Submitted:
03 December 2024
Posted:
04 December 2024
Read the latest preprint version here
Abstract
Wildfire is a key factor in regulating ecological processes in grassland ecosystems; however, changes in land use/cover have modified the intensity and frequency of fires as occurred naturally. Different factors have caused a rise in woody vegetation in these ecosystems, leading to changes in species composition, diversity, and biogeochemical cycles. The prescribed burns are a tool for controlling and eradicating shrubs; however, its effectiveness depends on vegetation composition, biomass availability, and the objectives of restoration. We evaluate the effectiveness of fire as a shrub controller in a semiarid grassland ecosystem. We measured several shrub dasometric pa-rameters and the percentage of damage in ten 2000 m2 plots, three months after a prescribed burning was performed. Both crown height and width, and total height were the main variables that explained the percentage of shrub damage by fire. Individuals with a height greater than 1.6 m and wide crowns, did not suffer damage. Moreover, even though 96% of the total shrubs presented some fire damage, 60% recovered after the rain period. Our results show that fire could be an effective strategy to control early-growing shrubs, but on overgrazed arid lands it would be difficult to have enough biomass for implementing burning programs.
Keywords:
shrub colonization
; woody encroachment
; biomass
; controlled fire
1. Introduction
Semi-arid natural grasslands are distributed from central Mexico to the southern United States of America, in the Chihuahuan desert region [1] (Huenneke et al., 2001). The grasslands are mainly covered by various species of grasses and some shrubs in low density [2] (Rzedowski, 2021). In recent years, grasslands have faced multiple threats, such as the alteration of natural fire cycles, the increase in CO2, conversion to agriculture, overuse by extensive livestock, the introduction of exotic grasses, and the increase in herbaceous and less desirable shrubs [3] (Barger et al., 2011). The presence of shrubs in the pastures is a normal state; however, these factors have triggered colonization processes of the native shrub and semi-shrub species, causing the displacement of grasses species and the consequent changes in species diversity, availability of forage for cattle, alteration in the cycle of nutrients, etc. [4] (Van Auken, 2000).
The woody species that are increasing their density in the Mexican central highlands are those of the genera Vachellia, Prosopis, Opuntia, Larrea, Brickellia and Isocoma. The increase in the density of shrub species has shown to reduce the availability of soil water for the dominant plant species [5] (Caldeira et al., 2015). In addition, these species have a high capacity for adaptation to degraded areas [6] (Kidron, 2013). It has been reported that the reduction of herbivores is a driver of the increase of shrubs [7] (Mata-González et al., 2007) and that excessive grazing has the same adverse effects; mainly due to the reduction in the amount of biomass (fuel) available to propitiate a natural fire with sufficient intensity to affect shrub species [8] (Munson et al., 2013). Therefore, the balance of herbaceous-woody species must be analyzed in the context of interactions of cattle grazing with other factors such as climate, topography, fire frequency, nurse plants, soil type, among others [9] (Sepp et al., 2021).
The colonization of shrubs is a process that is difficult to reverse. The decrease in the intensity of grazing does not necessarily reduce or even reverse the colonization of shrubs, but it is necessary to invest in different techniques that range from the application of mechanical, chemical, pyric, and sometimes also biological treatments [10] (Ibarra et al., 2014). On the other hand, fire, which naturally limited the growth and development of shrubs and shrub species centuries ago [11] (Osborn et al., 2002), has decreased in frequency and in many cases has been completely eradicated due to the reduction of above ground biomass or fuel in grassland ecosystems. Historically, grasslands have been subjected to periods of fire, which has led them to adapt and maintain themselves as fire-dependent species since their meristems are close to the ground surface and are less susceptible to fire [12] (Rego et al., 2021). When the natural fire frequency was modified, the shrub species increased their growth and density, becoming an ecological and economic problem in the region and displacing key species in the pastures [13] (Wright and Bayley, 1982). In this sense, prescribed burning programs have recently been applied in grassland ecosystems in order to eliminate shrub species and recover ecosystem productivity. However, some studies report that fire frequency every 3 to 4 years do not guarantee the control of the expansion of woody plants [14] (Nippert et al., 2021), since the intensity of regularly prescribed fires is less than natural fires in non-fragmented landscapes. By increasing fire intensity and frequency, short- and long-term post-fire soil responses could be improved, which favors a prompt restoration of natural pastures [15] (Hahn et al., 2021).
Shrub colonization in grassland ecosystems occurs gradually at first, until a point is reached where woody species spread rapidly [16] (Ratajczak et al., 2014). Once woody species reach considerable size and density, the resilience of grasslands deteriorates, resulting in a rapid increase in the abundance of woody plants [17] (Ratajczak et al., 2017). Taking into account that fire (prescribed burning) can be used as a tool to reverse the colonization and prevent the recruitment of new shrub seedlings, and that its effectiveness depends on the intensity to achieve greater mortality of woody plants, the objective of this study was to evaluate the effect of fire (by prescribed burnings) on shrub species in different growth stages in the Llanos de Ojuelos, Jalisco, as well as identify the main dasometric variables that influence the degree of fire damage caused by fire.
2. Materials and Methods
The Materials and Methods should be described with sufficient details to allow others to replicate and build on the published results. Please note that the publication of your manuscript implicates that you must make all materials, data, computer code, and protocols associated with the publication available to readers. Please disclose at the submission stage any restrictions on the availability of materials or information. New methods and protocols should be described in detail while well-established methods can be briefly described and appropriately cited.
2.1. Study Area
The study area is located in the arid and semi-arid region of the Central Plateau of Mexico, at the south of the Chihuahuan desert, in Santo Domingo ranch. Is an area dedicated to livestock production for 36 years. The production system is cattle, with grazing intensities ranging between 3 and 7 ha per animal unit and a forage consumption of less than 40% of the available aerial biomass. The study area is managed in a rotational grazing system with 8 paddocks of 60 ha.
Figure 1.
Changes in shrub abundance into the experimental site. Images correspond to years 2011 (left) and 2022 (right; Google, s.f.).
Figure 1.
Changes in shrub abundance into the experimental site. Images correspond to years 2011 (left) and 2022 (right; Google, s.f.).

2.2. Vegetation Cover
The predominant vegetation type in the Llanos de Ojuelos consists of scrub and grasslands. The grasslands are mainly composed of blue grama grass (Bouteloua gracilis), wolfstail (Lycurus phleoides), scorpion grama (Bouteloua scorpioides), three-bearded grass (Aristida divaricata), buffalo grass (Bouteloua dactyloides), silvery strawweed (Bothriochloa barbinodis), zacatón (Muhlenbergia rigida), and hairy grama (Bouteloua hirsuta). These grasslands are interspersed with shrubs such as huizache (Vachellia schaffneri), cat's claw (Mimosa biuncifera), and mesquite (Prosopis leaviagata), which, while native to scrublands, are considered invasive. The herbaceous vegetation includes species like foxtail (Brickellia spinulosa), sawtooth candyleaf (Stevia serrata), toad grass (Eryngium carlinae F. Delaroche), and trompillo (Solanum elaeagnifolium), among others.
2.3. Climate
The climate of the study area is characterized as dry and semi-dry, with precipitation levels consistently lower than potential evapotranspiration. Rainfall is highly variable, ranging between 300 and 500 mm annually, with a mean annual precipitation of 424 mm. Precipitation typically occurs from June to September. The average annual temperature varies between 16 and 18 °C. The soils are shallow, less than 50 cm deep, with minimal organic matter and a cemented layer (caliche, tepetate). Dominant soil types include durisols and phaeozem [18,19] (INEGI, 2007; WRB, 2014).
2.4. History and Grazing Management
The study site is a paddock planted with weeping lovegrass (Eragrostis curvula). Rainfed agriculture was practiced here until the early 1980s, after which the land was abandoned. For the past 15 years prior to burning, the pasture was primarily used for producing weeping lovegrass seed, with minimal cattle grazing. The production system on the Santo Domingo Ranch involves cattle grazing with intensities ranging from 3 to 7 hectares per animal unit and a forage consumption rate of less than 40% of the available aerial biomass. The area is managed using a rotational grazing system with 8 paddocks of 60 hectares each.
2.5. Experimental Design
To evaluate the effect of prescribed burning in grasslands invaded by Vachellia schaffneri and Mimosa biuncifera, four different sites were selected for the study. At Sites 1 and 3, four plots of 20 x 100 meters (2000 m²) each were established, while at Sites 2 and 4, three plots of the same dimensions were set up. Prescribed burns were applied to the plots at these sites. To prevent the spread of fire to neighboring areas, the plots were delimited with mineral lines and black lines. All plots had approximately 20% shrub cover with varying heights. Measurements of all shrubs within the plots were taken before the burn and three months afterward. Data collected included diameter at 30 cm above ground (base diameter), total height, crown height, crown diameter, number of branches, and the shrub’s condition (alive or dead). Additionally, the presence of regrowth and the percentage of damage, such as black burn marks on dead stems or the absence of aerial biomass, were recorded.
Each individual shrub was marked with a metal plate containing the plot number and the shrub number. It was assumed that individuals not located, or whose identification plates were missing, were consumed by the fire after the prescribed burn.
2.6. Vegetation Sampling
Aboveground biomass and species diversity were assessed using 1 m² plots, with a total of 20 plots sampled. To measure aboveground biomass, herbaceous vegetation was clipped, dried, and then weighed. The biomass was separated into two categories: living biomass, representing current year growth, and dead biomass, consisting of residual plant material from the previous year. Additionally, the percentage of grasses and herbaceous plants present in the plots was identified and quantified, providing a detailed assessment of plant composition.
Species diversity was evaluated by recording the number of species present in each plot, allowing for the calculation of diversity indices. These data were collected after the prescribed burn treatments. To compare the available biomass before the burn treatment, contiguous plots were sampled prior to the prescribed burn, which allowed for the assessment of pre-existing biomass in the area.
2.7. Application of Prescribed Burns
The prescribed burns were conducted on different dates to effectively control and manage the fire, minimizing risks and ensuring that the most favorable weather conditions were selected. The first prescribed burn was conducted on March 29, 2021, over an area of 10 hectares with a slope of less than 3%. A head-fire burn was applied with a fuel load of 11,160 t/ha, relative humidity exceeding 30%, and wind speeds below 10 km/h from the south-southeast. The second prescribed burn was performed on April 26, 2021, in another 10-hectare area. This burn had a fuel load of 12,220 t/ha, with relative humidity again exceeding 30% and wind speeds under 7 km/h from the south. The higher fuel load resulted in flame heights reaching approximately 10 meters.
The third prescribed burn was executed on April 19, 2022, over an area of 9 hectares. This site had grass cover and biomass accumulation similar to the previous sites. The recorded environmental conditions included a relative humidity of 69%, wind speeds of 3.7 km/h, and a temperature of 54°F (12°C). The fourth prescribed burn was conducted on April 22, 2022, over an area of 10 hectares with measurements showing a relative humidity of 47%, a temperature of 60°F (16°C), and wind speeds of 9 km/h.
Figure 2.
Aereal view of one of the four sites before (left) and after (right) the burning treatment. The black contour in the left image is the protection line which was made one week before the prescribed burning.
Figure 2.
Aereal view of one of the four sites before (left) and after (right) the burning treatment. The black contour in the left image is the protection line which was made one week before the prescribed burning.

2.8. Environmental Variables
Environmental variables were recorded to characterize aboveground and soil temperatures during the prescribed burn at site 4. This site had grass cover and biomass accumulation similar to the other treatment sites. Soil temperature was measured every second using four temperature sensors (TMC20-HD, Onset, Bourne, MA) connected to a data logger (U12-006, Onset, Bourne, MA). The sensors were placed at depths of 1, 3, 5, and 10 cm in the soil. Air temperature was monitored using three type K thermocouples positioned at heights of 50, 100, and 150 cm above the ground, while an additional three thermocouples were placed directly over tussocks to monitor air temperature at the crown level.
2.9. Analysis of Data
Species composition was evaluated by measuring the proportion of dry biomass of each species within the cutting plots. For shrub composition, we quantified the number of individual shrubs per species in the fire plots to assess changes in species abundance and diversity due to the burns.
To determine the variables that most significantly affected shrub species and to identify the thresholds for vegetation response to fire, we employed boosted regression trees (BRTs). This analytical method helps to uncover complex relationships between environmental variables and vegetation response. The "GBM" (Generalized Boosted Regression Models) library in R software [20] (R Core Team, 2020) was used for this analysis. BRTs were particularly useful for handling nonlinear and interactive effects of multiple variables on shrub response, providing insights into how different factors such as fire dasometric variables influence fire damage and shrub recovery and survival.
By using BRTs, we aimed to capture the most influential factors affecting shrub species and to identify critical thresholds for vegetation resilience to fire. This approach allows for a more nuanced understanding of fire’s impacts on different shrub species.
3. Results
3.1. Species Composition
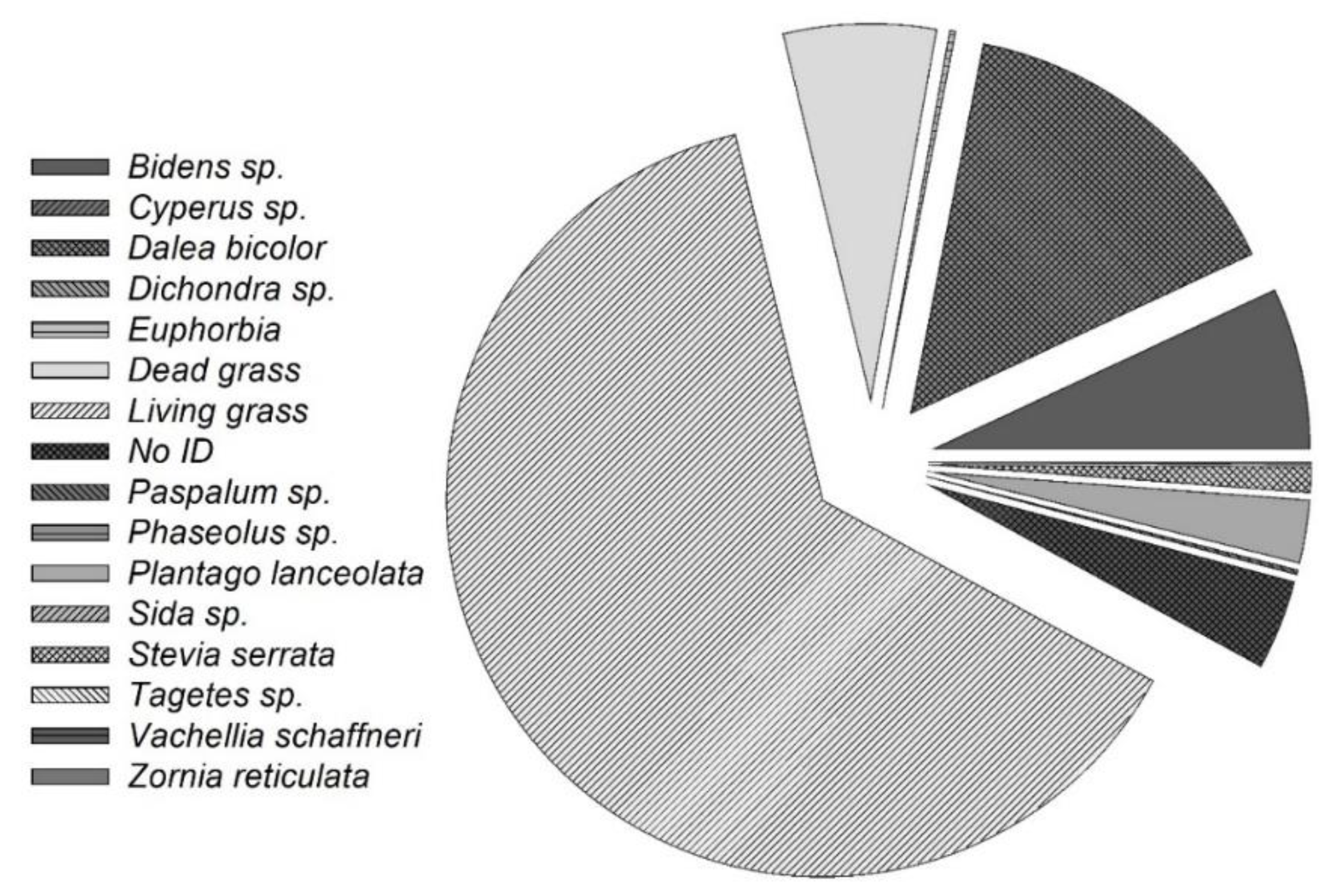
The herbaceous stratum at the study site consisted of 13 herbaceous species, including 8 native grasses and 1 introduced species, Eragrostis curvula (Figure 3). Native grasses dominated the biomass, comprising 75% of the total. The shrub layer was predominantly composed of Vachellia schaffneri, Mimosa biuncifera, and Prosopis species, with 97% of individuals identified as V. schaffneri. Shrub heights ranged from 30 cm to 3.5 m, with an average crown width of 3 m.
Figure 3.
Plant species composition and biomass contribution (%) in the study site.
3.2. Prescribed Burn Temperatures
Soil temperature did not show significant changes due to the fire effect (Figure 4a); however, larger variations were observed at a depth of 1 cm due to darkened soil surface by deposition of charcoal residues that remained on soil several days after the burning (Figure 4b; Figure 2). In contrast, tussocks experienced crown temperatures as high as 500°C during the burn, although this peak temperature lasted for less than a minute (Figure 4c). Air temperatures decreased with height, from 180°C at 50 cm to 100°C at 150 cm aboveground (Figure 4d).
3.3. Fire Effects on Shrubs
Ninety-seven percent of the total shrubs exhibited some level of damage, while the remaining shrubs were unaffected by fire. Of the damaged shrubs, 86% showed regrowth after the rainy season, even when the damage was complete (100%).
For Vachelia shrubs, those lower than 5 cm of DAP, with an average crown lower than 2 meters, and lower than 2 m height showed regrowth after the fire treatment. The most influential variable on fire damage was crown area, contributing 53% to the relative influence, followed by diameter and total height (24% and 22%, respectively; Table 1; Figure 5f). Crown and stem shape did not significantly affect damage levels and were therefore excluded from the final regression trees.
Shrubs with the highest percentage of damage were those with crown areas lower than 5 square meters, lower than 5 cm of DAP, and heights between 0 and 2 meters (Figure 4). This suggests that the fire caused total damage to younger Vachelia shrubs with smaller crowns. As shrub height and crown width increased, the fire inflicted little or no damage.
For Mimosa shrubs, the crown diameter was the variable that most explained the number of resprouts after fire treatment (60%), it was followed by DAP (30%) and plant height (10%). Since all individuals of Mimosa shrubs were damaged at 100%, this variable was not used for the boosted regression tree analysis (Table 1). Recovery of shrubs increased with the crown diameter, i.e. shrubs with smaller crowns produced a lower number of sprouts than the bigger ones. This same pattern was observed with the height of shrubs; taller plants produced more resprouts after the fire. The 99% of Mimosa shrubs were totally consumed by fire, but only 18% of these plants did not produce any resprout.
For both Vachellia and Mimosa, lower in stature and in crown area plants were more susceptible to fire (Figure 5). These variables were related to the number of resprouts after fire, which could be interpreted as resistance or the capacity of shrubs to recover from disturbances. Base diameter is related to the age of shrubs or vigor and is correlated with plant height. This suggests that smaller and younger shrubs were more susceptible to fire damage, while larger, more established individuals demonstrated higher resilience.
4. Discussion
Our results showed that the burning affected 97% (Vachellia) and 100% (Mimosa) of the shrubs; however, on average, 85% of the affected shrubs recovered after the first rains. The taller individuals with larger base diameters and with wide crowns did not suffer any effects from the burning. This indicates that as height and crown width increase, fire causes little or no damage to the shrubs. This quick recovery and low fire damage to large shrubs are likely explained by thicker bark at the base of the shrubs, lower grass biomass (fuel) at the base, and taller growth structures that lessen the effect of fire on adult plants. It also suggests that shrubs become less vulnerable to the effects of fire as they grow [21] (Archer et al., 2017). A larger crown area reduces grass cover due to shading just below the shrub crown, which lowers fire intensity by decreasing the availability of fuel. On the other hand, greater height allows the growth structures to escape from the fire [22,23] (Ludwig et al., 2001; Harrington and Kathol, 2009). This indicates that fire causes total damage to low-growing shrubs with small crowns.
Available biomass is a factor to consider during burning. In our plots, there was an average biomass of 12 tons per hectare, and although the fire reached flame heights of around 10 meters, its duration in certain areas was only a few seconds due to the speed of the fire’s movement (Figure 3). Our results showed that a single burn reduces shrub cover by 15%, primarily affecting young plants while having minimal impact on other herbaceous species that are senescent or found in the soil seed bank. One year after the burn, net primary productivity recovered 100%, and other studies in semi-arid grasslands suggest that full biomass recovery after burning takes about 3-5 years [24,25] (Bennett et al., 2003; Wells et al., 2021). Our study site is not the typical grassland in Mexico; overgrazing by cattle maintain very low standing biomass levels (<1 t ha-1), which prevents of severe wildfires, but that could not represent a threat for shrub survival.
In the early stage of shrub colonization, grasses can suppress shrub dominance through competition for near-surface soil resources, slowing shrub growth. However, when grasses are inactive, shrubs use these resources to accelerate growth [26] (Holdo and Brocato, 2015). In the later stages of the transition from grassland to shrubland, shrub-shrub competition does not slow the expansion rate of shrubs [27] (Pierce et al., 2019). An adult-stage shrub can reach heights of 3 to 4 meters and develop better roots, allowing for greater nutrient reserves [28] (Bond and Midgley, 2003), which could translate into greater fire resistance and the ability to regrow even after 100% damage. Woody plants benefit from disturbance regimes as they are capable of resprouting, though these sprouts tend to produce fewer seeds than older branches [28,29] (Bond and Midgley, 2003; Li et al., 2022). This has implications for management and conservation programs that use fire to control shrubs.
Our results reveal that the diameter at 30 cm is an important factor for lowering fire damage and for favoring resprouting. This finding can be explained by several ecological and morphological factors. First, the diameter at 30 cm from the ground serves as an indicator of the shrub’s age and vigor. Shrubs with a larger diameter, which are typically older and more vigorous, have a greater capacity to generate biomass and support a more extensive branching structure [29] (Bond and Midgley, 2003). This is because a larger diameter provides a more robust base capable of supporting a greater number of branches. On the other hand, the crown area is closely related to the shrub's photosynthetic capacity, which influences branch formation. A shrub with a larger crown can capture more sunlight and produce more energy, facilitating the development of a greater number of branches [30] (Weber-Grullon et al., 2022). In contrast, total height is not necessarily linked to branching, as some species can grow in height without proportionally increasing the number of branches [31] (Yang et al., 2024). Vachellia and Mimosa showed similar responses; even though Mimosa are lower in stature than Vachellia, burned plants resprouted and only a minimum percentage of plants died. Fire and grazing have modeled plant communities for millennia, and environmental conditions have selected species with morphological characteristics for deep water exploration and root reserves. This suggests that shrub morphology and branching capacity have been determined by environmental conditions and disturbances, with high capacity of resistance and survival [32] (Scholtz et al., 2017). Post-fire resilience of shrubs may be associated with their branching capacity; shrubs with a larger diameter and bigger crown have more resources to recover after a disturbance such as fire [33] (Capozzelli et al., 2020). Base diameter and crown area are an indicative of recovery capacity, as more robust shrubs with a larger crown seem better equipped to withstand damage and regenerate branches more easily [34] (Jordan, 2024).
As brush encroachment increases, grasses and other flammable fuels decrease, so reintroducing fire after prolonged suppression may not necessarily be beneficial [35] (Wright, 1974). It is critical to determine the frequency of fire needed to prevent shrubs from reaching a crown height or width at which fire is no longer an effective control tool. Other factors, such as fire frequency and intensity's effects on biodiversity, also need consideration [36] (Jones and Tingley, 2021).
Interestingly, the relationship between the percentage of damage and crown diameter and height is not linear, indicating that prolonged periods without fire pressure or more favorable climatic events for shrub growth (e.g., winter rains; [37] Biederman et al., 2018) could lead to "no return" colonization stages that are not controllable with fire. The woody species that resprout are the most difficult to control with fire alone and require additional management strategies to reduce their abundance and eventually remove them from the ecosystem. In agreement with some studies, combining prescribed burns with the browsing of ruminant species can reduce shrub density by up to 90% and promote grass cover by improving the light environment for herbaceous species [38,39,40] (Ascoli et al., 2013; Pausas et al., 2016; O'Connor et al., 2020). Although controlled grazing can be a valuable supplementary technique, it cannot replace fire for controlling brush in these systems [23,40] (Harrington and Kathol, 2009; O'Connor et al., 2020). Properly timed prescribed burns can be a key ally in controlling shrubs before they become a serious problem.
Our study shows some weaknesses and limitations, such as the lack of spatial and temporal variability, which limits its general applicability. On the other hand, the observed recovery in 85% of the shrubs after the first rains suggests that post-fire climatic conditions, such as the amount and frequency of rainfall, were favorable for regeneration. In scenarios with lower water availability or more extreme climatic conditions, the results could be different. Although primary productivity recovery is mentioned within a year, longer-term effects, such as changes in species composition or ecosystem dynamics, are not fully explored. These effects could have more significant implications for long-term management.
5. Conclusions
Fire primarily affects young and small shrubs. When shrubs exceed large crown areas, base diameter or height, the fire causes minimal or no damage. Species within the Vachellia and Mimosa genera do not show differences in their degree of fire damage, indicating that the species itself does not significantly affect the percentage of damage incurred. Vachellia spp. and Mimosa spp. exhibit resilience to adverse environmental conditions, including fire. Based on our results, it is beneficial to apply fire treatments to control these species during their early growth stages, when their characteristics make them more susceptible to fire. Continuous prescribed burning could potentially improve shrub control [41] (Taylor, 2005). However, in overgrazed arid lands, it may be challenging to maintain sufficient biomass for a continuous two-year burning program.
Author Contributions
T. Alfaro-Reyna: Conceptualization, Methodology, Formal analysis, Investigation, Data curation, Writing – original draft, Writing – review & editing; J. de la Cruz Domínguez: Methodology, Data curation, Writing – original draft; C.A. Aguirre-Gutierrez: Investigation; Writing – original draft, Writing – review & editing; M. Luna-Luna: Methodology, Investigation, Writing – review & editing; D. Flores-Rentería: Writing – original draft, Writing – review & editing; J. Delgado-Balbuena: Methodology, Formal analysis, Investigation Writing – original draft, Writing – review & editing, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CONAHCYT with project reference CF 320641.
Data Availability Statement
Data will be available from authors under reasonable request.
Acknowledgments
Authors thank National Confederation of Livestock Organizations (CNOG), and to Miguel Luna-Luna, the manager of Santo Domingo Ranch, for the facilities to carry out this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Huenneke, L. F.; Clason, D.; Muldavin, E. Spatial heterogeneity in Chihuahuan Desert vegetation: implications for sampling methods in semi-arid ecosystems. Journal of Arid Environments 2001, 47, 257–270. [Google Scholar] [CrossRef]
- Rzedowski, J. 2021 Flora of the Valley of Mexico. 2nd edition. Institute of Ecology, Mexico.
- Barger, N. N.; Archer, S. R.; Campbell, J. L.; Huang, C. Y.; Morton, J. A.; Knapp, A. K. Woody plant proliferation in North American drylands: a synthesis of impacts on ecosystem carbon balance. Journal of Geophysical Research: Biogeosciences. [CrossRef]
- Van Auken, O. W. Shrub invasions of North American semiarid grasslands. Annual review of ecology and systematics 2000, 31, 197–215 . [Google Scholar] [CrossRef]
- Caldeira, M. C.; Lecomte, X.; David, T. S.; Pinto, J. G.; Bugalho, M. N.; Werner, C. Synergy of extreme drought and shrub invasion reduce ecosystem functioning and resilience in water-limited climates. Scientific reports 2015, 5, 15110. [Google Scholar] [CrossRef] [PubMed]
- Kidron, G. J.; Gutschick, V. P. Soil moisture correlates with shrub–grass association in the Chihuahuan Desert. Catena 2013, 107, 71–79. [Google Scholar] [CrossRef]
- Mata-González, R.; Figueroa-Sandoval, B.; Clemente, F.; Manzano, M. Vegetation changes after livestock grazing exclusion and shrub control in the southern Chihuahuan Desert. Western North American Naturalist 2007, 67, 63–70. [Google Scholar] [CrossRef]
- Munson, S. M.; Muldavin, E. H.; Belnap, J.; Peters, D. P.; Anderson, J. P.; Reiser, M. H. . & Christiansen, T. A. Regional signatures of plant response to drought and elevated temperature across a desert ecosystem. Ecology 2013, 94, 2030–2041 . [Google Scholar] [CrossRef]
- Sepp, S. K.; Davison, J.; Moora, M.; Neuenkamp, L.; Oja, J.; Roslin, T. . & Zobel, M. Woody encroachment in grassland elicits complex changes in the functional structure of above-and belowground biota. Ecosphere 2021, 12, e03512 . [Google Scholar] [CrossRef]
- Ibarra Flores, F. A.; Martín Rivera, S.; Moreno Medina, M.; Denogean Ballesteros, F. G.; Martinez Duran, A. B.; Retes López, R. ;... & Aguilar Valdes, A. Beneficios económicos asociados con el control de invasiones de uña de gato en el pastizal mediano abierto de Cananea, Sonora, México. Revista Mexicana de Agronegocios 2016, 34(1345-2016-104467). [Google Scholar] [CrossRef]
- Osborn, S.; Wright, V. (2002). Understanding and managing invasive plants in wilderness and other natural areas: An annotated reading list (Vol. 4). US Department of Agriculture, Forest Service, Rocky Mountain Research Station.
- Rego, F.; Rigolot, E.; Fernandes, P.; Montiel, C.; Silva, J. S. Towards integrated fire management. EFI Policy Brief 2010, 4, 16. [Google Scholar]
- Wright, H.A.; and A.W. Bailey. 1982. Fire Ecology. United States and Southern Canada. John Wiley & Sons, Inc.
- Nippert, J. B.; Telleria, L.; Blackmore, P.; Taylor, J. H.; O'Connor, R. C. Is a prescribed fire sufficient to slow the spread of woody plants in an infrequently burned grassland? A case study in tallgrass prairie. Rangeland Ecology & Management 2021, 78, 79–89. [Google Scholar] [CrossRef]
- Hahn, G. E.; Coates, T. A.; Aust, W. M. Soil chemistry following single-entry, dormant season prescribed fires in the Ridge and Valley Province of Virginia, USA. Communications in Soil Science and Plant Analysis 2021, 52, 2065–2073. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J. B.; Ocheltree, T. W. Abrupt transition of mesic grassland to shrubland: evidence for thresholds, alternative attractors, and regime shifts. Ecology 2014, 95, 2633–2645 . [Google Scholar] [CrossRef]
- Ratajczak, Z.; D'Odorico, P.; Nippert, J. B.; Collins, S. L.; Brunsell, N. A.; Ravi, S. Changes in spatial variance during a grassland to shrubland state transition. Journal of Ecology 2017, 105, 750–760 . [Google Scholar] [CrossRef]
- INEGI, Conjunto de datos vectorial edafológico. Escala 1:250000. Serie II. Continuo Nacional. 2007, Instituto Nacional de Estadística y Geografía: Aguascalientes, México.
- WRB, 2014. World Reference Base for Soil Resources 2015. International soil classification system for naming soils and creating legends for soil maps, 106. FAO, Rome, Italy, 203 pp.
- R Core Team (2020). A: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
- Archer, S. R.; Andersen, E. M.; Predick, K. I.; Schwinning, S.; Steidl, R. J.; Woods, S. R. Woody plant encroachment: causes and consequences. Rangeland systems: Processes, management and challenges 2017, 25–84. [Google Scholar] [CrossRef]
- Ludwig, F.; de Kroon, H.; Prins, H. H.; Berendse, F. Effects of nutrients and shade on tree-grass interactions in an East African savanna. Journal of Vegetation Science 2001, 12, 579–588 . [Google Scholar] [CrossRef]
- Harrington, J. A.; Kathol, E. Responses of shrub midstory and herbaceous layers to managed grazing and fire in a North American savanna (oak woodland) and prairie landscape. Restoration Ecology 2009, 17, 234–244. [Google Scholar] [CrossRef]
- Bennett, L. T.; Judd, T. S.; Adams, M. A. Growth and nutrient content of perennial grasslands following burning in semi-arid, sub-tropical Australia. Plant Ecology 2003, 164, 185–199. [Google Scholar] [CrossRef]
- Wells, A. G.; Munson, S. M.; Sesnie, S. E.; Villarreal, M. L. Remotely sensed fine-fuel changes from wildfire and prescribed fire in a semi-arid grassland. Fire 2021, 4, 84 . [Google Scholar] [CrossRef]
- Holdo, R. M.; Brocato, E. R. Tree–grass competition varies across select savanna tree species: a potential role for rooting depth. Plant Ecology 2015, 216, 577–588. [Google Scholar] [CrossRef]
- Pierce, N. A.; Archer, S. R.; Bestelmeyer, B. T. Competition suppresses shrubs during early, but not late, stages of arid grassland–shrubland state transition. Functional Ecology 2019, 33, 1480–1490 . [Google Scholar] [CrossRef]
- Bond, W. J.; Midgley, J. J. The evolutionary ecology of sprouting in woody plants. International Journal of Plant Sciences 2003, 164(S3), S103–S114. [Google Scholar] [CrossRef]
- Li, J.; Ravi, S.; Wang, G.; Van Pelt, R. S.; Gill, T. E.; Sankey, J. B. Woody plant encroachment of grassland and the reversibility of shrub dominance: Erosion, fire, and feedback processes. Ecosphere 2022, 13, e3949 . [Google Scholar] [CrossRef]
- Weber-Grullon, L.; Gherardi, L.; Rutherford, W. A.; Archer, S. R.; Sala, O. E. Woody-plant encroachment: Precipitation, herbivory, and grass-competition interact to affect shrub recruitment. Ecological Applications 2022, 32, e2536 . [Google Scholar] [CrossRef]
- Yang, J.; Will, R.; Zhai, L.; Zou, C. Future climate change shifts the ranges of major encroaching woody plant species in the Southern Great Plains, USA. Earth's Future 2024, 12, e2024EF004520 . [Google Scholar] [CrossRef]
- Scholtz, R.; Smit, I. P. J.; Coetsee, C.; Kiker, G. A.; Venter, F. J. Legacy effects of top–down disturbances on woody plant species composition in semi-arid systems. Austral Ecology 2017, 42, 72–83 . [Google Scholar] [CrossRef]
- Capozzelli, J. F.; Miller, J. R.; Debinski, D. M.; Schacht, W. H. Restoring the fire–grazing interaction promotes tree–grass coexistence by controlling woody encroachment. Ecosphere 2020, 11, e02993. [Google Scholar] [CrossRef]
- Jordan, S. (2024). The Interactive Effects of Precipitation and Disturbance on the Functioning of Dryland Ecosystems as Modulated by Mean Annual Precipitation. Arizona State University.
- Wright, H. A. Range burning. Rangeland Ecology & Management/Journal of Range Management Archives 1974, 27, 1–11. [Google Scholar]
- Jones, G. M.; Tingley, M. W. Pyrodiversity and biodiversity: A history, synthesis, and outlook. Diversity and Distributions 2022, 28, 386–403 . [Google Scholar] [CrossRef]
- Biederman, J. A.; Scott, R. L.; Arnone III, J. A.; Jasoni, R. L.; Litvak, M. E.; Moreo, M. T. . & Vivoni, E. R. Shrubland carbon sink depends upon winter water availability in the warm deserts of North America. Agricultural and Forest Meteorology 2018, 249, 407–419. [Google Scholar] [CrossRef]
- Ascoli, D.; Lonati, M.; Marzano, R.; Bovio, G.; Cavallero, A.; Lombardi, G. Prescribed burning and browsing to control tree encroachment in southern European heathlands. Forest Ecology and Management 2013, 289, 69–77. [Google Scholar] [CrossRef]
- Pausas, J. G.; Pratt, R. B.; Keeley, J. E.; Jacobsen, A. L.; Ramirez, A. R.; Vilagrosa, A. . & Davis, S. D. Towards understanding resprouting at the global scale. New Phytologist 2016, 209, 945–954 . [Google Scholar] [CrossRef]
- O’Connor, R. C.; Taylor, J. H.; Nippert, J. B. Browsing and fire decreases dominance of a resprouting shrub in woody encroached grassland. Ecology 2020, 101, e02935 . [Google Scholar] [CrossRef]
- Taylor, C. 2005. Fire ecology in the central Texas and Edwards Plateau regions of Texas. 32-41 In: Symposium for land managers. Kerrville, TX.
Figure 4.
Soil temperature at 1, 3, 5, and 10 cm depth during the burning treatments (a), and soil temperature at three depths five days after fire treatment. The air temperature at three different heights over soil (b), and the temperature over the grass tussocks during the fire treatment (d).
Figure 4.
Soil temperature at 1, 3, 5, and 10 cm depth during the burning treatments (a), and soil temperature at three depths five days after fire treatment. The air temperature at three different heights over soil (b), and the temperature over the grass tussocks during the fire treatment (d).
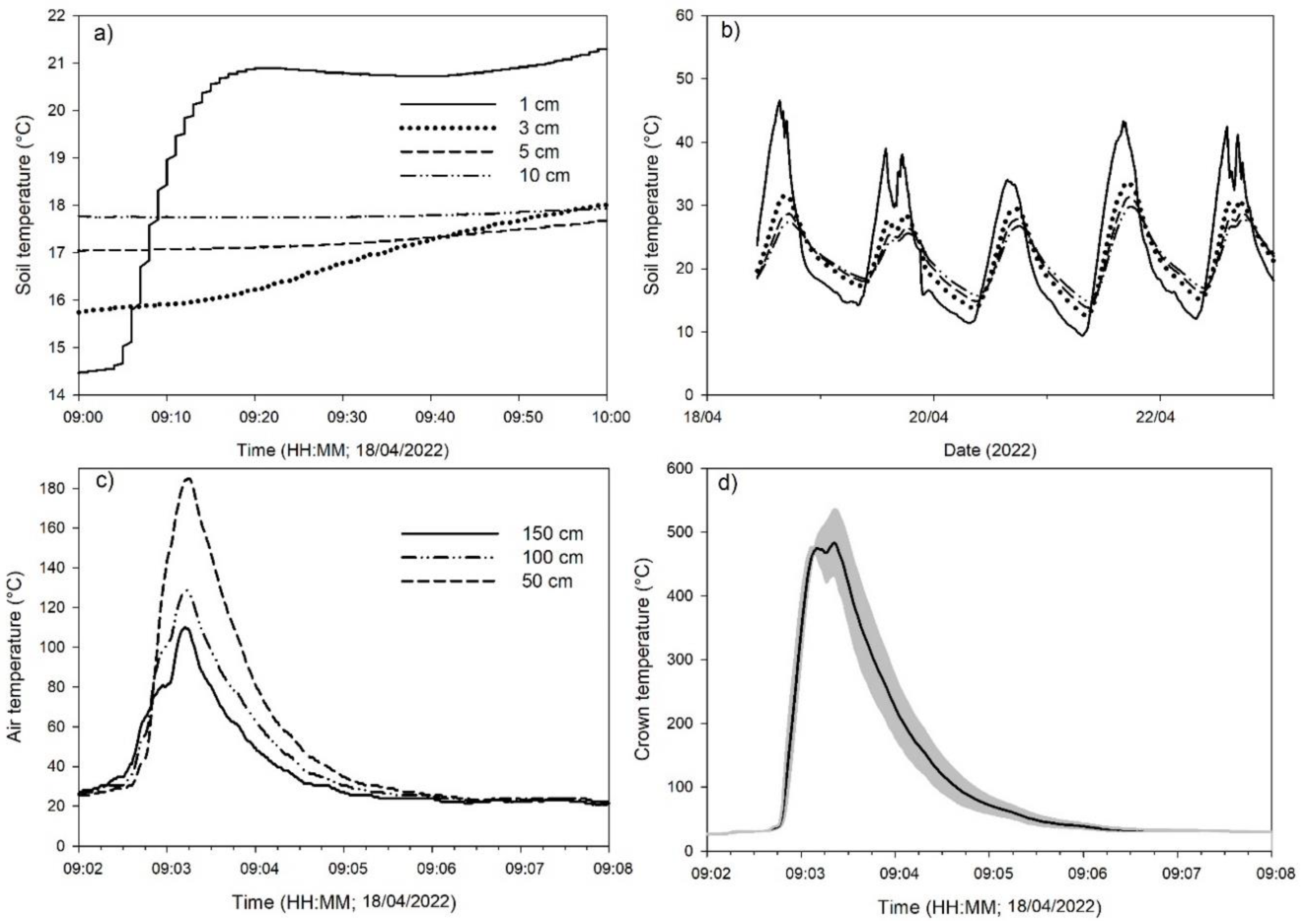
Figure 5.
Fitted functions (normalized units) of relationships between fire response and dasometric variables (crown area, base diameter (diameter at 30 cm), and total height). Number of resprouts for Mimosa biuncifera (a-c) and for Vachelia Schafneri (d-f), and the percent of fire damage for V. schafneri (g-i).
Figure 5.
Fitted functions (normalized units) of relationships between fire response and dasometric variables (crown area, base diameter (diameter at 30 cm), and total height). Number of resprouts for Mimosa biuncifera (a-c) and for Vachelia Schafneri (d-f), and the percent of fire damage for V. schafneri (g-i).
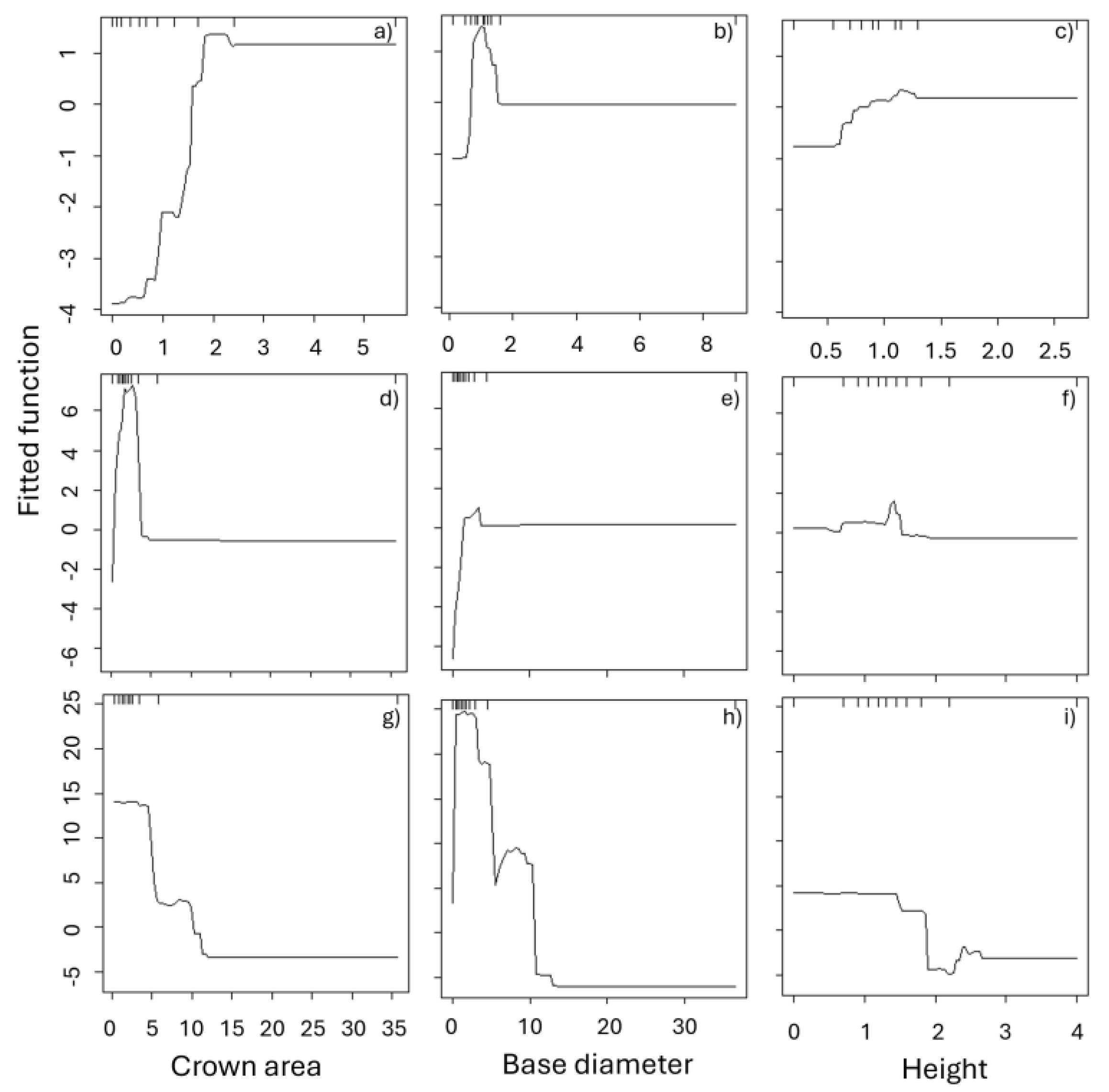

Table 1.
Relative importance of variables for explaining fire damage and number of resprouts after fire treatment in two shrub species.
Table 1.
Relative importance of variables for explaining fire damage and number of resprouts after fire treatment in two shrub species.
| Species | Explanatory variable | Relative importance (%) | |
| Fire damage | Number of resprouts | ||
| Vachellia schafneri | |||
| Crown diameter | 53.1 | 28.7 | |
| Base diameter | 24.7 | 65.2 | |
| Height | 22.2 | 6.0 | |
| Mimosa biuncifera | |||
| Crown diameter | 60.0 | ||
| Base diameter | 30.1 | ||
| Height | 9.8 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



