
Submitted:
05 December 2024
Posted:
09 December 2024
You are already at the latest version
Abstract
European beech (Fagus sylvatica L.) has a wide distribution range through Central and Western Europe, and is tolerant to a range of environmental factors and shade. Due to the wood quality and growth rate, there is increasing interest in this species in Northern Europe, which is further boosted by closer to nature forestry management practices, where shade tolerant species are essential. Latvia is located to the north of the natural range of European beech, but stands have been successfully established in the 19th century, using reproductive material of unknown origin. This study investigated the natural regeneration, genetic diversity and population structure, and provenance of Latvian F.sylvatica using nuclear microsatellite markers. Stands from two areas in western Latvia were analysed – Skede and Kaleti. Parent-offspring analyses did not identify a decrease in genetic diversity parameters in the naturally regenerated offspring, indicating that the introduced F.sylvatica populations have adapted to Latvian conditions, and natural regeneration has not decreased the genetic viability of these populations. Genetic diversity was low in Kaleti stand, probably as a result of the reproductive material used to establish this stand. Bayesian clustering analysis indicated that the most likely provenance of the Skede stand in Latvia was from southern Germany.
Keywords:
European beech
; parental assignment
; adaptation
; provenance
; microsatellite markers
; assisted migration
; climate change
1. Introduction
Rapid global climate changes are already affecting the distribution and composition of forest tree species. The potential distribution ranges of species are constrained by climate and are expected to shift toward higher latitudes and elevations as global warming intensifies [1,2] Trees have long life spans and delayed reproduction, and thus are not able to swiftly react to changing environmental conditions, as adaptation, migration and related mutation accumulation are long-term events [3]. Intraspecific genetic variation and local adaptation are essential for survival of populations growing at species distribution range limits [4], and research concerning local genetic diversity and species potential under current conditions, while also considering predicted changes in the future is essential [5]. Assisted migration is the deliberate movement of species or populations to accelerate natural range expansion as a part of climate change mitigation management processes [6]. It is considered as one way to maintain forest productivity and health under changing climactic conditions [7], and assessment of the potential benefits of assisted migration of forest tree species is becoming more relevant with regard to sustainable forest management and future planning [8]. The global scientific community is assessing potential risks and benefits of such actions. In this study, an existing case of assisted migration of European beech (Fagus sylvatica L.) in Latvia is investigated.
F.sylvatica is a deciduous, monoecious broad leaved tree species, that reaches reproductive age after 40-50 years [9]. It is mainly outcrossing, with self-pollination mostly resulting in empty seeds, and selfing rates are estimated to be less than 10 % [10,11]. F.sylvatica has a wide distribution range through Central and Western Europe [12], and is considered as a successful forest tree species because of tolerance to variable environmental factors and shade [13,14], and due to its ability to form both large single-species stands in suitable areas as well as productive admixed stands [15]. However, currently a large proportion of areas suitable for F.sylvatica are used for agriculture or are reforested with Scots pine (Pinus sylvestris L.) [16]. The potential use of selective cutting forestry management practices for F.sylvatica and the high quality of beech wood as well as good growth characteristics has led to growing interest in increasing F.sylvatica stand areas in Northern Europe [17,18]. The interest to use this species is further boosted by the intent to implement closer to nature forestry management practices, where shade tolerant species are essential [19].
Latvia is located beyond the northern natural range limit of F.sylvatica, and no natural introductions or migration of this species have been identified. During the 19th century, German foresters successfully introduced F.sylvatica in western Latvia, using germplasm of unknown origin [20], and in the last decades, natural regeneration in these stands has been observed [21]. In 1969, the F.sylvatica forest at Skede covered 11 ha, with the oldest trees ranging from 60 to 80 years old (the seedling count after a mast seed year was approximately 360000 ha-1 [21]. Tree ring widths of F.sylvatica in Latvia were not supressed, indicating adequate growth conditions under current climactic parameters [22]. The distribution of F.sylvatica is constrained by winter temperatures and summer precipitation [23]. However, the distribution range may be larger than currently delineated, and forest management practices in the past may have contributed to a reduction of the area where European beech is currently distributed [24]. Climate change projections predict the northward migration of European beech [25], and an increase in its competitiveness at northern range margins [26].
F.sylvatica populations been intensively studied in Central Europe using various marker types [9,11,27,28,29,30,31,32]. In general, isozymes and nuclear markers revealed high levels of diversity, and low differentiation between populations. Chloroplast marker analyses have revealed higher levels of population differentiation, indicating that gene flow by seed dispersal is relatively low [33,34]. SSR markers have been the most widely used markers system in studies of F.sylvatica [31]. They are also widely used in population genetics studies as they are codominant, highly polymorphic, and technically relatively easy to implement [35]. Studies utilising DNA markers have indicated that forest management techniques such as shelter wood systems have little or no impact on the genetic diversity of beech stands [36,37]. Climate change can be expected to have negative effects on beech stands, mainly in the southern part of its distribution range, due to temperature and water regime changes. However, it has been suggested that intra-population genetic diversity, gene flow, and phenotypic plasticity can mitigate these threats [38] and can be utilized to assist the shift of the distribution range of European beech northwards.
Studies of F.sylvatica have shown that it has strong family structure - up to 30 m radius [39], but spatial genetic structure can also be influenced by wind, gravity or bird and animal activity, and a 27% seed immigration rate has been reported in F.sylvatica stands in France [11]. Research has shown that up to 90% of F.sylvatica seeds are dispersed in a 25 m radius [21,40], but some studies have reported even more limited dispersal [11]. As seed dispersal distances are short, pollen flow ensures longer distance gene transfer, but these distances also can be limited in comparison with other broad leaved tree species, being approximately 42 meters [11].
The overall genetic diversity of F.sylvatica in Europe is reduced due to intensive forestry practices over centuries, and as high genetic diversity is vital for long term evolutionary and adaptive changes [41], an assessment of the current situation and conservation of existing genetic resources is important for the future. A sufficient level of genetic diversity in the parent population is also important to recommend the wide-scale use of locally adapted reproductive material [22] for propagation at the fringes of the current distribution range. A recent study in Lithuania revealed a significant association between phenology and stem quality of introduced European beech populations with significant implications for forest management practices, as planting of stands can disrupt spatial genetic structure and grouping of related individuals, thus enhancing genetic diversity [42]. European beech stands in Latvia have been studied from dendrochronological [43], ecological [44], and silvicultural [14] aspects. However, genetic analyses of these Latvian F.sylvatica stands have not been done. The aim of this study was to investigate the genetic structure and diversity of existing F.sylvatica stands in Latvia and to determine the proportion of mature F.sylvatica trees that are involved in regeneration to evaluate perspectives for F.sylvatica stands in Latvia propagated with locally adapted material. Parental-offspring analyses were conducted in two stands to assess the genetic aspects of natural regeneration processes. Genetic diversity and population structure was assessed in four Latvian F.sylvatica stands planted with imported reproductive material in the 19th century and the beginning of the 20th century. In addition, the potential provenance of the reproductive material introduced to Latvia was investigated by comparing them to F.sylvatica stands in Southern Germany, Poland and Sweden. Historical records indicate that the material used for establishing the F.sylvatica stands in Latvia was sourced from unknown areas in Germany [20]. Poland and Sweden are the closest countries to Latvia which have regions within the natural distribution range of F.sylvatica. A previous study from Lithuania, also outside of the natural distribution range, and immediately to the south of Latvia, indicated that at least one F. sylvatica stand was established using reproductive material from Poland [45]. The results of these analyses will assist in the management of the Latvian F.sylvatica stands, determining if natural regeneration processes are maintaining genetic diversity in subsequent generations. Assessment of the genetic diversity and population structure of mature F.sylvatica stands provides information for the efficient management of forest genetic resources. Identifying the origin of the reproductive material used to establish these Latvian stands forms the basis for an ecogeographical assessment of the long-range transfer of F. sylvatica to areas outside its current natural distribution and makes a valuable contribution to research on assisted migration.
2. Materials and Methods
2.1. Characteristics of Latvian F.sylvatica stands and sample collection
Samples for parental-offspring analysis were collected from two stands (Skede 20 and Kaleti), aged 112-118 years, located in the western part of Latvia. Sampling design was adapted from Vranckx et al. [46]. In the centre of each stand one circular sample plot with area 0.78 ha was established. All mature trees (potential parents/mothers) in a circular sample plot were sampled. Young trees (offspring) were sampled from a 20 x 20 m subplot in the centre of the circular plot. Leaves from young trees and woody tissue from mature trees were collected for DNA extraction. F.sylvatica was the dominant species in the Skede stand (57°15’N, 22°44’E), with admixture of several conifer and deciduous tree species, age of the trees was 112-118 years, area 2.44 ha, basal area 42 m2 ha -1. It is situated on flat terrain on fertile, well-drained clay soil, elevation is approximately 90 metres The Kaleti stand (56⁰36’N, 21⁰48’E) was dominated by F.sylvatica with some admixture of deciduous tree species. The age of the mature trees in the Kaleti stand was 112 years, stand area was 5.8 ha, basal area 36 m2 ha -1. It is situated on flat terrain, well-drained clay soil, elevation approximately 30 metres.
For genetic diversity and population structure analysis, samples from mature individuals only were collected from two additional stands within the larger Skede population – Skede 21 (43 individuals) and Skede 23 (42 individuals) (121-125 years old).
For the provenance analysis, fifty mature individuals from the Skede 23 stand were analysed with the same SSR markers as used previously to compare introduced European beech stands in Lithuania with potential source and reference populations in Germany, Poland and Sweden [45].
Locations of analysed F.sylvatica stands and populations are shown in Figure 1, and details provided in Supplementary file 1.
2.2. DNA extraction and genotyping
DNA was extracted from leaves and woody tissue using a CTAB buffer based method [47]. For parental-offspring and diversity and structure analysis of the Skede stands, genotyping was done using 18 nuclear SSR markers. Each forward primer was fluorescently labelled for detection using capillary electrophoresis. PCR reactions for the eleven SSR markers described by Lefèvre et al. (2012) (csolfagus_31, sfc_0036, csolfagus_06, csolfagus_19, csolfagus_29, EEU75_A_0, EJV8T_A_0, EMILY_A_0, ERHBI_A_0, DZ447_A_0, DE576_A_0) were carried out according to the published protocol. The PCR reactions for the seven SSR markers described in Pastorelli et al. (2003) (FS1-15, FS1-25, FS1-03, FS1-11, FS3-04, FS4-46, FCM5) were carried out in 20μL reactions consisting of 0.2 mM dNTPs, 2.5 mM MgCl2, 1.5 µl DNA solution, 1x Taq buffer and 1U of recombinant Taq DNA polymerase (Thermo Scientific). PCR conditions were: initial denaturation - 95 °C for 5 min, 30 cycles of 95 °C for 30 s, 52 °C for 45 s (56 °C for M2-30), and 72 °C for 60 s with a final extension step of 72 °C for 10 min. PCR products were separated by capillary electrophoresis using an ABI 3100XL capillary sequencer (Applied Biosystems) and genotyped using GeneMapper 4.0 software (Applied Biosystems).
For provenance and phylogeographic analysis, samples from the stand in Latvia were genotyped with the same ten SSR markers (FS1-15, FS3-04, csolfagus_31, csolfagus_19, DE576_A_0, MFC11, MFC5, MFC7, sfc_0036, MFS11) as in a previous phylogeographic study [45]. Genotype data from this previous study of the German, Polish and Swedish populations were compared to the genotypes of the Skede 23 stand.
Sequences of the primers used for PCR amplifications are provided in Supplementary file 2.
2.3. Data analysis
The possible presence of null alleles was determined using the program MICRO-CHECKER version 2.2 [48].
Null alleles are genetic variants where PCR amplification fails due to mutations, deletions in primer binding sites, or large indels, reducing apparent genetic diversity and complicating parent-offspring analyses.
Genetic diversity parameters, as well as pairwise relatedness within each candidate parental and offspring population (using the Ritland estimator [49]) were calculated using GenAlEx [50]. Principal coordinates analysis (PCoA) of pairwise Nei genetic distances was also done using GenAlEx. The genetic diversity parameters calculated were the number of alleles (Na), the effective number of alleles (Ne), information index (I), observed heterozygosity (Ho), expected heterozygosity (He), and fixation index (F). Na is the total number of alleles, and Ne is the number of equally frequent alleles needed for the same expected heterozygosity. The Information Index, similar to the Shannon-Weaver index, is an unbounded alternative to PIC or expected heterozygosity. Observed heterozygosity is calculated from codominant markers, while expected heterozygosity measures genetic variation. The Fixation Index (-1 to +1) indicates inbreeding or null alleles (positive) and excess heterozygosity (negative).
Offspring and mother tree groups were analysed separately in each population, and loci with potential null alleles in both offspring and mother tree groups were excluded from pedigree analysis. Assignment of parentage was done using the Cervus software package [51], which uses likelihood ratios to assign parentage, and simulations to determine the level of confidence in parental assignments. As F.sylvatica is a monoecious species, the candidate mothers and fathers were the same individuals. Parental assignment was done for one parent (i.e. maternity or paternity), as well as for parent pairs, with a confidence level of >0.95. Identification of full- and half-sib families was done using the COLONY 2.0.5 software package [52], which implements a maximum likelihood to estimate full- and half-sib relationships. The allelic dropout rate and the rate of other kinds of genotyping errors (e.g. false alleles, miscalling) of each marker were both set at 0.005.
Bayesian analysis can be used to infer the genetic clustering of individuals by grouping individuals with similar patterns of genetic variation [53]. No prior grouping or population information about the individuals to be analysed is required, but can be provided (e.g. sampling location, ecology or other characteristics) to shape the clustering analysis. STRUCTURE 2.3.4 was used to determine clustering of the populations using a 150000 burnin period, followed by 300000 MCMC steps, K was determined for 1 to 15 with 10 runs for each value of K. The optimal value of K was determined using the ΔK method [54] using StructureSelector [55]. In addition, to check for correlations between pairwise genetic and geographic distances, or isolation by distance (IBD), the pairwise genetic and geographic distance matrices of individuals from the populations within the natural distribution range of F.sylvatica (i.e. the German, Polish and Swedish populations) were compared using a Mantel test with 999 permutations in the GenAlEx software package.
3. Results
Parental-offspring analyses
To increase sensitivity and accuracy, samples with missing genotypes at more than three loci were discarded from further analysis. Possible null alleles were identified in each population and group. Loci with potential null alleles that were identified in both mother and offspring groups were not utilised for further pedigree analysis to avoid false exclusion of potential kin relationships. In the Skede 20 population, three loci were excluded (FS1-15, FS4-46, FSM5), and two loci were excluded in the Kaleti population (FS4-46, FSM5). In addition, four pairs of individuals in the Kaleti offspring group had identical multilocus genotypes, therefore one of the individuals in each pair was excluded from further analysis. The sibling probability of identity (PIsibs), which provides an estimation of the average probability that two siblings will by chance have the same multilocus genotype, was 2.4x10-5 (using all analysed loci). The more likely explanation of these identical multilocus genotypes is these individuals were clones from root suckers, which has also been observed while assessing regeneration in other sites in Latvia [21].
After exclusion of individuals and loci as described above, pedigree analysis in the Skede population was done with 15 loci and 63 potential parent trees and 101 offspring. In the Kaleti population, 16 loci were retained, and 45 potential parent trees and 106 offspring.
Skede 20 stand parental-offspring analysis
Of the 101 offspring in the Skede stand, 54 were assigned to single parent trees from the collected mature individuals. A total of 20 parent trees were identified from the 63 candidate parent individuals. For parent pair analysis, 15 offspring individuals were assigned to parent pairs from the sampled candidate parent individuals. In total, combining the assigned parents from the parental assignment analysis, 22 parent trees were identified from the 63 candidate parental individuals, indicating that approximately 30% of the parents contributed to the regeneration of the sampled offspring plot. Of the 20 parent trees assigned with a confidence level of >0.95, 8 were assigned to only one offspring individual, while the remaining 12 parent trees were assigned to between 2-13 offspring individuals (Figure 2).
The Colony software analysis identified 8 full sib families (p>0.90), most with two members, but two families were identified with 4 and one with 5 individuals. An additional 4 potential full-sib families were identified, with p<0.90.
The origin of the reproductive material utilised to establish the two stands was unknown, therefore a kinship analysis of the parent trees was also done. In the Skede stand, of the 63 parent trees analysed, only two pairs of individuals were inferred as being full-sibs (p>0.95). In addition, most likely number of both paternal and maternal half-sib families was between 58-63, with the majority of likely half-sib configurations falling within that range (>0.95 of both inferred paternal and maternal half-sib families).
The pairwise relatedness in the Skede population candidate parent group and the offspring group were calculated, and there was no significant difference in the average pairwise relatedness (-0.001 in both groups).
Kaleti stand parental-offspring analysis
Of the 106 offspring in the Kaleti stand, 44 were assigned to single parent trees from the collected mature individuals. A total of 18 parent trees were identified from the 45 candidate parent individuals. For parent pair analysis, 6 offspring individuals were assigned to parent pairs from the sampled mature individuals. In total, combining the assigned parents from the parental assignment analysis, 22 parent trees were identified, indicating that approximately 50% of the parents contributed to the regeneration of the sampled offspring plot. Of the 18 parent trees assigned with a confidence level of >0.95, 10 were assigned to only one offspring individual, while the remaining 8 parent trees were assigned to between 2-11 offspring individuals (Figure 3).
The Colony software analysis identified 4 full sib families (p>0.90), containing 2, 3, 6 and 14 individuals). An additional 6 potential full-sib families were identified, with p<0.90.
Kinship analysis of the parent trees in the Kaleti stand revealed a higher level of relatedness among them. Two full-sib families with more than two members (p>0.95) were identified, consisting of two and seven members. However, a large full-sib family of 15 members was inferred with a p-value of 0.931. The most likely number of full-sib families (including families with only one inferred member) was 15-19 (0.985 of inferred full sib families). In addition, most likely number of both paternal and maternal half-sib families was between 12-19, with the majority of likely half-sib configurations falling within that range (>0.95 of both inferred paternal and maternal half-sib families).
The pairwise relatedness in the Kaleti population candidate parent group and the offspring group were calculated, and the average pairwise relatedness in the candidate parent group (0.012) was significantly higher than in the offspring group (0.000).
Comparison of genetic diversity between the parent and offspring groups
Genetic diversity indicators (number of alleles, effective number of alleles, expected heterozygosity) were similar between the parental and offspring groups in each analysed stand. Unique (private) alleles were found in each parent and offspring group in both stands. The unique alleles in the offspring group are probably due to pollen flow from outside the sample plots. The genetic diversity was lower in the Kaleti population, in comparison to the Skede 20 population (Table 1, Figure 4). While the origin of the reproductive material utilised to establish these stands is not known, kinship and pairwise relatedness analysis indicated a higher degree of relatedness between the parental trees sampled from the Kaleti stand.
Genetic diversity and structure of mature stands
Comparison of the mature individuals from four Latvian stands (Skede_20, Skede_21, Skede_23 and Kaleti), revealed that the number of alleles, effective number of alleles and information index was lower in the Kaleti stand. However, the observed heterozygosity was highest in the Kaleti stand, and the fixation (inbreeding) index was negative for the Kaleti stand, but was positive for the three Skede stands analysed (Table 2).
The Kaleti stand was genetically differentiated from the three Skede stands. Principal Coordinates Analysis (PCoA) of the pairwise Nei genetic distances is shown in Figure 5. Pairwise Nei genetic distance values between the three Skede stands were 0.028 (Skede_20 – Skede_21), 0.017 (Skede_20 – Skede_23) and 0.031 (Skede_21 – Skede_23). The pairwise Nei genetic distance values between the Kaleti stand and the Skede_20, Skede_21 and Skede_23 stands were 0.213, 0.231 and 0.220, respectively. Pairwise Fst values between the three Skede stands were 0.006 (Skede_20 – Skede_21), 0.004 (Skede_20 – Skede_23) and 0.007 (Skede_21 – Skede_23). The pairwise Fst values between the Kaleti stand and the Skede_20, Skede_21 and Skede_23 stands were 0.047, 0.052 and 0.051, respectively.
Provenance of the Skede stand
Bayesian clustering analysis using the STRUCTURE software package of the Skede 23 stand and F.sylvatica stands from Southern Germany, Poland and Sweden was done using the Structure software program. The most likely number of genetic clusters determined by the deltaK method was eight, but there was also a high deltaK value for K=4 (Figure 6)
The coefficients of membership of each of the predefined populations for K=8 and K=4 are shown in Figure 7. The K=4 clustering broadly divided the analysed populations according to geography, with the populations from southern Germany in one group, together with the Latvian population. The populations from Poland were more heterogeneously clustered into three groups, with the exception of the south east populations, which were in one group. The populations with the highest similarity to the Latvian population are the populations from southern Germany, in particular the DE03_BA, DE_K_BA and DE09_OR populations for the K=8 clustering. Comparison of pairwise genetic and geographic distance matrices at both the individual and population level of the populations within the natural distribution range of F.sylvatica (i.e. the German, Polish and Swedish populations) indicated a significant positive correlation (p=0.001 for both individuals and population comparisons). This genetic isolation by distance is not surprising, given the large geographic scale of the analysed populations. However, long-range gene dispersal is probably effective in F.sylvatica, as the analysed Polish population are less structured. The results of this clustering analysis clearly indicate that the analysed Skede stand was established with germplasm originating from southern Germany, rather than the more geographically closer Polish F.sylvatica populations.
This clustering was confirmed by running STRUCTURE analysis on a subset containing only the Skede (LAT) and the southern German populations (allele frequencies correlated, no admixture model, burn in period of 15000 repetitions, 30000 MCMC repetitions after burn in, K limited to 2 groups, 10 iterations. When the number of clusters is set to two, the similarity of the Skede (LAT) population with the DE03_BA, DE_K_BA and DE09_OR populations is clearly defined (Figure 8).
4. Discussion
Based on the experimental design, and previous reports on seed dispersal distance in Fagus, it could be expected that a high proportion the sampled candidate parent group would be the parents of the sampled offspring group in each stand. In the Skede stand, 54 offspring of a total of 101 sampled offspring were assigned to at least one parent, while in the Kaleti stand 44 of 106 offspring were assigned to at least one parent. These assignments were at a high confidence level (>0.95), and with less stringent conditions, a majority of the offspring from each stand were assigned to sampled parents. The lower level of assigned parents in the Kaleti stand was also due to the lower level of genetic diversity in this stand, which meant that the parental assignments were with a lower confidence level compared to the Skede stand. At a confidence level of >0.80, 73 of 101 Skede offspring were assigned to parents, and 79 of 106 Kaleti offspring. The unassigned offspring could be attributed to seed dispersal from outside the parental sample plots.
As the parental stands were established using imported reproductive material, no spatial genetic structure (SGS) would be expected in the mature individuals. However, further investigation of SGS in the offspring generation could provide additional information about regeneration processes in these introduced stands. Significant SGS has been reported to develop in beech as soon as after one generation, but that pollen from outside of the local forest stand can decrease the degree of SGS in subsequent generations [56]. Given that there are no F.sylvatica stands surrounding these stands established in Latvia using imported reproductive material, and that the genetic base of the seeds used for stand establishment is narrow, high SGS could be expected in the naturally regenerated offspring generations. This information would be useful for management of these F.sylvatica stands in Latvia.
The Skede parent population did not exhibit a high degree of relatedness, and the offspring were assigned to approximately 30% of the candidate parents sampled. This could be due to seed dispersal or exclusion of parentage due the presence of null alleles (even though loci with potential null alleles were excluded). The Kaleti parent population was structured, and showed extensive relatedness. This is probably due to the reproductive material utilised. Nevertheless, the offspring group did not show higher levels of relatedness or lower genetic diversity than the parent group. The seed source and details of the establishment of the Kaleti stand are not known. One possibility is that seeds from a small number of individuals were used leading to the higher relatedness between the analysed mature individuals. The higher observed heterozygosity in the could be a result of the low selfing rate in F.sylvatica [11,57] among the limited number of individuals from which the seeds were sampled [58].
In the Skede stand, approximately 30% of the sampled parents contributed to the regeneration of the sampled offspring plot, while in the Kaleti stand, approximately 50% of the sampled parents were identified as parents of the sampled offspring. However, the genetic diversity parameters of the offspring were similar (in Skede) or higher (in Kaleti) than those of the sampled mature trees. Further investigations are required to determine if the parents who contribute to the next generation varies between years due to various factors, e.g. flowering time, seed set etc. However, from a forest management perspective, these results indicate that genetic diversity is being maintained via natural regeneration processes.
The differences in population structure and diversity between the parental populations in the Kaleti and Skede stands are probably due to the methods used to collect seed (or seedlings) for stand establishment. A previous study investigated the provenance and population structure of mature F.sylvatica stands in Lithuania [45]. Lithuania is also outside of the northern limit of the F.sylvatica distribution in Europe, and similarly to the situation in Latvia, beech stands have been established in the previous centuries using imported seed material, but their origin and methodologies are not known [23]. The Lithuanian study [45] found high allelic diversity in the mature beech stands, but also high inbreeding levels, and the authors suggested that this could be due to the collection of seeds from a number of different, and genetically differentiated beech stands, but from only a few, high-quality individuals from each stand. These results are similar to the results from the Skede stand, which had high allelic diversity parameters, but had higher inbreeding levels compared to the Kaleti stand and the Skede offspring. The information about genetic diversity and population structure can be used to improve forest genetic resource management strategies. The three analysed Skede stands are genetically very similar, and were probably established with the same reproductive material. Further investigation of the Kaleti stand is needed to determine quality and other parameters. While the genetic diversity is low in this stand, which was probably established using a narrow range or germplasm, the criteria used for selection of this material are not known, and perhaps this stand contains trees with unique traits. This can inform the selection of genetic resource stands, either for in situ or ex situ conservation approaches.
Bayesian clustering analysis indicated that the most likely provenance of the Skede stand in Latvia was from southern Germany. This is in agreement with results from a study in Lithuania [45], where Lithuanian F.sylvatica populations were compared to the same southern German, Polish and Swedish populations as in this study. Five of the ten analysed Lithuanian populations also clustered with the DE03_BA, DE_K_BA and DE09_OR populations. Of these five Lithuanian populations, three were forest stands, and two were manor parks. This suggests that the reproductive material introduced into the Baltic states in the 19th century was largely of southern German origin, and possibly collected from the same or similar sources for introduction into Latvia and Lithuania. Some of the other analysed Lithuanian populations also clustered with the Polish populations, indicating that in some cases, reproductive material from Poland was also in introduced to Lithuania. In Latvia, the Skede population is the only large and well-established F.sylvatica population in Latvia. The Kaleti population, located in south-western Latvia close to the Lithuanian border is small, and as shown in this study, likely to have been established with reproductive material from a very limited number of individuals. Probably due to this founder effect, the Kaleti population was genetically differentiated from all other F.sylvatica populations analysed in this study, including the Skede population, and identifying a provenance region for the Kaleti population was not possible. The determination of the provenance of the F.sylvatica material introduced into Latvia approximately 100 years ago enables an assessment of the long-range transfer of germplasm outside of the natural distribution of this species. The successful survival and reproduction of this material indicates that these long-range transfers can result in the establishment of viable forest stands in the long-term. This study provides genetic data on the introduced material used to establish the European beech stands in Latvia. In terms of climate change predictions for north-eastern Europe, beech is sensitive to drought [59], but the response of European beech provenances to environmental constraints varies, and the phenotypic plasticity and evolutionary adaptability of this species may not be fully appreciated [24]. However, genetic diversity is the foundation of adaptation in populations, and so this information is crucial for the long-term management of tree genetic resources. More detailed analyses of growth rates (e.g. by using dendrochronological (tree ring) analysis), can provide additional information to the responses of this introduced germplasm to long-term local climactic and environmental conditions. This can contribute data for modelling and other analyses to inform decisions about assisted migration in the future.
This study provides a significant contribution to understanding the genetic diversity, population structure, and provenance of Fagus sylvatica stands in Latvia. The findings lay a crucial foundation for future research into fine-scale spatial genetic structure, which will enhance strategies for managing these introduced stands. Additionally, integrating growth and quality assessments with genetic analyses offers valuable insights into the genetic architecture and control of silviculturally important traits. This approach also opens new opportunities for selective breeding, utilizing the germplasm introduced to Latvia over a century ago, while advancing the potential for selective genomics and assisted migration strategies.
Author Contributions
Conceptualization, D.E.R., D.D. and Ā.J.; methodology, D.E.R., D.D., R.K., L.J., M.K. and Ā.J.; formal analysis, D.E.R., D.D., R.K. and M.K.; resources, D.D., R.K., L.J. and M.K.; writing—original draft preparation, D.E.R.; writing—review and editing, D.E.R., D.D., M.K., Ā.R.; project administration, Ā.J.; funding acquisition, Ā.J. All authors have read and agreed to the published version of the manuscript.
Funding
Assessment of Latvian beech stands was carried out in the projects ‘Support for Forest Tree Seed Production’ (funded by Latvia’s State Forests (LVM)) and ‘Climate smart solutions for enhanced biodiversity through increased presence of broadleaved trees in landscape FORESTBAND’ (INTERREG, No EE-LV00188).
Data Availability Statement
Genotyping data is provided in Supplementary file 3.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation During the 20th Century. Science. 2008, 320, 1768–1771. [CrossRef]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [CrossRef]
- Cortés, A.J.; Restrepo-Montoya, M.; Bedoya-Canas, L.E. Modern Strategies to Assess and Breed Forest Tree Adaptation to Changing Climate. Front. Plant Sci. 2020, 11. [CrossRef]
- Kreyling, J.; Buhk, C.; Backhaus, S.; Hallinger, M.; Huber, G.; Huber, L.; Jentsch, A.; Konnert, M.; Thiel, D.; Wilmking, M.; et al. Local adaptations to frost in marginal and central populations of the dominant forest tree Fagus sylvatica L. as affected by temperature and extreme drought in common garden experiments. Ecol. Evol. 2014, 4, 594–605. [CrossRef]
- Graudal, L.; Aravanopoulos, F.; Bennadji, Z.; Changtragoon, S.; Fady, B.; Kjær, E.D.; Loo, J.; Ramamonjisoa, L.; Vendramin, G.G. Global to local genetic diversity indicators of evolutionary potential in tree species within and outside forests. For. Ecol. Manage. 2014, 333, 35–51. [CrossRef]
- Williams, M.I.; Dumroese, R.K. Preparing for Climate Change: Forestry and Assisted Migration. J. For. 2013, 111, 287–297. [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Aubin, I.; Beardmore, T.; Beaulieu, J.; Iverson, L.; O’Neill, G.A.; Winder, R.S.; Ste-Marie, C. Placing Forestry in the Assisted Migration Debate. Bioscience 2012, 62, 835–842. [CrossRef]
- Ste-Marie, C. Adapting Sustainable Forest Management to Climate Change: A Review of Assisted Tree Migration and its Potential Role in Adapting Sustainable Forest Management to Climate Change. 2014, 28.
- Magri, D. Patterns of post-glacial spread and the extent of glacial refugia of European beech (Fagus sylvatica). J. Biogeogr. 2008, 35, 450–463. [CrossRef]
- Merzeau, D.; Comps, B.; Thiébaut, B.; Cuguen, J.; Letouzey, J. Genetic structure of natural stands of Fagus sylvatica L. (beech). Heredity (Edinb). 1994, 72, 269–277. [CrossRef]
- Oddou-Muratorio, S.; Bontemps, A.; Klein, E.K.; Chybicki, I.; Vendramin, G.G.; Suyama, Y. Comparison of direct and indirect genetic methods for estimating seed and pollen dispersal in Fagus sylvatica and Fagus crenata. For. Ecol. Manage. 2010, 259, 2151–2159. [CrossRef]
- von Wühlisch, G. Technical Guidelines for genetic conservation and use for European beech (Fagus sylvatica).; Bioversity International: Rome, 2008; ISBN 978-92-9043-787-1.
- Leuschner, C.; Meier, I.C.; Hertel, D. On the niche breadth of Fagus sylvatica: Soil nutrient status in 50 Central European beech stands on a broad range of bedrock types. Ann. For. Sci. 2006, 63, 355–368. [CrossRef]
- Matisone, I.; Jaunslaviete, I.; Adamovičs, A.; Matisons, R.; Jansons, Ā. Response of underplanted European beech to shelterwood thinning at the northeasternmost forpost plantation in Europe. New For. 2024, 55, 1267–1281. [CrossRef]
- Pretzsch, H.; del Río, M.; Ammer, C.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; et al. Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2015, 134, 927–947. [CrossRef]
- Peters, R. Beech Forests; Springer Netherlands: Dordrecht, 1997; ISBN 978-90-481-4824-0.
- Gailis, A.; Šmaukstelis, E. Oak and beech resources in Latvia. In First EUFORGEN Meeting on Social Broadleaves, Bordeaux, France, 23–25 October 1997; Turok, J., Kremer, A., De Vries, S.M.G., Eds.; International Plant Genetic Resources Institute: Rome, 1998; pp. 121–126.
- Matisons, R.; Šņepsts, G.; Puriņa, L.; Donis, J.; Jansons, Ā. Dominant height growth of European beech at the northeasternmost stands in Europe. Silva Fenn. 2018, 52. [CrossRef]
- 2023; European Commission Guidelines on closer-to-nature forest management; Publications Office of the European Union, 2023;
- Dreimanis, A. Dižskābaržu audžu ražība Šķēdes mežu novadā [Productivity of European beech in Šķede forest district]. LLU Raksti 2006, 16, 97–100.
- Puriņa, L.; Neimane, U.; Džeriņa, B.; Jansons, Ā. Eiropas dižskābarža (Fagus sylvatica L.) atjaunošanos ietekmējošie faktori [Factors affecting regeneration of European beech (Fagus sylvatica L.)]. Mežzinātne 2013, 27, 67–76.
- Jansons, Ā.; Matisons, R.; Puriņa, L.; Neimane, U.; Jansons, J. Relationships between climatic variables and Tree-Ring width of European beech and European larch growing outside of their natural distribution area. Silva Fenn. 2015, 49, 1–8. [CrossRef]
- Augustaitis, A.; Kliučius, A.; Marozas, V.; Pilkauskas, M.; Augustaitiene, I.; Vitas, A.; Staszewski, T.; Jansons, A.; Dreimanis, A. Sensitivity of European beech trees to unfavorable environmental factors on the edge and outside of their distribution range in northeastern Europe. iForest - Biogeosciences For. 2016, 9, 259–269. [CrossRef]
- Bolte, A.; Czajkowski, T.; Kompa, T. The north-eastern distribution range of European beech - A review. Forestry 2007, 80, 413–429. [CrossRef]
- Buras, A.; Menzel, A. Projecting Tree Species Composition Changes of European Forests for 2061–2090 Under RCP 4.5 and RCP 8.5 Scenarios. Front. Plant Sci. 2019, 9. [CrossRef]
- Bolte, A.; Hilbrig, L.; Grundmann, B.; Kampf, F.; Brunet, J.; Roloff, A. Climate change impacts on stand structure and competitive interactions in a southern Swedish spruce–beech forest. Eur. J. For. Res. 2010, 129, 261–276. [CrossRef]
- Gallois, A.; Audran, J.C.; Burrus, M. Assessment of genetic relationships and population discrimination among Fagus sylvatica L. by RAPD. Theor. Appl. Genet. 1998, 97, 211–219. [CrossRef]
- Pastorelli, R.; Smulders, M.J.M.; Van’t Westende, W.P.C.; Vosman, B.; Giannini, R.; Vettori, C.; Vendramin, G.G. Characterization of microsatellite markers in Fagus sylvatica L. and Fagus orientalis Lipsky. Mol. Ecol. Notes 2003, 3, 76–78.
- Bilela, S.; Dounavi, A.; Fussi, B.; Konnert, M.; Holst, J.; Mayer, H.; Rennenberg, H.; Simon, J. Natural regeneration of Fagus sylvatica L. adapts with maturation to warmer and drier microclimatic conditions. For. Ecol. Manage. 2012, 275, 60–67. [CrossRef]
- Lefèvre, S.; Wagner, S.; Petit, R.J.; De Lafontaine, G. Multiplexed microsatellite markers for genetic studies of beech. Mol. Ecol. Resour. 2012, 12, 484–491. [CrossRef]
- Stefanini, C.; Csilléry, K.; Ulaszewski, B.; Burczyk, J.; Schaepman, M.E.; Schuman, M.C. A novel synthesis of two decades of microsatellite studies on European beech reveals decreasing genetic diversity from glacial refugia. Tree Genet. Genomes 2023, 19, 3. [CrossRef]
- Kempf, M.; Konnert, M. Distribution of genetic diversity in Fagus sylvatica at the north-eastern edge of the natural range. Silva Fenn. 2016, 50, 1–17. [CrossRef]
- Demesure, B.; Comps, B.; Petit, R.J. Chloroplast DNA Phylogeography of the Common Beech (Fagus sylvatica L.) in Europe. Evolution (N. Y). 1996, 50, 2515. [CrossRef]
- Vettori, C.; Vendramin, G.G.; Anzidei, M.; Pastorelli, R.; Paffetti, D.; Giannini, R. Geographic distribution of chloroplast variation in Italian populations of beech (Fagus sylvatica L.). Theor. Appl. Genet. 2004, 109, 1–9. [CrossRef]
- Ramesh, P.; Mallikarjuna, G.; Sameena, S.; Kumar, A.; Gurulakshmi, K.; Reddy, B.V.; Reddy, P.C.O.; Sekhar, A.C. Advancements in molecular marker technologies and their applications in diversity studies. J. Biosci. 2020, 45, 123. [CrossRef]
- Buiteveld, J.; Vendramin, G.G.; Leonardi, S.; Kamer, K.; Geburek, T. Genetic diversity and differentiation in European beech (Fagus sylvatica L.) stands varying in management history. For. Ecol. Manage. 2007, 247, 98–106. [CrossRef]
- Westergren, M.; Bozic, G.; Ferreira, A.; Kraigher, H. Insignificant effect of management using irregular shelterwood system on the genetic diversity of European beech (Fagus sylvatica L.): A case study of managed stand and old growth forest in Slovenia. For. Ecol. Manage. 2015, 335, 51–59. [CrossRef]
- Frank, A.; Pluess, A.R.; Howe, G.T.; Sperisen, C.; Heiri, C. Quantitative genetic differentiation and phenotypic plasticity of European beech in a heterogeneous landscape: Indications for past climate adaptation. Perspect. Plant Ecol. Evol. Syst. 2017, 26, 1–13. [CrossRef]
- Vornam, B.; Decarli, N.; Gailing, O. Spatial Distribution of Genetic Variation in a Natural Beech Stand (Fagus sylvaticaL.) Based on Microsatellite Markers. Conserv. Genet. 2004, 5, 561–570. [CrossRef]
- Kramer, K.; Buiteveld, J.; Forstreuter, M.; Geburek, T.; Leonardi, S.; Menozzi, P.; Povillon, F.; Schelhaas, M.J.; Teissier du Cros, E.; Vendramin, G.G.; et al. Bridging the gap between ecophysiological and genetic knowledge to assess the adaptive potential of European beech. Ecol. Modell. 2008, 216, 333–353. [CrossRef]
- Rajendra, K.C.; Seifert, S.; Prinz, K.; Gailing, O.; Finkeldey, R. Subtle human impacts on neutral genetic diversity and spatial patterns of genetic variation in European beech (Fagus sylvatica). For. Ecol. Manage. 2014, 319, 138–149. [CrossRef]
- Kembrytė, R.; Danusevičius, D.; Baliuckas, V.; Buchovska, J. Phenology Is Associated with Genetic and Stem Morphotype Variation in European Beech (Fagus sylvatica L.) Stands. Forests 2022, 13, 664. [CrossRef]
- Matisons, R.; Puriņa, L.; Adamovičs, A.; Robalte, L.; Jansons, Ā. European beech in its northeasternmost stands in Europe: Varying climate-growth relationships among generations and diameter classes. Dendrochronologia 2017, 45, 123–131. [CrossRef]
- Jansone, D.; Matisons, R.; Kārše, V.; Bāders, E.; Kaupe, D.; Jansons, Ā. Structural Heterogeneity of European Beech (Fagus sylvatica L.) Stands at Its Northernmost Limits. Sustainability 2023, 15, 14681. [CrossRef]
- Kembrytė, R.; Danusevičius, D.; Buchovska, J.; Baliuckas, V.; Kavaliauskas, D.; Fussi, B.; Kempf, M. DNA-based tracking of historical introductions of forest trees: the case of European beech (Fagus sylvatica L.) in Lithuania. Eur. J. For. Res. 2021, 140, 435–449. [CrossRef]
- Vranckx, G.; Jacquemyn, H.; Mergeay, J.; Cox, K.; Kint, V.; Muys, B.; Honnay, O. Transmission of genetic variation from the adult generation to naturally established seedling cohorts in small forest stands of pedunculate oak (Quercus robur L.). For. Ecol. Manage. 2014, 312, 19–27. [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Report. 1997. [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [CrossRef]
- Ritland, K. Estimators for pairwise relatedness and individual inbreeding coefficients. Genet. Res. 1996, 67, 175–185. [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program cervus accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [CrossRef]
- Jones, O.R.; Wang, J. COLONY: A program for parentage and sibship inference from multilocus genotype data. Mol. Ecol. Resour. 2010, 10, 551–555. [CrossRef]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, Á.; Lareu, M. V. An overview of STRUCTURE: applications, parameter settings, and supporting software. Front. Genet. 2013, 4. [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [CrossRef]
- Li, Y.; Liu, J. StructureSelector : A web-based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 2018, 18, 176–177. [CrossRef]
- Sandurska, E.; Ulaszewski, B.; Meyza, K.; Sztupecka, E.; Burczyk, J. Factors determining fine-scale spatial genetic structure within coexisting populations of common beech (Fagus sylvatica L.), pedunculate oak (Quercus robur L.), and sessile oak (Q. petraea (Matt.) Liebl.). Ann. For. Sci. 2024, 81, 3. [CrossRef]
- Merzeau, D.; Comps, B.; Thiebaut, B.; Letouzey, J. Estimation of Fagus sylvatica L mating system parameters in natural populations. Ann. des Sci. For. 1994, 51, 163–173. [CrossRef]
- Balloux, F. Heterozygote excess in small populations and the heterozygote-excess effective population size. Evolution (N. Y). 2004, 58, 1891–1900. [CrossRef]
- Leuschner, C. Drought response of European beech (Fagus sylvatica L.)—A review. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125576. [CrossRef]
Figure 1.
Locations of analysed F.sylvatica stands and populations.
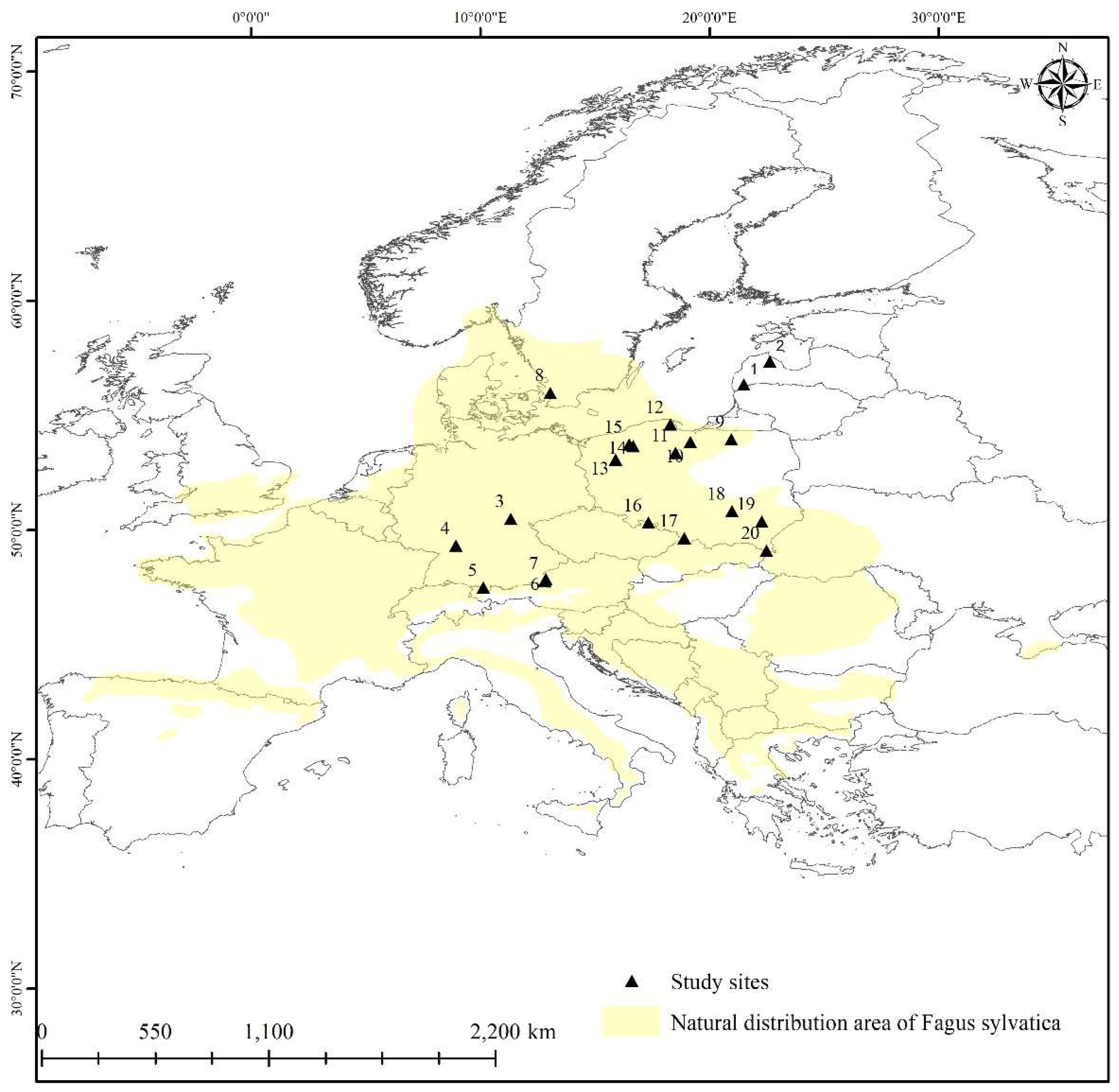
Figure 2.
Assignment of offspring to parents in the Skede stand.
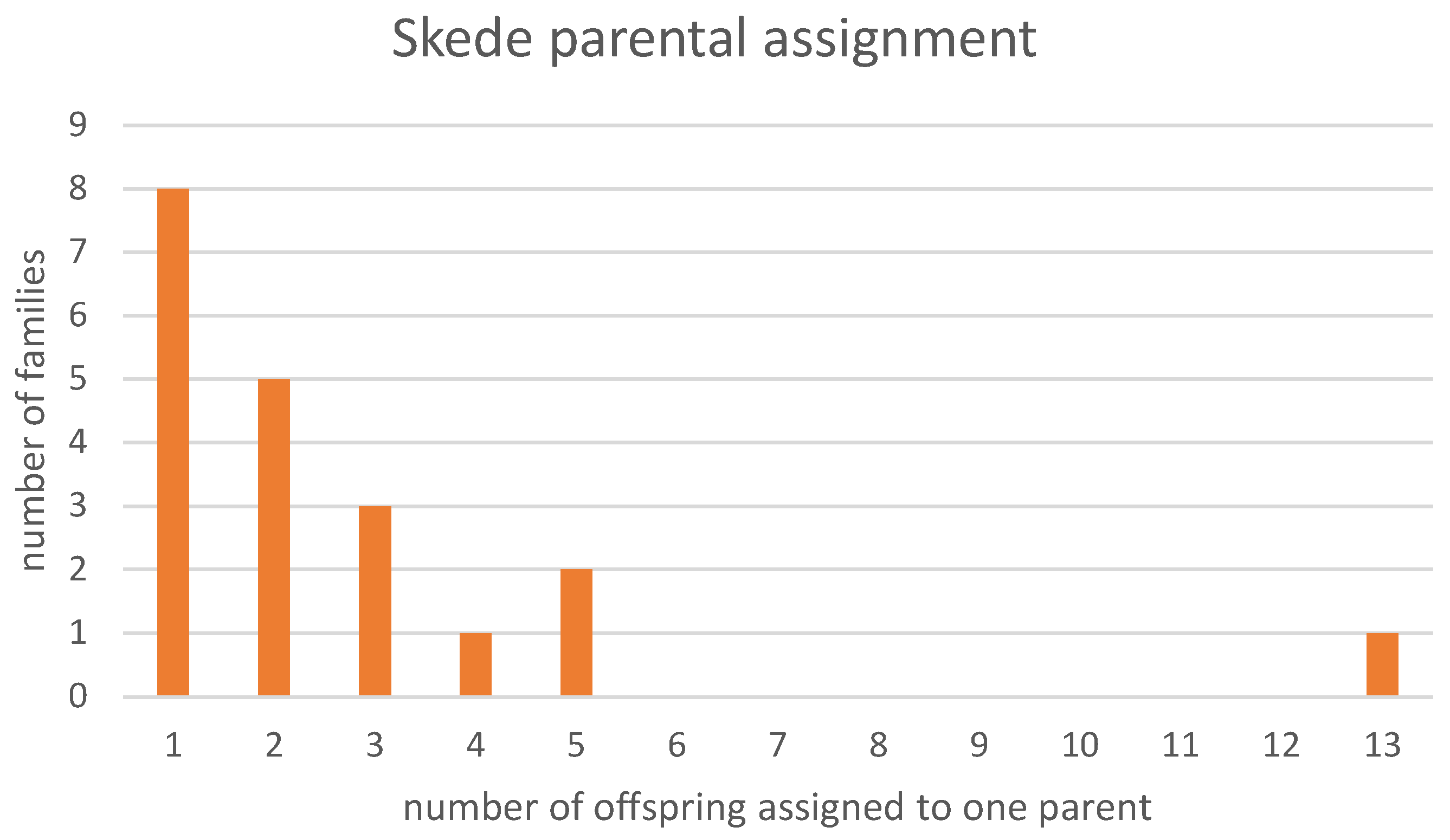
Figure 3.
Assignment of offspring to parents in the Kaleti stand.
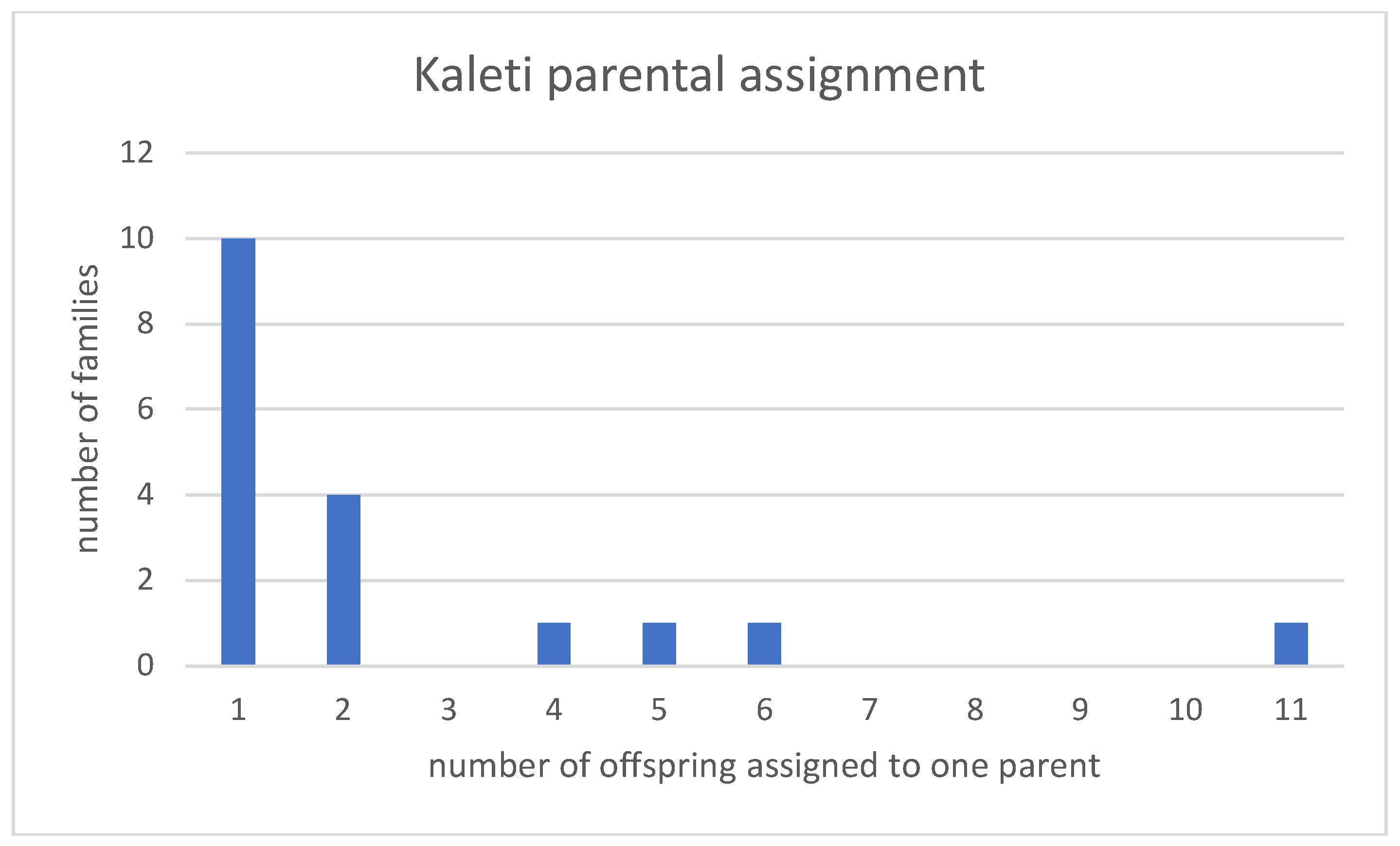
Figure 4.
Genetic diversity parameters in the Kaleti and Skede 20 parental and offspring populations. Na – mean number of alleles, Ne – mean number of effective alleles, I – mean Shannon’s Information Index, He – mean expected heterozygosity.
Figure 4.
Genetic diversity parameters in the Kaleti and Skede 20 parental and offspring populations. Na – mean number of alleles, Ne – mean number of effective alleles, I – mean Shannon’s Information Index, He – mean expected heterozygosity.
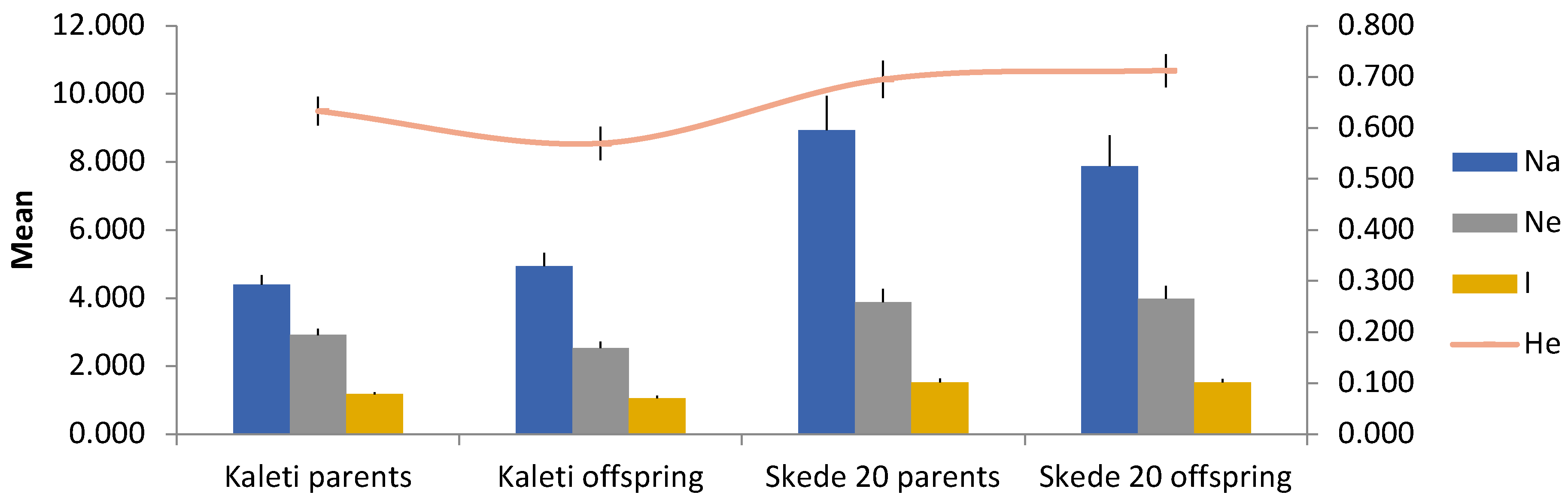
Figure 5.
Principal Coordinates Analysis (PCoA) of the pairwise Nei genetic distances between the four analysed Latvian European beech stands. Percentage of variation explained by axis 1: 98.36%, axis 2: 1.31%.
Figure 5.
Principal Coordinates Analysis (PCoA) of the pairwise Nei genetic distances between the four analysed Latvian European beech stands. Percentage of variation explained by axis 1: 98.36%, axis 2: 1.31%.
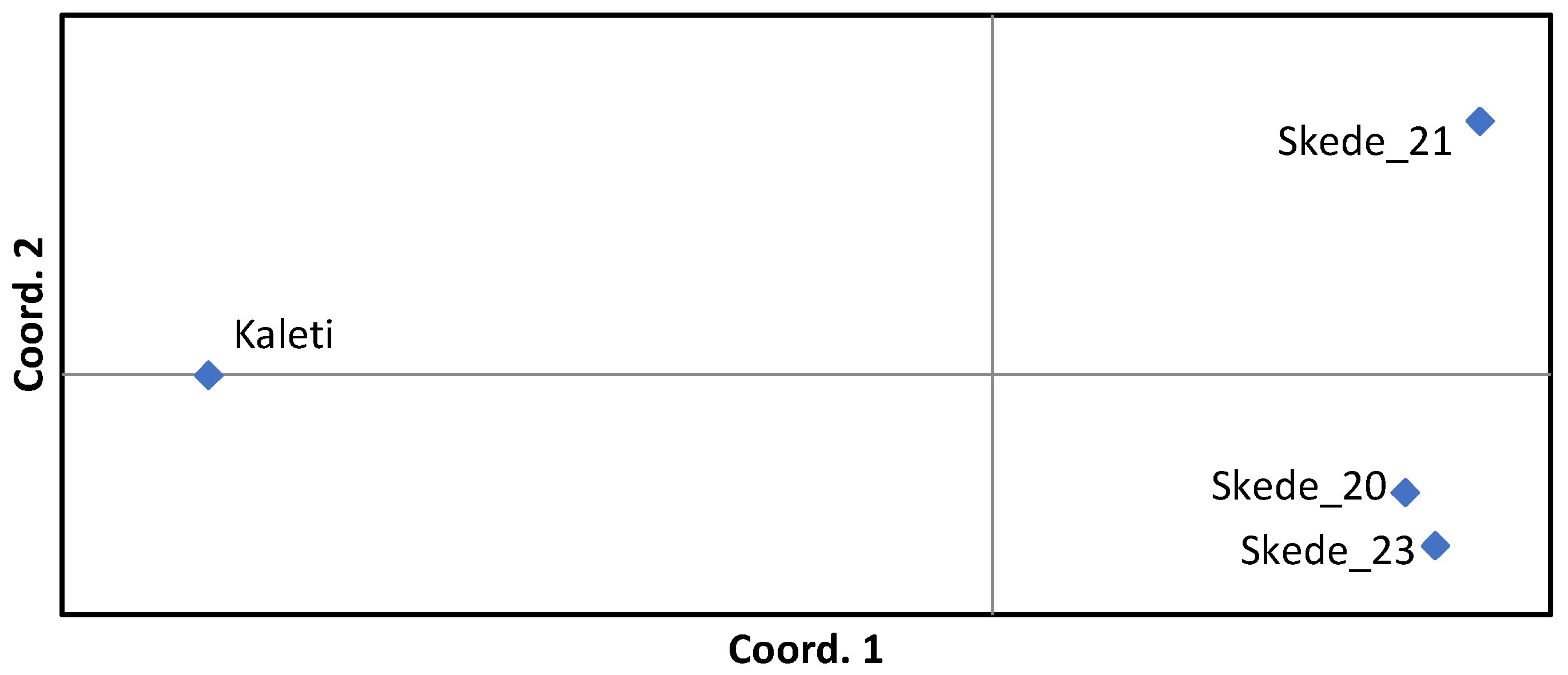
Figure 6.
DeltaK plot for the analysed individuals from southern Germany, Poland, Sweden and Latvia.
Figure 6.
DeltaK plot for the analysed individuals from southern Germany, Poland, Sweden and Latvia.
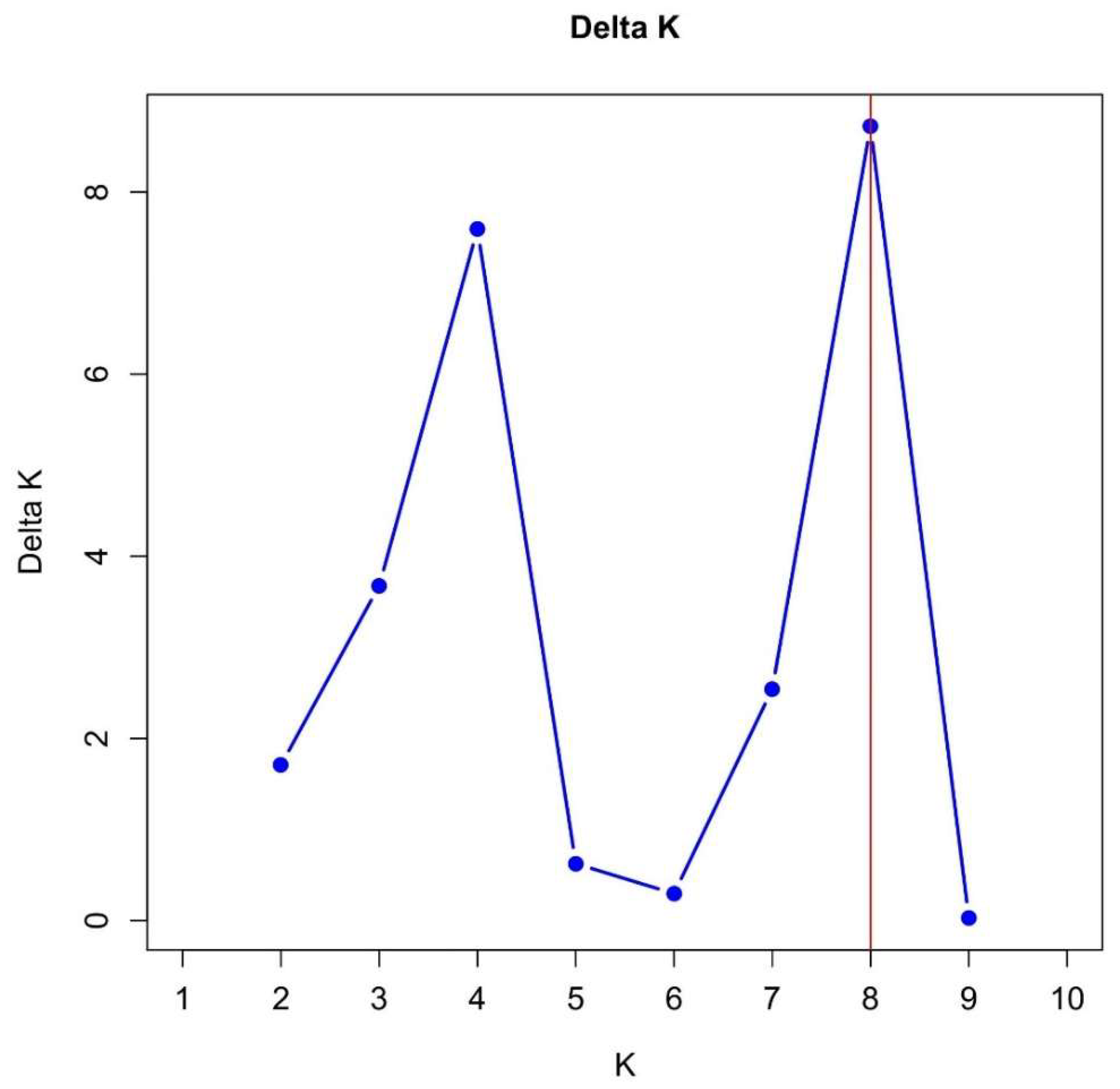
Figure 7.
Coefficients of membership (Y axis) of each population for K=4 and K=8 for the 19 predefined F.sylvatica populations.
Figure 7.
Coefficients of membership (Y axis) of each population for K=4 and K=8 for the 19 predefined F.sylvatica populations.

Figure 8.
Results of the STRUCTURE clustering on a data subset containing the five southern German populations and the Skede (LAT) population. Each vertical bar represents one individual, the Y axis indicates the membership coefficients to the two clusters.
Figure 8.
Results of the STRUCTURE clustering on a data subset containing the five southern German populations and the Skede (LAT) population. Each vertical bar represents one individual, the Y axis indicates the membership coefficients to the two clusters.
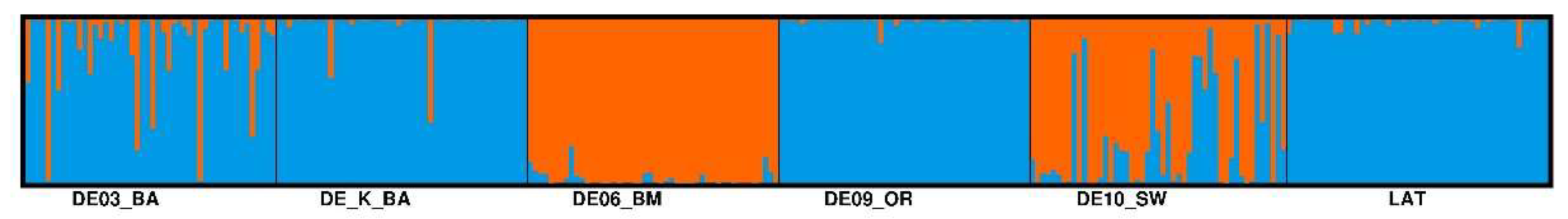

Table 1.
Genetic diversity parameters in the Kaleti and Skede 20 parental and offspring populations.
Table 1.
Genetic diversity parameters in the Kaleti and Skede 20 parental and offspring populations.
| Pop | Na (SE) | Ne (SE) | I (SE) | Ho (SE) | He (SE) | F (SE) |
|---|---|---|---|---|---|---|
| Kaleti parents | 4.400 (0.273) | 2.912 (0.184) | 1.172 (0.059) | 0.697 (0.041) | 0.633 (0.028) | -0.100 (0.036) |
| Kaleti offspring | 4.933 (0.396) | 2.524 (0.197) | 1.059 (0.065) | 0.588 (0.035) | 0.569 (0.033) | -0.040 (0.031) |
| Skede 20 parents | 8.933 (1.007) | 3.881 (0.389) | 1.516 (0.114) | 0.672 (0.037) | 0.695 (0.036) | 0.030 (0.027) |
| Skede 20 offspring | 7.867 (0.920) | 3.983 (0.378) | 1.523 (0.104) | 0.718 (0.040) | 0.712 (0.032) | -0.012 (0.038) |
Na – mean number of alleles, Na Freq. >= 5% - mean number of alleles with a frequency over 0.05, Ne – mean number of effective alleles, I – mean Shannon’s Information Index, He – mean expected heterozygosity, SE – standard error.
Table 2.
Genetic diversity parameters of mature individuals in the Kaleti and Skede stands.
| Stand | Na (SE) | Ne (SE) | I (SE) | Ho (SE) | He (SE) | F (SE) |
|---|---|---|---|---|---|---|
| Skede_20 | 8.500 (0.976) | 3.722 (0.382) | 1.474 (0.114) | 0.669 (0.040) | 0.685 (0.037) | 0.021 (0.027) |
| Skede_21 | 7.286 (0.952) | 3.713 (0.386) | 1.456 (0.124) | 0.649 (0.043) | 0.676 (0.044) | 0.031 (0.029) |
| Skede_23 | 7.000 (0.864) | 3.553 (0.413) | 1.396 (0.125) | 0.618 (0.047) | 0.658 (0.045) | 0.058 (0.03) |
| Kaleti | 4.286 (0.266) | 2.956 (0.192) | 1.177 (0.063) | 0.707 (0.042) | 0.638 (0.030) | -0.107 (0.038) |
Na – mean number of alleles, Ne – mean number of effective alleles, I – mean Shannon’s Information Index, Ho – mean observed heterozygosity, He – mean expected heterozygosity, F – mean Fixation Index, SE – standard error.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



