Submitted:
03 December 2024
Posted:
04 December 2024
You are already at the latest version
Abstract
Chlorophyll is the primary pigment responsible for capturing light energy during photosynthesis, while carotenoids assist in light absorption and provide photoprotection by dissipating excess energy. Both pigments are essential for plant growth and development, playing distinct and complementary roles in maintaining photosynthetic efficiency and protecting plants from oxidative stress. Because of their function in photosynthesis and photoprotection, chlorophyll and carotenoid accumulation are strongly associated with light conditions, especially blue and red lights which play key roles in regulating their metabolisms. Despite advancements in understanding pigment metabolism, there remains a limited comprehensive overview of how various light qualities influence these pathways throughout the entire process. The effects of other light qualities, such as green light, far-red light, and UV, are not yet fully understood. This review aims to synthesize recent findings about the regulatory network of chlorophyll and carotenoid pathways under different light spectra, emphasizing the interplay between light- regulated transcription factors and genes involved in their biosynthesis and degradation.
Keywords:
chlorophyll
; carotenoid
; light spectra
1. Introduction
Chlorophylls are important molecules in photosynthetic processes and are especially responsible for light energy absorption. Chlorophyll absorbs photons in light sources, excites electrons to a higher energy state, and transfers energy into the photosystem, driving the process of photosynthesis. In plants, chlorophyll metabolism is highly coordinated with a series of cooperative reactions and the involvement of various transcription regulators. Chlorophylls are initially biosynthesized from glutamate, then converted to 5-aminolevulinic acid (ALA) and further to protochlorophyllide (Pchlide) by numerous genes such as HEMA1 (a glutamyl-tRNA reductase protein) and CHCL (protoporphyrin IX magnesium chelatase, subunit H) [1]. In plants growing under dark conditions, chlorophyll biosynthesis halts at the step of Pchlide, the immediate precursor of chlorophyllide (Chlide), while light triggers the photoactivity of NADPH-protochlorophyllide oxidoreductase (PORs), catalyzing the conversion of PChlide to Chlide [1,2], and follows by chlorophyll cycle to generate chlorophyll a and chlorophyll b. In the degradation process, chlorophyll breaks down into non-toxic compounds, primarily through the actions of enzymes like chlorophyll b reductase (CBR) and pheophorbide a oxygenase (PAO)[3]. Regulation of the levels of chlorophyll and its derivatives is extremely important. Chlorophyll metabolism not only affects the photosynthetic machinery, but it is also involved in other growth processes such as programmed cell death, the ‘stay-green’ phenomenon, and chloroplast-nucleus communication [4]. Besides, over-accumulated Pchlide, a strong photosensitizer, can lead to the generation of reactive oxygen species (ROS), which accelerate growth retardation or cell death [2,4]. Chlorophyll degradation prevents the accumulation of potentially harmful byproducts and is often triggered by stress or aging [3].
Carotenoids are isoprenoid pigments which are essential for plant life. In plants, they provide yellow, orange, and red colors to leaves, flowers, fruits, and some roots [5]. From the MEP pathway, carotenoid biosynthesis begins with the production of precursor molecules like geranylgeranyl diphosphate (GGPP), which undergoes multiple enzyme-driven steps to form carotenoids such as lutein, zeaxanthin, and β-carotene [6]. Key enzymes in this pathway include phytoene synthase (PSY), phytoene desaturase (PDS), and lycopene β-cyclase (LYCB). The biosynthesis process is tightly regulated by transcription factors responsive to light and environmental cues, promoting carotenoid accumulation, particularly under red and blue light [7]. In the context of carotenoid degradation, carotenoids are broken down by carotenoid cleavage dioxygenases (CCDs), converting them into smaller molecules like abscisic acid (ABA) and strigolactones [8]. These degradation products serve as essential molecules in plant growth, stress responses, and signaling. The balance between biosynthesis and degradation ensures that carotenoids support light absorption, photoprotection, and growth regulation while avoiding excess accumulation that could disrupt cellular processes. Apart from providing colors, carotenoids play an important role in protecting photosynthetic apparatus from photooxidative damage [6]. Carotenoids bind with chlorophylls in photosystem (PS) I and PSII and are arranged in the lipid bilayers of the thylakoid membranes, therefore inhibiting uncontrolled interaction of excited pigments with oxygen that can generate ROS. Additionally, carotenoids act as accessory pigments, absorb light spectra that are outside of the chlorophyll absorption range, and provide extra energy for photosynthesis.
Because of their role in photosynthesis and photoprotection, chlorophyll and carotenoid accumulation are strongly associated with light conditions. Light is one of the important factors in controlling the growth and development of plants and absorption of water and nutrients. The sensing of light in plants is conducted by various light photoreceptors, such as phytochrome (phy), cryptochrome (cry), and UV RESISTANCE LOCUS8 (UVR8) [9]. By perceiving light at different compositions, plants can adjust growth through physiological processes such as photosynthesis, accumulation and transport of photoproduct [10]. Therein, modulating plant pigment metabolism, including chlorophylls and carotenoids, is an essential mechanism to respond to different light spectra (Figure 1). Blue and red lights are critical attention in understanding physiological mechanisms of chlorophylls and carotenoids due to the maximum absorption wavelength, and thus directly lead to the accumulation of these pigments [11]
Despite recent studies on pigment metabolism, there is limited comprehensive overview of how different light spectra influence these pathways, particularly the interaction between transcription factors and key metabolic genes. Especially with carotenoids, many studies reveal a large number of regulators in the transcriptional control of carotenogenic genes [5,12,13,14,15], however, these results provided fragmented and unassociated insights, and some of TFs mechanism and function in carotenoid pathways still remain unverified, e.g., NAC, MYB [7]. The objective of this review is to share recent knowledge of the effect of different light qualities on chlorophyll and carotenoid pathways, focusing on synthesizing the interplay between light-mediate regulators and biosynthesis- and degradation-involved genes.
2. Chlorophyll Pathway Under Different Light Spectra
Different light spectra were found to be directly involved in the start of the chlorophyll biosynthesis pathway, in which LONG HYPOCOTYL 5 (HY5) and GOLDEN 2-LIKE (GLK) play vital role in the convergence of blue and red lights (Figure 2). In the dark, the COP1-SPA E3 ligase complex directly enhances HY5 degradation, resulting in a low level of HY5 in plants. With the onset of light, Cryptochromes 1/2 (Cry1/2) inhibits COP1-SPA complex formation and compete with HY5 to COP1 binding, thus increasing the HY5 accumulation [16,17]. HY5 directly binding to the G-box of GLK promoter and enhances its expression level [18]. Two start genes HEMA1, encoding for glutamyl-tRNA reductase (GluTR) activity, and CHLH, responsible for magnesium insertion into the protoporphyrin IX ring in chlorophyll branch, are both positively enhanced by HY5 and GLK [19,20]. On the other hand, monochromatic red light showed a more complex effect on chlorophyll metabolism through activities of phytochrome B (PhyB), a red-light receptor (Figure 3). PhyA/B is also involved in disrupting the formation of the COP1-SPA complex, therefore, regulating the activity of HY5 [16]. Besides, PhyB is an inactive repressor of light responses such as phytochrome-interacting factor 1 and 3 (PIF1, PIF3), which depress either HEMA1 or CHLH, thus partly inducing the start of chlorophyll biosynthesis [21,22]. However, in the later pathway, red light acts as a negative regulator for the chlorophyll cycle. Three NADPH-protochlorophyllide oxidoreductase (POR) genes, PORA, PORB, and PORC, induce the conversion of Phlide to Chlide under light conditions, however, are downregulated by red light through the inhibiting activity of their positive regulator such as RVE1, PIF1, and EIN3/EIL1 [23,24,25]. PhyB takes the role as a repressor of EIN3/EIL1 and PIF4/5, resulting in depressing chlorophyll reduction genes like NYC1, NYE1, and PAO [3,26]. Interestingly, the combination of red and blue light significantly increased the expression levels of genes involved in the chlorophyll cycle and degradation such as CAO, CBR, and PAO (with a ratio of red to blue light at 3:1), PPH (at a 1:1 ratio) [20]. By contrast, white light significantly boosted the expression level of CBR compared to other red-blue combinations [20], suggesting a possible role for green light in regulating downstream genes. In plants, green light can be absorbed by the lower leaves in complex capony, involved in plant responses to shade, which causes increased synthesis of chlorophyll b [27,28]. However, the effect of green light on chlorophyll pathways remains unclear and requires further investigation.
Recent research suggested that UV-B light also upregulated the expression of upstream genes. In tea leaves, UV-B irradiation enhanced the activity of CsGLK1/2, directly leading to higher levels of its target genes CsHEMA1, CsCHLH and CsPORA [29]. Although the expression of CSGLK1/2 decreased significantly under UV-A, that of CsPORA increases under both UV-A and UV-B lights. Thus, revealing the possible existence of other regulators involved in UV light signal transduction pathways that are responsible for control CsPORA expression.
3. Carotenoid Pathway Under Different Light Spectra
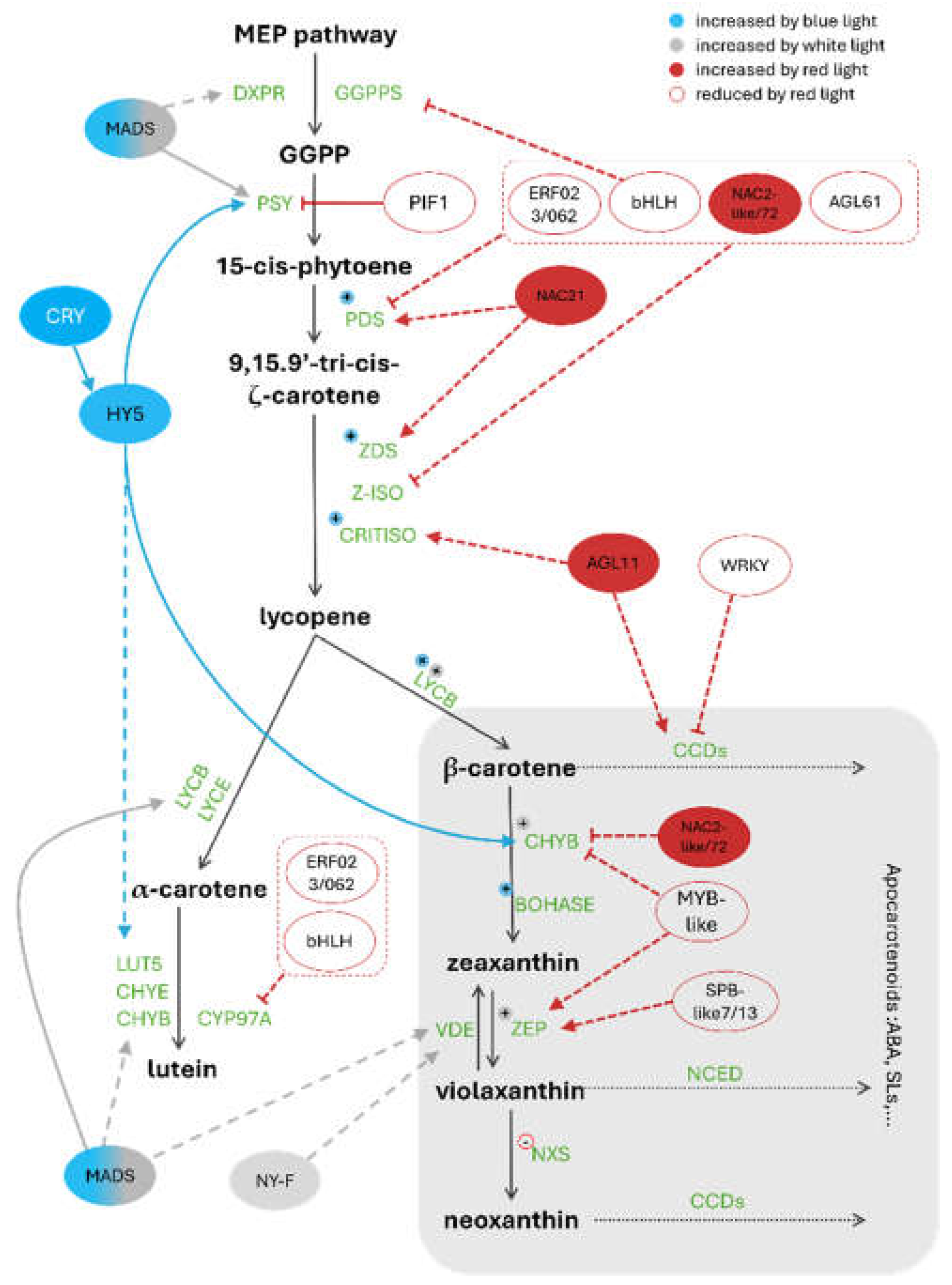
Red light is positively involved in regulating carotenoid accumulation through a complicated interplay between various transcription factors and carotenoid biosynthesis- and cleavage-related genes (Figure 3). Transcriptome analysis revealed that transcription factors from different families, such as bHLH, NAC, MYB AGL, ERF, WRKY, SPB and PIF, directly or indirectly modulates the expression of carotenogenic genes, including GGPPS2, PDS, Z-ISO, ZDS2/7, CRTISO3, CYP97A, CHYB, ZEP2, CCD1-2 under red light treatment [12,30]. Red light downregulates some transcription factors that are negatively correlated with carotenoid biosynthesis genes, such as bHLH128 as a GGPPS- and Z-ISO-depressor, ERF023/062 and AGL61 as PDS- and Z-ISO-depressor, MYB-like and SBPlike-7/13 as ZEP-depressor [14,30]. In addition, phyB inhibits PIF1, which directly targets to the promoter of phytoene synthase gene (PSY) and reduces its expression level [31,32]. In other ways, red light enhances the positive regulators like NAC21 and AGL11, both induced lycopene formation [33]. Carotenoid cleavage dioxygenases (CCDs), which catalyzed carotenoid breakdowns to apocarotenoids, were found to be regulated by WRKY transcription factors in Osmanthus fragrans [34]. Under red light treatment, WRKY was downregulated in the ripening grapefruit which was accompanied by the reduction of CCD1-2 transcription expression.
Compared to red light and dark conditions, blue and white light were reported to significantly increase total carotenoid accumulation, with the greatest enhancements of lutein and β-carotene concentrations [12]. Both these light qualities upregulate the activity of HY5 and MADS, which were positively active upstream genes like DXPR and PSY [32,35,36]. HY5 can directly bind to the CHYB promoter and increase its expression. HY5 and MADS also were reported to be positively correlated with increased expression of key carotenoid biosynthesis genes, including LYCB, LYCE, LUT5, CHYB, and βOHASE. MAD and HY5 can directly bind to LYCB and CHYB promoters, respectively, and increase their expression [37]. In maize sprouts, blue light promoted the upregulation of PDS, ZDS, CRITISO, while white light significantly upregulated genes like CHXB, ZEP and VDE through the activity of MADS and NY-F compared to red or blue light only [12], leading to higher levels of lutein, zeaxanthin, and β-carotene.
4. Conclusions and Perspective
Overall, chlorophyll and carotenoid metabolisms are strongly influenced by light spectra, with blue and red lights playing key roles in regulating their biosynthesis and degradation. Blue light promotes chlorophyll accumulation, while the effect of red light varies with its combination with blue light. The combination of red and blue lights is recognized as enhancing an abundance in carotenoid
Understanding the interplay between light spectra and plant pigments has transformative applications in agriculture production. Chlorophyll and carotenoids play vital roles in maintaining photosynthetic efficiency and protecting plants from oxidative stress, and indirectly involved in other light-induce processes. With the use of artificial lighting as a supplement or alternative to natural light, farmers nowadays can tailor their lighting system to enhance specific light spectrums, such as blue or UV light. Therefore, modulating chlorophyll and carotenoid pathways through light regime could enable precise control over plant development and stress tolerance, thus promoting crop yield and quality.
These insights also support the enhancement of nutritional and aesthetic qualities in crops during the cultivation and and storage period. For instance, in vegetable leaves, blue light irradiation can increase the carotenoid content, especially lutein and β-carotene, which provide vitamin A and act as antioxidants in the human body [6]. Post-harvest applications include using specific light treatments to stabilize pigments, and maintaining color, flavor, and nutritional value during storage and transport [38,39].
Author Contributions
writing—original draft preparation, TNP.N; writing—review and editing, J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the “Research Program for Agriculture Science & Technology Development (Project No. RS-2023-00215864)”, Rural Development Administration, Republic of Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tripathy, B.C.; Pattanayak, G.K. Chlorophyll Biosynthesis in Higher Plants. In; 2012; pp. 63–94.
- Reinbothe, C.; Bakkouri, M. El; Buhr, F.; Muraki, N.; Nomata, J.; Kurisu, G.; Fujita, Y.; Reinbothe, S. Chlorophyll Biosynthesis: Spotlight on Protochlorophyllide Reduction. Trends Plant Sci 2010, 15, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, J.; Qiu, K.; Kuai, B. Phytohormone and Light Regulation of Chlorophyll Degradation. Front Plant Sci 2017, 8. [Google Scholar] [CrossRef]
- Tanaka, A.; Tanaka, R. Chlorophyll Metabolism. Curr Opin Plant Biol 2006, 9, 248–255. [Google Scholar] [CrossRef]
- Quian-Ulloa, R.; Stange, C. Carotenoid Biosynthesis and Plastid Development in Plants: The Role of Light. International Journal of Molecular Sciences 2021, 22, 1184. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, J.; Ramanan, R.N.; Raghunandan, M.E.; Galanakis, C.M.; Krishnamurthy, N.P. Carotenoids. In Nutraceutical and Functional Food Components: Effects of Innovative Processing Techniques; Elsevier Inc., 2017; pp. 259–296. ISBN 9780128052570. [Google Scholar]
- Stanley, L.; Yuan, Y.-W. Transcriptional Regulation of Carotenoid Biosynthesis in Plants: So Many Regulators, So Little Consensus. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Mol Plant 2015, 8, 68–82. [Google Scholar] [CrossRef]
- Kong, S.-G.; Okajima, K. Diverse Photoreceptors and Light Responses in Plants. J Plant Res 2016, 129, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Eng, C.; Wei, Z.; Teo, N.; Shen, L.; Yu, H. Seeing the Lights for Leafy Greens in Indoor Vertical Farming. Trends Food Sci Technol 2020, 106, 48–63. [Google Scholar] [CrossRef]
- Huché-Thélier, L.; Crespel, L.; Gourrierec, J. Le; Morel, P.; Sakr, S.; Leduc, N. Light Signaling and Plant Responses to Blue and UV Radiations—Perspectives for Applications in Horticulture. Environ Exp Bot 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Xiang, N.; Zhao, Y.; Wang, S.; Guo, X. The Modulation of Light Quality on Carotenoids in Maize (Zea Mays L.) Sprouts. Food Chemistry: Molecular Sciences 2022, 5. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, L.; Stange, C. Light-Dependent Regulation of Carotenoid Biosynthesis in Plants. Cienc Investig Agrar 2009, 36. [Google Scholar] [CrossRef]
- Mohanty, B.; Lakshmanan, M.; Lim, S.-H.; Kim, J.K.; Ha, S.-H.; Lee, D.-Y. Light-Specific Transcriptional Regulation of the Accumulation of Carotenoids and Phenolic Compounds in Rice Leaves. Plant Signal Behav 2016, 11, e1184808. [Google Scholar] [CrossRef]
- Llorente, B.; Martinez-garcia, J.F.; Stange, C.; Rodriguez-concepcion, M. Illuminating Colors : Regulation of Carotenoid Biosynthesis and Accumulation by Light. Curr Opin Plant Biol 2017, 37, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chu, L.; Zhang, Y.; Bian, Y.; Xiao, J.; Xu, D. HY5: A Pivotal Regulator of Light-Dependent Development in Higher Plants. Front Plant Sci 2022, 12. [Google Scholar] [CrossRef]
- Li, Y.; Shi, Y.; Li, M.; Fu, D.; Wu, S.; Li, J.; Gong, Z.; Liu, H.; Yang, S. The CRY2–COP1–HY5–BBX7/8 Module Regulates Blue Light-Dependent Cold Acclimation in Arabidopsis. Plant Cell 2021, 33, 3555–3573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, R.; Zeng, X.-Y.; Lee, S.; Ye, L.-H.; Tian, S.-L.; Zhang, Y.-J.; Busch, W.; Zhou, W.-B.; Zhu, X.-G.; et al. GLK Transcription Factors Accompany ELONGATED HYPOCOTYL5 to Orchestrate Light-Induced Seedling Development in Arabidopsis. Plant Physiol 2024, 194, 2400–2421. [Google Scholar] [CrossRef] [PubMed]
- McCormac, A.C.; Terry, M.J. Light-signalling Pathways Leading to the Co-ordinated Expression of HEMA1 and Lhcb during Chloroplast Development in Arabidopsis Thaliana. The Plant Journal 2002, 32, 549–559. [Google Scholar] [CrossRef]
- Liu, X.; Yang, M.; Xie, X.; ABM, K.; ATAK, A.; Zhong, C.; Li, D. Effect of Light on Growth and Chlorophyll Development in Kiwifruit Ex Vitro and in Vitro. Sci Hortic 2022, 291, 110599. [Google Scholar] [CrossRef]
- Stephenson, P.G.; Fankhauser, C.; Terry, M.J. PIF3 Is a Repressor of Chloroplast Development. Proceedings of the National Academy of Sciences 2009, 106, 7654–7659. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.; Zhong, S. Interplay between Light and Plant Hormones in the Control of Arabidopsis Seedling Chlorophyll Biosynthesis. Front Plant Sci 2017, 8. [Google Scholar] [CrossRef]
- Jiang, Z.; Xu, G.; Jing, Y.; Tang, W.; Lin, R. Phytochrome B and REVEILLE1/2-Mediated Signalling Controls Seed Dormancy and Germination in Arabidopsis. Nat Commun 2016, 7, 12377. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Shen, X.; Liu, R.; Xue, C.; Wei, N.; Deng, X.W.; Zhong, S. The Red Light Receptor Phytochrome B Directly Enhances Substrate-E3 Ligase Interactions to Attenuate Ethylene Responses. Dev Cell 2016, 39, 597–610. [Google Scholar] [CrossRef]
- Zhong, S.; Zhao, M.; Shi, T.; Shi, H.; An, F.; Zhao, Q.; Guo, H. EIN3/EIL1 Cooperate with PIF1 to Prevent Photo-Oxidation and to Promote Greening of Arabidopsis Seedlings. Proceedings of the National Academy of Sciences 2009, 106, 21431–21436. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Jeong, J.; Kang, M.-Y.; Kim, J.; Paek, N.-C.; Choi, G. Phytochrome-Interacting Transcription Factors PIF4 and PIF5 Induce Leaf Senescence in Arabidopsis. Nat Commun 2014, 5, 4636. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front Plant Sci 2021, 12. [Google Scholar] [CrossRef]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t Ignore the Green Light: Exploring Diverse Roles in Plant Processes. J Exp Bot 2017, 68, 2099–2110. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cheng, X.; Cao, J.; Zhu, W.; Wan, X.; Liu, L. GOLDEN 2-LIKE Transcription Factors Regulate Chlorophyll Biosynthesis and Flavonoid Accumulation in Response to UV-B in Tea Plants. Hortic Plant J 2023, 9, 1055–1066. [Google Scholar] [CrossRef]
- Huang, X.; Hu, L.; Kong, W.; Yang, C.; Xi, W. Red Light-Transmittance Bagging Promotes Carotenoid Accumulation of Grapefruit during Ripening. Commun Biol 2022, 5, 1–10. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Rodríguez-Concepción, M. Direct Regulation of Phytoene Synthase Gene Expression and Carotenoid Biosynthesis by Phytochrome-Interacting Factors. Proceedings of the National Academy of Sciences 2010, 107, 11626–11631. [Google Scholar] [CrossRef]
- XIE, B. xing; WEI, J. jing; ZHANG, Y. ting; SONG, S. wei; SU, W.; SUN, G. wen; HAO, Y. wei; LIU, H. cheng Supplemental Blue and Red Light Promote Lycopene Synthesis in Tomato Fruits. J Integr Agric 2019, 18, 590–598. [Google Scholar] [CrossRef]
- Fu, C.; Han, Y.; Kuang, J.; Chen, J.; Lu, W. Papaya CpEIN3a and CpNAC2 Co-Operatively Regulate Carotenoid Biosynthesis-Related Genes CpPDS2/4, CpLCY-e and CpCHY-b During Fruit Ripening. Plant Cell Physiol 2017, 58, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wu, M.; Cao, L.; Yuan, W.; Dong, M.; Wang, X.; Chen, W.; Shang, F. Characterization of OfWRKY3, a Transcription Factor That Positively Regulates the Carotenoid Cleavage Dioxygenase Gene OfCCD4 in Osmanthus Fragrans. Plant Mol Biol 2016, 91, 485–496. [Google Scholar] [CrossRef]
- Quian-Ulloa, R.; Stange, C. Carotenoid Biosynthesis and Plastid Development in Plants: The Role of Light. Int J Mol Sci 2021, 22, 1184. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Ye, J.; Zhu, K.; Zhang, Y.; Zhang, M.; Xu, Q.; Deng, X. A Fruit Ripening-Associated Transcription Factor CsMADS5 Positively Regulates Carotenoid Biosynthesis in Citrus. J Exp Bot 2021, 72, 3028–3043. [Google Scholar] [CrossRef]
- Zhu, K.; Chen, H.; Mei, X.; Lu, S.; Xie, H.; Liu, J.; Chai, L.; Xu, Q.; Wurtzel, E.T.; Ye, J.; et al. Transcription Factor CsMADS3 Coordinately Regulates Chlorophyll and Carotenoid Pools in Citrus Hesperidium. Plant Physiol 2023, 193, 519–536. [Google Scholar] [CrossRef] [PubMed]
- Ngamwonglumlert, L.; Devahastin, S.; Chiewchan, N.; Raghavan, V. Plant Carotenoids Evolution during Cultivation, Postharvest Storage, and Food Processing: A Review. Compr Rev Food Sci Food Saf 2020, 19, 1561–1604. [Google Scholar] [CrossRef]
- Wang, T.; Liu, S.; Tian, S.; Ma, T.; Wang, W. Light Regulates Chlorophyll Biosynthesis via ELIP1 during the Storage of Chinese Cabbage. Sci Rep 2022, 12, 11098. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Summary introduction.
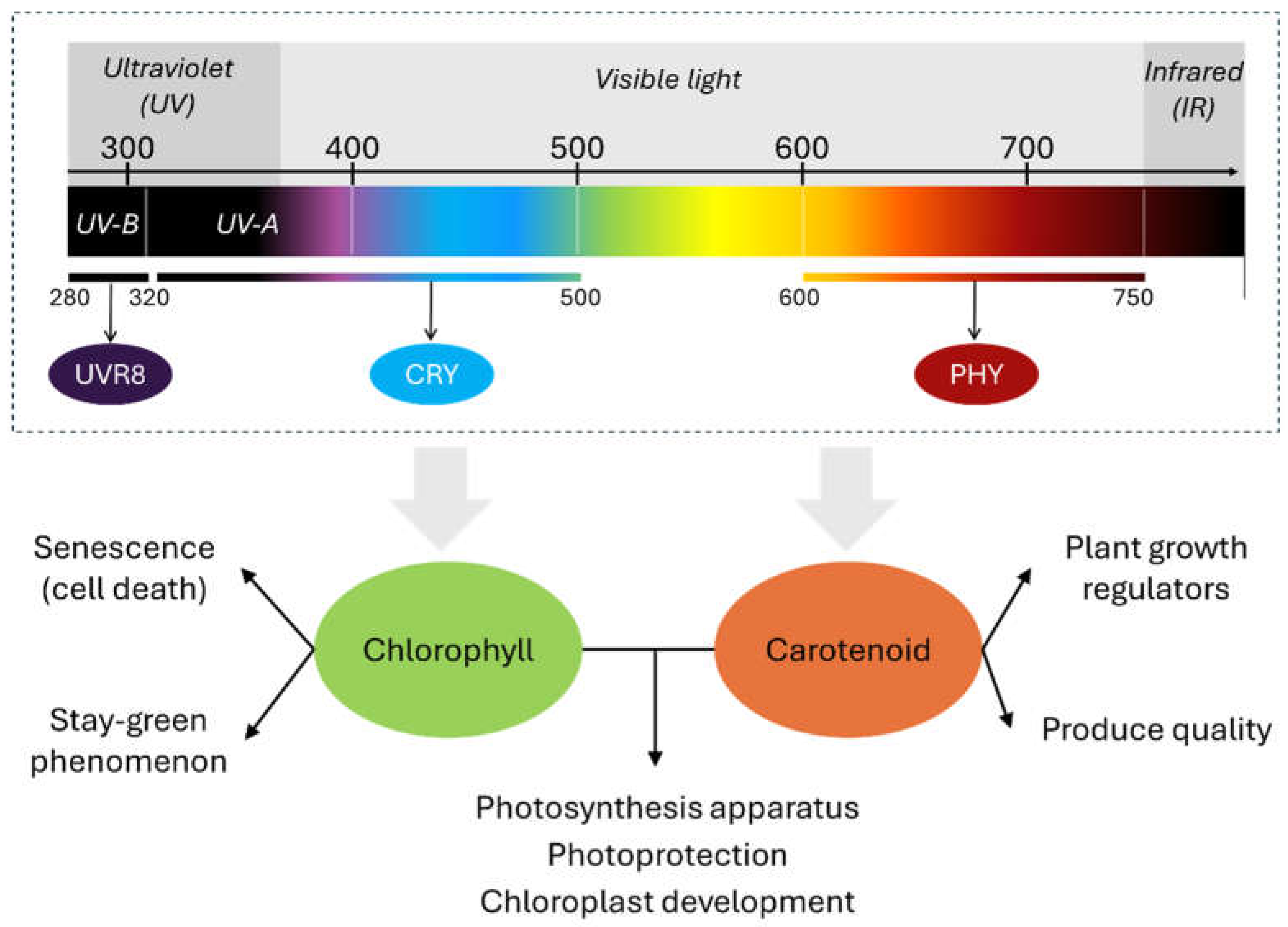
Figure 2.
Regulatory network of chlorophyll biosynthesis and degradation pathways under different light spectra and qualities. Transcription factors involved are shown in colored oval shapes, with the fill color or border color indicating the positive or negative effect of the corresponding light spectrum.
Figure 2.
Regulatory network of chlorophyll biosynthesis and degradation pathways under different light spectra and qualities. Transcription factors involved are shown in colored oval shapes, with the fill color or border color indicating the positive or negative effect of the corresponding light spectrum.
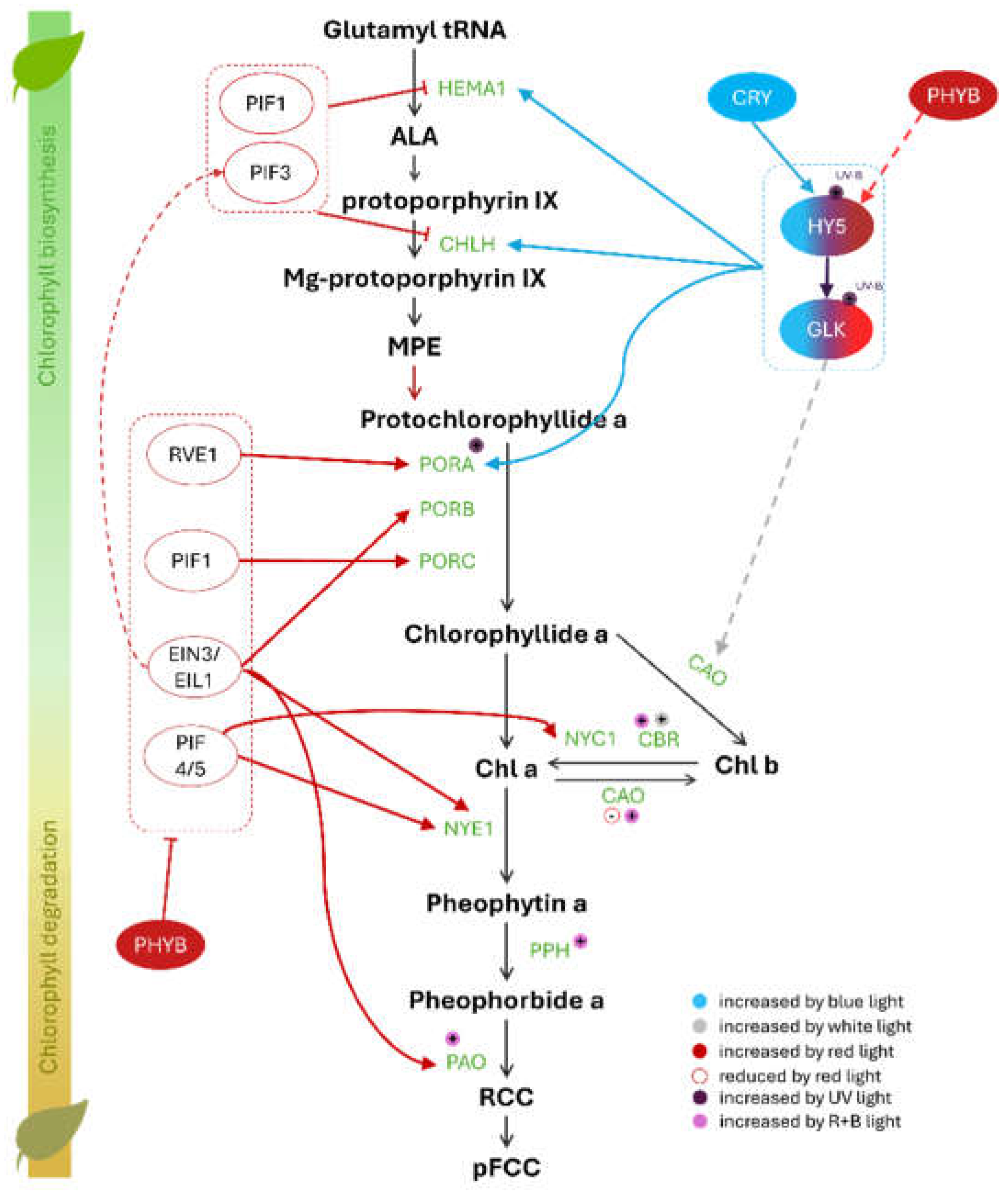
Figure 3.
Regulatory network of carotenoid biosynthesis and degradation pathways under different light spectra and qualities. Transcription factors involved are shown in colored oval shapes, with the fill color or border color indicating the positive or negative effect of the corresponding light spectrum.
Figure 3.
Regulatory network of carotenoid biosynthesis and degradation pathways under different light spectra and qualities. Transcription factors involved are shown in colored oval shapes, with the fill color or border color indicating the positive or negative effect of the corresponding light spectrum.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



