
Submitted:
19 December 2024
Posted:
20 December 2024
You are already at the latest version
Abstract
The present pilot study aimed to investigate whether common single nucleotide polymorphisms (SNPs) in the gene encoding glutathione S-transferase omega 1 (GSTO1), both individually and in combination with variants of the catalytic subunit of glutamate cysteine ligase (GCLC) gene and environmental risk factors, are associated with the risk of psoriasis. The research included a total of 944 participants, comprising 474 individuals diagnosed with psoriasis and 470 healthy control subjects. Five common SNPs in the GSTO1 gene—specifically, rs11191736, rs34040810, rs2289964, rs11191979, and rs187304410—were genotyped in the study groups using the MassARRAY-4 system. The allele rs187304410-A (OR=0.19, 95% CI 0.04-0.86, Pperm=0.02) and the genotype rs187304410-G/A (OR=0.19, 95% CI 0.04-0.85, Pperm=0.01) were found to be associated with psoriasis in females. The model-based multifactor dimensionality reduction approach facilitated the identification of higher-order epistatic interactions between the variants of the GSTO1 and GCLC genes (Pperm<0.0001). These interactions, along with the risk factor of alcohol abuse, collectively contribute to the pathogenesis of psoriasis. This study is the first to demonstrate that polymorphisms in the GSTO1 gene, both individually and in combination with variants of the GCLC gene and alcohol abuse, are associated with an increased risk of psoriasis.
Keywords:
Psoriasis
; genetic susceptibility
; oxidative stress
; glutathione metabolism
; glutathione S-transferase omega 1
; GSTO1
; single nucleotide polymorphism
; alcohol abuse
; gene–gene interactions
; gene–environment interactions
1. Introduction
Psoriasis is a chronic dermatosis characterized by immune-mediated inflammation, resulting in the formation of thickened, scaly erythematous plaques [1]. The global prevalence of psoriasis is approximately 2%, with psoriasis vulgaris, also known as plaque-type psoriasis, representing the most common subtype and accounting for around 90% of all cases [2,3]. Psoriasis is commonly understood as a complex multifactorial disorder resulting from the interplay between genetic and environmental factors [4]. In recent decades, substantial progress has been achieved in elucidating the genetic mechanisms underlying psoriasis, primarily facilitated by genome-wide association studies that have identified more than sixty regions associated with disease susceptibility [4]. These investigations have highlighted the pathogenic significance of genes involved in the activation of Th17 cells and have also uncovered rare genetic variants associated with distinct forms of psoriasis [4,5]. Numerous extrinsic factors—such as mechanical stress, air pollutants, sun exposure, medications, vaccinations, infections, and lifestyle choices—and intrinsic factors—including obesity, diabetes mellitus, dyslipidemia, hypertension, and mental stress—have been identified as triggers and exacerbating factors for psoriasis [6]. Research conducted on extensive twin cohorts indicates that genetic factors account for 60-75% of the variability in susceptibility to psoriasis, while the remaining variation is attributed to nonshared environmental influences [7]. Despite the well-established role of environmental factors in the pathogenesis of psoriasis [8], exploring the mechanisms by which these factors disrupt the body’s equilibrium and contribute to the onset and progression of the disease remains a significant challenge.
In recent years, a growing body of research has emerged that underscores the significant role of chemical exposure, particularly air pollutants, in the etiology of psoriasis. Numerous air pollutants, including polycyclic aromatic hydrocarbons, volatile organic compounds, oxides, particulate matter, ozone, heavy metals, and ultraviolet radiation, have been shown to adversely affect the skin and increase the risk of psoriasis [9,10]. A recent large prospective cohort study involving 474,055 participants demonstrated that long-term exposure to air pollution—measured in terms of nitrogen dioxide, nitrogen oxides, fine particulate matter with a diameter of less than 2.5 µm, and particulate matter with a diameter of less than 10 µm, is associated with an increased risk of psoriasis [11]. Additionally, this study found an interaction between air pollution and genetic susceptibility that jointly contributes to the risk of psoriasis [11]. Furthermore, environmental exposure to toxic metals such as barium, cesium, antimony, uranium, lead, mercury, and cadmium has been found to compromise immunity and increase inflammation, thereby predisposing individuals to the development and exacerbation of psoriasis [12,13,14,15,16]. In order to mitigate the harmful effects of chemical pollutants, the skin expresses enzymes that are involved in xenobiotic biotransformation, with glutathione S-transferases (GST) playing a particularly significant role [17,18]. The primary biological function of glutathione S-transferases is to facilitate the detoxification of chemicals by catalyzing their conjugation with reduced glutathione [19]. Numerous studies have demonstrated an increased expression of glutathione S-transferases in affected skin regions compared to unaffected tissues in patients with psoriasis [20,21,22]. This observation suggests, first, the critical involvement of these enzymes in the pathogenesis of the disease, and second, that the elevated levels of GSTs in the affected skin may serve a protective function by neutralizing toxic substances that accumulate in the dermal layers. Since the genetic variability of glutathione S-transferases can influence the activity and expression of these enzymes, thereby determining individual characteristics related to skin protection against chemical exposures, polymorphisms in the GST genes have become a focal point for genetic studies on psoriasis in recent years. The vast majority of studies on psoriasis have concentrated on the deletion polymorphisms of the glutathione S-transferase genes GSTM1 and GSTT1 [23,24,25,26,27,28,29,30,31]. These studies have demonstrated associations with susceptibility to psoriasis, its clinical features, and the efficacy of various therapeutic approaches for the condition. The present pilot study aimed to investigate whether common single nucleotide polymorphisms (SNPs) in the gene encoding glutathione S-transferase omega 1 (GSTO1), both individually and in combination with variants of the catalytic subunit of glutamate cysteine ligase (GCLC) gene and environmental risk factors, are associated with the risk of psoriasis.
2. Materials and Methods
2.1. Study Patients
All phases of data collection and analysis were conducted in accordance with the principles outlined in the Declaration of Helsinki. Informed written consent was obtained from human participants in compliance with protocols approved by the Ethics Review Committee of Kursk State Medical University. The study involved a cohort of 944 unrelated individuals of European ancestry, primarily consisting of Russians from the Kursk region. This group included 474 patients diagnosed with psoriasis and 470 healthy control subjects. Patient enrollment for the study involving individuals with psoriasis occurred at Medvenka Central District Hospital in the Kursk region, the Center for Medical Examinations and Prevention in Kursk, and the Kursk Regional Multidisciplinary Clinical Hospital, covering the period from September 2018 to December 2021. The control group, comprising subjects without chronic diseases, was drawn from our previous research efforts [32,33,34].
2.2. Diagnosis of Psoriasis
The diagnosis of psoriasis was confirmed by certified dermatologists through the assessment of the characteristic clinical presentation of skin lesions and their specific locations [35]. The research included individuals diagnosed with various forms of psoriasis, such as classic plaque psoriasis, palmoplantar psoriasis, seborrheic psoriasis, scalp psoriasis, the von Zumbusch variant of generalized pustular psoriasis, inverse psoriasis, guttate psoriasis, and erythrodermic psoriasis. Additionally, the study examined patients with comorbid conditions associated with psoriasis, including psoriatic arthritis and onychodystrophy. The participants in the study were not diagnosed with chronic infectious diseases, such as HIV or hepatitis, nor did they exhibit severe chronic conditions that predated the onset of psoriasis. Additionally, individuals undergoing biologic therapy at the time of recruitment, as well as pregnant women, were excluded from the study. The Psoriasis Area and Severity Index (PASI) was utilized as a clinical tool for assessing the severity of psoriasis progression.
2.3. Interviewing of Patients
Participants in the study completed a validated questionnaire administered by a physician [36] to evaluate risk factors associated with psoriasis, including cigarette smoking, alcohol consumption, and reduced consumption of fresh fruits and vegetables [37]. Interest in these risk factors stems from their detrimental effect on the levels of endogenous glutathione reserves, which are crucial for the detoxification of xenobiotics via the action of glutathione S-transferase enzymes [38]. The alcohol consumption, cigarette smoking and dietary habits of the study participants were evaluated as previously described [39,40].
2.4. SNP Selection
Single nucleotide polymorphisms of the GSTO1 gene were selected for this study based on the following criteria: 1) a minor allele frequency of ≥ 1% in the European population (Ensembl data, www.ensembl.org); 2) the presence of a SNP with GSTO1 expression quantitative trait loci (eQTL) and/or splicing quantitative trait loci (sQTL) in the skin, specifically in sun-exposed areas (lower leg) and non-sun-exposed areas (suprapubic), according to data from the GTEx portal (https://gtexportal.org); 3) linkage disequilibrium (r² < 0.8) among the SNPs of interest (HapMap data for the European population); and 4) the feasibility of joint SNP detection in a multiplex panel using the MALDI-TOF mass spectrometry method. As a result, five common SNPs of the GSTO1 gene were selected for genotyping: rs11191736, rs34040810, rs2289964, rs11191979 (tagSNP), and rs187304410.
2.5. Genetic Analysis
All genetic investigations were performed at the Research Institute for Genetic and Molecular Epidemiology, situated at Kursk State Medical University in Kursk, Russia. Venous blood samples were obtained from the cubital vein of the study participants and collected in EDTA-coated tubes. These samples were promptly frozen and stored at −20 °C until they were processed. Genomic DNA was isolated utilizing the conventional methods of phenol/chloroform extraction followed by ethanol precipitation. Genotyping of the selected polymorphisms was conducted using the MALDI-TOF mass spectrometry iPLEX platform on the MassArray-4 system (Agena Bioscience, Inc., San Diego, CA, USA). The methodologies for sample preparation and single nucleotide polymorphism (SNP) genotyping using the MassArray-4 system are thoroughly detailed in our latest publication [41]. The primer sequences utilized for genotyping can be provided upon request. In order to ensure quality control, 5% of the DNA samples were subjected to duplicate genotyping, with the researchers remaining unaware of the case-control status. The concordance rate of the control genotyping was greater than 99%.
2.6. Statistical and Bioinformatics Analysis
The estimation of statistical power for the association analysis was conducted using the GAS power calculator, as documented in previous research [39]. Fisher’s exact test was employed to evaluate the distribution of genotype frequencies in relation to the Hardy-Weinberg equilibrium (HWE). The frequencies of alleles and genotypes within the study cohorts, along with their correlations to the risk of developing psoriasis, were analyzed using PLINK software version 1.9 [42]. Logistic regression analysis was employed to investigate the relationships between polymorphisms in the GSTO1 gene and the risk of psoriasis. The unadjusted odds ratios (OR) and 95% confidence intervals (95% CI) were calculated to assess the associations between single nucleotide polymorphisms (SNPs) and the corresponding phenotypes. In the context of SNP-disease associations, various genetic models—including allelic, recessive, dominant, and log-additive models—were assessed. Additionally, haplotype analysis of the GSTO1 gene was conducted using Haploview software, version 4.2. P-values (Pperm) for associations involving alleles, genotypes, and haplotypes were calculated using adaptive permutation methods with PLINK and Haploview software. The model with the lowest Pperm was considered the best for SNP-disease association. The relationships between pairwise genotype combinations (diplotypes) and the risk of developing psoriasis were evaluated using the chi-squared test. Gene-gene (SNP-SNP) and gene-environment interactions were analyzed using a model-based multifactor dimensionality reduction method (mbmdr) [43]. The replication of associations between polymorphisms in the GSTO1 gene and psoriasis was conducted using the Gene ATLAS database from the UK Biobank (http://geneatlas.roslin.ed.ac.uk, accessed on September 10, 2024).
3. Results
3.1. Association Between GSTO1 Gene Polymorphisms and the Risk of Psoriasis
The genotype frequencies for all single nucleotide polymorphisms of GSTO1 were found to be in HWE in both the case and control groups (P>0.05). We conducted an analysis of the relationships between polymorphisms in the GSTO1 gene and the risk of developing psoriasis, examining both the overall population and subgroups stratified by sex. A summary of the associations between GSTO1 gene polymorphisms and psoriasis risk for both the entire cohort and the sex-stratified groups is presented in Table 1. No statistically significant associations were found between SNPs of the GSTO1 gene and the risk of developing psoriasis when comparing the overall case and control groups. The sex-stratified analysis presented in Table 1 indicates that two specific polymorphisms are significantly associated with the risk of psoriasis: rs11191736 (allelic model) in males and rs187304410 (dominant model) in females.
Table 2 presents the genotype and allele frequencies of the GSTO1 gene, along with their associations to the risk of psoriasis, analyzed for both the entire group and stratified by sex. In the male groups, allele rs11191736-T was found only among patients with psoriasis (Pperm=0.017). Allele rs187304410-A (OR=0.19 95% CI 0.04-0.86, Pperm=0.02) and genotype rs187304410-G/A (OR=0.19 95% CI 0.04-0.85, Pperm = 0.01) were found to confer protection against the risk of psoriasis in females.
Subsequently, we conducted a replication analysis to investigate the associations between polymorphisms of the GSTO1 gene and susceptibility to psoriasis in the large cohort from the UK Biobank. None of the polymorphisms we studied demonstrated a statistically significant association with the risk of developing psoriasis in this cohort.
The difficulty in replicating associations between SNPs and diseases may be partially attributed to genetic variations across different populations, as we recently demonstrated [44,45]. This observation prompted us to conduct an association analysis of psoriasis in relation to all SNPs of the GSTO1 gene that were genotyped within the UK Biobank cohort. As a result, 40 SNPs of the GSTO1 gene within the UK Biobank cohorts have been identified as being associated with an increased risk of psoriasis, with a P-value of ≤ 0.05, as presented in Supplementary Table 1.
3.2. The Combined Impact of GSTO1 Gene Polymorphisms on Psoriasis Risk
The combined influence of polymorphisms in the GSTO1 gene on the risk of developing psoriasis was assessed using haplotype and diplotype analyses. Table 3 presents the GSTO1 haplotypes and their correlation with the risk of psoriasis, analyzed for both the overall population and stratified by sex. The study identified four prevalent haplotypes of GSTO1, designated as H1–H4, which exhibited a frequency exceeding 2% within the study populations. As shown in Table 3, none of the GSTO1 haplotypes were found to be associated with the risk of psoriasis (Pperm>0.05). SNP rs11191736 exhibited a positive linkage disequilibrium (P<0.001) with the polymorphisms rs34040810 (D’=0.497), rs2289964 (D’=0.441), and rs187304410 (D’ = 0.426). Polymorphism rs34040810 was associated with rs2289964 (D’=0.992) and rs187304410 (D’=0.605). Additionally, rs187304410 showed a correlation with rs2289964 (D’=0.358).
Supplementary Table 2 presents the genotype combinations (diplotypes) of GSTO1 and their associations with the risk of psoriasis. Two diplotypes, rs11191736C/C × rs34040810C/C (OR=0.21, 95% CI 0.04-0.95, P=0.03) and rs2289964C/T × rs11191979T/T (OR=2.50, 95% CI 1.06-5.92, P=0.05), along with three diplotypes—rs11191736C/C × rs187304410G/G (OR=5.99, 95% CI 1.34-26.89, P=0.01), rs34040810C/C × rs187304410G/G (OR=3.91, 95% CI 1.09-14.05, P=0.02), and rs34040810C/C × rs187304410G/A (OR=0.24, 95% CI 0.06-0.98, P=0.05)—were found to be associated with the risk of psoriasis risk in males and females, respectively. Nevertheless, none of these associations remained significant after applying the Benjamini–Hochberg procedure for multiple testing corrections.
Considering that glutathione S-transferases utilize glutathione for the detoxification of xenobiotics, it is essential to investigate the potential interactive effects of polymorphisms in the GSTO1 gene and the gene encoding the catalytic subunit of glutamate cysteine ligase (GCLC) on susceptibility to psoriasis. Genotyping data for six common polymorphisms of the GCLC gene in psoriasis patients and controls were obtained from our previous study [39]. As shown in Supplementary Table 3, diplotypes GSTO1 rs34040810C/C × GCLC rs648595G/G (OR=0.70 95% CI 0.50-0.99, P=0.04), GSTO1 rs11191979T/T × GCLC rs542914C/C (OR=1.44 1.02-2.02, P=0.04), GSTO1 rs11191979T/C × GCLC rs542914C/C (OR=0.69 95% CI 0.48-0.99, P=0.04), and GSTO1 rs11191979T/C × GCLC rs648595G/G (OR=0.55 95% CI 0.33-0.94, P=0.03) were associated with the risk of psoriasis. These associations also did not survive after adjusting for multiple testing.
3.3. The Role of Gene-Gene and Gene-Environment Interactions in the Risk of Psoriasis
The comprehensive assessment of gene-by-gene interactions poses a challenge commonly known as the “curse of dimensionality.” This phenomenon arises in the context of epistatic analysis, where the number of SNPs increases exponentially, significantly limiting the effectiveness of traditional parametric statistical methods [46,47,48]. In order to address this challenge, we employed the model-based multifactor dimensionality reduction approach, as outlined by Calle with colleagues [43], to assess the interactions between genes (G×G) and between genes and environmental factors (G×E) that are associated with susceptibility to psoriasis. The mbmdr methodology facilitates the simultaneous evaluation of multiple variables, including gene polymorphisms and environmental factors, by reducing the dimensionality of the calculated parameters. This reduction improves the detection of non-linear or non-additive interactions among the variables being studied [43]. Among the environmental risk factors contributing to psoriasis, particular attention has been directed toward those that adversely affect endogenous levels of glutathione. These factors include excessive alcohol consumption, tobacco use, and a dietary deficiency of fresh fruits and vegetables [39]. Two-, three-, and four-order G×G and G×E models, which include combinations of SNPs at the GSTO1 and GCLC genes, along with the aforementioned risk factors, were analyzed to identify associations with the risk of psoriasis. Table 4 summarizes the number of mbmdr models that demonstrate significant associations between SNP × risk factor interactions and the risk of developing psoriasis. A total of 12 two-order, 68 three-order, and 239 four-level statistically significant (Pperm<0.05) mbmdr models associated with the risk of psoriasis were established for the entire group. The primary finding indicated that the vast majority of models linking the risk of psoriasis were established due to gene-environment interactions, with alcohol abuse emerging as a significant risk factor. Notably, the highest number of gene-environment interactions was identified between alcohol abuse and the polymorphisms of the GSTO1 and GCLC genes. Furthermore, many complex (three- and four order) mbmdr models comprised cigarette smoking along with the interactions of gene polymorphisms and alcohol abuse. Distinct characteristics of gene-environment interactions in males and females have been identified. Notably, among the top five n-order mbmdr models, the risk factor of smoking was associated with 11 G×E interactions in females, while only one gene-smoking interaction (ALCOHOL × SMOKE × GCLC rs542914 × GSTO1 rs11191979) was identified in males.
As illustrated in Table 4, statistically significant differences between sexes were observed in both three-order and four-order models. In particular, the number of models involving ALCOHOL × GCLC (1 SNP) × GSTO1 (1 SNP) was twice as high in males compared to females. The ALCOHOL × GSTO1 (2 SNPs) × GCLC (1 SNP) model accounted for nearly 36% of the four-order models, with no corresponding model identified in females. In contrast, the number of ALCOHOL × SMOKE × GCLC (1 SNP) × GSTO1 (1 SNP) and ALCOHOL × SMOKE × GSTO1 (2 SNPs) mbmdr models was higher in females than in males. The top five n-order mbmdr G×E models associated with the risk of psoriasis in both males and females are presented in Table 5 and Table 6, respectively. Polymorphisms of the GSTO1 gene were identified in 19 of the best mbmdr models for males and 14 models for females. Notably, the two-order mbmdr models for females are exclusively composed of polymorphisms in the GSTO1 gene. In contrast, the models for males are influenced by both gene-environment interactions, incorporating three SNPs of GSTO1 and two variants of GCLC. Additionally, SNPs of the GCLC gene were detected in 9 models for males and 5 models for females.
4. Discussion
Glutathione S-transferases (GSTs) play a crucial role in detoxifying harmful xenobiotics, inactivating endogenous compounds resulting from oxidative stress, and synthesizing essential biological molecules such as leukotrienes, prostaglandins, testosterone, and progesterone [19]. In the context of xenobiotic biotransformation, GSTs utilize the antioxidant glutathione to facilitate the detoxification of both exogenous and endogenous compounds [19,38]. GSTs also exhibit a degree of antioxidant activity, particularly in relation to the inactivation of end products resulting from lipid peroxidation [49,50]. Hence, the association of these enzymes with oxidative stress can be attributed to the depletion of glutathione levels, which are utilized for neutralizing foreign chemical compounds. The omega class glutathione S-transferases (GSTO) are cytosolic enzymes that have been identified in various species, including humans [51]. In contrast to other classes of GSTs typically contain tyrosine or serine residues in their active sites, the active sites of GSTO enzymes feature an N-terminal cysteine residue that can bind to glutathione [52]. There is a growing body of evidence indicating that GSTO enzymes play a significant role in the detoxification of various exogenous stressors. In vitro studies indicate that human GSTO1 modulates the ryanodine receptor in the sarcoplasmic/endoplasmic reticulum, which is essential for Ca2+ release during excitation-contraction coupling in cardiac and skeletal muscles [53]. Furthermore, these studies indicated that human GSTO1 modulates the signaling pathway in c-Jun N-terminal kinase (JNK)-mediated apoptosis and activates interleukin-1β, a crucial mediator of inflammation [51]. Human GSTO1 has also been shown to regulate lipopolysaccharide-induced inflammatory responses in macrophages [54]. GSTO1 plays a critical role in redox homeostasis by influencing the glutathionylation and deglutathionylation of target proteins [51].
Oxidative stress is vital in psoriasis pathogenesis, prompting studies on the link between functionally significant glutathione S-transferase gene polymorphisms and disease risk. Certain studies indicate significant correlations between these variants and an increased susceptibility to psoriasis; however, other research has not found any associations with the risk of the disease [55,56,57,58]. The present study is the first to investigate whether polymorphisms in the GSTO1 gene contribute to the risk of developing psoriasis. No statistically significant correlations were found between polymorphisms of the GSTO1 gene and psoriasis risk when analyzing the overall case and control groups. The sex-stratified analysis revealed that two polymorphisms—specifically, rs11191736 in males and rs187304410 in females—are associated with the risk of psoriasis. Although the associations of psoriasis with the rs11191736 and rs187304410 polymorphisms could not be replicated in the overall population of the UK Biobank, 40 other SNPs in the GSTO1 gene have been identified as nominally associated with the risk of psoriasis (Supplementary Table 1). These findings clearly highlight the significant role of GSTO1 gene polymorphisms in psoriasis susceptibility. Analyses of haplotypes and diplotypes indicated that there were either no significant joint effects or only weak associations between the combinations of GSTO1 genotypes and the risk of developing psoriasis. Nonetheless, bioinformatics modeling of the non-linear interactions between GSTO1 and GCLC gene polymorphisms using the mbmdr method has revealed the existence of higher-order epistatic interactions between these genes, as well as with environmental risk factors, in relation to the risk of developing psoriasis. Moreover, the vast majority of mbmdr GxE models incorporated the combined effects of alcohol abuse and the polymorphisms of the GSTO1 and GCLC genes on the risk of developing the disease. Research indicates that alcohol abuse significantly affects both the onset and progression of psoriasis [59,60]. Our recent study identified the joint effects of alcohol consumption and GCLC gene polymorphisms on the psoriasis risk [39]. Apparently, these gene-environment interactions can be elucidated by the fact that, on one hand, the consumption of substantial amounts of alcohol is linked to the production of a significant number of reactive oxygen species and oxidative stress [61,62]. This process depletes the reserves of reduced glutathione and causes damage to liver tissue, where the majority of the body’s glutathione is synthesized [63,64]. On the other hand, the development of oxidative stress may be exacerbated by reduced glutathione synthesis resulting from decreased expression of the GCLC gene and/or increased utilization of reduced glutathione by the GSTO1 enzyme.
To understand the mechanisms by which polymorphisms are associated with the onset of psoriasis, it is crucial to analyze the functional annotation data of the GSTO1 polymorphisms. SNP rs187304410, which has been associated with a reduced risk of psoriasis in females according to the GTEx portal (https://www.gtexportal.org/home/, accessed on December 8, 2024), correlates with alternative splicing events of the GSTO1 and GSTO2 genes in suprapubic and lower leg skin, as well as in cultured skin fibroblasts (https://www.gtexportal.org/home/, accessed on December 8, 2024). One of the alternative splicing sites is situated within intron 9 and is associated with a reduction in the expression of one GSTO1 isoform. Conversely, the other site, located in intron 7, is linked to an elevated production of a different isoform of the enzyme. It is well established that alternative splicing can lead to the generation of multiple isoforms, some of which remain unknown [65]. To address the question regarding the properties and functions of the presumed GSTO1 isoforms with altered functional activity, it is essential to conduct experimental studies aimed at determining enzyme activity, substrate specificity, and other characteristics. In the blood, the polymorphism rs187304410 (allele A) is strongly associated with the expression levels of GSTO2, according to data from the eQTLGen consortium (https://www.eqtlgen.org/cis-eqtls.html, accessed on December 8, 2024). Other SNPs were found to be correlated with the expression levels of the GSTO1 and GSTO2 genes in the skin. The GSTO1 and GSTO2 genes are situated in proximity to one another on the same chromosomal segment, 10q25.1. Their transcriptional activity in skin, specifically in foreskin fibroblast primary cells, seems to be regulated by shared enhancers found in the adjacent genomic region, 104470800-104472000, as indicated by the Super-Enhancer database (http://www.licpathway.net/sedb/, accessed on December 8, 2024).
As mentioned above, GSTOs are involved in the biotransformation of xenobiotics and play a crucial role in protecting the skin from damage caused by environmental chemicals that penetrate both the superficial and deeper layers of the skin through transcutaneous and systemic routes [10,66]. Hence, the skin is one of the primary targets of environmental pollutants, and the disruption of the skin barrier due to inadequate xenobiotic biotransformation may potentially cause or exacerbate psoriasis. In particular, certain chemicals, especially drugs such as hydroxychloroquine [67], imiquimod [68], and lithium [69], have been identified as agents that can induce and exacerbate psoriasis. Per- and polyfluoroalkyl substances (PFAS), a class of synthetic chemicals extensively used in various consumer products, have been found to contribute to the risk of psoriasis [70]. Human exposure to PFAS primarily occurs through ingestion and inhalation, with the main sources being food, drinking water, and airborne dust. GSTO1 and GSTO2 are involved in the biotransformation of inorganic arsenic and play a crucial role in the reduction of monomethylarsonic acid, a highly toxic intermediate in the methylation process of inorganic arsenic [71,72]. Monomethylarsonic acid, a metabolite of arsenic, is the active ingredient in over 600 herbicides used in various agricultural products, including cotton, almonds, and oranges, as well as in landscaping applications [73]. GSTO2, like GSTO1, exhibits glutathione-dependent thiol transferase activity and shows significantly higher dehydroascorbate reductase activity—approximately 70 to 100 times greater than that of GSTO1 [72]. This enhanced activity may contribute to the regeneration of ascorbic acid, thereby providing protection against oxidative stress. This function of GSTO2 aligns with research findings indicating that ascorbic acid alleviates symptoms associated with skin lesions in psoriasis [37,74].
Our study has both strengths and limitations. This investigation is the first to explore the associations between GSTO1 gene polymorphisms and the risk of psoriasis. It is also one of the largest studies conducted on GST gene polymorphisms and their susceptibility to psoriasis. The application of the mbmdr methods enabled the identification of epistatic interactions between the polymorphisms of the GSTO1 gene and variants of GCLC in determining the risk of psoriasis. Given that our research is a pilot study, it is essential to conduct further investigations in independent populations to validate the associations identified between GSTO1 gene polymorphisms and the risk of psoriasis. Furthermore, as our study examined a relatively limited number of single nucleotide polymorphisms, it is advisable for future genetic association studies to explore the relationship between psoriasis and a broader array of polymorphisms, including those that have demonstrated associations with psoriasis in the UK Biobank cohorts (as detailed in the Supplementary Table 1). Additionally, since the polymorphisms analyzed in the GSTO1 gene are located in noncoding regions, any phenotypic implications should be approached with caution, as no assessments of gene expression in skin biopsies from the study participants have been conducted.
5. Conclusions
The present study demonstrates, for the first time, that polymorphisms in the gene encoding glutathione S-transferase omega 1 serve as novel genetic markers for susceptibility to psoriasis. Bioinformatics analysis has revealed that polymorphisms in the GSTO1 gene exhibit a significant epistatic interaction with polymorphisms in the GCLC gene. Collectively, these genetic variations, along with patterns of alcohol abuse, contribute to an increased susceptibility to psoriasis. Potential mechanisms by which GSTO1 gene polymorphisms may contribute to the development of psoriasis are summarized in Figure 1.
Our proposed hypothesis suggests that the predisposition to psoriasis is influenced by significant interactions between the polymorphisms of the GSTO1 and GCLC genes. The effect of these genetic variations on the development of the disease is mediated by the presence of risk factors, with alcohol consumption identified as a critical factor (as illustrated in the central portion of the figure). Furthermore, the risk of developing psoriasis exhibits a degree of sexual dimorphism, which is evident in the SNP-phenotypic associations observed. In females, the influence of polymorphisms in the GSTO1 gene is more pronounced than that of the GCLC gene. In contrast, in males, the polymorphisms of both genes analyzed contribute almost equally to the risk. Sexual dimorphism in the associations of GSTO1 polymorphisms with psoriasis—specifically, the main effects of SNP rs187304410 observed exclusively in women—may be attributed to the greater sensitivity of women to the toxic effects of chemical pollutants, as demonstrated by epidemiological studies [15,75]. The phenotypic effects associated with GSTO1 gene polymorphisms are likely mediated by alterations—specifically reductions—in the expression levels of the GSTO1 and GSTO2 genes, as well as other adjacent genes that may be regulated by shared enhancers located within the genomic region 10q25.1. In addition, functional SNP annotation data indicate that polymorphisms in GSTO1 and GSTO2 are sites for alternative splicing of these genes in the skin. This may lead to the production of functionally distinct enzyme isoforms with varying activities concerning the conjugation of cutaneous xenobiotics and free radicals with glutathione. The aforementioned metabolic disorders may be exacerbated by a reduction in cellular glutathione levels, which can occur due to diminished synthesis associated with decreased expression of the GCLC gene, the catalytic subunit of glutamate cysteine ligase. Additionally, increased consumption of glutathione by glutathione S-transferases and glutathione peroxidases, which neutralize free radicals and xenobiotics, along with the detrimental effects of alcohol, smoking, and other external factors, contribute to this depletion. Consequently, these disorders lead to the accumulation of free radicals and toxic substances, thereby initiating oxidative stress and causing damage to skin tissue. It is unequivocal that oxidative stress acts as a mechanistic pathway in the pathogenesis of psoriasis and represents a fundamental mechanism underlying the detrimental effects of environmental chemicals [9]. Gaining deeper insights into how the polymorphisms of the GSTO1 gene interact with other genes associated with glutathione metabolism could pave the way for innovative, evidence-based strategies for treating and preventing psoriasis.
Author Contributions
Conceptualization, A.P.; methodology, A.P., R.S., E.E., O.B. and M.C.; software, A.P. and M.S.; validation, A.P. and M.C.; formal analysis, A.P., R.S. and O.B.; investigation, A.P., R.S., E.E., and O.B.; resources, R.S. and E.E.; data curation, A.P. and M.S.; writing—original draft preparation, A.P. and M.S.; writing—review and editing, A.P., M.S. and M.C.; visualization, A.P.; supervision, A.P.; project administration, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of Kursk State Medical University (protocol No. 4, 9.04.2018).
Informed Consent Statement
Informed consent was obtained from all participants involved in the study.
Data Availability Statement
Data supporting reported results are available upon request.
Acknowledgments
We thank all psoriasis patients, healthy volunteers, and hospital staff in the Kursk region for their participation in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Campanati A, Marani A, Martina E, Diotallevi F, Radi G, Offidani A. Psoriasis as an Immune-Mediated and Inflammatory Systemic Disease: From Pathophysiology to Novel Therapeutic Approaches. Biomedicines. 2021 Oct 21;9(11):1511. [CrossRef]
- Parisi R, Symmons DP, Griffiths CE, Ashcroft DM; Identification and Management of Psoriasis and Associated ComorbidiTy (IMPACT) project team. Global epidemiology of psoriasis: a systematic review of incidence and prevalence. J Invest Dermatol. 2013 Feb;133(2):377-85. [CrossRef]
- Rendon A, Schäkel K. Psoriasis Pathogenesis and Treatment. Int J Mol Sci. 2019 Mar 23;20(6):1475. [CrossRef]
- Capon, F. The Genetic Basis of Psoriasis. Int J Mol Sci. 2017 Nov 25;18(12):2526. [CrossRef]
- Yang C, Chen M, Huang H, Li X, Qian D, Hong X, Zheng L, Hong J, Hong J, Zhu Z, Zheng X, Sheng Y, Zhang X. Exome-Wide Rare Loss-of-Function Variant Enrichment Study of 21,347 Han Chinese Individuals Identifies Four Susceptibility Genes for Psoriasis. J Invest Dermatol. 2020 Apr;140(4):799-805.e1. [CrossRef]
- Kamiya K, Kishimoto M, Sugai J, Komine M, Ohtsuki M. Risk Factors for the Development of Psoriasis. Int J Mol Sci. 2019 Sep 5;20(18):4347. [CrossRef]
- Lønnberg AS, Skov L, Skytthe A, Kyvik KO, Pedersen OB, Thomsen SF. Heritability of psoriasis in a large twin sample. Br J Dermatol. 2013 Aug;169(2):412-6. [CrossRef]
- Zeng J, Luo S, Huang Y, Lu Q. Critical role of environmental factors in the pathogenesis of psoriasis. J Dermatol. 2017 Aug;44(8):863-872. [CrossRef]
- Puri P, Nandar SK, Kathuria S, Ramesh V. Effects of air pollution on the skin: A review. Indian J Dermatol Venereol Leprol. 2017 Jul-Aug;83(4):415-423. [CrossRef]
- Araviiskaia E, Berardesca E, Bieber T, Gontijo G, Sanchez Viera M, Marrot L, Chuberre B, Dreno B. The impact of airborne pollution on skin. J Eur Acad Dermatol Venereol. 2019 Aug;33(8):1496-1505. [CrossRef]
- Wu J, Ma Y, Yang J, Tian Y. Exposure to Air Pollution, Genetic Susceptibility, and Psoriasis Risk in the UK. JAMA Netw Open. 2024 Jul 1;7(7):e2421665. [CrossRef]
- Liaw FY, Chen WL, Kao TW, Chang YW, Huang CF. Exploring the link between cadmium and psoriasis in a nationally representative sample. Sci Rep. 2017 May 11;7(1):1723. [CrossRef]
- Wacewicz-Muczyńska M, Socha K, Soroczyńska J, Niczyporuk M, Borawska MH. Cadmium, lead and mercury in the blood of psoriatic and vitiligo patients and their possible associations with dietary habits. Sci Total Environ. 2021 Feb 25;757:143967. [CrossRef]
- Bellinato F, Adami G, Vaienti S, Benini C, Gatti D, Idolazzi L, Fassio A, Rossini M, Girolomoni G, Gisondi P. Association Between Short-term Exposure to Environmental Air Pollution and Psoriasis Flare. JAMA Dermatol. 2022 Apr 1;158(4):375-381. [CrossRef]
- Wu J, Chen H, Yang R, Yu H, Shang S, Hu Y. Short-term exposure to ambient fine particulate matter and psoriasis: A time-series analysis in Beijing, China. Front Public Health. 2022 Oct 13;10:1015197. [CrossRef]
- Chen Y, Pan Z, Shen J, Wu Y, Fang L, Xu S, Ma Y, Zhao H, Pan F. Associations of exposure to blood and urinary heavy metal mixtures with psoriasis risk among U.S. adults: A cross-sectional study. Sci Total Environ. 2023 Aug 20;887:164133. [CrossRef]
- Götz C, Pfeiffer R, Tigges J, Blatz V, Jäckh C, Freytag EM, Fabian E, Landsiedel R, Merk HF, Krutmann J, Edwards RJ, Pease C, Goebel C, Hewitt N, Fritsche E. Xenobiotic metabolism capacities of human skin in comparison with a 3D epidermis model and keratinocyte-based cell culture as in vitro alternatives for chemical testing: activating enzymes (Phase I). Exp Dermatol. 2012 May;21(5):358-63. [CrossRef]
- van Eijl S, Zhu Z, Cupitt J, Gierula M, Götz C, Fritsche E, Edwards RJ. Elucidation of xenobiotic metabolism pathways in human skin and human skin models by proteomic profiling. PLoS One. 2012;7(7):e41721. [CrossRef]
- Hayes JD, Flanagan JU, Jowsey IR. Glutathione transferases. Annu Rev Pharmacol Toxicol. 2005;45:51-88. [CrossRef]
- Ryu J, Park SG, Park BC, Choe M, Lee KS, Cho JW. Proteomic analysis of psoriatic skin tissue for identification of differentially expressed proteins: up-regulation of GSTP1, SFN and PRDX2 in psoriatic skin. Int J Mol Med. 2011 Nov;28(5):785-92. [CrossRef]
- Karadag AS, Uzunçakmak TK, Ozkanli S, Oguztuzun S, Moran B, Akbulak O, Ozlu E, Zemheri IE, Bilgili SG, Akdeniz N. An investigation of cytochrome p450 (CYP) and glutathione S-transferase (GST) isoenzyme protein expression and related interactions with phototherapy in patients with psoriasis vulgaris. Int J Dermatol. 2017 Feb;56(2):225-231. [CrossRef]
- Akbulak O, Karadag AS, Akdeniz N, Ozkanli S, Ozlu E, Zemheri E, Oguztuzun S. Evaluation of oxidative stress via protein expression of glutathione S-transferase and cytochrome p450 (CYP450) ısoenzymes in psoriasis vulgaris patients treated with methotrexate. Cutan Ocul Toxicol. 2018 Jun;37(2):180-185. [CrossRef]
- Reich K, Westphal G, Schulz T, Müller M, Zipprich S, Fuchs T, Hallier E, Neumann C. Combined analysis of polymorphisms of the tumor necrosis factor-alpha and interleukin-10 promoter regions and polymorphic xenobiotic metabolizing enzymes in psoriasis. J Invest Dermatol. 1999 Aug;113(2):214-20. [CrossRef]
- Richter-Hintz D, Their R, Steinwachs S, Kronenberg S, Fritsche E, Sachs B, Wulferink M, Tonn T, Esser C. Allelic variants of drug metabolizing enzymes as risk factors in psoriasis. J Invest Dermatol. 2003 May;120(5):765-70. [CrossRef]
- Gambichler T, Kreuter A, Susok L, Skrygan M, Rotterdam S, Höxtermann S, Müller M, Tigges C, Altmeyer P, Lahner N. Glutathione-S-transferase T1 genotyping and phenotyping in psoriasis patients receiving treatment with oral fumaric acid esters. J Eur Acad Dermatol Venereol. 2014 May;28(5):574-80. [CrossRef]
- Solak B, Karkucak M, Turan H, Ocakoğlu G, Özemri Sağ Ş, Uslu E, Yakut T, Erdem T. Glutathione S-Transferase M1 and T1 Gene Polymorphisms in Patients with Chronic Plaque-Type Psoriasis: A Case-Control Study. Med Princ Pract. 2016;25(2):155-8. [CrossRef]
- Hruska P, Rybecka S, Novak J, Zlamal F, Splichal Z, Slaby O, Vasku V, Bienertova-Vasku J. Combinations of common polymorphisms within GSTA1 and GSTT1 as a risk factor for psoriasis in a central European population: a case-control study. J Eur Acad Dermatol Venereol. 2017 Oct;31(10):e461-e463. [CrossRef]
- Srivastava DSL, Jain VK, Verma P, Yadav JP. Polymorphism of glutathione S-transferase M1 and T1 genes and susceptibility to psoriasis disease: A study from North India. Indian J Dermatol Venereol Leprol. 2018 Jan-Feb;84(1):39-44. [CrossRef]
- Guarneri F, Sapienza D, Papaianni V, Marafioti I, Guarneri C, Mondello C, Roccuzzo S, Asmundo A, Cannavò SP. Association between genetic polymorphisms of glutathione S-transferase M1/T1 and psoriasis in a population from the area of the strict of messina (Southern Italy). Free Radic Res. 2020 Jan;54(1):57-63. [CrossRef]
- Tawfik, Noha Z.a. AbdallahHoda Y.bAbdullahMona E.aAlshaarawyHagar F.a.AtwaMona A.a.Glutathione S-transferase M1 and T1 gene polymorphisms in psoriasis patients: a pilot case-control study. Egyptian Journal of Dermatology and Venereology 43(3):p 200-207, September-December 2023. [CrossRef]
- Dursun HG, Dursun R, Çınar Ayan İ, Zamani AG, Yıldırım MS. Relationship between Glutathione S-transferase gene polymorphisms and clinical features of psoriasis: A case-control study in the Turkish population. Turkderm-Turk Arch Dermatol Venereol. 2024;58:75-82. [CrossRef]
- Klyosova, E.; Azarova, I.; Polonikov, A. A Polymorphism in the Gene Encoding Heat Shock Factor 1 (HSF1) Increases the Risk of Type 2 Diabetes: A Pilot Study Supports a Role for Impaired Protein Folding in Disease Pathogenesis. Life 2022a, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
- Lazarenko, V.; Churilin, M.; Azarova, I.; Klyosova, E.; Bykanova, M.; Ob’edkova, N.; Churnosov, M.; Bushueva, O.; Mal, G.; Povetkin, S.; et al. Comprehensive Statistical and Bioinformatics Analysis in the Deciphering of Putative Mechanisms by Which Lipid-Associated GWAS Loci Contribute to Coronary Artery Disease. Biomedicines 2022, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Kobzeva, K.A.; Shilenok, I.V.; Belykh, A.E.; Gurtovoy, D.E.; Bobyleva, L.A.; Krapiva, A.B.; Stetskaya, T.A.; Bykanova, M.A.; Mezhenskaya, A.A.; Lysikova, E.A.; et al. C9orf16 (BBLN) gene, encoding a member of Hero proteins, is a novel marker in ischemic stroke risk. Res. Results Biomed. 2022, 8, 278–292. [Google Scholar] [CrossRef]
- Di Meglio, P.; Villanova, F.; Nestle, F.O. Psoriasis. Cold Spring Harb. Perspect. Med. 2014, 4, a015354. [Google Scholar] [CrossRef] [PubMed]
- Klyosova, E.Y.; Azarova, I.E.; Sunyaykina, O.A.; Polonikov, A.V. Validity of a brief screener for environmental risk factors of age-related diseases using type 2 diabetes and coronary artery disease as examples. Res. Results Biomed. 2022b, 8, 130–137. [Google Scholar]
- Garbicz J, Całyniuk B, Górski M, Buczkowska M, Piecuch M, Kulik A, Rozentryt P. Nutritional Therapy in Persons Suffering from Psoriasis. Nutrients. 2021 Dec 28;14(1):119. [CrossRef]
- Mazari AMA, Zhang L, Ye ZW, Zhang J, Tew KD, Townsend DM. The Multifaceted Role of Glutathione S-Transferases in Health and Disease. Biomolecules. 2023 Apr 18;13(4):688. [CrossRef]
- Efanova E, Bushueva O, Saranyuk R, Surovtseva A, Churnosov M, Solodilova M, Polonikov A. Polymorphisms of the GCLC Gene Are Novel Genetic Markers for Susceptibility to Psoriasis Associated with Alcohol Abuse and Cigarette Smoking. Life (Basel). 2023 Jun 2;13(6):1316. [CrossRef]
- Azarova I, Klyosova E, Polonikov A. Association between RAC1 gene variation, redox homeostasis and type 2 diabetes mellitus. Eur J Clin Invest. 2022 Aug;52(8):e13792. [CrossRef]
- Stetskaya TA, Kobzeva KA, Zaytsev SM, et al. HSPD1 gene polymorphism is associated with an increased risk of ischemic stroke in smokers. Research Results in Biomedicine. 2024;10(2):175-186. [CrossRef]
- Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, Maller J, Sklar P, de Bakker PI, Daly MJ, Sham PC. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007 Sep;81(3):559-75. [CrossRef]
- Calle ML, Urrea V, Malats N, et al. mbmdr: an R package for exploring gene-gene interactions associated with binary or quantitative traits. Bioinformatics. 2010;26(17):2198-9.
- Polonikov A, Bocharova I, Azarova I, Klyosova E, Bykanova M, Bushueva O, Polonikova A, Churnosov M, Solodilova M. The Impact of Genetic Polymorphisms in Glutamate-Cysteine Ligase, a Key Enzyme of Glutathione Biosynthesis, on Ischemic Stroke Risk and Brain Infarct Size. Life (Basel). 2022 Apr 18;12(4):602. [CrossRef]
- Zhabin S, Lazarenko V, Azarova I, Klyosova E, Bykanova M, Chernousova S, Bashkatov D, Gneeva E, Polonikova A, Churnosov M, Solodilova M, Polonikov A. The Joint Link of the rs1051730 and rs1902341 Polymorphisms and Cigarette Smoking to Peripheral Artery Disease and Atherosclerotic Lesions of Different Arterial Beds. Life (Basel). 2023 Feb 10;13(2):496. [CrossRef]
- Ritchie, M.D.; Hahn, L.W.; Roodi, N.; Bailey, L.R.; Dupont, W.D.; Parl, F.F.; Moore, J.H. Multifactor-Dimensionality Reduction Reveals High-Order Interactions among Estrogen-Metabolism Genes in Sporadic Breast Cancer. Am. J. Hum. Genet. 2001, 69, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay A, Lu TP. Gene-gene interaction: the curse of dimensionality. Ann Transl Med. 2019 Dec;7(24):813. [CrossRef]
- Lazarenko V, Churilin M, Azarova I, Klyosova E, Bykanova M, Ob’edkova N, Churnosov M, Bushueva O, Mal G, Povetkin S, Kononov S, Luneva Y, Zhabin S, Polonikova A, Gavrilenko A, Saraev I, Solodilova M, Polonikov A. Comprehensive Statistical and Bioinformatics Analysis in the Deciphering of Putative Mechanisms by Which Lipid-Associated GWAS Loci Contribute to Coronary Artery Disease. Biomedicines. 2022 Jan 25;10(2):259. [CrossRef]
- Sharma R, Yang Y, Sharma A, Awasthi S, Awasthi YC. Antioxidant role of glutathione S-transferases: protection against oxidant toxicity and regulation of stress-mediated apoptosis. Antioxid Redox Signal. 2004 Apr;6(2):289-300. [CrossRef]
- Singhal SS, Singh SP, Singhal P, Horne D, Singhal J, Awasthi S. Antioxidant role of glutathione S-transferases: 4-Hydroxynonenal, a key molecule in stress-mediated signaling. Toxicol Appl Pharmacol. 2015 Dec 15;289(3):361-70. [CrossRef]
- Kim Y, Cha SJ, Choi HJ, Kim K. Omega Class Glutathione S-Transferase: Antioxidant Enzyme in Pathogenesis of Neurodegenerative Diseases. Oxid Med Cell Longev. 2017;2017:5049532. [CrossRef]
- Board PG, Menon D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochim Biophys Acta. 2013 May;1830(5):3267-88. [CrossRef]
- Lanner JT, Georgiou DK, Joshi AD, Hamilton SL. Ryanodine receptors: structure, expression, molecular details, and function in calcium release. Cold Spring Harb Perspect Biol. 2010 Nov;2(11):a003996. [CrossRef]
- Menon D, Coll R, O’Neill LA, Board PG. Glutathione transferase omega 1 is required for the lipopolysaccharide-stimulated induction of NADPH oxidase 1 and the production of reactive oxygen species in macrophages. Free Radic Biol Med. 2014 Aug;73:318-27. [CrossRef]
- Hruska P, Rybecka S, Novak J, Zlamal F, Splichal Z, Slaby O, Vasku V, Bienertova-Vasku J. Combinations of common polymorphisms within GSTA1 and GSTT1 as a risk factor for psoriasis in a central European population: a case-control study. J Eur Acad Dermatol Venereol. 2017 Oct;31(10):e461-e463. [CrossRef]
- Dursun HG, Dursun R, Çınar Ayan İ, Zamani AG, Yıldırım MS. Relationship between Glutathione S-transferase gene polymorphisms and clinical features of psoriasis: A case-control study in the Turkish population. Turkderm-Turk Arch Dermatol Venereol. 2024;58:75-82. [CrossRef]
- Wang ZM, Wang DZ, Zhang KY, Huang W, Liu JJ. Systematic evaluation of association between the microsomal glutathione S-transferase 2 common variation and psoriasis vulgaris in Chinese population. Arch Dermatol Res. 2006 Aug;298(3):107-12. [CrossRef]
- Campione E, Mazzilli S, Di Prete M, Dattola A, Cosio T, Lettieri Barbato D, Costanza G, Lanna C, Manfreda V, Gaeta Schumak R, Prignano F, Coniglione F, Ciprani F, Aquilano K, Bianchi L. The Role of Glutathione-S Transferase in Psoriasis and Associated Comorbidities and the Effect of Dimethyl Fumarate in This Pathway. Front Med (Lausanne). 2022 Feb 8;9:760852. [CrossRef]
- Farkas A, Kemény L. Alcohol, liver, systemic inflammation and skin: a focus on patients with psoriasis. Skin Pharmacol Physiol. 2013;26(3):119-26. [CrossRef]
- Farkas A, Kemény L, Széll M, Dobozy A, Bata-Csörgo Z. Ethanol and acetone stimulate the proliferation of HaCaT keratinocytes: the possible role of alcohol in exacerbating psoriasis. Arch Dermatol Res. 2003 Jun;295(2):56-62. [CrossRef]
- Wu D, Cederbaum AI. Alcohol, oxidative stress, and free radical damage. Alcohol Res Health. 2003;27(4):277-84.
- Michalak A, Lach T, Cichoż-Lach H. Oxidative Stress-A Key Player in the Course of Alcohol-Related Liver Disease. J Clin Med. 2021 Jul 6;10(14):3011. [CrossRef]
- Lu, SC. Glutathione synthesis. Biochim Biophys Acta. 2013 May;1830(5):3143-53. [CrossRef]
- Vairetti M, Di Pasqua LG, Cagna M, Richelmi P, Ferrigno A, Berardo C. Changes in Glutathione Content in Liver Diseases: An Update. Antioxidants (Basel). 2021 Feb 28;10(3):364. [CrossRef]
- Yamaguchi K, Ishigaki K, Suzuki A, Tsuchida Y, Tsuchiya H, Sumitomo S, Nagafuchi Y, Miya F, Tsunoda T, Shoda H, Fujio K, Yamamoto K, Kochi Y. Splicing QTL analysis focusing on coding sequences reveals mechanisms for disease susceptibility loci. Nat Commun. 2022 Aug 24;13(1):4659. [CrossRef]
- Lowe ME, Akhtari FS, Potter TA, Fargo DC, Schmitt CP, Schurman SH, Eccles KM, Motsinger-Reif A, Hall JE, Messier KP. The skin is no barrier to mixtures: Air pollutant mixtures and reported psoriasis or eczema in the Personalized Environment and Genes Study (PEGS). J Expo Sci Environ Epidemiol. 2023 May;33(3):474-481. [CrossRef]
- Sachdeva M, Mufti A, Maliyar K, Lytvyn Y, Yeung J. Hydroxychloroquine effects on psoriasis: A systematic review and a cautionary note for COVID-19 treatment. J Am Acad Dermatol. 2020 Aug;83(2):579-586. [CrossRef]
- van der Fits L, Mourits S, Voerman JS, Kant M, Boon L, Laman JD, Cornelissen F, Mus AM, Florencia E, Prens EP, Lubberts E. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J Immunol. 2009 May 1;182(9):5836-45. [CrossRef]
- Jafferany, M. Lithium and psoriasis: what primary care and family physicians should know. Prim Care Companion J Clin Psychiatry. 2008;10(6):435-9. [CrossRef]
- Zhang Q, Zhang M, Zhao C. Exposure to Per- and Polyfluoroalkyl Substances and Risk of Psoriasis: A Population-Based Study. Toxics. 2024; 12(11):828. [CrossRef]
- Zakharyan RA, Sampayo-Reyes A, Healy SM, Tsaprailis G, Board PG, Liebler DC, Aposhian HV. Human monomethylarsonic acid (MMA(V)) reductase is a member of the glutathione-S-transferase superfamily. Chem Res Toxicol. 2001 Aug;14(8):1051-7. [CrossRef]
- Schmuck EM, Board PG, Whitbread AK, Tetlow N, Cavanaugh JA, Blackburn AC, Masoumi A. Characterization of the monomethylarsonate reductase and dehydroascorbate reductase activities of Omega class glutathione transferase variants: implications for arsenic metabolism and the age-at-onset of Alzheimer’s and Parkinson’s diseases. Pharmacogenet Genomics. 2005 Jul;15(7):493-501. [CrossRef]
- Barnett, J. B. , and K. M. Brundage. “Immunotoxicology of pesticides and chemotherapies.” In Comprehensive Toxicology (Second Edition) Elsevier Science (2010): 467-487.
- Kitahata K, Matsuo K, Hara Y, Naganuma T, Oiso N, Kawada A, Nakayama T. Ascorbic acid derivative DDH-1 ameliorates psoriasis-like skin lesions in mice by suppressing inflammatory cytokine expression. J Pharmacol Sci. 2018 Dec;138(4):284-288. [CrossRef]
- Fares-Medina S, Díaz-Caro I, García-Montes R, Corral-Liria I, García-Gómez-Heras S. Multiple Chemical Sensitivity Syndrome: First Symptoms and Evolution of the Clinical Picture: Case-Control Study/Epidemiological Case-Control Study. Int J Environ Res Public Health. 2022 Nov 29;19(23):15891. [CrossRef]
Figure 1.
Proposed mechanisms for the involvement of GSTO1 and GCLC gene polymorphisms in the pathogenesis of psoriasis. The central section of the figure provides a schematic representation of the interactions between single nucleotide polymorphisms and their relationships with environmental factors, specifically alcohol abuse and cigarette smoking. These factors are the most significantly correlated with the risk of psoriasis, as determined by the top five n-order mbmdr models. The blue lines in the central section represent interactions among males, while the pink lines indicate interactions among females. The left and right sections of the figure display violin plots of expression quantitative trait loci (eQTLs) and splicing quantitative trait loci (sQTLs) for the studied SNPs obtained from the GTEx portal (https://www.gtexportal.org/home/, accessed on December 8, 2024). Below the plots are photographs of psoriasis patients from our own collection. Comments regarding the figure are provided in the main text of the article.
Figure 1.
Proposed mechanisms for the involvement of GSTO1 and GCLC gene polymorphisms in the pathogenesis of psoriasis. The central section of the figure provides a schematic representation of the interactions between single nucleotide polymorphisms and their relationships with environmental factors, specifically alcohol abuse and cigarette smoking. These factors are the most significantly correlated with the risk of psoriasis, as determined by the top five n-order mbmdr models. The blue lines in the central section represent interactions among males, while the pink lines indicate interactions among females. The left and right sections of the figure display violin plots of expression quantitative trait loci (eQTLs) and splicing quantitative trait loci (sQTLs) for the studied SNPs obtained from the GTEx portal (https://www.gtexportal.org/home/, accessed on December 8, 2024). Below the plots are photographs of psoriasis patients from our own collection. Comments regarding the figure are provided in the main text of the article.
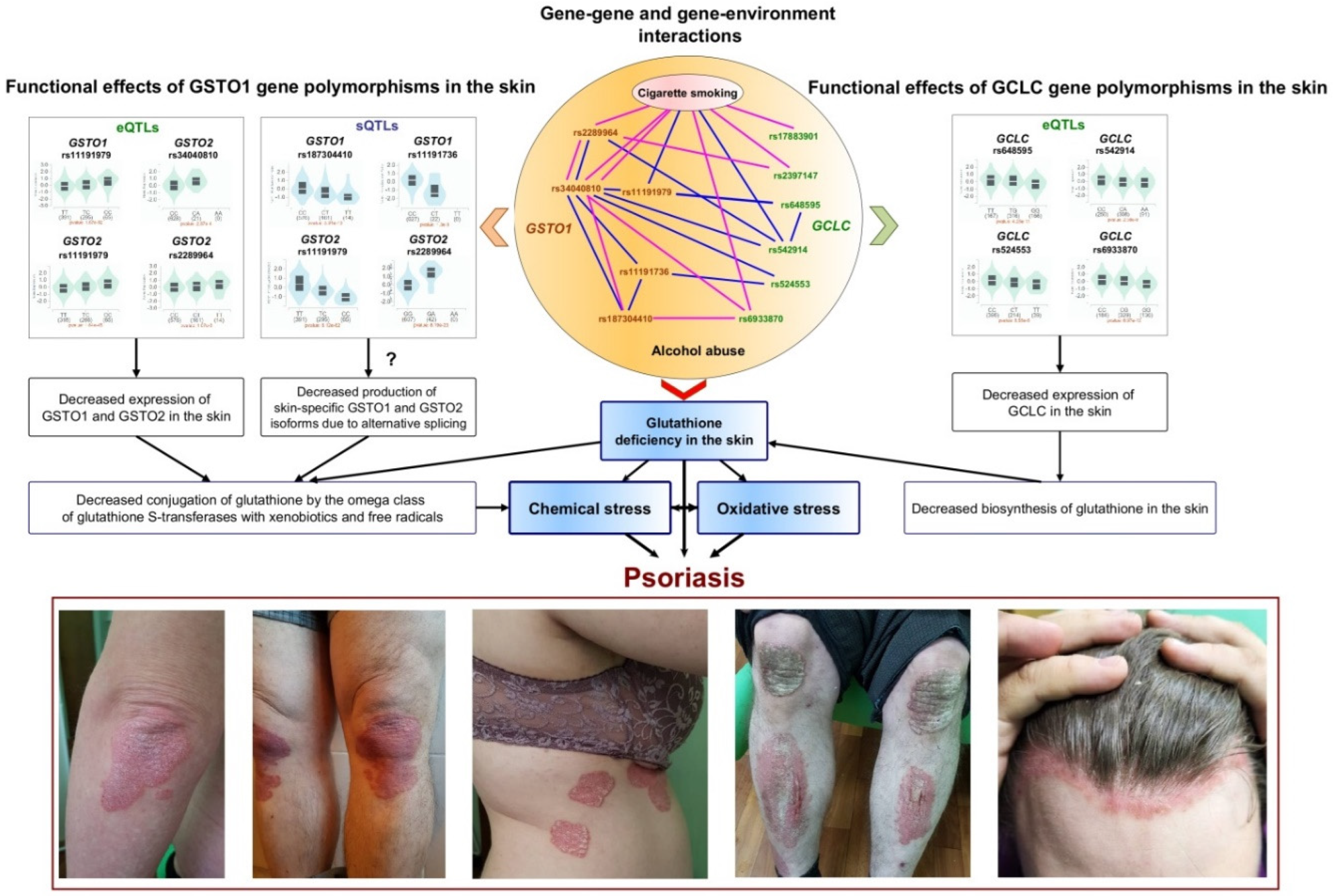

Table 1.
A summary of associations between GSTO1 gene polymorphisms and psoriasis risk in the entire and sex-stratified groups.
Table 1.
A summary of associations between GSTO1 gene polymorphisms and psoriasis risk in the entire and sex-stratified groups.
| SNP ID | Minor allele | N | Permutation P-values (Pperm) estimated for genetic models of SNP-disease associations* | |||
| Allelic | Additive | Dominant | Recessive | |||
| Entire groups | ||||||
| rs11191736 | T | 901 | 0.26 | 0.22 | 0.19 | NA |
| rs34040810 | A | 944 | 0.45 | 0.29 | 0.39 | NA |
| rs2289964 | T | 941 | 0.86 | 1.00 | 0.78 | 0.86 |
| rs11191979 | C | 930 | 0.59 | 1.00 | 0.67 | 0.73 |
| rs187304410 | A | 944 | 0.86 | 0.31 | 0.52 | NA |
| Males | ||||||
| rs11191736 | T | 467 | 0.02 | NA | NA | NA |
| rs34040810 | A | 486 | 0.34 | 0.39 | 0.64 | NA |
| rs2289964 | T | 486 | 0.78 | 0.64 | 0.55 | 0.59 |
| rs11191979 | C | 479 | 0.71 | 0.67 | 0.52 | 0.59 |
| rs187304410 | A | 486 | 0.46 | 0.31 | 0.29 | NA |
| Females | ||||||
| rs11191736 | T | 434 | 0.25 | NA | NA | NA |
| rs34040810 | A | 458 | 0.86 | 0.63 | 0.55 | NA |
| rs2289964 | T | 455 | 0.33 | 0.33 | 0.29 | NA |
| rs11191979 | C | 451 | 0.86 | 0.86 | 1.00 | 1.00 |
| rs187304410 | A | 458 | 0.02 | 0.012 | 0.011 | NA |
Significance of SNP-disease associations was assessed by adaptive permutations using the PLINK software v.1.9. NA, not available.
Table 2.
Genotype and allele frequencies of the GSTO1 gene and their associations with psoriasis risk in the entire and sex-stratified groups.
Table 2.
Genotype and allele frequencies of the GSTO1 gene and their associations with psoriasis risk in the entire and sex-stratified groups.
| SNP | Genotype/ allele |
Healthy Controls1 | Patients with psoriasis1 | OR2 (95% CI) | Pperm3 | ||
| n | % | n | % | ||||
| Entire groups | |||||||
| rs11191736 | C/C | 454 | 99.3 | 438 | 98.6 | 2.07 (0.52-8.34) | 0.19D |
| C/T | 3 | 0.7 | 6 | 1.4 | |||
| T/T | 0 | 0.0 | 0 | 0.0 | |||
| T | 3 | 0.3 | 6 | 0.7 | 2.07 (0.52-8.29) | 0.26 | |
| rs34040810 | C/C | 465 | 98.9 | 466 | 98.3 | 1.60 (0.52-4.92) | 0.29A |
| C/A | 5 | 1.1 | 8 | 1.7 | |||
| A/A | 0 | 0.0 | 0 | 0.0 | |||
| A | 5 | 0.5 | 8 | 0.8 | 1.59 (0.52-4.88) | 0.45 | |
| rs2289964 | C/C | 360 | 76.6 | 365 | 77.5 | 0.95 (0.70-1.29) | 0.78D |
| C/T | 103 | 21.9 | 99 | 21.0 | |||
| T/T | 7 | 1.5 | 7 | 1.5 | |||
| T | 117 | 12.4 | 113 | 12.0 | 0.96 (0.73-1.26) | 0.86 | |
| rs11191979 | T/T | 236 | 51.3 | 248 | 52.8 | 0.94 (0.73-1.22) | 0.53D |
| T/C | 192 | 41.7 | 183 | 38.9 | |||
| C/C | 32 | 7.0 | 39 | 8.3 | |||
| C | 256 | 27.8 | 261 | 27.8 | 1.00 (0.81-1.22) | 0.59 | |
| rs187304410 | G/G | 451 | 96.0 | 459 | 96.8 | 0.78 (0.39-1.55) | 0.31A |
| G/A | 19 | 4.0 | 15 | 3.2 | |||
| A/A | 0 | 0.0 | 0 | 0.0 | |||
| A | 19 | 2.0 | 15 | 1.6 | 0.78 (0.39-1.54) | 0.86 | |
| Males | |||||||
| rs11191736 | C/C | 226 | 100.0 | 235 | 97.5 | NA | NA |
| C/T | 0 | 0.0 | 6 | 2.5 | |||
| T/T | 0 | 0.0 | 0 | 0.0 | |||
| T | 0 | 0.0 | 6 | 1.2 | NA | 0.017 | |
| rs34040810 | C/C | 231 | 98.7 | 245 | 97.2 | 2.20 (0.56-8.6) | 0.39A |
| C/A | 3 | 1.3 | 7 | 2.8 | |||
| A/A | 0 | 0.0 | 0 | 0.0 | |||
| A | 3 | 0.6 | 7 | 1.4 | 2.18 (0.56-8.49) | 0.34 | |
| rs2289964 | C/C | 185 | 79.1 | 192 | 76.2 | 1.18 (0.77-1.81) | 0.55D |
| C/T | 42 | 17.9 | 54 | 21.4 | |||
| T/T | 7 | 3.0 | 6 | 2.4 | |||
| T | 56 | 12.0 | 66 | 13.1 | 1.11 (0.76-1.62) | 0.78 | |
| rs11191979 | T/T | 111 | 48.5 | 129 | 51.6 | 0.88 (0.62-1.26) | 0.52D |
| T/C | 100 | 43.7 | 98 | 39.2 | |||
| C/C | 18 | 7.9 | 23 | 9.2 | |||
| C | 136 | 29.7 | 144 | 28.8 | 0.96 (0.72-1.27) | 0.71 | |
| rs187304410 | G/G | 226 | 96.6 | 239 | 94.8 | 1.54 (0.63-3.78) | 0.29D |
| G/A | 8 | 3.4 | 13 | 5.2 | |||
| A/A | 0 | 0.0 | 0 | 0.0 | |||
| A | 8 | 1.7 | 13 | 2.6 | 1.52 (0.63-3.71) | 0.46 | |
| Females | |||||||
| rs11191736 | C/C | 228 | 98.7 | 203 | 100.0 | NA | NA |
| C/T | 3 | 1.3 | 0 | 0.0 | |||
| T/T | 0 | 0.0 | 0 | 0.0 | |||
| T | 3 | 0.6 | 0 | 0.0 | NA | 0.25 | |
| rs34040810 | C/C | 234 | 99.2 | 221 | 99.5 | 0.53 (0.05-5.88) | 0.55D |
| C/A | 2 | 0.8 | 1 | 0.5 | |||
| A/A | 0 | 0.0 | 0 | 0.0 | |||
| A | 2 | 0.4 | 1 | 0.2 | 0.53 (0.05-5.87) | 0.86 | |
| rs2289964 | C/C | 175 | 74.2 | 173 | 79.0 | 0.76 (0.49-1.18) | 0.29D |
| C/T | 61 | 25.8 | 45 | 20.5 | |||
| T/T | 0 | 0.0 | 1 | 0.5 | |||
| T | 61 | 12.9 | 47 | 10.7 | 0.81 (0.54-1.21) | 0.33 | |
| rs11191979 | T/T | 125 | 54.1 | 119 | 54.1 | 1.03 (0.77-1.39) | 0.86A |
| T/C | 92 | 39.8 | 85 | 38.6 | |||
| C/C | 14 | 6.1 | 16 | 7.3 | |||
| C | 120 | 26.0 | 117 | 26.6 | 1.03 (0.77-1.39) | 0.86 | |
| rs187304410 | G/G | 225 | 95.3 | 220 | 99.1 | 0.19 (0.04-0.85) | 0.012D |
| G/A | 11 | 4.7 | 2 | 0.9 | |||
| A/A | 0 | 0.0 | 0 | 0.0 | |||
| A | 11 | 2.3 | 2 | 0.5 | 0.19 (0.04-0.86) | 0.02 | |
* The table shows the best genetic models for SNP-disease associations. 1 Absolute number and percentage of individuals and chromosomes with a particular genotype and allele, respectively. 2 Odds ratio with 95% confidence intervals (crude analysis) estimated for the best SNP-disease association model. 3 P-value estimated for the best SNP-disease association model through adaptive permutations. Superscripts denote the models: R, recessive; D, dominant; A, additive. Bold depicts statistically significant P-values and Odds Ratios. NA, not available.
Table 3.
Haplotypes of the GSTO1 gene and their association with psoriasis risk in the entire and sex-stratified groups.
Table 3.
Haplotypes of the GSTO1 gene and their association with psoriasis risk in the entire and sex-stratified groups.
| Haplotypes | SNP | Patients with psoriasis | Healthy Controls | Chi Square | P-value | ||||
| rs11191736 | rs34040810 | rs2289964 | rs11191979 | rs187304410 | |||||
| Entire gropus | |||||||||
| H1 | C | C | C | T | G | 0.671 | 0.672 | 0.005 | 0.999 |
| H2 | C | C | C | C | G | 0.200 | 0.190 | 0.314 | 0.972 |
| H3 | C | C | T | C | G | 0.076 | 0.085 | 0.589 | 0.951 |
| H4 | C | C | T | T | G | 0.032 | 0.030 | 0.070 | 0.993 |
| Rare | - | - | - | - | - | 0.021 | 0.023 | - | - |
| Males | |||||||||
| H1 | C | C | C | T | G | 0.645 | 0.657 | 0.118 | 1.000 |
| H2 | C | C | C | C | G | 0.208 | 0.215 | 0.097 | 1.000 |
| H3 | C | C | T | C | G | 0.076 | 0.084 | 0.089 | 1.000 |
| H4 | C | C | T | T | G | 0.036 | 0.026 | 0.436 | 0.953 |
| Rare | - | - | - | - | - | 0,035 | 0,018 | - | - |
| Females | |||||||||
| H1 | C | C | C | T | G | 0.701 | 0.687 | 0.233 | 1.000 |
| H2 | C | C | C | C | G | 0.187 | 0.165 | 0.733 | 1.000 |
| H3 | C | C | T | C | G | 0.077 | 0.089 | 0.438 | 1.000 |
| H4 | C | C | T | T | G | 0.028 | 0.032 | 0.094 | 1.000 |
| Rare | - | - | - | - | - | 0.007 | 0.027 | - | - |
Estimation of haplotype frequencies and significance of haplotype-disease associations was done using the Haploview software, v.4.2.
Table 4.
Summary of the number of mbmdr-models of SNP × risk factor interactions significantly associated with psoriasis risk.
Table 4.
Summary of the number of mbmdr-models of SNP × risk factor interactions significantly associated with psoriasis risk.
| Mbmdr-models of SNP × risk factors interactions | Entire groups1 | Males1 | Females1 | |||
| N | % | N | % | N | % | |
| Two-order models | N=12 | N=16 | N=13 | |||
| ALCOHOL × GSTO1 (1 SNP) | 5 | 41.7 | 5 | 31.3 | 5 | 38.5 |
| ALCOHOL × GCLC (1 SNP) | 6 | 50.0 | 6 | 37.5 | 6 | 46.2 |
| ALCOHOL × SMOKE | 1 | 8.3 | 1 | 6.3 | 1 | 7.7 |
| GSTO1 (1 SNP) × GCLC (1 SNP) | 0 | 0.0 | 4 | 25.0 | 0 | 0.0 |
| GSTO1 (1 SNP) × GSTO1 (1 SNP) | 0 | 0.0 | 0 | 0.0 | 1 | 7.7 |
| Three-order models | N=68 | N=83 | N=28 | |||
| ALCOHOL × GSTO1 (1 SNP) × GSTO1 (1 SNP) | 10 | 14.7 | 10 | 12.0 | 8 | 28.6 |
| ALCOHOL × GCLC (1 SNP) × GSTO1 (1 SNP)* | 30 | 44.1 | 45 | 54.2 | 8 | 28.6 |
| ALCOHOL × GCLC (1 SNP) × GCLC (1 SNP) | 15 | 22.1 | 15 | 18.1 | 1 | 3.6 |
| ALCOHOL × SMOKE × GSTO1 (1 SNP) | 5 | 7.4 | 5 | 6.0 | 5 | 17.9 |
| ALCOHOL × SMOKE × GCLC (1 SNP) | 6 | 8.8 | 6 | 7.2 | 6 | 21.4 |
| Other models | 2 | 2.9 | 2 | 2.4 | 0 | 0.0 |
| Four-order models | N=239 | N=148 | N=19 | |||
| ALCOHOL × GSTO1 (3 SNPs) | 10 | 4.2 | 9 | 6.1 | 0 | 0.0 |
| ALCOHOL × GSTO1 (2 SNPs) × GCLC (1 SNP)** | 60 | 25.1 | 53 | 35.8 | 0 | 0.0 |
| ALCOHOL × GCLC (2 SNPs) × GSTO1 (1 SNP) | 95 | 39.7 | 6 | 4.1 | 1 | 5.3 |
| ALCOHOL × GCLC (3 SNPs) | 20 | 8.4 | 14 | 9.5 | 0 | 0.0 |
| ALCOHOL × SMOKE × GCLC (2 SNPs) | 10 | 4.2 | 12 | 8.1 | 1 | 5.3 |
| ALCOHOL × SMOKE × GCLC (1 SNP) × GSTO1 (1 SNP)** | 30 | 12.6 | 36 | 24.3 | 12 | 63.2 |
| ALCOHOL × SMOKE × GSTO1 (2 SNPs)* | 10 | 4.2 | 10 | 6.8 | 5 | 26.3 |
| Other models | 4 | 1.7 | 8 | 5.4 | 0 | 0.0 |
1 The absolute number and percentage of a specific mbmdr model within a study group. Asterisk indicates that the mbmdr models demonstrated quantitative differences between males and females. (*P<0.05, **P<0.01). ALCOHOL, alcohol abuse; SMOKE, cigarette smoking.
Table 5.
The best n-order mbmdr-models of SNP × risk factor interactions significantly associated with psoriasis risk in males.
Table 5.
The best n-order mbmdr-models of SNP × risk factor interactions significantly associated with psoriasis risk in males.
| Mbmdr-models of SNP × risk factors interactions | NH | β-H | WH | NL | β-L | WL | Pperm | |
| Two-order models | ||||||||
| 1 | ALCOHOL × GSTO1 rs34040810 | 2 | 0.322 | 34.87 | 1 | -0.322 | 34.87 | <0.0001 |
| 2 | ALCOHOL × GSTO1 rs11191736 | 1 | 0.299 | 26.12 | 1 | -0.313 | 30.68 | <0.0001 |
| 3 | ALCOHOL × GSTO1 rs187304410 | 1 | 0.300 | 27.14 | 1 | -0.292 | 29.57 | <0.0001 |
| 4 | ALCOHOL × GCLC rs648595 | 3 | 0.308 | 29.57 | 1 | -0.259 | 14.33 | <0.0001 |
| 5 | ALCOHOL × GCLC rs542914 | 3 | 0.308 | 29.57 | 2 | -0.183 | 14.68 | <0.0001 |
| Three-order models | ||||||||
| 1 | ALCOHOL × GCLC rs524553 × GSTO1 rs34040810 | 2 | 0.297 | 25.88 | 3 | -0.322 | 34.30 | <0.0001 |
| 2 | ALCOHOL × GCLC rs542914 × GSTO1 rs34040810 | 4 | 0.322 | 34.30 | 2 | -0.203 | 18.31 | <0.0001 |
| 3 | ALCOHOL × GSTO1 rs34040810 × GSTO1 rs11191736 | 1 | 0.299 | 26.12 | 1 | -0.323 | 33.91 | <0.0001 |
| 4 | ALCOHOL × GSTO1 rs11191979 × GSTO1 rs34040810 | 4 | 0.317 | 33.31 | 2 | -0.267 | 27.36 | <0.0001 |
| 5 | ALCOHOL × GSTO1 rs2289964 × GSTO1 rs34040810 | 3 | 0.316 | 32.45 | 1 | -0.172 | 12.53 | <0.0001 |
| Four-order models | ||||||||
| 1 | ALCOHOL × GCLC rs648595 × GCLC rs542914 × GSTO1 rs11191979 | 4 | 0.301 | 16.69 | 6 | -0.303 | 38.90 | <0.0001 |
| 2 | ALCOHOL × GCLC rs524553 × GSTO1 rs34040810 × GSTO1 rs11191736 | 2 | 0.288 | 22.76 | 3 | -0.322 | 33.34 | <0.0001 |
| 3 | ALCOHOL × GCLC rs542914 × GSTO1 rs2289964 × GSTO1 rs34040810 | 5 | 0.313 | 29.13 | 3 | -0.167 | 11.98 | <0.0001 |
| 4 | ALCOHOL × GSTO1 rs187304410 × GSTO1 rs34040810 × GSTO1 rs11191736 | 1 | 0.292 | 23.98 | 1 | -0.292 | 28.65 | <0.0001 |
| 5 | ALCOHOL × SMOKE × GCLC rs542914 × GSTO1 rs11191979 | 5 | 0.291 | 26.44 | 4 | -0.279 | 28.03 | <0.0001 |
NH—number of interacting high-risk variables (SNP and risk factors); β-H—regression coefficient for high-risk interactions identified in step 2 of the analysis. WH—Wald statistics for high-risk interactions; NL is the number of interacting low-risk variables; β-L—regression coefficient for low-risk interactions identified in step 2 of the analysis; WL—Wald statistics for low-risk interactions. Pperm—permutation significance levels for the models associated with psoriasis risk.
Table 6.
The best n-order mbmdr-models of SNP × risk factor interactions significantly associated with psoriasis risk in females.
Table 6.
The best n-order mbmdr-models of SNP × risk factor interactions significantly associated with psoriasis risk in females.
| Mbmdr-models of SNP × risk factors interactions | NH | β-H | WH | NL | β-L | WL | Pperm | |
| Two-order models | ||||||||
| 1 | ALCOHOL × SMOKE | 3 | 0.285 | 22.22 | 1 | -0.285 | 22.22 | <0.0001 |
| 2 | ALCOHOL × GSTO1 rs187304410 | 1 | 0.319 | 14.02 | 2 | -0.319 | 14.02 | <0.0001 |
| 3 | ALCOHOL × GSTO1 rs34040810 | 1 | 0.317 | 13.42 | 1 | -0.288 | 11.62 | <0.0001 |
| 4 | ALCOHOL × GSTO1 rs11191736 | 1 | 0.327 | 12.65 | 1 | -0.291 | 10.29 | <0.0001 |
| 5 | ALCOHOL × GSTO1 rs2289964 | 2 | 0.323 | 14.27 | 1 | -0.159 | 6.21 | <0.0001 |
| Three-order models | ||||||||
| 1 | ALCOHOL × SMOKE × GSTO1 rs2289964 | 3 | 0.263 | 15.51 | 2 | -0.289 | 22.82 | <0.0001 |
| 2 | ALCOHOL × SMOKE × GSTO1 rs187304410 | 3 | 0.285 | 22.22 | 2 | -0.285 | 22.22 | <0.0001 |
| 3 | ALCOHOL × SMOKE × GSTO1 rs34040810 | 3 | 0.282 | 21.52 | 1 | -0.270 | 20.04 | <0.0001 |
| 4 | ALCOHOL × SMOKE × GCLC rs17883901 | 3 | 0.261 | 16.24 | 2 | -0.187 | 10.81 | <0.0001 |
| 5 | ALCOHOL × SMOKE × GCLC rs542914 | 3 | 0.320 | 14.56 | 1 | -0.236 | 8.39 | <0.0001 |
| Four-order models | ||||||||
| 1 | ALCOHOL × SMOKE × GCLC rs2397147 × GSTO1 rs2289964 | 2 | 0.292 | 8.24 | 3 | -0.238 | 22.18 | <0.0001 |
| 2 | ALCOHOL × SMOKE × GSTO1 rs187304410 × GSTO1 rs34040810 | 3 | 0.282 | 21.52 | 2 | -0.270 | 20.04 | <0.0001 |
| 3 | ALCOHOL × SMOKE × GCLC rs6933870 × GSTO1 rs187304410 | 4 | 0.342 | 21.24 | 2 | -0.166 | 9.87 | <0.0001 |
| 4 | ALCOHOL × SMOKE × GSTO1 rs2289964 × GSTO1 rs34040810 | 3 | 0.263 | 15.51 | 2 | -0.274 | 20.61 | <0.0001 |
| 5 | ALCOHOL × SMOKE × GCLC rs6933870 × GSTO1 rs34040810 | 4 | 0.340 | 20.56 | 1 | -0.165 | 9.84 | <0.0001 |
NH—number of interacting high-risk variables (SNP and risk factors); β-H—regression coefficient for high-risk interactions identified in step 2 of the analysis. WH—Wald statistics for high-risk interactions; NL is the number of interacting low-risk variables; β-L—regression coefficient for low-risk interactions identified in step 2 of the analysis; WL—Wald statistics for low-risk interactions. Pperm—permutation significance levels for the models associated with psoriasis risk.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



