
Submitted:
13 December 2024
Posted:
16 December 2024
You are already at the latest version
Abstract
In the last years, major changes in the biosystem related to the industrial development and envi-ronmental modifications have had a direct impact on human and animal fertility, as well as on biodiversity. It is widely demonstrated that all these changes impair in the reproductive function. Several studies have connected the increase of reactive oxygen species (ROS) generated in mito-chondria to the recently identified decline of fertility due to various factors, including heat stress. The study of antioxidants and especially of mitochondria targeted antioxidants, has been ad-dressed to identify more efficient and less toxic therapies that could circumvent the problem of in-fertility in mammals. These antioxidants can be obtained from natural compounds used in the diet and converted into more effective forms to mitochondria, which will be a much more natural therapy. The use of mitochondriotropic diet-based antioxidants in Assisted Reproductive Tech-nologies (ART) may be an important way to circumvent the low fertility, allowing the conserva-tion of biodiversity in animal species, including domestic breeds. This paper provides a concise re-view of the current state of the art on this topic, with a particular focus on antioxidants: Mitoqui-none, AntiOxBEN2, AntiOxCIN4, Urolithin A and the most recent Piperine.
Keywords:
fertility
; reproduction
; gamete
; embryos
; oxidative stress
; antioxidants
; mitochondria
1. Introduction
In recent years, pollution levels have increased significantly, largely as a consequence of industrialization during the 20th century. Agricultural fertilizers, manure, sewage, heavy metals, plastics, chemicals and industrial discharges are polluting coastal regions, oceans and continental ecosystems. Today, many of these compounds are found in the air, drinking water, soil, food, plants and in the bodies of animals and humans [1,2,3,4]. Poor air quality has become a global problem not limited to industrialized cities. All these changes and exposure to other specific environmental factors, such as temperature, climate, radiation and nutrition create an environment with increasing stress factors (Figure 1). These factors have a serious impact on the adaptation, reproduction and survival of living organisms by changing the ecosystem and biodiversity, with negative consequences, namely the growth of degenerative diseases and the decrease of fertility [5,6,7]. Environmental pollution has revealed to impair fertility in all mammal species [2,8]. Many of these toxic compounds are endocrine disruptors or have endocrine active substances that can damage the reproductive performance, with important social and economic consequences [3,4,9,10].
The most frequent cause of infertility has been related to the low number and quality of germ cells, namely azoospermia and oligospermia in the male or ovulation failure caused by hormonal disorders or behavioral changes in the female. It can also be caused by problems in the pelvic cavities such as endometriosis, Sertoli cell disorders or obstruction of the genital tract [11,12,13,14]. Infertility can also be linked to other factors, not only genetic or pathological, but also issues related to the age of the parents, nutritional problems, the environment, stress and the use of certain drugs [1,4,8,15]. This issue, compounded by factors such as population growth, climate change, socio-economic shifts, economic globalization, and financial constraints, has underscored the urgency of conducting research and developing strategies to manage fertility and maintain biodiversity. These efforts include the preservation of endangered local breeds, which are critical for sustaining ecological balance and genetic diversity [6,16]. It is estimated that a considerable number of species will become extinct in a near future [5].
Environmental changes, often caused by human activity, have repercussions on reproductive effectiveness and in the maintenance of species and subspecies. Guillette & Edwards (2005) showed that alligators in a lake contaminated with pesticides with nitrogenous compounds suffered from reproductive problems. Problems were also found in rats exposed to methoxychlor pesticide [1]. The relationship between hormonal system dysregulation and environmental contaminants has been proven over time [1,17].
Likewise, heat stress, closely associated with climate change, can also affect reproductive effectiveness. Different studies have showed that oocytes collected during warmer weather were of lower quality than those collected in winter, especially in extreme heat conditions [9,18] and that spermatogenesis is also affected, leading to low fertilization rates and embryonic development [19]. According to Leroy et al. (2008) the fertility of dairy cows has been declining since the mid-80s. The low quality of oocytes and embryos has been described as the main reason for low birth rates and high embryonic death rates. Low fertility rates are also associated with high production and feeding and it has been described that balanced diets, with a good supply of protein and energy, are essential for good energy stability and a consequent improvement in gamete quality. For instance, diets rich in starch can improve energy status and, therefore, ovarian activity in the early postpartum period. However, oocyte and embryonic quality can suffer from these diets, and diets with a high protein content can raise ammonia and urea concentrations in the blood, leading to modifications in the female reproductive system [20,21].
It is, therefore, urgent to find solutions to mitigate the deleterious effects of environmental and other changes, on fertility and welfare of animals and humans. This review briefly summarizes the current problems associated with infertility, focused on bovines and other animals, mainly related to stress factors that lead to increased oxidative stress and possible therapeutic approaches based on antioxidants, whether they are of natural origin or manipulated to act on the mitochondria.
2. Technological Applications to Improve Fertility
The commercialization of bovine embryos is widespread globally, and despite the economic challenges posed by the COVID-19 pandemic, the embryo industry has continued to grow. A deep analysis of data on the international trade of embryos is particularly useful to understand that these technologies have been adopted in 1/3 of the countries, which represent more than half of the world's cattle population [22]. The commercial value of this activity for the meat and milk trade is directly linked to genetic quality and crossbreeding schemes, which can be obtained by using oocytes, semen and embryos from selected donors, as well as, through the implementation of reproductive management strategies to improve pregnancy rates. Another advantage is related to the number of "cheap" embryos that can be produced in vitro and used for research [23,24] or infertility treatments such as embryo transfer for repeat-breeding cows [24,25,26]. Although some limitations associated to the in vitro fertilization technique were detected specifically linked to the low rate of produced embryos and some associated problems in the offspring it is an excellent tool for solving infertility problems and implementing studies related to fertility/infertility [23,27,28].
A huge scientific and technological advance has resulted from the extensive research carried out on several studies including reproduction and epigenetic alterations [29,30], oxidative stress in fertilization [31,32,33,34], among others. It should also be noted that ART practiced on different livestock species is a very successful business worldwide and as such, plays an important role in the livestock farming development [35,36].
2.1. Assisted Reproductive Technologies (ART)
Artificial insemination (AI) is the oldest technique used in assisted reproduction for both animals and humans. Initially introduced for animals of zootechnical interest in the late 1930s, AI had a strong sanitary focus. However, its large-scale adoption was driven primarily by economic factors. Following World War II, the development of semen cryopreservation greatly facilitated its distribution and expanded its commercial potential, particularly in the dairy industry. Additionally, AI has been applied to address issues related to sexual dysfunction and infertility [37,38,39].
The development of ART has opened new possibilities for the preservation of germplasm and fertility [6]. However, significant challenges remain, including technical limitations in laboratory procedures and escalating costs. To address these issues, concerted and sustained investment in research is essential to enhance these technologies and ensure their accessibility on a global scale [27,40]. Despite great efforts to improve the quality of both oocytes and in vitro produced embryos, the quality of these blastocysts is still inferior to those produced in vivo. These differences can be seen in morphology, metabolism, gene expression, tolerance to cryopreservation and persist throughout the development [41,42,43].
ART cover all in vivo and in vitro treatments or procedures on oocytes and sperm or embryos to establish a pregnancy, and have been widespread throughout the world, even in developing countries [22,40]. These include, among others, superovulation treatments and IA, sexing, In Vitro Fertilization (IVF), Intra Cytoplasmic Sperm Injection (ICSI), embryo transfer, and cryopreservation of gametes and embryos [11,24,37]. IVF was developed for use in humans and cattle, allowing embryo transfer worldwide [44]. Nevertheless, to be successful, IVF requires rigorous control and manipulation of the reproductive cycle to select the greatest number of oocytes and spermatozoa, assessing their quality, as well as, the quality of the embryos [43,45]. The ICSI, which consists of the microinjection of a single sperm into the cytoplasm of the oocyte, is currently the most widely used method to overcome serious male infertility problems and when the IVF method is unsuccessful, making it the most widely used in human and equine ART [38,45,46]. In horses, ICSI can be used when the number or quality of sperm is low, making it possible to overcome infertility problems in these animals. Also, oocyte retrieval and ICSI make it possible to manage normally fertile mares and stallions, reproducing and storing embryos of high genetic quality, creating a highly competitive market worldwide [38,39]. However, ICSI raises questions about the health of the offspring as it goes against natural selection [47], unlike IVF, where insemination itself allows the union of the two gametes to be brought closer to natural. The great success of embryo transfer and even IVF rates in cattle has spread its use worldwide, while ICSI is rarely used in this species [35].
As mentioned, ART comprises several stages, which can include ovarian stimulation, semen collection and freezing, IVF or ICSI, and also embryo culture, occasional biopsy, and finally embryo transfer, which involves significant changes to the gametes and the embryonic environment. It is now known that the early stages of mammalian embryonic development are very sensitive to their microenvironment [48]. Only 70% of the oocytes fertilized in vitro reach cleavage after three days of culture, and of these only 30%-50% give rise to blastocysts, falling short of development in vivo, even after the technique has been improved over the years [31,44,48]. For instance, pregnancy rates following the transfer of bovine in vitro produced embryos are 10-30% lower than those of embryos developed in vivo [48,49,50].
Deterioration in the quality of oocytes is one of the factors associated with the failure of ART, since their quality is a determining factor of the potential for development of the embryo after fertilization. Ovulation asynchrony and aged oocytes have been reported to be detrimental to the success of AI and embryo production programs in mare, cow and sheep, implying significant economic losses [20,46,51]. Any disturbance in the follicular or other environment affects the maturation of the female gamete and reduces its quality. In vitro maturation models have even shown that some of these metabolic alterations reduce the suitability of the oocyte, with repercussions on the development and quality of the embryo. Therefore, a technique to ensure optimal oocyte maturation is needed, as studies indicate, that adverse conditions for oocyte growth and maturation can also jeopardize the health and performance of the offspring [29,48,51,52].
In order to replicate the conditions of the maternal reproductive system, in vitro embryo production systems techniques must recreate the in vivo environment, with correct regulation of oxygen (O2) concentration and composition of culture media [53,54,55]. Several physicochemical factors affect oocytes and embryos, such as temperature control, maintenance of osmolality, pH and protection against oxidative stress and toxic substances [55,56]. De Munck et al. (2019), Leite (2017) and Amin et al (2014) showed a better efficacy of 5% O2 during embryo culture. Also, for cattle oocyte maturation and IVF the temperature should be 38.8ºC, in atmospheric conditions saturated with humidity and with 5% carbon dioxide (CO2)[53,57]. The culture media recreate the carbohydrate concentrations of the fallopian tubes and uterus by adding pyruvate, lactate, glucose, and proteins normally albumin, glutamine, and other amino acids [55,58,59]. Antioxidants have been reported to be also necessary [60,61]. Nevertheless, the culture conditions must be adapted to the stage of ART and species, existing protocols that differ between laboratories [6,43,57].
3. The Role of Mitochondria in Gametes Functionality
Environmental stressors, including heat stress, significantly affect the developmental competence of gametes. Furthermore, the mitochondrial response to these stressors has been identified as a major cause of reduced oocyte quality (Figure 2) and spermatozoa functionality [9,31,51,60,62]. Mitochondria are cellular organelles, responsible for several metabolic processes including oxidative phosphorylation (OXPHOS) and fatty acid metabolism, while also regulating cytosolic calcium concentration, the production of reactive oxygen species (ROS) and regulation of cell death pathways [63,64]. In addition to the nucleus, mitochondria are the only organelles in animal cells that contain their own DNA, known as mitochondrial DNA (mtDNA), which carries essential genetic information [44,65].
The mitochondria use the energy released by the oxidation of glucose and other sugars to synthesize adenosine triphosphate (ATP) from adenosine diphosphate (ADP). Electrons from nicotinamide adenine dinucleotide (NADH) or succinate produced in the mitochondrial matrix during the Krebs cycle are transferred to molecular oxygen by a series of protein complexes in the mitochondrial inner membrane (or in the cristae membranes), creating a transmembrane electrochemical gradient, which is used by the ATP synthase to produce ATP from ADP and inorganic phosphate [66]. This process is called OXPHOS and occurs under aerobic conditions. In addition to energy conversion, this process gives rise to the generation of ROS as by-products of electron transport chain (ETC) activity [67].
Within the oocyte, mitochondria are involved in ATP generation, calcium homeostasis, regulation of cytoplasmic redox status, signal transduction and apoptosis [9]. As oocyte maturation requires a large amount of ATP for continuous transcription and translation, the availability of the right number of functional mitochondria is essential. In addition, during oocyte maturation, the mtDNA copy number increases dramatically and the distribution of mitochondria changes significantly [44,65].There is therefore a correlation between oocyte quality and mtDNA copy number and ATP concentration [44,65,68].
In spermatozoa, mitochondria are also susceptible to loss of membrane potential and electron leakage during OXPHOS, reducing their energy production capacity [19,69,70,71]. Several studies have analyzed mitochondrial function in sperm cells establishing a positive correlation between sperm mitochondrial membrane potential (MMP) and other parameters required for sperm functionality such as motility, viability, capacitation status, acrosome, and chromatin integrity, suggesting that mitochondrial status reflects the quality of the sperm functionality [60,62]. Moreover, according to Jorge et al. (2024), differences among male sperm bioenergetic parameters can be used to predict spermatozoa functionality and developmental potential and therefore for the selection of bull breeders.
On the other hand, if IVF conditions are not optimal, there may be changes in the morphology of mitochondria and in the coding of proteins associated with their function, which can lead to a reduced ability to counteract ROS production, leading to oxidative stress [65]. Heat stress is closely associated with alterations in mitochondrial distribution and MMP, disrupting the expression of genes linked to mitochondrial function. This includes genes involved in the transcription and replication of mtDNA and those encoding OXPHOS complexes essential for ATP production [9].
Moreover, Soares et al (2020) have reported significant age-related alterations in bovine oocytes namely in the cytoplasmic volume, mitochondrial aggregation pattern, and mitochondria-produced H2O2 levels [72]. Aged oocytes have also presented reduced nuclear maturation progression, MMP, developmental competence, and altered gene expression levels [51]. Those unique characteristics make mitochondria appealing to study fertility/infertility and to test new therapeutic strategies aiming to improve the mitochondrial function of gametes and embryos.
4. Reactive Oxygen Species (ROS) in Gametes and Embryos
Oxidation-reduction reactions are a basic component of the systems of living beings and the free radicals, including oxygen ions and hydrogen peroxide that result from them, are also essential for nuclear maturation or sperm capacitation. These radicals are the result of cellular metabolism and enzymatic reactions that occur in the body and require a balance between the loss and gain of electrons [9,60,61,73,74]. As previously stated, during OXPHOS, a residual production of ROS occurs naturally as a consequence of electron transfer. Additional studies focusing on specific complexes embedded in the mitochondrial inner membrane have established that complexes I and III are the sites with the greatest capacity for generating ROS due to electron slippage [67,75] (Figure 3).
ROS plays direct and indirect roles in a very wide range of physiological processes. However, when there is an imbalance in their homeostasis, i.e. an imbalance between oxidants and antioxidants in favor of the oxidants, leads to a disruption of redox signaling and control and/or molecular damage [60,61,73,74,76]. Oxidative stress and mitochondrial dysfunction have been associated with metabolic diseases and age.
The presence of excess ROS within the ovary and endometrium has significant physiological and pathological implications for conceiving [74]. In IVF there are various sources of ROS, including the cells themselves, such as oocytes and spermatozoa, as well as the composition of the culture media used. Likewise, the laboratory culture conditions practiced during maturation and insemination, namely excess light, temperature variation, and high oxygen tension, increase the production of ROS and can have detrimental effects on post-fertilization development and assisted reproduction results, as they have a detrimental effect on mitochondria, DNA, RNA and fertilization [31,33,61].
The increase in ROS is well documented in oocytes exposed to heat stress or environmental toxicants [9,18] and in spermatozoa, causes damage to membrane lipids and decreased sperm motility. The exposure of DNA to both ROS and apoptosis enzymes during the cell division of spermatogenesis makes spermatozoa vulnerable, impairing not only fertilization but also subsequent embryonic development [19,70,77]. In addition, this increase in ROS production and consequent oxidative stress affects the quality of gametes, their environment, and ultimately, their interaction which can not only harm IVF success rates but, can also result in epigenetic and genetic changes in the embryo, resulting in transgenerational effects [31,61,78,79].
Marques et al. (2014) detected the production of mitochondria-specific ROS (mtROS) in sperm, using flow cytometry, with a mitochondria-specific superoxide fluorescent probe MitoSOX Red. In this study, they have shown that ejaculates were heterogeneous in mtROS production, with three detectable subpopulations, where the sperm subpopulation that produces the least amount of mtROS presented the most functional subset of male gametes, which were correlated with the largest number of live gametes and non-apoptotic sperm. In addition, this subpopulation was clearly more effective in samples that gave rise to pregnancies after assisted reproduction. Recently, Nakai et al. (2024) have shown that increased ROS production were associated with oocyte maturation inhibition, and demonstrated that the loss of a protein membrane (Mul1) was directly connected with the increasing of ROS concentrations in oocytes, resulting in an abnormal preimplantation embryogenesis. They reinforced that manipulating the mitochondrial ROS levels in oocytes may be a potential therapeutic approach to target infertility [80]
Several studies have therefore concluded that the use of antioxidants is an effective approach to mitigate oxidative stress and improve fertility [31,81,82]. Agarwal et al. (2005) state that infertility treatment strategies should focus on oral supplementation with antioxidants or, in the case of IVF, supplementation of the culture media with antioxidants to reduce oxidative stress.
5. Antioxidants in the Reproductive Medicine
Antioxidants are substances that inhibit oxidation by neutralizing free radicals found in the environment, reducing their effects on various diseases such as cancer, diabetes, degenerative diseases aging, and infertility [83]. Eukaryotic cells have antioxidant strategies and multifaceted mechanisms to maximize energy production while limiting the negative and toxic effects of ROS production [67]. There are several natural antioxidants in the body, including enzymatic antioxidants such as catalase, SOD, glutathione peroxidase [74,84]. In addition, non-enzymatic antioxidants such glutathione (GSH), vitamins C and E, hypotaurine, and taurine, among others provide maternal protection from oxidative stress to oocytes and embryos [41].
Excessive ROS production, if not counteracted by intrinsic defense mechanisms, will cause oxidative damage to cellular components, and, as mentioned above, can lead to DNA damage and cell death by necrosis or apoptosis [9,73,85,86,87]. Thus, antioxidant supplementation can be a strategy to improve fertility. Moreover, the use of antioxidants has been reported to be necessary for ART [60,61].
5.1. Endogenous Antioxidants
Reduced glutathione (GSH) is an endogenous tripeptide antioxidant (Figure 4) present in both male and female gametes that plays a very important role in fertility. GSH has a role in intracellular defense against oxidative stress and contributes to the regeneration of other antioxidants. [88,89]. A study by Ogata et al. (2022) has shown positive effects on the early formation of male pronuclei without negative effects on DNA integrity and cell number in blastocyst stage embryos. Although the effect was dependent on the bull and GSH concentration, semen supplementation has improved in vitro embryo production from frozen semen [88]. In this study, after GSH supplementation (0, 1, 5 and 10 mM), sperm quality varied between bulls. One bull had decreased total sperm motility, and two other bulls had decreased sperm DNA integrity. The GSH supplementation had positive effects on embryonic development in all three bulls. Two of them showed improved cleavage rates and blastocyst formation, while the other only showed a higher cleavage rate. Besides GSH, to counteract the harmful effects of ROS, many studies have included vitamin E in the semen extenders or embryo culture media to improve cryosurvival parameters, sperm cell viability, and embryo production [90,91,92]. Vitamin E is a well-established naturally-occurring lipid-soluble chain-breaking antioxidant that scavenges oxygen radicals within the membranes, acting in synergy with vitamin C, and protecting germlines [92,93].
Coenzyme Q10 (CoQ10) or ubiquinone (Figure 4), a free radical scavenger and a very important component of the mitochondrial, ETC has also been pointed out to impact male and female gametes [9,18,94]. CoQ10 antioxidant function (ubiquinol) works by inhibiting lipid peroxidation in vitro and in vivo. In addition to inhibiting lipid peroxidation without the mediation of vitamin E, ubiquinol can also amplify the antioxidant effect of this vitamin. It should be noted that ubiquinol is the only known fat-soluble antioxidant that can be synthesized in animal cells and regenerated through the mitochondrial electron transport system to its reduced antioxidant form. These characteristics, together with its high degree of hydrophobicity and its general occurrence in biological membranes and low-density lipoproteins, suggest a very important role for ubiquinol in cellular defense against oxidative damage [95,96].
Gendelman & Roth (2012) found that oocyte maturation medium supplemented with 50μM of CoQ10 induced changes in mitochondrial distribution within the oocytes and increased the proportion of polarized mitochondria. In addition, CoQ10 incorporation into oocytes induced changes in gene transcription involved in the mitochondrial ETC and a higher proportion of embryos that developed into blastocysts. However, CoQ10 did not improve oocyte development in winter and summer, which suggests that this antioxidant only contributes beneficially in phases with moderate climate damage, as there was an improvement in the fall [18]. Even so, these authors have suggested the widespread use of this antioxidant to increase the in vitro production of bovine embryos.
In 2017, Yang et al. tested the action of the natural antioxidant melatonin (10-9 M) (Figure 4) in IVF with positive results in the maturation of bovine oocytes and an increase in the number of produced embryos, concluding that melatonin improves the distribution of mitochondria and preserves the ATP production [41].
5.2. Exogenous Antioxidants
Natural antioxidants derived from the diet have antioxidative characteristics with the potential to prevent diseases caused by oxidative stress [83]. In fact, antioxidants in fruit, vegetables, and drinks play an important role in mammalian health, such as preventing cancer and cardiovascular disease and reducing the incidence of different diseases [97]. The study of these natural compounds that are available in food/feed will contribute to increasing their use, rather than the use of artificial drugs.
5.2.1. Naturally-Occurring Antioxidants
Piperine is a simple alkaloid found in black pepper seeds (Figure 5), whose biological properties have been extensively studied at the pharmacological level and has shown great therapeutic potential as an antioxidant, anticancer, anti-inflammatory, antihypertensive, hepatoprotective, neuroprotective and in improving fertility. It also can alter gastrointestinal disorders and drug metabolizing enzymes [98,99].
Piperine induces specific toxicity in cancer cells, but does not affect normal cells, and has also been shown to restore the function of aged cells around cancer cells [100]. In addition, piperine improves the efficacy of current cancer therapies and represents a good adjuvant to certain phytochemical compounds such as curcumin or resveratrol.
The use of piperine as a bioactive compound is still in its infancy, so studies on this compound are still essential for the formulation, improvement and discovery of new drugs. Recently our team has used piperine during bovine oocyte maturation. It was demonstrated that 1 and 10µM piperine improved the number of mature oocytes (metaphase II.) Also, higher MMP was attained with 1µM concentration compared to the control [101].
Polyphenols are secondary plant metabolites mostly involved in defense against oxidative stress and are found largely in fruits, vegetables, cereals and beverages present in the diet [102,103,104]. Over the last few years, several studies have found that foods rich in polyphenols protect against age-related diseases such as atherosclerosis, cardiovascular disease, cancer, arthritis, cataracts, osteoporosis, type 2 diabetes mellitus, hypertension, Alzheimer's disease, and diseases with mitochondrial etiology [102,105].
In 2024, Yang et al. showed the positive effect of the supplementation of the natural antioxidant astaxanthin (Figure 5) to porcine granulosa cells culture identifying positive variations in morphology, apoptosis, ROS levels, and the expression of apoptosis and anti-oxidation-related genes [106]. Astaxanthin also has ameliorated oxidative stress and reproductive outcomes after assisted reproduction [107].
Another example of an antioxidant of natural origin is Urolithin A (Figure 5), which is a metabolite resulting from the transformation by intestinal bacteria of ellagitannins, ellagic acid and polyphenols, found in foods such as pomegranates, berries and nuts. Several studies have shown that it plays an important role in preventing ageing and various diseases [51,108,109]. Fonseca et al. (2021) have tested the use of this antioxidant in the development of oocytes from old and young cows and concluded that supplementing this antioxidant in the oocyte maturation medium prevented oocyte aging and improved embryonic development. Later, Jorge et al. (2024) reinforced the potential therapeutic value of Urolithin A for addressing reproductive sub/infertility problems and improving ART outcomes. Their results have showed that Urolithin A has improved sperm motility quality, increasing ATP production while reducing oxidative stress levels in a dose-dependent manner.
5.2.2. Synthetic Mitochondriotropic Antioxidants
For several years, different approaches have been established to prevent oxidative damage to mitochondria, including the development of ETC inhibitors, OXPHOS, mitochondrial Ca2+ modulators, and mitochondriotropic antioxidants [86]. The pharmacological activities of new naturally derived and synthetic molecules have been extensively studied [83]. Currently, the prevention of mitochondrial oxidative damage through pharmacological solutions is recognized as an indispensable tool [51,102], to strengthen mitochondria's antioxidant power. Mitochondrial function can be improved through the use of antioxidants and various types of antioxidants have already been tested in animal and human oocytes with promising results [65], especially when targeted at the mitochondria [59,60,110].
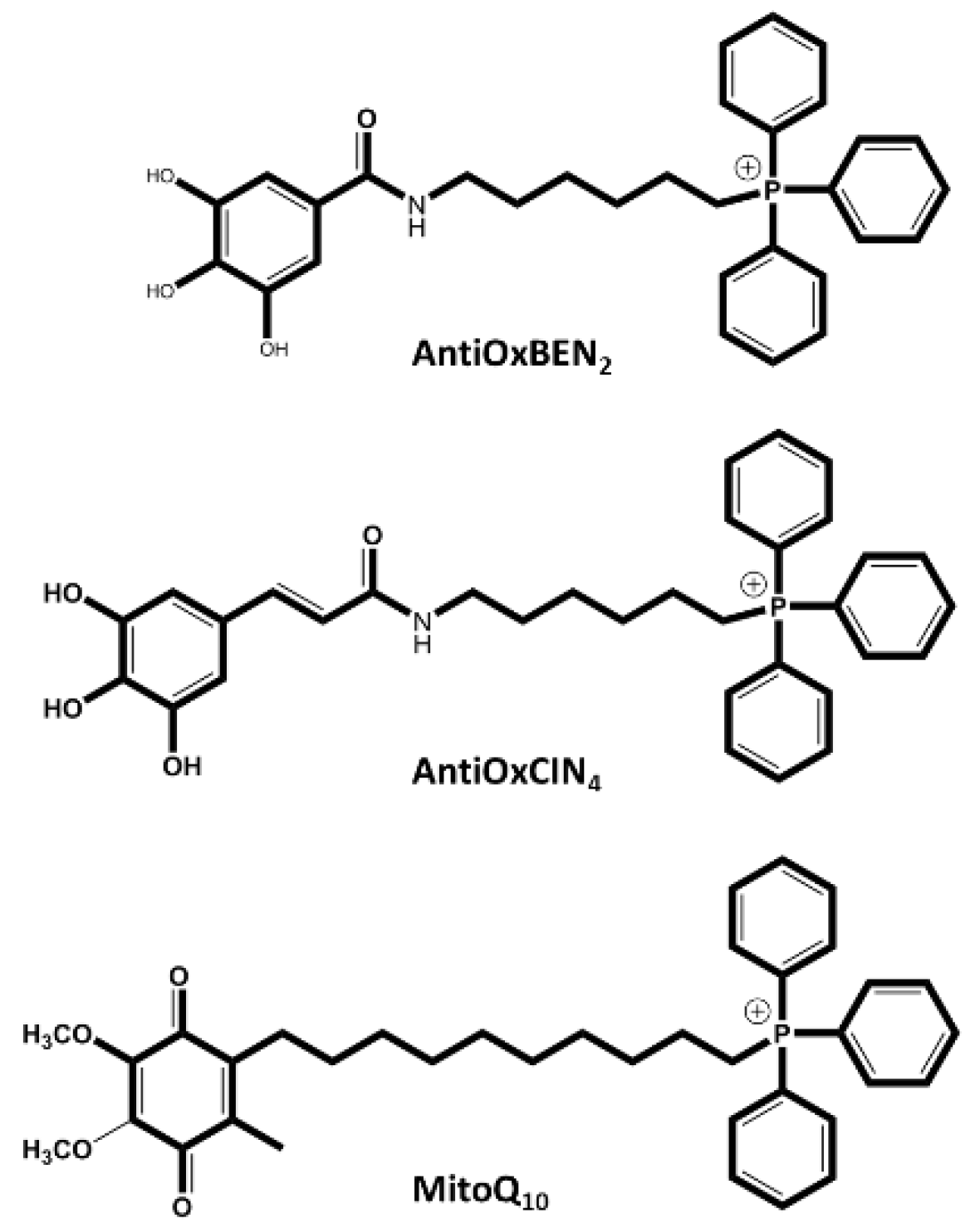
Mitoquinone (MitoQ) is one of the most widely studied mitochondriotropic antioxidants. MitoQ has shown encouraging pre-clinical results in numerous studies on isolated mitochondria, cells, and tissues subjected to oxidative stress and death by apoptosis, as it can reproduce the role of the endogenous mitochondrial antioxidant CoQ10. This coenzyme is covalently linked to a triphenylphosphonium cation (TPP) by a 10-carbon alkyl chain (dTPP), a lipophilic spacer (Figure 6) that allows the molecule to cross mitochondrial membranes and considerably increases its antioxidant capacity. This antioxidant represents the first clinical attempt to deliver an antioxidant to the mitochondria. The results of the clinical trials with MitoQ were very important for understanding the relevance of a mitochondria-oriented approach [102,111].
A study in bovine oocytes demonstrated that MitoQ (1µM) when supplemented to the culture medium improved mitochondrial function and enhanced embryo development. This effect was achieved by reducing mitochondrial ROS levels below the critical thresholds that trigger apoptosis [112]. Another study carried out during the maturation of rat oocytes also concluded that the addition of MitoQ (0, 0.01, 0.02 and 0.04µM) to IVF culture media improved oocyte fertilization and the subsequent development of blastocysts [113]. An increased GSH level and membrane potential of oocytes concomitant with reduced intracellular ROS concentrations were identified. Also, MitoQ supplementation (25, 50, and 100nM) to a stallion semen extender before cryopreservation showed the positive effect of this antioxidant on sperm mobility, especially at 25nM. The highest concentration of 200nM harmed the mobility and viability parameters of frozen-thawed semen. Nevertheless, none of the used concentrations affect the plasma membrane, acrosome, DNA integrity, MMP or intracellular ROS concentrations [114]. Conversely, the addition of MitoQ (0, 0.2, 2 and 20nM) to the semen extender before cryopreservation was unable to improve the post-thaw quality of bull sperm [115]. According to some studies, MitoQ supplementation has harmed the bioenergetic function of mitochondria and did not contribute to the inhibition of iron toxicity [102,116]
AntiOxBEN2 is a mitochondria-targeted antioxidant derived from gallic acid, a hydroxybenzoic acid. AntiOxBEN2 was synthesized by conjugating the antioxidant gallic acid with the lipophilic triphenylphosphonium cation (TPP+) (Figure 6) through a 6-carbon aliphatic chain, designed to specifically accumulate within the mitochondrial matrix [102,117,118]. Results in in vitro models show that AntiOxBEN2, unlike MitoQ10, proved capable of chelating ferrous iron upregulating antioxidant systems, and improving mitochondria function by activating the Nrf2/Keap1 pathway [119]. Teixeira et al. (2020) have studied the effect of mitochondriotropic antioxidant AntiOxBEN2 on the prevention of oxidative stress in bovine oocytes and embryos by supplementing it to the oocyte maturation medium with concentrations of 10, 20, 50 and 100μM. AntiOxBEN2 improved oocyte maturation and embryo production in a dose-dependent manner. These authors have suggested its future use at a concentration of 10μM during the oocyte maturation process. Later, Santos et al. (2022) supplemented culture media during IVF of bovine oocytes with AntiOxBEN2, demonstrating its benefits in reducing ROS, increasing spermatozoa MMP and consequently sperm quality. When the sperm capacitation and the fertilization media were supplemented with a concentration of 1µM of AntiOxBEN2, an improvement in embryo development, due to an increase of the number of cleaved embryos and blastocysts, was seen.
AntiOxCIN4 is a mitochondriotropic antioxidant based on caffeic acid that has been described to prevent oxidative stress-related events through the activation of endogenous ROS-protective pathways in normal primary human fibroblasts (PHSF) and in PHSF from sporadic Parkinson's disease patients (Figure 6). AntiOxCIN4 also increased cell stress resistance in human hepatoma-derived cells (HepG2) by activating the Nrf2-p62-Keap1 axis, leading to up-regulation of antioxidant defenses, triggering macroautophagy and/or mitochondrial autophagy (mitophagy) and mitochondrial biogenesis [116,120]. Additionally, AntiOxCIN4 supplementation was shown to improve steatotic liver in a metabolic disease mice model [121]. Teixeira et al. (2020) have supplemented the oocyte maturation medium with concentrations of 10, 20, 50 and 100μM of AntiOxCIN4, improving oocyte maturation in a dose-dependent manner. Recently, the supplementation of AntiOxCIN4 to the capacitation and fertilization media, at concentrations of 0.1 and 1μM, was studied. It was found that the supplementation of 1μM AntiOxCIN4 during the sperm capacitation process improved some of the functional characteristics of the spermatozoa and that both concentrations (0.1 and 1μM) have increased the number of good quality embryos. However, the study is still ongoing for a more robust approach [122].
Finally, AntiOxBEN2 and AntiOxCIN4 antioxidants at concentrations of 1, 2.5 and 10μM have also been tested in the embryo culture media to prevent oxidative stress. The concentration of 2.5μM improved the quality of the produced embryos. Based on these results, this concentration was used to further study the resistance of these embryos to the vitrification process. The data have shown that AntiOxCIN4 and especially AntiOxBEN2 had a beneficial effect on embryo development and cryopreservation survival, pointing to a possible therapy to prevent oxidative stress in ART [123,124].
6. Conclusions
This review has shown that infertility problems are increasing worldwide, both in humans and in animals. The reasons given range from human-induced environmental changes such as the industrial revolution of the 20th century, with its repercussions on pollution and all its surroundings, to climate change and lifestyles adopted, which encompass a large number of changes. Presently, ART procedures are imperative to overcome infertility problems, but also to maintain animal productivity and biodiversity. The study of natural antioxidants targeting mitochondria is essential, since mitochondria play a predominant role in the formation of ROS and oxidative stress. These compounds have been shown to present several different and positive roles, not only in terms of fertility, but also in the prevention and therapy of ageing, cancer, and neurodegenerative diseases, among others. Some studies have already been carried out with these antioxidants in ART, with promising results. However, this is still an area to be explored and more robust data are needed to consolidate the existing ones.
Funding
This research was funded by FCT UIDB/00276/2024 and Cryostore projects http://dx.doi.org/10.13039/100018693.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to acknowledge the contribution of Paulo Oliveira in critically reviewing the manuscript.
Conflicts of Interest
Fernanda Borges is cofounder of the CNC-UP spin-off company MitoTAG (Coimbra, Portugal). This SME had no involvement in the data collection, analysis, interpretation, writing of the manuscript, and decision to submit it for publication. All authors declare no conflict of interest.
References
- Luoma, J. Challenged Conceptions: Environmental Chemicals and Fertility. In Proceedings of the Understanding Environmental Contaminants and Human Fertility: Science and Strategy; Stanford University School of Medicine’s: Stanford, October 2005. [Google Scholar]
- Canipari, R.; De Santis, L.; Cecconi, S. Female Fertility and Environmental Pollution. Int J Environ Res Public Health 2020, 17, 8802. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Mao, R.; Wang, D.; Yu, P.; Zhou, C.; Liu, J.; Li, S.; Nie, Y.; Liao, H.; Peng, C. Association of Plasma Metal Levels with Outcomes of Assisted Reproduction in Polycystic Ovary Syndrome. Biol Trace Elem Res 2024, 202, 4961–4977. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Mao, Y.; Zhang, H.; Lou, H.; Zhang, L.; Moreira, J.P.; Jin, F. Exposure of Women Undergoing In-Vitro Fertilization to per-and Polyfluoroalkyl Substances: Evidence on Negative Effects on Fertilization and High-Quality Embryos. Environmental Pollution 2024, 359, 124474. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Boni, R.; Tosti, E. Gamete Quality in a Multistressor Environment. Environ Int 2020, 138. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Marques, C.C.; Pimenta, J.; Barbas, J.P.; Baptista, M.C.; Diniz, P.; Torres, A.; Lopes-da-Costa, L. Assisted Reproductive Technologies (ART) Directed to Germplasm Preservation. In Advances in Animal Health, Medicine and Production; Springer International Publishing, 2020; pp. 199–215.
- Sakali, A.-K.; Bargiota, A.; Bjekic-Macut, J.; Macut, D.; Mastorakos, G.; Papagianni, M. Environmental Factors Affecting Female Fertility. Endocrine 2024, 86, 58–69. [Google Scholar] [CrossRef] [PubMed]
- LaPointe, S.; Lee, J.C.; Nagy, Z.P.; Shapiro, D.B.; Chang, H.H.; Wang, Y.; Russell, A.G.; Hipp, H.S.; Gaskins, A.J. Air Pollution Exposure in Vitrified Oocyte Donors and Male Recipient Partners in Relation to Fertilization and Embryo Quality. Environ Int 2024, 193, 109147. [Google Scholar] [CrossRef]
- Roth, Z. Symposium Review: Reduction in Oocyte Developmental Competence by Stress Is Associated with Alterations in Mitochondrial Function. J Dairy Sci 2018, 101, 3642–3654. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A.J. Usefulness of Bovine and Porcine IVM/IVF Models for Reproductive Toxicology. Reproductive Biology and Endocrinology 2014, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, S. Infertility. In Encyclopedia of Mental Health: Second Edition; Elsevier Inc., 2016; pp. 375–383 ISBN 9780123970459.
- Kiesswetter, M.; Marsoner, H.; Luehwink, A.; Fistarol, M.; Mahlknecht, A.; Duschek, S. Impairments in Life Satisfaction in Infertility: Associations with Perceived Stress, Affectivity, Partnership Quality, Social Support and the Desire to Have a Child. Behavioral Medicine 2020, 46, 130–141. [Google Scholar] [CrossRef]
- Szkodziak, P.; Wozniak, S.; Czuczwar, P.; Wozniakowska, E.; Milart, P.; Mroczkowski, A.; Paszkowski, T. Infertility in the Light of New Scientific Reports – Focus on Male Factor. Annals of Agricultural and Environmental Medicine 2016, 23, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, A.A.; Reijo Pera, R.A. Infertility. Brenner’s Encyclopedia of Genetics: Second Edition 2013, 71–74. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Chavarro, J.E. Diet and Fertility: A Review. Am J Obstet Gynecol 2018, 218, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Peng, W.F.; Hu, X.J.; Zhao, Y.X.; Lv, F.H.; Yang, J. Global Genomic Diversity and Conservation Priorities for Domestic Animals Are Associated with the Economies of Their Regions of Origin. Sci Rep 2018, 8. [Google Scholar] [CrossRef]
- Yawer, A.; Sychrová, E.; Labohá, P.; Raška, J.; Jambor, T.; Babica, P.; Sovadinová, I. Endocrine-Disrupting Chemicals Rapidly Affect Intercellular Signaling in Leydig Cells. Toxicol Appl Pharmacol 2020, 404, 115177. [Google Scholar] [CrossRef]
- Gendelman, M.; Roth, Z. Incorporation of Coenzyme Q10 into Bovine Oocytes Improves Mitochondrial Features and Alleviates the Effects of Summer Thermal Stress on Developmental Competence. Biol Reprod 2012, 87. [Google Scholar] [CrossRef]
- Capela, L.; Leites, I.; Romão, R.; Lopes-Da-costa, L.; Pereira, R.M.L.N. Impact of Heat Stress on Bovine Sperm Quality and Competence. Animals 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Leroy, J.L.M.R.; Van Soom, A.; Opsomer, G.; Goovaerts, I.G.F.; Bols, P.E.J. Reduced Fertility in High-Yielding Dairy Cows: Are the Oocyte and Embryo in Danger? Part II. Mechanisms Linking Nutrition and Reduced Oocyte and Embryo Quality in High-Yielding Dairy Cows. Reproduction in Domestic Animals 2008, 43, 623–632. [Google Scholar] [CrossRef]
- Nandi, S.; Tripathi, S.K.; Gupta, P.S.P.; Mondal, S. Nutritional and Metabolic Stressors on Ovine Oocyte Development and Granulosa Cell Functions in Vitro. Cell Stress Chaperones 2018, 23, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Hm Viana, J. 2020 Statistics of Embryo Production and Transfer in Domestic Farm Animals; 2021. [Google Scholar]
- Galli, C.; Lazzari, G. Practical Aspects of IVM/IVF in Cattle. Anim Reprod Sci 1996, 42, 371–379. [Google Scholar] [CrossRef]
- Parker Gaddis, K.L.; Dikmen, S.; Null, D.J.; Cole, J.B.; Hansen, P.J. Evaluation of Genetic Components in Traits Related to Superovulation, in Vitro Fertilization, and Embryo Transfer in Holstein Cattle. J Dairy Sci 2017, 100, 2877–2891. [Google Scholar] [CrossRef] [PubMed]
- Yaginuma, H.; Funshima, N.; Tanikawa, N.; Miyamura, M.; Tsuchiya, H.; Noguchi, T.; Iwata, H.; Kuwayama, T.; Shirasuna, K.; HAMANO, S. Improvement of Fertility in Repeat Breeder Dairy Cattle by Embryo Transfer Following Artificial Insemination: Possibility of Interferon Tau Replenishment Effect. Journal of Reproduction and Development 2019, 65, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Funeshima, N.; Noguchi, T.; Onizawa, Y.; Yaginuma, H.; Mitamura, M.; Tsuchiya, H.; Iwata, H.; Kuwayama, T.; Hamano, S.; Shirasuna, K. The Transfer of Parthenogenetic Embryos Following Artificial Insemination in Cows Can Enhance Pregnancy Recognition via the Secretion of Interferon Tau. Journal of Reproduction and Development 2019, 65, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Realizing the Promise of IVF in Cattle—an Overview. Theriogenology 2006, 65, 119–125. [Google Scholar] [CrossRef]
- Siqueira, L.G.B.; Dikmen, S.; Ortega, M.S.; Hansen, P.J. Postnatal Phenotype of Dairy Cows Is Altered by in Vitro Embryo Production Using Reverse X-Sorted Semen. J Dairy Sci 2017, 100, 5899–5908. [Google Scholar] [CrossRef] [PubMed]
- Leroy, J.L.M.R.; Rizos, D.; Sturmey, R.; Bossaert, P.; Gutierrez-Adan, A.; Van Hoeck, V.; Valckx, S.; Bols, P.E.J. Intrafollicular Conditions as a Major Link between Maternal Metabolism and Oocyte Quality: A Focus on Dairy Cow Fertility. Reprod Fertil Dev 2012, 24, 1–12. [Google Scholar]
- Malhi, P.S.; Adams, G.P.; Mapletoft, R.J.; Singh, J. Oocyte Developmental Competence in a Bovine Model of Reproductive Aging. Reproduction 2007, 134, 233–239. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R. Oxidative Stress and Its Implications in Female Infertility - A Clinician’s Perspective. Reprod Biomed Online 2005, 11, 641–650. [Google Scholar]
- Song, P.; Liu, C.; Sun, M.; Liu, J.; Lin, P.; Wang, A.; Jin, Y. Oxidative Stress Induces Bovine Endometrial Epithelial Cell Damage through Mitochondria-Dependent Pathways. Animals 2022, 12. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative Stress in an Assisted Reproductive Techniques Setting. Fertil Steril 2006, 86, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Leite, R.F.; Annes, K.; Ispada, J.; de Lima, C.B.; dos Santos, É.C.; Fontes, P.K.; Gouveia Nogueira, M.F.; Milazzotto, M.P. Corrigendum to “Oxidative Stress Alters the Profile of Transcription Factors Related to Early Development on in Vitro Produced Embryos” (Oxidative Medicine and Cellular Longevity (2017) 2017 (1502489). Oxid Med Cell Longev 2018, 2018. [CrossRef]
- Moore, S.G.; Hasler, J.F. A 100-Year Review: Reproductive Technologies in Dairy Science. J Dairy Sci 2017, 100, 10314–10331. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Marques, C. Produção e Transferência de Embriões: Uma Técnica Em Expansão; Vida Rural, 2017. 1892. 28-29.
- Verberckmoes, S.; Soom, A. Van; De Kruif, A. Intra-Uterine Insemination in Farm Animals and Humans. Reproduction in Domestic Animals 2004, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.H.A. The Development of in Vitro Embryo Production in the Horse. Equine Vet J 2018, 50, 712–720. [Google Scholar] [CrossRef]
- Hinrichs, K. Assisted Reproductive Techniques in Mares. Reproduction in Domestic Animals 2018, 53, 4–13. [Google Scholar] [CrossRef]
- Inhorn, M.C.; Patrizio, P. Infertility around the Globe: New Thinking on Gender, Reproductive Technologies and Global Movements in the 21st Century. Hum Reprod Update 2014, 21, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tao, J.; Chai, M.; Wu, H.; Wang, J.; Li, G.; He, C.; Xie, L.; Ji, P.; Dai, Y.; et al. Melatonin Improves the Quality of Inferior Bovine Oocytes and Promoted Their Subsequent IVF Embryo Development: Mechanisms and Results. Molecules 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; de La Fuente, J.; Lonergan, P.; Gutiérrez-Adán, A. Consequences of In Vitro Culture Conditions on Embryo Development and Quality. Reproduction in Domestic Animals 2008, 43, 44–50. [Google Scholar] [CrossRef]
- Pereira, R.M.; Marques, C.C. Animal Oocyte and Embryo Cryopreservation. Cell Tissue Bank 2008, 9, 267–277. [Google Scholar] [CrossRef]
- Kageyama, M.; Ito, J.; Shirasuna, K.; Kuwayama, T.; Iwata, H. Mitochondrial Reactive Oxygen Species Regulate Mitochondrial Biogenesis in Porcine Embryos. 2021. [CrossRef]
- Palermo, G.D.; Neri, Q. V.; Rosenwaks, Z. To ICSI or Not to ICSI. Semin Reprod Med 2015, 33, 92–102. [Google Scholar] [CrossRef]
- Carnevale, E.M.; Maclellan, L.J.; Stokes, J.A.E. In Vitro Culture of Embryos from Horses. In Methods in Molecular Biology; Humana Press Inc., 2019; Vol. 2006, pp. 219–227.
- Bedoschi, G.; Roque, M.; Esteves, S.C. ICSI and Male Infertility: Consequences to Offspring. In Male Infertility; Springer International Publishing: Cham, 2020; pp. 767–775. [Google Scholar]
- Duranthon, V.; Chavatte-Palmer, P. Long Term Effects of ART: What Do Animals Tell Us? Mol Reprod Dev 2018, 85, 348–368. [Google Scholar] [CrossRef] [PubMed]
- Pontes, J.H.F.; Nonato-Junior, I.; Sanches, B.V.; Ereno-Junior, J.C.; Uvo, S.; Barreiros, T.R.R.; Oliveira, J.A.; Hasler, J.F.; Seneda, M.M. Comparison of Embryo Yield and Pregnancy Rate between in Vivo and in Vitro Methods in the Same Nelore (Bos Indicus) Donor Cows. Theriogenology 2009, 71, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Block, J.; Seidel, G.E.; Brink, Z.; McSweeney, K.; Farin, P.W.; Bonilla, L.; Hansen, P.J. Pregnancy Rates of Lactating Cows after Transfer of in Vitro Produced Embryos Using X-Sorted Sperm. Theriogenology 2013, 79, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, É.; Marques, C.C.; Pimenta, J.; Jorge, J.; Baptista, M.C.; Gonçalves, A.C.; Pereira, R.M.L.N. Anti-Aging Effect of Urolithin a on Bovine Oocytes in Vitro. Animals 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, H.; Takahashi, T.; Takahashi, E.; Tezuka, N.; Nakahara, K.; Takahashi, K.; Kurachi, H. Aged Mouse Oocytes Fail to Readjust Intracellular Adenosine Triphosphates at Fertilization. Biol Reprod 2005, 72, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.M.; Marques, C.C.; Baptista, M.C.; Vasques, M.I.; Horta, A.E.M. Embryos and Culture Cells: A Model for Studying the Effect of Progesterone. Anim Reprod Sci 2009, 111, 31–40. [Google Scholar] [CrossRef]
- De Munck, N.; Janssens, R.; Segers, I.; Tournaye, H.; Van De Velde, H.; Verheyen, G. Influence of Ultra-Low Oxygen (2%) Tension on in-Vitro Human Embryo Development. Human Reproduction 2019, 34, 228–234. [Google Scholar] [CrossRef]
- Cohen, J.; Rieger, D.; Mastenbroek, S.; Meintjes, M.; Janssens, R.; Catt, J.; Morbeck, D.; Mortimer, D.; Fawzy, M.; Alikani, M.; et al. ‘There Is Only One Thing That Is Truly Important in an IVF Laboratory: Everything’ Cairo Consensus Guidelines on IVF Culture Conditions. Reprod Biomed Online 2020, 40, 33–60. [Google Scholar] [CrossRef]
- Mortimer, D.; Cohen, J.; Mortimer, S.T.; Fawzy, M.; McCulloh, D.H.; Morbeck, D.E.; Pollet-Villard, X.; Mansour, R.T.; Brison, D.R.; Doshi, A.; et al. Cairo Consensus on the IVF Laboratory Environment and Air Quality: Report of an Expert Meeting. In Proceedings of the Reproductive BioMedicine Online; Elsevier Ltd, June 1 2018; Vol. 36; pp. 658–674. [Google Scholar]
- Marques, C.C.; Santos-Silva, C.; Rodrigues, C.; Matos, J.E.; Moura, T.; Baptista, M.C.; Horta, A.E.M.; Bessa, R.J.B.; Alves, S.P.; Soveral, G.; et al. Bovine Oocyte Membrane Permeability and Cryosurvival: Effects of Different Cryoprotectants and Calcium in the Vitrification Media. Cryobiology 2018, 81, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Tay, J.I.; Rutherford\, A.J.; Killick, S.R.; Maguiness, S.D.; Partridge, R.J.; Leese, H.J. Human Tubal Fluid: Production, Nutrient Composition and Response to Adrenergic Agents. Human Reproduction vol 1997, 12, 2451–2456. [Google Scholar] [CrossRef]
- Jorge, M.; Ferreira, F.C.; Marques, C.C.; Batista, M.C.; Oliveira, P.J.; Lidon, F.; Duarte, S.C.; Teixeira, J.; Pereira, R.M.L.N. Effect of Urolithin A on Bovine Sperm Capacitation and In Vitro Fertilization. Animals 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Montezinho, L.; Cagide, F.; Borges, F.; Oliveira, P.J.; Pereira, R.M.L.N. Effect of a Novel Hydroxybenzoic Acid Based Mitochondria Directed Antioxidant Molecule on Bovine Sperm Function and Embryo Production. Animals 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.L.M.; Day, M.L.; Morris, M.B. Redox Regulation and Oxidative Stress in Mammalian Oocytes and Embryos Developed in Vivo and in Vitro. Int J Environ Res Public Health 2021, 18. [Google Scholar] [CrossRef]
- Sousa, A.P.; Amaral, A.; Baptista, M.; Tavares, R.; Campo, P.C.; Peregrín, P.C.; Freitas, A.; Paiva, A.; Almeida-Santos, T.; Ramalho-Santos, J. Not All Sperm Are Equal: Functional Mitochondria Characterize a Subpopulation of Human Sperm with Better Fertilization Potential. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Agrawal, A.; Mabalirajan, U. Rejuvenating Cellular Respiration for Optimizing Respiratory Function: Targeting Mitochondria. Am J Physiol Lung Cell Mol Physiol 2016, 310, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C.; Fan, W.; Procaccio, V. Mitochondrial Energetics and Therapeutics. Annual Review of Pathology: Mechanisms of Disease 2010, 5, 297–348. [Google Scholar] [CrossRef]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial Respiratory Chain Complexes. In; 2018; pp. 167–227.
- Schofield, J.H.; Schafer, Z.T. Mitochondrial Reactive Oxygen Species and Mitophagy: A Complex and Nuanced Relationship. Antioxid Redox Signal 2021, 34, 517–530. [Google Scholar] [CrossRef]
- Qi, L.; Chen, X.; Wang, J.; Lv, B.; Zhang, J.; Ni, B.; Xue, Z. Mitochondria: The Panacea to Improve Oocyte Quality? Ann Transl Med 2019, 7, 789–789. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Whiting, S.; De Iuliis, G.N.; McClymont, S.; Mitchell, L.A.; Baker, M.A. Electrophilic Aldehydes Generated by Sperm Metabolism Activate Mitochondrial Reactive Oxygen Species Generation and Apoptosis by Targeting Succinate Dehydrogenase. Journal of Biological Chemistry 2012, 287, 33048–33060. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and Consequences of Oxidative Stress in Spermatozoa. Reprod Fertil Dev 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Koppers, A.J.; De Iuliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of Mitochondrial Reactive Oxygen Species in the Generation of Oxidative Stress in Spermatozoa. Journal of Clinical Endocrinology and Metabolism 2008, 93, 3199–3207. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.; Sousa, A.P.; Fernandes, R.; Ferreira, A.F.; Almeida-Santos, T.; Ramalho-Santos, J. Aging-Related Mitochondrial Alterations in Bovine Oocytes. Theriogenology 2020, 157, 218–225. [Google Scholar] [CrossRef]
- Aitken, R.J. Impact of Oxidative Stress on Male and Female Germ Cells: Implications for Fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef] [PubMed]
- Showell, M.G.; Brown, J.; Clarke, J.; Hart, R.J. Antioxidants for Female Subfertility. Cochrane Database of Systematic Reviews 2013, 2013. [Google Scholar] [CrossRef]
- Ishigami, M. Superoxide Dismutase. Nippon rinsho. Japanese journal of clinical medicine 1998, 56 Suppl 3, 160–168. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Concept and Some Practical Aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Teng, S.; Saunders, P.T.K. A Single, Mild, Transient Scrotal Heat Stress Causes Hypoxia and Oxidative Stress in Mouse Testes, Which Induces Germ Cell Death. Biol Reprod 2009, 80, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Gad, A.; Salilew-Wondim, D.; Prastowo, S.; Held, E.; Hoelker, M.; Rings, F.; Tholen, E.; Neuhoff, C.; Looft, C.; et al. Bovine Embryo Survival under Oxidative-Stress Conditions Is Associated with Activity of the NRF2-Mediated Oxidative-Stress-Response Pathway. Mol Reprod Dev 2014, 81, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Leite, R.F.; Annes, K.; Ispada, J.; De Lima, C.B.; Dos Santos, É.C.; Fontes, P.K.; Nogueira, M.F.G.; Milazzotto, M.P. Oxidative Stress Alters the Profile of Transcription Factors Related to Early Development on in Vitro Produced Embryos. Oxid Med Cell Longev 2017, 2017. [Google Scholar] [CrossRef]
- Nakai, A.; Fukushima, Y.; Yamamoto, A.; Amatsu, Y.; Chen, X.; Nishigori, M.; Yoshioka, Y.; Kaneko, M.; Koshiba, T.; Watanabe, T. Increased ROS Levels in Mitochondrial Outer Membrane Protein Mul1-deficient Oocytes Result in Abnormal Preimplantation Embryogenesis. FEBS Lett 2024, 598, 1740–1752. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.T.; Lysiak, J.J. Oxidative Stress: A Common Factor in Testicular Dysfunction. J Androl 2008, 29, 488–498. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Scarlata, E. Oxidative Stress and Reproductive Function: The Protection of Mammalian Spermatozoa against Oxidative Stress. Reproduction 2022, 164, F67–F78. [Google Scholar] [CrossRef] [PubMed]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal Prospects of Antioxidants: A Review. Eur J Med Chem 2019, 178, 687–704. [Google Scholar] [CrossRef]
- Burton, G.J.; Hempstock, J.; Jauniaux, E. Oxygen, Early Embryonic Metabolism and Free Radical-Mediated Embryopathies. Reprod Biomed Online 2003, 6, 84–96. [Google Scholar] [CrossRef]
- Grimm, A.; Eckert, A. Brain Aging and Neurodegeneration: From a Mitochondrial Point of View. J Neurochem 2017, 143, 418–431. [Google Scholar] [CrossRef]
- Smith, R.A.J.; Hartley, R.C.; Cochemé, H.M.; Murphy, M.P. Mitochondrial Pharmacology. Trends Pharmacol Sci 2012, 33, 341–352. [Google Scholar] [CrossRef]
- Pagano, G.; Aiello Talamanca, A.; Castello, G.; Cordero, M.D.; D’Ischia, M.; Gadaleta, M.N.; Pallardó, F. V.; Petrović, S.; Tiano, L.; Zatterale, A. Oxidative Stress and Mitochondrial Dysfunction across Broad-Ranging Pathologies: Toward Mitochondria-Targeted Clinical Strategies. Oxid Med Cell Longev 2014, 2014. [Google Scholar] [CrossRef]
- Ogata, K.; Imai, A.; Sato, S.; Nishino, K.; Watanabe, S.; Somfai, T.; Kobayashi, E.; Takeda, K. Effects of Reduced Glutathione Supplementation in Semen Freezing Extender on Frozen-Thawed Bull Semen and in Vitro Fertilization. Journal of Reproduction and Development 2022, 68, 2022. [Google Scholar] [CrossRef] [PubMed]
- Adeoye, O.; Olawumi, J.; Opeyemi, A.; Christiania, O. Review on the Role of Glutathione on Oxidative Stress and Infertility. J Bras Reprod Assist 2018, 22, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.M.; Baptista, M.C.; Vasques, M.I.; Horta, A.E.M.; Portugal, P.V.; Bessa, R.J.B.; Silva, J.C. e; Pereira, M.S.; Marques, C.C. Cryosurvival of Bovine Blastocysts Is Enhanced by Culture with Trans-10 Cis-12 Conjugated Linoleic Acid (10t,12c CLA). Anim Reprod Sci 2007, 98, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.-H.; Zhao, X.-L.; Tian, W.-Q.; Zan, L.-S.; Li, Q.-W. Effects of Vitamin E Supplementation in the Extender on Frozen-Thawed Bovine Semen Preservation. Animal 2011, 5, 107–112. [Google Scholar] [CrossRef]
- Arkoun, B.; Galas, L.; Dumont, L.; Rives, A.; Saulnier, J.; Delessard, M.; Rondanino, C.; Rives, N. Vitamin E but Not GSH Decreases Reactive Oxygen Species Accumulation and Enhances Sperm Production during In Vitro Maturation of Frozen-Thawed Prepubertal Mouse Testicular Tissue. Int J Mol Sci 2019, 20, 5380. [Google Scholar] [CrossRef]
- NIKI, E. Interaction of Ascorbate and A-Tocopherol. Ann N Y Acad Sci 1987, 498, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, S. Coenzyme Q10 One Antioxidant, Many Promising Applications. Alternative & Complementary Therapies 2009, 9, 111–116. [Google Scholar] [CrossRef]
- Ernster, L.; Forsmark, P.; Nordenbrand1, K. The Mode of Action of Lipid-Soluble Antioxidants in Biological Membranes. Relationship between the Effects of Ubiquinol and Vitamin E as Inhibitors of Lipid Peroxidation in Submitochondrial Particles. 1992. [Google Scholar]
- Yildirim, R.M.; Seli, E. Mitochondria as Therapeutic Targets in Assisted Reproduction. Human Reproduction 2024, 39, 2147–2159. [Google Scholar] [CrossRef] [PubMed]
- Oroian, M.; Escriche, I. Antioxidants: Characterization, Natural Sources, Extraction and Analysis. Food Research International 2015, 74, 10–36. [Google Scholar] [CrossRef]
- Chavarria, D.; Silva, T.; MagalhãesE Silva, D.; Remiaõ, F.; Borges, F. Lessons from Black Pepper: Piperine and Derivatives Thereof. Expert Opin Ther Pat 2016, 26, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Ray, A.K.; Mishra, S.K. Molecular and Pharmacological Aspects of Piperine as a Potential Molecule for Disease Prevention and Management: Evidence from Clinical Trials. Beni Suef Univ J Basic Appl Sci 2022, 11. [Google Scholar] [CrossRef]
- Lim, J.S.; Lee, D.Y.; Lim, J.H.; Oh, W.K.; Park, J.T.; Park, S.C.; Cho, K.A. Piperine: An Anticancer and Senostatic Drug. Frontiers in Bioscience - Landmark 2022, 27. [Google Scholar] [CrossRef]
- Ferreira, F.; Oliveira, A.; Lindon, F.; Oliveira, P.O.; Teixeira, J.; Pereira, R.M.L.N. ;. Effect of the Natural Antioxidant Piperine on Maturation of Bovine Oocyte Production and Embryo Production. In Proceedings of the XIV Congresso Ibérico sobre Recursos Genéticos Animais; Vila Real, 2024. p. 104.
- Teixeira, J.; Oliveira, C.; Amorim, R.; Cagide, F.; Garrido, J.; Ribeiro, J.A.; Pereira, C.M.; Silva, A.F.; Andrade, P.B.; Oliveira, P.J.; et al. Development of Hydroxybenzoic-Based Platforms as a Solution to Deliver Dietary Antioxidants to Mitochondria. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Leopoldini, M.; Russo, N.; Toscano, M. The Molecular Basis of Working Mechanism of Natural Polyphenolic Antioxidants. Food Chem 2011, 125, 288–306. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am J Clin Nutr 2004, 79, 727–774. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Li, S.; Lin, C.C. Effect of Resveratrol and Pterostilbene on Aging and Longevity. BioFactors 2018, 44, 69–82. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, D.; Gao, L.; Wang, Y.; Wang, Y.; Jia, R.; Bai, Y.; Shi, D.; Lu, F. Effects of Astaxanthin on the Physiological State of Porcine Ovarian Granulose Cells Cultured In Vitro. Antioxidants 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Rostami, S.; Alyasin, A.; Saedi, M.; Nekoonam, S.; Khodarahmian, M.; Moeini, A.; Amidi, F. Astaxanthin Ameliorates Inflammation, Oxidative Stress, and Reproductive Outcomes in Endometriosis Patients Undergoing Assisted Reproduction: A Randomized, Triple-Blind Placebo-Controlled Clinical Trial. Front Endocrinol (Lausanne) 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F.; Lei, J.; Li, Q.; Zhou, B. Activation of the MiR-34a-Mediated SIRT1/MTOR Signaling Pathway by Urolithin A Attenuates d-Galactose-Induced Brain Aging in Mice. Neurotherapeutics 2019, 16, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, D.; Andreux, P.A.; Valdés, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol Med 2021, 27, 687–699. [Google Scholar] [CrossRef]
- Teixeira, C.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Cagide, F.; Borges, F.; Montezinho, L.; Oliveira, P.; Pereira, R.M.L.N. Beneficial Effect of Mitochondriotropic Antioxidants on Oocyte Maturation and Embryo Production. Eur J Clin Invest 2020, 50, 34–35. [Google Scholar] [CrossRef]
- Tauskela JS MitoQ--a Mitochondria-Targeted Antioxidant. IDrugs 2007, 10, 399–412.
- Marei, W.F.A.; Van Den Bosch, L.; Pintelon, I.; Mohey-Elsaeed, O.; Bols, P.E.J.; Leroy, J.L.M.R. Mitochondria-Targeted Therapy Rescues Development and Quality of Embryos Derived from Oocytes Matured under Oxidative Stress Conditions: A Bovine in Vitro Model. Human Reproduction 2019, 34, 1984–1998. [Google Scholar] [CrossRef]
- Shirzeyli, M.H.; Amidi, F.; Shamsara, M.; Nazarian, H.; Eini, F.; Shirzeyli, F.H.; Zolbin, M.M.; Novin, M.G.; Joupari, M.D. Exposing Mouse Oocytes to Mitoq during in Vitro Maturation Improves Maturation and Developmental Competence. Iran J Biotechnol 2020, 18, 12–21. [Google Scholar] [CrossRef]
- Elkhawagah, A.R.; Donato, G.G.; Poletto, M.; Martino, N.A.; Vincenti, L.; Conti, L.; Necchi, D.; Nervo, T. Effect of Mitoquinone on Sperm Quality of Cryopreserved Stallion Semen. J Equine Vet Sci 2024, 141, 105168. [Google Scholar] [CrossRef] [PubMed]
- Câmara, D.R.; Ibanescu, I.; Siuda, M.; Bollwein, H. Mitoquinone Does Not Improve Sperm Cryo-resistance in Bulls. Reproduction in Domestic Animals 2022, 57, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.; Cagide, F.; Benfeito, S.; Soares, P.; Garrido, J.; Baldeiras, I.; Ribeiro, J.A.; Pereira, C.M.; Silva, A.F.; Andrade, P.B.; et al. Development of a Mitochondriotropic Antioxidant Based on Caffeic Acid: Proof of Concept on Cellular and Mitochondrial Oxidative Stress Models. J Med Chem 2017, 60, 7084–7098. [Google Scholar] [CrossRef]
- Oliveira, C.; Cagide, F.; Teixeira, J.; Amorim, R.; Sequeira, L.; Mesiti, F.; Silva, T.; Garrido, J.; Remião, F.; Vilar, S.; et al. Hydroxybenzoic Acid Derivatives as Dual-Target Ligands: Mitochondriotropic Antioxidants and Cholinesterase Inhibitors. Front Chem 2018, 6. [Google Scholar] [CrossRef]
- Benfeito, S.; Oliveira, C.; Fernandes, C.; Cagide, F.; Teixeira, J.; Amorim, R.; Garrido, J.; Martins, C.; Sarmento, B.; Silva, R.; et al. Fine-Tuning the Neuroprotective and Blood-Brain Barrier Permeability Profile of Multi-Target Agents Designed to Prevent Progressive Mitochondrial Dysfunction. Eur J Med Chem 2019, 167, 525–545. [Google Scholar] [CrossRef]
- Teixeira, J.; Basit, F.; Willems, P.H.G.M.; Wagenaars, J.A.; van de Westerlo, E.; Amorim, R.; Cagide, F.; Benfeito, S.; Oliveira, C.; Borges, F.; et al. Mitochondria-Targeted Phenolic Antioxidants Induce ROS-Protective Pathways in Primary Human Skin Fibroblasts. Free Radic Biol Med 2021, 163, 314–324. [Google Scholar] [CrossRef]
- Amorim, R.; Cagide, F.; Tavares, L.C.; Simões, R.F.; Soares, P.; Benfeito, S.; Baldeiras, I.; Jones, J.G.; Borges, F.; Oliveira, P.J.; et al. Mitochondriotropic Antioxidant Based on Caffeic Acid AntiOxCIN4 Activates Nrf2-Dependent Antioxidant Defenses and Quality Control Mechanisms to Antagonize Oxidative Stress-Induced Cell Damage. Free Radic Biol Med 2022, 179, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Videira, A.J.C.; Veloso, C.D.; Benfeito, S.; Soares, P.; Martins, J.D.; Gonçalves, B.; Duarte, J.F.S.; Santos, A.M.S.; Oliveira, P.J.; et al. Cytotoxicity and Mitochondrial Effects of Phenolic and Quinone-based Mitochondria-targeted and Untargeted Antioxidants on Human Neuronal and Hepatic Cell Lines: A Comparative Analysis. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Ferreira FC; Sousa A; Marques CC; Baptista MC; Teixeira J; Cagide F; Borges F; Oliveira P; Pereira RMLN Effect of a Mitochondriotropic Antioxidant Based on Caffeic Acid (AntiOxCIN4) on Spermatozoa Capacitation and in Vitro Fertilization (Poster). In Proceedings of the Congresso CIISA “Inovação em Pesquisa Animal, Veterinária e Biomédica” - 10-11 novembro; 2022. Pp 10.
- Lourenço B; Ferreira F; Marques CC; Batista MC; Teixeira J; Cagide F; Borges F; Oliveira P; Pereira RMLN Natural Derived Mitochondriotropic Molecules Improve Embryo Quality and Cryosurvival (Poster). In Proceedings of the XIII CONGRESO IBÉRICO SERGA/SPREGA SOBRE RECURSOS GENÉTICOS ANIMALES - 21-23 outubro; 2022.
- Lourenço, B. Effect of Mitochondriotropic Molecules in Reducing Oxidative Stress in Assisted Reprodution Techniques (Tese de Mestrado), Universidade de Lisboa - Faculdade de Ciências: Lisboa, 2020.
Figure 1.
Impact of environmental stress on animal welfare and fertility, as well as on biodiversity (Created in Biorender.com).
Figure 1.
Impact of environmental stress on animal welfare and fertility, as well as on biodiversity (Created in Biorender.com).
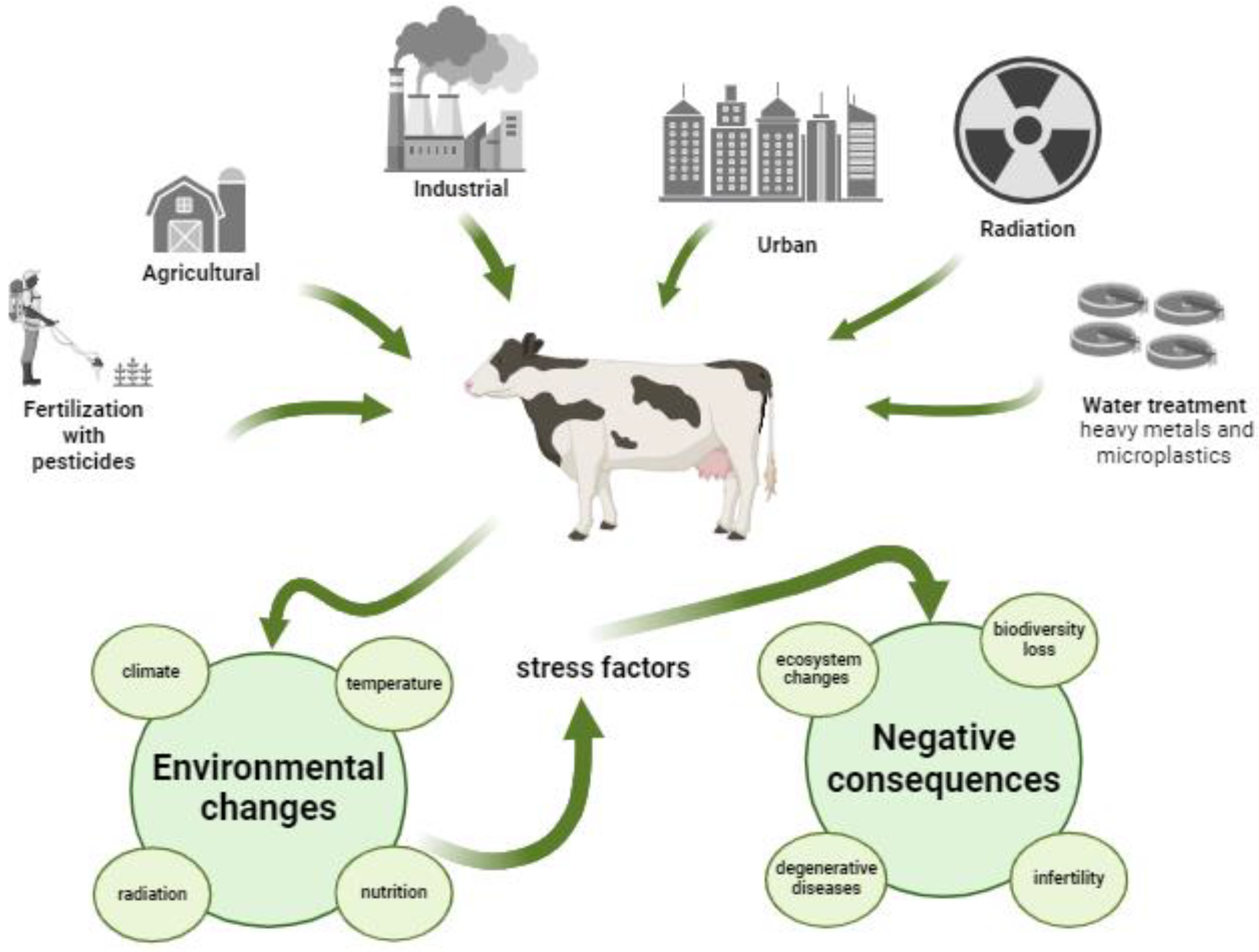
Figure 2.
Effects of environmental stress factors on oocyte mitochondria. Stress induces cellular and molecular changes in the oocyte, which in turn can reduce the gamete's developmental competence. It has been shown that supplementation with mitochondrial agents, antioxidants [epigallocatechin gallate, melatonin], or injection of mitochondria into oocytes, attenuates these effects and improves competence for subsequent development (adapted from [9] and created in Biorender.com).
Figure 2.
Effects of environmental stress factors on oocyte mitochondria. Stress induces cellular and molecular changes in the oocyte, which in turn can reduce the gamete's developmental competence. It has been shown that supplementation with mitochondrial agents, antioxidants [epigallocatechin gallate, melatonin], or injection of mitochondria into oocytes, attenuates these effects and improves competence for subsequent development (adapted from [9] and created in Biorender.com).
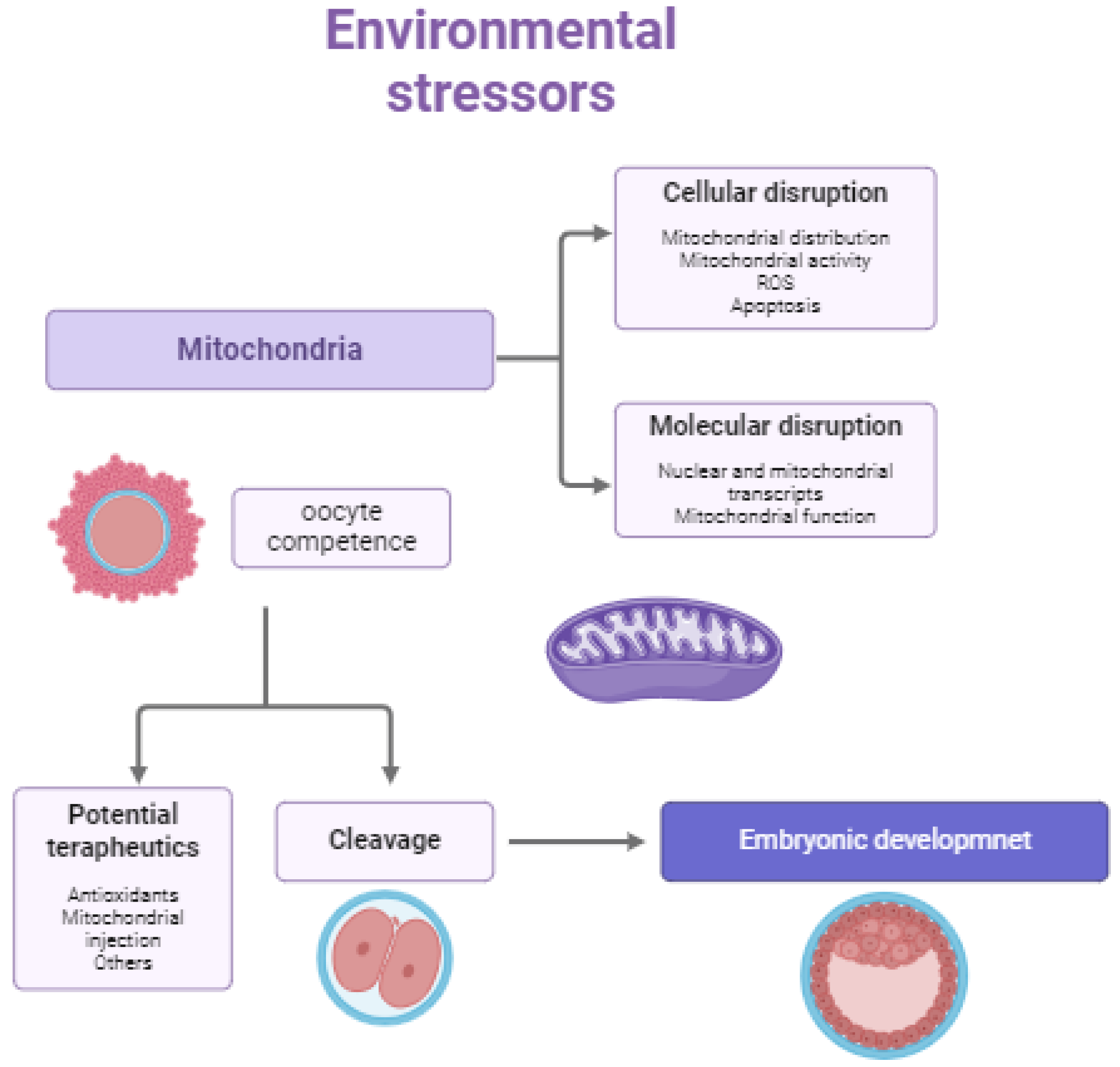
Figure 3.
Respiratory chain in mitochondria - During aerobic respiration, electrons from NADH and succinate are transferred to molecular O2 through a chain of redox enzymes (Complexes I-IV) embedded in the inner mitochondrial membrane, which is accompanied by pump protons across the inner membrane, generating an electrochemical gradient. This proton gradient is used for the mechanical work needed to produce ATP from ADP and inorganic phosphate (Created in Biorender.com).
Figure 3.
Respiratory chain in mitochondria - During aerobic respiration, electrons from NADH and succinate are transferred to molecular O2 through a chain of redox enzymes (Complexes I-IV) embedded in the inner mitochondrial membrane, which is accompanied by pump protons across the inner membrane, generating an electrochemical gradient. This proton gradient is used for the mechanical work needed to produce ATP from ADP and inorganic phosphate (Created in Biorender.com).
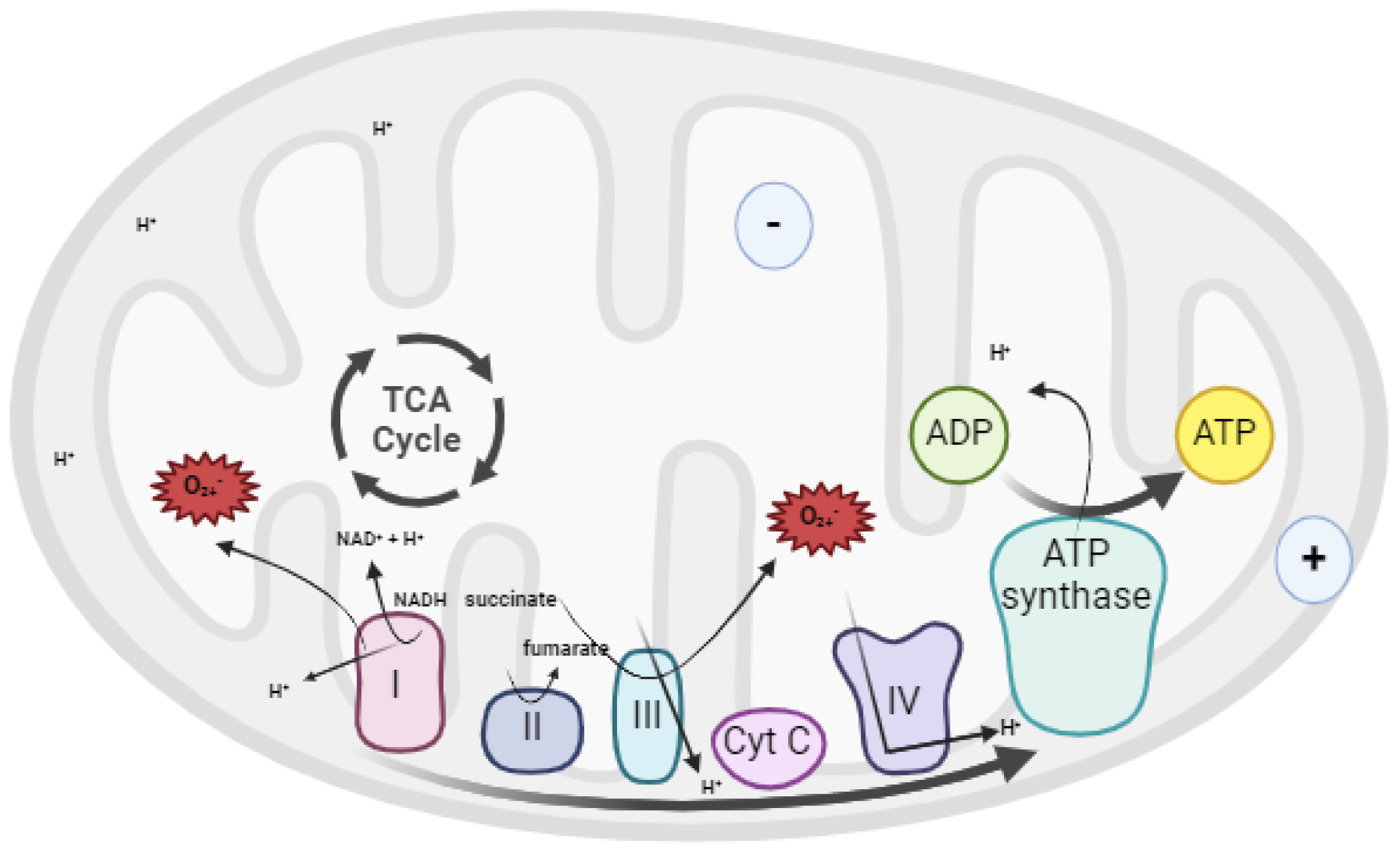
Figure 4.
Chemical structures of endogenous antioxidants evaluated in ART context. Glutathione (GSH), Ubiquinone (CoQ10), Melatonin, and Vitamin E have been tested in bovine germplasm to improve fertility.
Figure 4.
Chemical structures of endogenous antioxidants evaluated in ART context. Glutathione (GSH), Ubiquinone (CoQ10), Melatonin, and Vitamin E have been tested in bovine germplasm to improve fertility.
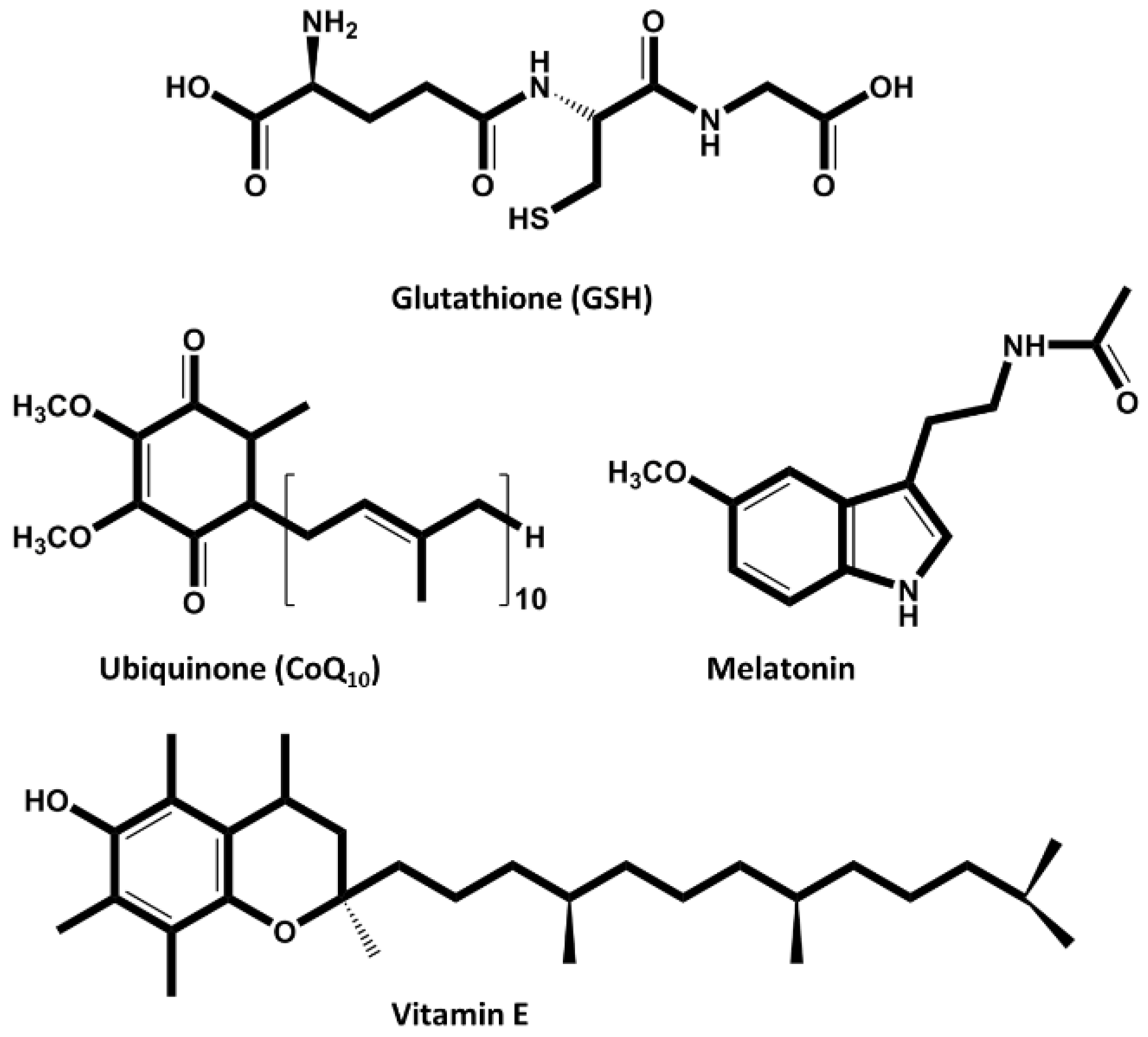
Figure 5.
Exogenous naturally occurring antioxidants used to improve ART outcomes – Piperine, Astahantine and Urolithin A have been tested in animal germplasm to improve fertility (created in BioRender.com).
Figure 5.
Exogenous naturally occurring antioxidants used to improve ART outcomes – Piperine, Astahantine and Urolithin A have been tested in animal germplasm to improve fertility (created in BioRender.com).
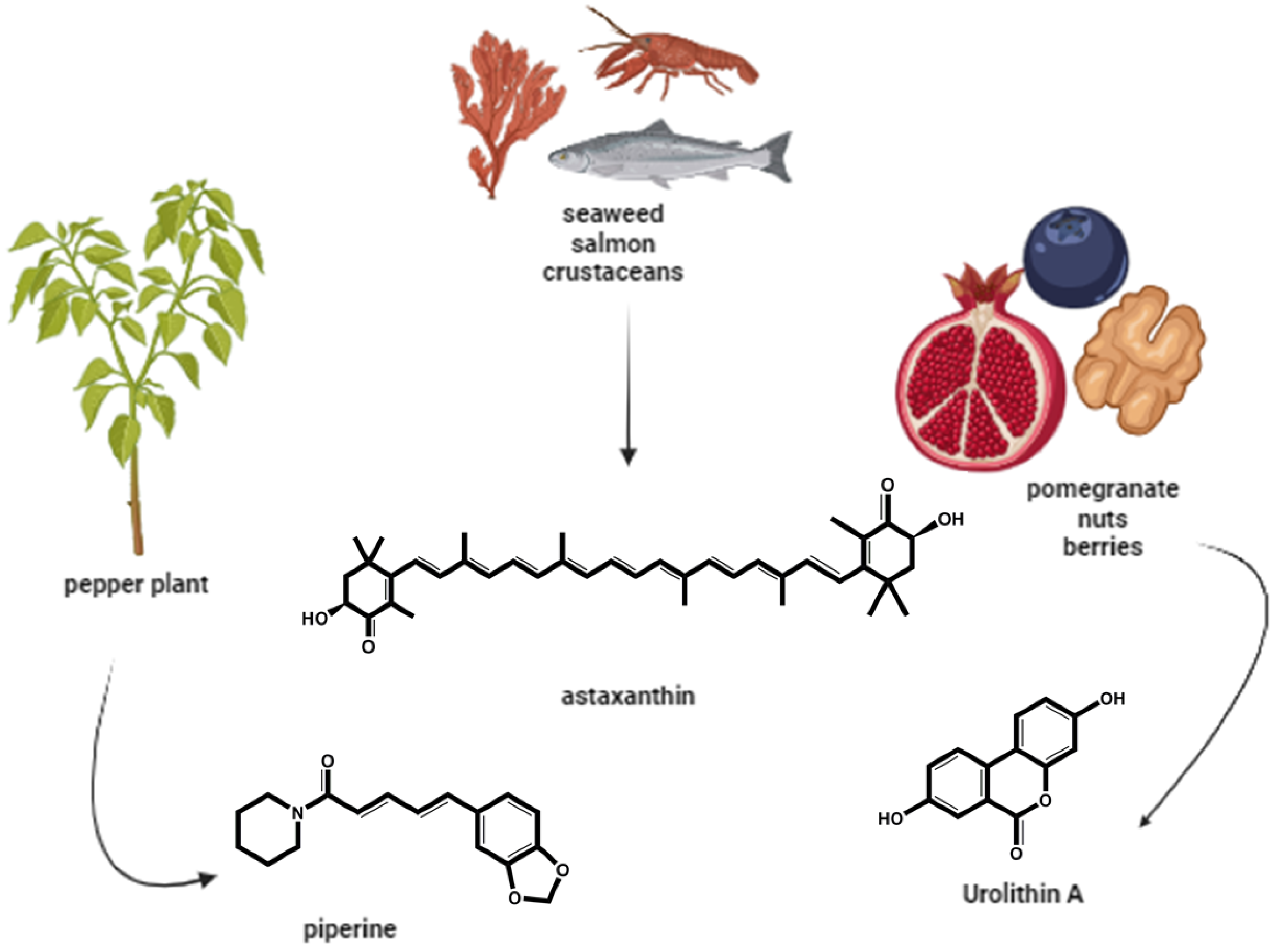
Figure 6.
Chemical structures of synthetic mitochondriotropic antioxidants evaluated in ART context. AntiOxBEN2, AntiOxCIN4, and MitoQ10 have been tested in bovine cells to decreased ROS and increase the results in ART.
Figure 6.
Chemical structures of synthetic mitochondriotropic antioxidants evaluated in ART context. AntiOxBEN2, AntiOxCIN4, and MitoQ10 have been tested in bovine cells to decreased ROS and increase the results in ART.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



