
Submitted:
13 December 2024
Posted:
13 December 2024
You are already at the latest version
Abstract
A new species of Asian horned toads Boulenophrys is described from Yongzhou City, Hunan Province, China. The species phylogenetic sister to B. yunkaiensis based on 16S rRNA and COI genes. The new species differs from its congeners by following combination of characters: (1) small body size, SVL 37.6–40.2 mm (38.9 ± 1.3, n = 7) in adult males and SVL 41.8–45.9 mm (43.6 ± 2.1, n = 3) in adult females; (2) tympanum boundary clear, TD/ED 0.48–0.57 in males, 0.47–0.57 in females; (3) presence of small horn-like tubercle at the edge of upper eyelid; (4) vomerine ridge present, vomerine teeth absent; (5) margin of tongue rounded, not notched posteriorly; (6) dorsal skin rough, a discontinuous “V”-shaped ridge with two discontinuous dorsolateral ridges on two side on the back, dense tubercles on skin of ventral surface of dorsal shank and thigh, spiny tubercles surrounding the cloaca; (7) hindlimbs slender, heels overlapping when the flexed hindlimbs are held at right angles to the body axis, tibio-tarsal articulation reaching forward between anterior margin of tympanum and posterior corner of eye when leg stretched forward; (8) relative finger length IV < II < I < III, a subarticular tubercle present at the base of each finger; (9)supernumerary tubercles below the base of I and II toes distinct; (10) toes without lateral fringes and with only rudimentary webbing, webbing formula: I1 – 1-II1 – 2-III2 – 3IV3- – 2V.
Keywords:
Asian horned toads
; taxonomy
; Hunan Province
; Boulenophrys dupanglingensis sp. nov
1. Introduction
The subfamily Megophryinae is endemic to Asia, with a distribution spanning the Himalayas, northeastern India, southern China, and extending southwards to the islands of Southeast Asia [1,2,3]. The classification of Megophryinae has been a subject of debate for decades. Molecular phylogenetic studies consistently indicate that it is a monophyletic group [4,5,6]. However, recent years have been numerous alternations in the generic relationships and divisions within this subfamily [4,5,6,7,8,9,10]. Lyu et al. proposed a new classification system, identifying ten genera: Atympanophrys Tian and Hu, 1983, Brachytarsophrys Tian and Hu, 1983, Grillitschia Dubois, Ohler, and Pyron, 2021, Sarawakiphrys Lyu and Wang, 2023, Jingophrys Lyu and Wang, 2023, Xenophrys Günther, 1864, Megophrys Kuhl and Van Hasselt, 1822, Pelobatrachus Beddard, 1907, Ophryophryne Boulenger, 1903, Boulenophrys Fei, Ye, and Jiang, 2016[7,10,11,12,13,14,15,16]. Among these, Boulenophrys is the most diverse, comprising 69 recognized species to date (Table 1).
Dupangling National Nature Reserve is located in the southwest of Yongzhou City, Hunan Province, within the Nanling Mountains. During herpetological surveys conducted in 2022, we discovered an undescribed species in Daoxian County and Jiangyong County. Based on the diagnostic characters established by Lyu et al.[10], these specimens belong to the genus Boulenophrys, however, they could not be assigned to any recognized species due to their unique combination of morphological characteristics. Subsequent molecular phylogenetic analysis confirmed that these specimens represent a distinct evolutionary lineage. Accordingly, we describe them here as a new species within the genus Boulenophrys.
2. Materials and Methods
2.1. Sampling
Field surveys were conducted in Dupangling National Nature Reserve, Hunan Province, China (Figure 1), in April 2022. A total of ten specimens were newly collected. All specimens were initially fixed in 10% formalin and then transferred to 75% ethanol for storage. Before being fixed in formalin, liver tissues were sampled from four specimens and preserved in 95% ethanol for DNA extraction. Nine specimens were deposited in the Animal Museum of Life Sciences College of Hunan Normal University (HUNU), and one museum specimen was preserved in 75% ethanol and deposited at Chengdu Institute of Biology (CIB), Chinese Academy of Sciences, Chengdu City, Sichuan Province, China.
2.2. Molecular and Phylogenetic Analyses
Genomic DNA was extracted from 95% ethanol-preserved liver tissues using a TSINGKE DNA extraction kit. The mitochondrial genes, specifically the partial 16S ribosomal RNA gene (16S rRNA) and partial cytochrome c oxidase 1 gene (COI) were sequenced for three samples (HUNU 22SA05, HUNU 22SA07, HUNU 22SA08 and CIB2016050802). Polymerase Chain Reaction (PCR) amplifications were conducted following Liu et al. [5]. All PCR products were sequenced on an ABI 3730 XL Genetic Analyzer at the Sangon Biotech Co., Ltd (Shanghai), and the resulting sequences were deposited in GenBank.
For phylogenetic analyses, sequences were obtained from GenBank, including 68 known Boulenophrys species (except B.shuichengensis for which no sequence information is availablle) and two outgroup species (Xenophrys glandulosa and X. mangshanensis) (Table S1). All sequences were aligned using the Clustal W algorithm with default parameters [54] and trimmed with the partial gaps deletion in MEGA 11 [55]. In highly variable regions, all gaps were removed. The best-fit evolutionary model (GTR+G+I) was selected using PartitionFinder v. 2.1.1 under the AIC criterion [56]. The sequenced data were analyzed using Bayesian inference (BI) with MrBayes 3.2 [57], and phylogenetic trees were generated using maximum likelihood (ML) methods implemented in PhyML v. 3.0[58]. The BI analysis was conducted with 10,000,000 generations, sampling every 1000 generations, with the first 25% of samples discarded as burn-in, resulting in a potential scale reduction factor (PSRF) of < 0.005. In the ML analysis, a bootstrap consensus tree inferred from 1000 replicates was generated.
2.3. Morphological Analysis
The definition of morphological characters follows to Lyu et al. [10]. The following measurements were taken with digital calipers to the nearest 0.1 mm : SVL, snout-vent length (from the tip of snout to the posterior edge of the vent); HDL, head length (from the tip of snout to the articulation of the jaw); HDW, head width (head width at the commissure of the jaws); SNT, snout length (from the tip of snout to the anterior corner of the eye); IND, internasal distance (distance between nares); IOD, interorbital distance (minimum distance between upper eyelids); ED, eye diameter (from the anterior corner of the eye to posterior corner of the eye); TD, tympanum diameter (horizontal diameter of tympanum); TED, tympanum-eye distance (from the anterior edge of tympanum to posterior corner of the eye); HND, hand length (from the proximal border of the outer palmar tubercle to the tip of digit III); RAD, radio-ulna length (from the flexed elbow to the proximal border of the outer palmar tubercle); FTL, foot length (from the distal end of the shank to the tip of digit IV); TIB, tibial length (from the outer surface of the flexed knee to the heel). The webbing was expressed using the webbed formula[59], where Roman numerals refer to the fingers and toes. Sex was determined by direct observation of calls or the presence of internal vocal sac openings in males. Presence or absence of nuptial pads was examined under a dissecting microscope. Diagnostic characters distinguishing all 69 species of Boulenophrys are shown in Table 2.
3. Results
3.1. Molecular Phylogenetic Analyses
A total of 540 base pair (bp) of 16S and 636 bp of COI, were concatenated seriatim into a single1196 bp single sequence. Phylogenetic analyses using ML and BI methods produced nearly identical results, with most major nodes well supported on both the ML and Bayesian trees. The Bayesian tree is presented in (Figure 2), with ML bootstrap values (BS) ≥ 70 and BI posterior probabilities (BPP) ≥ 0.90 considered indicative of strong support for major nodes. In different analyses, the newly collected samples were clustered into a monophyletic lineage with robust nodal support (BS 100, BPP 1.00) and exhibited low intraspecific genetic divergence (mean p-distances 4.2 % in COI). Additionally, these specimens can be distinguished from all congeners by a unique combination of morphological characters (see Taxonomic account below).
3.2. Morphological Comparisons
Comparative data of Boulenophrys dupanglingensis sp. nov. with other recognized species of the genus Boulenophrys are listed in Table 2.
Boulenophrys dupanglingensissp. nov. is phylogenetically closed to B. yunkaiensis. It differs from B. yunkaiensis by (1) relatively shorter shanks, TIB/SVL 0.37–0.39 (0.38 ± 0.01, n=7) in adult males (vs. TIB/SVL 0.40–0.48, 0.44 ± 0.04, n=3), 0.32–0.36 in adult female(0.34± 0.02, n=3) (vs. TIB/SVL 0.42–0.46 in adult, 0.44 ±0.02, n=3 for B. yunkaiensis); (2) distinct supernumerary tubercles below the base of I and II toes (vs. indistinct under all toes in B. yunkaiensis); (3) rough dorsum skin with several dense small granules on the flanks (vs. sparse larger granules on the flanks) (Figure 3). These characteristics make Boulenophrys dupanglingensis sp. nov. easily distinguished from B. yunkaiensis.
With relatively longer shanks, and heels overlapping when the flexed hindlimbs are held at right angles to the body axis, Boulenophrys dupanglingensis sp. nov. is easily distinguished from the following 16 congeners, B. acuta, B. brachykolos, B. daoji, B. dongguanensis, B. fengshunensis, B. hungtai, B. hengshanesis, B. insularis, B. kuatunensis, B.lichun, B. nankunensis, B. obesa, B. ombrophila, B. puningensis and B. wugongensis (vs. all of which have relatively shorter shanks with the heels not meeting), B. xuefengmontis (vs. just meeting).
Having an unnotched tongue, Boulenophrys dupanglingensis sp. nov. can be distinguished from the following 9 congeners, B. baolongensis, B. binlingensis, B. boettgeri, B. cheni, B. lushuiensis, B. minor, B.mufumontana, B.qianbeiensis and B. sanmingensis (vs. tongue notched posteriorly in these species).
Having a subarticular tubercle present at the base of each finger, differs from the following 7 congeners, B. angka, B. chishuiensis, B. leishanensis, B. lishuiensis, B. tuberogranulatu, B. wuliangshanensis and B. yaoshanensis (vs. just the first and second finger present or not a finger in sight).
Lacking vomerine teeth, vomerine ridge present, Boulenophrys dupanglingensis sp. nov. differs from the following 15 congeners, B. daiyunensis, B. daweimontis, B.elongata, B. fansipanensis, B. frigida, B. hoanglienensis, B. jinggangensis, B. jiulianensis, B. nanlingensis, B. palpebralespinosa, B.pepe, B. rubrimera, B. shimentaina, B. tongboensis, and B. yingdeensis (vs. presence of vomerine teeth in these species).
Lacking lateral fringes on toes, Boulenophrys dupanglingensis sp. nov. differs from the following 9 congeners, B. anlongensis, B. baishanzuensis, B. binchuanensis, B. congjiangensis, B. lini, B. xiangnanensis, B. xianjuensis, and B. yangmingensis (vs. presence of lateral fringes on toes in these species); and from B. wushanensis (vs. presence of wide lateral fringes on toes in males while lacking in females).
In compared to other congeners, Boulenophrys dupanglingensis sp. nov. has a smaller body size with SVL 37.6–40.2 mm (38.9 ±1.3, n=7) in adult males and SVL 41.8–45.9mm (43.7 ±2.1, n=3) in adult females. It is significantly different from the 9 congeners whose SVL ≥ 50 mm in adult males or females, including B. caudoprocta (81.3 mm in males), B.fangjingmontis (58.6–63.8 in males), B. jingdongensis (53.0–56.5 mm in males), B. liboensis (60.5–67.7 mm in males), B. mirabilis (55.8–61.4 mm in males), B. omeimontis (56.0–59.5 mm in males), B. sangzhiensis (53.0–60.8 mm in male), B. shuichengensis(102.0–118.3 mm in males), and B. spinata (47.2–54.4 mm in males).
For the remaining congeners, Boulenophrys dupanglingensis sp. nov. can be further distinguished by having two subarticular tubercle at the base of each finger (vs. three metacarpal tubercles in B.jiangi), and tibio-tarsal articulation reaching forward between anterior margin of tympanum and posterior corner of eye (vs. tibio-tarsal articulation reaching to tip of snout in B. shunhuangensis, or the middle of eye in B. caobangensis)
3.3. Taxonomic Account
Based on the results of molecular phylogenetic and morphological comparisons, the specimens distinct from all other congeners of the Boulenophrys. The description of this new species is provided below.
Boulenophrys dupanglingensis Xi & Mo, sp. nov.
Dupangling Horned Toad (in English)/ (都庞岭角蟾 in Chinese)
Figures 4, Figures 5
Holotype. Adult male, HUNU 22SA01 (Figure 4), collected by Xiaoyang Mo on 20 April 2022, from Dajiangyuan Village (111.36477082º E, 25.45793412º N; 380 m a.s.l.), Dupangling National Nature Reserve, Daoxian County, Yongzhou City, Hunan Province, China.
Paratypes: nine adults: six adult males (HUNU 22SA02–06, CIB2016050802) from the same locality as the holotype on 21 April 2022 and 23 June 2016; one adult female (HUNU 22SA07), which was collected from Daguyuan Village, Jiangyong County (111.337095º E, 25.435091º N; 442 m a.s.l.) on 23 April 2022 (Figure 5); two adult females (HUNU 22SA08–09), which were collected from Daguyuan Village, Jiangyong County (112.245310º E, 25.328928º N; 451 m a.s.l.) on 24 April 2022. The Paratypes collected by Xiaoyang Mo, Bin Wang, Jiayan Xi, Shengchao Shi, Qi Li, Hui Li, Leqiang Zhu, Ayinuer Maimaiti, Yalan Xu, and Fan Zhou.
Etymology: The specific epithet dupanglingensis refers to the type locality of the new species, Dupangling National Nature Reserve, Southern Hunan Province, China.
Diagnosis. (1) small body size, SVL 37.6–40.2 mm (38.9 ± 1.3, n = 7) in adult males and SVL 41.8–45.9 mm (43.6 ± 2.1, n = 3) in adult females; (2) tympanum boundary clear, TD/ED 0.48–0.57 in males, 0.47–0.57 in females; (3) presence of small horn-like tubercle at the edge of upper eyelid; (4) vomerine ridge present, vomerine teeth absent; (5) margin of tongue rounded, not notched posteriorly; (6) dorsal skin rough, a discontinuous “V”-shaped ridge with two discontinuous dorsolateral ridges on two side on the back, dense tubercles on skin of ventral surface of dorsal shank and thigh, spiny tubercles surrounding the cloaca; (7) hindlimbs slender, heels overlapping when the flexed hindlimbs are held at right angles to the body axis, tibio-tarsal articulation reaching forward between anterior margin of tympanum and posterior corner of eye when leg stretched forward; (8) relative finger length IV < II < I < III, a subarticular tubercle present at the base of each finger; (9)supernumerary tubercles below the base of I and II toes distinct; (10) toes without lateral fringes and with only rudimentary webbing, webbing formula: I1 – 1-II1 – 2-III2 – 3IV3- – 2V.
Coloration of holotype in life. Forearms and hindlimbs are marked with dark brown transverse bands; supratympanic ridges are light ivory with dark spot in the posterior edge; a dark vertical band extend below the eye, from the inferior margin of the eye to the upper lip; numerous brown patches scattered across the lateroventral surface of flanks; groin red-orange; ventral surface of throat and chest are light salmon in color with dark brown patches and dark orange blotches; a distinct longitudinal stripe on throat; ventral surface of body light salmon in color with brown patches and white spots; ventral surface of limbs light salmon in color with dark brown spots and blotches; ventral surfaces of hands and feet are brown, with tips of digits pale brown; metacarpal tubercle and metatarsal tubercles are reddish; Pectoral glands and femoral glands are white; Iris yellowish brown.
Coloration of holotype in preservative. Dark brown faded to greyish brown dorsally. Triangular marking is present between the eyes, accompanied by a “V” shaped marking on the mid-dorsum, transverse bands on dorsal sides of the forearms and hind limbs become indistinct. The ventral surface faded to greyish white, with all previously distinct bands and spots becoming less pronounced or indistinguishable.
Variation and sexual dimorphism. Measurement data for the type series are listed in Table 3. Morphological diagnostic characters observed in all paratypes were consistent with those of the holotype. However, there were variations in coloration and stripe patterns between individuals (Figure 4). For example, adult male paratype (HUNU 22SA03) with a smaller body size (SVL 37.8 mm); has light brown back and a black-brown belly; ventral surfaces of limbs primarily black-brown; have a prominent black spot at the right upper eyelids; In contrast, adult female paratype (HUNU 22SA09) with a larger body size (SVL 41.8mm); had a yellowish brown dorsal coloration; chest and anterior abdomen exhibited dark orange blotches on a light brown background; ventral surfaces of limbs were predominantly orange red; fewer markedly enlarged tubercles were present on the flanks.
Sexual Size Dimorphism: Females (SVL 43.6 ± 2.1 mm, n = 3) are distinctly larger than males (SVL 38.8 ± 0.9 mm, n = 6). Adult males possess a single internal subgular vocal sac. Breeding males develop prominent black nuptial spines on dorsal surfaces at the bases of first and second fingers.
Reproductive Characteristics: The eggs are creamy white, with an approximate diameter of 2.6 mm. The observed clutch size consisted of approximately 258 eggs. (Female, HUNU 22SA08, Figure 6B)
Distribution and habitats. Currently, Boulenophrys dupanglingensis sp. nov. is known only from Dupangling National Nature Reserve, southern Hunan, China. Individuals were found in evergreen secondary forest, near mountain streams, and within leaf litter on the forest floor at elevations from 380–450 m (Figure 7). Male calls were recorded during fieldwork conducted from April to June, suggesting that their breeding season corresponds to this period. The collected female specimens contained mature, creamy white oocytes. No tadpoles were observed during the fieldwork.
4. Discussion
In this study, we described a new species of the genus Boulenophrys from Hunan Province, which brings the total number of recognized Boulenophrys species in Hunan Province to 10, including B. caudoprocta, B. dupanglingensis sp. nov., B.hengshanensi, B. mufumontana, B.Sangzhiensis, B. shunhuangensis, B. tuberogranulatus, B. xiangnanensis, B. xuefengmontis, B. yangmingensis [10,33,60].
The Nanling Mountain range spans the provinces of Guangxi, Hunan, Guangdong and Jiangxi, forming a natural barrier between the Yangtze River and Pearl River basins. This region exhibits high species diversity, strong forest dependence, and overlapping distributions of the Boulenophry genus [36]. These patterns highlight the need for further research into speciation processes, niche differentiation, and coexistence mechanisms is warranted [23]. Moreover, Climatic fluctuations, habitat diversity, and the dynamics of montane forests likely drive diversification within Boulenophrys, influencing the evolution of complex phenotypes.
5. Conclusions
Combining the molecular phylogenetic and morphological analysis results, a new Boulenophry species, B. dupanglingensis sp. nov. is described from Yongzhou City, Hunan Province, China. This research brings the total number of recognized Boulenophrys species in Hunan Province to 10. This discovery highlights the biodiversity within the Nanling Mountain range and further emphasizes the importance of the region as a biodiversity hotspot.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Localities, voucher information, and GenBank accession numbers for all samples used in this study.
Author Contributions
Conceptualization, B. X., J.Y. X. and X.Y. M.; methodology, B. X., S.C. S.; software, B. X., J.Y. X. and L.Q. Z.; validation, B.X., S.C. S. and B. W.; investigation, all authors; visualization, L.Q. Z., J.Y. X., S.C. S and B. X.; data curation, all authors; writing—original draft preparation, B. X., J.Y. X. and X.Y. M.; writing—review and editing, B. X., S.C. S., and X.Y. M.; All authors have read and agreed to the published version of the manuscript.
Funding
Biodiversity Survey Project of Dupangling National Nature Reserve.
Institutional Review Board Statement
All animal protocols in this study were reviewed and approved by the Animal Ethical and Welfare Committee of Hunan Normal University (permit number:2024728).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors have declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Acknowledgments
We would like specially thank for the support and investigate assistance by Prof. Xuejian Deng of HUNU. We are grateful for help by Lu Wang, Yinyong Wang, Shuo Qi, Yinmeng Hou for the assistance with specimen examination, and suggestions on manuscript.
References
- AmphibiaWeb: information on amphibian biology and conservation, University of California, Berkeley, California. Available online: http://amphibiaweb.org/ (accessed on 18 Nov 2024).
- Bonaparte, C.L. Conspectus Systematum Herpetologiae et Amphibiologiae; Brill EJ, Leiden, 1850.
- Frost, D.R.; Amphibian Species of the World: an Online Reference. Version 6.2 (Date of access). Available online: doi.org/10.5531/db.vz.0001 (accessed on 18 Nov 2024).
- Chen, J.M. , Zhou, W.W., Poyarkov, N. J., Stuart, B.L., Brown, R.M., Lathrop, A., Wang, Y.Y., Yuan, Z.Y., Jiang, K.; H.M., C., H.M., Suwannapoom, C.,; Nguyen, S.N., Duong, T.V., Papenfuss, T.J., Murphy, R.W., Zhang, Y.P., Che, J. A novel multilocus phylogenetic estimation reveals unrecognized diversity in Asian horned toads, genus Megophrys sensu lato (Anura: Megophryidae). Molecular Phylogenetics and Evolution 2017, 106, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y. , Chen, G.L., Zhu, T.Q., Zeng, Z.C., Lyu, Z.T, Wang, J., Messenger, K., Greenberg, A. J., Guo, Z., Yang, Z.H., Shi, S.H., Wang, Y.Y. Prevalence of cryptic species in morphologically uniform taxa - Fast speciation and evolutionary radiation in Asian frogs. Molecular Phylogenetics and Evolution 2018, 127, 723–731. [Google Scholar] [CrossRef]
- Mahony, S. , Foley, N.M. Biju, S. D. Teeling, E. C. Evolutionary History of the Asian Horned Frogs (Megophryinae): Integrative Approaches to Timetree Dating in the Absence of a Fossil Record. Molecular Biology and Evolution 2017, 34, 744–771. [Google Scholar] [CrossRef] [PubMed]
- Dubois, A. , Ohler, A., Pyron, R.A. New concepts and methods for phylogenetic taxonomy and nomenclature in zoology, exemplified by a new ranked cladonomy of recent amphibians (Lissamphibia). Megataxa 2021, 5, 1–738. [Google Scholar] [CrossRef]
- Frost, D.R. , Grant, T., Faivovich, J., Bain, R. H., Haas, A., Haddad, C. F.B., De Sá, R. O., Channing, A., Wilkinson, M., Donnellan, S. C.,; Raxworthy, C.J., Campbell, J. A., Blotto, B. L., Moler, P., Drewes, R. C., Nussbaum, R. A., Lynch, J. D., Green, D. M., Wheeler, W. C. The amphibian tree of life. Bulletin of the American Museum of Natural History 2006, 8–370. [Google Scholar] [CrossRef]
- Li, S.Z. , Lu, N. N., Liu, J., Wang, B. Description of a new Megophrys Kuhl & Van Hasselt, 1822 (Anura, Megophryidae) from Guizhou Province, China. Zookeys 2020, 986, 101–126. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.T. , Qi, S., Wang, J. Zhang, S. Y., Zhao, J., Zeng, Z. C., Wan, H., Yang, J. H., Mo, Y. M., Wang, Y. Y. Generic classification of Asian horned toads (Anura: Megophryidae: Megophryinae) and monograph of Chinese species. Zool Research 2023, 44, 380–450. [Google Scholar] [CrossRef] [PubMed]
- Beddard, F.E. Contributions to the knowledge of the anatomy of the batrachian family Pelobatid. Proceedings of the Zoological Society of London 1907, 871–911. [Google Scholar] [CrossRef]
- Boulenger, G.A. Descriptions of three new batrachians from Tonkin. Annals and Magazine of Natural History, Series 7 1903, 12, 186–188. [Google Scholar] [CrossRef]
- Fei, L. , Ye, C. Y. Amphibians of China; Chengdu Institute of Biology, Chinese Academy of Sciences. Science Press: Beijing, China, 2016; Volume 1. [Google Scholar]
- Günther, A.C.L.G. The reptiles of British India; Ray Society: London, 1864; pp. 414–415. [Google Scholar]
- Kuhl, H. , Van Hasselt, J. C. Uittreksels uit breieven van de Heeren Kuhl en van Hasselt, aan de Heeren C. J. Temminck, Th. van Swinderen en W. de Haan. Algemeene Konst-en Letter-Bode 1822, 7, 99–104. [Google Scholar]
- Tian, W.S. , Hu, Q. X. Taxonomic study on genus Megophrys, with descriptions of two new genera. Acta Herpetologica Sinica 1983, 2, 41–48. [Google Scholar]
- Li, Y.L. , Jin, M. J., Zhao, J., Liu, Z. Y., Wang, Y. Y., Pang, H. Description of two new species of the genus Megophrys (Amphibia: Anura: Megophryidae) from Heishiding Nature Reserve, Fengkai, Guangdong, China, based on molecular and morphological data. Zootaxa 2014, 3795, 449–471. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H. , Suwannapoom, C., Poyarkov, N. A., Jr., Pawangkhanant, P., Xu, K., Jin, J. Q., Murphy, R. W., Che, J. A new species of the genus Xenophrys Anura Megophryidae from northern Thailand. Zool Res 2019, 40, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. , Zhang, D.D., Lyu, Z.T., Wang, J., Li, Y. L., Liu, Z. Y., Chen, H.H., Rao, D.Q., Jin, Z.F., Zhang, C.Y., Wang, Y.Y. Review of the genus Brachytarsophrys (Anura: Megophryidae), with revalidation of Brachytarsophrys platyparietus and description of a new species from China. Zoological Research 2020, 41, 105–122. [Google Scholar] [CrossRef]
- Ye, C.Y. , Fei, L., Xie, F. A new species of Megophryidae–Megophyrys baolongensis from China (Amphibia, Anura). Acta Herpetologica Sinica 2007, 11, 38–41. [Google Scholar]
- Nguyen, T.Q. , Pham, C. T., Nguyen, T. T., Luong, A. M., Ziegler, T. A new species of Megophrys (Amphibia: Anura: Megophryidae) from Vietnam. Zootaxa 2020, 4722, 401–422. [Google Scholar] [CrossRef]
- Shen, Y.H. , Yang, D. D., Mo, X. Y., Li, H. H., Chen, D. The Fauna of Hunan: Amphibia; Hunan Science and Technology Press: Changsha, 2014. [Google Scholar]
- Luo, T. , Wang, Y., Wang, S., Lu, X., Wang, W., Deng, H., Zhou, J. A species of the genus Panophrys (Anura, Megophryidae) from southeastern Guizhou Province, China. Zookeys 2021, 1047, 27–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y. , Zhao, J., Yang, J.H., Zhou, Z., Chen, G.L., Liu, Y. Morphology, molecular genetics, and bioacoustics support two new sympatric Xenophrys toads (Amphibia: Anura: Megophryidae) in southeast China. PLoS One 2014, 9, e93075. [Google Scholar] [CrossRef]
- Xu, N. , Li, S. Z., Liu, J., Wei, G., Wang, B. A new species of the horned toad Megophrys Kuhl & Van Hasselt, 1822 (Anura, Megophryidae) from southwest China. Zookeys 2020, 943, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.T. , Zeng, Z. C., Wang, J., Liu, Z. Y., Huang, Y. Q., Li, W. Z., Wang, Y. Y. Four new species of Panophrys (Anura, Megophryidae) from eastern China, with discussion on the recognition of Panophrys as a distinct genus. Zootaxa 2021, 4927, 009–040. [Google Scholar] [CrossRef]
- Wang, J. , Lyu, Z. T., Liu, Z. Y., Liao, C. K., Zeng, Z. C., Zhao, J., Li, Y. L., Wang, Y. Y. Description of six new species of the subgenus Panophrys within the genus Megophrys (Anura, Megophryidae) from southeastern China based on molecular and morphological data. Zookeys, 2019; 851, 113–164. [Google Scholar] [CrossRef]
- Zeng, Z.C. , Wang, J., Chen, H.H., Xiao, W.W., Zhan, B.B., Li, Y.H., Lin, S.S. A New Species of the Genus <italic>Boulenophrys</italic> (Anura, Megophryidae) from Eastern Guangdong, China. Asian Herpetological Research 2024, 15, 12–21. [Google Scholar] [CrossRef]
- Zhang, L. , Liang, L.,Ran, H., Shen, Z.X. Megophrys binlingensis fanjingmontis: A New Subspecies of Megophryidae from Guizhou,China. Chinese Journal of Zoology 2012, 47, 135–138. [Google Scholar] [CrossRef]
- Tapley, B. , Cutajar, T., Nguyen, L.T., Nguyen, C.T., Harding, L., Portway, C., Van Luong, H. & Rowley, J.J. A new locality and elevation extension for Megophrys rubrimera (Tapley et al., 2017) in Bat Xat Nature Reserve, Lao Cai Province, northern Vietnam. Herpetology Notes 2018, 11, 865–868. [Google Scholar]
- Wang, J. , Zeng, Z. C., Lyu, Z. T., Qi, S., Liu, Z. Y., Chen, H. H., Lu, Y. H.,; Xiao, H.W., Lin, C. R., Chen, K., Wang, Y. Y. Description of three new Boulenophrys species from eastern Guangdong, China, emphasizing the urgency of ecological conservation in this region (Anura, Megophryidae). Zootaxa 2022, 5099, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Tapley, B. , Cutajar, T., Nguyen, L. T., Portway, C.,Mahony, S., Nguyen, C.T.,Harding, L.,Luong, H.V., Rowley, J. L. A new potentially Endangered species of Megophrys (Amphibia: Megophryidae) from Mount Ky Quan San, north-west Vietnam. Journal of Natural History 2020, 54, 2543–2575. [Google Scholar] [CrossRef]
- Qian, T.Y. , Hu, K., Mo, X.Y., Gao, Z.W., Zhang, N., Yang, D.D. A new species of Boulenophrys from central Hunan Province, China (Anura: Megophryidae). Vertebrate Zoology 2023, 73, 915–930. [Google Scholar] [CrossRef]
- Tapley, B. , Cutajar, T., Mahony, S., Nguyen, C.T., Dau, V.Q., Luong, A.M., Le, D.T., Nguyen, T.T., Nguyen, T.Q., Portway, C., Luong, H. V., Rowley, J. L. Two new and potentially highly threatened Megophrys Horned frogs (Amphibia: Megophryidae) from Indochina's highest mountains. Zootaxa 2018, 4508, 301–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Z.Y.; Lyu, Z.T.; Zeng, Z.C.; Wang, Y.Y. A new species of the genus Xenophrys (Amphibia: Anura: Megophryidae) from an offshore island in Guangdong Province, southeastern China. Zootaxa 2017, 4324, 541–556. [Google Scholar] [CrossRef]
- Liu, J. , Li, S. Z., Wei, G., Xu, N., Cheng, Y. L., Wang, B., Wu, J. A New Species of the Asian Toad Genus Megophrys sensu lato (Anura: Megophryidae) from Guizhou Province, China. Asian Herpetological Research 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Wang, Y.Y., Zhang, T. D., Zhao, J., Sung, Y. H., Yang, J. H., Pang, H., ; Zhang, Z. Description of a new species of the genus Xenophrys Gunther, 1864 (Amphibia: Anura: Megophryidae) from Mount Jinggang, China, based on molecular and morphological data. Zootaxa, 2012; 53–67.
- Li, S.Z.; Xu, N.; Liu, J.; Jiang, J.P.; Wei, G.; Wang, B. A New Species of the Asian Toad Genus Megophrys sensu lato (Amphibia: Anura: Megophryidae) from Guizhou Province, China. Asian Herpetological Research 2018, 9, 224–239. [Google Scholar] [CrossRef]
- Lin, S.S. , Chen, H. H., Li, Y. H., Peng, Z. N., Zeng, Z. C., Wang, J. A new Boulenophrys species (Anura, Megophryidae) from the coastal hills of eastern Fujian Province, China. Zookeys 2024, 1216, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.C. , Li, D.H., Zhu, W.B., Wang, J., Jiang, J.P., Ye, C.Y., Fei, L., Wang, B. Description of a new toad of Megophrys Kuhl & Van Hasselt, 1822 (Amphibia: Anura: Megophryidae) from western Yunnan Province, China. Zootaxa 2021, 4942, 351–381. [Google Scholar] [CrossRef]
- Zhang, Y.N. , Li, G., Xiao, N., Li, J.Q., Pan, T., Wang, H., Zhang, B.W., Zhou, J. A New Species of the Genus Xenophrys (Amphibia: Anura: Megophryidae) from Libo County, Guizhou, China. Asian Herpetological Research 2017, 8, 75. [Google Scholar] [CrossRef]
- Wang, L. , Deng, X.J., Liu, Y., Wu, Q.Q., Liu, Z. A new species of the genus Megophrys (Amphibia: Anura: Megophryidae) from Hunan, China. Zootaxa 2019, 4695, 301–330. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.T. , Li, Y.Q., Zeng, Z.C., Zhao, J., Liu, Z.Y., Guo, G.X., Wang, Y.Y. Four new species of Asian horned toads (Anura, Megophryidae, Megophrys) from southern China. ZooKeys 2020, 942, 105–140. [Google Scholar] [CrossRef] [PubMed]
- Messenger, K.R. , Dahn, H.A., Liang, Y., Xie, P., Wang, Y., Lu, C. A new species of the genus Megophrys Gunther, 1864 (Amphibia: Anura: Megophryidae) from Mount Wuyi, China. Zootaxa 2019, 4554, 561–583. [Google Scholar] [CrossRef]
- Wang, J. , Lin, S.S., Gan, J.S., Chen, H.H., Yu, L.M., Pan, Z., Xiao, J.J., Zeng, Z.C. A new species of the genus Boulenophrys from South China (Anura, Megophryidae). Zootaxa 2024, 5514, 451–468. [Google Scholar] [CrossRef] [PubMed]
- Su, H.J. , Shi, S.C., Wu, Y.Q., Li, G.R., Yao, X.G., Wang, B., Li, S.Z. Description of a new horned toad of Megophrys Kuhl & Van Hasselt, 1822 (Anura, Megophryidae) from southwest China. Zookeys, 2020; 131–159. [Google Scholar] [CrossRef]
- Tapley, B. , Cutajar, T., Mahony, S., Nguyen, C. T., Dau, V. Q., Nguyen, T. T., Luong, H. V., Rowley, J. J. L. The Vietnamese population of Megophrys kuatunensis (Amphibia: Megophryidae) represents a new species of Asian horned frog from Vietnam and southern China. Zootaxa 2017, 4344, 465–492. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.P. , Ye, C.Y., Fei, L. A New Horn Toad Megophrys sangzhiensis from Hunan, China (Amphibia,Anura). Zoological Research 2008, 29, 219–222. [Google Scholar] [CrossRef]
- Tian, Y.Z. , Gu, X. M., Sun, A. Q. A new species of Xenophrys in China (Amphibia: Pelobatidae). Acta Zootaxonomica Sinica 2000, 25, 462–466. [Google Scholar]
- Mo, X.Y. , Shen, Y.H., Li, H.H., Wu, X.S. A new species of Megophrys (Amphibia: Anura: Megophryidae) from the northwestern Hunan Province, China. Current Zoology 2010, 56, 432–436. [Google Scholar] [CrossRef]
- Fei, L. Atlas of Amphibians in China. Field Edition; Henan Science and Technology Press: Zhengzhou, p. 432.
- Wang, B. , Wu, Y. Q., Peng, J. W., Shi, S. C., Lu, N. N., Wu, J. A new Megophrys Kuhl & Van Hasselt (Amphibia, Megophryidae) from southeastern China. Zookeys 2020, 904, 35–62. [Google Scholar] [CrossRef]
- Qi, S. , Lyu, Z. T., Wang, J. Mo, Y. M., Zeng, Z. C., Zeng, Y. J., Dai, K. Y., Li, Y. Q., Grismer, L. L., Wang, Y. Y. Three new species of the genus Boulenophrys (Anura, Megophryidae) from southern China. Zootaxa 2021, 5072, 401–438. [Google Scholar] [CrossRef]
- Thompson, J.D. , Gibson, T. J., Plewniak, F., Jeanmougin, F., Higgins, D. G. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic acids research 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Tamura, K. , Stecher, G., Kumar, S. MEGA11 Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Lanfear, R. , Calcott, B., Ho, S.Y. W., Guindon, S. PartitionFinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Molecular Biology and Evolution 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Ronquist, F. , Teslenko, M., van der Mark, P., Ayres, D.L., Darling, A., Hohna, S., Larget, B., Liu, L., Suchard, M.A., Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Systematic Biology 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Guindon, S. , Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W., Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Systematic Biology 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Savage, J.M. , Heyer, W.R. Variation and distribution in the tree-frog genus Phyllomedusa. Beitra¨ge zur Neotropischen Fauna 1967, 5, 111–131. [Google Scholar] [CrossRef]
- Gao, Z.W. , Qian, T. Y., Jiang, J. P., Hou, D. J., Deng, X. J., Yang, D. D. Species diversity and distribution of amphibians and reptiles in Hunan Province, China. Biodiversity Science 2022, 30, 21290. [Google Scholar] [CrossRef]
Figure 1.
Distribution map showing the type localities of Boulenophrys species phylogenetically close to B. dupanglingensis sp. nov.
Figure 1.
Distribution map showing the type localities of Boulenophrys species phylogenetically close to B. dupanglingensis sp. nov.
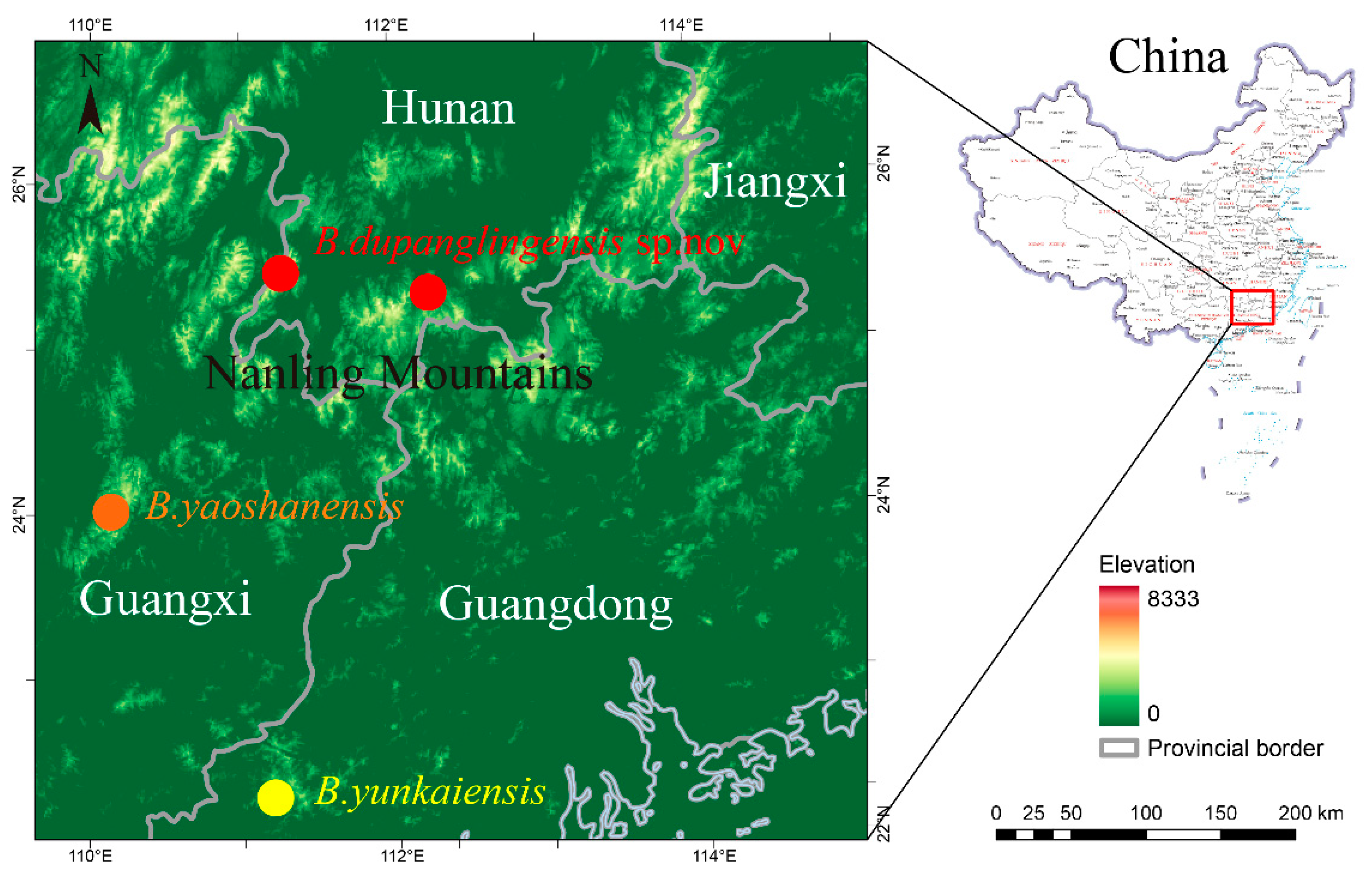
Figure 2.
Bayesian topology based on the partial DNA sequences of the mitochondrial 16S rRNA and COI genes. with Bootstrap Support (BS) / Bayesian Posterior Probabilities (BPP) indicated at nodes. A dash ‘-’ denotes BS < 70 or BPP < 90.
Figure 2.
Bayesian topology based on the partial DNA sequences of the mitochondrial 16S rRNA and COI genes. with Bootstrap Support (BS) / Bayesian Posterior Probabilities (BPP) indicated at nodes. A dash ‘-’ denotes BS < 70 or BPP < 90.
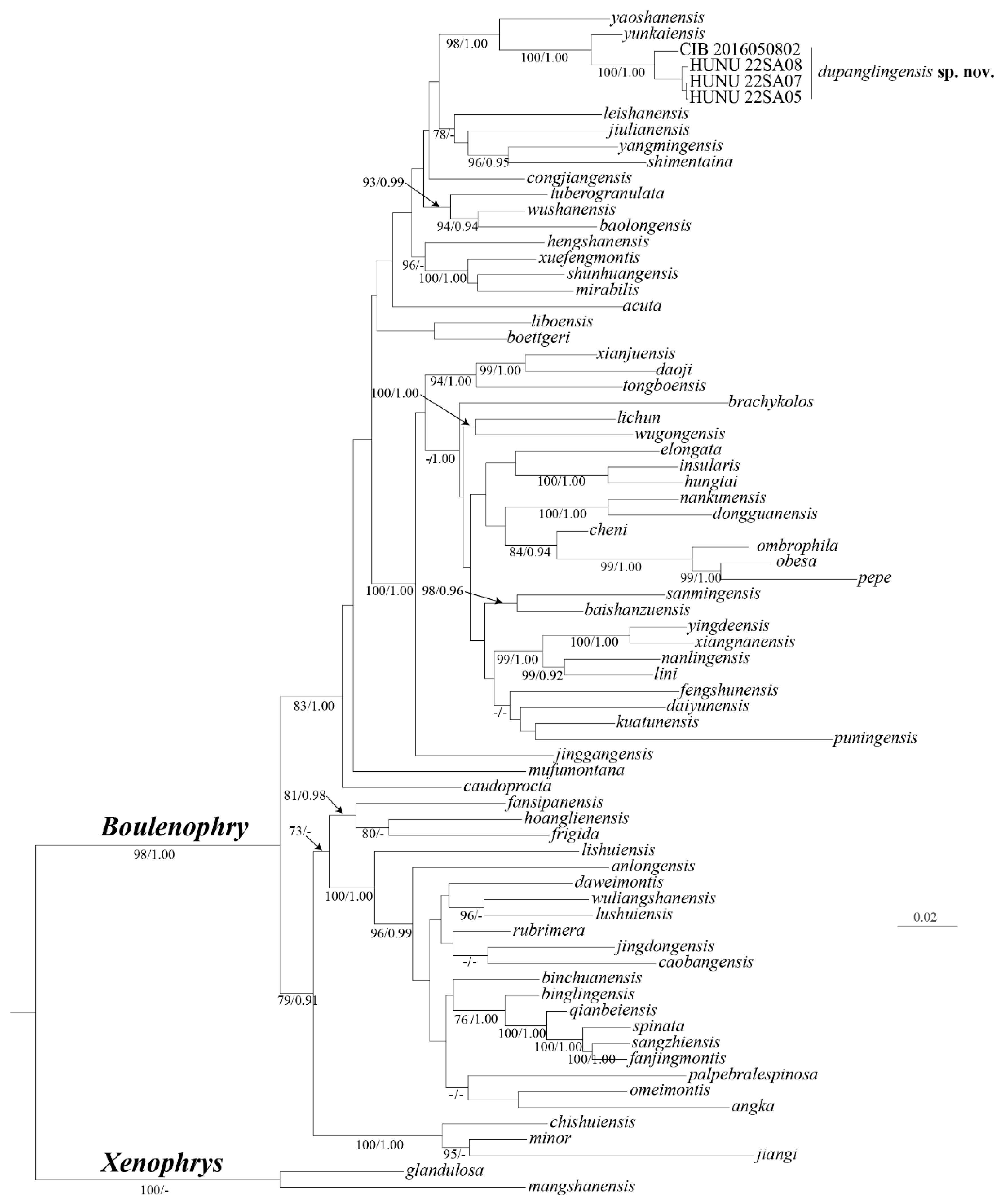
Figure 3.
Comparisons of Boulenophrys dupanglingensis sp. nov. (CIB2016050802) and B. yunkaiensis (SYS a004986) in preservative. White arrows indicate differences between closely related congeners. Boulenophrys dupanglingensis sp. nov.: A: dorsolateral view B: plantar view of foot; B. yunkaiensis: C: dorsolateral view D: plantar view of foot. photo by Le-Qiang Zhu.
Figure 3.
Comparisons of Boulenophrys dupanglingensis sp. nov. (CIB2016050802) and B. yunkaiensis (SYS a004986) in preservative. White arrows indicate differences between closely related congeners. Boulenophrys dupanglingensis sp. nov.: A: dorsolateral view B: plantar view of foot; B. yunkaiensis: C: dorsolateral view D: plantar view of foot. photo by Le-Qiang Zhu.
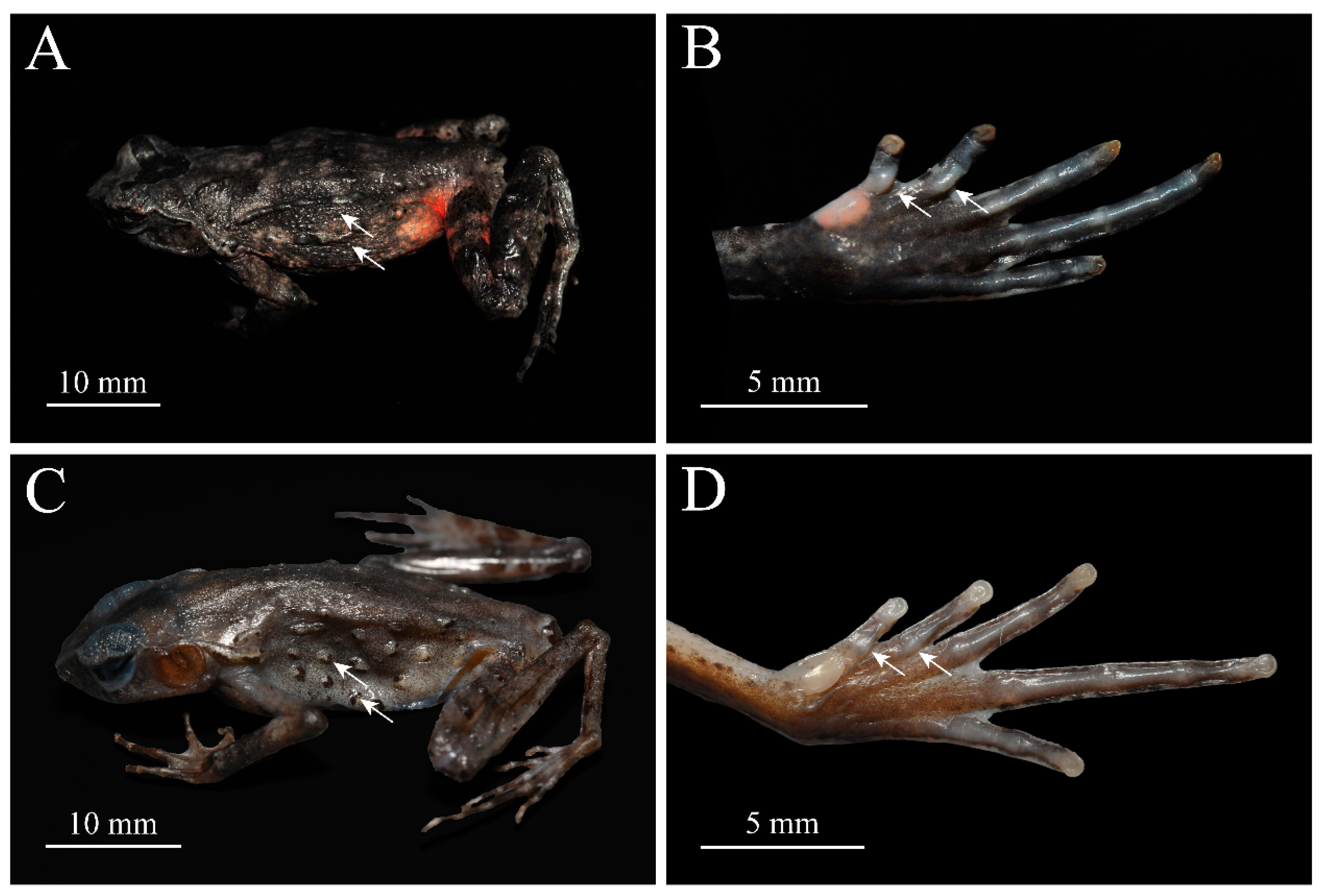
Figure 4.
Holotype of Boulenophrys dupanglingensis sp. nov. (HUNU 22SA01) in life: A: lateral view; B: ventral view; C: dorsal view; D: volar view of left hand; E: plantar view of left foot. photo by Jia-Yan Xi.
Figure 4.
Holotype of Boulenophrys dupanglingensis sp. nov. (HUNU 22SA01) in life: A: lateral view; B: ventral view; C: dorsal view; D: volar view of left hand; E: plantar view of left foot. photo by Jia-Yan Xi.

Figure 5.
Paratypes of Boulenophrys dupanglingensis sp. nov. in life. Male paratype (HUNU 22SA003): A, C, E; Female paratype (HUNU 22SA009): B, D, F. photo by Jia-Yan Xi.
Figure 5.
Paratypes of Boulenophrys dupanglingensis sp. nov. in life. Male paratype (HUNU 22SA003): A, C, E; Female paratype (HUNU 22SA009): B, D, F. photo by Jia-Yan Xi.

Figure 6.
(A) edge of the tongue; (B) eggs of living paratype. photo by Jia-Yan Xi.
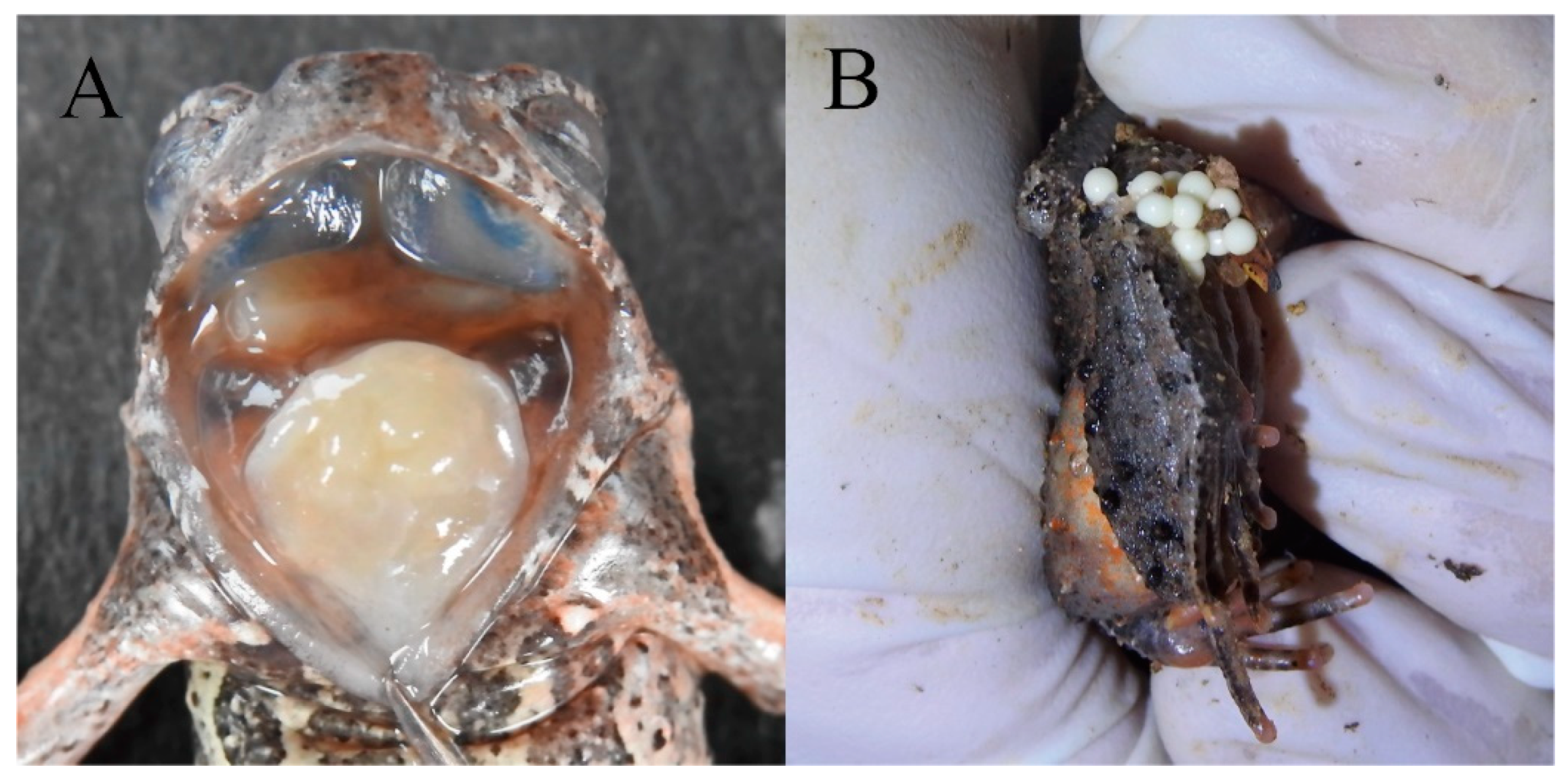
Figure 7.
Habitat of Boulenophrys dupanglingensis sp. nov. A: landscape of montane forests in the type locality B: a mountain stream where the new species collected. photo by Sheng-Chao Shi.
Figure 7.
Habitat of Boulenophrys dupanglingensis sp. nov. A: landscape of montane forests in the type locality B: a mountain stream where the new species collected. photo by Sheng-Chao Shi.


Table 1.
Literature for morphological characters of 69 recognized species of Boulenophrys.
| ID | Boulenophrys species | References |
|---|---|---|
| 1 | B. acuta (Wang, Li & Jin, 2014) | Li et al. 2014[17] |
| 2 | B. angka (Wu, Suwannapoom, Poyarkov, Pawangkhanant, Xu, Jin, Murphy & Che, 2019) | Wu et al. 2019[18] |
| 3 | B. anlongensis (Li, Lu, Liu & Wang, 2020) | Li et al. 2020a[19] |
| 4 | B. baishanzuensis (Wu, Li, Liu, Wang & Wu, 2020) | Wu et al. 2020[18] |
| 5 | B. baolongensis (Ye, Fei & Xie, 2007) | Ye et al. 2007[20] |
| 6 | B. binchuanensis (Ye & Fei, 1995) | Fei and Ye 2016[13] |
| 7 | B. binlingensis (Jiang, Fei & Ye, 2009) | Fei and Ye 2016[13] |
| 8 | B. boettgeri (Boulenger, 1899) | Fei and Ye 2016[13] |
| 9 | B. brachykolos (Inger & Romer, 1961) | Fei and Ye 2016[13] |
| 10 | B. caobangensis (Nguyen, Pham, Nguyen, Luong & Ziegler, 2020) | Nguyen et al. 2020[21] |
| 11 | B. caudoprocta (Shen, 1994) | Fei and Ye 2016[22] |
| 12 | B. congjiangensis (Luo, Wang, Wang, Lu, Wang, Deng & Zhou, 2021) | Luo et al. 2021[23] |
| 13 | B. cheni (Wang & Liu, 2014) | Wang et al. 2014[24] |
| 14 | B. chishuiensis (Xu, Li, Liu, Wei & Wang, 2020) | Xu et al. 2020[25] |
| 15 | B. daiyunensis (Lyu, Wang & Wang, 2021) | Lyu et al. 2021[26] |
| 16 | B. daoji (Lyu, Zeng, Wang & Wang, 2021) | Lyu et al. 2023[10] |
| 17 | B. daweimontis (Rao & Yang, 1997) | Fei & Ye 2016[13] |
| 18 | B. dongguanensis (Wang & Wang, 2019) | Wang et al. 2019a[27] |
| 19 | B. elongata (Zeng, Wang, Chen, Xiao, Zhan, Li & Lin 2024) | Zeng et al. 2024[28] |
| 20 | B. fanjingmontis (Zhang, Liang, Ran & Shen 2012) | Zhang et al.2012[29] |
| 21 | B. fansipanensis (Tapley, Cutajar, Mahony, Nguyen, Dau, Luong, Le, Nguyen, Nguyen, Portway, Luong & Rowley, 2018) | Tapley et al. 2018[30] |
| 22 | B. fengshunensis (Wang, Zeng, Lyu, & Wang, 2022) | Wang et al. 2022[31] |
| 23 | B. frigida (Tapley, Cutajar, Nguyen, Portway, Mahony, Nguyen, Harding, Luong & Rowley, 2021) | Tapley et al. 2020[32] |
| 24 | B. hengshanensis (Qian, Hu, Mo, Gao, Zhang, & Yang, 2023) | Qian et al. 2023[33] |
| 25 | B. hoanglienensis (Tapley, Cutajar, Mahony, Nguyen, Dau, Luong, Le, Nguyen, Nguyen, Portway, Luong & Rowley, 2018) | Tapley et al. 2018[34] |
| 26 | B. hungtai (Wang, Zeng, Lyu, Xiao, & Wang, 2022) | Wang et al. 2022[31] |
| 27 | B. insularis (Wang, Liu, Lyu, Zeng & Wang, 2017) | Wang et al. 2017a[35] |
| 28 | B. jiangi (Liu, Li, Wei, Xu, Cheng, Wang & Wu, 2020) | Liu et al. 2020[36] |
| 29 | B. jingdongensis (Fei & Ye, 1983) | Fei & Ye 2016[13] |
| 30 | B. jinggangensis (Wang, 2012) | Wang et al. 2012[37] |
| 31 | B. jiulianensis (Wang, Zeng, Lyu & Wang, 2019) | Lyu et al. 2023[10] |
| 32 | B. kuatunensis (Pope, 1929) | Fei & Ye 2016; [13] |
| 33 | B. leishanensis (Li, Xu, Liu, Jiang, Wei & Wang, 2018) | Li et al. 2018[38] |
| 34 | B.lichun (Lin, Chen, Li, Peng, Zeng &Wang, 2024) | Lin et al. 2024[39] |
| 35 | B. lushuiensis (Shi, Li, Zhu, Jiang, Jiang & Wang, 2021) | Shi et al. 2021[40] |
| 36 | B. liboensis (Zhang, Li, Xiao, Li, Pan, Wang, Zhang & Zhou, 2017) | Zhang et al. 2017[41] |
| 37 | B. lini (Wang & Yang, 2014) | Wang et al. 2014[24] |
| 38 | B. lishuiensis (Wang, Liu & Jiang, 2017) | Wang et al. 2017b[42] |
| 39 | B. minor (Stejneger, 1926) | Fei & Ye 2016[13] |
| 40 | B. mirabilis (Lyu, Wang & Zhao, 2020) | Lyu et al. 2020[43] |
| 41 | B. mufumontana (Wang, Lyu & Wang, 2019) | Wang et al. 2019a[27] |
| 42 | B. nankunensis (Wang, Zeng & Wang, 2019) | Wang et al. 2019a[27] |
| 43 | B. nanlingensis (Lyu, Wang, Liu & Wang, 2019) | Wang et al. 2019a[27] |
| 44 | B. obesa (Wang, Li & Zhao, 2014) | Li et al. 2014[17] |
| 45 | B. ombrophila (Messenger & Dahn, 2019) | Messenger et al. 2019[44] |
| 46 | B. omeimontis (Liu, 1950) | Fei & Ye 2016[13] |
| 47 | B. palpebralespinosa (Bourret, 1937) | Fei & Ye 2016[13] |
| 48 | B. pepe (Wang & Zeng, 2024) | Wang et al. 2024[45] |
| 49 | B. puningensis(Wang, Zeng, Lyu, Xiao, & Wang, 2022) | Wang et al. 2022[31] |
| 50 | B. qianbeinsis (Su, Shi, Wu, Li, Yao, Wang & Li, 2020) | Su et al. 2020[46] |
| 51 | B. rubrimera (Tapley, Cutajar, Mahony, Chung, Dau, Nguyen, Luong & Rowley, 2017) | Tapley et al. 2017[47] |
| 52 | B. sangzhiensis (Jiang, Ye & Fei, 2008) | Jiang et al. 2008[48] |
| 53 | B. sanmingensis (Lyu & Wang, 2021) | Lyu et al. 2023[26] |
| 54 | B. shimentaina (Lyu, Liu & Wang, 2020) | Lyu et al. 2023[10] |
| 55 | B. shuichengensis (Tian and Sun, 1995) | Tian et al. 2000[49] |
| 56 | B. shunhuangensis (Wang, Deng, Liu, Wu & Liu, 2019) | Wang et al. 2019b[42] |
| 57 | B. spinata (Liu & Hu, 1973) | Fei & Ye 2016[13] |
| 58 | B. tongboensis (Wang & Lyu, 2021) | Lyu et al. 2023[10] |
| 59 | B. tuberogranulatus (Shen, Mo & Li, 2010) | Mo et al. 2010[50] |
| 60 | B. wugongensis (Wang, Lyu & Wang, 2019) | Wang et al. 2019a[27] |
| 61 | B. wuliangshanensis (Ye & Fei, 1995) | Fei and Ye 2020[51] |
| 62 | B. wushanensis (Ye & Fei, 1995) | Fei and Ye 2020[51] |
| 63 | B. xiangnanensis (Lyu, Zeng & Wang, 2020) | Lyu et al. 2023[10] |
| 64 | B. xianjuensis (Wang, Wu, Peng, Shi, Lu & Wu, 2020) | Wang et al. 2020[52] |
| 65 | B. xuefengmontis (Lyu & Wang, 2023) | Lyu et al. 2023[10] |
| 66 | B. yangmingensis (Lyu, Zeng & Wang, 2020) | Lyu et al. 2023[10] |
| 67 | B. yaoshanensis (Qi, Mo, Lyu, Wang & Wang, 2021) | Qi et al. 2021[53] |
| 68 | B. yingdeensis (Qi, Lyu, Wang & Wang, 2021) | Qi et al. 2021[53] |
| 69 | B. yunkaiensis (Qi, Wang, Lyu & Wang, 2021) | Qi et al. 2021[53] |
Table 2.
Diagnostic characters separating all 69 species of Boulenophrys. “/” indicates that data are not available.
Table 2.
Diagnostic characters separating all 69 species of Boulenophrys. “/” indicates that data are not available.
| Species | SVL in males (in mm) | SVL in females (in mm) | horn-like tubercle at upper eyelid: slightly large (2), small (1) |
Vomerine teeth: present (1), or absent (0) |
Tongue: notched (1), or not notched (0) |
Heels overlapping (2) just meeting (1), or not meeting (0) |
lateral fringes on toes: wide (2), narrow (1), or absent (0) | Webs on toes: more than one-fourth (2), rudimentary (1), or lacking (0) | |
| 1 | B. dupanglingensissp. nov. | 37.8-40.2 | 41.8-45.9 | 1 | 0 | 0 | 2 | 0 | 1 |
| 2 | B. acuta | 27.1–33.0 | 28.1–33.6 | 2 | 0 | 0 | 0 | 1 | 1 |
| 3 | B. angka | 31.2–32.1 | 37.5–39.2 | 1 | 0 | 0 | 1 or 2 | 0 | 1 |
| 4 | B. anlongensis | 40.0–45.5 | 48.9–51.2 | 1 | 0 | 0 | 2 | 1 | 1 |
| 5 | B. baishanzuensis | 28.4–32.4 | / | 1 | 0 | 0 | 2 | 1 | 0 |
| 6 | B. baolongensis | 42.0–45.0 | / | 1 | 0 | 1 | 1 | 0 | 0 |
| 7 | B. binchuanensis | 32.0–36.0 | 40.2–42.5 | 1 | 0 | 0 or 1 | 1 | 2 | 1 |
| 8 | B. binlingensis | 45.1–51.0 | / | 1 | 0 | 1 | 2 | / | 1 |
| 9 | B. boettgeri | 34.5–37.8 | 39.7–46.8 | 1 | 0 | 1 | 1 | 2 | 1 |
| 10 | B. brachykolos | 33.7–39.3 | 33.9–45.9 | 1 | 0 | 0 | 0 | 0 | 1 |
| 11 | B. caobangensis | 34.9–38.9 | / | 1 | 0 | 0 | / | 0 | 1 |
| 12 | B. caudoprocta | 81.3 | / | 2 | 1 | 0 | 1 | / | 1 |
| 13 | B. congjiangensis | 28.6–33.4 | 38.4–40.2 | 1 | 0 | 0 | 2 | 1 | 1 |
| 14 | B. cheni | 26.2–29.5 | 31.8–34.1 | 1 | 0 | 1 | 2 | 2 | 1 |
| 15 | B. chishuiensis | 43.4–44.1 | 44.8–49.8 | 1 | 0 | 0 | 2 | 0 | 1 |
| 16 | B. daiyunensis | 27.6–28.7 | 33.7–35.6 | 1 | 1 | 0 | 1 or 2 | 1 | 1 |
| 17 | B. daoji | 32.6–33.6 | 37.5–41.4 | 1 | 0 | 0 | 0 | 1 | 1 |
| 18 | B. daweimontis | 34.0–37.0 | 40.0–46.0 | 1 | 1 | / | / | 0 | 0 |
| 19 | B. dongguanensis | 30.2–39.3 | / | 1 | 1 | 0 | 0 | 0 | 1 |
| 20 | B. elongata | 28.2–28.5 | 35.1–37.6 | 1 | 1 | 0 | 1 | 1 | 0 |
| 21 | B. fanjingmontis | 58.2–63.6 | 62.8–72.2 | 1 | 1 | 1 | 1 | 2 | 1 |
| 22 | B. fansipanensis | 30.9–44.3 | 41.7–42.5 | 1 | 1 | 0 or 1 | / | 0 | 0 |
| 23 | B. fengshunensis | 34.3–39.4 | 42.5–44.9 | 1 | 1 | 0 | 0 | 0 | 1 |
| 24 | B. frigida | 30.3–31.8 | / | 1 | 1 | 0 or 1 | / | 0 | 0 |
| 25 | B.hengshanensis | 35.7–41.2 | 37.5–50.2 | 1 | 0 | 0 | 0 | 0 | 0 |
| 26 | B. hoanglienensis | 37.4–47.6 | 59.6 | 1 | 1 | 1 | / | 0 | 1 or 0 |
| 27 | B. hungtai. | 25.8–33.3 | / | 1 | 0 | 0 | 0 | 0 | 0 |
| 28 | B. insularis | 36.8–41.2 | 47.1 | 1 | 1 | 1 | 0 | 0 | 1 |
| 29 | B. jiangi | 34.4–39.2 | 39.5–40.4 | 1 | 0 | 0 | 2 | 0 | 1 |
| 30 | B. jingdongensis | 53.0–56.5 | 63.5 | 1 | 1 | 1 | 2 | 2 | 2 |
| 31 | B. jinggangensis | 35.1–36.7 | 38.4–41.6 | 2 | 1 | 0 | 2 | 1 | 1 |
| 32 | B. jiulianensis | 30.4–33.9 | 34.1–37.5 | 1 | 1 | 1 | 2 | 0 | 1 |
| 33 | B. kuatunensis | 26.2–31.4 | 26.6–37.3 | 1 | 0 | 1 | 0 or 2 | 0 or 1 | 0 |
| 34 | B. leishanensis | 30.4–38.7 | 42.3 | 1 | 0 | 0 | 2 | 0 | 1 |
| 35 | B. lichun | 33.5–37.0 | 47.1 | 1 | 1 | 0 | 0 | 0 | 0 |
| 36 | B. liboensis | 60.5–67.7 | 60.8–70.6 | 2 | 1 | 1 | 2 | 2 | 1 |
| 37 | B. lini | 34.1–39.7 | 37.0–39.9 | 1 | 0 | 0 | 2 | 2 | 1 |
| 38 | B. lishuiensis | 30.7–34.7 | 36.9–40.4 | 1 | 0 | 0 | / | 0 | 0 |
| 39 | B. lushuiensis | 31.0–34.8 | / | 1 | 0 | 1 | / | 1 | 1 |
| 40 | B. minor | 34.5–41.2 | / | 1 | 0 | 1 | 1 or 2 | 0 | 1 |
| 41 | B. mirabilis | 55.8–61.4 | 68.5–74.8 | 2 | 0 | 0 | 2 | 1 | 1 |
| 42 | B. mufumontana | 30.1–30.8 | 36.3 | 1 | 0 | 0 | 2 | 1 | 1 |
| 43 | B. nankunensis | 29.9–34.9 | 39.4–41.9 | 1 | 1 | 0 | 0 | 0 | 1 |
| 44 | B. nanlingensis | 30.5–37.3 | / | 1 | 1 | 1 | 2 | 1 | 1 |
| 45 | B. obesa | 35.6 | 37.5–41.2 | 1 | 0 | 0 | 0 | 0 | 1 |
| 46 | B. ombrophila | 27.4–34.5 | 32.8–35.0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 47 | B. omeimontis | 56.0–59.5 | 68.0–72.5 | 1 | 1 | 1 | 2 | 1 | 1 |
| 48 | B. palpebralespinosa | 36.2–38.0 | / | 2 | 1 | 0 | 2 | 2 | 2 |
| 49 | B. pepe | 35.3–36.4 | / | 1 | 0 | 1 | 0 | 0 | 0 |
| 50 | B. puningensis | 31.7–34.6 | 37.8–38.3 | 1 | 1 | 0 | 0 | 0 | 1 |
| 51 | B. qianbeinsis | 49.3–58.2 | / | 1 | 1 | 1 | 2 | 2 | 2 |
| 52 | B. rubrimera | 26.6–30.8 | / | 1 | 1 | 0 or 1 | / | 1 | 0 |
| 53 | B. sangzhiensis | 54.7 | / | 1 | 1 | 1 | 2 | 1 | 1 |
| 54 | B. sanmingensis | 27.0–29.5 | 29.5 | 1 | 0 | 1 | 2 | 2 | 1 |
| 55 | B. shimentaina | 28.0–30.6 | / | 1 | 1 | 0 | 2 | 1 | 1 |
| 56 | B. shuichengensis | 102.0–118.3 | 99.8–115.6 | 2 | 0 | 1 | / | 2 | 2 |
| 57 | B. shunhuangensis | 30.3–33.7 | 37.6 | 1 | 0 | 0 | 2 | 0 | 1 |
| 58 | B. spinata | 47.2–54.4 | 54.0–55.0 | 1 | 0 | 1 | 2 | 2 | 2 |
| 59 | B. tongboensis | 26.5–31.5 | / | 1 | 1 | 1 | 2 | 0 | 0 |
| 60 | B. tuberogranulatus | 33.2–39.0 | 50.5 | 1 | 0 | 0 | / | 0 | 1 |
| 61 | B. wugongensis | 31.0–34.1 | 38.5–42.8 | 1 | 0 | 0 | 0 | 0 | 1 |
| 62 | B. wuliangshanensis | 27.3–31.6 | 41.3 | 1 | 0 | 0 or 1 | 2 | 0 | 0 |
| 63 | B. wushanensis | 30.4–35.5 | 38.4 | 1 | 0 | 0 | 1 | 0(in female), 2(in male) |
1 |
| 64 | B. xiangnanensis | 38.6–42.0 | 44.4 | 1 | 0 | 0 | 1 | 2 | 1 |
| 65 | B. xianjuensis | 31.0–36.3 | 41.6 | 1 | 0 | 0 | 2 | 1 | 1 |
| 66 | B. xuefengmontis | 37.0–38.3 | 45.3–48.9 | 1 | 0 | 0 | 1 or 2 | 0 | 0 |
| 67 | B. yaoshanensi | 32.5–42.6 | 46.6–47.4 | 1 | 0 | 0 | 1 or 2 | 0 | 1 |
| 68 | B. yangmingensis | 33.2–37.1 | 45.2 | 1 | 0 | 0 | 2 | 1 | 1 |
| 69 | B. yingdeensis | 33.2–35.3 | 36.3–45.8 | 1 | 1 | 0 | 1 or 2 | 0 | 1 |
| 70 | B. yunkaiensis | 35.3–40.0 | 45.3–46.1 | 1 | 0 | 0 | 1 or 2 | 0 | 1 |
Table 3.
Measurements (in mm) and ratios of the type series of Boulenophrys dupanglingensis sp. nov. * indicates the holotype.
Table 3.
Measurements (in mm) and ratios of the type series of Boulenophrys dupanglingensis sp. nov. * indicates the holotype.
| HUNU 22SA01* | HUNU 22SA02 | HUNU 22SA03 | HUNU 22SA04 | HUNU 22SA05 | HUNU 22SA06 | CIB2016050802 | HUNU 22SA07 | HUNU 22SA08 | HUNU 22SA09 | |
| Sex | Male | Male | Male | Male | Male | Male | Male | Female | Female | Female |
| SVL | 39.6 | 38.1 | 37.8 | 40.2 | 38.8 | 38.5 | 37.6 | 43.0 | 45.9 | 41.8 |
| HDL | 13.3 | 12.8 | 13.2 | 12.1 | 12.1 | 12.1 | 11.3 | 13.3 | 13.1 | 13.2 |
| HDW | 13.7 | 13.2 | 13.4 | 12.7 | 13.4 | 13.2 | 12.6 | 14.2 | 14.9 | 14.0 |
| SNT | 3.3 | 3.3 | 3.2 | 3.4 | 3.2 | 3.1 | 4.4 | 3.2 | 3.4 | 3.1 |
| IND | 4.4 | 4.1 | 3.7 | 4.2 | 4.2 | 3.9 | 4.2 | 4.4 | 4.7 | 4.4 |
| IOD | 3.1 | 3.3 | 3.3 | 3.7 | 3.2 | 3.3 | 3.8 | 3.6 | 3.8 | 3.7 |
| ED | 6.3 | 5.6 | 5.2 | 6.0 | 5.6 | 5.8 | 4.2 | 5.5 | 6.0 | 5.3 |
| TD | 3.3 | 3.2 | 2.9 | 2.9 | 3.1 | 3.1 | 2.3 | 2.6 | 3.1 | 3.0 |
| TED | 1.8 | 1.8 | 1.8 | 1.8 | 1.4 | 1.6 | 2.4 | 2.0 | 2.5 | 2.1 |
| HND | 9.7 | 9.2 | 9.7 | 9.5 | 9.2 | 9.2 | 8.4 | 10.3 | 11.8 | 9.8 |
| RAD | 8.0 | 7.4 | 7.8 | 7.7 | 7.6 | 7.2 | 8.9 | 8.0 | 8.3 | 7.1 |
| TIB | 15.1 | 14.8 | 14.9 | 15.5 | 14.8 | 14.4 | 14.6 | 15.7 | 18.6 | 15.8 |
| FTL | 23.8 | 23.1 | 23.2 | 23.6 | 23.2 | 22.8 | / | 25.4 | 27.3 | 26.7 |
| TD/ED | 0.52 | 0.57 | 0.48 | 0.48 | 0.55 | 0.53 | 0.54 | 0.53 | 0.47 | 0.57 |
| RAD/SVL | 0.20 | 0.19 | 0.21 | 0.19 | 0.20 | 0.19 | 0.24 | 0.19 | 0.18 | 0.17 |
| TIB/SVL | 0.38 | 0.39 | 0.39 | 0.39 | 0.38 | 0.37 | 0.39 | 0.37 | 0.41 | 0.38 |
| FTL/SVL | 0.60 | 0.61 | 0.61 | 0.59 | 0.60 | 0.59 | / | 0.59 | 0.59 | 0.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



