
Submitted:
18 December 2024
Posted:
19 December 2024
You are already at the latest version
Abstract
Africa – like the rest of the world – periodically experiences drought, which may become further exacerbated because of climate change, leading to severe impacts on crop production and food security in the region. The genus Tylosema consists of five distinct species, namely T. humifusum, T. argentum, T. angolense, T. esculentum, and T. fassoglense. Tylosema fassoglense (Schweinf.) Torre & Hillc. has the widest distribution across Africa in diverse habitats, while Tylosema esculentum (Burch.) A. Schreib. is limited to arid regions in southern Africa. T. fassoglense holds unique nutritional composition and bioactive properties, and thus could offer unique uses that are not found in T. esculentum. Consequently, enhancing drought tolerance in T. fassoglense can contribute to food security and sustain its medicinal applications and thus mitigate the impacts of climate change on vulnerable populations in the region. The present study aimed at comparing the physiological and biochemical responses between the two species to elucidate the underlying mechanisms of differences in drought tolerance between T. esculentum and T. fassoglense. We compared water status, growth performance, oxidative stress levels as well as the responses of selected antioxidant components under water deficit in the two species. Our results showed that the water deficit caused higher reductions in leaf water potential and relative water content in T. fassoglense than in T. esculentum, which translated to a higher loss of leaf and root fresh weights in T. fassoglense than in T. esculentum. A higher increase in free proline content, hydrogen peroxide content and lipid peroxidation level were observed in T. fassoglense when compared to T. esculentum. Distinct differences were observed in the antioxidant enzyme activities for superoxide dismutase, catalase, ascorbate peroxidase, and glutathione reductase between these two species in response to water deficit. The observed differences in antioxidant enzyme responses demonstrate their role in the differences in water deficit tolerance between the two species. Further detailed ‘omics’-based analyses will aid in understanding the molecular basis of drought responses in these two species and potentially towards improving drought tolerance in T. fassoglense.
Keywords:
antioxidant enzyme
; drought
; genus Tylosema
; marama
; proline
1. Introduction
Drought is one of the most significant abiotic stress factors affecting crop production and water resources in South Africa [1]. Prolonged exposure of plants to drought leads to increased cellular levels of reactive oxygen species (ROS), which can be deleterious to cells if they accumulate beyond appropriate thresholds [2,3]. To abate oxidative stress, plants mobilize an array of antioxidant defense systems [4]. Tolerance to oxidative stress is mostly associated with increased antioxidant activity. Several studies have investigated the response of the plant antioxidant system to drought stress. However, such studies have mostly focused on common bean, soybean, cowpea, and mung bean for cultivated grain legumes [5,6,7,8,9,10]. The only study that investigated drought responses in the genus Tylosema was limited to the evaluation of physiological responses [11].
Leguminous crops are beneficial to human nutrition because they serve as a good source of protein, dietary fiber, vitamins, and minerals [12]. Marama bean (genus Tylosema) grows in the semi-arid tropical/sub-tropical regions of Southern and Eastern Africa [13]. Amongst the five different species of Tylosema that are endemic to sub-Saharan Africa [14], Tylosema esculentum (Burch.) A. Schreib. and Tylosema fassoglense (Schweinf). Torre & Hillc. are the predominant Tylosema species in South Africa [14,15,16,17,18,19]. These species grow in diverse soil types, some of which are sandy and have limited water-holding capacity, and they can withstand drought far better than most of the currently cultivated legumes [11]. Although T. esculentum grows in more arid environments compared to the tropical and subtropical climates favored by T. fassoglense, the latter has a wider range of distribution across diverse habitats and may thrive in regions or environments where T. esculentum struggles to adapt. T. fassoglense occurs in eastern and central tropical Africa, from Sudan and Ethiopia southwards to Angola and Namibia and the north-eastern part of South Africa, while T. esculentum is restricted to southern Africa [14]. T. esculentum mostly grows in sandy plains with altitudes ranging from 900 to 1100 m, while T. fassoglense can thrive in diverse ecosystems including open woodlands, forest edges, cultivated fields, clay, sandy or rocky soils, and occasionally in periodically flooded areas, occurring at altitudes ranging from 30 to 2100 m [14]. Marama bean pods produce one to four seeds per pod, with varying color and shape. These edible seeds are round and brownish-black or flattened and chestnut-brown in T. esculentum and T. fassoglense, respectively [17,20,21]. In addition to their nutritional importance, these plants also have medicinal properties [22]. Although marama beans have nutritional and medicinal importance, they are not recognized as a staple food because it is not cultivated but rather found in the wild and only consumed by a small percentage of the population in the regions of Africa where it grows. If domesticated and properly maintained, marama bean could be a key drought resilient crop that could assist in addressing the impacts of climate change on food security. A comparative study on nutrient composition indicated that T. fassoglense possesses higher mineral levels and protein content compared to T. esculentum [23]. Distinct medicinal uses of these two species were reported. For example, in southern Africa, local communities use T. esculentum as medicine to treat asthma and diarrhea [24,25]. T. fassoglense is used to improve postpartum recovery, and to treat hypertension related diseases and impotence [25,26]. Local communities in Kenya use the boiled tuber of T. fassoglense to treat infertility in women, epilepsy, renal disease, and cancer [27]. In Uganda, it was also reported that extracts from T. fassoglense tuber are used as a treatment for various types of cancer [28]. T. fassoglense could offer unique nutritional and medicinal benefits that may not be found in T. esculentum. Therefore, improving drought tolerance in T. fassoglense is crucial for unlocking its full potential to ensure sustainable utilization of its nutritional and medicinal properties, contributing to enhanced food security and human health. On these bases, our study aimed at investigating differences in the physiological and biochemical responses between T. esculentum and T. fassoglense to water deficit stress. Understanding these responses in the two species will provide valuable insights towards improving drought tolerance in T. fassoglense and potentially other leguminous crops.
2. Materials and Methods
2.1. Plant growth and treatment
Seeds of T. esculentum (Burch.) A. Schreib. and T. fassoglense (Schweinf). Torre & Hillc. were obtained from the wild in South Africa near Tosca for T. esculentum (-25°49'37.5"S 23°42'30.3"E) and near Tzaneen for T. fassoglense (-23°52'12.8"S 30°06'05.8"E). The seeds were stored at room temperature at the Plant Stress Tolerance Laboratory at the University of the Western Cape until further use. The seeds were surface sterilized using chlorine gas and germinated as described by [29]. Soil was prepared by moistening Promix Organic with tap water, followed by supplementation with nutrients as described by [29]. The initial water potential of the soil was adjusted to − 0.3 ± 0.025 MPa for all pots. Germinated seeds were transplanted 5 cm deep into the soil (6 liters of soil in pots with diameter of 25 cm).
The pots were divided such that twelve were for well-watered (WW) plants and 12 were for plants that were to be subjected to water deficit (WD). The transplanted seeds were allowed to grow by placing the pots in a greenhouse at the University of the Western Cape (-33.930917, 18.624778; i.e. 33°55'51.3"S 18°37'29.2"E); wherein the greenhouse was set to remain at a temperature of 29 °C for 16 hours (during the day) and a temperature of 18 °C during the night (8 hours dark period). Lighting during the day was supplemented with high pressure sodium lamps providing a photon flux density of 600 µmol m-2 s-1. Soil for WW plants was kept moist by irrigating the soil with 300 ml of tap water once a week (every Friday at midday) until the time of harvesting. The soil for WD plants did not receive water until the plants were harvested. WW and WD plants from both species were harvested 42 days after transplanting the germinated seeds into the soil, which was the time when reduction in leaf tissue turgor (apparent partial flaccidity of some of the leaves) was observed in some of the WD plants of both species.
2.2. Physiological and biochemical analyses
2.2.1. Soil water potential
The WP4C water potentiometer (Meter Group, Pullman, WA, USA) was used to measure soil water potential at the day of harvesting for the WW and the WD plants. Water potential measurement in the soil were done according to the instructions in the operating manual of the WP4C instrument in precise mode. Soil samples were selected randomly from WW and WD plants in triplicates for each Tylosema species for the measurements on the WP4C.
2.2.2. Leaf water potential
Leaf water potential was measured at midday on the day of harvesting in three of each of the WW and WD plants from each of the Tylosema species, using the Model 1505D Pressure Chamber Instrument (PMS Instruments, Albany, OR, USA), according to the instructions provided in the manual for the pressure chamber. For the leaf water potential, three biological replicates of the second youngest leaf from each treatment were harvested and used in the pressure chamber instrument.
2.2.3. Measurements of shoot and root weights
The WW and WD plants of each of the two Tylosema species were used to measure the fresh weights of roots and shoots of the plants by weighing them on a scale (Kern® PLE 4200-2N precision scale, Kern and Sohn GmbH, Balingen, Germany).
2.2.4. Relative water content
The relative water content of the leaves of WW and WD T. esculentum and T. fassoglense (three biological replicates for each) was determined based on the formula:
The remaining leaf material (the first, third, fourth, and fifth youngest leaves) and roots of each plant were harvested, ground in liquid nitrogen and stored as a fine powder at -80 °C until further use.
2.2.5. Proline content
Free proline content was measured in extracts from ground leaf material in three biological replicates. Each biological replicate was generated by combining leaf material from 4 plants of each species under each treatment. The procedure described by Ali et al. [30] was used for the spectrophotometry-based measurement of free proline.
2.2.6. Hydrogen peroxide and malondialdehyde contents
Hydrogen peroxide (H2O2) content was measured from extracts from the same leaf material used for the proline measurements. The procedure used for H2O2 measurements was performed as described by Ali and Ludidi [31]. The same extract used for H2O2 measurement was used to measure the malondialdehyde content in the leaves, which is indicative of the extent of leaf lipid peroxidation, as described by Ali and Ludidi [31].
2.2.7. Antioxidant enzyme activities
The activities of superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT) and glutathione reductase (GR) enzymes were measured in protein extracts in triplicates using the spectrophotometric procedures described by Ali and Ludidi [31]. Each of the root samples used in these measurements were a pool from 4 plants from each species under each treatment. In these spectrophotometric measurements, the SOD assay was based on Beauchamp and Fridovich [32], APX activity was measured based on Nakano and Asada [33], the method followed for CAT activity was adapted from Lück [34], whereas the GR measurement was based on the method of Foyer and Halliwell [35].
Characterization of the responses of each of the antioxidant enzyme isoforms to water deficit was assessed based on changes in the intensity of bands in the images of gels obtained after native polyacrylamide gel electrophoresis (PAGE). The pixel intensities of the isoforms were calculated using AlphaEase 4.0 (Alpha Innotech Corporation, San Leonardo, CA). The native PAGE for SOD was conducted as described by Beauchamp and Fridovich [32], APX activity was assessed using the method of Seckin et al. [36], the occurrence of the CAT isoforms was detected based on Yamashita et al. [37], and GR isoforms were identified following the method of Rao et al. [38].
2.2.7. Statistical analysis
Results from each of the experiments described were analyzed using GraphPad Prism 6.0 (GraphPad Software, Boston, MA) using one-way analysis of variance (ANOVA), with the level of significance at 5% determined based on the built-in Tukey-Kramer test.
To explore the relationship between H2O2 contents, MDA levels and antioxidant enzyme activates in the two Tylosema species under water deficit stress, an R script was used to investigate the correlation between these parameters. The fold change of each parameter was calculated for both T. esculentum and T. fassoglense to determine the changes of these parameters within the two species. Subsequently, a fold change ratio for each parameter was computed by dividing the fold change of T. esculentum by that of T. fassoglense. The fold change ratios indicate how the studied parameters differ between the two species in response to water deficit stress. These fold change ratios were used to determine the correlation between the studied parameters using Pearson correlation coefficient.
3. Results
3.1. Plant water status and growth
Limitation of water supply to the soil mixture reduced soil water potential to equivalent levels in the soil in which the two Tylosema species were grown. At the time of harvesting, the soil water potential for the WW plants corresponded to approximately -0.24 MPa for T. esculentum and -0.26 MPa for T. fassoglense. However, for the WD plants, the soil water potential dropped to -0.83 and -0.81 MPa for T. esculentum and T. fassoglense plants, respectively (Figure 1A). Soil water deficit resulted in a reduction in the water potential of the leaves of both species, but the reduction in the leaf water potential in T. fassoglense was more prominent than in T. esculentum. The leaf water potential decreased from -0.66 to -1.1 MPa (1.6-fold) in response to water deficit in T. esculentum whereas it decreased from -0.65 to -1.5 MPa (2.3-fold) in T. fassoglense in response to water deficit (Figure 1B).
The decreased leaf water potential was accompanied by reduced shoot fresh weight in both Tylosema species (Figure 1C) and a reduced root fresh weight only for T. fassoglense (Figure 1D) in response to WD. The decrease in shoot fresh weight was more pronounced in T. fassoglense (1.5-fold, from 7.6 to 5.1 g/plant) than in T. esculentum (1.3-fold, from 4.8 to 3.7 g/plant) in response to water deficit (Figure 1C). Water deficit decreased root fresh weight by 1.5-fold (5.3 to 3.6 g/plant) in T. fassoglense, whereas no significant change was observed for T. esculentum root fresh weight between WD and WW conditions (Figure 1D).
WD reduced the relative water content from 96 % to 85 % (1.1-fold) in the leaves of T. esculentum, whereas the water deficit-induced decrease in T. fassoglense was from 96 % to 75 % (1.3-fold) (Figure 2A). These changes in leaf relative water content were accompanied by corresponding changes in leaf free proline content, in which a more pronounced increase in proline content occurred for T. fassoglense (3.3-fold, from 9.6 to 31.3 mmol.mg-1 FW) than for T. esculentum (1.3-fold, from 9.4 to 12.3 mmol.mg-1 FW) in response to WD (Figure 2B).
3.2. Oxidative stress and antioxidant responses
Water deficit (WD) did not significantly affect leaf hydrogen peroxide (H2O2) content in T. esculentum, whereas it caused an increase of 1.7-fold (299 to 518 nmol.g-1 FW) in H2O2 content in the leaves of T. fassoglense (Figure 3A). The H2O2 response corresponded to similar changes in malondialdehyde (MDA) content, where no change was triggered by water deficit in T. esculentum but an increase of 1.2-fold (89.7 to 104 nmol.g-1 FW) occurred in T. fassoglense in response to WD (Figure 3B).
Superoxide dismutase (SOD) activity increased by 1.5-fold (1.5 to 2.3 units.mg-1 protein) in the leaves of T. esculentum, while only 1.2-fold increase (2.5 to 3.1 units.mg-1 protein) was observed in T. fassoglense in response to WD (Figure 4A). For roots, WD increased the superoxide dismutase activity in T. esculentum by 4.5-fold (0.3 to 1.1 units.mg-1 protein) but had no significant effect on the superoxide dismutase activity of T. fassoglense roots (Figure 4B). Leaf tissue showed a total of six superoxide dismutase isoforms in both T. esculentum and T. fassoglense, with noticeable increase in the intensities of SOD 2 (1.3-fold, from 7.3 to 9.3 arbitrary units) and SOD 3 (1.3-fold, 7.7 to 10.3 arbitrary units) in response to WD in T. esculentum, whereas a noticeable increase for SOD 1 (1.7-fold, from 6.3 to 10.7 arbitrary units), SOD 2 (1.2-fold, from 9.3 to 11.3 arbitrary units) and SOD 6 (1.2-fold, from 9.3 to 10.7 arbitrary units) were observed for T. fassoglense (Figure 4C, Supplementary Table 1). Roots of both species showed a total of two SOD isoforms, namely SOD 3 and SOD 4 (Figure 4C, Supplementary Table 1). No SOD isoforms were visible for T. esculentum under well-watered conditions in roots, but WD induced the appearance of SOD 3 by 2.2-fold (5.7 to 12.3 arbitrary units) and SOD 4 by 1.4-fold (5.7 to 7.7 arbitrary units) in the roots of this species (Figure 4C, Supplementary Table 1). No clear change in the superoxide dismutase isoforms occurred in roots of T. fassoglense in response to WD (Figure 4C, Supplementary Table 1).
Catalase (CAT) activity increased in the leaves and roots of both species, whereby the leaf CAT activity for T. esculentum increased by 2-fold (75 to 150 µmol.min-1.mg-1 protein) and that for T. fassoglense increased by 2.7-fold (45 to 102 µmol.min-1.mg-1 protein) (Figure 5A), whereas the root CAT activity showed an increase of 1.8-fold (6.7 to 12.2 µmol.min-1.mg-1 protein) and 2.6-fold (3.5 to 8.6 µmol.min-1.mg-1 protein) in T. esculentum and T. fassoglense, respectively (Figure 5B). Two CAT isoforms were observed in the leaves of the two Tylosema species, wherein the water deficit increased CAT 1 and CAT 2 in both species (Figure 5C, Supplementary Table 2). T. fassoglense exhibited a higher intensity increase for these isoforms, with a 1.5-fold increase (41 to 61 arbitrary units) for CAT 1, and an increase of 1.4-fold (71 to 97.7 arbitrary units) for CAT 2. In contrast, T. esculentum, showed an increase of 1.3-fold (68.3 to 86.3 arbitrary units) for CAT 1 intensity and a 1.2-fold increase for CAT 2 (90.3 to 111 arbitrary units) intensity (Figure 5C, Supplementary Table 2). Three CAT isoforms were detected in the roots of the two species, with T. esculentum showing enhanced intensity of 1.3-fold for CAT 2 (6.3 to 8.3 arbitrary units) and 1.7-fold for CAT 3 (5.3 to 9.0 arbitrary units) in response to WD, while T. fassoglense showed an increased intensity only for CAT 2 (1.4-fold, from 5.7 to 7.7 arbitrary units). Root CAT activity was notably lower than leaf CAT activity, wherein CAT 3 and CAT 2 were the most prominent root isoforms in T. esculentum and T. fassoglense, respectively (Figure 5C, Supplementary Table 2).
Leaf ascorbate peroxidase (APX) activity exhibited a 1.2-fold increase (2.5 to 3.0 µmol.min-1.mg-1 protein) in T. esculentum and a 1.5-fold increase (2.4 to 3.6 µmol.min-1.mg-1 protein) in T. fassoglense (Figure 6A), whereas the increase was not significant in either T. esculentum or T. fassoglense roots (Figure 6B). Five APX isoforms (APX 1, 2, 3, 4, and 5) were observed in the leaves and roots of the two Tylosema species, with the most prominent isoform being APX 5, as the intensity of the rest of the isoforms was low, although APX 1 was also prominent in the roots of both species (Figure 6C, Supplementary Table 3).
The assessment of glutathione reductase (GR) activity revealed that WD augmented leaf GR activity only in T. fassoglense from 15 to 21 nmol.min-1.mg-1 protein (1.4-fold), but not in T. esculentum (Figure 7A). However, GR activity was increased in response to WD in the T. esculentum roots by 1.3-fold (34 to 45 nmol.min-1.mg-1 protein) but decreased by 1.7-fold (44 to 26 nmol.min-1.mg-1 protein) in the roots of T. fassoglense (Figure 7B). Five isoforms of GR (GR 1, 3, 5, 6, and 7) were observed in the leaves of the two Tylosema species and six (GR 1, 2, 3, 4, 5, and 6) were observed in their roots (Figure 7C). In the leaves, only the intensity (and thus activity) of GR 7 was increased in T. esculentum (1.2-fold, from 6.0 to 7.3 arbitrary units) in response to WD (Figure 7C, Supplementary Table 4). In T. fassoglense leaves, only GR 1 and GR 5 activities were up-regulated in response to WD, with an increase of 1.4-fold (6.7 to 9.7 arbitrary units) for GR1 and an increase of 1.3-fold (6.7 to 8.7 arbitrary units) for GR 5, whereas the rest of the isoforms were not affected (Figure 7C, Supplementary Table 4). GR 1, GR 2, GR 3, and GR 4 were increased by 1.2-fold, 1.6-fold, 1.3 and 1.4-fold (21 to 26, 14 to 23, 8.3 to 11 and 7.7 to 11, arbitrary units), respectively in T. esculentum roots in response to WD. The intensities of all GR isoforms were down-regulated in T. fassoglense roots, except for the T. fassoglense GR 5 isoform, which was induced by 1.3-fold (6.0 to 8.0 arbitrary units) under WD (Figure 7C, Supplementary Table 4).
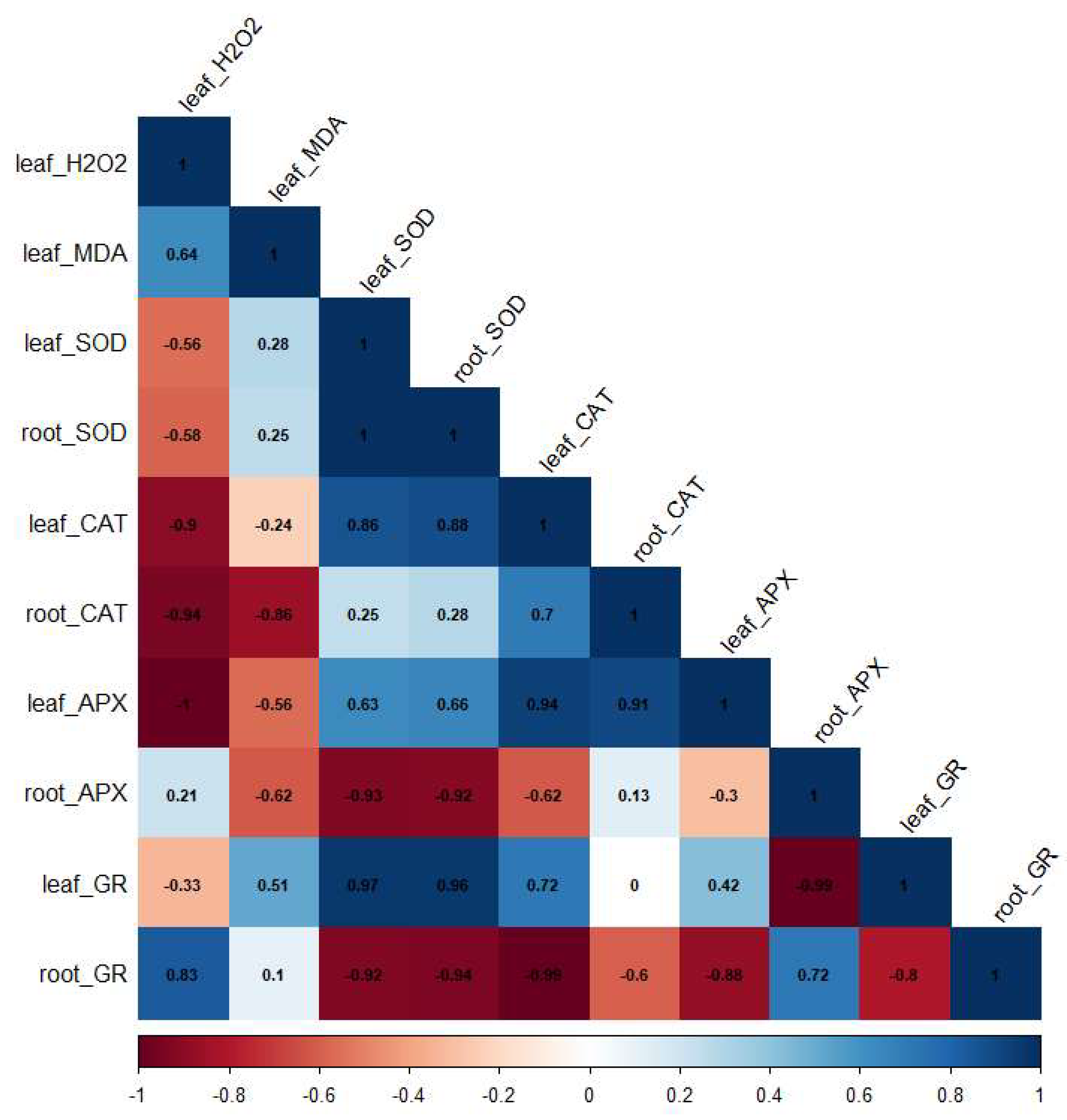
Pearson correlation analyses revealed relationships among various biochemical responses to water deficit in the two Tylosema species, as illustrated in Figure 8. A moderate positive correlation was observed between leaf H2O2 and MDA contents. H2O2 exhibited a moderate negative correlation with SOD activity in both leaf and root tissues. A strong negative correlation was observed between H2O2 and CAT activity in both leaves and roots. Notably, H2O2 exhibited a perfect negative correlation with APX activity in the leaves and a strong positive correlation with root GR activity.
4. Discussion
Members of the genus Tylosema are known for their exceptional drought tolerance, but the mechanisms of their adaptation to drought vary and appear to depend on the specific species and their natural habitat [16]. Several physiological responses of Tylosema esculentum to drought, high temperature and high radiation were reported by Mitchell et al. [11], and by Karamanos and Travlos [39]. However, very little is known about the comparative responses between T. esculentum and T. fassoglense under similar water deficit (WD) stress. In this study, we exposed T. esculentum and T. fassoglense to well-watered (WW) and WD treatments under greenhouse conditions. Following the treatment period, we observed no significant differences in the soil water potential of the WW soil in which T. esculentum and T. fassoglense were grown (approximately -0.25 MPa). When we imposed WD conditions, the soil water potential dropped to approximately -0.8 MPa for the treated Tylosema species. This result demonstrated that the two Tylosema species experienced the same level of soil WD conditions at the time of harvesting.
Tylosema plants exhibit adaptation to reduce transpiration-induced water loss through early stomatal closure [11]. In this study, we measured the leaf water potential of the two species under contrasting water regimes. The two species did not show any differences in leaf water potential when grown under WW conditions. However, both species showed reductions in leaf water potential when exposed to WD conditions. The reduction in leaf water potential was even higher in T. fassoglense than in T. esculentum. We thus conclude that T. esculentum maintains leaf water status better than T. fassoglense when both plants are exposed to soil WD. Karamanos and Travlos [39] observed similar leaf water potential levels when T. esculentum, collected from Botswana, were exposed to WD stress under greenhouse conditions. Karamanos and Travlos [39] concluded that leaf behavior plays a vital role in the regulation of the water status in T. esculentum under WD stress. The observed changes in leaf relative water content in the two Tylosema species indicate that T. esculentum has better leaf water retention capacity than T. fassoglense.
Maintaining a larger root biomass under drought stress is part of the adaptive responses of plants to cope with limited water availability [40]. Our results show that T. esculentum underwent less leaf biomass reduction than T. fassoglense, and that WD stress did not alter the root fresh weight of T. esculentum, whereas significant root weight reductions occurred in T. fassoglense. This suggests that the lower reduction in shoot biomass observed in T. esculentum could be attributed to its ability to maintain root biomass under WD stress. It was established that drought-tolerant plants can maintain their root length and root volume to improve their capacity to take up the limited water in the soil [41].
Proline often accumulates in plants under drought stress, wherein it functions as an osmoprotectant and ROS scavenger [42]. In our study, T. fassoglense leaves accumulated more proline than the T. esculentum leaves, which could indicate that the T. fassoglense leaves experienced higher WD stress levels than T. esculentum leaves. The higher accumulation of proline observed in T. fassoglense did not assist the plant to maintain leaf biomass, which suggests that differences in WD stress tolerance between these two species is not associated proline content. Lv et al. [43] reported similar findings where no correlation between proline content and stress tolerance was observed in 25 rice cultivars.
Hydrogen peroxide (H2O2) accumulates to high levels in drought-exposed plants and leads to overaccumulation of malonaldehyde (MDA), resulting in lipid membrane damage [44]. In our study, we observed no changes in H2O2 and MDA contents in T. esculentum leaves exposed to WD stress. However, we observed a significant increase in H2O2 and MDA in the leaves of T. fassoglense exposed to WD. We suggest that the increase in leaf H2O2 led to more membrane damage, as evidenced by the increased MDA levels in T. fassoglense under WD stress than in T. esculentum. From this result, we conclude that T. esculentum limits ROS accumulation better under WD than T. fassoglense, which leads to better WD stress tolerance in T. esculentum than in T. fassoglense. Keyster et al. [45] demonstrated that the capacity to control ROS content is a key determinant of sensitivity and tolerance of plants to stress. In this study, we measured various ROS-antioxidant enzymes in order to further understand the differential ROS and MDA accumulation in T. esculentum and T. fassoglense under WD. We measured the activities of superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), and glutathione reductase (GR), as both total enzymatic activity and isoform analysis by native-PAGE staining.
SOD regulates the dismutation of superoxide radicals into H2O2 in different plant organelles [46]. Therefore, SOD is a key enzyme in the regulation of intracellular ROS levels and maintains redox homeostasis under oxidative stress caused by drought [47]. In this study, in response to WD, both leaf and root total SOD enzymatic activity was increased in T. esculentum, while no change was observed in the root total SOD enzymatic activity of T. fassoglense, which indicates that T. fassoglense does not have sufficient SOD capacity in the roots to control excessive ROS accumulation under WD condition. Isoform analysis indicated that SOD 2 and SOD 3 contribute the most to the total SOD increase in T. esculentum leaves under WD, whereas SOD 1, SOD 2 and SOD 6 account for the total SOD increase in T. fassoglense leaves. Furthermore, in T. esculentum roots, the increase in total SOD activity is mostly regulated by SOD 3 and SOD 4. One the other hand, these isoforms remained unchanged in T. fassoglense roots. This implies that the regulation of SOD isoform activity differs between T. esculentum and T. fassoglense under WD. Huseynova et al. [48] concluded that the differential regulation of a Mn-SOD in wheat cultivars contributed to differential sensitivity to drought stress. Wang et al. [49] observed similar results when exposing two rice cultivars to drought stress conditions where the drought-tolerant cultivar had a higher SOD activity than the drought-sensitive one. This observation was also reported by Singh et al. [50], who showed that drought-tolerant cotton cultivars had a higher SOD activity than cotton cultivars that are sensitive to drought stress.
CAT and APX have contrasting affinities for H2O2, indicating the existence of complementary metabolic roles of these two antioxidant enzymes in plants [51]. Corpas [52] proposed that APX may be an important enzyme for fine-tuning and scavenging low concentrations of H2O2, whereas CAT seems to be a key scavenger enzyme when H2O2 concentrations are high. Our results showed that although WD led to a higher increase in leaf and root total CAT enzymatic activity in T. fassoglense than in T. esculentum, the overall activity was higher in T. esculentum under both water conditions. This suggests that T. esculentum possesses a higher basal capacity for oxidative stress management, potentially contributing to its better WD tolerance. In addition, we observed a more pronounced increase in the CAT 2 and CAT 3 isoforms in T. esculentum compared to T. fassoglense leaves under WD. Therefore, our results suggest that these two CAT isoforms are important in regulating the level of H2O2; and the capacity of T. esculentum in increasing the activity of these isoforms under WD conditions is likely to contribute to its drought tolerance. These two isoforms may have distinct substrate affinities or subcellular localizations, allowing them to efficiently scavenge H2O2, highlighting the complex interplay between total antioxidant enzyme activity, isoform-specific activity and ROS regulation. Our APX data showed a higher increase in total leaf APX activity in T. fassoglense compared to T. esculentum. However, we noticed no significant difference in the total root APX activity of both species in response to WD. We hypothesize that leaf APX activity is much more important than root APX activity in H2O2 regulation under WD stress in Tylosema species. We also suggest that the regulation of APX 5 activity is important for WD responses because T. esculentum experienced a much lower decrease in APX 5 activity in roots under WD stress, which could also be correlated to its better H2O2 detoxification capacity. These results suggest that H2O2 scavenging capacity differs in the two Tylosema species under WD stress. Chugh et al. [53] observed that differences in CAT and APX activities correlated with the tolerance or sensitivity to drought stress in maize cultivars.
GR maintains the intracellular glutathione (GSH) pool in the reduced state [54]. GSH can function as a direct antioxidant (non-enzymatically) or an indirect antioxidant by recycling ascorbic acid [55]. Ahmad et al. [56] concluded that differential regulation of sulfur metabolism by GR in leaves and roots was associated with enhanced drought stress response in maize. In our study, total GR activity was increased in the leaves of T. fassoglense but no significant change was observed in T. esculentum in response to WD stress. Contrastingly, we observed an increase in the root GR activity in T. esculentum, while a decrease in GR activity was observed in the roots of T. fassoglense in response to WD stress. The GR isoform analysis indicated that GR 1 and GR 5 are the isoforms likely responsible for the increased GR activity in the leaves of T. fassoglense in response to WD. Furthermore, isoforms GR 1, GR 2, GR 3, and GR 4 may be responsible for the increase in the GR activity of T. esculentum roots in response to WD. We suggest that the increased GR activity in the roots of T. esculentum possibly contributed to its greater maintenance of root biomass and oxidative stress tolerance than T. fassoglense. Similar findings were reported by Khanna et al. [57]. These authors observed an increase in GR activity in the roots of a tolerant maize genotype in response to heat stress and a decrease in the activity in the leaves, while no significant change in GR activity was observed in both leaves and roots of the sensitive genotype. Zhang and Kirkham [58] observed that sorghum plants maintained a much higher increase in GR activity under drought stress when compared to sunflower.
The correlation coefficients provided insights into how the studied parameters are interconnected, contributing to a deeper understanding of the adaptive responses in the two Tylosema species to water deficit stress. The observed positive correlation between H2O2 and MDA contents suggests that the increase in H2O2 contributed to the increase in MDA content observed in the leaves of the two Tylosema species under water deficit stress. The negative correlation observed between H2O2 and CAT activity in both leaf and root tissues demonstrates the significance of CAT in mitigating excessive H2O2 levels in Tylosema species under water deficit. The perfect negative correlation of leaf APX activity with H2O2 further highlights the crucial role of leaf APX in the regulation of H2O2 in Tylosema species under water deficit stress. The positive correlation observed between GR activity in the roots and H2O2 contents could be linked to its key role in oxidative stress regulation via the regeneration of reduced glutathione (GSH), which gets oxidized by several pathways linked to H2O2 scavenging [59]. This correlation highlights the key role of GR activity in drought responses of Tylosema species and further indicates that the observed increase in GR activity in the roots of T. esculentum potentially contributes to its better efficiency in H2O2 detoxification than T. fassoglense.
5. Conclusions
This comparative study revealed that T. esculentum maintains greater water retention compared to T. fassoglense under the water deficit. Accumulation of proline under WD was not translated into enhanced water status in T. fassoglense, suggesting alternative adaptive mechanisms could govern water relations in these two Tylosema species. However, the higher water retention in T. esculentum could be attributed to its ability to maintain root biomass under WD. Furthermore, this study highlights the critical role of antioxidant enzyme activities in mediating tolerance to WD stress in the Tylosema species. Our findings indicate that T. esculentum demonstrated a higher SOD activity, which led to an effective control of ROS levels. The higher baseline of CAT total activity in both leaf and root tissues of T. esculentum plays an important role in its greater ability of oxidative stress management. The decrease in APX 5 activity was much lower in T. esculentum, indicating its potential role in mediating drought responses in the two species. Moreover, GR activity also exhibited differing patterns between the two species, with T. fassoglense showing an increase in GR activity in leaves but a decrease in roots, while T. esculentum displayed increased GR activity in the roots. This differing GR activity was associated with different levels of H2O2 detoxification, indicating that upregulation of GR activity, particularly in the roots, plays a key role against WD-induced oxidative damage in T. esculentum. These findings emphasize the complexity of the antioxidant defense system, suggesting that more finely tuned regulatory mechanisms may play a significant role in ROS detoxification under WD stress.
The present work offers a foundation for further research in elucidating the genetic basis of drought adaptation in the genus Tylosema. Understanding the molecular adaptive mechanisms underlying the extreme drought tolerance in T. esculentum and how they differ in T. fassoglense could offer potential avenues for enhancing drought tolerance in T. fassoglense and other leguminous crops. This enhancement of crop tolerance to drought is essential for sustainable agriculture and global food security.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Pixel intensities of SOD isoforms in the leaves (a) and roots (b) of T. esculentum and T. fassoglense as detected on native PAGE (Figure 4C); Table S2: Pixel intensities of CAT isoforms in the leaves (a) and roots (b) of T. esculentum and T. fassoglense as observed on native PAGE (Figure 5C); Table S3: Pixel intensities of APX isoforms in the leaves (a) and roots (b) of T. esculentum and T. fassoglense as detected on native PAGE (Figure 6C); Table S4: Pixel intensities of GR isoforms in the leaves (a) and roots (b) of T. esculentum and T. fassoglense as observed on native PAGE (Figure 7C).
Author Contributions
AEEA performed the protein extractions, Bradford assays, GR native PAGE, measured the pixel intensities presented in the supplementary tables, performed correlation analyses and measured the soil and leaf water potentials; EK, AM, IME, ANM, GB, MK, and NL collected the seeds of T. esculentum and T. fassoglense; MOA, GB, and AEEA measured the growth parameters and the leaf relative water content; GB performed the assays for H2O2, MDA, and proline contents; AEEA, ANM, and AM grew the plants; EK performed the CAT native PAGE; AMB performed the GR and CAT spectrophotometric assays; MOA performed the APX spectrophotometric assay; EK performed the catalase native PAGE; YB and AEEA performed the APX native PAGE; RM performed the SOD native PAGE; IME, UG, and NL contributed to the conceptualization of the project; NL designed the experiments; UG and NL raised funding. All authors contributed to the writing of the original draft of the manuscript, AEEA and NL revised the original draft to write the final version of the submitted manuscript.
Funding
This work was supported by research grant funding from the DSI-NRF Centre of Excellence in Food Security (Grant ID 22202 to NL) and a joint research grant from the Swiss National Science Foundation (310030L_204528 to UG) and the National Research Foundation South Africa Research Cooperation Programme (GUN 137753 to NL).
Data Availability Statement
All data are available upon request in addition to data provided in the supplementary material.
Acknowledgments
We thank the Department of Economic Development, Environment and Tourism in the province of Limpopo, and the Department of Economic Development, Environment, Conservation and Tourism of the province of the North West in South Africa for permitting us to collect seeds of T. fassoglense and T. esculentum, respectively.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Mathivha, F.I.; Mabala, L.; Matimolane, S.; Mbatha, N. El Niño-Induced Drought Impacts on Reservoir Water Resources in South Africa. Atmosphere 2024, 15, 249. [Google Scholar] [CrossRef]
- Grant, J.J.; Loake, G.J. Role of reactive oxygen intermediates and cognate redox signalling in disease resistance. Plant Physiol 2000, 124, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Z.; Zhou, Q.; Wang, X.; Song, S.; Dong, S. Physiological response of soybean plants to water deficit. Front. Plant Sci 2022, 12, 809692. [Google Scholar] [CrossRef] [PubMed]
- Arya, H.; Mohan, B.S.; Prem, L.B. Towards developing drought-smart soybeans. Front. Plant Sci 2021, 12, 750664. [Google Scholar] [CrossRef]
- Dong, S.; Jiang, Y.; Dong, Y.; Wang, L.; Wang, W.; Ma, Z.; Yan, C.; Ma, C.; Liu, L. A study on soybean responses to drought stress and rehydration. Saudi J. Biol. Sci 2019, 26, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- Morosan, M.; Hassan, M.; Naranjo, M.; López-Gresa, M.P.; Boscaiu, M.; Vicente, O. Comparative analysis of drought responses in Phaseolus vulgaris (common bean) and P. coccineus (runner bean) cultivars. The Euro. Biotech. J 2017, 1, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Alsamadany, H. Physiological, biochemical and molecular evaluation of mung bean genotypes for agronomical yield under drought and salinity stresses in the presence of humic acid. Saudi J. Biol. Sci 2022, 29, 103385. [Google Scholar] [CrossRef]
- Sedlar, A.; Kidrič, M.; Šuštar-Vozlič, J.; Pipan, B.; Zadražnik, T.; Meglič, V. Drought stress response in agricultural plants: A case study of common bean (Phaseolus vulgaris L.). In Drought-Detection and Solutions; Intech Open: London, UK, 2019. [Google Scholar] [CrossRef]
- Mitchell, R.A.C.; Keys, A.J.; Madgwick, P.J.; Parry, M.A.J.; Lawlor, D.W. Adaptation of photosynthesis in marama bean Tylosema esculentum (Burchell A. Schreiber) to a high temperature, high radiation, drought-prone environment. Plant Physiol. Biochem 2005, 43, 969–976. [Google Scholar] [CrossRef]
- Ebert, A.W. Potential of underutilized traditional vegetables and legume crops to contribute to food and nutritional security, income and more sustainable production systems. Sustainability 2014, 6, 319–335. [Google Scholar] [CrossRef]
- Dakora, F.D. Biogeographic distribution, nodulation and nutritional attributes of underutilized indigenous African legumes. Acta. Hortic 2013, 979, 53–64. [Google Scholar] [CrossRef]
- Castro, S.; Paulo, S.; Antonio, P.C.; Estrela, F. Systematic studies in Tylosema (Leguminosae). Bot. J. Linnean Soc 2005, 147, 99–115. [Google Scholar] [CrossRef]
- Omotayo, A.O.; Aremu, A.O. Marama bean Tylosema esculentum (Burch.) A. Schreib: an indigenous plant with potential for food, nutrition, and economic sustainability. Food Funct 2021, 12, 2389–2403. [Google Scholar] [CrossRef] [PubMed]
- Cullis, C.; Chimwamurombe, P.; Barker, N.; Kunert, K.; Vorster, J. Orphan legumes growing in dry environments: Marama bean as a case study. Front. Plant Sci 2018, 9, 1199. [Google Scholar] [CrossRef]
- Holse, M.; Husted, S.; Hansen, A. Chemical composition of Marama bean (Tylosema esculentum): A wild African bean with unexploited potential. J Food Compos. Anals 2010, 23, 648–657. [Google Scholar] [CrossRef]
- Nepolo, E.; Takundwa, M.; Chimwamurombe, P.M.; Cullis, C.A.; Kunert, K. A review of geographical distribution of marama bean [Tylosema esculentum (Burchell) Schreiber] and genetic diversity in the Namibian germplasm. Afr. J. Biotechnol 2009, 8, 2088–2093. [Google Scholar] [CrossRef]
- Powell, A.M. Marama bean (Tylosema esculentum, Fabaceae) seed crop in Texas. Econ. Bot 1987, 41, 216–220. [Google Scholar] [CrossRef]
- Li, J.; Cullis, C. The multipartite mitochondrial genome of Marama (Tylosema esculentum). Front. Plant Sci 2021, 12, 787443. [Google Scholar] [CrossRef]
- Brink, M.; Belay, G. Plant Resources of Tropical Africa 1. Cereals and Pulses. Fondation PROTA; Backhuys Publishers: Leiden, Netherlands, 2006. [Google Scholar]
- Anywar, G.; Kakudidi, E.; Byamukama, R.; Mukonzo, J.; Schubert, A.; Oryem-Origa, H. Medicinal plants used by traditional medicine practitioners to boost the immune system in people living with HIV/AIDS in Uganda. Eur. J. Integr. Med 2020, 35, 1876–3820. [Google Scholar] [CrossRef]
- Amonsou, E.; Taylor, J.; Minnaar, A. Microstructure of protein bodies in marama bean species. LWT-Food Science and Technology 2011, 44, 42–47. [Google Scholar] [CrossRef]
- Chingwaru, W.; Majinda, R.T.; Yeboah, S.O.; Jackson, J.C.; Kapewangolo, P.T.; Kandawa-Schulz, M.; Cencic, A. Tylosema esculentum (Marama) tuber and bean extracts are strong antiviral agents against rotavirus infection. Evidence-Based Complementary and Alternative Medicine 2011, 2011, 284795. [Google Scholar] [CrossRef] [PubMed]
- Mapesa, W.A.; Waweru, M.P.; Bukachi, F.; Wafula, K.D. Aqueous tuber extracts of Tylosema fassoglense (kotschy ex schweinf.) torre and hillc.(fabaceae). Possess significant in-vivo antidiarrheal activity and ex-vivo spasmolytic effect possibly mediated by modulation of nitrous oxide system, voltage-gated calcium channels, and muscarinic receptors. Front. Pharmacol 2021, 12, 636879. [Google Scholar] [CrossRef]
- Munialo, S.; Gasparatos, A.; Ludidi, N.; Ali, A.E.E.; Keyster, E.; Akanbi, M.O.; Emmambux, M.N. Systematic Review of the Agro-Ecological, Nutritional, and Medicinal Properties of the Neglected and Underutilized Plant Species Tylosema fassoglense. Sustainability 2024, 16, p6046. [Google Scholar] [CrossRef]
- Kigen, G.; Maritim, A.; Some, F.; Kibosia, J.; Rono, H.; Chepkwony, S.; Kipkore, W.; Wanjoh, B. Ethnopharmacological survey of the medicinal plants used in Tindiret, Nandi County, Kenya. Afr. J. Tradit. Complement. Altern. Med 2016, 13, 156–168. [Google Scholar] [CrossRef]
- Kudamba, A.; Kasolo, J.N.; Bbosa, G.S.; Lugaajju, A.; Wabinga, H.; Niyonzima, N.; Ocan, M.; Damani, A.M.; Kafeero, H.M.; Ssenku, J.E.; Alemu, S.O. Medicinal plants used in the management of cancers by residents in the Elgon Sub-Region, Uganda. BMC complement. med. Ther 2023, 23, 450. [Google Scholar] [CrossRef]
- Kolo, Z.; Majola, A.; Phillips, K.; Ali, A.E.E.; Sharp, R.E.; Ludidi, N. Water Deficit-Induced Changes in Phenolic Acid Content in Maize Leaves Is Associated with Altered Expression of Cinnamate 4-Hydroxylase and p-Coumaric Acid 3-Hydroxylase. Plants 2023, 12, 101. [Google Scholar] [CrossRef]
- Ali, A.E.E.; Husselmann, L.H.; Tabb, D.L.; Ludidi, N. Comparative Proteomics Analysis between Maize and Sorghum Uncovers Important Proteins and Metabolic Pathways Mediating Drought Tolerance. Life 2023, 13, 170. [Google Scholar] [CrossRef]
- Ali, A.E.E.; Ludidi, N. Antioxidant responses are associated with differences in drought tolerance between maize and sorghum. J. Oasis Agric. Sustain 2021, 3, pp.1–12. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem 1971, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; and Asada, K. . Purification of ascorbate peroxidase in spinach chloroplasts: its inactivation in ascorbate depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol 1987, 28, 131–140. [Google Scholar] [CrossRef]
- Lück, H. Catalase. In Method of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York and London, 1965; pp. 885–894. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Seckin, B.; Turkan, I.; Sekmen, A.H.; Ozdan, C. The role of antioxidant defense systems at differential salt tolerance of Hordeum marinum Huds. (Sea barley grass) and Hordeum vulgare L. (cultivated barley). Environ. Exp. Bot 2010, 69, 76–85. [Google Scholar] [CrossRef]
- Yamashita, K.; Shiozawa, A.; Banno, S.; Fukumori, F.; Ichiishi, A.; Kimura, M.; Fujimura, M. Involvement of OS-2 MAP kinase in regulation of the large-subunit catalases CAT-1 and CAT-3 in Neurospora crassa. Genes Genet. Syst 2007, 82, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B-and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Karamanos, A.J.; Travlos, I.S. The water relations and some drought tolerance mechanisms of the marama bean. J. Agron 2012, 104, 65–72. [Google Scholar] [CrossRef]
- Yan, S.; Weng, B.; Jing, L.; Bi, W. Effects of drought stress on water content and biomass distribution in summer maize (Zea mays L.). Front. Plant Sci 2023, 14, 1118131. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.S.; Li, F.M.; Xu, H. Deficiency of water can enhance root respiration rate of drought-sensitive but not drought-tolerant spring wheat. Agric. Water Manag 2004, 64, 41–48. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, Y.; Yadav, V.; Zhao, W.; He, Y.; Zhang, X.; Wei, C. Drought-induced proline is mainly synthesized in leaves and transported to roots in watermelon under water deficit. Hortic. Plant J 2022, 8, 615–626. [Google Scholar] [CrossRef]
- Lv, B.S.; Ma, H.Y.; Li, X.W.; Wei, L.X.; Lv, H.Y.; Yang, H.Y.; Jiang, C.J.; Liang, Z.W. Proline accumulation is not correlated with saline-alkaline stress tolerance in rice seedlings. J. Agron 2015, 107, 51–60. [Google Scholar] [CrossRef]
- Cao, Y.; Luo, Q.; Tian, Y.; Meng, F. Physiological and proteomic analyses of the drought stress response in Amygdalus mira (Koehne) Yü et Lu roots. BMC Plant Biol 2017, 17, .1–16. [Google Scholar] [CrossRef] [PubMed]
- Keyster, M.; Klein, A.; Du Plessis, M.; Jacobs, A.; Kappo, A.; Kocsy, G.; Galiba, G.; Ludidi, N. Capacity to control oxidative stress-induced caspase-like activity determines the level of tolerance to salt stress in two contrasting maize genotypes. Acta Physiol Plant 2013, 35, 31–40. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Sohrabi, F.; Fasihfar, E.; Baniasadi, F.; Riasat, M.; Mozafari, A.A. Superoxide dismutase (SOD) as a selection criterion for triticale grain yield under drought stress: a comprehensive study on genomics and expression profiling, bioinformatics, heritability, and phenotypic variability. BMC Plant Biol 2021, 21, 1–19. [Google Scholar] [CrossRef]
- Mao, H.; Chen, M.; Su, Y.; Wu, N.; Yuan, M.; Yuan, S.; Brestic, M.; Zivcak, M.; Zhang, H.; Chen, Y. Comparison on photosynthesis and antioxidant defense systems in wheat with different ploidy levels and octoploid triticale. Int. J. Mol. Sci 2018, 19, 3006. [Google Scholar] [CrossRef]
- Huseynova, I.M.; Aliyeva, D.R.; Aliyev, J.A. Subcellular localization and responses of superoxide dismutase isoforms in local wheat varieties subjected to continuous soil drought. Plant Physiol. Biochem 2014, 81, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Sci. Rep 2019, 9, 8543. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.K.; Rajkumar, B.K.; Kumar, V. Differential responses of antioxidants and osmolytes in upland cotton (Gossypium hirsutum) cultivars contrasting in drought tolerance. Plant Stress 2021, 2, 100031. [Google Scholar] [CrossRef]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant catalases: peroxisomal redox guardians. Arch. Biochem. Biophys 2012, 525, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J. What is the role of hydrogen peroxide in plant peroxisomes? Plant Biol 2015, 17, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Chugh, V.; Kaur, N.; Gupta, A.K. Evaluation of oxidative stress tolerance in maize (Zea mays L.) seedlings in response to drought. Indian J Biochem Biophys 2011, 48, 47–53. [Google Scholar]
- Carvalho, M.D. Drought stress and reactive oxygen species. Plant Signal Behav 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R. L-ascorbic acid: a multifunctional molecule supporting plant growth and development. Scientifica 2013, 2013, 795964. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought stress in maize causes differential acclimation responses of glutathione and sulfur metabolism in leaves and roots. BMC Plant Biol 2016, 16, 1–15. [Google Scholar] [CrossRef]
- Khanna, P.; Kaur, K.; Gupta, A.K. Salicylic acid induces differential antioxidant response in spring maize under high temperature stress. Indian J. Exp. Biol 2016, 54, 386–393. [Google Scholar] [PubMed]
- Zhang, J.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol 1996, 132, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Rahantaniaina, M.S.; Li, S.; Chatel-Innocenti, G.; Tuzet, A.; Mhamdi, A.; Vanacker, H.; Noctor, G. Glutathione oxidation in response to intracellular H2O2: key but overlapping roles for dehydroascorbate reductases. Plant Signal. Behav 2017, 12, 1356531. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of water deficit on soil water status, plant water status, and plant growth. (A) The water potential of the soil in which Tylosema esculentum and Tylosema fassoglense were grown was measured on the day of harvesting, from three randomly selected pots of each water regime (well-watered, WW; water deficit, WD) from each of the two Tylosema species. (B) The water potential of the leaves of T. esculentum and T. fassoglense. Water potential was measured on the day of harvesting, from the second youngest leaf of three plants in each water regime (WW, WD) from each species. Fresh weights were measured from shoots (C) and roots (D) of T. esculentum and T. fassoglense. Data was captured and analysed by 1-way ANOVA on GraphPad Prism 6.0, with different letters above the bars indicating statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 1.
Effect of water deficit on soil water status, plant water status, and plant growth. (A) The water potential of the soil in which Tylosema esculentum and Tylosema fassoglense were grown was measured on the day of harvesting, from three randomly selected pots of each water regime (well-watered, WW; water deficit, WD) from each of the two Tylosema species. (B) The water potential of the leaves of T. esculentum and T. fassoglense. Water potential was measured on the day of harvesting, from the second youngest leaf of three plants in each water regime (WW, WD) from each species. Fresh weights were measured from shoots (C) and roots (D) of T. esculentum and T. fassoglense. Data was captured and analysed by 1-way ANOVA on GraphPad Prism 6.0, with different letters above the bars indicating statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
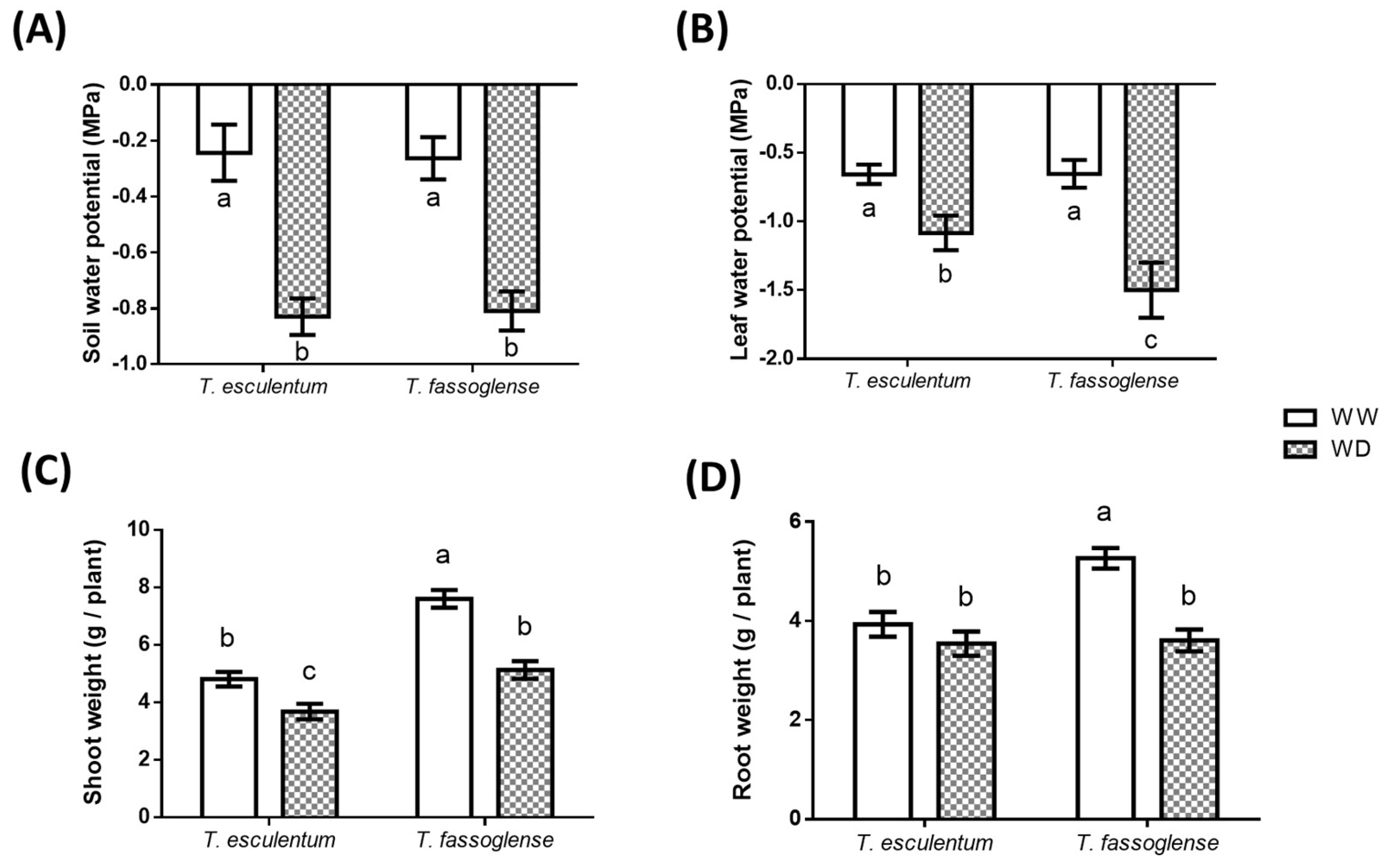
Figure 2.
Water deficit-induced changes in leaf relative water content (A) and leaf free proline content (B) in Tylosema esculentum and Tylosema fassoglense. Data represent means analysed by 1-way ANOVA on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 2.
Water deficit-induced changes in leaf relative water content (A) and leaf free proline content (B) in Tylosema esculentum and Tylosema fassoglense. Data represent means analysed by 1-way ANOVA on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
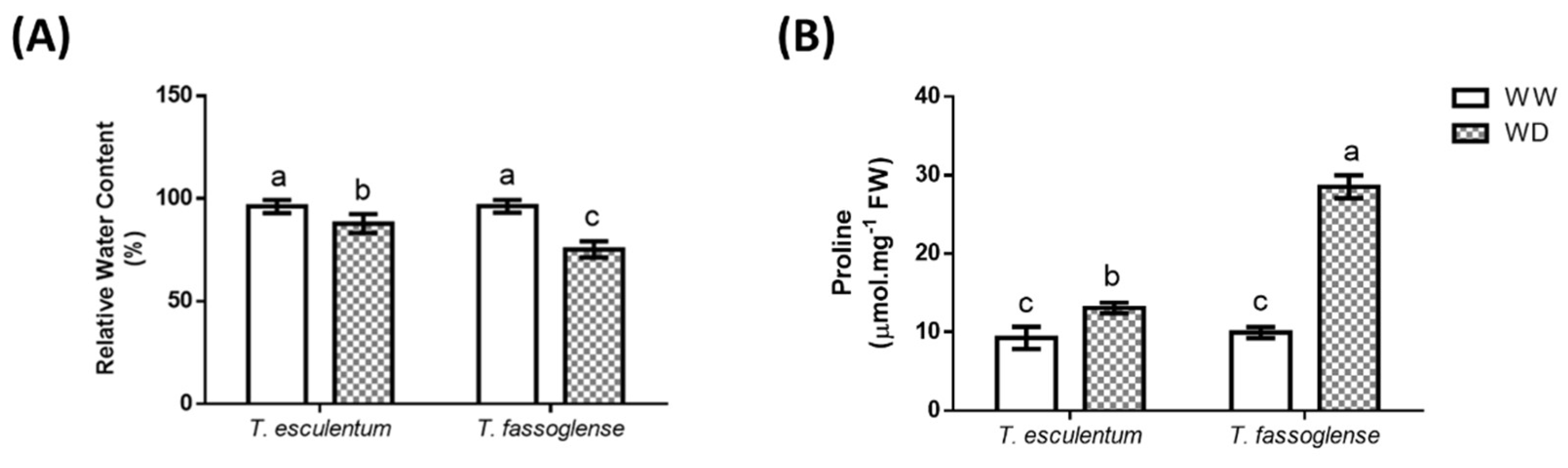
Figure 3.
Water deficit-induced changes in leaf hydrogen peroxide content (A), and malondialdehyde content (B) in Tylosema esculentum and Tylosema fassoglense. Data represent means analysed by 1-way ANOVA on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 3.
Water deficit-induced changes in leaf hydrogen peroxide content (A), and malondialdehyde content (B) in Tylosema esculentum and Tylosema fassoglense. Data represent means analysed by 1-way ANOVA on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
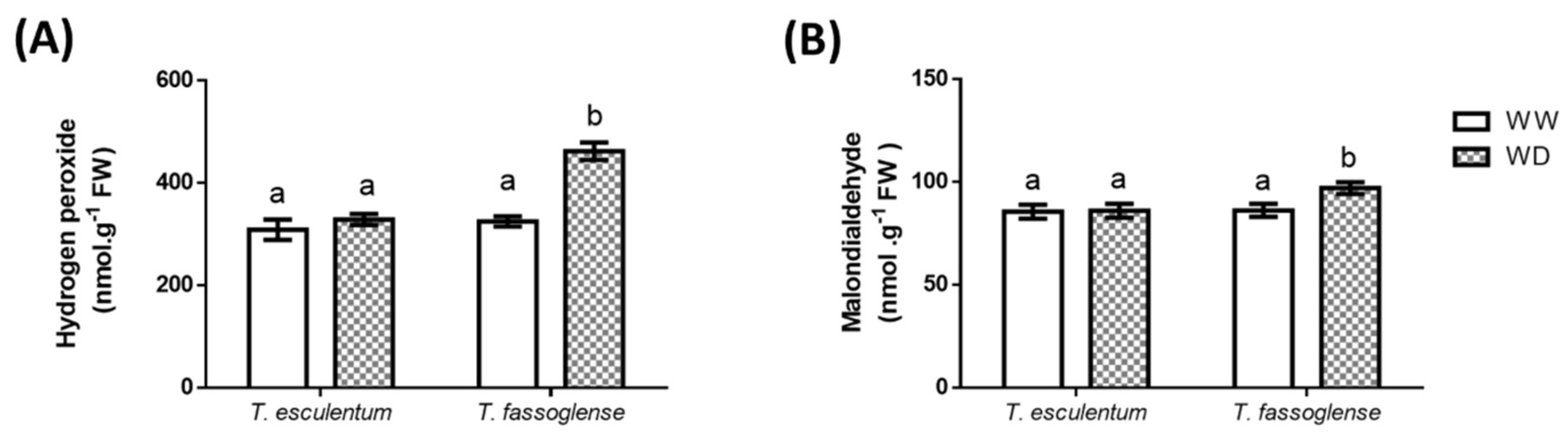
Figure 4.
Superoxide dismutase (SOD) activities measured in leaves and roots of Tylosema esculentum and Tylosema fassoglense. Water deficit-induced changes in leaf (A) and root (B) SOD activities were spectrophotometrically measured in the two Tylosema species. Changes triggered by water deficit on individual SOD enzyme isoforms (C) were detected using a native gel SOD activity assay in the leaves and roots. Data presented are from three independent experiments and, for numerical data, were subjected to 1-way ANOVA on GraphPad Prism 6.0. Letters above the bars indicate statistically similar (same letters) or statistically different (different letters) means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 4.
Superoxide dismutase (SOD) activities measured in leaves and roots of Tylosema esculentum and Tylosema fassoglense. Water deficit-induced changes in leaf (A) and root (B) SOD activities were spectrophotometrically measured in the two Tylosema species. Changes triggered by water deficit on individual SOD enzyme isoforms (C) were detected using a native gel SOD activity assay in the leaves and roots. Data presented are from three independent experiments and, for numerical data, were subjected to 1-way ANOVA on GraphPad Prism 6.0. Letters above the bars indicate statistically similar (same letters) or statistically different (different letters) means (P ≤ 0.05) based on the Tukey-Kramer test.
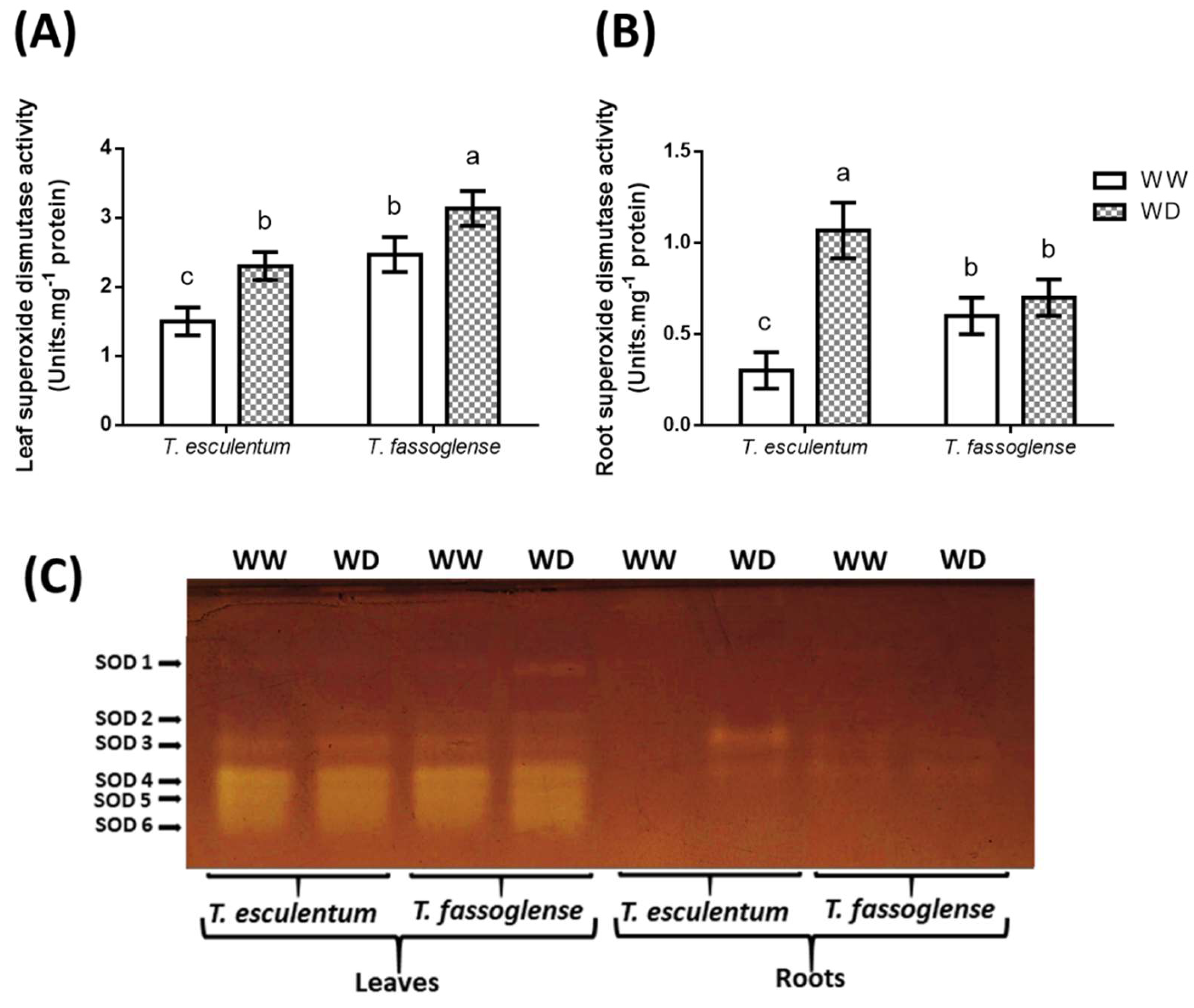
Figure 5.
Water deficit-induced changes in catalase enzymatic activity in the leaves (A) and roots (B) of the two Tylosema species. The effects of water deficit on individual CAT (C) isoforms were detected using native gel activity assay. For numerical data, 1-way ANOVA was used on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 5.
Water deficit-induced changes in catalase enzymatic activity in the leaves (A) and roots (B) of the two Tylosema species. The effects of water deficit on individual CAT (C) isoforms were detected using native gel activity assay. For numerical data, 1-way ANOVA was used on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
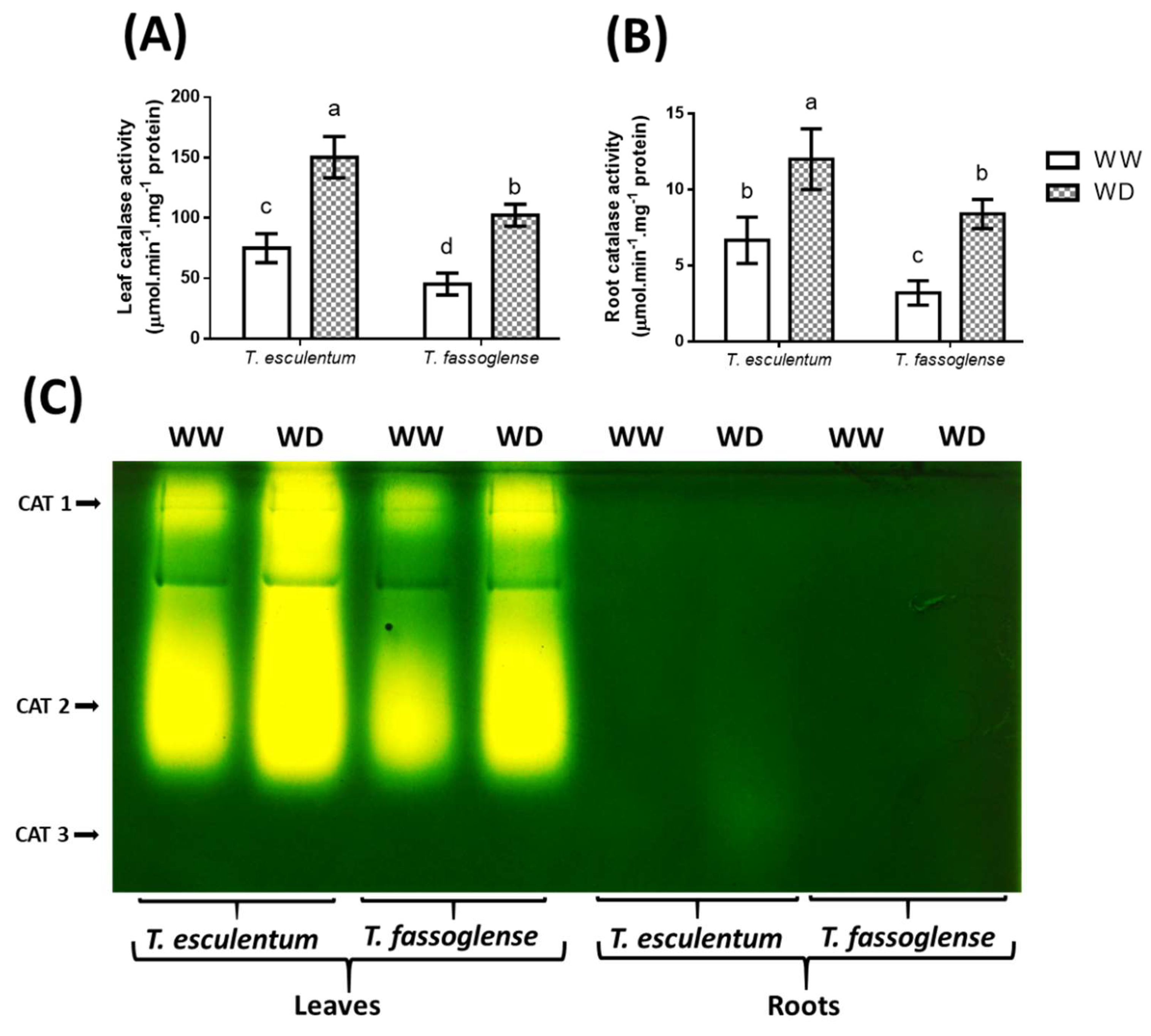
Figure 6.
Water deficit-induced changes in ascorbate peroxidase enzymatic activity in the leaves (A) and roots (B) of the two Tylosema species. The effects of water deficit on individual APX (C) isoforms were detected using native gel activity assay. For numerical data, 1-way ANOVA was used on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 6.
Water deficit-induced changes in ascorbate peroxidase enzymatic activity in the leaves (A) and roots (B) of the two Tylosema species. The effects of water deficit on individual APX (C) isoforms were detected using native gel activity assay. For numerical data, 1-way ANOVA was used on GraphPad Prism 6.0. Different letters above the bars indicate statistically different means (P ≤ 0.05) based on the Tukey-Kramer test.
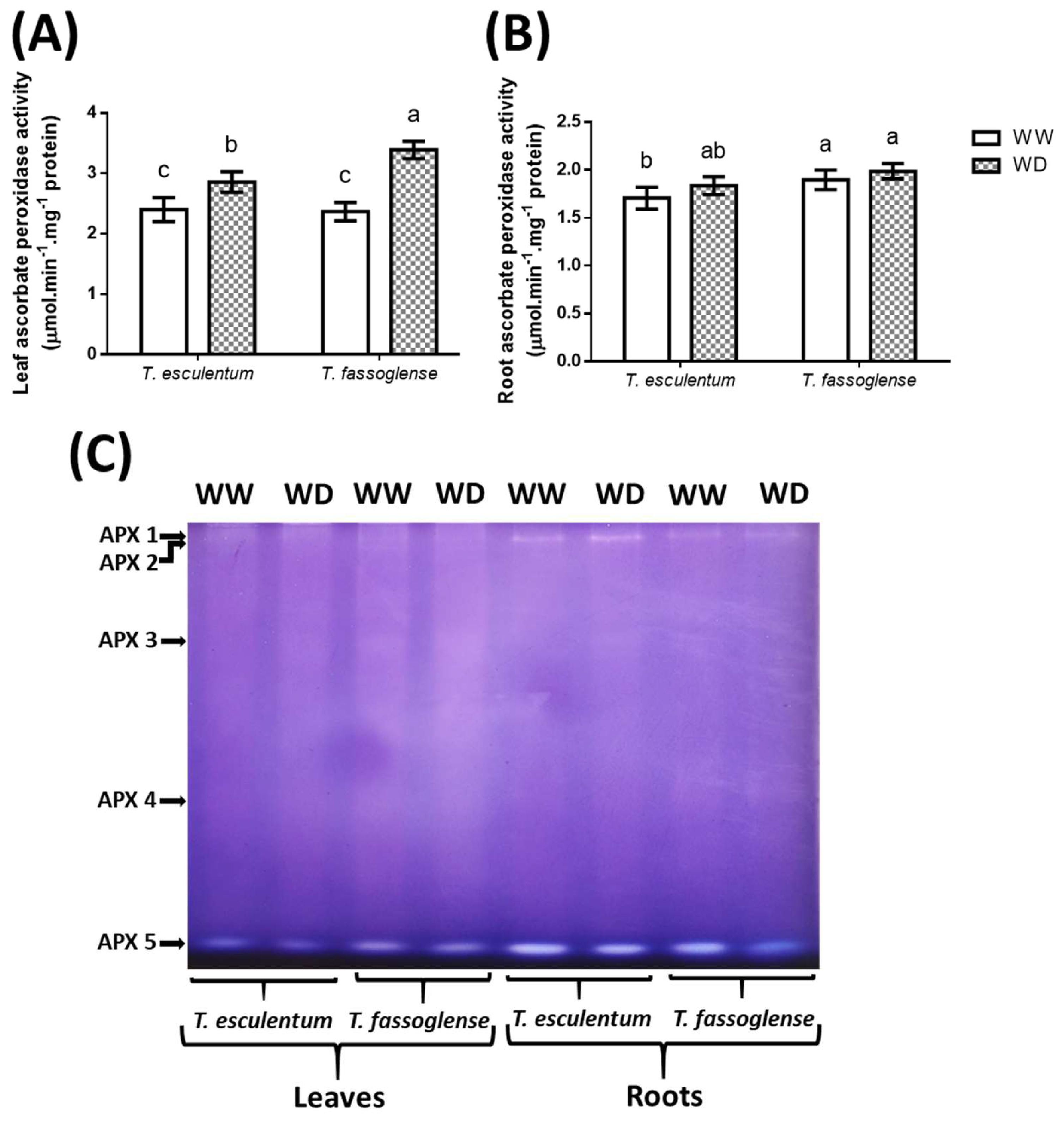
Figure 7.
Water deficit-induced changes in leaf (A) and root (B) GR activities in the two Tylosema species. Changes caused by water deficit on individual GR enzyme isoforms (C) were detected using a native gel GR activity assay in the leaves and roots. Data presented are from three independent experiments. For numerical data, means were subjected to 1-way ANOVA on GraphPad Prism 6.0. Letters above the bars indicate statistically similar (same letters) or statistically different (different letters) means (P ≤ 0.05) based on the Tukey-Kramer test.
Figure 7.
Water deficit-induced changes in leaf (A) and root (B) GR activities in the two Tylosema species. Changes caused by water deficit on individual GR enzyme isoforms (C) were detected using a native gel GR activity assay in the leaves and roots. Data presented are from three independent experiments. For numerical data, means were subjected to 1-way ANOVA on GraphPad Prism 6.0. Letters above the bars indicate statistically similar (same letters) or statistically different (different letters) means (P ≤ 0.05) based on the Tukey-Kramer test.
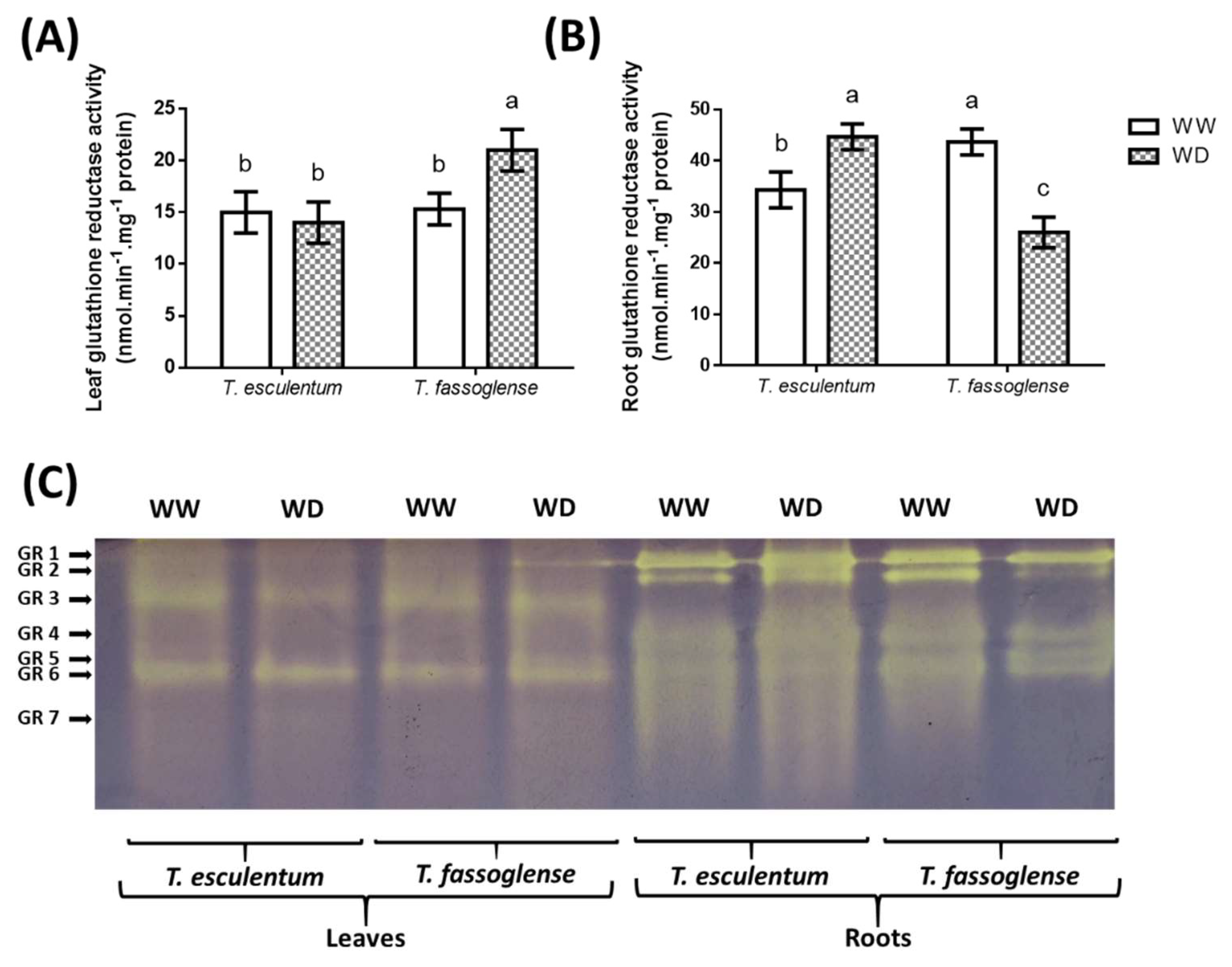
Figure 8.
The correlation among H2O2, MDA and antioxidant enzymes SOD, CAT, APX and GR) in Tylosema species in response to water deficit stress. The colour of the square and the coefficient values indicate the strength and direction of the relationships between parameters. A correlation coefficient of 1 indicates a perfect correlation; 0.75-1 indicates a strong correlation; 0.5-0.75 indicates a moderate correlation and 0.0-0.5 indicates a week correlation. A positive correlation indicates that as one parameter increases or decreases, the other moves in the same direction, while a negative correlation indicates an opposite relationship. H2O2: hydrogen peroxide; MDA: malondialdehyde; SOD: superoxide dismutase; CAT: catalase; APX: ascorbate peroxidase; GR: glutathione reductase.
Figure 8.
The correlation among H2O2, MDA and antioxidant enzymes SOD, CAT, APX and GR) in Tylosema species in response to water deficit stress. The colour of the square and the coefficient values indicate the strength and direction of the relationships between parameters. A correlation coefficient of 1 indicates a perfect correlation; 0.75-1 indicates a strong correlation; 0.5-0.75 indicates a moderate correlation and 0.0-0.5 indicates a week correlation. A positive correlation indicates that as one parameter increases or decreases, the other moves in the same direction, while a negative correlation indicates an opposite relationship. H2O2: hydrogen peroxide; MDA: malondialdehyde; SOD: superoxide dismutase; CAT: catalase; APX: ascorbate peroxidase; GR: glutathione reductase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



