
Submitted:
09 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
Wildfire is a key factor in regulating ecological processes in grassland ecosystems; however, changes in land use/cover have modified the intensity and frequency of fires as they occurred naturally. Different factors have caused a rise in woody vegetation in these ecosystems, leading to changes in species composition, diversity, and biogeochemical cycles. The prescribed burns are a tool for controlling and eradicating shrubs; however, their effectiveness depends on vegetation composition, biomass availability, and the objectives of restoration. We evaluate the effectiveness of fire as a shrub controller in a semiarid grassland ecosystem. We measured several shrub dasometric parameters and the percentage of damage in ten 2000 m² plots three months after a prescribed burning was performed. Both crown height and width and total height were the main variables that explained the percentage of shrub damage by fire. Individuals with a height greater than 1.6 m and wide crowns did not suffer damage. Moreover, even though 97% of the total shrubs presented some fire damage, 86% recovered after the rain period. Our results show that fire could be an effective strategy to control early-growing shrubs, but on overgrazed arid lands it would be difficult to have enough biomass for implementing burning programs.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Study Area
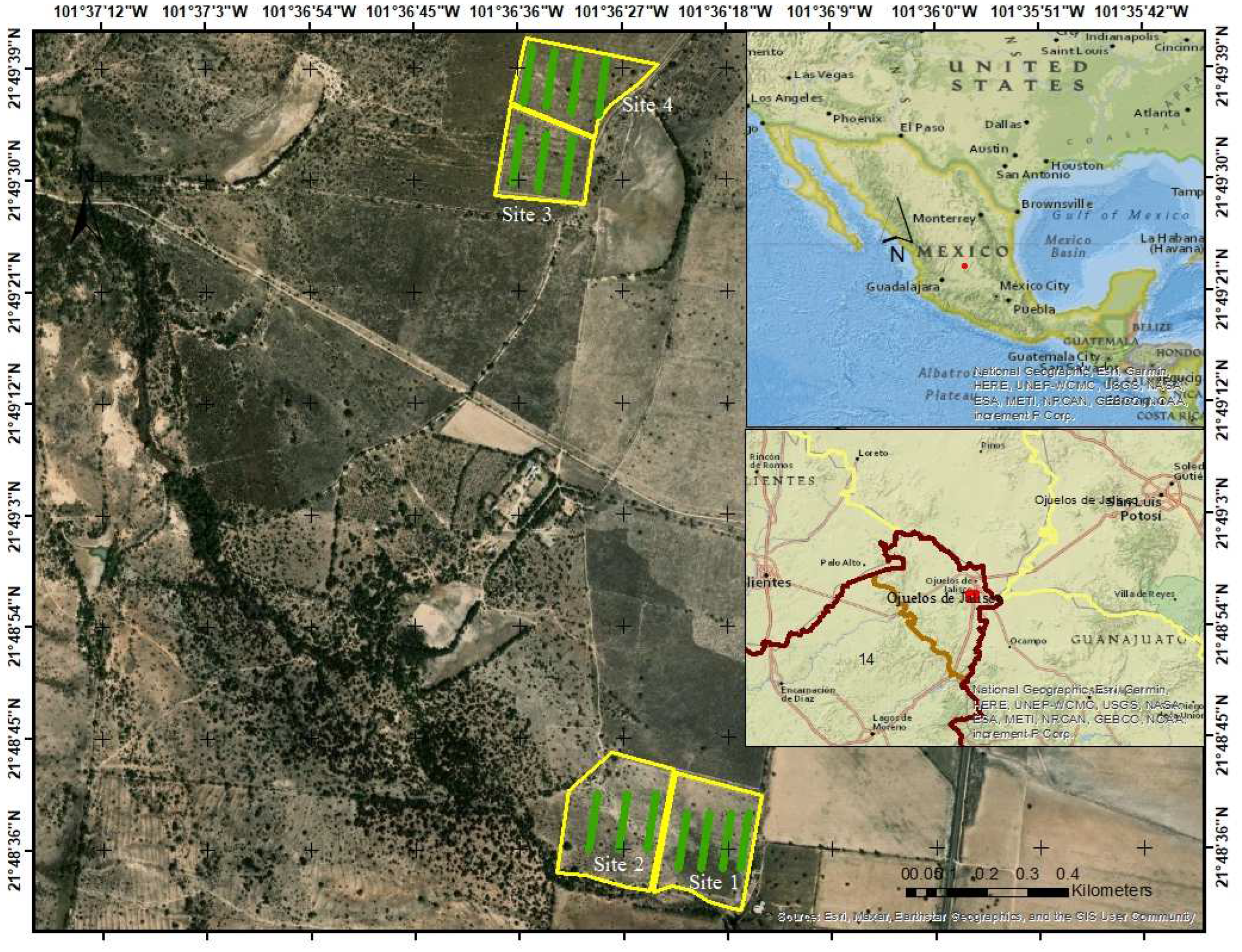
2.2. Vegetation Cover
2.3. Climate
2.4. History and Grazing Management
2.5. Experimental Design
2.6. Vegetation Sampling
2.7. Application of Prescribed Burns

2.8. Environmental Variables
2.9. Analysis of Data
3. Results
3.1. Species Composition
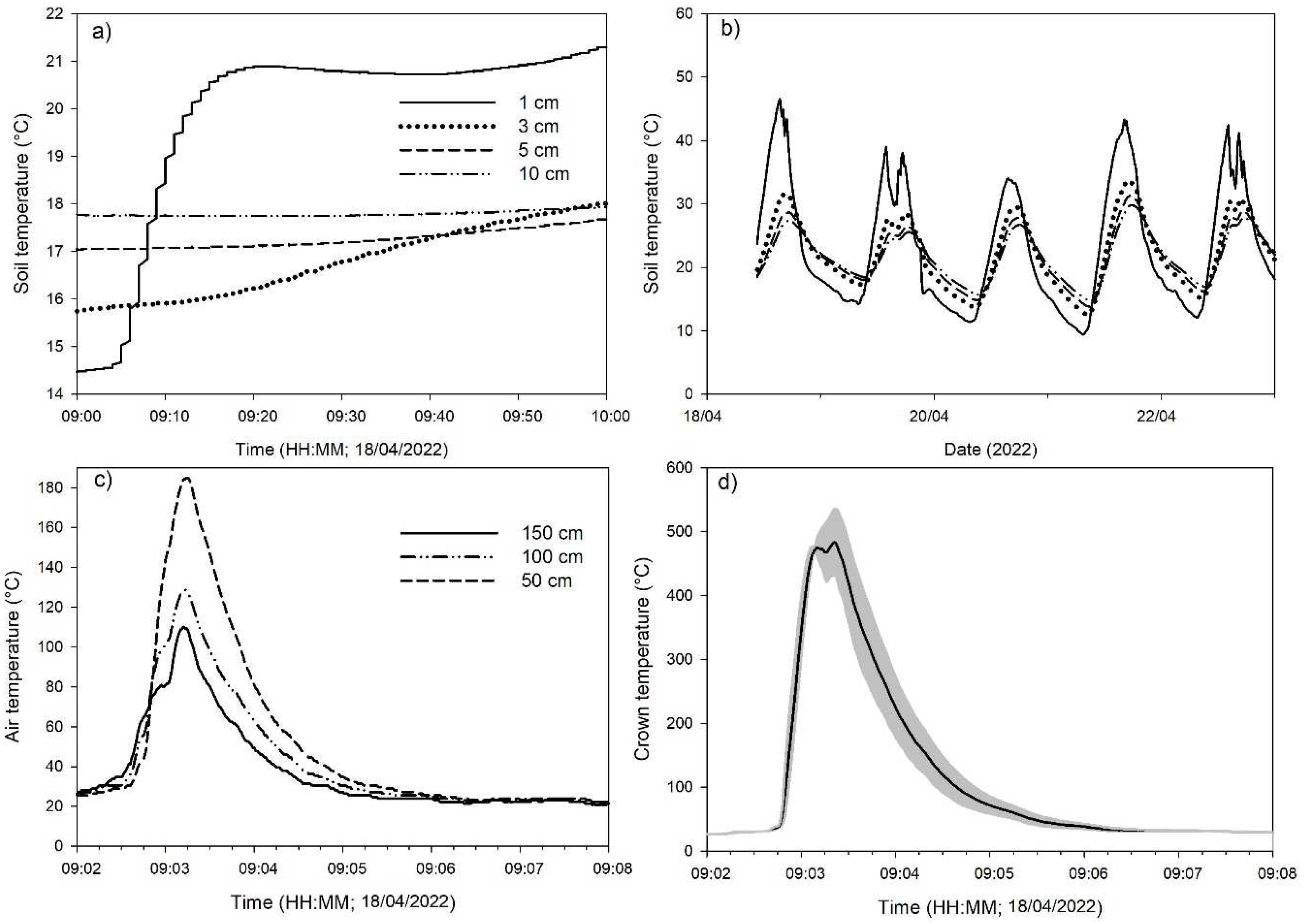
3.2. Temperatures in Prescribed Burnings
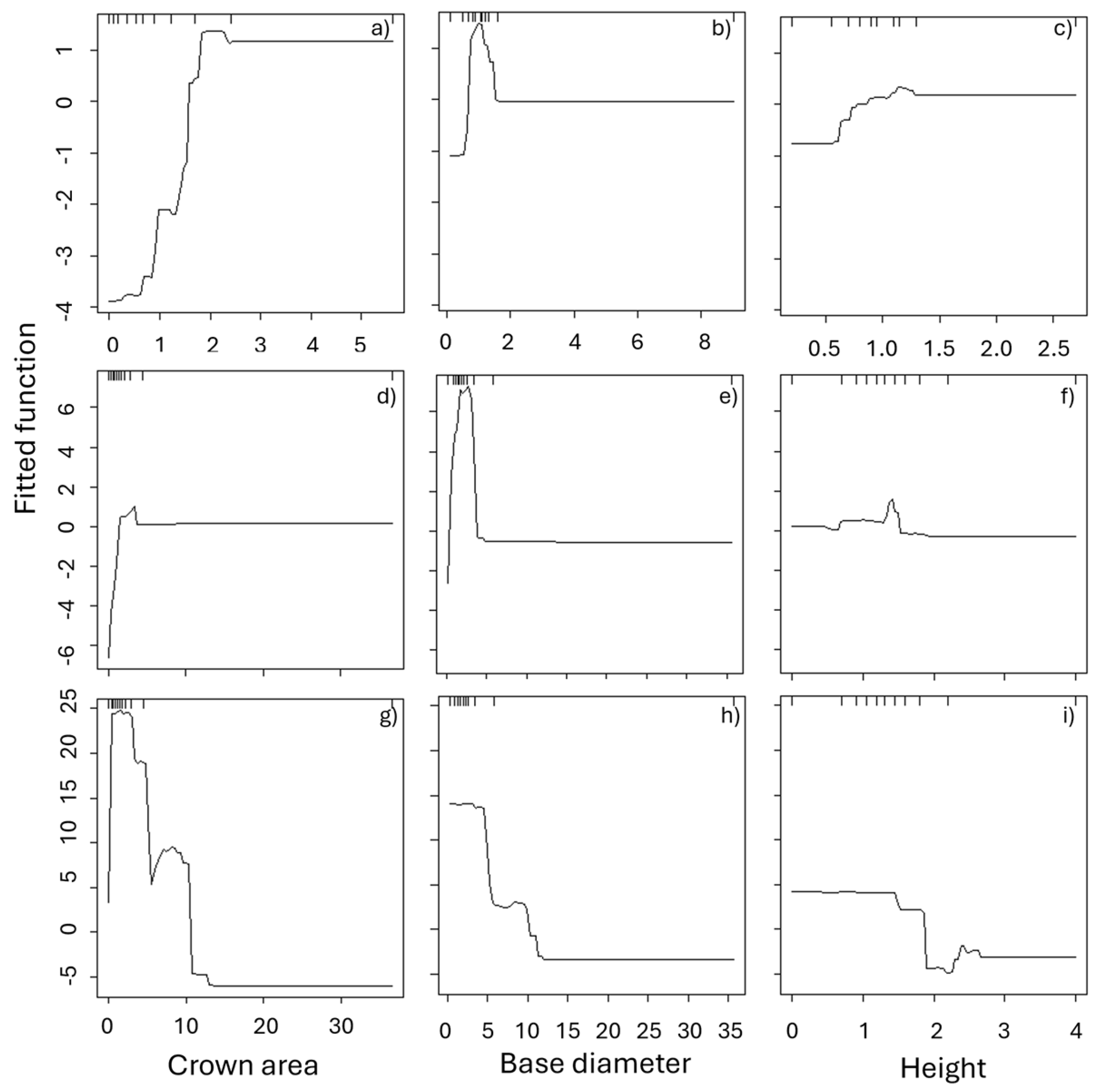
3.3. Fire Effects on Shrubs

| Species | Explanatory variable | Relative importance (%) | |
|---|---|---|---|
| Fire damage | Number of resprouts | ||
| Vachellia schafneri | |||
| Crown diameter | 53.1 | 28.7 | |
| Base diameter | 24.7 | 65.2 | |
| Height | 22.2 | 6.0 | |
| Mimosa biuncifera | |||
| Crown diameter | 60.0 | ||
| Base diameter | 30.1 | ||
| Height | 9.8 | ||
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huenneke, L.F.; Clason, D.; Muldavin, E. Spatial heterogeneity in Chihuahuan Desert vegetation: implications for sampling methods in semi-arid ecosystems. J. Arid. Environ. 2001, 47, 257–270. [Google Scholar] [CrossRef]
- Rzedowski, J. 2021 Flora of the Valley of Mexico, 2nd ed.; Institute of Ecology: Mexico.
- Barger, N.N.; Archer, S.R.; Campbell, J.L.; Huang, C.-Y.; Morton, J.A.; Knapp, A.K. Woody plant proliferation in North American drylands: A synthesis of impacts on ecosystem carbon balance. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef]
- Van Auken, O.W. Shrub Invasions of North American Semiarid Grasslands. Annu. Rev. Ecol. Syst. 2000, 31, 197–215. [Google Scholar] [CrossRef]
- Caldeira, M.C.; Lecomte, X.; David, T.S.; Pinto, J.G.; Bugalho, M.N.; Werner, C. Synergy of extreme drought and shrub invasion reduce ecosystem functioning and resilience in water-limited climates. Sci. Rep. 2015, 5, 15110–15110. [Google Scholar] [CrossRef] [PubMed]
- Kidron, G.J.; Gutschick, V.P. Soil moisture correlates with shrub–grass association in the Chihuahuan Desert. CATENA 2013, 107, 71–79. [Google Scholar] [CrossRef]
- Mata-González, R.; Figueroa-Sandoval, B.; Clemente, F.; Manzano, M. Vegetation changes after livestock grazing exclusion and shrub control in the southern Chihuahuan Desert. West. North Am. Nat. 2007, 67, 63–70. [Google Scholar] [CrossRef]
- Munson, S.M.; Muldavin, E.H.; Belnap, J.; Peters, D.P.C.; Anderson, J.P.; Reiser, M.H.; Gallo, K.; Melgoza-Castillo, A.; Herrick, J.E.; Christiansen, T.A. Regional signatures of plant response to drought and elevated temperature across a desert ecosystem. Ecology 2013, 94, 2030–2041. [Google Scholar] [CrossRef] [PubMed]
- Sepp, S.; Davison, J.; Moora, M.; Neuenkamp, L.; Oja, J.; Roslin, T.; Vasar, M.; Öpik, M.; Zobel, M. Woody encroachment in grassland elicits complex changes in the functional structure of above- and belowground biota. Ecosphere 2021, 12, e03512. [Google Scholar] [CrossRef]
- Ibarra Flores, F.A.; Martín Rivera, S.; Moreno Medina, M.; Denogean Ballesteros, F.G.; Martinez Duran, A.B.; Retes López, R.; De La Toba García, V.M.; Aguilar Valdes, A. Beneficios económicos asociados con el control de invasiones de uña de gato en el pastizal mediano abierto de Cananea, Sonora, México. Revista Mexicana de Agronegocios 2014, 34. [Google Scholar] [CrossRef]
- Osborn, S.; Wright, V. Understanding and managing invasive plants in wilderness and other natural areas: An annotated reading list. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, 2002; Volume 4. [Google Scholar]
- Rego, F.C.; Rigolot, E.; Alexandrian, D.; Fernandes, P. EU project FIRE PARADOX: Moving towards integrated Fire Management.
- Wright, H.A.; Bailey, A.W. Fire Ecology. John Wiley & Sons, Inc.: United States and Southern Canada, 1982. [Google Scholar]
- Nippert, J.B.; Telleria, L.; Blackmore, P.; Taylor, J.H.; O'Connor, R.C. Is a Prescribed Fire Sufficient to Slow the Spread of Woody Plants in an Infrequently Burned Grassland? A Case Study in Tallgrass Prairie. Rangel. Ecol. Manag. 2021, 78, 79–89. [Google Scholar] [CrossRef]
- Hahn, G.E.; Coates, T.A.; Aust, W.M. Soil Chemistry following Single-Entry, Dormant Season Prescribed Fires in the Ridge and Valley Province of Virginia, USA. Commun. Soil Sci. Plant Anal. 2021, 52, 2065–2073. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Ocheltree, T.W. Abrupt transition of mesic grassland to shrubland: evidence for thresholds, alternative attractors, and regime shifts. Ecology 2014, 95, 2633–2645. [Google Scholar] [CrossRef]
- Ratajczak, Z.; D'Odorico, P.; Nippert, J.B.; Collins, S.L.; Brunsell, N.A.; Ravi, S. Changes in spatial variance during a grassland to shrubland state transition. J. Ecol. 2016, 105, 750–760. [Google Scholar] [CrossRef]
- INEGI. Conjunto de datos vectorial edafológico Escala 1:250000. Serie II. Continuo Nacional; Instituto Nacional de Estadística y Geografía: Aguascalientes, México, 2007. [Google Scholar]
- WRB. World Reference Base for Soil Resources 2015. International soil classification system for naming soils and creating legends for soil maps, 106. FAO: Rome, Italy, 203 pp.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 11 December 2023).
- Rangeland Systems; Springer Nature: Dordrecht, GX, Netherlands, 2017.
- Ludwig, F.; de Kroon, H.; Prins, H.H.; Berendse, F. Effects of nutrients and shade on tree-grass interactions in an East African savanna. J. Veg. Sci. 2001, 12, 579–588. [Google Scholar] [CrossRef]
- Harrington, J.A.; Kathol, E. Responses of Shrub Midstory and Herbaceous Layers to Managed Grazing and Fire in a North American Savanna (Oak Woodland) and Prairie Landscape. Restor. Ecol. 2009, 17, 234–244. [Google Scholar] [CrossRef]
- Bennett, L.; Judd, T.; Adams, M. Growth and nutrient content of perennial grasslands following burning in semi-arid, sub-tropical Australia. Plant Ecol. 2003, 164, 185–199. [Google Scholar] [CrossRef]
- Wells, A.G.; Munson, S.M.; Sesnie, S.E.; Villarreal, M.L. Remotely Sensed Fine-Fuel Changes from Wildfire and Prescribed Fire in a Semi-Arid Grassland. Fire 2021, 4, 84. [Google Scholar] [CrossRef]
- Holdo, R.M.; Brocato, E.R. Tree–grass competition varies across select savanna tree species: a potential role for rooting depth. Plant Ecol. 2015, 216, 577–588. [Google Scholar] [CrossRef]
- Pierce, N.A.; Archer, S.R.; Bestelmeyer, B.T. Competition suppresses shrubs during early, but not late, stages of arid grassland–shrubland state transition. Funct. Ecol. 2019, 33, 1480–1490. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. The Evolutionary Ecology of Sprouting in Woody Plants. Int. J. Plant Sci. 2003, 164, S103–S114. [Google Scholar] [CrossRef]
- Li, J.; Ravi, S.; Wang, G.; Van Pelt, R.S.; Gill, T.E.; Sankey, J.B. Woody plant encroachment of grassland and the reversibility of shrub dominance: Erosion, fire, and feedback processes. Ecosphere 2022, 13. [Google Scholar] [CrossRef]
- Weber-Grullon, L.; Gherardi, L.; Rutherford, W.A.; Archer, S.R.; Sala, O.E. Woody-plant encroachment: Precipitation, herbivory, and grass-competition interact to affect shrub recruitment. Ecol. Appl. 2022, 32, e2536. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Will, R.; Zhai, L.; Zou, C. Future Climate Change Shifts the Ranges of Major Encroaching Woody Plant Species in the Southern Great Plains, USA. Earth's Futur. 2024, 12. [Google Scholar] [CrossRef]
- Scholtz, R.; Smit, I.P.J.; Coetsee, C.; Kiker, G.A.; Venter, F.J. Legacy effects of top-down disturbances on woody plant species composition in semi-arid systems. Austral Ecol. 2016, 42, 72–83. [Google Scholar] [CrossRef]
- Capozzelli, J.F.; Miller, J.R.; Debinski, D.M.; Schacht, W.H. Restoring the fire–grazing interaction promotes tree–grass coexistence by controlling woody encroachment. Ecosphere 2020, 11, e02993. [Google Scholar] [CrossRef]
- Jordan, S. The Interactive Effects of Precipitation and Disturbance on the Functioning of Dryland Ecosystems as Modulated by Mean Annual Precipitation. Arizona State University, 2024. [Google Scholar]
- Wright, H.A. Range burning. Rangeland Ecology & Management. Journal of Range Management Archives 1974, 27, 1–11. [Google Scholar]
- Jones, G.M.; Tingley, M.W. Pyrodiversity and biodiversity: A history, synthesis, and outlook. Divers. Distrib. 2021, 28, 386–403. [Google Scholar] [CrossRef]
- Biederman, J.A.; Scott, R.L.; Iii, J.A.A.; Jasoni, R.L.; Litvak, M.E.; Moreo, M.T.; Papuga, S.A.; Ponce-Campos, G.E.; Schreiner-McGraw, A.P.; Vivoni, E.R. Shrubland carbon sink depends upon winter water availability in the warm deserts of North America. Agric. For. Meteorol. 2018, 249, 407–419. [Google Scholar] [CrossRef]
- Ascoli, D.; Lonati, M.; Marzano, R.; Bovio, G.; Cavallero, A.; Lombardi, G. Prescribed burning and browsing to control tree encroachment in southern European heathlands. For. Ecol. Manag. 2013, 289, 69–77. [Google Scholar] [CrossRef]
- Pausas, J.G.; Pratt, R.B.; Keeley, J.E.; Jacobsen, A.L.; Ramirez, A.R.; Vilagrosa, A.; Paula, S.; Kaneakua-Pia, I.N.; Davis, S.D. Towards understanding resprouting at the global scale. New Phytol. 2015, 209, 945–954. [Google Scholar] [CrossRef]
- O’connor, R.C.; Taylor, J.H.; Nippert, J.B. Browsing and fire decreases dominance of a resprouting shrub in woody encroached grassland. Ecology 2019, 101, e02935. [Google Scholar] [CrossRef]
- Taylor, C. Fire ecology in the central Texas and Edwards Plateau regions of Texas. 32-41. Symposium for Land Managers: Kerrville, TX, 2005. [Google Scholar]
| Species | Biomass contribution (%) |
|---|---|
| Living grass standing biomass | 63.305 |
| Dalea Bicolor | 14.961 |
| Bidens sp. | 6.936 |
| Dead grass standing biomass | 6.545 |
| No ID | 3.837 |
| Plantago lanceolata | 2.695 |
| Stevia serrata | 1.075 |
| Euphorbia | 0.243 |
| Paspalum sp. | 0.190 |
| Zornia reticulata | 0.079 |
| Phaseolus sp. | 0.057 |
| Dichondra sp. | 0.041 |
| Tagetes sp. | 0.013 |
| Cyperus sp. | 0.012 |
| Sida sp. | 0.010 |
| Vachellia schaffneri | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



