renewable Energy Generation. World J Microbiol Biotechnol 2024, 40, 337. [Google Scholar] [CrossRef] [PubMed]
- Koreti, D.; Kosre, A.; Jadhav, S.K.; Chandrawanshi, N.K. A Comprehensive Review on Oleaginous Bacteria: An Alternative Source for Biodiesel Production. Bioresour Bioprocess 2022, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Antonopoulou, I.; Enman, J.; Rova, U.; Christakopoulos, P.; Matsakas, L. Lipids Detection and Quantification in Oleaginous Microorganisms: An Overview of the Current State of the Art. BMC Chemical Engineering 2019, 1, 13. [Google Scholar] [CrossRef]
- Gorte, O.; Kugel, M.; Ochsenreither, K. Optimization of Carbon Source Efficiency for Lipid Production with the Oleaginous Yeast Saitozyma Podzolica DSM 27192 Applying Automated Continuous Feeding. Biotechnol Biofuels 2020, 13, 181. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Ham, S.; Jeong, J.; Ku, H.; Kim, H.; Lee, C. Temperature Matters: Bacterial Response to Temperature Change. Journal of Microbiology 2023, 61, 343–357. [Google Scholar] [CrossRef]
- Zhu, L.; Cheng, J.; Luo, B.; Feng, S.; Lin, J.; Wang, S.; Cronan, J.E.; Wang, H. Functions of the Clostridium Acetobutylicium FabF and FabZ Proteins in Unsaturated Fatty Acid Biosynthesis. BMC Microbiol 2009, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Tanaka, A.; Kawamoto, J.; Kurihara, T. Purification and Characterization of 1-Acyl-Sn-Glycerol-3-Phosphate Acyltransferase with a Substrate Preference for Polyunsaturated Fatty Acyl Donors from the Eicosapentaenoic Acid-Producing Bacterium Shewanella Livingstonensis Ac10. The Journal of Biochemistry 2018, 164, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Gao, Z.; Shi, T.-Q.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. Reactive Oxygen Species-Mediated Cellular Stress Response and Lipid Accumulation in Oleaginous Microorganisms: The State of the Art and Future Perspectives. Front Microbiol 2017, 8. [Google Scholar] [CrossRef]
- Paulsen, I.T. Carbon Metabolism and Its Regulation in Streptomyces and Other High GC Gram-Positive Bacteria. Res Microbiol 1996, 147, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Bucca, G.; Pothi, R.; Hesketh, A.; Möller-Levet, C.; Hodgson, D.A.; Laing, E.E.; Stewart, G.R.; Smith, C.P. Translational Control Plays an Important Role in the Adaptive Heat-Shock Response of Streptomyces Coelicolor. Nucleic Acids Res 2018, 46, 5692–5703. [Google Scholar] [CrossRef]
- Narberhaus, F.; Waldminghaus, T.; Chowdhury, S. RNA Thermometers. FEMS Microbiol Rev 2006, 30, 3–16. [Google Scholar] [CrossRef]
- Kalscheuer, R.; Steinbüchel, A. A Novel Bifunctional Wax Ester Synthase/Acyl-CoA:Diacylglycerol Acyltransferase Mediates Wax Ester and Triacylglycerol Biosynthesis InAcinetobacter Calcoaceticus ADP1. Journal of Biological Chemistry 2003, 278, 8075–8082. [Google Scholar] [CrossRef] [PubMed]
- Wältermann, M.; Stöveken, T.; Steinbüchel, A. Key Enzymes for Biosynthesis of Neutral Lipid Storage Compounds in Prokaryotes: Properties, Function and Occurrence of Wax Ester Synthases/Acyl-CoA:Diacylglycerol Acyltransferases. Biochimie 2007, 89, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.A.; Morais, M.A.B.; Terrett, O.M.; Lyczakowski, J.J.; Zanphorlin, L.M.; Ferreira-Filho, J.A.; Tonoli, C.C.C.; Murakami, M.T.; Dupree, P.; Souza, A.P. An Engineered GH1 β-Glucosidase Displays Enhanced Glucose Tolerance and Increased Sugar Release from Lignocellulosic Materials. Sci Rep 2019, 9, 4903. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Li, S.; Bello, A.; Liu, J.; Gao, L.; Fan, Z.; Wang, S.; Liu, L.; Ma, B.; et al. Mechanism of Differential Expression of β-Glucosidase Genes in Functional Microbial Communities in Response to Carbon Catabolite Repression. Biotechnology for Biofuels and Bioproducts 2022, 15, 3. [Google Scholar] [CrossRef]
Figure 1.
Rewiring metabolic pathway for lipid production in S. jeddahensis.
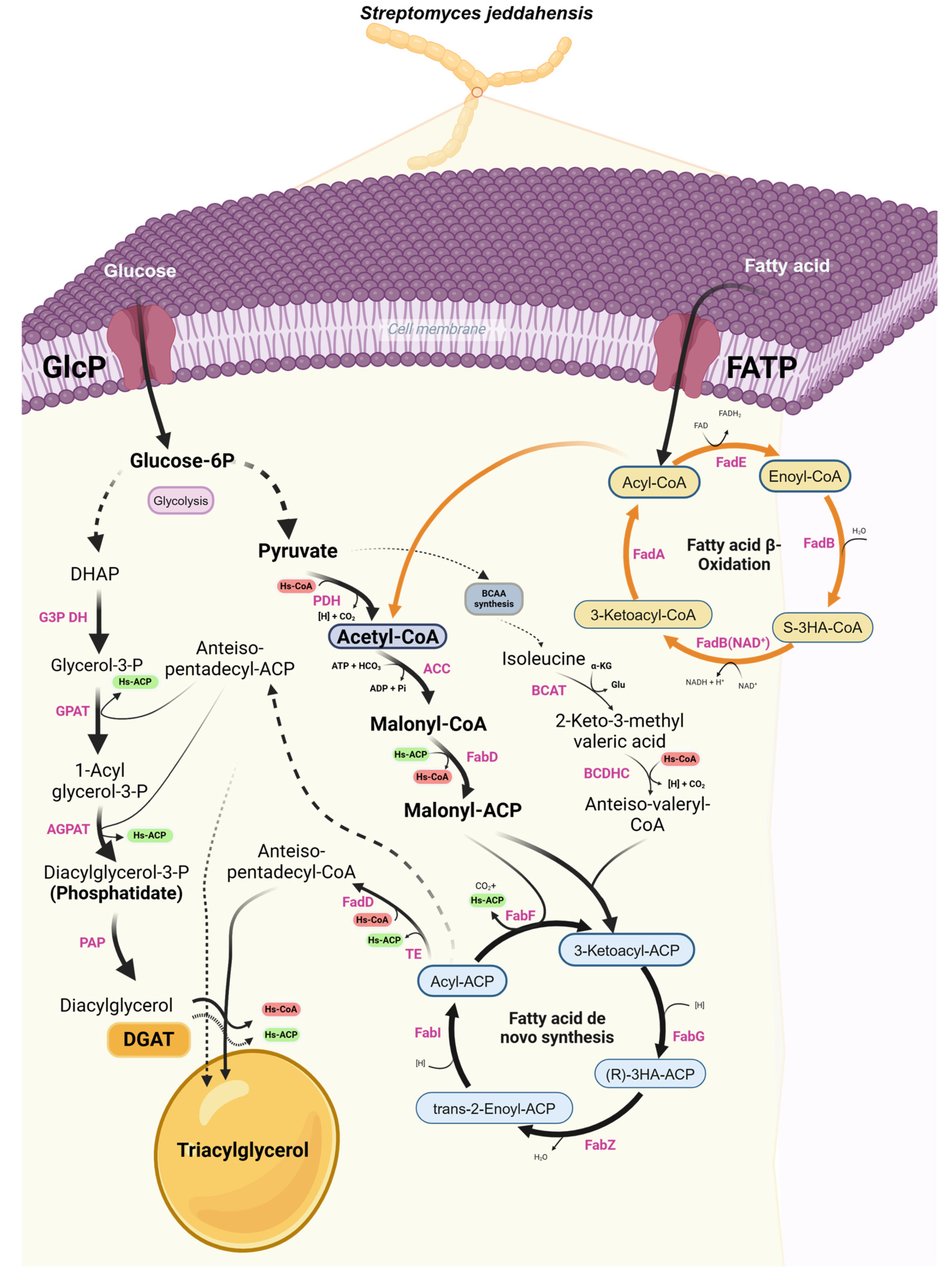
Figure 2.
Fermentations of S. jeddahensis strains, measured from 1 mL samples, and standard deviations were reported. The strain was cultivated for 48 h, with triplicate samples taken. (a) CDW was strain culture into medium MSM containing 2.5 g/L glucose. (b) Glucose consumption. (c) CDW was strains culture into medium MSM containing 2.5 g/L sodium hexanoate (d) sodium hexanoate consumption. (e) CDW was strains culture into medium MSM containing 2.5 g/L sodium octanoate. (f) sodium octanoate consumption. (g) CDW was strains culture into medium MSM containing 2.5 g/L sodium decanoate. (h) sodium decanoate consumption.
Figure 2.
Fermentations of S. jeddahensis strains, measured from 1 mL samples, and standard deviations were reported. The strain was cultivated for 48 h, with triplicate samples taken. (a) CDW was strain culture into medium MSM containing 2.5 g/L glucose. (b) Glucose consumption. (c) CDW was strains culture into medium MSM containing 2.5 g/L sodium hexanoate (d) sodium hexanoate consumption. (e) CDW was strains culture into medium MSM containing 2.5 g/L sodium octanoate. (f) sodium octanoate consumption. (g) CDW was strains culture into medium MSM containing 2.5 g/L sodium decanoate. (h) sodium decanoate consumption.
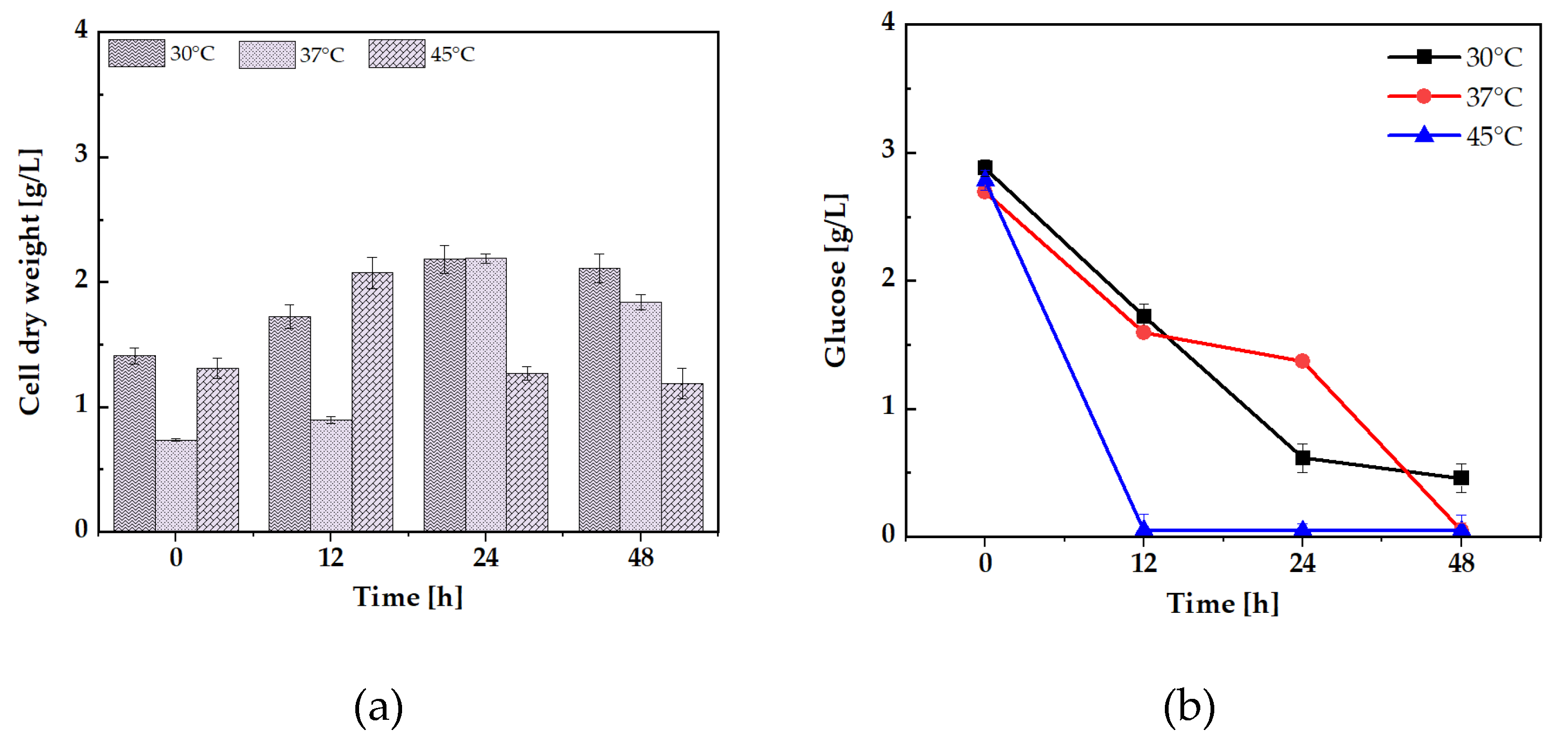
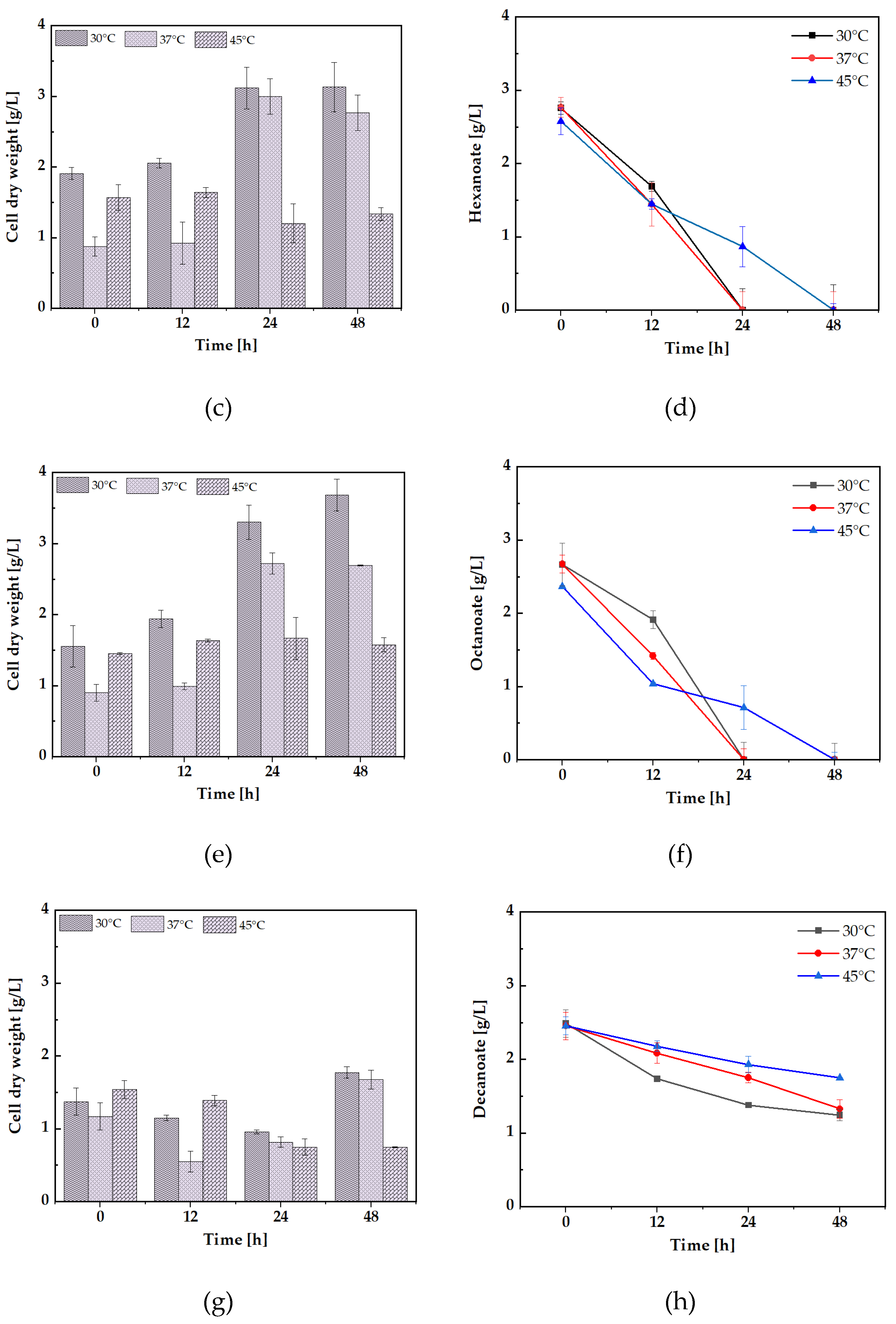
Figure 3.
Observation of lipid accumulation in S. jeddahensis., culture process on 37°C. The panel displays images of both strains as observed through Bodipy staining, phase-contrast microscopy (Brightfield), and merged (bars, 10µm). (A) strains culture into medium MSM containing 2.5 g/L glucose, (B) strains culture into medium MSM containing 2.5 g/L sodium hexanoate, (C) strains culture into medium MSM containing 2.5 g/L sodium octanoate, (D) strains culture into medium MSM containing 2.5 g/L sodium decanoate.
Figure 3.
Observation of lipid accumulation in S. jeddahensis., culture process on 37°C. The panel displays images of both strains as observed through Bodipy staining, phase-contrast microscopy (Brightfield), and merged (bars, 10µm). (A) strains culture into medium MSM containing 2.5 g/L glucose, (B) strains culture into medium MSM containing 2.5 g/L sodium hexanoate, (C) strains culture into medium MSM containing 2.5 g/L sodium octanoate, (D) strains culture into medium MSM containing 2.5 g/L sodium decanoate.
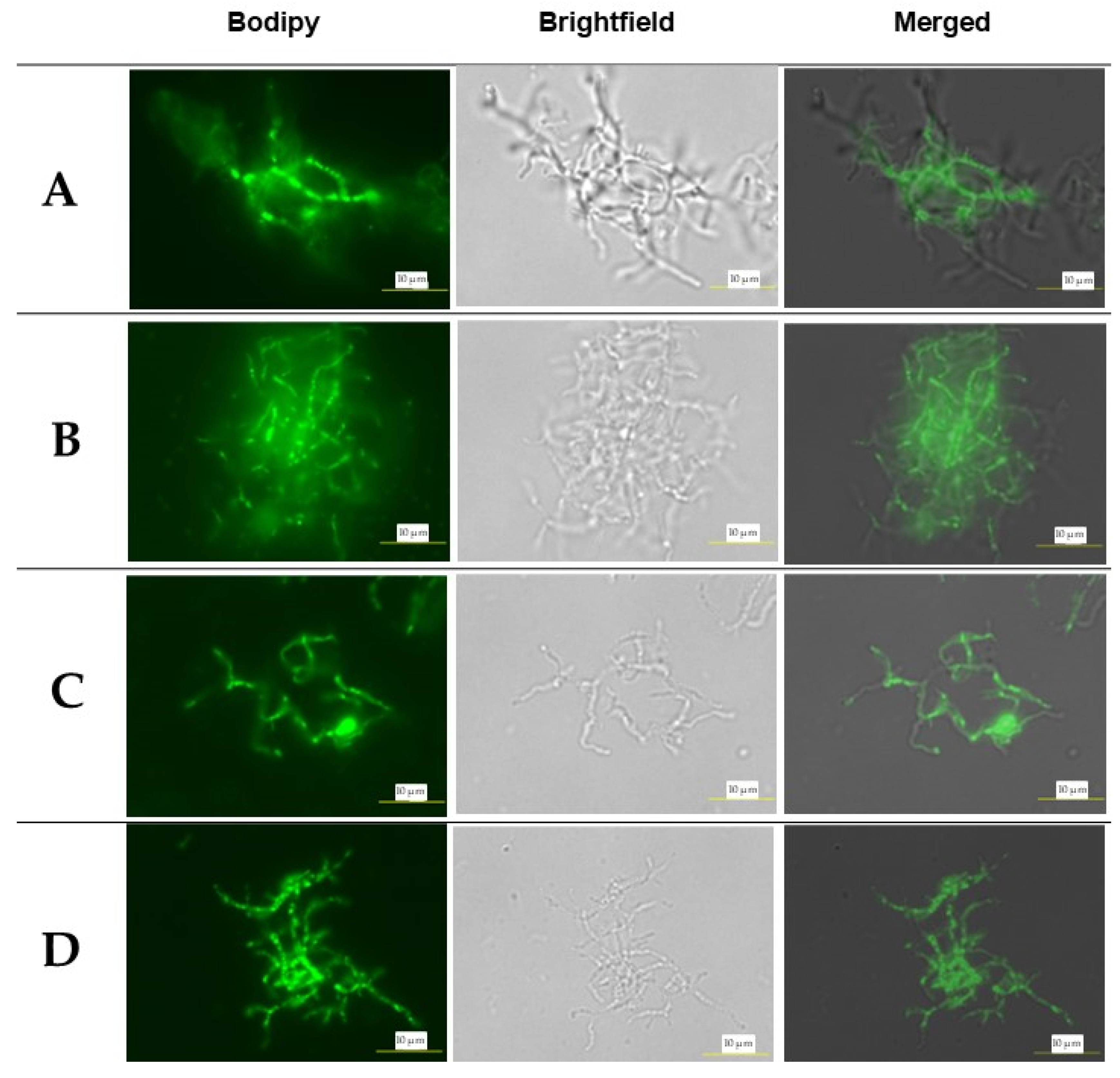
Figure 4.
The bioconversion of various carbon sources into lipids was evaluated by cultivating S. jeddahensis for 48 hours in MSM medium supplemented with different carbon sources at 30°C, 37°C, and 45°C. The lipid content (%, w/w) is represented as a bar, showing the average value and standard deviation (n = 3).
Figure 4.
The bioconversion of various carbon sources into lipids was evaluated by cultivating S. jeddahensis for 48 hours in MSM medium supplemented with different carbon sources at 30°C, 37°C, and 45°C. The lipid content (%, w/w) is represented as a bar, showing the average value and standard deviation (n = 3).
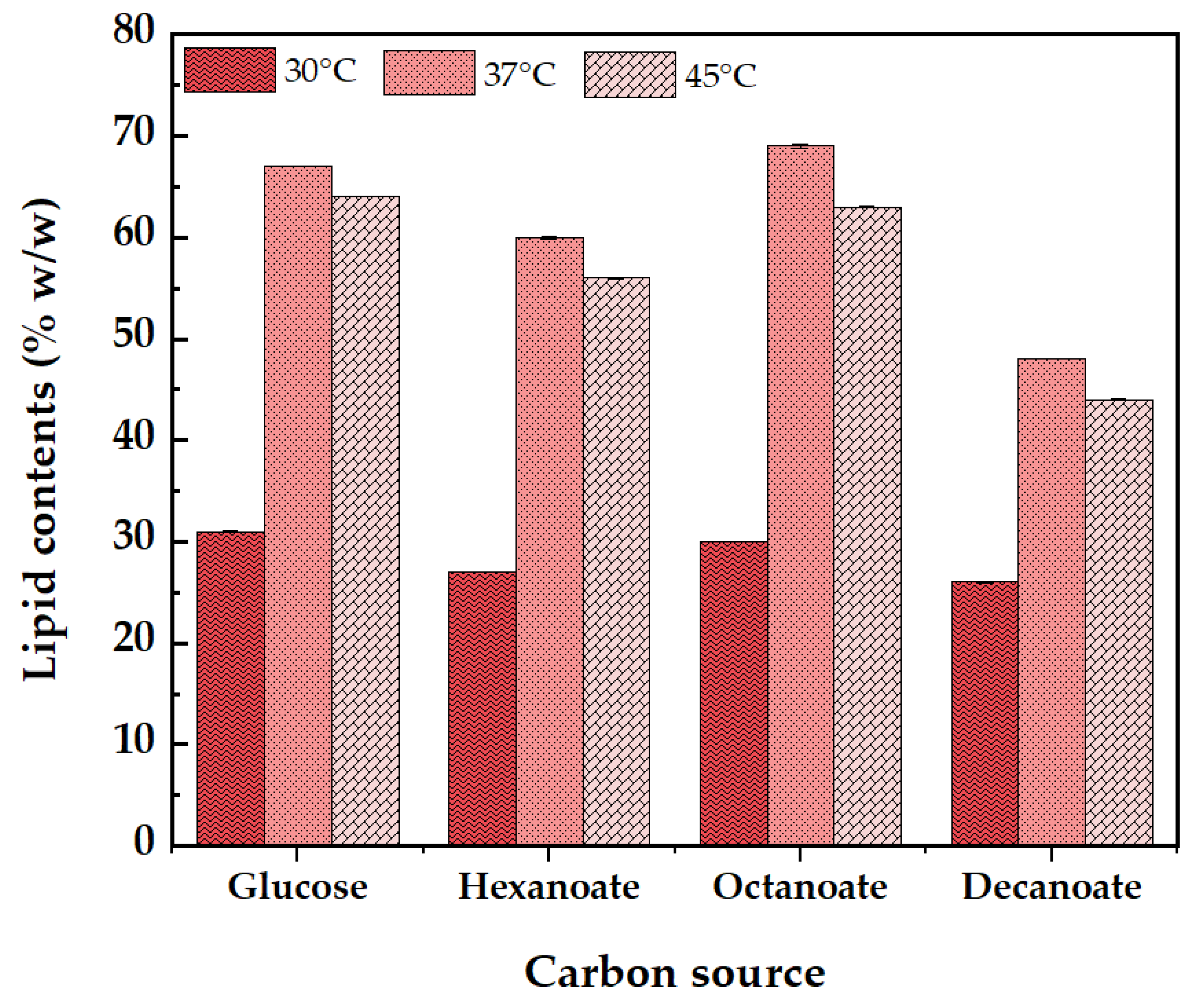
Figure 5.
Total fatty acid (TFA) contents of S. jeddahensis strains cultivated in minimal salt medium (MSM) supplemented with different carbon sources (2.5 g/L glucose, sodium hexanoate, sodium octanoate, or sodium decanoate) and incubated at varying temperatures (30°C, 37°C, and 45°C) for 48 hours. Experimental conditions are as follows: MSM + glucose at (30°C, 37°C, 45°C), MSM + sodium hexanoate at (30°C, 37°C), MSM + sodium octanoate at (30°C, 37°C, 45°C), MSM + sodium decanoate at (30°C, 37°C, 45°C). Results highlight the impact of carbon source and temperature on Total FA production under defined growth conditions.
Figure 5.
Total fatty acid (TFA) contents of S. jeddahensis strains cultivated in minimal salt medium (MSM) supplemented with different carbon sources (2.5 g/L glucose, sodium hexanoate, sodium octanoate, or sodium decanoate) and incubated at varying temperatures (30°C, 37°C, and 45°C) for 48 hours. Experimental conditions are as follows: MSM + glucose at (30°C, 37°C, 45°C), MSM + sodium hexanoate at (30°C, 37°C), MSM + sodium octanoate at (30°C, 37°C, 45°C), MSM + sodium decanoate at (30°C, 37°C, 45°C). Results highlight the impact of carbon source and temperature on Total FA production under defined growth conditions.
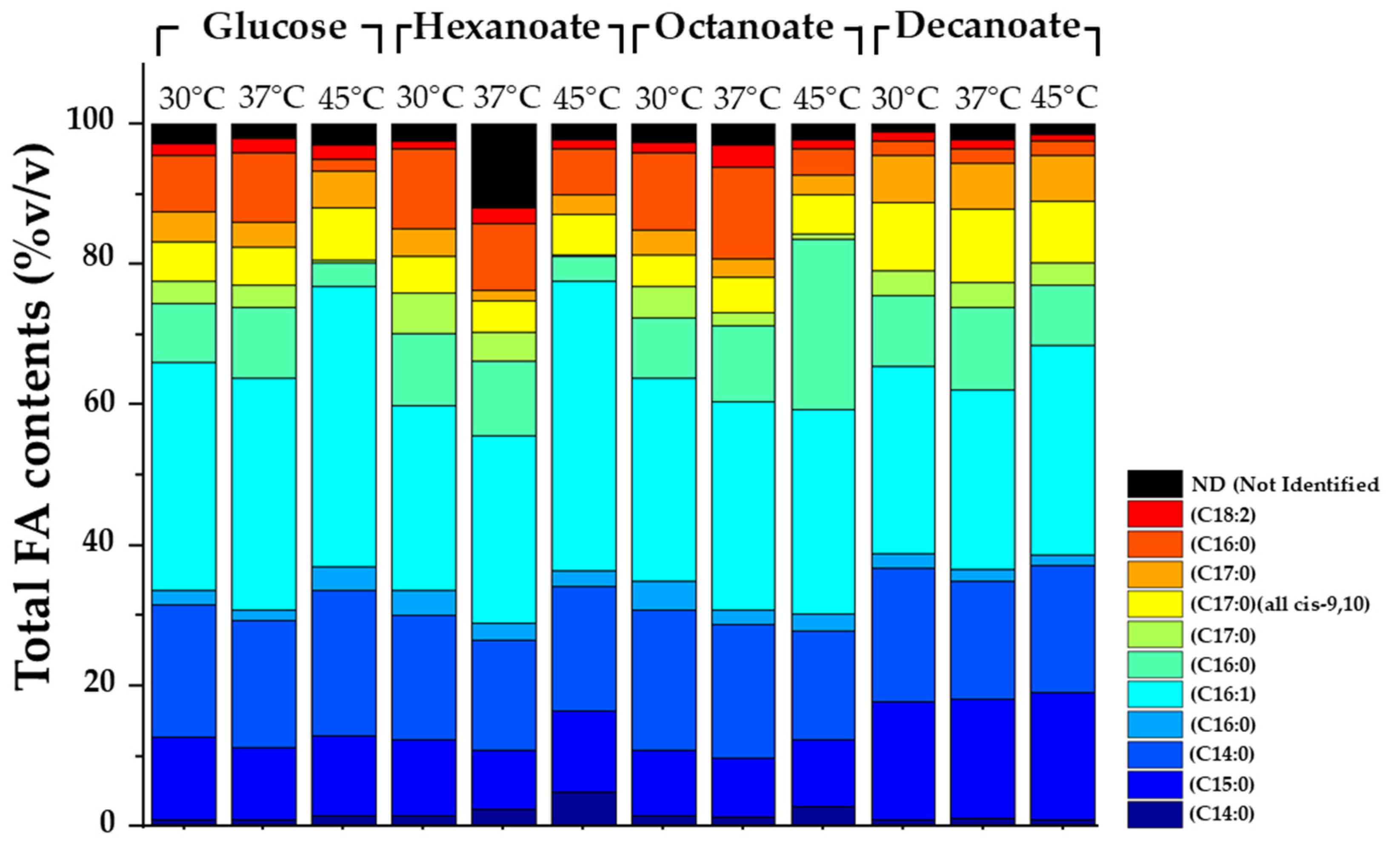

Table 1.
List of Primers Used for Measuring Gene Expression via Quantitative PCR.
| Gene | Forward primer (5’→3’) | Reverse primer (5’→3’) |
|---|---|---|
| STSP_00760 (plsC_1) | CCGTGAACCACTCCCACAA | ACCTTCAGCTGGCCGATTC |
| STSP_00850 | GCCAGTGCGTACCGAGATAG | CTCGCCTATATGGCCTCGAC |
| STSP_01000 (inhA_1) | CAGGTTGCAGCGGATGTTCT | CGCAGTACGACTGGATGGG |
| STSP_01010 (fabG1_1) | CTCCAGTGATGTACGAGGCG | TCGACACCGACATGACCAAG |
| STSP_03080 (DGAT) | GATCGACGGGGTCCATCAC | TTCGCCGTGCTGTTCAAGTT |
| STSP_12530 | TAGCCCCACTCGAAGTTGTC | CGGACCGTATCGCGTATCTG |
| STSP_17580 (bkdA_1) | GTTGACACCACGGAACATGC | GACGACTACGTCTTCCCGAC |
| STSP_19060 (gpsA) | GATGGTCTCCTCCAGGGTCA | CATGACCTTCTCCGGACTCG |
| STSP_28680 (accD5_1) | CATCAAGCAGCTCCTGTCGT | CTCGATGACCTCGTGCATGT |
| STSP_45110 (tesB_2) | GTACTTCGGCAGTGACTCGG | GCAGCCGATCTTCCATCTGT |
| STSP_45150 (accA1_2) | CGGAGAGCTATCTGTCGGTG | CTCGGCGAGGAATCCGTATC |
| STSP_50580 (FabZ) | CCGACCTCTTTGGCGAACTT | GCCCAGACCTTCAACGTGAC |
| STSP_61390 (ilvE) | TGACGATCAAGTGGACGGAG | CGAAGATCTCCTGGGCGTAG |
| STSP_67970 (fabD) | GCGTACGTCTCGAACAAGGA | TCTGGAACGTCTCCATGCAC |
| STSP_67980 (fabH_2) | AGCTGATCAAGCAGACCGTG | GGAACTTGATGTCGCCGTTG |
| STSP_68010 (fabF) | CGATCGAGATGATCCGCACC | TGTTCTTGGACATCGCCATCA |
| STSP_72400 (lcfB-5) | CACGAGACGGTCTACATCGG | ATCGTCACAGCTCAACTCGG |
| STSP_74190 | TACCTGGTCCGTCAGGTTGA | AAGTGGACGATCCAGTTCGG |
| rpoB1 | GTAGTTGTGACCCTCCCACG | GACTACATCACCACCGCCAA |
1 Housekeeping gen.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



