
Submitted:
09 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
Background: The patterns of inbreeding coeficient (FIS) and fine-spatial genetic structure have been evaluated regarding mating system and inbreeding depression of food-deceptive orchid populations, Dactylorhiza majalis, Dactylorhiza incarnata var. incarnata, and Dactylorhiza fuchsii from NE Poland. Methods: We used 455 individuals representing nine populations of three Dactylorhiza taxa and AFLP markers to estimate percent polymorphic loci (P%) and Nei’s gene diversity (H), calculated using the Bayesian method, FIS, FST, the spatial autocorrelation analysis with the pairwise kinship coefficient Fij, and AMOVA in populations. The genetic diversity parameters were discussed applying data from fruit set, in vitro seed germination and inbreeding depression of three Dactylorhiza. Results: We detected a relatively high proportion of polymorphic fragments (P = 40.4.5–68.4%) and Nei’s gene diversity indices (H = 0.140–0.234). The overall FIS was relatively low-to-moderate 0.071–0.312. The average Fij for the populations of three Dactylorhiza taxa showed significantly positive values, which were observed in the short-distance classes (1-10m (20 m)). FST values were significant in each Dactylorhiza taxon, ranging from the lowest values in D. fuchsii and D. majalis (0.080-0.086, p < 0.05), and the higher value (0.163, p < 0.05) in D. incarnata var. incarnata. The molecular variance was highest within populations (AMOVA: 76.5-86.6%; p < 0.001). Conclusions: We observed the concordant genetic diversity patterns in food-deceptive, allogamous and pollinator-dependent, although also self-compatible with the mix-mating system three Dactylorhiza. In our survey, FIS is often substantially higher than Fij at the first class of SGS, suggesting that selfing (meaning of geitonogamy) is at least responsible for homozygosity. The strong SGS may have additional and unexplored evolutionary consequences in Dactylorhiza, and combined with low inbreeding depression, may strongly influence establishing the inbred lines in the case of D. majalis and D. incarnata var. incarnata.
Keywords:
Dactylorhiza fuchsii
; Dactylorhiza incarnata var. incarnata
; Dactylorhiza majalis
; FIS
; spatial genetic structure
1. Introduction
The mating system affects plant population genetic structure by modifying the drift/migration equilibrium characterized by the effective size of a population [1,2]. The mating system is also related to inbreeding in a complex way. Theoretical and experimental studies often supported that pollen transfer in outcrossers leads to lower genetic structure and higher genetic diversity compared to selfers resulting from a strong founder event, which increases the inbreeding [2,3,4]. However, pollen flow may act in concert with typical life-history traits related to dispersal mechanisms, such as seeds dispersed by wind or animals, and they can show together high power to predict the magnitude of FSGS within populations [5,6]. When pollen and seed dispersal are limited, resulting in strong intra-population structure, biparental inbreeding can also affect FIS. Therefore, the strong fine spatial genetic structure (FSGS) is frequent even within allogamous or potentially allogamous plants following an isolation-by-distance model [6]. The higher levels of fine-spatial genetic structure (FSGS) have also been highlighted for selfing and clonal species and in low-density populations [6,7,8]. Variations in mating systems and different edaphic and climatic may additionally translate into substantial differences in FSGS conditions [9]. FIS also reflects inbreeding in previous generations of perennials, resulting in the Wahlund effect on the population [10]. Mating system and seed dispersal also influence FST via its effect on pollen-mediated and short and/or leptokurtic gene flow and effective population size, especially in selfing and mating between relatives by increasing inbreeding, which enhances genetic drift. A summary report by Duminil et al. [11] based on data from 263 plant species revealed that the FIS observed at the adult plant stage allows testing of the impact of both biparental inbreeding and inbreeding depression on population genetic structure. In the earliest stages of the plant life cycle, inbreeding depression mainly affects inbred progeny. Therefore, the FIS of adult plants includes information on the selfing rate and inbreeding depression.
In Orchidaceae, the mating system and gene flow within and among populations can generate common genetic diversity patterns and FSGS [12]. First, pollinator-mediated gene flow among populations, e.g. is higher in deceptive than in rewarding orchids [13,12]. The deceived pollinators typically visit only a few flowers among plants within populations, thereby promoting cross-pollination and reducing the likelihood of inbreeding [14,15,16,17,19]. Therefore, it can be hypothesized that FSGS of these orchids is weak. In contrast, in rewarding ones, within-population genetic structure could be stronger due to geitonogamy and mating among close relatives. However, based on earlier orchid studies, the dispersal of dusty-like seeds seems to be limited [e.g. 20,21,22,23,24,25]. These results agree with studies that have investigated FSGS both of deceptive orchids e.g. Caladenia tentaculata [26], Cephalanthera longibracteata [22], Orchis cyclochila [23], Orchis purpurea [27], and Orchis mascula [28,29], Cymbidium goeringii [30] and rewarding ones e.g. Gymnadenia conopsea [31], Pogonia ophioglossoides [32], Epipactis thunbergii [33]. Moreover, orchid germination success has been reported to be higher in the vicinity of mother plants because a mycorrhizal could favour the establishment of seedlings [34,35].
In this study, we focused on Dactylorhiza taxa, which are food-deceptive orchids with no rewards for their pollinators [36]. This genus can be considered as a model one due to plant-pollinator interactions, natural selection, and consequent female reproductive success and their impact on genetic structure in food-deceptive plant groups [37,38,39]. In this context, the mating system and ID in food-deceptive Dactylorhiza majalis, Dactylorhiza incarnata var. incarnata, and Dactylorhiza fuchsii populations from NE Poland have been documented in detail comprehensively [40,41,4243]. A mixed mating system was observed in all three studied Dactylorhiza taxa similar to Hedrén and Nordström study [44]. Ostrowiecka et al. [40] found that pollinator behavior in D. majalis likely promotes geitonogamy, explaining the development of selfed seeds in fruits at different levels of the inflorescence with germination potential similar to that of outcrosses within populations. Vallius et al. [45] and Hedrén and Nordstöm [44] argued that different D. incarnata varieties maintain a high level of inbreeding, and populations might consist of several inbred lines that were fixed for characters such as flower colour, leaf shape, and leaf spot. Wróblewska et al. [42] results corroborate with previous studies on Dactylorhiza, concerning the low or medium level of fruit set ranged from 7.4% to 77.5% [36,46,45]. In vitro experiments revealed that seed germination of three Dactylorhizataxa both from natural pollination and hand-treatments (selfing and outcrossing) shaped at a relatively low level, up to 35% (exception of D. fuchsii, outcrossing experiments) [42]. The in vitro asymbiotic seed germination was similar or slightly higher in selfing than crossing experiments in D. incarnata var. incarnata and D. majalis, while it was reversed in D. fuchsii [42]. Spontaneous autogamy in three Dactylorhiza taxa existed in < 1% of pollination in the studied populations and most likely did not affect reproductive success [47,48]. Taxa are assumed to be terrestrial, long-lived, self-compatible, and tuberous perennial orchids that reproduce either by seeds or (rarely) vegetatively [49,40]. Pollination occurs by different taxonomical groups of insects (Hymenoptera, Diptera, and/or Coleoptera, mostly bees and bumblebees) [40,41]. Molecular markers such as cpDNA (trnL, trnF and psbC–trnK), internal transcribed spacer (ITS) sequences, and flow cytometry data confirmed the taxonomic status of the studied three orchids [41].
Based on estimates of earlier ecological survey, e.g. natural fruit set, mix-mating system, inbreeding depression from controlled crosses treatment and of studied orchid taxa from NE Poland [40,42,43] and genetic report of Hedrén and Nordstöm [44], Naczk et al. [49] we tested the hypotheses that that inbreeding coefficient is shaped at a high level the food-deceptive orchids D. majalis, D. incarnata var. incarnata, and D. fuchsii. We also assumed that seed dispersal is mainly a short distance in orchids close to the mother plant, as was observed by many authors who researched experimentally seed dispersal; therefore, the fine-scale genetic structure is stronger due to the effect of inbreeding and short-distance dispersal. Finally, the purpose of this study is (1) to estimate the inbreeding coefficient and the intensity of FSGS using AFLP markers, (2) to discuss how similar mating systems and different ID shape genetic diversity patterns of three food-deceptive Dactylorhiza taxa.
2. Materials and Methods
2.1. Study Sites
The present study was performed from May to July in 2014 to 2017 in three D. majalis populations (KA, SKI, and SKII), three D. incarnata var. incarnata populations (ZB, RO, and MR), and three D. fuchsii populations (BR, CM, and GR) located in northeastern Poland (Figure 1). D. majalis grows in wet meadows with abundant, entomophilous, and rewarding plants. The study sites differed in the abundance of D. majalis individuals, from ca. 120–200 flowering individuals in SKI and SKII, to ca. 1,000 flowering individuals in KA. All meadows were extensively used, mown every year in late July or early August, and not artificially fertilized. Three populations of D. incarnata var. incarnata had similar sizes (MA, ca. 68–100 flowering plants; ZB, ca. 30–100 flowering plants; and RO, ca. 35–200 flowering plants; Figure 1). The populations in the Biebrza Valley and Rospuda Valley occupied sedge communities with a low cower of the herb layer by rewarding plant species (ca. 10%). D. fuchsii was observed in open hornbeam forests, with a low number of rewarding plants, in the Białowieża Primeval Forest and its vicinity (CM and BR, population sizes ca. 84–133 flowering plants). One D. fuchsii population (GR, population size ca. 140–193 flowering plants) was located in the Biebrza Valley [42].
The study was based on samples comprising 455 individuals representing nine populations of three Dactylorhiza taxa (162, 129 and 164 individuals of DM, DI and DF, respectively; Table 1; Figure 1). Even though Dactylorhiza rarely regenerates clonally, one leaf sample was taken from a single shoot distance of 1 m from one another within each population to avoid the effects of the population substructure. In all populations each sample was mapped in the grid coordinate system, using a hand-held GPS (Garmin GPSMAP 65s) to calculate the distance between samples.
2.2. AFLP Analysis
Genomic DNA was extracted from dry leaf tissues with the Genomic Mini AX Plant kit (A & A Biotechnology, Poland), and then samples were genotyped for AFLP markers. The AFLP procedure described by Vos et al. [50] was modified according to the Applied Biosystems protocol (AFLPTM Plant Mapping). First, 12 primer pair combinations were tested on four selected samples from each Dactylorhiza taxon. The fluorescence-labeled selective amplification products were mixed with a 500 Liz-labeled size standard (Applied Biosystems) and run on an ABI 3130. From this analysis, we chose seven primer combinations that produced polymorphic, clear, reproducible fragments of homogeneous intensity in three Dactylorhiza taxa (D. majalis EcoR1-ACC/MseI-CAG; EcoR1-AGG/MseI-CAC; D. incarnata var. incarnata EcoR1-ACA/MseI-CAG; EcoR1-ACA/MseI-CTA; D. fuchsii EcoR1-AGG/MseI-CAG, EcoR1-ACC/MseI-CAT, EcoR1-ACC/MseI-CTA). Variable fragments in the 70–500 bp size range were scored as present (1) or absent (0) using Genemapper 4.0 (Applied). To test the repeatability of the AFLP results, three individuals from each population were completely replicated, starting from the restriction/ligation reaction of the AFLP. Potential resampling of clones was checked with the AFLPdat R-script but was insignificant, thus, not corrected for.
To assess the levels of genetic diversity, the proportion of polymorphic fragments (PLpoly) and Nei’s gene diversity (H) were calculated using the Bayesian method with a nonuniform prior distribution of allele frequencies proposed by Zhivotovsky [51], as implemented in AFLP-Surv ver. 1.0 [52]. The F statistic was determined by analysis of the molecular variance (AMOVA) using the program Arlequin 3.11 [53]. The significance of the variance components was determined using 1,000 independent permutation runs.
The FSGS was described by conducting spatial autocorrelation analysis using the pairwise kinship coefficient Fij for dominant markers [54]. Mean Fij estimates over pairs of individuals for given distance classes were calculated and plotted against distance on a logarithmic scale using the software SPAGeDi 1.4 [54,6]. Separate distance classes (m) were created for each population of D. majalis, D. incarnata var. incarnata, and D. fuchsii due to different patterns of plant distribution in space. To test the significance of FSGS, the regression slopes (b) of kinship coefficients and the
natural logarithm of distance were compared with the slopes obtained for permutations of individual genotypes (10,000 random permutations). The extent of FSGS was quantified using the Sp statistic proposed by Vekemans and Hardy [6] and calculated as Sp = −b / (1 − F1), where b is the regression slope and F1 is the average Fij between individuals. For each spatial distance class, the 99% confidence interval was computed using 1,000 permutations (with SPAGeDi) [55]. The probability value (P) was computed for each spatial distance class and coefficient.
To investigate the FIS, the Metropolis–Gibbs algorithm was applied to I4A software based on dominant markers [56]. The data were run using the prior values of beat-distribution equal to α = β = 1.0 (corresponding to an ‘uninformative’ flat distribution) and 60,000 repetitions, including a 10,000 step burn-in.
3. Results
Overall, 193, 215, and 263 polymorphic bands were scored in D. majalis, D. incarnata var. incarnata, and D. fuchsii, respectively. Considering the error rates (2%, 1.3%, and 1.5%, respectively), none of the samples may have represented clones.
A relatively high proportion of polymorphic fragments (P = 40.4.5–68.4%) and Nei’s gene diversity indices (H = 0.140–0.234) were detected among the three orchid species (Table 1). The overall FIS was relatively low-to-moderate and equaled 0.071–0.224 in D. incarnata var. incarnata, 0.079–0.134 in D. fuchsii, and reached the highest values of 0.192–0.312 in D. majalis.
Correlograms of the average Fij values for the populations of three Dactylorhiza taxa showed significantly positive values, which were observed in the short-distance classes. In D. majalis the relatives were noted at a distance from 1 m to 10 m (Figure 2a), and the values were significantly negative at the longer distance classes (52–72 m) in the two other populations. Similar observations were made in two of the three populations of D. incarnata var. incarnata and D. fuchsii. Significant positive values were observed at a distance from 2 m to 20 m in D. incarnata var. incarnata (Figure 2b) and from 2 m to 10 m in D. fuchsii (Figure 2c). The bF values for D. majalis (−0.051–0.009), D. incarnata var. incarnata (−0.055–0.002), and D. fuchsii (−0.026–0.002) were almost all significant (permutation test, p < 0.05) (Table 1). The highest Sp values were observed for D. incarnata var. incarnata (0.063) and D. majalis (0.056) (Table 1).
4. Discussion
The associations of mating system, inbreeding depression and biparental inbreeding with FIS and FST have been confirmed sparsely in plant surveys ([11], and literature herein). Baskin and Baskin [57]) summarized the effects of inbreeding depression on seed germination in 743 cases of 233 species in 64 families. They demonstrated that in 50.1% of cases, inbred and outbred seeds germinated at a similar frequency, and 8.1% of inbred seeds germinated better than outbred seeds. Notably, the authors did not find a strong relationship between the decrease in germination and an increase in FIS or between an increase in germination and an increase in population genetic diversity. However, we observed the concordant genetic diversity patterns in food-deceptive, allogamous and pollinator-dependent, although also self-compatible with the mix-mating system D. majalis, D. incarnata var. incarnata, and D. fuchsii. Genetic diversity within studied Dactylorhiza populations was shaped at a relatively high level comparable with the data reported by Naczk et al. [49] as well Hedrén and Nordström [44,58], suggesting that studied Dactylorhiza populations can be found by multiple, genetically diverse individuals and/or by gene flow (leptokurtic dispersal) from the surrounding populations. The genetic differentiation among them was low and significant (0.080-0.163), showing that gene flow (historical) in NE Poland was relatively high or population were established from one source. However, the isolation processes of these Dactylorhiza population were observed, leading to the formation of substructure.
Furthermore, the inbreed was shaped on moderate to high levels in three Dactylorhiza taxa similar to studies Hedrén and Nordström [44], Filippov et al. [59] and Naczk et al. [49]. Meanwhile, in D. majalis as the allotetraploid, the FIS has a wide range of values in populations reported by Balao et al. [59], Hedrén and Nordström [44] and Naczk and Ziętara [61]. If we predict that inbreeding is solely the result of mating among neighboring plants, we will expect FIS to be approximately equal to Fij at the smallest distance interval in studied Dactylorhiza populations [6]. In our survey, FIS is substantially higher than Fij at the first class of spatial distance in the majority of Dactylorhiza populations, suggesting that selfing is at least partially responsible for homozygosity [56,62]. However, the spontaneous autogamy in three Dactylorhiza taxa existed until 1% of pollination in the studied populations [47,48]. Hence, the only explanation of selfing in three Dactylorhiza taxa is the pollinator behaviour of bumblebees and other pollinators, which were known to promote geitonogamy and/or autogamy, explaining the development of selfed seeds in fruits [40,41]. The important factor shaping FIS was the slightly higher frequency of selfed than outcross seeds germinated in vitro treatments in D. majalis, and D. incarnata var. incarnata, while in D. fuchsii, the germination pattern was reversed [42]. This phenomenon suggested that inbred and outbred D. majalis, and D. incarnata var. incarnata seeds germinated at a similar or even slightly higher frequency. We stress the careful interpretation of the relationship between seed germination and FIS. This needs to be confirmed by further studies, including the following stages of growth and mortality observations of plants germinated from selfed and outcross seeds. However, these data were shown for a single studied Dactylorhiza species; we can suppose that in D. fuchsii, a similar pattern existed in two out of three populations like in D. majalis and D. incarnata var. incarnata. In the CM and BR populations, high inbreeding and slightly lower kinship coefficient supported the possibility of selfing (geitonogamy). The interesting question is whether biparental inbreeding can exist in food-deceptive Dactylorhiza taxa, even though pollinators spend a short time on flowers and inflorescence and learn to avoid the deceptive flowers. They typically visit probe fewer flowers per plant only and/or a few flowers between inflorescences within populations, promoting cross-pollination and skipping more plants between plant visits. In the light of this outcrossing hypothesis [15], the biparental inbreeding is rather unlikely. Ostrowiecka et al. [40] observed using videotaping in D. majalis that A. mellifera visited three to five flowers on the same inflorescence over a period of 11 s to 40 s contributing to geitonogamy. We observed that A. mellifera pollinators never repeatedly returned to the same flowers and never visited all the flowers on the inflorescences. On the other hand, pollinaria bending is a mechanism that prevents geitonogamy and biparental inbreeding between individuals, especially in proximity. In Dactylorhiza, the mean bending is 39–54 s, considered a relatively long time for deceptive plants and similar to other deceptive Dactylorhiza taxa [63]. The banding time span in each studied D. majalis population ranged from 8 s to 2 min 5 s [40]. This short bending time can likely provide an opportunity for geitonogamy. This phenomenon and the bending times in the studied populations support our hypothesis that we cannot completely exclude the possibility of geitonogamy than biparental inbreeding in deceptive orchids. Dactylorhiza appears to have a more generalised pollination system and many pollinator were described and studied in detail. These pollinators can spend different time on the flowers promoting geitonogamy.
However, hand pollination treatment with emasculated flowers was also used as the level of apparent selfing in plants [64,65]. A previous fruit set observation from control pollination in three Dactylorhiza documented a moderate level of fruit set (35.4-40.5%), while the emasculation experiments in their populations showed a significant decrease in fruiting between these treatments (D. majalis, 28.2% fruit set from emasculated flower, paired t = 2.68, df = 8, p < 0.002; D. incarnata var. incarnata, 14.6% fruit set from emasculated flower, paired t = 3.46, df = 10, p < 0.006; D. fuchsii, 28.2% fruit set from emasculated flower, paired t = 4.83, df = 10, p < 0.0007; Wróblewska et al. unpublished data). This study concludes that selfing in three Dactylorhiza occurs mainly through geitonogamy. Kropf and Renner [18] have also pointed out the high levels of geitonogamous pollination in Dactylorhiza. In this fact, measuring biparental inbreeding can be challenging in deceptive plants.
The FIS observed at the adult stage allowed testing of the impact of inbreeding depression on the population's genetic structure. In long-living plants, FIS reflects inbreeding not only in the current generation but also in previous overlapping generations. However, other factors, such as the long lifespan of plants, can affect inbreeding depression [11]. In D. majalis, the selfing (e.g., geitonogamy) and/or progeny, and likely seed dispersal very close to the mother plant, can manifest most strongly in a spatial genetic structure. In the case of D. majalis, the results of the present study are inconsistent with those of Husband and Schemske [66], who concluded that purging is a significant evolutionary force in natural populations. Without actually reducing the genetic load, such fixation could reduce inbreeding depression [66,67]. However, inbreeding depression may be lower in long-standing populations with inbreeding than in populations with outcrossing populations where selection may have purged the genome of its genetic load [68,66,67,69]. These two alternative approaches should be tested in a laboratory at a later stage of the life cycle of D. majalis, such as in seedlings and adult reproductive individuals. The selfing (meaning of geitonogamy) and the strong fine-scale genetic structure may have additional and unexplored evolutionary consequences in Dactylorhiza, and combined with low inbreeding depression may strongly influence to establish the inbred lines in the case of D. majalis and D. incarnata var. incarnata. Today, we cannot state in Dactylorhiza that inbreeding depression may be widely viewed as the primary selective factor allowing transition to complete selfing.
Author Contributions
Conceptualization, A.W; methodology, A.W. and B.O.; software, A.W.; validation, A.W; formal analysis, A.W; investigation, A.W, B.O, E.J, and I. T.; writing—original draft preparation, A.W.; writing—review and editing, A.W.; visualization, X.X.; supervision, X.X.; project administration, X.X.; funding acquisition, Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Center in Poland (no. 2013/09/B/NZ8/03350).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data cited in the study are publicly available.
Acknowledgments
We thank Emilia Brzosko and Paweł Mirski for their help and support during fieldwork.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hamrick, J.L.; Godt, M.J.W. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. B: Biol. Sci. 1996, 351, 1291–1298, . [CrossRef]
- Hamrick, J.L.; Linhart; V.B.; Mitfon, J. Relationships between life history characteristics and electrophoretically detectable genetic variation in plants. Annual Review of Ecology; Evolution; and Systematics, 1979, 10, 173–200.
- Loveless, M.D.; Hamrick, J.L. Ecological Determinants of Genetic Structure in Plant Populations. Annu. Rev. Ecol. Syst. 1984, 15, 65–95, . [CrossRef]
- Nybom, H.; Bartish, I.V. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 93–114, . [CrossRef]
- Petit, R.J.; Duminil, J.; Fineschi, S.; Hampe, A.; Salvini, D.; Vendramin, G.G. INVITED REVIEW: Comparative organization of chloroplast, mitochondrial and nuclear diversity in plant populations. Mol. Ecol. 2004, 14, 689–701, . [CrossRef]
- Vekemans, X.; Hardy, O.J. New insights from fine-scale spatial genetic structure analyses in plant populations. Mol. Ecol. 2004, 13, 921–935, . [CrossRef]
- Volis, S.; Zaretsky, M.; Shulgina, I. Fine-scale spatial genetic structure in a predominantly selfing plant: role of seed and pollen dispersal. Heredity 2009, 105, 384–393, . [CrossRef]
- Binks, R.M.; Millar, M.A.; Byrne, M. Not all rare species are the same: contrasting patterns of genetic diversity and population structure in two narrow-range endemic sedges. Biol. J. Linn. Soc. 2015, 114, 873–886, . [CrossRef]
- Mosca, E.; Di Pierro, E.A.; Budde, K.B.; Neale, D.B.; González-Martínez, S.C. Environmental effects on fine-scale spatial genetic structure in four Alpine keystone forest tree species. Mol. Ecol. 2017, 27, 647–658, . [CrossRef]
- Wahlund, S. Zusammensetzung von Population und Korrelationserscheinung vom Standpunkt der Vererbungslehre aus betrachtet. Hereditas, 1928, 11, 65–106.
- Duminil, J.; Hardy, O.J.; Petit, R.J. Plant traits correlated with generation time directly affect inbreeding depression and mating system and indirectly genetic structure. BMC Evol. Biol. 2009, 9, 177–177, . [CrossRef]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification. Biol. J. Linn. Soc. 2004, 84, 1–54, . [CrossRef]
- Cozzolino, S.; Schiestl, F.P.; Müller, A.; De Castro, O.; Nardella, A.M.; Widmer, A. Evidence for pollinator sharing in Mediterranean nectar-mimic orchids: absence of premating barriers?. Proc. R. Soc. B: Biol. Sci. 2005, 272, 1271–1278, . [CrossRef]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671, . [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219–235, . [CrossRef]
- Juillet, N.; Dunand-Martin, S.; Gigord, L.D.B. Evidence for Inbreeding Depression in the Food-Deceptive Colour-Dimorphic Orchid Dactylorhiza sambucina (L.) Soò. Plant Biology, 2006, 9(1), 147-51.
- Kropf, M.; Renner, S.S. Pollinator-mediated selfing in two deceptive orchids and a review of pollinium tracking studies addressing geitonogamy. Oecologia 2007, 155, 497–508, . [CrossRef]
- Jacquemyn, H.; Brys, R. Lack of strong selection pressures maintains wide variation in floral traits n a food-deceptive orchid. Ann. Bot. 2020, 126, 445–453, . [CrossRef]
- Machon, N.; Bardin, P.; Mazer, S.J.; Moret, J.; Godelle, B.; Austerlitz, F. Relationship between genetic structure and seed and pollen dispersal in the endangered orchidSpiranthes spiralis. New Phytol. 2003, 157, 677–687, . [CrossRef]
- Jersáková, J.; Malinová, T. Spatial aspects of seed dispersal and seedling recruitment in orchids. New Phytol. 2007, 176, 237–241, . [CrossRef]
- Brzosko, E.; Ostrowiecka, B.; Kotowicz, J.; Bolesta, M.; Gromotowicz, A.; Gromotowicz, M.; Orzechowska, A.; Orzołek, J.; Wojdalska, M. Seed dispersal in six species of terrestrial orchids in Biebrza National Park (NE Poland). Acta Soc. Bot. Pol. 2017, 86, . [CrossRef]
- Chung, M.Y.; Nason, J.D.; Chung, M.G. Spatial genetic structure in populations of the terrestrial orchid Cephalanthera longibracteata (Orchidaceae). American Journal of Botany, 2004, 91, 52–57.
- Nason, J.D.; Chung, M.G. Spatial genetic structure in populations of the terrestrial orchid Orchis cyclochila (Orchidaceae). Plant Syst. Evol. 2005, 254, 209–219, . [CrossRef]
- Trapnell, D.W.; Hamrick, J.L. Mating patterns and geneflow in the Neotropical epiphytic orchid; Laelia rubescens. Molecular Ecology, 2005, 14; 75–84.
- Trapnell, D.W.; Hamrick, J.L.; Nason, J.D. Three-dimensional fine-scale genetic structure of the Neotropicalepiphytic orchid; Laelia rubescens. Molecular Ecology, 2004,13 1111–1118.
- Peakall, R.; Beattie, A.J. Ecological and genetic consequences of pollination by sexual deception in the orchid Calladenia tentaculata. Evolution, 1996, 50, 2207–2220.
- Jacquemyn, H.; Brys, R.; Vandepitte, K.; Honnay, O.; Roldán-Ruiz, I. Fine-scale genetic structure of life history stages in the food-deceptive orchid Orchis purpurea. Molecular Ecology, 2006, 15, 2801–2808.
- Jacquemyn, H.; Wiegand, T.; Vandepitte, K.; Brys, R.; Roldán-Ruiz, I.; Honnay, O. Multigenerational analysis of spatial structure in the terrestrial, food-deceptive orchid Orchis mascula. J. Ecol. 2009, 97, 206–216, . [CrossRef]
- Helsen, K.; Meekers, T.; Vranckx, G.; Roldán-Ruiz, I.; Vandepitte, K.; Honnay, O. A direct assessment of realized seed and pollen flow within and between two isolated populations of the food-deceptive orchidOrchis mascula. Plant Biol. 2015, 18, 139–146, . [CrossRef]
- Chung, M.Y.; Nason, J.D.; Chung, M.G. Significant fine-demographic and scale genetic structure in expanding and senescing populations of the terrestrial orchid Cymbidium goeringii (Orchidaceae). American Journal of Botany, 2011, 98, 2027–2039.
- Sletvold, N.; Grindeland, J.M.; Zu, P.; Ågren, J. Fine-scale genetic structure in the orchid Gymnadenia conopsea is not associated with local density of flowering plants. American Journal of Botany, 2024, 111 (2), e16273.
- Pandey, M.; Sharma, J. Efficiency of microsatellite isolation from orchids via next generation sequencing. Open J. Genet. 2012, 02, 167–172, . [CrossRef]
- Chung, M.Y.; Chung, M.G. Extremely low levels of genetic diversity in the terrestrial orchid Epipactis thunbergii (Orchidaceae) in South Korea, implications for conservation Botanical Journal of the Linnean Society, 2007, 155, 161–169.
- Diez, J.M. Hierarchical patterns of symbiotic orchid germination linked to adult proximity and environmental gradients. J. Ecol. 2006, 95, 159–170, . [CrossRef]
- Jacquemyn, H.; Wiegand, T.; Vandepitte, K.; Brys, R.; Roldán-Ruiz, I.; Honnay, O. Spatial variation in below-ground seed germination and divergent mycorrhizal associations correlate with spatial segregation of three co-occurring orchid species. Journal of Ecology, 2012, 100 (6), 1328–1337.
- Claessens, J.; Kleynen, J. The flower of the European orchid, form and function. 2011. Voerendaal, Jean Claessens and Jacques Kleynen.
- Mattila, E.; Kuitunen, T.M. Nutrient versus pollination limitation in Platanthera bifolia and Dactylorhiza incarnata (Orchidaceae). OIKOS, 2000, 89, 360–366.
- Sletvold, N.; Grindeland, J.M.; Ågren, J. Pollinator-mediated selection on floral display, spur length and flowering phenology in the deceptive orchid Dactylorhiza lapponica. New Phytol. 2010, 188, 385–392, . [CrossRef]
- Trunschke, J.; Sletvold, N.; Ågren, J. Interaction intensity and pollinator-mediated selection. New Phytol. 2017, 214, 1381–1389, . [CrossRef]
- Ostrowiecka, B.; Tałałaj, I.; Brzosko, E.; Jermakowicz, E.; Mirski, P.; Kostro-Ambroziak, A.; Mielczarek, Ł.; Lasoń, A.; Kupryjanowicz, J.; Kotowicz, J.; Wróblewska, A. Pollinators and visitors of the generalized food-deceptive orchid Dactylorhiza majalis in North-Eastern Poland; Biologia, 2019, 74, 1247–1257.
- Wróblewska, A.; Szczepaniak, L.; Bajguz, A.; Jędrzejczyk, I.; Tałałaj, I.; Ostrowiecka, B.; Brzosko, E.; Jermakowicz, E.; Mirski, P. Deceptive strategy in Dactylorhiza orchids: multidirectional evolution of floral chemistry. Ann. Bot. 2019, 123, 1005–1016, . [CrossRef]
- Wróblewska, A.; Ostrowiecka, B.; Brzosko, E.; Jermakowicz, E.; Tałałaj, I.; Mirski, P. The patterns of inbreeding depression in food-deceptive Dactylorhiza orchids. Front. Plant Sci. 2024, 15, 1244393, . [CrossRef]
- Wróblewska, A.; Ostrowiecka, B.; Kotowicz, J.; Jermakowicz, E.; Tałałaj, I.; Szefer, P. What are the drivers of female success in food-deceptive orchids? Ecology and Evolution, 2024b, 14 (4); 11233.
- Hedrén, M.; Nordström, S. Polymorphic populations of Dactylorhiza incarnata s.l. (Orchidaceae) on the Baltic island of Gotland, Morphology; habitat preference and genetic differentiation. Annals of Botany, 2009, 104 (3), 527–542.
- Vallius, E.; Salonen, V.; Kull, T. Factors of divergence in co-occurring varieties of Dactylorhiza incarnata (Orchidaceae). Plant Syst. Evol. 2004, 248, 177–189, . [CrossRef]
- Kindlmann, P.; Jersáková, J. Effect of floral display on reproductive success in terrestrial orchids. Folia Geobot. 2006, 41, 47–60, . [CrossRef]
- Tałałaj, I.; Kotowicz, J.; Brzosko, E.; Ostrowiecka, B.; Aleksandrowicz, O.; Wróblewska, A. Spontaneous caudicle reconfiguration in Dactylorhiza fuchsii, A new self-pollination mechanism for Orchideae; Plant Systematics and Evolution, 2019, 305, 269–280.
- Siudek, K. The role of pollinarium reconfiguration as the mechanism of selfing in Dactylorhiza majalis and Dactylorhiza incarnata. 2020. PhD dissertation. University of Bialystok; Faculty of Biology; pp. 68.
- Naczk, A.M.; Chybicki, I.J.; Ziętara, M.S. Genetic diversity of Dactylorhiza incarnata (Orchidaceae) in northern Poland. Acta Soc. Bot. Pol. 2016, 85, . [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van De Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 21, 4407–4414, doi:10.1093/nar/23.21.4407.
- Zhivotovsky, L.A. Estimating population structure in diploids with multilocus dominant DNA markers. Mol. Ecol. 1999, 8, 907–913, . [CrossRef]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 2002, 11, 139–151, . [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50.
- Hardy, O.J. Estimation of pairwise relatedness between individuals and characterization of isolation-by-distance processes using dominant genetic markers. Mol. Ecol. 2003, 12, 1577–1588, . [CrossRef]
- Manly, B. Randomization; Bootstrap and Monte Carlo Methods in Biology. 2007. Third Edition; Chapman and Hall/CRC; London.
- Chybicki, I.J.; Oleksa, A.; Burczyk, J. Increased inbreeding and strong kinship structure in Taxus baccata estimated from both AFLP and SSR data. Heredity 2011, 107, 589–600, . [CrossRef]
- Baskin, J.M.; Baskin, C.C. Inbreeding depression and the cost of inbreeding on seed germination. Seed Sci. Res. 2015, 25, 355–385, . [CrossRef]
- Hedrén M; Nordström S. High levels of genetic diversity in marginal populations of the marsh orchid Dactylorhiza majalis subsp. majalis. Nordic Journal of Botany, 2018, e01747.
- Filippov, E.G.; Andronova, E.V.; Kazlova, V.M. Genetic structure of the populations of Dactylorhiza ochroleuca and D. incarnata (Orchidaceae) in the area of their joint growth in Russia and Belarus. Russ. J. Genet. 2017, 53, 661–671, . [CrossRef]
- Balao, F.; Tannhäuser, M.; Lorenzo, M.T.; Hedrén, M.; Paun, O. Genetic differentiation and admixture between sibling allopolyploids in the Dactylorhiza majalis complex. Heredity 2015, 116, 351–361, . [CrossRef]
- Naczk, A.M.; Ziętara, M.S. Genetic diversity in Dactylorhiza majalis subsp. majalis populations (Orchidaceae) of northern Poland. Nordic Journal of Botany, 2019, e01989.
- Geremew, A.; Stiers, I.; Sierens, T.; Kefalew, A.; Triest, L. Clonal growth strategy; diversity and structure, A spatiotemporal response to sedimentation in tropical Cyperus papirus swamps. PLoS ONE, 2018, 13(1), e0190810.
- Niiniaho, J. The role of geitonogamy in the reproduction success ofa nectarless Dactylorhiza maculata (Orchidaceae). 2011. Dissertation; University of Jyväskylä.
- Eckert, C.G. Contributions of autogamy and geitonogamy to self-fertilization in a mass-flowering; clonal plant. Ecology, 2000, 81(2), 532–542.
- Hayashi, T.; Ayre, B.M.; Bohman, B.; Brown, G.R.; Reiter, N.; Phillips, R.D. Pollination by multiple species of nectar foraging Hymenoptera in Prasophyllum innubum, a critically endangered orchid of the Australian Alps. Aust. J. Bot. 2024, 72, . [CrossRef]
- Husband, B.C.; Schemske, D.W. Evolution of the magnitude and timing of inbreeding depression in plants. Evolution, 1996, 50, 54–70.
- Johnston, M.O.; Schoen, D.J. Correlated Evolution of Self-Fertilization and Inbreeding Depression: An Experimental Study of Nine Populations of Amsinckia (Boraginaceae). Evolution 1996, 50, 1478, . [CrossRef]
- Angeloni, F.; Ouborg, N.J.; Leimu, R. Meta-analysis on the association of population size and life history with inbreeding depression in plants. Biol. Conserv. 2010, 144, 35–43, . [CrossRef]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends in Ecology & Evolution, 2002, 17(5), 230-241.
Figure 1.
Localities of nine Dactylorhiza populations in north-eastern Poland. D. majalis (DM), KA, SKI, and SKII; D. incarnata var. incarnata (DI), ZB, MR, and RO; D. fuchsii (DF) CM, BR, and GR.
Figure 1.
Localities of nine Dactylorhiza populations in north-eastern Poland. D. majalis (DM), KA, SKI, and SKII; D. incarnata var. incarnata (DI), ZB, MR, and RO; D. fuchsii (DF) CM, BR, and GR.
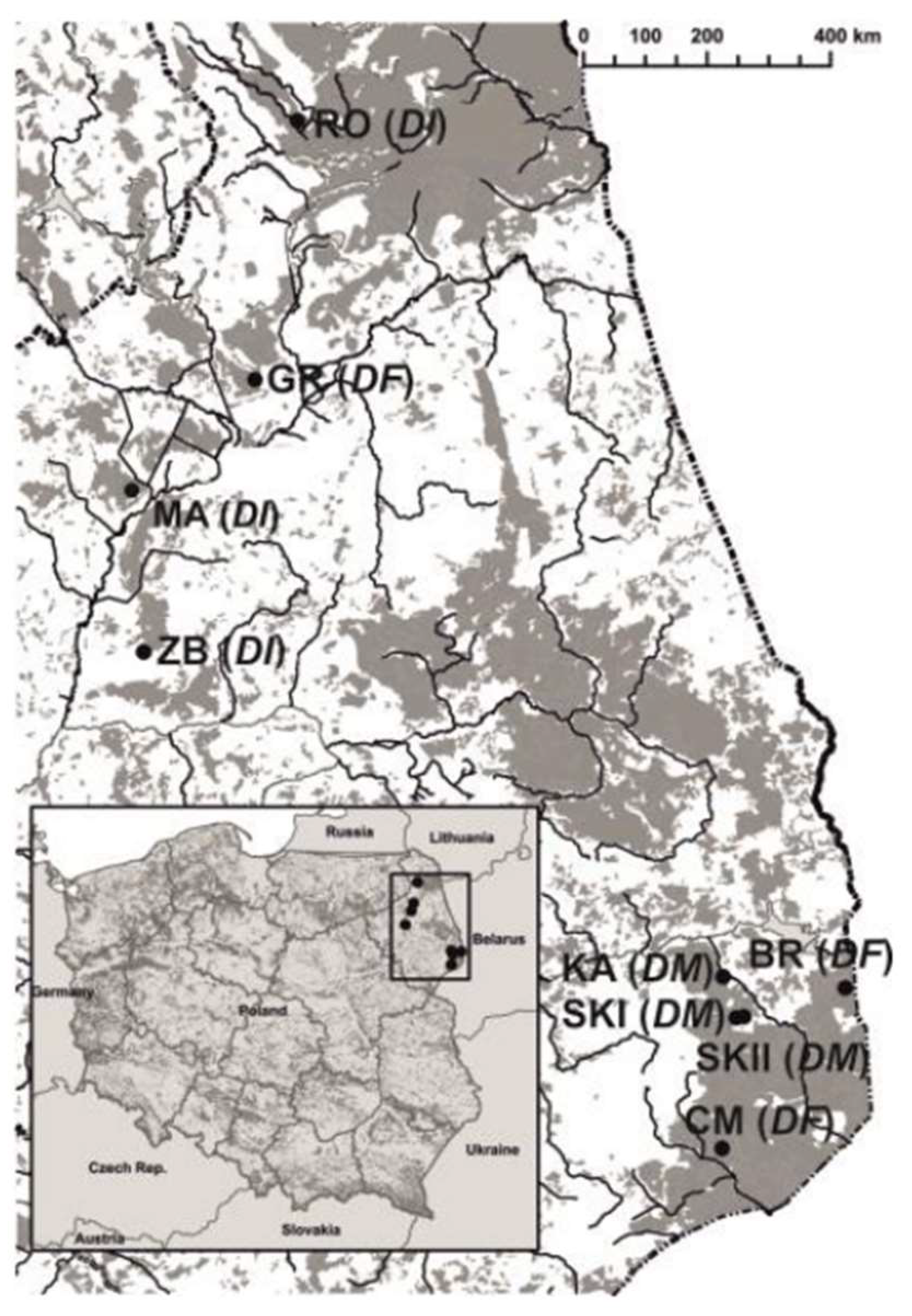
Figure 2.
Spatial correlograms for Dactylorhiza majalis, D. incarnata var. incarnata, and D. fuchsii populations with mean pairwise kinship coefficients (Fij) of distance classes for AFLPs around the hypothesis of random genetic structure obtained by permuting individual spatial locations as implemented in the SPAGeDi 1.4 [6]. The dotted lines indicate the 95% confidence intervals obtained from 10,000 permutations of genotypes). Codes of populations (KA, SKI, SKII, ZA, MR, RO, CM, BR, GR, see Table 1); * p < 0.05.
Figure 2.
Spatial correlograms for Dactylorhiza majalis, D. incarnata var. incarnata, and D. fuchsii populations with mean pairwise kinship coefficients (Fij) of distance classes for AFLPs around the hypothesis of random genetic structure obtained by permuting individual spatial locations as implemented in the SPAGeDi 1.4 [6]. The dotted lines indicate the 95% confidence intervals obtained from 10,000 permutations of genotypes). Codes of populations (KA, SKI, SKII, ZA, MR, RO, CM, BR, GR, see Table 1); * p < 0.05.
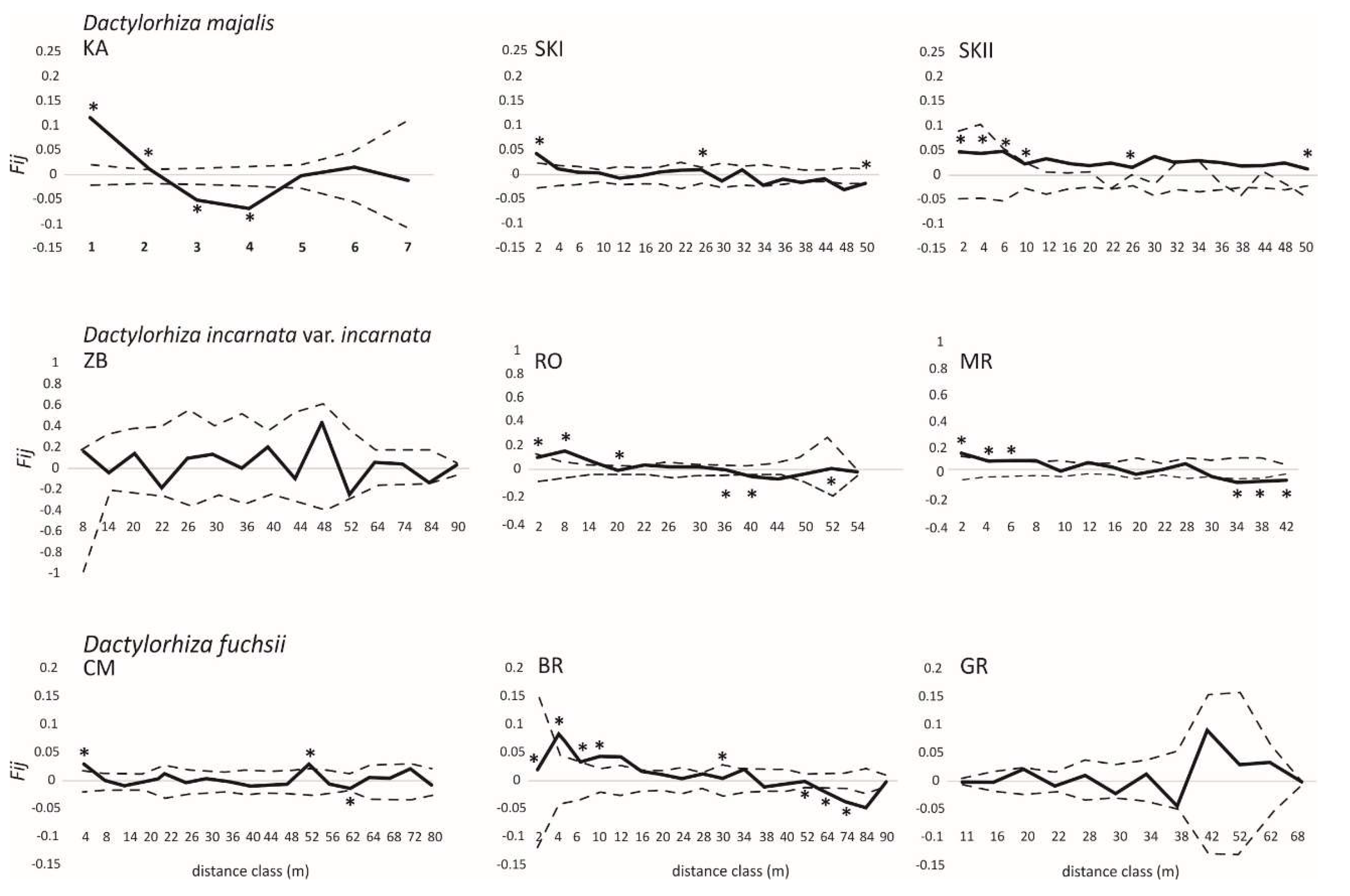

Table 1.
Locations of Dactylorhiza majalis (DM), D. incarnata var. incarnata (DI) and D. fuchsii (DF) populations in NE Poland and summary statistic of the genetic diversity and spatial genetic structure estimated using SPAGeDi 1.4 (Veckemans and Hardy 2004). N – number of AFLP samples, P% - frequency of polymorphic loci, H –Nei’s gene diversity, FIS – inbreeding coefficient, CI - the upper and lower 95% confidence interval values, Fij(1) – mean pairwise kinship coefficient among individuals at the first distance class, b1 - regression slope of pairwise kinship at the first distance, Sp – the intensity of FSGS according Veckemans and Hardy [6]. Vouchers specimens have been collected by Ada Wróblewska and deposited in the herbarium of the Faculty of Biology, University of Bialystok, Poland.
Table 1.
Locations of Dactylorhiza majalis (DM), D. incarnata var. incarnata (DI) and D. fuchsii (DF) populations in NE Poland and summary statistic of the genetic diversity and spatial genetic structure estimated using SPAGeDi 1.4 (Veckemans and Hardy 2004). N – number of AFLP samples, P% - frequency of polymorphic loci, H –Nei’s gene diversity, FIS – inbreeding coefficient, CI - the upper and lower 95% confidence interval values, Fij(1) – mean pairwise kinship coefficient among individuals at the first distance class, b1 - regression slope of pairwise kinship at the first distance, Sp – the intensity of FSGS according Veckemans and Hardy [6]. Vouchers specimens have been collected by Ada Wróblewska and deposited in the herbarium of the Faculty of Biology, University of Bialystok, Poland.
| Taxa | Population | GPS | N | P% | H | FIS (CI) | Fij(1) | b1 | Sp |
|---|---|---|---|---|---|---|---|---|---|
| DM | KA | 52°53’00’’N 23°40’29’’E |
49 | 62.2 | 0.205 | 0.293 (0.000-1.000) | 0.095* | -0.051* | 0.056 |
| SKI | 52°49’50’’N 23°43’10’’E |
59 | 59.6 | 0.205 | 0.312 (0.000-1.000) | 0.038* | -0.009* | 0.001 | |
| SKII | 52°49’50’’N 23°43’10’’E |
54 | 40.4 | 0.140 | 0.192 (0.000-1.000) | 0.071* | -0.021* | 0.022 | |
| DI | ZB | 53°29’02’’N 22°59’28’’E |
48 | 58.6 | 0.217 | 0.179 (0.101-0.284) | 0.008 | -0.002 | 0.0002 |
| RO | 53°54’39’’N 22°56’32’’E |
48 | 58.6 | 0.197 | 0.071 (0.022-0.149) | 0.224* | -0.055* | 0.063 | |
| MR | 53°47’25’’N 22°57’22’’E |
33 | 58.1 | 0.206 | 0.098 (0.032-0.218) | 0.092* | -0.037* | 0.041 | |
| DF | CM | 52°41’03’’N 23°39’07’’E |
58 | 68.4 | 0.234 | 0.113 (0.034-0.244) | 0.078* | -0.021* | 0.019 |
| BR | 52°50’59’’N 23°53’40’’E |
57 | 63.9 | 0.211 | 0.134 (0.068-0.226) | 0.084* | -0.026* | 0.028 | |
| GR | 53°60’68” E 22°84’68”N | 49 | 56.7 | 0.197 | 0.079 (0.024-0.169) | -0.008 | 0.002 | -0.0002 |
Almost all FST values were significant in each Dactylorhiza taxon, ranging from the lowest values in D. fuchsii and D. majalis (0.080 and 0.086, p < 0.05, permutation test), and the higher value (0.163, p < 0.05, permutation test) in D. incarnata var. incarnata (Table 2). The amount of molecular variance was highest within populations and was maintained at relatively higher and similar levels in D. majalis, D. incarnata var. incarnata, and D. fuchsii (AMOVA: 76.5%, 85.5%, and 86.6%; p < 0.001, respectively).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



