
Submitted:
10 December 2024
Posted:
11 December 2024
You are already at the latest version
Abstract
Growing demands for environmentally-friendly and sustainable crop pest management increased the interest for biopesticides as an alternative to the synthetic chemical pesticides. This review presents the current status of bioacaricides defined as commercial biopesticide products based on microorganisms (microbial acaricides) and biologically active substances of microbial, plant or animal origin (biochemicals and semiochemicals), used in crop protection against spider mites (Tetranychidae) and other plant-feeding mites. The most important microbial bioacaricides are mycopesticides, products manufactured from living propagules of Beauveria bassiana s.l. and several other acaropathogenic fungi. Products based on avermectins and milbemycins, secondary metabolites of actinomycetes, are well-known examples of biochemicals of microbial origin. Among biochemicals of plant origin, the most widely used have been the products based on pyrethrum, obtained from the Dalmatian daisy, Tanacetum cinerariifolium (Asteraceae), and azadirachtin, obtained from the Indian neem tree, Azadirachta indica (Meliaceae). In the recent years, products based on the essential oils from aromatic plants belonging to the families Lamiaceae, Myrtaceae, Rutaceae and others, have also gained increasing importance in the market. Special emphasis in this review is given on the compatibility of bioacaricides with predatory mites from the family Phytoseiidae as natural enemies used in the integrated management of plant-feeding mites.
Keywords:
biopesticides
; bioacaricides
; plant-feeding mites
; pest management
1. Introduction
Crop plants are threatened by a variety of pests, harmful organisms that reduce their productivity, such as animal pests (insects, mites, nematodes, molluscs and some vertebrates), microbial plant pathogens (bacteria, fungi, viruses), and weeds [1]. Considering that crop losses due to the pests can be substantial (higher than 80% in some crops) pest control measures are required. The use of synthetic chemical pesticides for this purpose has increased dramatically since the mid-20th century. The application of synthetic pesticides improved crop yield and quality, boosted food security and increased farmers' income [2,3]. On the other hand, the widespreadl adoption of synthetics has also led to negative outcomes such as development of pest resistance to pesticides, adverse effects on beneficial and non-target organisms, environmental contamination and increased risks to human health [4,5].
Growing public demands for environmentally-friendly, safe and sustainable crop pest management, along with increasingly stringent pesticide regulatory requirements has boosted the interest for pesticides of biological origin – biopesticides - as an alternative to synthetic chemicals [6,7,8]. Biopesticides are not a novelty in pest control and crop protection. Several plant-based products (e.g. nicotine, pyrethrum, rotenone) were commercialized for use against animal pests in Western Europe and the United States during the 19th and early 20th centuries [9,10]. After the World War II development of biopesticides continued, albeit in the shadow of a large-scale production and use of synthetic chemical pesticides. The most important commercialized biopesticides included the products based on the entomopathogenic bacterium Bacillus thuringiensis Berliner, fermentation products from soil actinomycetes, mycopesticides based on the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff), as well as azadirachtin and other plant-based products derived from the neem tree (Azadirachta indica A. Juss). Despite ongoing development, biopesticides had a minor role in crop protection. However, since the late 20th anderaly 21th centuries, the biopesticide sector has experienced faster growth in the global market, compared to the sector of synthetic pesticides [6,7,11].
The family of spider mites (Tetranychidae) includes several major pests of crop plants, which the most important being the two-spotted spider mite, Tetranychus urticae Koch, the citrus red mite, Panonychus citri (McGregor), and the European red mite, Panonychus ulmi (Koch). Economically important pests are also found in other mite families, such as Eriophyidae [citrus rust mite, Phyllocoptruta oleivora (Ashmead), and coconut mite, Aceria guerreronis Kiefer], Tarsonemidae [broad mite, Polyphagotarsonemus latus (Banks), and cyclamen mite, Phytonemus pallidus (Banks)], and Tenuipalpidae (flat mites, Brevipalpus spp).
Acaricides, pesticide products used against plant-feeding mites, remain a vital component of integrated pest management (IPM) programs in crop protection. It should be noted that a considerable number of active substances used against mites are actually insecto-acaricides, i.e. their spectra of toxic activity include both insects and mites. This overlap occurs because these substances typically act on molecular target sites common to both insects and mites [12,13,14,15]. In this review we use the term acaricide to refer to all pesticides intended to control mites, regardless of whether they are also labeled for insect control. In the context of IPM, the crucial point is acaricide selectivity i.e. their compatibility with natural enemies (predatory mites and insects) used as biological control agents (BCA) of tetranychid and other plant-feeding mites. Among contemporary acaricides there are not so many products of biological origin. Considering recent trends in global pesticide market, however, an increase in the number of acaricidal products of biological origin (bioacaricides) could be expected. This review will focus on properties and effects of contemporary bioacaricide products intended for use in crop protection. To do the review on bioacaricides, however, we first need to clarify what we actually talk about when we talk about biopesticides in the modern world?
2. Biopesticides in the Modern World
2.1. Definitions and Classifications
There is no global consensus on the definition of the term biopesticide (Figure 1). The Organisation for Economic Co-operation and Development (OECD) distinguished four types of biological pesticides as products used for pest control: microbial pesticides (microbials)- microorganisms (bacteria, fungi, viruses etc.) and any associated metabolites to which the effects of pest control are attributed; botanical pesticides (botanicals) - active substances obtained by processing plant material (plant extracts and essential oils, and/or their components); semiochemicals - chemical substances emitted by animals, plants, and other organisms that evoke a response of individuals of the same or other species (allelochemicals, pheromones); and invertebrates - insects, mites and other animals as the BCA used in pest control [16,17,18,19]. The Food and Agriculture Organization of the United Nations (FAO) and the World Health Organization (WHO) define biopesticides as substances derived from nature that may be formulated and applied in a manner similar to chemical pesticides. This definition includes microbials, botanicals and semiochemicals, but not invertebrates [20].
On the other hand, the Environmental Protection Agency of the United States (US EPA) distinguishes three classes of biopesticides (defined as pesticides derived from natural materials): microbial pesticides (microorganisms as active ingredients); biochemical pesticides (natural substances that control pests by non-toxic action), and plant incorporated protectants (pesticidal substances that plants produce from genetic material that has been added to the plant). Biochemical pesticides include insect pheromones, growth regulators, repellents, attractants, induced resistance promoters, suffocating agents, desiccants [21,22]. The US EPA has established a special committee to determine which substances meet the criteria to be classified as biochemical pesticides. Considering their toxic action, microbial metabolites produced at the industrial scale (e.g. fermentation products of soil actinomycetes) and some botanicals (e.g. pyrethrum) are not included in biochemical pesticides. This class corresponds to semiochemicals and a part of botanicals from the OECD and FAO-WHO definitions (Figure 1), with the exception of substances of animal origin and some other natural substances.
In China, biopesticides are divided into five types: microbial pesticides, plant-derived pesticides, biochemical pesticides, agricultural antibiotics, and natural enemies [23]. The first, second, and fifth types match the OECD's first, second and fourth types, while agricultural antibiotics match microbial metabolites produced at the industrial scale (Figure 1). Biochemical pesticides are defined similarly to the U'S EPAs classification, with the difference that plant-based growth regulators (e.g. azadirachtin) are included to the second type.
In the European Union (EU) biopesticides are not formally recognized as a distinct category. The EU legislation recognizes microorganisms as one type of active substance used in plant protection products [24]. In addition to microorganisms, the EU Pesticide Database [25] includes active substances that can be classified as botanicals, semiochemicals, or microbial metabolites produced at the industrial scale (Figure 1). The types of low-risk substances and basic substances are also recognized in the EU legislation on plant protection products [24] and many of these substances belong to biopesticides [26].
According to a widely accepted definition, proposed by Bailey et al. [27] and Chandler et al. [28] biopesticides are mass-produced and biologically based pest control products that fall into three main categories: microorganisms (microbial biopesticides), semiochemicals, and biochemicals (natural substances of microbial, plant, and animal origin with toxic action). The last category, with the exception of few animal-derived biochemicals, matches botanicals and microbial metabolites produced at the industrial scale (Figure 1). Taking into account what is common to all above definitions, biopesticides can be broadly defined as pesticides manufactured from living organisms and their biologically active products.
2.2. Global Market, Advantages and Constraints
Due to the lack of a globally agreed definition of biopesticides, as well as various methods of data collection and processing, estimates of the global biopesticide market can vary widely. An additional challenge arises from the difficulty in separating data on biopesticide use in public health programs from use in crop protection. Nevertheless, it is clear that the biopesticide sector represents a small portiont of the global pesticide market, as well as that this sector has been growing rapidly: from a mere 1% of global market in 1998 to appoximately 10% in 2022 [6,8,29]. This increased adoption of biopesticides is attributed to technological advancements in this sector, as well as restrictions on the use of synthetic pesticides, broader implementation of the IPM programs and rising demands for organic food in developed countries [8,30].
Low mammalian toxicity, zero or low harvest and re-entry restrictions, higher safety to human health, non-target organisms and the environment, lower risk of target pests developing resistance, have been the most frequently highlighted advantages of biological pesticides over the synthetic chemical ones [6,28]. Although there are good grounds for such general perception, there are also data indicating that a comparison of biopesticides and synthetics isn't simply a black and white issue. Notable examples include the toxic and adverse sublethal effects of the fermentation products of soil actinomycetes (abamectin, spinosad), botanicals (essential oils, neem-based products) and mycopesticides on polinators and natural enemies of insect and mite pests [31,32,33,34], as well as the harmful effects of fungal biopesticides on mycorrhizal and saprophytic fungi, soil bacteria, plants [35]. Another often neglected issue is that biopesticides are also vulnerable to resistance evolution. According to the Arthropod Pesticide Resistance Database (APRD), 28 insect pest species and 19 insect and mite pest species have developed resistance to spinosad and abamectin, respectively [36]. Resistance to antibiotics (another group of fermentation products of actinomycetes) has been recorded in some plant pathogenic bacteria [37], while several lepidopteran pest species have developed resistance to Bt-toxins [38,39].
Further expansion of global biopesticide market depends on successful overcoming of various constraints. The most important technical constraints include lower and variable efficacy of biopesticide products and their greater susceptibility to unfavourable environmental conditions, compared to the synthetics, and the issue of quality and availability of resources [28,30]. From a socio-political point of view, regulatory barriers and inconsistent policies and regulatory procedures across regions, along with limited awareness of biopesticides among the end-users [8,30,40] are the major constraints to the wider use of biopesticides.
3. Bioacaricides in Contemporary Crop Protection
Within the extensive recent scientific literature on biopesticides there have been few reviews specifically dealing with bioacaricides in crop protection. Flamini [41] published a comprehensive review of plant-derived compounds used against harmful species from the subclass Acari in agriculture, veterinary and human medicine covering the last decade of the 20th century. Other reviews have mainly focused on the use of bioacaricides against T. urticae. Maniania et al. [42] reviewed the use of fungal pathogens in the inundative, conservation and classical biological control of T. urticae and Tetranychus evansi Baker & Pritchard, the red tomato spider mite. Attia et al. [43] and Rincón et al. [44] reviewed research on botanicals, while Jakubowska et al. [45] briefly presented examples of plant extracts, microorganisms, and natural enemies used in the control of T. urticae.
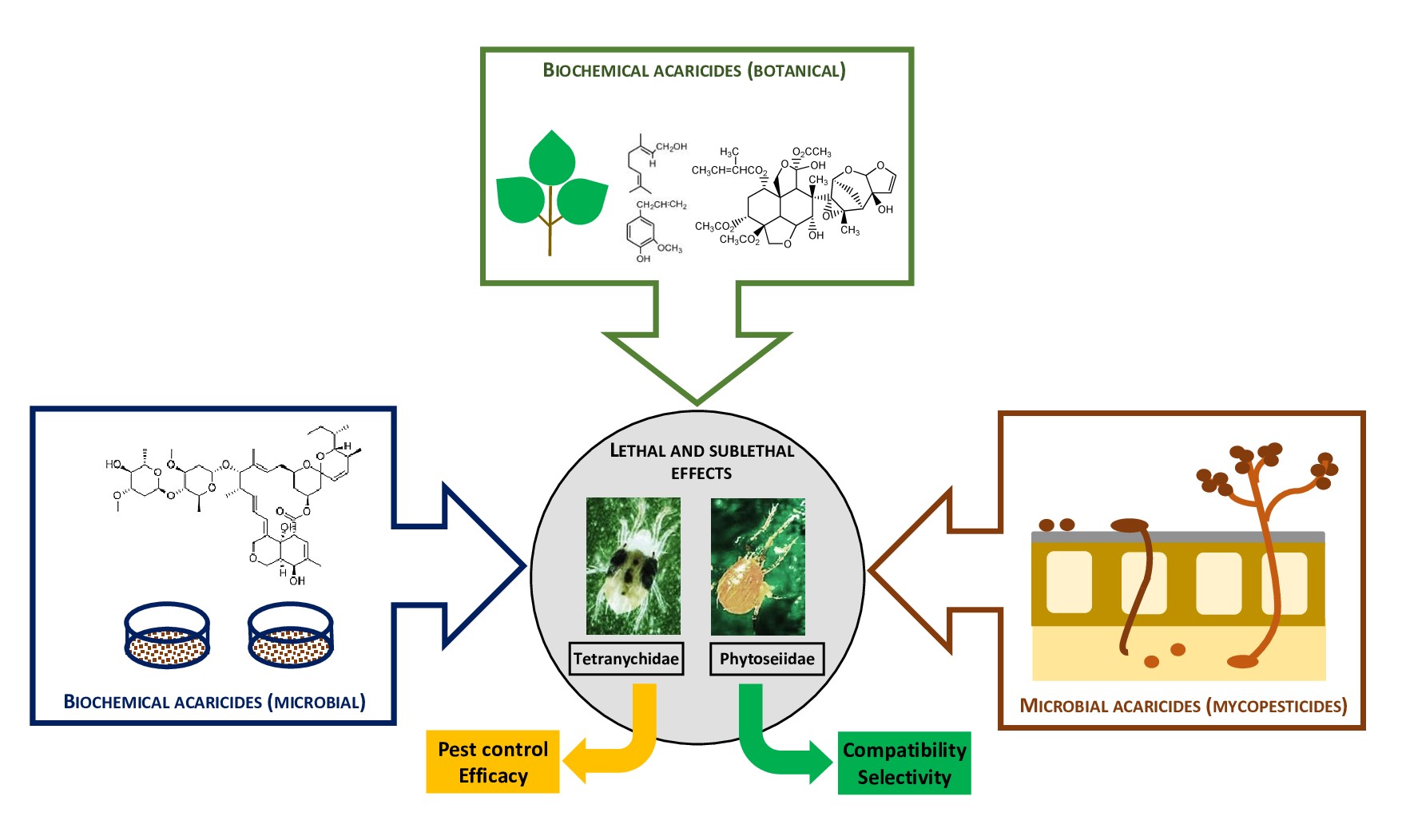
Below we provide an overview of the properties, effects, and uses of contemporary bioacaricides. They are defined as commercialized crop protection products used in the control of plant-feeding mites, based on microorganisms (microbial acaricides); biochemical active substances with toxic action (biochemical acaricides) and with non-toxic action (semiochemicals); based on their origin, biochemical acaricides are further divided into microbial biochemical acaricides and botanical biochemical acaricides. Special emphasis will be given on their compatibility with predatory mites from the family Phytoseiidae which serve as natural enemies in the integrated management of plant-feeding mites and insects.
3.1. Microbial Acaricides
Microbial biopesticides account for around 60% of the global biopesticide market and they are dominated by products based on δ-endotoxins (crystal proteins) from B. thuringiensis (Bt) [8,46] The spectrum of activity of these products includes lepidopteran, coleopteran and dipteran insect pests, but not mites. Some Bt strains are capable of producing non-proteinaceous β-exotoxins, such as thuringiensin. Besides insecticidal, thuringiensin also shows considerable acaricidal activity against spider mites [47,48,49]. Due to the mode of toxic action of β-exotoxins (inhibition of DNA-dependent RNA polymerase) which may affect mammals, their absence is a requirement for Bt products in Europe, the US and Canada. In some countries, such as Russia, β-exotoxin-containing products are still used [50,51,52].
In the shadow of Bt, two betaproteobacterial species have recently been used by Marrone Bio Innovation (USA) as the basis for development of crop protection products with insecticidal and acaricidal activity. The betaproteobacterium Chromobacterium subtsugae Martin et al., isolated from forest soil in the USA, is the source of the commercial product “Grandevo”, which contains the strain PRAA4-1T. This product is labeled for controlling insect pests fromthe orders Lepidoptera, Hemiptera and Thysanoptera, as well as plant-feeding mites. During fermentation the bacterium produces violacein and other secondary metabolites that contribute to its toxicity [8,53]. Another commercially successful betaproteobacterial species is Burkholderia rinojensis Cordova-Kreylos et al., isolated from a soil sample in Japan (strain A396). Its commercial product “Venerate” is labeled for use against several hemipteran species and plant-feeding mites. Various bacterial metabolites formed during the fermentation are involved in its toxic effect [8,54]. In addition to the USA, where these products were developed, they have also been registered in Canada, Mexico, Chile, New Zealand, and some other countries. Both products contain inactivated bacteria cells and spent fermenation media. These are slow acting products that take several days to achieve their full toxic effect. Therefore, they should be used early, when mite populations are still at a low level, with consecutive applications recommended on a 7-day schedule, preferably rotated with other products. Laboratory bioassays have shown that “Grandevo” exhibits low to moderate toxicity to T. urticae female adults [55,56,57], as well as a significant reduction in fecundity [57]. The product “Venerate” was also moderately toxic to the females [56]. Field evaluations showed moderate effectiveness of “Grandevo” in controlling P. latus on pepper and P. oleivora on orange trees [58,59], as well as T. urticae on strawberry [60].
Among microbial acaricides, mycopesticides (insecticides and acaricides) manufactured from living propagules of entomo- and acaropathogenic fungi [61,62] form a large group of commercial products. However, it should be mentioned that the majority of them are based on two fungal species, B. bassiana s.l. and M. anisopliae s.l. Other commercially important species include Isaria fumosorosea Wize, Lecanicillium muscarium (Petch) Zare & Gams, Lecanicillium lecanii (Zimmermann) Gams and Zare and Hirsutella thompsonii Fisher [46,63,64]. Their pathogenic mode of action against insects and mites is based on penetration of the host integument (by mechanical force and cuticle degrading by hydrolytic enzymes), colonization and production of toxic metabolites, leading to mycosis and death. Successful application of mycopesticides relies particularly on relative humidity (RH): their efficacy increases with raising of RH [65,66,67].
The majority of mycopesticides are intended for use in horticultural crops, such as greenhouse and field vegetables, ornamentals and fruit crops. Only a smaller portion of these products is labeled for controlling both insects and mites, while the vast majority are registered solely as mycoinsecticides. For example, more than 20 strains of B. bassiana have been used for manufacturing mycoinsecticides [63,64], yet less than one-third of them are also registered as mycoacaricides (Table 1). The target pest range of these mycoacaricides varied. Someproducts are labeled only for the control of T. urticae and other tetranychid mites, while the others are also used against Tarsonemidae (P. latus, P. pallidus), Eriophyidae (P. oleivora), and Tenuipalpidae. Certainproducts are labeled for use against all plant-feeding mites. Unlike most mycopesticides, the products based on H. thompsonii, a mite-specific fungus, are registered exclusively as mycoacaricides in India targeting A. guerreronis and various spider mites [68]. In addition, there are also some differences between countries in the intended use of products derived from the same fungal strains. For example, products based on the strains ATCC 74040, GHA and PPRI 5339 of B. bassiana and the strains Apopka 97 and FE9901 of I. fumosorosea are registered for controlling sucking insect pests and plant-feeding mites in the USA, while in the EU they are not intended to use as mycoacaricides. The only mycoinsecticide and mycoacaricide registered both in the EU and in the USA is a product based on Metarhizium brunneum (Petch) strain F52 [25,69].
Field and greenhouse trials have shown moderate to high effectiveness of mycopesticide products in controlling tetranychid and other plant-feeding mites in various countries and regions. Typically, at least two treatments at 7-14 days intervals are required for effective control, which may prove uneconomical unlessused in higher value crops. Variable effectiveness of the mycopesticides is largely attributed to their high sensitivity to ultraviolet radiation and other environmental conditions, which is less pronounced in greenhouses [75,76,77,78,79,80,81]. In addition to appropriate formulations, the sustainability of mycopesticide use in the control of plant-feeding mites can be improved by including them in the IPM programs. Within these programs, mycopesticides can be combined with other bioacaricides or synthetic acaricides, as well as predatory mites and insects [42,46,63,82].
3.2. Biochemical Acaricides
3.2.1. Microbial Biochemical Acaricides
Secondary metabolites produced by fermentation from soil actinomycetes have been one of the most important sources for mass production of biopesticides. Products based on avermectins and milbemycins, macrocyclic lactones obtained from Streptomyces avermitilis (Burg et al.) Kim and Goodfellow and Streptomyces hygroscopicus subsp. aureolacrimosus Takiguchi et al., respectively, are very well-known examples of biopesticides commercially successful at the global level [6,12]. Abamectin (a mixture of avermectins B1a and B1b) and milbemectin (a mixture of milbemicins A3 and A4) are used against tetranychid, eriophyid and tarsonemid mite pests. Abamectin is also labeled for use against a broad range of insect pests, while milbemectin is primarily used against mites. They have neurotoxic mode of action - allosteric modulation of glutamate-gated chloride channels, causing paralysis of mites and insects [15]. Abamectin is marketed under a wide variety of trade names, such as “Agri-Mek”, “Dynamec” and “Vertimec” from Syngenta, the largest manufacturer. Mitsui Chemicals, company that developed milbemectin, sells it under the trade names “Milbeknock” and “Koromite”. Together, abamectin and milbemectin occupy third place in the global market of products targeting insects and mites [83]. Despite being sensitive to photolytic degradation, these biochemical acaricides provide residual activity due to rapid penetration into the leaves and translaminar movement. Their residual action against mites can be further improved by mixing with various spray adjuvants [59,84,85].
Another globally successful biopesticide is spinosad, a mixture of spinosyns A and D, derived from the fermentation of actinomycete Saccharopolyspora spinosa Mertz and Yao. It is also a neurotoxic compound, acting as an allosteric modulator of nicotinic acetylcholine receptors in the insect central nervous system [15]. Spinosad is highly effective against a wide range of insect pests, such as thrips, caterpillars and leaf miners [7]. Some of its products are also labeled for spider mite control, such as “Conserve SC” (Corteva Agriscience). The recommended application rate for controlling spider mites with this product (180 g a.i/l) is 2-3 times higher than those for insect control. The acaricidal properties of spinosad have been supportedby several bioassays. In acute toxicity bioassays with T. urticae females [66,86,87,88] the LC50 estimations were found to be far below the recommended rate. Van Leeuwen et al. [86] demonstrated the possibility of systemic use of spinosad to control spider mites, applied to the roots of tomato plants grown in rockwool. The instructions for using the product labeled for spider mites recommend an early application (before mite populations have become abundant), reapplication after several days and addition of nonionic adjuvant to improve spray coverage. Poor spray coverage and late application may, at least partially, explain variable results of spider mite control in field trials with spinosad [89,90].
Due to their toxic action, the US EPA doesn't consider abamectin, milbemycin and spinosad as biopesticides. Also, they are not addressed in the OECD and FAO-WHO guidelines on biopesticides [17,20,22,91]. On the other hand, Bailey et al. [27] and Chandler et al. [28] considered them as biochemical pesticides and they are classified as biopesticides in India and China [23,30].
3.2.2. Botanical Biochemical Acaricides
The history of biopesticides of plant origin (botanicals) intended to control insects and mites has been largely a tale of two plants: the Dalmatian daisy, Tanacetum cinerariifolium (Trevir.) Sch. Bip. (Asteraceae), and the Indian neem tree, Azadirachta indica A. Juss. (Meliaceae). The most widely used botanicals are commercial products that have come from these two plant species. Since the early 2000s, essential oil-based products have also gained increasing importance in the market of botanicals [10,40,62].
Dried flowers of T. cinerariifolium are a source of oleoresin pyrethrum, a crude flower dust containing a mixture of six related esters as active ingredients. The most important ones are pyrethrin I (an ester of pyrethrolone and chrysanthemic acid) and pyrethrin II (an ester of pyrethrolone and pyrethric acid). Technical grade pyrethrum used in formulating commercial products contains 20-25% pyrethrins [10]. Pyrethrins are neurotoxic: they act as sodium channel modulators keeping the channels open and causing hyperexcitation [15]. Although it has been on the market for more than a century, pyrethrum is still one of the leading botanicals, partly due to its non-agricultural use (structural pest control, public health), and because of the relatively low toxicity to humans and mammals [92,93]. Eastern African countries had long been world leaders in the cultivation of T. cinerariifolium and pyrethrum production before Australia (Tasmania) took the lead in the past two decades [40,94]. Pyrethrum-based products are recommended for use against a range of insect and mite pests in home gardens, as well as in commercial crop and ornamental production (Table 2).
With regard to A. indica, its seeds are a rich source of neem oil that contains a numerous active compounds, including the four most important tetranortriterpenoids (limonoids): azadirachtin, salannin, meliantriol and nimbin. Azadirachtin is a slow acting insect and mite growth regulator and a potent feeding and oviposition deterrent. Its exact mode of action on growth and developmental processes at the molecular level is still unclear [97,98,99,100,101]. Various types of products based on azadirachtin and neem oil are currently available on the market. In addition to insect pests, most products are also labeled for the control of mites, primarily tetranychids (Table 2). A plethora of products have been registered in India, where the neem tree is native. The current global status of azadirachtin-based biopesticides is primarily due to the registration of a great number of such products in many developing countries of Africa and Latin America (as a consequence of the introduction of the neem trees), as well as in China [23,40,97]. Azadirachtin-based products have shown moderate to high efficacy in controlling of spider mites and other plant-feeding mites [102,103,104,105]. The formulation type and production technology have a great influence on their effectiveness [10].
A great increase in research interest in botanicals over the past two decades has resulted in moderate success in commercialization of new products intended to control insect and mite crop pests [40,94,96]. Far behind pyrethrum and azadirachtin/neem, the third place in global market is held by crop protection products based on plant essential oils and/or their biologically active constituents. Essential oils, complex mixtures of volatile secondary metabolites, are obtained by distillation from aromatic plant species belonging to the families Lamiaceae, Myrtaceae, Rutaceae, and several others. Terpenoids (monoterpenes and sesquiterpenes) and, to a lesser extent, phenylpropanoids are the main constituents of essential oils. Some terpenoids can also be obtained by economically viable industrial synthesis. Essential oil-based commercial products contain either a single oil/constituent or a mixture of oils/constituents. The products act rapidly by direct contact and the vapour phase. Owing to their volatile nature, they are effective fumigants in closed spaces. Their lethal effects on insects and mites are most likely a consequence of neurotoxic action of the constituents on one or more molecular targets, such as acetylcholinesterase, octopamine receptors and GABA-gated chloride channels. Being often complex mixtures of terpenoids, the toxic action of essential oils appears to be a result of synergy among their constituents. In addition to the lethal effect, essential oil-based products also cause behavioral (repellent and deterrent) effects [10,40,94,96,106].
The most common essential oils that have been used for manufacturing commercial products intended to control mite pests are rosemary oil, peppermint oil, orange oil, tea tree oil, cinnamon oil (Table 2). The acaricidal activity of the essential oil-based products is based on their constituents such as monoterpenes carvacrol, 1.8-cineole, citronellol, d-limonene, eugenol, geraniol, menthol, and phenylpropanoid cynnamaldehyde. One terpenoid can have several botanical sources [40,96]. It is interesting, as Isman [94] pointed out, that US EPA includes into biochemical biopesticides, defined as non-toxic substances, several monoterpenes (citronellol, eugenol, geraniol, methol) and essential oils (eucalyptus oil, orange oil, tea tree oil) even though they are clearly neurotoxic to insects and mites.
The essential oil-based products are usually recommended for use against a range of insect and mites, primarily tetranychids. In some products, essential oil is combined with various non-volatile plant (vegetable) oils, such as castor oil and cottonseed oil. Some of these products (“Akabrown”, “Mitexstream”, “Biomite”) are only labeled for the control of mite pests. Greenhouse and field evaluations have shown that these products can control mites as effectively as other botanical biochemical acaricides and even synthetic chemical acaricides [107,108,109,110]. Considering their low persistence, repeated treatments are usually recommended. On the other hand, the extended efficacy of essential oil-based products observed in some field trials suggests that their behavioral effects may contribute considerably to the overall impact [96].
One of the most important recently introduced biopesticide products is “Requiem”, based on monoterpenes α-terpinene, d-limonene and p-cymene as active ingredients (Table 2). It is a contact insecticide and acaricide with strong repellent and deterrent activity [111,112,113]. The product was initially manufactured from the extract of American wormseed, Chenopodium ambrosioides L. near ambrosioides (Chenopodiaceae), containing these three monoterpenes as the major constituents. Since Bayer Crop Science took over the product, it has been made from the blend of synthesized monoterpenes, intended to mimic the naturally-occurring extract [40,96]. The product is considered for use against a broad range of plant-feeding mites and insect pests, mostly hemipteran.
Some newer botanical biochemical acaricides have alkaloids as active ingredients (Table 2). Products based on the extracts from Sophora flavescens Aiton (Fabaceae), a Chinese medicinal herb, contain matrine, oxymatrine, and other quinolizidine alkaloids. Besides insect pests, these products have proven effective against spider mites and flat mites [78,101,114,115,116]. Veratrine and related cevadine-type alkaloids are active ingredients in the products extracted from Veratrum nigrum L. (Melanthiaceae), an herb also used in Chinese traditional medicine. Another recent product is “Captiva”, based on a combination of canola oil and garlic oil with capsaicin, an alkaloid found in the oleoresin of red pepper, Capsicum annuum L. (Solanaceae). The product is labeled for use against spider mites, P. oleivora, P. latus, Brevipalpus phoenicis (Geijskes), and a broad range of insect pests [40,94,96].
In addition to combining with essential oils, vegetable oils are also used independently to formulate commercial biopesticide products. Canola oil and rapeseed oil are two vegetable oils widely used as biopesticides (Table 2). They come from the same botanical source (Brassica napus L., Brassica rapa L.), but canola oil is produced from cultivars with low content of erucic acid and glucosinolates. The main active constituents of these oils are fatty acids, such as oleic acid and linoleic acid [7,117]. Oleic acid is also the main constituent of both cottonseed oil and safflower oil, while ricinoleic acid is the main constituent of castor oil (Table 2). Prevention of gas exchange (suffocation) is widely accepted theory of the major mode of action of plant oils against soft-bodied insects and mites, while the interference of fatty acids with the cell membrane constituents, leading to its disruption, is of lesser importance [7]. However, Takeda et al. [118] indicated inhibition of larval hatching, instead of suffocation, as the major mode of action of plant oils against T. urticae eggs. Canola oil and rapeseed oil are also used to formulate commercial biopesticide products in combination with other botanicals, such as pyrethrum and azadirachtin/neem [23,25,69].
Botanical biochemical acaricides can reduce mite populations to the level comparable to those achieved by synthetic chemical acaricides. However, due to the low persistence of botanicals, achieving such results requires application of relatively high concentrations and repeated treatments. Tank mix application of botanical biochemical acaricides with synthetic chemical products is one of the ways to increase their effectiveness. Another way is to combine two botanicals or botanicals and microbials. In order to achieve long-term and sustainable use of biopesticides, it is necessary to include them into the IPM programs [40,95,96,100].
3.3. Semiochemicals as Bioacaricides
Semiochemical biopesticides include commercial crop protection products based on behavior-modifying signalling chemicals of plant or animal origin that carry information between individuals, intended for the control of insect and mite pests. They are divided into pheromones, inducers of behavioral response in the receiving individuals of the same species, and allelochemicals, involved in interspecific interactions [28]. The most widely used semiochemical biopesticides are insect sex pheromones: a lot of pheromone-based products are applied in mating disruption, mass trapping, attract-and-kill, and other control strategies [119].
Unlike the situation with insects, sex pheromones of plant-feeding mites have negligible importance as biopesticides. Nerolidol and farnesol, two structurally related sesquiterpenes found in immature T. urticae females, act as sex pheromones to attract males for mating [6,120]. They are also constituents of some plant essential oils. As active ingredients of the product “Biomite” (Tab. 2) nerolidol and farnesol increase activity of mites and consequently their exposure to co-formulated toxic monoterpenes geraniol and citronellol.
Allelochemicals are divided into allomones, kairomones or synomones, depending on whether the response is favourable to the emitter, the receiver, or both. Plants produce various secondary metabolites (allomones) to repel or deter insects and mites. Repellent effects of essential oils and feeding deterrent effects of azadirachtin/neem are well known, but these effects are of secondary importance to the control of crop pests [96,100]. There have been no commercially successful products based unequivocally on repellency or feeding deterrence as the major activity. Kairomones are volatile plant secondary metabolites, induced by plant-feeding insects and mites, that attract their natural enemies as the receivers. Methyl salicylate, one of the best-known attractants, is synthesized and available as a slow-release dispenser (commercial product “PredaLure”, manufactured by AgBio, USA). The product demonstrated effective attraction of phytoseiid predators in several field trials [120].
4. Side Effects of Bioacaricides on Predatory Mites
The IPM paradigm in crop protection defines pesticide selectivity as compatibility of pesticides with natural enemies of plant-feeding pests, and it is based on evaluation of detrimental effects of pesticides on natural enemies as the BCAs of the pests [121]. The most important BCAs of spider mites and other plant-feeding mites, as well as some insect pests (whiteflies, thrips) are predatory mites of the family Phytoseiidae. Among more than 30 commercialized phytoseiids, the major species are Amblyseius swirskii Athias-Henriot, Neoseiulus californicus (Garman), Neoseiulus cucumeris (Oudemans) and Phytoseiulus persimilis Athias-Henriot, which account for 60% of the global market [122]. The selectivity of pesticides with the BCA has two aspects. Physiological selectivity is based on toxicokinetic and toxicodynamic mechanisms that ensure a lower sensitivity of the BCAs compared to harmful species. Ecological selectivity is the result of limiting the exposure of the BCA to pesticides in time and space [121,123].
Various methods have been used for the evaluation of pesticide selectivity. Since the 1970s the Working Group (WG) “Pesticides and Beneficial Organisms” of the International Organization for Biological Control – Western Palearctic Regional Section (IOBC-WPRS) has developed standardized methods for over 30 beneficial species, natural enemies of insect and mite crop pests, and has tested nearly 400 pesticides [124,125]. The IOBC methods constitute a programme for sequential testing of pesticide effects on beneficials in which the decision to perform field trials depends on the outcome of laboratory bioassays. A harmless classification implies that further testing is not necessary i.e. pesticide is considered compatible [126,127]. Initial toxicity bioassay is carried out on glass plates where the most susceptible life stage is exposed to fresh deposite of pesticides applied at the highest recommended rates (the “worst case” scenario). The other laboratory bioassays include modification such as exposure of less susceptible stages on leaves or leaf discs (extended laboratory bioassay), exposure to field aged pesticide residues (persistence bioassay) [124,128].
In the laboratory bioassays, the IOBC ranking for classifying harmfulness is based on reduction in beneficial capacity, as a consequence of mortality and reduced fecundity. It is expressed by the coefficient of toxicity or the total effect (E, %), calculated using the formula: E = 100% - (100% - M) × R, where M is the percentage of mortality corrected for mortality in the control, and R is the ratio between the number of eggs produced by treated females and the number of eggs produced by females in the control [129]. Instead of the number of eggs laid (fecundity), in some studies [130,131,132] the number of hatched eggs (fertility) was recorded.
The IOBC ranking includes four categories, based on the values of E (%): 1 = harmless (E < 30%), 2 = slightly harmful (E = 30-80%), 3 = moderately harmful (E = 80-99%), and 4 = harmful (E > 99%). If a pesticide is classified in categories 2-4, further testing in semi-field and field trials is necessary. To calculate E in these trials the effects of pesticides on population size and dynamics are recorded, and a pesticide is considered harmless (E < 25%) slightly harmful (E = 25-50%), moderately harmful (E = 50-75%), and harmful (E > 75%) [128].
The IOBC-WPRS database on effects of plant protection products on beneficial arthropods (set up in the early 2000s, currently under revision) compiled data concerning the compatibility of pesticides and beneficials, published in scientific journals and proceedings of the WG conferences, as well as in the reports on regulatory testing in the European Union. The database includes 1768 test results on the selectivity of 379 pesticides to 16 phytoseiid species [125,133]. Acaricides are represented with 567 results for 98 compounds. Besides acaricides, other pesticides have also been evaluated, including 143 herbicides and 111 fungicides, as well as 27 insecticides. With 346 tested pesticides (76% of total results and 60% of acaricide results) the species Typhlodromus pyri Scheuten stands out among phytoseiids (followed by P. persimilis, Amblyseius andersoni (Chant) and Euseius finlandicus (Oudemans). The predominance of T. pyri is mainly a consequence of its status as an indicator species since over 86% of the results originate from reports on the regulatory testing of pesticides in European Union [133].
The database contains test results of 11 bioacaricides and 10 phytoseiid mites with a total of 65 results (55 from laboratory bioassays and 10 from field trials), 69% of which are for T. pyri. Among bioacaricides, more than a half of the results refer to abamectin, spinosad and B. bassiana (Table 3a). In laboratory bioassays abamectin was shown mostly to be moderately harmful to harmful, with the expected variation in results depending on predator species and/or abamectin concentrations. The persistence bioassay showed that compatibility could be achieved by exposure of N. californicus to residues aged at least 15 days. In field trials abamectin was slightly harmful to A. swirskii and harmless to T. pyri. Spinosad results were similar to abamectin results, while B. bassiana was slightly harmful or harmless to T. pyri and other phytoseiids (Table 3b).
Most of the data on compatibility of acaricides with phytoseiid mites have not come from the IOBC-WPRS database. A number of papers were not included in the database because they used methods that more or less deviate from the standard characteristics [124,125]. The experimental design of many other studies, carried out almost exclusively on leaves or leaf discs, have included new species of phytoseiid mites, different developmental stages, a greater number of acaricide concentrations, new ways of exposure and new toxicity parameters and endpoints [123,134]. Concentrations lower than the recommended ones have also been tested, and some of the studies have been carried out to estimate LC50 and the other LCs. In addition to mortality for eggs, juveniles and/or adults, many studies include assessment of sublethal effects of pesticides on developmental time, fecundity, fertility, and other life history traits of phytoseiid mites. Different ways of exposure to pesticides have been applied in the bioassays individually or in combinations, such as triple exposure: a combination of direct treatment, residual exposure and feeding of predators with treated prey. Some studies also included evaluation of comparative toxicity predator-prey [130,135,136,137,138]. In addition to the total effect and the IOBC classification of toxicity, new parameters and criteria for compatibility evaluation were introduced. Beers & Schmidt [139] and Schmidt-Jeffris & Beers [140] proposed that the effect of acaricides is expressed by calculating the cumulative effect on the survival of treated females, their fecundity and fertility (egg hatching), as well as on the survival of hatched larvae. For predator-prey comparisons, the selectivity index is calculated as the difference between the cumulative effects of acaricides on predator and prey.
Recently, the evaluation of compatibility of pesticides with phytoseiid mites has increasingly been based on laboratory bioassays that estimate their effect at the population level. In the demographic bioassay the effect is expressed by a change in the value of the intrinsic rate of population increase (rm) as a consequence of the pesticide impact on developmental time, fecundity, longevity and other life history traits of the mites. The demographic bioassay is based on construction of the life table from data on survival and reproduction. Two types of the life table have been used: the female fertility life table, constructed from data on female survival and fertility (production of female offspring) [141,142] and the age-stage two-sex life table, which includes juvenile development and survival data for both sexes, and female fecundity [143].
Examples of the aforementioned studies are presented in Table 4, Table 5, Table 6 and Table 7. Most of the results of these studies refer to abamectin, spinosad and azadirachtin. In addition to the four major commercialized species of global importance (A. swirskii, N. californicus, N. cucumeris and P. persimilis) these studies have included the other important phytoseiid species, such as Galendromus occidentalis (Nesbitt), Neoseiulus womersleyi (Schicha), Phytoseiulus macropilis (Banks) and Phytoseiulus longipes Evans. Both commercialized and native strains have been evaluated. Mycopesticides based on B. bassiana and H. thompsonii were mostly compatible with phytoseiids (Table 4).

Table 4.
Examples of evaluation of the compatibility of microbial bioacaricides (mycopesticides) and phytoseiid mites in various laboratory bioassays; Comp = compatibility conclusion: + positive (compatible); - negative (not compatible).
Table 4.
Examples of evaluation of the compatibility of microbial bioacaricides (mycopesticides) and phytoseiid mites in various laboratory bioassays; Comp = compatibility conclusion: + positive (compatible); - negative (not compatible).
| Mycopesticides | Phytoseiid mites | Methodology | Comp | References | |||
| Exposure | Endpoints | ||||||
| Beauveria bassiana GHA | Amblyseius swirskii | c | rc; Td-re | Sf, Fec, Fer | + | [144] | |
| Beauveria bassiana ATCC 74040 | Neoseiulus californicus | n | rc; Td-re | Se, Sf, Fec, Fer, E ● | - | [130] | |
| Phytoseiulus persimilis | n | rc; Td-re | Se, Sf, Fec, Fer, E ● | + | [132] | ||
| Hirsutella thompsonii | Phytoseiulus longipes | n | rc; Td-re | Sf, Fec, Fer, E, IOBC | + | [145] | |
| rc; Tre (4-31) | + Tre (10) | ||||||
c = commercial strains, n = native populations; rc = recommended concentrations; Td-re = direct (topical) treatment and residual exposure; Tre (x-y) = exposure to residues aged x-y days; Se = egg hatching; Sf = adult female survival; Fec = fecundity (eggs produced by treated females); Fer = fertility (hatched eggs produced by treated females); E = the coefficient of toxicity [129]; IOBC = classification of toxicity according the IOBC-WPRS guidelines [128]; ● comparative toxicity predator – prey.
Abamectin and milbemectin proved to be not compatible after direct treatment followed by residual exposure and consumption of treated prey. Abamectin was considered compatible mostly when predatory mites were exposed to its aged residues. This is an example of ecological selectivity that can be achieved by temporal separation between acaricides and phytoseiids [123]. Spinosad results were variable; similar to abamectin, its low persistence allowed ecological compatibility (Table 5). Azadirachtin was shown to be compatible with phytoseiids, regardless of the route of exposure, while pyrethrum was not compatible after direct treatment and residual exposure. Similar to abamectin and spinosad, low persistence of oxymatrine allowed its compatibility (Table 6). In research that includes LC50 as one of the bioassay endpoints [137,146,147] conclusions were made based on the comparison of the LC50 values with the recommended concentrations.
Table 5.
Examples of evaluation of the compatibility of microbial biochemical acaricides and phytoseiid mites in various laboratory bioassays; Comp = compatibility conclusion: + positive (compatible); - negative (not compatible); → further research needed .
Table 5.
Examples of evaluation of the compatibility of microbial biochemical acaricides and phytoseiid mites in various laboratory bioassays; Comp = compatibility conclusion: + positive (compatible); - negative (not compatible); → further research needed .
| Bioacaricides | Phytoseiid mites | Methodology | Comp | References | ||
| Exposure | Endpoints | |||||
| abamectin | Amblyseius swirskii | n | rc, rc+/-; Td-re | Se, Sl, Sf, Fec, LC50 | → | [146] |
| Euseius scutalis | n | rc, rc+/-; Td-re | Se, Sl, Sf, Fec, LC50 | → | [147] | |
| Galendromus occidentalis | c | rc; Tre (3-37) | Sf, Fec, Fer, E, IOBC | + Tre (6) | [148] | |
| Neoseiulus barkeri | c | rc, rc+/-; Tre (0) | Sf, Fec, Fer, LC50, E, IOBC ● | + | [149] | |
| Neoseiulus californicus | c | rc; Tre (0-21) | Sf | + Tre (7) | [150] | |
| n | rc; Tre (0-21) | Sf ● | + Tre (7) | [151] | ||
| c | rc; Td-re + Tp | Sf, Fec, Fer, SI ● | - | [138] | ||
| Neoseiulus cucumeris | n | rc, rc+/-; Tre (0) | Sf, LC50 ● | + | [152] | |
| c | rc; Tre (0) | Sf | - | [153] | ||
| Neoseiulus fallacis | c | rc; Td-re + Tp | Sf, Fec, Fer, SI ● | - | [138] | |
| Phytoseiulus longipes | n | rc; Td-re | Sf, Fec, Fer, E, IOBC | - | [145] | |
| rc; Tre (4-31) | + Tre (10) | |||||
| Phytoseiulus persimilis | c | rc; Tre (3-37) | Sf, Fec, Fer, E, IOBC | + Tre (14) | [148] | |
| n | rc; Td-re | Sf, Fec, Fer, E, IOBC | - | [131] | ||
| c | rc; Tre (0-21) | Sf | + Tre (14) | [150] | ||
| c | rc; Td-re + Tp | Sf, Fec, Fer, SI ● | - | [138] | ||
| milbemectin | Neoseiulus womersleyi | n | rc; Td-re | Sel-a, Sf, Fec ● | - | [154] |
| Phytoseiulus persimilis | n | rc; Td-re | Sel-a, Sf, Fec ● | - | [155] | |
| spinosad | Amblyseius swirskii | n | rc, rc+/-; Td-re | Se, Sl, Sf, Fec, LC50 | - | [156] |
| Galendromus occidentalis | n | rc; Td-re + Tp | Se, Sf, Fec | + | [136] | |
| Kampimodromus aberrans | n | rc; Tre (0) | Sf, Fec, Fer, E | - | [157] | |
| Neoseiulus cucumeris | c | rc; Tre (0-6) + Tp | Sf, IOBC | + Tre (4) | [158] | |
| Neoseiulus fallacis | n | rc, rc+/-; Td-re + Tp | Se, Sf, Fec, LC50 | → | [137] | |
| n | rc; Tre (0), Tp | Sel-a, Sf, Fec | - | [159] | ||
| Phytoseiulus persimilis | n | rc; Td-re | Sf, Fec, Fer, E, IOBC | + | [131] | |
| Typhlodromus montdorensis | c | rc; Tre (0-6) + Tp | Sf, IOBC | + Tre (5) | [158] | |
c = commercial strains, n = native populations; rc = recommended concentrations; rc+/- concentrations higher/lower than rc; Td-re = direct (topical) treatment and residual exposure; Tre (0) = exposure to fresh residues; Tre (x-y) = exposure to residues aged x-y days; Td = direct (topical) treatment and transfer to untreated surface; Tp = consumption of treated prey; Se = egg hatching; Sel-a = egg hatching and larval survival to the adulthood; Sl = larval survival; Sl-a = larval survival to the adulthood; Sf = adult female survival; Fec = fecundity (eggs produced by treated females); Fer = fertility (hatched eggs produced by treated females); E = the coefficient of toxicity [129]; IOBC = classification of toxicity according the IOBC-WPRS guidelines [128]; SI = selectivity index; ● comparative toxicity predator – prey.
Table 6.
Examples of evaluation of the compatibility of botanical biochemical acaricides and phytoseiid mites in various laboratory bioassays; Comp = compatibility conclusion: + positive (compatible); - negative (not compatible).
Table 6.
Examples of evaluation of the compatibility of botanical biochemical acaricides and phytoseiid mites in various laboratory bioassays; Comp = compatibility conclusion: + positive (compatible); - negative (not compatible).
| Bioacaricides | Phytoseiid mites | Methodology | Comp | References | ||
| Exposure | Endpoints | |||||
| azadirachtin | Amblyseius andersoni | n | rc, Td-re | Sf, Fec, E | + | [160] |
| Neoseiulus barkeri | c | rc, Tre (0) | Sf, Fec, Fer, E, IOBC | + | [149] | |
| Neoseiulus californicus | n | rc; Td-re | Se, Sf, Fec, Fer, E ● | + | [130] | |
| c | rc; Td-re | Sf, Fec, Fer | + | [161] | ||
| Neoseiulus cucumeris | c | rc; Td, Tre (0) + Tp | Sel-a, Sf, Fec | + | [162] | |
| Phytoseiulus longipes | n | rc; Td-re | Sf, Fec, Fer, E, IOBC | + | [145] | |
| rc; Tre (4-31) | + Tre (4) | |||||
| Phytoseiulus macropilis | c | rc; Td-re | Sf, Fec, Fer | + | [161] | |
| Phytoseiulus persimilis | c | rc; Td, Tre (0) + Tp | Sel-a, Sf, Fec | + | [149] | |
| n | rc; Td-re | Se, Sf, Fec, Fer, E ● | + | [132] | ||
| oxymatrine | Phytoseiulus longipes | n | rc; Td-re | Sf, Fec, Fer, E, IOBC | - | [145] |
| rc; Tre (4-31) | + Tre (10) | |||||
| pyrethrum | Amblyseius andersoni | n | rc, Td-re | Se, Sf, Fec, E | - | [160] |
| Neoseiulus californicus | n | rc; Td-re | Se, Sf, Fec, Fer, E ● | - | [130] | |
| Phytoseiulus persimilis | n | rc; Td-re | Se, Sf, Fec, Fer, E ● | - | [132] | |
| rosemary oil | Phytoseiulus persimilis | c | rc, rc+/-; Td-re, Tre (0) | Sf, LC50 ● | + | [107] |
| soybean oil | n | rc; Td-re | Se, Sf ● | + | [135] | |
| + fatty acids | ||||||
| + caraway oil | ||||||
c = commercial strains, n = native populations; rc = recommended concentrations; rc+/- concentrations higher/lower than rc; Td-re = direct (topical) treatment and residual exposure; Tre (0) = exposure to fresh residues; Tre (x-y) = exposure to residues aged x-y days; Td = direct (topical) treatment and transfer to untreated surface; Tp = consumption of treated prey; Se = egg hatching; Sel-a = egg hatching and larval survival to the adulthood; Sl = larval survival; Sl-a = larval survival to the adulthood; Sf = adult female survival; Fec = fecundity (eggs produced by treated females); Fer = fertility (hatched eggs produced by treated females); E = the coefficient of toxicity [129]; IOBC = classification according the IOBC-WPRS guidelines [128]; ● comparative toxicity predator – prey .
Investigation of the comparative toxicity to predators and their prey has an important place in this evaluation of the selectivity of bioacaricides. This type of investigation is the basis for assessing physiological selectivity [123]. The lower comparative toxicity to phytoseiids, however, does not itself ensure compatibility. If the bioacaricide significantly reduces the prey population, the lack of food can cause decline and disappearance of the predator population and followed by an outbreak of the prey population [163]. Also, the consequences of approximately equal harmfulness of the recommended concentrations for predator and prey should be interpreted depending on the level of harmful effect of a bioacaricide. Bergeron & Schmidt-Jeffris [138], for example, calculated abamectin selectivity indices for N. fallacis, N. californicus and P. persimilis to be zero. These values resulted from approximately equal but high harmfulness (96 - 100%) for both predators and their prey (T. urticae), so this bioacaricide is not rated as compatible. The predator-prey comparison using LC50 as the endpoint requires additional interpretation of the results, because significantly lower toxicity for the predator compared to the prey has practical significance if the LC50 for the predator is higher than the recommended concentration [152]. When investigating physiological selectivity, the phenomenon of acaricide resistance should be taken into account as well [123]. Regarding bioacaricides, the only example is resistance to abamectin and milbemectin, widespread in T. urticae populations [164] and also observed in other tetranychids [165,166].
Evaluations using demographic bioassays (Table 7) showed that the recommended concentrations of several tested bioacaricides reduce population growth of phytoseiid mites. Lower concentrations can also cause reduction, as shown by the treatment of Phytoseius plumifer (Canestrini & Fanzago) with abamectin applied at LC20 against adult females [167]. On the other hand, the finding that low concentrations do not cause reduction [168,169] should be interpreted bearing in mind the recommended concentrations. Comparative demographic studies would allow a deeper insight, but there have been no examples of such research with bioacaricides.
Table 7.
Examples of evaluation of the compatibility of bioacaricides and phytoseiid mites in demographic bioassays (rm = the intrinsic rate of increase: ↓ = reduction, ns = non significant effect).
Table 7.
Examples of evaluation of the compatibility of bioacaricides and phytoseiid mites in demographic bioassays (rm = the intrinsic rate of increase: ↓ = reduction, ns = non significant effect).
| Bioacaricides | Phytoseiid mites | Methodology | Effect on rm | References | |
| Beauveria bassiana GHA | Phytoseiulus persimilis | c | rc, ATlt, F0 | ↓ | [170] |
| Hirsutella thompsonii | Phytoseiulus longipes | n | rc, ATlt, F0 | ↓ | [145] |
| abamectin | Phytoseiulus longipes | n | rc, ATlt, F0 | ↓ | [145] |
| Neoseiulus baraki | n | rc, Flt, F0 F1 | ↓ F0 | [171] | |
| Phytoseius plumifer | n | LC10, 20, Flt, F1 | ↓ LC20 | [167] | |
| milbemectin | Amblyseius swirskii | n | LC5, 15, 25, ATlt, F1 | ns | [169] |
| azadirachtin | Neoseiulus baraki | n | rc, Flt, F0 F1 | ns | [171] |
| Phytoseiulus longipes | n | rc, ATlt, F0 | ↓ | [145] | |
| geraniol + citronellol | Neoseiulus californicus | c | LC10, 20, ATlt, F1 | ns | [168] |
| + nerolidol + farnesol | |||||
| oxymatrine | Phytoseiulus longipes | n | rc, ATlt, F0 | ↓ | [145] |
c = commercial strains, n = native populations; rc = recommended concentrations; ATlt = age-stage two-sex life tables; Flt = fertility life tables; F0 = treatment and assessment in F0 generation; F1 = treatment in F0 generation, assessment in F1 generation; F0 F1 = treatment in F0 generation, assessment in F0 and F1 generations.
An alternative and less labour- and time-consuming approach for assessing effects at the population level is based on the calculation of the instantaneous rate rate of increase (ri) from the number of live individuals at the beginning and the end of bioassay [172]. Using this approach Tsolakis & Ragusa [135] compared the effects of a commercial mixture of vegetable and essential oils and potassium salts of fatty acids on population growth of T. urticae and P. persimilis; the biopesticide significantly reduced the ri values of the former but not of the latter species. Lima et al. [173] observed significant reduction of the ri values following treatment of Neoseiulus baraki (Athias-Henriot) with abamectin, while Silva et al. [149] found no reduction in Neoseiulus barkeri Hughes treated with abamectin and azadirachtin.
The aforementioned examples show that life history traits have been in the focus of evaluation of sublethal effects of bioacaricides on phytoseiids. Few studies have been focused on evaluation of behavioral effects. Lima et al. [174] investigated repellence (avoiding acaricide without making direct contact with its residues) and irritancy (moving away from the treated area after making the contact) of azadirachtin for N. baraki and found significant effects. The authors emphasized that these effects on walking behavior potentially reduce the exposure of predators, but also can cause their dispersal and decrease the efficiency of biological control. On the other hand, Bostanian et al. [136] and Beers & Schmidt-Jeffris [175] observed low level of repellency of spinosad on G. occidentalis. Bioacaricides can also affect feeding and reproduction behavior of phytoseiid mites. Investigations of the effects on N. baraki revealed that azadirachtin impaired copulation [176] and abamectin lowered the attack rate [177], while both bioacaricides impaired prey location [178]. These effects hinder predator-prey interaction and can compromise biological control.
The largest amount of data on the effects of acaricides on phytoseiids comes from laboratory bioassays. Therefore, the extrapolation of the results from laboratory to field is of fundamentally important, especially for the IOBC-WPRS programme for sequential testing. Translating laboratory results to field conditions has been a major challenge considering that laboratory bioassays most often do not take into account complexity and variability of environmental factors, heterogeneity of spray coverage in time and space, indirect effects through food supply, population dynamics [123,179]. In addition to these general limitations, the IOBC-WPRS testing has been critically discussed regarding the realism of the “worst case” scenario, and a rigid implementation of the trigger values for toxicity classification, which may affect accuracy of predictions drawn from the laboratory bioassay alone [180,181,182]. Methodological issues aside, the usefulness of the IOBC database is limited by the need to evaluate compatibility of pesticides (acaricides) with predatory species and strains important in local environments [123,179,182].
Investigating compatibility in greenhouse and/or field trials is necessary, not only because of the problems that arise when extrapolating results from the laboratory. The compatibility of acaricides with phytoseiids should be tested and proven under realistic conditions of the complex action of various factors. Large-scale field trials enable an assessment of the long-term impact of operational factors (timing, procedures and amounts of acaricide application, predator augmentation, habitat management measures) and biological-ecological factors (trophic relationships and interactions, migrations, host plant features, refugia) on the dynamics of predator and prey populations and the effectiveness of biological control under variable environmental conditions. On the other hand, the complexity and variability of factors in the field can make it difficult to interpret the results and draw clear conclusions [123,180,182].
There have been not many examples of studies evaluating the compatibility of bioacaricides and phytoseiids in greenhouse and/or field trials. In some of these studies, the dynamics of both predator and prey populations was monitored in order to explore possibility of combining predator activity and bioacaricide application. Greenhouse trials showed that mycopesticides could be successfully applied against T. urticae in combination with P. persimilis release on tomato plants [75] and N. californicus release on rose plants [183], as well as against mixed infestation of chrysanthemums with T. urticae and the western flower thrips, Frankliniella occidentalis (Pergande), in combination with A. swirskii and N. cucumeris [184]. Also, spinosad application at rates recommended for thrips control was compatible with P. persimilis release against T. urticae on ivy geranium [185]. On the other hand, application of spinosad in an apple orchard was detrimental to Kampimodromus aberrans (Oudemans), a predator of P. ulmi [157].
In other studies only the predator population was monitored. Jacas Miret and Garcia-Mari [186] rated abamectin as moderately harmful and azadirachtin as harmless to Euseius stipulatus (Athias-Henriot) following treatments in a citrus orchard. Castagnoli et al. [160] conducted field trials in an apple orchard in which azadirachtin and pyrethrum were applied twice on a bi-weekly basis. Azadirachtin did not affect population of A. andersoni; pyrethrum significantly reduced the population density after the second treatment, but the population recovered in a few days. Miles and Dutton [187] investigated the effects of single and repeated applications of spinosad on phytoseiid populations in vines and apples. Maximum reduction of 43% and 21% was found in T. pyri and K. aberrans populations in vines, respectively, and 41% in A. andersoni population in apples, after single treatment of spinosad. Repeated application caused maximum reduction of 75% in A. andersoni population in apples, and this effect was rated as harmful. On the other hand, de Andrade et al. [115] found no significant population reduction in Iphiseoides zuluagai Denmark & Muma (a predator of the citrus leprosis mite, Brevipalpus yothersi Baker) treated with oxymatrine in a commercial citrus grove.
Considering the advantages and limitations of different types of bioassays, a complementary approach to the evaluation of selectivity [123,160,180], which integrates laboratory and field data, is needed as a sustainable solution for exploiting physiological and/or ensuring ecological selectivity. This integrative approach implies more further research in the IPM context, on an expanded range of evaluated bioacaricides and phytoseiid species and strains, both commercialized and native.
5. Bioacaricides – Potential and Perspectives
According to a long-term estimate [11] biopesticides will equalize with synthetic pesticides in terms of market size by the early 2050s. Whether biopesticide sector will meet the great expectations, depends on its response to various growth challenges. The most important challenge to the increase of use of biopesticides in crop protection is the need for further improvements in their development, mass production, formulation and application [8,30].
Increasing the share of biopesticides on the global market can be achieved by the development of novel products, as well as the improvement of products that are already on the market. As far as bioacaricides are concerned, new commercial products of microbial origin could be found among new strains and isolates of Beauveria, Metarhizium and other entomopathogenic fungi [188,189,190,191]. Candidates for new bioacaricides could be also based on bacterial species that are symbiotically associated with entomopathogenic nematodes (EPN). Recent evaluation of cell suspensions (containing viable bacterial cells and toxins) and cell-free supernatants (containing only toxins) showed high acaropathogenic activity of several EPN-symbiotic species from the genera Photorhabdus and Xenorhabdus against spider mites [192,193,194]. The results of ongoing research have indicated new botanical sources for terpenoids/essential oils as potential bioacaricides [195,196,197]. Promising new ingredients for botanical bioacaricide products have also been found, such as coumarin scopoletin that can be isolated from many plants [198], acetogenins from Rollinia mucosa (Jacq.) Baill (Annonaceae) [199], humilinolids from Swietenia humilis Zucc. (Lamiaceae) [200] and carlina oxide from Carlina acaulis L. (Asteraceae) [201].
Mass production of biopesticides depends on the availability and quality of biological resources. This issue is particularly important for botanicals, given that plant biomass has been a limitation to the commercialization of many potentially useful biopesticide agents. The shortage of biomass can be mitigated by cultivation of plants containing bioactive compounds, as well as by utilization of agro-industrial byproducts and waste. A good example of the latter is orange oil, obtained by cold pressing of discarded citrus skin [40,94]. Inconsistent field efficacy of mycopesticides, which is a consequence of sensitivity of entomopathogenic fungi to abiotic stresses and naturally low virulence, can be overcome by genetic engineering [202,203].
Development of optimal formulations is required in order to ensure stability and increase efficacy of biopesticides. A recent approach to solving this issue has been through micro- and nanoencapsulation of biopesticides using various matrices and carrier systems [204,205,206]. For example, Ebadollahi et al. [207] showed that loading of Thymus eriocalyx (Ronniger) Jalas and Thymus kotschyanus Boiss. & Hohen essential oils in a mesoporous material MCM-41 increased persistence and toxicity of these oils to T. urticae, while Ahmadi et al. [208] found that encapsulation of Satureja hortensis L. essential oil using chitosan nanoparticles improved its toxicity to this pest. Microencapsulated formulations increased effectiveness of B. bassiana against T. urticae and even low conidial concentrations of the fungus resulted in high mortality rates [209]
In addition to these technological challenges, regulation of biopesticides and end-users’ perception about biopesticide effectiveness have been often emphasized as important impediments to their commercialization [8,40]. In order to overcome shortcomings in biopesticide regulation it is necessary to accelerate and simplified processes involved in product registration. There is also a need for global harmonization of biopesticide regulation [40,210,211]. More education, training and demonstrations are required to raise awareness among end-users about effectiveness and benefits of biopesticides requires [8].
There is no doubt that further technological advances, legislation expediting registration and increased adoption, as well as wider implementation of IPM and expansion of organic agriculture, will contribute to future growth of the biopesticide sector. However, the key driver of biopesticide success in the global market is and will be profit [28,62]. An agrochemical company – large ones have recently moved into the biopesticide sector [8] – will produce biopesticides only if there is a profit to be gained. The end-users (growers and farmers) will choose biopesticides only if they feel confident that the choice does not compromise their earnings.
Author Contributions
Conceptualization, D.M. and I.D.; Literature search, data analysis and interpretation, D.M., I.D., H.T.; writing—original draft preparation, D.M., I.D. and H.T.; writing—review and editing, D.M., I.D. and H.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia (Contract No. 451-03-66/2024-03/ 200214), Cukurova University Scientifc Research Foundation Units (CU-BAP), grant number: FBA-2022-14342, and the European Union – Next Generation EU - PNRR M4 - C2 -Investimento 1.1: Fondo per il Programma Nazionale di Ricerca e Progetti di Rilevante Interesse Nazionale (PRIN) - PRIN 2022 cod. 202274BK9L_001 dal titolo "Bioformulations for controlled release of botanical pesticides for sustainable agriculture" CUP B53D23008570006.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31-43. [CrossRef]
- Cooper, J.; Dobson, H. The benefits of pesticides to mankind and the environment. Crop Prot. 2007, 26, 1337–1348. [CrossRef]
- Popp, J.; Pető, K.; Nagy, J. Pesticide productivity and food security. A review. Agron. Sustain. Dev. 2013, 33, 243–255. [CrossRef]
- Pimentel, D.; Acquay, H.; Biltonen, M.; Rice, P.; Silva, M.; Nelson, J.; Lipner, V.; Giordano, S.; Horowitz, A.; D'Amore, M. Environmental and economic costs of pesticide use. BioScience 1992, 42, 750-760. [CrossRef]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48-60. [CrossRef]
- Copping, L.G;. Menn, J.J. Biopesticides: a review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651-676. [CrossRef]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524-554. [CrossRef]
- Marrone, P.G. Pesticidal natural products – status and future potential. Pest Manag. Sci. 2019, 75, 2325-2340. [CrossRef]
- Flint, M.L.; van den Bosch, R. Introduction to Integrated Pest Management; Plenum Press: New York and London, 1981; 240 pp.
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects - a review. Plant Prot. Sci. 2016, 52, 229-241. [CrossRef]
- Olson, S. An analysis of the biopesticide market now and where it is going. Outlooks Pest Manag. 2015, 26, 203-206. [CrossRef]
- Marčić, D. Acaricides in modern management of plant-feeding mites. J. Pest Sci. 2012, 85, 395-408. [CrossRef]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem.Physiol. 2015, 121, 12-21. [CrossRef]
- Umetsu, N.; Shirai, Y. Development of novel pesticides in the 21st century. J. Pestic. Sci. 2020, 45, 54-74. [CrossRef]
- IRAC (Insecticide Resistance Action Committee) – The IRAC Mode of action classification online. Available online: http://www.irac-online.org/modes-of-action/ (accessed on 31/05/ 2024).
- OECD (Organisation for Economic Co-operation and Development) Series on pesticides No. 12: Guidance for registration requirements for pheromones and other semiochemicals used for arthropod pest control; OECD - Environment Directorate: Paris, France, 2001; 25 pp.
- OECD (Organisation for Economic Co-operation and Development) Series on pesticides No. 18: Guidance for registration requirements for microbial pesticides; OECD - Environment Directorate: Paris, France,. 2003; 51 pp.
- OECD (Organisation for Economic Co-operation and Development) Series on pesticides No. 21: Guidance for information requirements for regulation of invertebrates as biological control agents (IBCAs); OECD - Environment Directorate: Paris, France, 2004; 22 pp.
- OECD (Organisation for Economic Co-operation and Development) Series on pesticides No. 90: Guidance document on botanical active substances used in plant protection products; OECD - Environment Directorate: Paris, France, 2017; 38 pp.
- FAO (Food and Agriculture Organization of the United Nations); WHO (World Health Organization) International code of conduct on pesticide management: Guidelines for the registration of miucrobial, botanical and semiochemical pest control agents for plant protection and public health issues. FAO & WHO: Rome, Italy, 2017; 76 pp.
- Marrone, P.G. Biopesticide commercialization in North America: state of the art and future opportunities. In Development and commercialization of biopesticides - costs and benefits; Koul, O., Ed.; Academic Press: London, UK, 2023; pp. 173 – 202. [CrossRef]
- US EPA (United States Environmental Protection Agency) - Biopesticides. Available online https://www.epa.gov/pesticides/biopesticides (accessed on 28/02/2024).
- Wang, Q;.Wang, Z. Biopesticides in China: development, commercialization, and regulation. In Development and commercialization of biopesticides - costs and benefits; Koul, O., Ed.; Academic Press: London, UK, 2023; pp. 203-212. [CrossRef]
- EC (European Commission) Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 concerning the placing of plant protection products on the market and repealing Council Directives 79/117/EEC and 91/414/EEC. Official Journal of the European Union, L 309/1.
- EC (European Commission) EU Pesticide Database – Active substances, safeners and synergists. Available online https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/start/screen/active-substances (accessed on 28/02/2024).
- Karamaouna, F.; Economou, L.P.; Lykogianni, M.; Mantzoukas, S.;. Eliopoulos, P.A. Biopesticides in the EU: state of play and perspectives after the Green Deal for agriculture. In Development and commercialization of biopesticides - costs and benefits; Koul, O., Ed.; Academic Press: London, UK, 2023; pp. 213-239. [CrossRef]
- Bailey, A.; Chandler, D.; Grant, W.P.; Greaves, J.; Prince, G.; Tatchell, M. Biopesticides: Pest Management and Regulation. CAB International : Wallingford, UK, 2010; 232 pp.
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B, 2011, 366, 1987-1998. [CrossRef]
- Marrone, P.G. Status of the biopesticide market and prospects for new bioherbicides. Pest Manag. Sci. 2024, 80, 81-86. [CrossRef]
- Koul, O. Biopesticides: commercial opportunities and challenges. In Development and commercialization of biopesticides - costs and benefits; Koul, O., Ed.; Academic Press: London, UK, 2023; pp. 1-23. [CrossRef]
- Biondi, A.; Mommaerts, V.; Smagghe, G.; Viuela, L.; Zappala, L.; Desneux, N. The non-target impact of spinosyns on beneficial arthropods. Pest Manag. Sci. 2012, 68, 1523-1536. [CrossRef]
- Cappa, F.; Baracchi, D.; Cervo, R. Biopesticides and insect pollinators: Detrimental effects, outdated guidelines, and future directions. Sci. Total Environ. 2022, 837, 155714. [CrossRef]
- Giunti, G.; Benelli, G.; Palmeri, V.; Laudani, F.; Ricupero, M.; Ricciardi, R.; Maggi, F.; Lucchi, A.; Guedes, R.N.C.; Desneux, N.; Campolo, O. Non-target effects of essential oil-based biopesticides for crop protection: Impact on natural enemies, pollinators, and soil invertebrates. Biol. Control, 2022, 176, 105071. [CrossRef]
- Toledo-Hernández, E.; Peňa-Chora, G.; Hernández-Velázquez, V.M.; Lormendez, C.C.; Toribio-Jimenéz, J.; Romero-Ramírez, Y.; León-Rodríguez, R. The stingless bees (Hymenoptera: Apidae: Meliponini): a review of the current threats to their survival. Apidologie, 2022, 53, 8. [CrossRef]
- Brimner, T.A.; Boland, G.J. A review of the non-target effects of fungi used to biologically control plant diseases. Agric. Ecosyst. Environ. 2003, 100, 3–16. [CrossRef]
- Mota-Sanchez, D.; Wise, J.C. The Arthropod Pesticide Resistance Database. Available online www.pesticideresistance.org (accessed on 31/05/2024).
- Sundin, G.W.; Wang, N. Antibiotic resistance in plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [CrossRef]
- Siegwart, M.; Graillot, B.; Lopez, C.B.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bio-insecticides or how to enhance their sustainability: a review. Front. Plant Sci. 2015, 6, 381. [CrossRef]
- Mangan, R., Bussière, L.F., Polanczyk, R.A. & Tinsley, M.C. Increasing ecological heterogeneity can constrain biopesticide resistance evolution. Trends Ecol. Evol. 2023, 38, 605-614. [CrossRef]
- Isman, M.B. Botanical insecticides in the twenty-first century: fulfilling the promise? Annu. Rev. Entomol. 2020, 65, 233-249. [CrossRef]
- Flamini, G. Acaricides of natural origin, personal experiences and review of literature (1990-2001). Studies in Natural Products Chemistry 2003, 28, 381-451. [CrossRef]
- Maniania, N.K.; Bugeme, D.M.; Wekesa, V.W.; Delalibero Jr, I.; Knapp, M. Role of entomopathogenic fungi in the control of Tetranychus evansi and Tetranychus urticae (Acari: Tetranychidae), pests of horticultural crops. Exp. Appl. Acarol. 2008, 46, 259-274. [CrossRef]
- Attia, S.; Grissa, K.L.; Lognay, G.; Bitume, E.; Hance, T.; Mailleux, A.C. A review of the major biological approaches to control the worldwide pest Tetranychus urticae with special reference to natural pesticides. J. Pest Sci. 2013, 86, 361-386. [CrossRef]
- Rincón, R.A.; Rodriguez, D.; Coy-Barrera, E. Botanicals against Tetranychus urticae Koch under laboratory conditions: a survey of alternatives for controlling pest mites. Plants 2019, 8, 272. [CrossRef]
- Jakubowska, M.; Dobosz, R.; Zawada, D.; Kowalska, J. A review of crop protection methods against the twospotted spider mite - Tetranychus urticae Koch (Acari: Tetranychidae) - with special reference to alternative methods. Agriculture 2022, 12, 898. [CrossRef]
- Smagghe, F.; Spooner-Hart, R.; Chen, Z.H.; Donovan-Mak, M. Biological control of arthropod pests in protected cropping by employing entomopathogens: Efficiency, production and safety. Biol. Control 2023, 186, 105337. [CrossRef]
- Royalty, R.N.; Hall, F.R.; Taylor, R.A.J. Effects of thuringiensin on Tetranychus urticae (Acari: Tetranychidae) mortality, fecundity, and feeding. J. Econ. Entomol. 1990, 83, 792-798. [CrossRef]
- Royalty, R.N.; Hall, F.R.; Lucius, B.A. Effects of thuringiensin on European red mite (Acarina: Tetranychidae) mortality, oviposition rate and feeding. Pestic. Sci. 1991, 33, 383-391. [CrossRef]
- Vargas, R.; Chapman, B.; Penman, D.R. Toxicity of thurigiensin on immature and adult stages of Tetranychus urticae Koch and Panonychus ulmi (Koch) (Acarina: Tetranychidae). Agric. Téc. 2001, 61, 3-14. [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: an overview of their biocidal activity. Toxins 2014, 6, 3296-3325. [CrossRef]
- Zenkova, A.A.; Grizanova, E.V.; Andreeva, I.V.; Gerne D.Y.; Shatalova, E.I.; Cvetcova, V.P.; Dubovskiy, I.M. Effect of fungus Lecanicillium lecanii and bacteria Bacillus thuringiensis, Streptomyces avermitilis on two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae) and predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). J. Plant Prot. Res. 2020, 60, 415-419. [CrossRef]
- Belousova, M.E.; Malovichko, Y.V.; Shikov, A.E.; Nizhnikov, A.A.; Antonets, K.S. Dissecting the environmental consequences of Bacillus thuringiensis application for natural ecosystems. Toxins 2021, 13, 355. [CrossRef]
- Martin, P.A.W.; Gundersen-Rindal, D.; Blackburn, M.; Buyer, J. Chromobacterium subtsugae sp. nov., a betaproteobacterium toxic to Colorado potato beetle and other insect pests. Int. J. Syst. Evol. Microbiol. 2007, 57, 993–999. [CrossRef]
- Cordova-Kreylos, A.L.; Fernandez, L.E;, Koivunen, M.; Yang, A.; Flor-Weiler, L.; Marrone, P.G. Isolation and characterization of Burkholderia rinojensis sp. nov., a non-Burkholderia cepacia complex soil bacterium with insecticidal and miticidal activities. Appl. Environ. Microbiol. 2013, 79, 7669 –7678. [CrossRef]
- Ray, H.A.; Hoy, M.A. Effects of reduced-risk insecticides on three orchid pests and two predacious natural enemies. Fla Entomol. 2014, 97, 972-978. [CrossRef]
- Mertoglu, G. Evaluation of biorational insecticides against two-spotted spider mite (Acari: Tetranychidae) on tomato. Master thesis, Auburn University, Alabama, 12/12/2019.
- Golec, J.R.; Hoge, B.; Walgenbach, J.F. Effects of biopesticides on different Tetranychus urticae (Acari: Tetranychidae) life stages. Crop Prot. 2020, 128, 105006. [CrossRef]
- Stansly, P.A.; Kostyk, B.C. Control of broad mite on “bell” pepper with labeled and experimental insecticides. Arthropod Management Tests, 2014, 39, E67. [CrossRef]
- Stansly, P.A.; Kostyk, B.C. Acaricidal control of citrus rust mites, 2013. Arthropod Management Tests, 2014, 39, D17. [CrossRef]
- Dara, S.K.; Peck, D.; Murray, D. Chemical and non-chemical options for managing twospotted spider mite, western tarnished plant bug and other arthropod pests in strawberries. Insects, 2018, 9, 156. [CrossRef]
- Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: a comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control, 2007, 43, 237–256. [CrossRef]
- Marčić, D. Biopesticides for insect and mite pest management in modern crop protection. In Understanding Pests and Their Control Agents as the Basis for Integrated Plant Protection. Tanović, B., Nicot. P., Dolzhenko, V., Marčić, D. Eds.; IOBC-WPRS: Darmstadt, Germany,. 2021, pp. 77-88.
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [CrossRef]
- Jiang, Y.; Wang, J. The registration situation and use of mycopesticides in the world. J. Fungi 2023, 9, 940. [CrossRef]
- Ullah, M.S.; Lim, U.T. Laboratory bioassay of Beauveria bassiana against Tetranychus urticae (Acari: Tetranychidae) on leaf discs and potted bean plants. Exp. Appl. Acarol. 2015, 65, 307-318. [CrossRef]
- Basak, R.; Akter, M.; Tumpa, T.A.; Sharmin, D.; Ullah, M.S. Laboratory bioassay of six pesticides, an entomopathogenic fungus, and a botanical pesticide on two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Persian J. Acarol. 2021, 10, 269-280.
- Nahian-Al Sabri, Md;, Jahan, M.; Tetsuo, G.; Döker, I.; Ullah, M.S. Effect of relative humidity on the efficacy of entomopathogen Beauveria bassiana-based mycopesticide against red spider mite Tetranychus macfarlanei. Syst. Appl. Acarol. 2022, 27, 2414-2425. [CrossRef]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial biopesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74-81. [CrossRef]
- US EPA (United States Environmental Protection Agency) - Biopesticide Active Ingredients. Available online https://www.epa.gov/ingredients-used-pesticide-products/biopesticide-active-ingredients (accessed on 28/02/2024).
- Arthurs, S.; Dara, S. Microbial biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2019, 165, 13-21. [CrossRef]
- Brownbridge, M.; Buitenhuis, R. Integration of microbial biopesticides in greenhouse floriculture: The Canadian experience. J. Invertebr. Pathol. 2019, 165, 4-12. [CrossRef]
- Hatting, J.L.; Moore, S.D.; Malan, A.P. Microbial control of phytophagous invertebrate pests in South Africa: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 54-66. [CrossRef]
- Mascarin, G.M.; Lopes, R.B.; Delalibera Jr, Í.; Fernandes, É.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46-53. [CrossRef]
- US EPA (United States Environmental Protection Agency) - Pesticide Product and Label System. Available online https://ordspub.epa.gov/ords/pesticides/f?p=PPLS:1 (accessed on 28/02/2024).
- Chandler, D.; Davidson, G.; Jacobson R.J. Laboratory and glasshouse evaluation of entomopathogenic fungi against the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae), on tomato, Lycopersicon esculentum. Biocontrol Sci. Technol. 2005, 15, 37–54. [CrossRef]
- Sreerama Kumar, P.S. Hirsutella thompsonii as a mycoacaricide for Aceria guerreronis on coconut in India: research, development, and other aspects. In Trends in Acarology; Sabelis, M.W., Bruin, J. Eds.; Springer: Dordrecht, The Netherlands, 2010, pp. 441–444. [CrossRef]
- Gatarayiha, M.C.; Laing, M.D.; Miller, R.M. Field evaluation of Beauveria bassiana efficacy for the control of Tetranychus urticae Koch (Acari: Tetranychidae). J. Appl. Entomol. 2011, 135, 582-592. [CrossRef]
- Marčić, D.; Prijović, M.; Drobnjaković, T.; Međo, I.; Perić, P.; Milenković, S. Greenhouse and field evaluation of two biopesticides against Tetranychus urticae and Panonychus ulmi (Acari: Tetranychidae). Pestic. Phytomed. 2012, 27, 313-320. [CrossRef]
- Marčić, D.; Perić, P.; Petronijević, S.; Prijović, M.; Drobnjaković, T.; Milenković, S. Efficacy evaluation of Beauveria bassiana strain ATCC 74040 (Naturalis) against spider mites (Acari: Tetranychidae) in Serbia. IOBC-WPRS Bull. 2013, 93, 65-71.
- Wraight, S.P.; Lopes, R.B.; Faria, M. Microbial control of mite and insect pests of greenhouse crops. In Microbial control of insect and mite pests; Lacey, L.A. Ed.; Academic Press: London, UK, 2017, pp. 237-252.
- Chouikhi, S.; Assadi, B.H.; Grissa Lebdi, K.; Belkadhi, M.S. Efficacy of the entomopathogenic fungus, Beauveria bassiana and Lecanicillium muscarium against two main pests, Bemisia tabaci (Genn.) and Tetranychus urticae Koch, under geothermal greenhouses of Southern Tunisia. Egypt. J. Biol. Pest Control, 2022, 32, 125. [CrossRef]
- Ullah, M.S.; Lim, U.T. Synergism of Beauveria bassiana and Phytoseiulus persimilis in control of Tetranychus urticae on bean plants. Syst. Appl. Acarol. 2017, 22, 1924-1935. [CrossRef]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Cordova, D.; Early F. et al. Insecticides, biologics and nematicides: updates to IRAC's mode of action classfication – a tool for resistance management. Pestic. Biochem. Physiol. 2020, 167, 104587. [CrossRef]
- Collett, M.G.; Oakes, K.A.; Babu, R.; McLennan, A. Evaluation of adjuvants for abamectin in early-season control of two-spotted mite and European red mite in apples. In Spray Oils Beyond 2000: Sustainable Pest and Disease Management; Beattie, A. et al. Eds.; University of Western Sydney: Sydney,Australia, 2002, pp. 285-288.
- Marčić, D.; Perić, P.; Prijović, M.; Ogurlić, I. Field and greenhouse evaluation of rapeseed spray oil against spider mites, green peach aphid and pear psylla in Serbia. Bull. Insectol. 2009, 62, 159-167.
- Van Leeuwen, T.; Dermauw, W.; Van De Veire, M.; Tirry, L. Systemic use of spinosad to control the two-spotted spider mite (Acari: Tetranychidae) on tomatoes grown in rockwool. Exp. Appl. Acarol. 2005, 37, 93–105. [CrossRef]
- Ismail, M.S.M.; Soliman, M.F.M.; El Naggar, M.H.; Ghallab, M.M. Acaricidal activity of spinosad and abamectin against two-spotted spider mites. Exp. Appl. Acarol. 2007, 43, 129-135. [CrossRef]
- Međo, I.; Stojnić, B.; Marčić, D. Acaricidal activity and sublethal effects of the microbial pesticide spinosad on Tetranychus urticae (Acari: Tetranychidae). Syst. Appl. Acarol. 2017, 22, 1748-1762. [CrossRef]
- Cowles, R.S. Effect of spinosad formulations and other miticides on twospotted spider mite. Arthropod Management Tests, 1998, 23, 342–343. [CrossRef]
- Tjosvold, SA.; Chaney, W.E. Evaluation of reduced risk and other biorational miticides on the control of spider mites (Tetranychus urticae). Acta Horticulturae 2001, 547, 93–96.
- OECD (Organisation for Economic Co-operation and Development) Series on pesticides No. 43: Working document on the evaluation of microbials for pest control. OECD - Environment Directorate: Paris, France, 64 pp.
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to Pyrethroids. Toxicol. Rev. 2005, 24, 93-106. [CrossRef]
- Chrustek, A.; Hołynska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A.; Olszewska-Słonina, D. Current research on the safety of pyrethroids used as insecticides. Medicina 2018, 54, 61.
- Isman, M.B. Commercialization and regulation of botanical biopesticides: a global perspective. In Development and commercialization of biopesticides - costs and benefits; Koul, O. Ed.; Academic Press: London, UK, 2023, pp. 25-36. [CrossRef]
- Campos, E.V.R;, Proença, P.L.F.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.C.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483-495. [CrossRef]
- Isman, M.B. Commercial development of plant essential oils and their constituents as active ingredients in bioinsecticides. Phytochem. Rev. 2020, 19, 235-241. [CrossRef]
- Juma, P.; Njau, N.; Wacera, F.W.; Micheni, C.M.; Khan, H.A.; Mitalo, O.W.; Odongo, D. Trends in neem (Azadirachta indica)-based botanical pesticides. In New and future development in biopesticide research: biotechnological exploration; De Mandal, S., Ramkumar, G., Karthi, S., Jin, F. Eds.; Springer Nature: Singapore,, 2022, pp. 137-156.
- Tsolakis, H.; Ragusa, E.; Ragusa, S. Effects of neem oil (Azadirachta indica A. Juss) on Tetranychus urticae Koch (Acariformes, Tetranychidae) in laboratory tests. In Acarid phylogeny and evolution - Adaptations in mites and ticks; Bernini, F., Nannetti, R., Nuzzaci C., de Lillo, E. Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002, pp. 351-362.
- Marčić, D.; Međo, I. Sublethal effects of azadirachtin-A (NeemAzal-T/S) on Tetranychus urticae (Acari: Tetranychidae). Syst. Appl. Acarol. 2015, 20, 25-38. [CrossRef]
- Kilani-Morakchi, S.; Morakchi-Goudji, H.; Sifi, K. Azadirachtin-based insecticide: overview, risk assessments, and future directions. Front. Agron. 2021, 3, 676208. [CrossRef]
- Savi, P.J.; Martins, M.B.; de Moraes, G.J.; Hountondji, F.C.C.; de Andrade, D.J. Bioactivity of oxymatrine and azadirachtin against Tetranychus evansi (Acari: Tetranychidae) and their compatibility with the predator Phytoseiulus longipes (Acari: Phytoseiidae) on tomato. Syst. Appl. Acarol. 2021, 26, 1264-1279. [CrossRef]
- Marčić, D.; Ogurlić, I.; Prijović, M.; Perić, P. Effectiveness of azadirachtin (NeemAzal-T/S) in controlling pear psylla (Cacopsylla pyri) and European red mite (Panonychus ulmi). Pestic. Phytomed. 2009, 24, 123-132. [CrossRef]
- Soto, A.; Venzon, M.; Oliveira, R.M.; Oliveira, H.G.; Pallini, A. Alternative control of Tetranychus evansi Baker & Pritchard (Acari: Tetranychidae) on tomato plants grown in greenhouses. Neotrop. Entomol. 2010, 39, 638–644. [CrossRef]
- Bagde, A.S.; Pashte, V.V. Efficacy of neem bio-pesticides against eggs of coconut eriophyid mite, (Aceria guerreronis Keifer). Adv. Life Sci. 2016, 5, 1436-1441.
- Prasad, R.; Hembrom, L; Prasad, D. Evaluation of acaricidal efficacy of some botanicals and conventional acaricides for management of yellow mite (Polyphagotarsonemus latus Banks) of chilli. J. Eco-friendly Agric. 2017, 12, 50-52.
- Regnault-Roger, C., Vincent, C. & Arnason, J.T. Essential oils in insect control: low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–425. [CrossRef]
- Miresmailli, S.; Isman, M.B. Efficacy and persistance of rosemary oil as an acaricide against twospotted spider mite (Acari: Tetranychidae) on greenhouse tomato. J. Econ. Entomol. 2006, 99, 2015-2023. [CrossRef]
- Cloyd, R.A;, Galle, C.L.; Keith, S.R.; Kalscheur, N.A.; Kemp, K.E. Effect of commercially available plant-derived essential oil products on arthropod pests. J. Econ. Entomol. 2009, 102, 1567–1579. [CrossRef]
- Haviland, D.; Hill S. The effects of miticide treatments on Pacific spider mite in Almond 2018. Arthropod Management Tests, 2019, 44, tsz067. [CrossRef]
- Szczepaniec, A.; Lathrop-Melting, A.; Janecek, T.; Nachappa, P.; Cranshaw, W.; Alnajjar, G.; Axtell, A. Suppression of hemp russet mite, Aculops cannabicola (Acari: Eriophyidae), in industrial hemp in greenhouse and field. Environ. Entomol. 2024, 53, 18–25. [CrossRef]
- Chiasson, H.; Bostanian, N.J.; Vincent, C. Acaricidal properties of a Chenopodium-based botanical. J. Econ. Entomol. 2004, 97, 1373-1377. [CrossRef]
- Chiasson, H;, Bostanian, N,J,; Vincent, C. Insecticidal properties of a Chenopodium-based botanical. J. Econ. Entomol. 2004, 97, 1378–1383. [CrossRef]
- Musa, A.; Međo, I.; Marić, I.; Marčić, D. Acaricidal and sublethal effects of a Chenopodium-based biopesticide on the twospotted spider mite (Acari: Tetranychidae). Exp. Appl. Acarol. 2017, 71, 211–226. [CrossRef]
- Marčić, D.; Međo, I. Acaricidal activity and sublethal effects of an oxymatrine-based biopesticide on two-spotted spider mite (Acari: Tetranychidae). Exp. Appl. Acarol. 2014, 64, 375-391. [CrossRef]
- de Andrade, D.J.; Ribeiro,E.B.; de Morais, M.R.; Zanardi, O.Z. Bioactivity of an oxymatrine-based commercial formulation against Brevipalpus yothersi Baker and its effects on predatory mites in citrus groves. Ecotoxicol. Environ. Saf. 2019, 176, 339-345. [CrossRef]
- de Andrade, D.J.; da Rocha, C.M.; de Matos, S.T.S.; Zanardi, O.Z. Oxymatrine-based bioacaricide as a management tool against Oligonychus ilicis (McGregor) (Acari: Tetranychidae) in coffee. Crop Prot. 2020, 134, 105182. [CrossRef]
- Rousseau, D. Chemical and physical properties of canola and rapeseed oil. In Rapeseed and canola oil production, processing, properties and uses; Gunstone, F.D. Ed.; Blackwell Publishing: Oxford, UK, 2004, pp. 79-105.
- Takeda, N.; Takata, A.; Arai, Y.; Sasaya, K;, Noyama, S.; Wakisaka, S.; Ghazy, N.A.; Voigt, D.; Suzuki, T. A vegetable oil–based biopesticide with ovicidal activity against the two-spotted spider mite, Tetranychus urticae Koch. Eng. Life Sci. 2020, 20, 525–534. [CrossRef]
- Villarreal, G.U.; Campos, E.V.R.; de Oliveira, J.L.; Fraceto, L.F. Development and commercialization of pheromone-based biopesticides: a global perspective. In Development and commercialization of biopesticides - costs and benefits; Koul O. Ed.; Academic Press: London, UK, 2023, pp. 37-56. [CrossRef]
- Carr, A.L.; Roe, M. Acarine attractants: Chemoreception, bioassay, chemistry and control. Pestic. Biochem. Physiol. 2016, 131, 60-79. [CrossRef]
- Torres, J.B.; Bueno, A.F. Conservation biological control using selective insecticides – a valuable tool for IPM. Biol. Control, 2018, 126, 53-64. [CrossRef]
- Knapp, M.; van Houten, I.; van Baal, E.; Groot, T. Use of predatory mites in commercial biocontrol: current status and future prospects. Acarologia 2018, 58, 72-82. [CrossRef]
- Duso, C.; Van Leeuwen, T.; Pozzebon, A. Improving the compatibility of pesticides and predatory mites: recent findings on physiological and ecological selectivity. Curr. Opin. Insect Sci. 2020, 39, 63-68. [CrossRef]
- Jansen, J.P. Beneficial arthropods and pesticides: building selectivity lists for IPM. IOBC-WPRS Bull. 2010, 55, 23-47.
- IOBC - Pesticide Side Effects Database. Available online https://iobc-wprs.org/ip-tools/pesticide-side-effect-database/ (Assessed on 15/01/2021).
- Samsøe-Petersen, L. Sequences of standard methods to test effects of chemicals on terrestrial arthropods. Ecotoxicol. Environ. Saf. 1990, 19, 310-319. [CrossRef]
- Bakker, F.M.; Grove, A.; Blümel, S.; Calis, J.; Oomen, P. Side-effect tests for phytoseiids and their rearing methods. IOBC-WPRS Bull. 1992, 15, 61-81.
- Sterk, G.; Hassan, S.A.; Baillod, M.; Bakker, F.; Bigler, F.; Blümel, S.; et al. Results of the seventh joint pesticide testing programme carried out by the IOBC/WPRS-Working Group „Pesticides and Beneficial Organisms“. BioControl 1999, 44, 99–117. [CrossRef]
- Overmeer, W.P.J.; van Zon, A.Q.; A standardized method for testing the side effects of pesticides on the predacious mite, Amblyseius potentillae (Acarina: Phytoseiidae). Entomophaga 1982, 27, 357-364. [CrossRef]
- Castagnoli, M.; Liguori, M.; Simoni, S.; Duso, C. Toxicity of some insecticides to Tetranychus urticae, Neoseiulus californicus and Tydeus californicus. BioControl, 2005, 50, 611–622. [CrossRef]
- Duso, C.; Malagnini, V.; Pozzebon, A.; Buzzetti, F.M.; Tirello, P. A method to assess the effects of pesticides on the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae) in the laboratory. Biocontrol Sci. Technol. 2008, 18, 1027-1040. [CrossRef]
- Duso, C.; Malagnini, V.; Pozzebon, A.; Castagnoli, M.; Liguori, M.; Simoni, S. Comparative toxicity of botanical and reduced-risk insecticides to Mediterranean populations of Tetranychus urticae and Phytoseiulus persimilis (Acari Tetranychidae, Phytoseiidae). Biol. Control, 2008, 47, 16-21. [CrossRef]
- Međo, I.; Marić, I.; Marčić, D.; Döker, I. Pesticides and phytoseiid mites: what the IOBC-WPRS Pesticide Side Effects Database can(not) tell us about selectivity? IOBC-WPRS Bull. 2024, 169, 45-47.
- Schmidt-Jeffris, R.; Beers, E.; Sater, C. Meta-analysis and review of pesticide non-target effects on phytoseiids, key biological control agents. Pest Manag. Sci. 2021, 77, 4848-4862. [CrossRef]
- Tsolakis, H.; Ragusa, S. Effects of a mixture of vegetable and essential oils and fatty acid potassium salts on Tetranychus urticae and Phytoseiulus persimilis. Ecotoxicol. Environ. Saf. 2008, 70, 276-282. [CrossRef]
- Bostanian, N.J.; Thistlewood, H.A.; Hardman, J.M.; Laurin, M.C.; Racette, G. Effects of seven new orchard pesticides on Galendromus occidentalis in laboratory studies. Pest Manag. Sci. 2009, 65, 635-639. [CrossRef]
- Bostanian, N.J.; Hardman, J.M.; Thistlewood, H.A.; Racette, G. Effects of six selected orchard insecticides on Neoseiulus fallacis (Acari: Phytoseiidae) in the laboratory. Pest Manag. Sci. 2010, 66, 1263-1267. [CrossRef]
- Bergeron, P.E.; Schmidt-Jeffris, R.A. Not all predators are equal: miticide non-target effects and differential selectivity. Pest Manag. Sci. 2020, 76, 2170-2179. [CrossRef]
- Beers, E.H.; Schmidt, R.A. Impact of orchard pesticides on Galendromus occidentalis: lethal and sublethal effects. Crop Prot. 2014, 56, 16-24. [CrossRef]
- Schmidt-Jeffris, R.; Beers, E. Potential impacts of orchard pesticides on Tetranychus urticae: A predator-prey perspective. Crop Prot. 2018, 103, 56-64. [CrossRef]
- Birch, L.C. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol. 1948, 17, 15-26. [CrossRef]
- Carey, J.R. Applied demography for biologists with special emphasis on insects. Oxford University Press: Oxford, UK, 1993.
- Chi, H.; You, M.; Atlihan, R.; Smith, C.L.; Kavousi, A.; Őzgőkçe, M.S. et al. Age-stage, two-sex life table: an introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103-124. [CrossRef]
- Midthassel, A.; Leather, S.R.; Wright, D.J.; Baxter, I.H. Compatibility of Amblyseius swirskii with Beauveria bassiana: two potentially complimentary biocontrol agents. BioControl 2016, 61, 437-447. [CrossRef]
- Savi, P.J.; de Moraes, G.J.; Hountondji, F.C.C.: Nansen C.; de Andrade, D.J. Compatibility of synthetic and biological pesticides with a biocontrol agent Phytoseiulus longipes (Acari: Phytoseiidae). Exp. Appl. Acarol. 2024, 93, 273–295. [CrossRef]
- Döker, I.; Kazak, C. Non-target effects of five acaricides on a native population of Amblyseius swirskii (Acari: Phytoseiidae). Int. J. Acarol. 2019, 45, 69-74. [CrossRef]
- Döker, I.; Kazak, C. Toxicity and risk assessment of acaricides on the predatory mite Euseius scutalis (Athias-Henriot) (Acari: Phytoseiidae) under laboratory conditions Chemosphere 2020, 261, 127760. [CrossRef]
- Sáenz-de-Cabezón Irigaray, F.J.; Zalom, F.G.; Thompson, P.B. Residual toxicity of acaricides to Galendromus occidentalis and Phytoseiulus persimilis reproductive potential. Biol. Control 2007, 40, 153-159. [CrossRef]
- Silva, D.L.; Lima, J.E.D.; Souza, P.V.L.; Melo, J.W.S.; Oliveira, J.E. de M.; Lima, D.B. Selectivity of acaricides to Neoseiulus barkeri (Hughes) (Acari: Phytoseiidae). J. Appl. Entomol. 2023, 147, 1014-1023. [CrossRef]
- Argolo, P.S.; Jacas, J.A.; Urbaneja, A. Comparative toxicity of pesticides in three phytoseiid mites with different life-style occuring in citrus: Euseius stipulatus, Neoseiulus californicus and Phytoseiulus persimilis. Exp. Appl Acarol. 2014, 62, 33-46. [CrossRef]
- Uddin, M.N.; Alam, M.Z.; Miah, M.R.U.; Mian, M.I.H.; Mustarin, K.E. Toxicity of pesticides to Tetranychus urticae Koch (Acari: Tetranychidae) and their side effects on Neoseiulus californicus. Int. J. Acarol. 2015, 41, 688-693. [CrossRef]
- Cheng, S.; Lin, R.; You, Y.; Lin, T.; Zeng, Z.; Yu, C. Comparative sensitivity of Neoseiulus cucumeris and its prey Tetranychus cinnbarinus after exposed to nineteen pesticides. Ecotoxicol. Environ. Saf. 2021, 217, 112234. [CrossRef]
- You, Y.; Lin, T.; Wei, H.; Zeng, Z.; Fu, J.; Liu, X.; Lin, R.; Zhang Y. Laboratory evaluation of the sublethal effects of four selective pesticides on the predatory mite Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae). Syst. Appl. Acarol. 2016, 21, 1506-1514. [CrossRef]
- Kim, S.S.; Seo, S.G. Relative toxicity of some acaricides to hte predatory mite, Amblyseius womersleyi, and the twospotted spider mite, Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae). Appl. Entomol. Zool. 2001, 36, 509-514. [CrossRef]
- Kim, S.S.; Yoo, S.S. Comparative toxicity of some acaricides to the predatory mite, Phytoseiulus persimilis and the twospotted spider mite, Tetranychus urticae. BioControl 2002, 47, 563-573. [CrossRef]
- Döker, I.; Yavaş, H.; Shirvani, Z.; Karaca, M.; Karut, K.; Marčić, D.; Kazak, C. Toxicity and risk assessment of pesticides on the predatory mite, Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) under laboratory conditions. Phytoparasitica, 2024, 52, 97. [CrossRef]
- Duso, C.; Ahmad, S.; Tirello, P.; Pozzebon, A.; Klaric, V.; Baldessari, M. The impact of insecticides applied in apple orchards on the predatory mite Kampimodromus aberrans (Acari: Phytoseiidae). Exp. Appl. Acarol. 2014, 62, 391-414. [CrossRef]
- Rahman, T.; Spafford, H.; Broughton, S. Compatibility of spinosad with predaceous mites (Acari) used to control Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Pest Manag. Sci. 2011, 67, 993-1003. [CrossRef]
- Villanueva, R.T.; Walgenbach, J.F. Development, oviposition, and mortality of Neoseiulus fallacis (Acari: Phytoseiidae) in response to reduced-risk insecticides. J. Econ. Entomol. 2005, 98, 2114-2120. [CrossRef]
- Castagnoli, M.; Angeli, G.; Liguori, M.; Forti, D.; Simoni, S. Side effects of botanical insecticides on predatory mite Amblyseius andersoni (Chant). J. Pest Sci. 2002, 75, 122-127. [CrossRef]
- Bernardi, D.; Botton, M.; da Cunha, U.S.; Bernardi, O.; Malausa, T.; Garcia, M.S.; Nava, D.E. Effects of azadirachtin on Tetranychus urticae (Acari: Tetranychidae) and its compatibility with predatory mites (Acari: Phytoseiidae) on strawberry. Pest Manag. Sci. 2013, 69, 75-80. [CrossRef]
- Spollen, K.M.; Isman, M.B. Acute and sublethal effects of a neem insecticide on the commercial biological control agents Phytoseiulus persimilis and Amblyseius cucumeris (Acari: Phytoseiidae) and Aphidoletes aphidimyza (Diptera: Cecidomyiidae). J. Econ. Entomol. 1996, 89, 1379-1386. [CrossRef]
- Ghazy, N.A.; Osakabe, M.; Negm, M.W.; Schausberger, P.; Gotoh, T.; Amano, H. Phytoseiid mites under environmental stress. Biol. Control 2016, 96, 120–134. [CrossRef]
- Xue, W.; Snoeck, S.; Njiru, C.; Inak, E.; Dermauw, W.; & Van Leeuwen, T. Geographical distribution and molecular insights into abamectin and milbemectin cross-resistance in European field populations of Tetranychus urticae. Pest Manag. Sci. 2020, 76, 2569-2581. [CrossRef]
- Döker, I.; Kazak, C.; Ay, R. Resistance status and detoxification enzyme activity in ten populations of Panonychus citri (Acari: Tetranychidae) from Turkey. Crop Prot. 2021, 141, 105488. [CrossRef]
- Khajehali, J.; Alavijeh, E.S.; Ghadamyari, M.; Marčić, D. Acaricide resistance in Panonychus citri and P. ulmi (Acari: Tetranychidae): Molecular mechanisms and management implications. Syst. Appl. Acarol. 2021, 26, 1526-1542. [CrossRef]
- Hamedi, N.; Fathipour, Y.; Saber, M. Sublethal effects of abamectin on the biological performance of the predatory mite Phytoseius plumifer (Acari: Phytoseiidae). Exp. Appl. Acarol. 2011, 53, 29-40. [CrossRef]
- Havasi, M.; Kheradmand, K.; Mosallanejad, H.; Fathipour,Y. Life history traits and demographic parameters of Neoseiulus californicus McGregor (Acari: Phytoseiidae) treated with the Biomite. Syst. Appl. Acarol. 2020, 25, 125-138. [CrossRef]
- Mousavi, A.; Kheradmand, K; Fathipour, Y.; Mosallanejad, H.; Havasi, M. Sublethal effects of milbemectin on biological parameters of Amblyseius swirskii (Acari: Phytoseiidae). Syst. Appl. Acarol. 2022, 27, 1085-1097. [CrossRef]
- Ullah, M.S.; Lim, U.T. Laboratory evaluation of the effect of Beauveria bassiana on the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). J. Invertebr. Pathol. 2017, 148, 102-109. [CrossRef]
- Lima, D.B.; Melo, J.W.S.; Gondim, M.G.C.Jr; Guedes, R.N.C.; Oliveira, J.E.M. Population-level effects of abamectin, azadirachtin and fenpyroximate on the predatory mite Neoseiulus baraki. Exp. Appl. Acarol. 2016, 70, 165-177. DOI; [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505-519. [CrossRef]
- Lima, D.B.; Monteiro, V.B.; Guedes, R.N.C.; Siqueira, H.A.A.; Pallini, A.; Gondim, M.G.C.Jr. Acaricide toxicity and synergism of fenpyroximate to the coconut mite predator Neoseiulus baraki. BioControl 2013, 58, 595-605. [CrossRef]
- Lima, D.B.; Melo, J.W.S.; Guedes, R.N.C.; Siqueira, H.A.A.; Pallini, A.; Gondim, M.G.C.Jr. Survival and behavioural response to acaricides of the coconut mite predator Neoseiulus baraki. Exp. Appl. Acarol. 2013, 60, 381-393. DOI; [CrossRef]
- Beers E.H.; Schmidt-Jeffris R.A. Effects of orchard pesticides on Galendromus occidentalis (Acari: Phytoseiidae): repellency and irritancy. J. Econ. Entomol. 2015, 108, 259-265. [CrossRef]
- Lima, D.B.; Melo, J.W.S.; Gondim, M.G.C.Jr; Guedes, R.N.C.; Oliveira, J.E.M.; Pallini A. Acaricide-impaired functional predation response of the phytoseiid mite Neoseiulus baraki to the coconut mite Aceria guerreronis. Ecotoxicology 2015, 24, 1124-1130. [CrossRef]
- Lima, D.B.; Melo, J.W.S.; Guedes, N.M.P.; Gontijo, L.M.; Guedes, R.N.C.; Gondim, M.G.C.Jr. Bioinsecticide-predator interactions: azadirachtin behavioral and reproductive impairment of the coconut mite predator Neoseiulus baraki. PLoS ONE, 2015, 10, e0118343. [CrossRef]
- Lima, D.B.; Oliveira, H.K.V.; Melo, J.W.S.; Gondim, M.G.C.Jr; Guedes, R.N.C.; Pallini, A. et al. Acaricides impair prey location in a predatory phytoseiid mite. J. Appl. Entomol. 2017, 141, 141-149. [CrossRef]
- Schmidt-Jeffris, R. Nontarget pesticide impacts on pest natural enemies: progress and gaps in currrent knowledge. Curr. Opin. Insect Sci. 2023, 58, 101056. [CrossRef]
- Sterk, G.; Creemers, P.; Merckx, K. Testing the side effects of pesticides on the predatory mite Typhlodromus pyri (Acari: Phytoseiidae) in field trials. IOBC/WPRS Bull. 1994, 17, 27-40.
- Bakker, F.M.; Jacas, J.A. Pesticides and phytoseiid mites: strategies for risk assessment. Ecotoxicol. Environ. Saf. 1995, 32, 58-67. [CrossRef]
- Thomson L.J.; Hoffmann A.A. Ecologically-sustainable chemical recommendations for agricultural pest control? J. Econ. Entomol. 2007, 100, 1741-1750. [CrossRef]
- Numa Vergel, S.J.; Bustos, R.A.; Rodríguez, C.D.; Cantor, R.F. Laboratory and greenhouse evaluation of the entomopathogenic fungi and garlic–pepper extract on the predatory mites, Phytoseiulus persimilis and Neoseiulus californicus and their effect on the spider mite Tetranychus urticae. Biol. Control 2011, 57, 143-149. [CrossRef]
- Saito, T.; Brownbridge, M. Compatibility of foliage-dwelling predatory mites and mycoinsecticides, and their combined efficacy against western flower thrips Frankliniella occidentalis. J. Pest Sci. 2018, 91, 1291–1300. [CrossRef]
- Holt, K.M.; Opit, G.P.; Nechols, J.R.; Margolies, D.C. Testing for non-target effects of spinosad on twospotted spider mites and their predator Phytoseiulus persimilis under greenhouse conditions. Exp. Appl. Acarol. 2006, 38, 141-149. [CrossRef]
- Jacas Miret, J.A.; Garcia-Mari, F. Side-effects of pesticides on selected natural enemies occurring in citrus in Spain. IOBC/WPRS Bull. 2001, 24, 103 – 112.
- Miles, M.; Dutton, R. Testing the effects of spinosad to predatory mites in laboratory, extended laboratory, semi-field and field studies. IOBC/WPRS Bull. 2003, 26, 9-20.
- Konopická, J.; Bohatá, A.; Nermuť, J.; Jozová, E.; Mráček, Z.; Palevsky, E.; Zemek, R. Efficacy of soil isolates of entomopathogenic fungi against the bulb mite, Rhizoglyphus robini (Acari: Acaridae). Syst. Appl. Acarol. 2021, 26, 1149-1167. https://orcid.org/0000-0002-7574-7243. [CrossRef]
- Yucel, C. Effects of local isolates of Beauveria bassiana (Balsamo) Vuillemin on the twospotted spider mite, Tetranychus urticae (Koch) (Acari: Tetranychidae). Egypt. J. Biol. Pest Control 2021, 31, 63. [CrossRef]
- Chaithra, M.; Prameeladevi, T.; Prasad, L.; Kundu, A.; Bhagyasree, S.N.; Subramanian, S.; Deba, K. Metabolomic diversity of local strains of Beauveria bassiana (Balsamo) Vuillemin and their efficacy against the cassava mite, Tetranychus truncatus Ehara (Acari: Tetranychidae). PLoS ONE, 2022, 17, e0277124. [CrossRef]
- Deka, B.; Babu, A.; Pandey, A.K.; Kumhar, K.C.; Rajbongshi, H.; Dey, P.; Peter, A.J.; Amalraj, E.L.D.; Tallur, V.R. Potential of the entomopathogenic fungus, Metarhizium anisopliae s.l. for control of red spider mite Oligonychus coffeae Nietner on tea crop. Int. J. Acarol. 2022, 48, 121-129. [CrossRef]
- Eroglu, C.; Cimen, H.; Ulug, D.; Karagoz, M.; Hazir, S.; Cakmak, I. Acaricidal effect of cell-free supernatants from Xenorhabdus and Photorhabdus bacteria against Tetranychus urticae (Acari: Tetranychidae). J. Invertebr. Pathol. 2019, 160, 61-66. [CrossRef]
- Aswini, M.N.; Bhaskar, H.; Mathew, D.; Shilaya, M.R.; Girija D. Isolation and evaluation of bacteria associated with entomopathogenic nematode Heterorhabditis spp. against the spider mite, Tetranychus truncatus Ehara (Acari: Tetranychidae). Egypt. J. Biol. Pest Control 2022, 32, 87. [CrossRef]
- Menon, A.G.; Bhaskar, H.; Subramanian, S.M.; Gopal, S.K. Acaricidal effects of two native isolates of entomopathogenic bacteria Photorhabdus luminescens (Thomas & Poinar) against spider mite, Tetranychus truncatus Ehara (Acari: Tetranychidae). Int. J. Acarol. 2023, 49, 395-399. [CrossRef]
- Camara, C.A.; Moraes, M.M.; Melo, J.P.; Silva M.M. Chemical composition and acaricidal activity of essential oils from Croton rhamnifolioides Pax and Hoffm. in different regions of a caatinga biome in northeastern Brazil. J. Essent. Oil-Bear. Plants 2017, 20, 1434-1449. [CrossRef]
- Lee, M.W.; Lee, D.H.; Nam, I.; Lee, J.W.; Huh, M.J.; Park, I.K. Acaricidal and insecticidal activities of essential oils and constituents of Tasmannia lanceolata (Poir.) A.C.Sm. (Canellales: Winteraceae) against Tetranychus urticae Koch (Trombidiformes: Tetranychidae) and Myzus persicae Sulzer (Hemiptera: Aphididae). J. Econ. Entomol. 2022, 116, 447-455. [CrossRef]
- Pavan, A.M.; Da-Costa, T.; Marques, A.J.; Ethur, E.M.; Buhl, B.; Soares, G.L.G.; Ferla, N.J. The use of essential oils as an alternative for the control of Tetranychus urticae (Acari: Tetranychidae). Crop Prot. 2024, 184, 106862. [CrossRef]
- Ma, X.F.; Zhang, Y.Y.; Guo, F.Y.; Luo, J.X.; Ding,W.; Zhang, Y.Q. Molecular characterization of a voltage-gated calcium channel and its potential role in the acaricidal action of scopoletin against Tetranychus cinnabarinus. Pestic. Biochem. Physiol. 2020, 168, 104618. [CrossRef]
- Miotto, J.; Duarte, A.F.; Bernardi, D.; Ribeiro, L.P.; Andreazza, F.; Cunha, U.S. Toxicities of acetogenin-based bioacaricides against two-spotted spider mite and selectivity to its phytoseiid predators. Exp. Appl. Acarol. 2020, 81, 173–187. [CrossRef]
- Maldonaldo-Michael, M.A.; Muňis-Valencia, R.; Peraza-Campos, A.L.; Parra-Delgado, H.; Chan-Cupul, W. Acaricidal, ovicidal and fagoinhibition activities of seed extracts from Swietenia humilis against Tetranychus urticae under laboratory conditions. Ind. Crops Prod. 2022, 177, 114494. [CrossRef]
- Rizzo, R.; Ragusa, E.; Benelli, G.; Verde, G.L.; Zeni, V.; Maggi, P.; Petrelli, R.; Spinozzi, E.; Ferrati, M.; Sinacori, M.; Tsolakis, H. Lethal and sublethal effects of carlina oxide on Tetranychus urticae (Acari: Tetranychidae) and Neoseiulus californicus (Acari: Phytoseiidae). Pest Manag. Sci. 2024, 80, 967-977. [CrossRef]
- Lovett, B.; St. Leger, R.J. Genetically engineering better fungal biopesticides. Pest Manag. Sci. 2018, 74, 781-789. [CrossRef]
- Gressel, J. Four pillars are required to support a successful biocontrol fungus. Pest Manag. Sci. 2024, 80, 35-39. [CrossRef]
- de la Cruz-Quiroz, R.; Maldonado, J.J,C.; de Jesús Rostro Alanis, M.; Antonio Torres, J.; Saldívar, R.P. Fungi based biopesticides: shelf life preservation technologies used in commercial products. J. Pest Sci. 2019, 92, 1003–1015. [CrossRef]
- Giunti, G.; Campolo, O.; Laudani, F.; Palmeri, V.; Spinozzi, E.; Bonacucina, G.; Maggi, F.; Pavela, R.; Canale, A.; Lucchi, A.; Benelli, G. Essential oil-based nanoinsecticides: ecological costs and commercial potential. In Development and commercialization of biopesticides - costs and benefits; Koul O. Ed.; Academic Press: London, UK, 2023, pp. 375-402. [CrossRef]
- Machado, S.; Pereira, R.; Sousa, R.M.O.F. Nanobiopesticides: are they the future of phytosanitary treatments in modern agriculture? Sci. Total Environ. 2023, 896, 166401. [CrossRef]
- Ebadollahi, A.; Sendi, J. J.; Aliakbar, A. Efficacy of nanoencapsulated Thymus eriocalyx and Thymus kotschyanus essential oils by a mesoporous material MCM-41 against Tetranychus urticae (Acari: Tetranychidae). J. Econ. Entomol. 2017, 110, 2413-2420. [CrossRef]
- Ahmadi, Z.; Saber, M.; Akbari, A.; Mahdavinia, G.R. Encapsulation of Satureja hortensis L. (Lamiaceae) in chitosan/TPP nanoparticles with enhanced acaricide activity against Tetranychus urticae Koch (Acari: Tetranychidae). Ecotoxicol. Environ. Saf. 2018, 161, 111-119. [CrossRef]
- de Oliveira, J.L.; de Oliveira, T.N.; Savassa, S.M.; Della Vechia, J.F.; de Matos, S.T.S.; de Andrade, D.J.; Fraceto, L.F. Encapsulation of Beauveria bassiana in polysaccharide-based microparticles: a promising carrier system for biological control applications. ACS Agric. Sci. Technol. 2023, 3, 785-794. [CrossRef]
- Arora, N.K.; Verma, M.; Prakash, J.; Mishra, J. Regulation of biopesticides: global concerns and policies. In Bioformulations: for sustainable agriculture; Arora, N.K., Mehnaz, S.; Balestrini, R. Eds.; Springer: New Delhi, India, 2016; pp. 283-299. [CrossRef]
- Balog, A.; Hartel, T.; Loxdale, H.D.; Wilson, K. Differences in the progress of the biopesticide revolution between the EU and other major crop-growing regions. Pest Manag. Sci. 2017, 73, 2203-2208. [CrossRef]
Figure 1.
Biopesticides - definitions and classifications: INV = Invertebrates; BOT = Botanicals; SEC = Semiochemicals; MIC = Microbials (microbial pesticides); MIN = Microbial metabolites produced at the industrial scale (fermentation products of soil actinomycetes); PIP = Plant incorporated protectans; BCH = biochemical pesticides; OECD = Organisation for Economic Co-operation and Development, FAO = Food and Agriculture Organization; WHO = World Health Organization; EU = European Union; US EPA = United States Environmental Protection Agency; ● EU legislation on plant protection products recognizes active substances that match BOT, SEC and MIN; ● US EPA recognizes biochemical pesticides as a biopesticide class that matches SEC and BOT with non-toxic action; ● China recognizes biochemical pesticides similarly to US EPA (plant-based growth regulators are included in BOT).
Figure 1.
Biopesticides - definitions and classifications: INV = Invertebrates; BOT = Botanicals; SEC = Semiochemicals; MIC = Microbials (microbial pesticides); MIN = Microbial metabolites produced at the industrial scale (fermentation products of soil actinomycetes); PIP = Plant incorporated protectans; BCH = biochemical pesticides; OECD = Organisation for Economic Co-operation and Development, FAO = Food and Agriculture Organization; WHO = World Health Organization; EU = European Union; US EPA = United States Environmental Protection Agency; ● EU legislation on plant protection products recognizes active substances that match BOT, SEC and MIN; ● US EPA recognizes biochemical pesticides as a biopesticide class that matches SEC and BOT with non-toxic action; ● China recognizes biochemical pesticides similarly to US EPA (plant-based growth regulators are included in BOT).
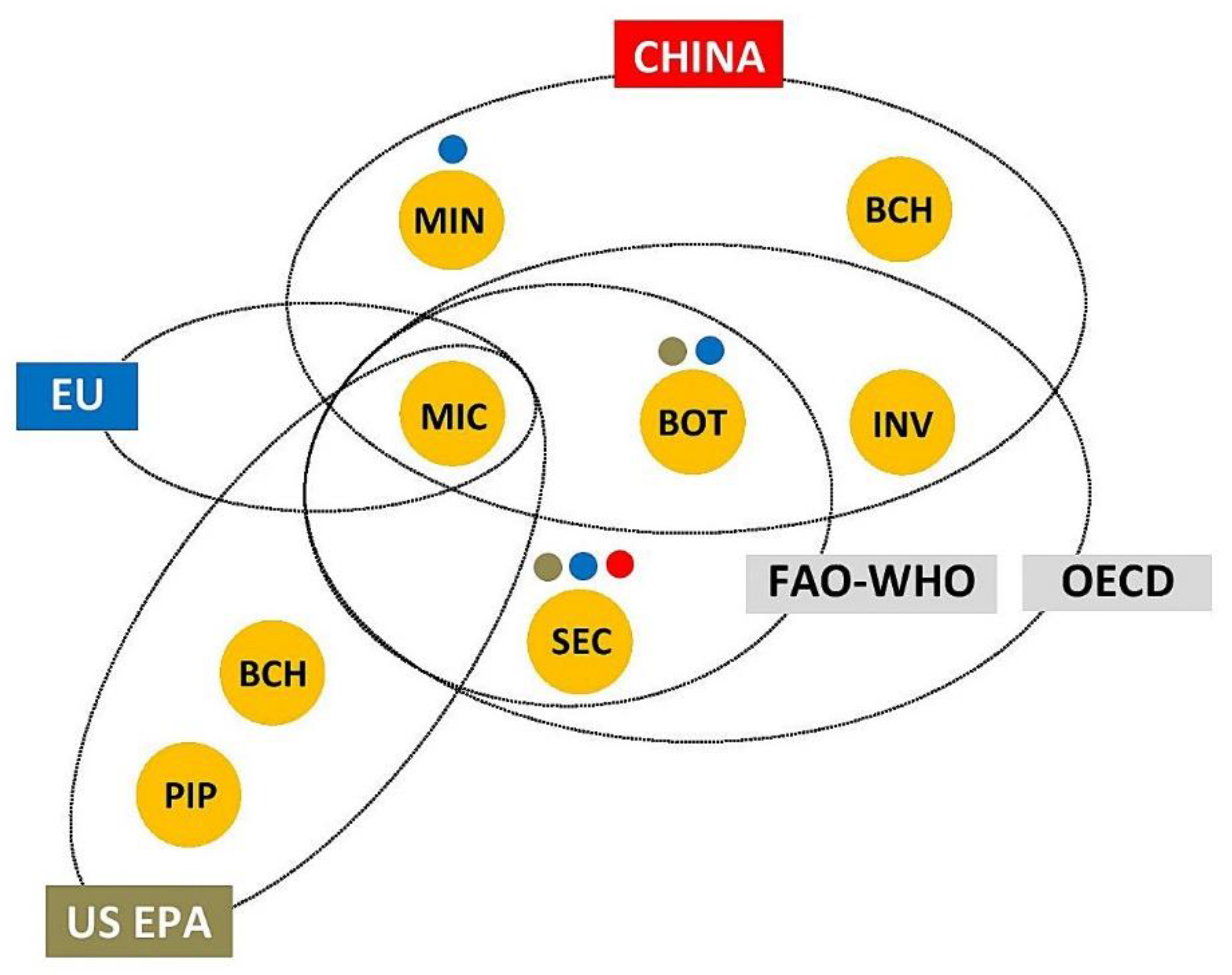
Table 1.
Examples of commercially available microbial acaricides - mycopesticides.
| Fungal species and strains | Commercial products | Manufacturers | Target mites |
|---|---|---|---|
| Beauveria bassiana | |||
| ATCC 74040 | Naturalis L | Danstar Ferment (USA) | Tet, Tar, Ten |
| GHA | Mycotrol, Botanigard | Certis (USA) | Tet, Tar, Eri |
| PPRI 5339 | Velifer | BASF Corporation (USA) | Pfm |
| Broadband | BASF (South Africa) | Tet | |
| R444 | Bb Protec, Eco-Bb | Andermatt (South Africa) | Tet |
| ESALQ-PL63 | Boveril | Koppert Biological Systems (Brazil) | Tur |
| IBCB-66 | Bovebio | Biofungi (Brazil) | Tur |
| Beauveria Oligos | Oligos Biotecnologia (Brazil) | Tur | |
| Eco-bass | Toyobo (Brazil) | Tur | |
| Not specified | Boverin | Cherkasy Biozakhyst (Ukraine) | Tet |
| Myco-Jaal | PCI (India) | Pfm | |
| Metarhizium brunneum * | |||
| F52/MA 43 | Lalguard M52 | Danstar Ferment (USA) | Pfm |
| Isaria fumosorosea ** | |||
| Apopka 97 | PFR 97 | Certis (USA) | Tet, Eri, Tar |
| FE9901 | Isarid | Koppert Biological Systems (USA) | Tet, Eri |
| PFA 011 | Paecilomite | AgriLife (India) | Pfm |
| Lecanicillium lecanii | |||
| MCC 0058 | Mealikil | AgriLife (India) | Pfm |
| V-80 | Biovert | Sibbiofarm (Russia) | Tet |
| Lecanicillium muscarium | |||
| Not specified | Verticillin | Cherkasy Biozakhyst (Ukraine) | Tet |
| Hirsutella thompsonii | |||
| HT 019 | Lancer | AgriLife (India) | Tet, Eri |
| Not specified | Almite | IPC Biologicals (India) | Tet, Eri |
This list was made by compiling data from several recent reviews of mycopesticides [46,63,64,68,70,71,72,73] official pesticide databases [25,69,74] and a number of manufacturer websites; Tur = T. urticae; Tet = Tetranychidae; Tar = Tarsonemidae; Eri = Eriophyidae; Ten = Tenuipalpidae; Pfm = (plant-feeding) mites; *formerly M. anisopliae var. anisopliae **formerly Paecilomyces fumosoroseus.
Table 2.
Examples of commercially available botanical biochemical acaricides.
| Active ingredients | Commercial products | Manufacturers | Target mites |
|---|---|---|---|
| pyrethrum (pyrethrin I and II)1 | Pyrethrum | PelGar International (UK) | Tet |
| Pyganic | MGK (USA) | Pfm | |
| Aphkiller | Kingbo Biotech (China) | Tet | |
| azadirachtin2 | NeemAzal T/S | Trifolio-M (Germany) | Tet |
| AzaGuard | BioSafe systems (USA) | Tet, Eri | |
| Azatin | Certis (USA) | Tet, Eri | |
| Ozomite | Ozone Biotech (India) | Tet | |
| Neem Kil | Kilpest (India) | Pfm | |
| Ecotin | PJ Margo (India) | Tet | |
| Azamax | UPL (Brazil) | Tur | |
| rosemary oil3 (1.8-cineole)* | EcoTrol | KeyPlex (USA) | Tet, Eri, Tar |
| + geraniol | |||
| + peppermint oil4 (menthol) | |||
| orange oil5 (d-limonene) | Prev-Am | Oro Agri (South Africa, USA) | Pfm |
| tea tree oil6 (terpinen-4-ol) | Eco-oil | OCP (Australia) | Tur |
| + eucalyptus oil7 (1.8-cineole) | |||
| cinnamon oil8 (cynnamaldehyde) | Akabrown | GreenCorp Biorganiks (Mexico) | Tet |
| + peppermint oil4 (menthol) | |||
| + clove oil9 (eugenol) | |||
| + oregano oil10 (carvacrol) | |||
| geraniol + citronellol | Mitexstream | Touchstone Enviro Solutions (USA) | Tet, Eri, Tar |
| geraniol + citronellol | Biomite | Arysta LifeScience (USA) | Tet |
| + nerolidol+ farnesol | |||
| α-terpinene + d-limonene + p-cymene | Requiem Prime | Bayer Crop Science (Germany) | Tet, Eri, Tar, Ten |
| matrine, oxymatrine11 | Matrine | Beijing Kingbo Biotech (China) | Pfm |
| veratrine12 | Marvee | Chengdu Newsun (China) | Tet |
| capsicum oleoresin13 (capsaicin) | Captiva | Gowan (USA) | Tet, Eri, Tar, Ten |
| + canola oil14 (oleic acid) | |||
| + garlic oil15 (allyl sulfides) | |||
| cottonseed oil16 (oleic acid) | GC-Mite | JH Biotech (Canada) | Pfm |
| + clove oil9 (eugenol) | |||
| + garlic oil15 (allyl sulfides) | |||
| castor oil17 (ricinoleic acid) | TetraCURB Max | Kemin (USA) | Tet |
| + rosemary oil3 (1.8-cineole) | |||
| + clove oil9 (eugenol) | |||
| + peppermint oil4 (menthol) | |||
| canola oil14 (oleic acid) | Neu1160 | Neudorff (Germany, USA) | Tet |
| rapessed oil14 (oleic acid) | Neu1160 I | Neudorff (Germany) | Tet |
| safflower oil18 (linoleic acid) | Suffoil | OAT Agrio )Japan) | Tet |
| + cottonseed oil16 (oleic acid) |
This list was made by compiling data from several recent reviews [10,23,26,40,94,95,96,97], official pesticide databases [25,69,74] and a number of manufacturer websites; Botanical sources: 1Tanacetum cinerariifolium (Asteraceae); 2Azadirachta indica (Meliaceae); 3Rosmarinus officinalis (Lamiaceae); 4Mentha piperita (Lamiaceae); 5Citrus sinensis (Rutaceae); 6Melaleuca alternifolia (Myrtaceae); 7Eucalyptus globulus (Myrtaceae); 8Cinnamomum verum (Lauraceae); 9Syzygium aromaticum (Myrtaceae); 10Origanum vulgare (Lamiaceae); 11Sophora flavescens (Fabaceae), 12Veratrum nigrum (Melanthiaceae); 13Capsicum annuum (Solanaceae); 14Brassica napus, B. rapa (Brassicaceae); 15Allium sativum (Amaryllidaceae); 16Gossypium hirsutum (Malvaceae); 17Ricinus communis (Euphorbiaceae); 18Carthamus tinctorium (Asteraceae); Tur = T. urticae; Tet = Tetranychidae; Tar = Tarsonemidae; Eri = Eriophyidae; Ten = Tenuipalpidae; Pfm = plant-feeding mites; * in brackets: the main active constituent.
Table 3a.
Evaluation of the compatibility of bioacaricides and phytoseiid mites according to the IOBC-WPRS standardized methods: the number of results listed in the database [125]; L = laboratory bioassays (○ initial toxicity bioassay; ● extended laboratory bioassay; ■ persistence bioassay); F = semi-field and field trials; T = total number of results
Table 3a.
Evaluation of the compatibility of bioacaricides and phytoseiid mites according to the IOBC-WPRS standardized methods: the number of results listed in the database [125]; L = laboratory bioassays (○ initial toxicity bioassay; ● extended laboratory bioassay; ■ persistence bioassay); F = semi-field and field trials; T = total number of results
| Bioacaricides | Aan | Ala | Asw | Ide | Kab | Nca | Ncu | Ppe | Ppl | Tpy | Σ | |||||||||||||||||||||||
| L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | T | ||||||||||||
| Microbial | ||||||||||||||||||||||||||||||||||
| B. bassiana | 1● | 1 | 8○● | 9 | 1 | 10 | ||||||||||||||||||||||||||||
| L. lecanii | 1● | 1 | 1○ | 1 | 2 | 2 | 4 | |||||||||||||||||||||||||||
| I. fumosorosea | 1 | 1 | 2 | 2 | ||||||||||||||||||||||||||||||
| M. brunneum | 1 | 1 | 1 | |||||||||||||||||||||||||||||||
| Biochemical | ||||||||||||||||||||||||||||||||||
| abamectin | 1○ | 1 | 2■ | 1● | 1● | 7● | 2 | 12 | 3 | 15 | ||||||||||||||||||||||||
| milbemectin | 7○● | 7 | 7 | |||||||||||||||||||||||||||||||
| spinosad | 1● | 1■ | 10○● | 12 | 12 | |||||||||||||||||||||||||||||
| azadirachtin | 2● | 2○● | 1 | 4 | 1 | 5 | ||||||||||||||||||||||||||||
| fatty acids | 1○ | 1○ | 2○● | 4 | 4 | |||||||||||||||||||||||||||||
| rapeseed oil | 1● | 3● | 4 | 4 | ||||||||||||||||||||||||||||||
| thymol | 1○ | 1 | 1 | |||||||||||||||||||||||||||||||
| 1 | 1 | 2 | 1 | 1 | 3 | 1 | 1 | 6 | 2 | 1 | 41 | 4 | 55 | 10 | 65 | |||||||||||||||||||
Aan= Amblyseius andersoni; Ala = A. largoensis; Asw = A. swirskii; Ide = Iphiseius degenerans; Kab = Kampimodromus aberrans; Nca = Neoseiulus californicus; Ncu = N. cucumeris; Ppe = Phytoseiulus persimilis; Ppl = Phytoseius plumifer; Tpy = Typhlodromus pyri.
Table 3b.
Evaluation of the compatibility of bioacaricides and phytoseiid mites according to the IOBC-WPRS standardized methods: IOBC-WPRS toxicity classification* [125]; L = laboratory bioassays, F = semi-field and field trials; T = total number of results.
Table 3b.
Evaluation of the compatibility of bioacaricides and phytoseiid mites according to the IOBC-WPRS standardized methods: IOBC-WPRS toxicity classification* [125]; L = laboratory bioassays, F = semi-field and field trials; T = total number of results.
| Bioacaricides | Aan | Ala | Asw | Ide | Kab | Nca | Ncu | Ppe | Ppl | Tpy | |||||||||||||||||||
| L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | L | F | ||||||||||
| Microbial | |||||||||||||||||||||||||||||
| B. bassiana | 2 | 1 | 1-2 | ||||||||||||||||||||||||||
| L. lecanii | 1 | 1 | 1 | 2 | |||||||||||||||||||||||||
| I. fumosorosea | 1 | 1 | |||||||||||||||||||||||||||
| M. brunneum | 1 | ||||||||||||||||||||||||||||
| Biochemical | |||||||||||||||||||||||||||||
| abamectin | 3 | 2 | 3-1a | 4 | 4 | 2-4 | 1 | ||||||||||||||||||||||
| milbemectin | 1-4 | ||||||||||||||||||||||||||||
| spinosad | 4 | 1b | 1-4 | ||||||||||||||||||||||||||
| azadirachtin | 1-4 | 1-3 | 1 | ||||||||||||||||||||||||||
| fatty acids | 3 | 1 | 1-3 | ||||||||||||||||||||||||||
| rapeseed oil | 1-3 | 1-3 | |||||||||||||||||||||||||||
| thymol | 1 | ||||||||||||||||||||||||||||
Aan= Amblyseius andersoni; Ala = A. largoensis; Asw = A. swirskii; Ide = Iphiseius degenerans; Kab = Kampimodromus aberrans; Nca = Neoseiulus californicus; Ncu = N. cucumeris; Ppe = Phytoseiulus persimilis; Ppl = Phytoseius plumifer; Tpy = Typhlodromus pyri *Laboratory bioassays: 1 = harmless (E < 30%), 2 = slightly harmful (E = 30 - 80%), 3 = moderately harmful (E = 80 - 99%), and 4 = harmful (E > 99%); Field trials: 1 = harmless ( E< 25%), 2 = slightly harmful (E = 25 - 50%), 3 = moderately harmful (E = 50 - 75%), and 4 = harmful (E > 75%); E = the total effect (%); Underlined toxicity classes refer to the persistence bioassay: a = residue aged 5 days (3) to 15.-30 days (1), b = residue aged 5 days.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



