
Submitted:
07 January 2025
Posted:
08 January 2025
You are already at the latest version
Abstract
Pectinases are essential enzymes and command a quarter of all food enzymes sold globally, and are recognised as excellent workhorses in plethora of industries. These enzymes are mainly sourced from animals, microorganisms and plants, with microbes serving as the main sources owing to the quality, quantity, ease of extraction and economic considerations. This research study isolated, screened, and identified five pectinase-producing fungi from the soil of decaying fruits and vegetables. The isolated fungal species were cultured and screened for highest pectinase production using pectinase screening agar medium containing 1% citrus pectin. Four isolates identified as Aspergillus niger, Fusarium species, Trichoderma species and Aspergillus flavus, exhibited very high values of pectinase hydrolysis based on clear zone method, which were 25, 23, 20, and 20 mm for the respective isolates. A secondary pectin hydrolysis screening exercise was conducted afterwards to obtain the highest level of reducing sugars and pectinase released by the isolates. Aspergillus niger, once again, proved to be the best as it recorded maximum reducing sugar of 3.92 mg/mL and pectinase enzyme activity of 36.23 U/mL. Soil have the potential to serve as the primary source of fungal species capable of degrading pectin to release pectinase enzymes, which have numerous applications in the production of pharmaceuticals, food, beverages, animal feed, textiles, detergents, protoplast fusion, pulp and paper and biofuels.
Keywords:
Pectinase
; fungal species
; Aspergillus niger
; pectin
; screening
1. Introduction
Pectin is a hecteropolysaccharide that serves as an important component of the cell wall and other internal components of higher plants. It confers mechanical support and flexibility to plants by acting as a cementing material that holds the cell walls of adjacent cells together [1,2]. Pectin is made up of numerous units that consists of d-galacturonic acids with carboxyl groups, mostly esterified with methanol, with the main chain being made up of α-(1, 4) glycosidic bonds. Pectin with its methyl groups removed is called pectate or pectic acid, which is also called polygalacturonic acid [3]. The structure of pectin contains homogalacturonan, which appears as a linear biopolymer and is made up of methyl-esterified or acetylated d-galacturonic acid residues or monomers containing α-(1, 4) glycosidic linkages. Pectin is formed when about 75% of carboxylic acids found in homogalacturonans become methylated. Pectic acid is also formed when less than 75% of the carboxylic acids are methylated polygalacturonic acid results when there is no methyl esterification [4]. Also, protopectin represents the pectic substances that are not soluble in water [5]. Soluble pectic substances can be formed after limited hydrolysis of protopectin [6]. Pectinic substances, which act as the methylated form of galacturonic acid are formed when there is enzymatic hydrolysis of protopectin in ripe fruits [7].
Pectin is non-toxic, biocompatible and biodegradable and can dissolve in water. It has crucial stabilizing, hydration, gelling, thickening and swelling properties, which makes it very useful in many commercial applications. A few of the commercial applications include food designs, the production of fruits juices, drinks, and fermented, confectionary pharmaceuticals and health products. Pectin is also utilized as a nano-encapsulator, drug delivery agent, constituent of pro-health formulations, dietary fiber, anti-cancer agent, and hydrogel formulation material. It is also used to lower blood cholesterol and low-density lipoprotein levels, as well as being used in the treatment of diarrhea and constipation [8,9,10]. The primary sources of pectin have been citrus fruits, apple pomace, mango peels and sugar beet pulp [11,12,13]. Figure 1 illustrates the structure and powdered form of pectin.
Pectin can be hydrolyzed by a group of enzymes called pectinases by splitting the glycosidic bonds existing between pectin molecules [15]. Pectinases are able to break down the pectin oligo-d-galacturonate present in plant cell walls into monogalacturonic acids. Hydrolysis is achieved when pectinolytic enzymes cleave the α-1, 4 glycosidic linkages of polygalacturonic acid [16]. Pectinases can be classified as extracellular and intracellular. Extracellular pectinase enzymes represent those secreted outside of the cell into the surrounding media in which the microbes live, whereas intracellular pectinase enzymes are those secreted and act within the cell membrane [17]. Nevertheless, extracellular and intracellular pectinases are also classified into three broader groups based on their mode of action on pectic substances [17,18], which include protopectinases, esterases, and depolymerases [19,20,21,22]. Pectin esterases are able to catalyze the de-esterification of methyl groups in pectin to form pectic acids. Pectin depolymerases, depending on their mechanism of action, are able to cleave the glycosidic bonds found in pectic acids. Also, protopectinases are able to break down protopectin [23]. In terms of their nature, pectinases can be categorized as acidic and alkaline. Acidic pectinases usually enjoy their applications in the wine, beverage and fruit juice industries for clarification and purification of fruit juices and wines [13,24]. Alkaline pectinases, on the other hand, find their applications in textile industries and wastewater treatments, [4,25,26].
Among the various pectinases, extracellular pectinases derived from fungal species have been known to possess excellent abilities compared to pectinases from other sources such as plants, animals and bacteria [27], and their method of extraction is very simple as the integrity of the cells remains intact. Other added advantages include high yields, economic feasibility, stability and easy genetic manipulation. Also, fungal-derived pectinolytic enzymes are generally regarded as having a safe (GRAS) status [23]. For these and many more reasons, extracellular pectinases isolated from fungal species especially that of Aspergillus sp. are in high demand in various applications [28,29]. These applications extend across food/feed, beverage, pulp and paper, biofuel, textile, oil extraction and waste and environmental remediation [13,22,30,31]. Pectinases employed in the fruit and wine industries can enhance taste, clarity and overall chromatic appearance compared to untreated ones [25].
The soil, as part of the natural environment, serves as a huge reservoir for fungal species that can be isolated and employed in the production of pectinase enzymes. Microorganisms isolated from the soil are mesophilic, can tolerate stress and have the ability to synthesize pectinase enzymes in large quantities in a wider spectrum of growth parameters [31]. Soils particularly found in plant food processing locations, markets and organic waste dumpsites have been proven to be very much dominant with pectin-degrading fungus [5]. Recent advances indicate that the trend to identify novel enzymes from microorganisms from such environments is accelerating due to their inherent stability and efficiency [32,33]. It is, therefore, hypothesized herein that, collecting soil composed of decaying organic (plant) matter and isolating from it, the fungal strains capable of degrading the pectin-rich components would be very beneficial to obtaining microbial strains tailored for pectinase production, biofuel production and bioremediation purposes. Also, the ability to identify and characterize such microbes isolated in this work would go a long way in providing more information on fungal strains with special and well-tailored capacities for industrial purposes. Hence, pectinolytic microbes were obtained from the soils exposed to high and low temperatures, and subsequent identification was carried out in this current investigation.
2. Materials and Methods
2.1. Collection of Soil Sample
Soil composed of plant-decayed materials was collected from the dumpsites of fruits, vegetables and plant materials in some markets in Kumasi, as these places contained many spoiled fruits and vegetable wastes and decaying plant materials. All the samples were pooled together, kept in sterile plastic containers and carried to the microbiology laboratories of Kwame Nkrumah University of Science and Technology, where fungal species were isolated from them.
2.2. Preparation of Medium for Isolation of Fungal Species
A sterile nutrient medium containing 1% citrus pectin, FeSO4.H2O (5 g/L), MgSO4.7H2O (0.02%), (NH4)2SO4 (0.14%), and K2HPO4 (0.2%) as well as a nutrient solution (0.1%) containing ZnSO4.7H2O (1.4 mg/L), MnSO4.H2O (1.6 mg/L) and CoCl2 (2.0 mg/L) was prepared, and mixed with 3% agar-agar (the gelling agent) and autoclaved at 121 °C for 20 min [21]. It was then allowed to cool to around 45 °C before being placed into Petri dishes to gel. To ensure sterility, the plates were incubated overnight in an incubator at 37 °C.
2.3. Preparation of Potato Dextrose Agar for Isolation of Fungal Species
Potato Dextrose Agar (PDA) was made from a commercial dehydrated OXOID product. Thirty-nine PDA powder (39.0 g) was dissolved in 1 L of distilled water, and sterilized at 121 °C for 20 min and at a pressure of 1.2 g/cm2 according to the product recommendations.
2.4. Preparation of 3, 5-Dinitrosalicylic Acid (DNS)
This protocol followed that of Miller [34]. To summarize, 1.0 g of DNS was dissolved by warming in 20 mL of 2N NaOH. A total of 30.0 g of potassium sodium tartarate was dissolved by warming in 50 mL of distilled water. After cooling, the two solutions were combined together and made up to 100 mL with distilled water.
2.5. Isolation and Sub-Culturing of Fungal Specie
The simple dilution plate method was adopted here. One (1) g of soil sample was diluted ten-fold in 9 mL of sterile distilled water. One (1) mL of the 6th dilution (10−6) was flooded onto three replicates of the previously prepared sterile Potato Dextrose Agar (PDA) nutrient medium, which had been supplemented with chloramphenicol antibiotic to inhibit bacterial growth. The plates were then incubated at room temperature (25 °C–28 °C) for 7 days, until visible fungal colonies appeared. All morphologically distinct colonies were purified by sub-culturing on different plates and streaking with a sterile inoculation loop. This process proceeded until pure fungal isolates were obtained. Pure cultures were maintained on PDA slants, refrigerated and sub-cultured at regular intervals throughout the study and were used as stock cultures. All pure fungal growths were counted with a colony counter, and identified appropriately. Their frequency of occurrence was subsequently established as well.
2.6. Identification of Fungal Isolates
The identification procedure for all the fungal isolates was based on macromorphological and micromorphological characteristics. Macromorphological identification was conducted by observing colony characteristics such as the color, texture, spore structure and the form of mycelia and/or pattern of growth. The micromorphological characteristics like separation, and spore shapes, among others were determined using the usual lactophenol cotton blue procedure (LPCB). All the fungal isolates had been defined in accordance with the manual for determining the identity of fungi [35] and Illustrated Genera of Imperfect Fungi by Barnett and Hunter, [36]. In summary, a three-day-old pure culture was utilized to prepare slides for microscopic examination. A small amount of mycelia was placed on the slide, along with a drop of lactophenol cotton blue reagent. A cover slip was laid over it, and the specimen was examined under a light microscope at ×400 magnification. Identification of the species was accomplished by comparing features to the micrographs, and the photographs taken included at the end of this section.
2.7. Qualitative (Primary) Screening for Pectinolytic Fungal Species
The process of screening for pectinase synthesis was carried out with the use of the prepared cultivation medium (in the previous section) by applying the spot inoculation method. In brief, pure cultures of the various fungal species were introduced into the already prepared sterile and solidified pectin agar medium. The plates that were inoculated were then incubated at a temperature of 30 °C for four days (96 h), and following that, a clear observable colony of fungi could be seen on every Petri dish. Thereafter, the Petri dishes were soaked with iodine-potassium iodide solutions (a mixture of 1.0 g of iodine and 5.0 g of potassium iodide dissolved in 330 mL of distilled water) for about 10 min. This was done in order to boost visibility for measuring the zone of clearance/carbohydrate hydrolysis [21]. A distinct zone that formed surrounding the pectinase-producing colonies was observed and subsequently measured. The fungal isolates with the most promising clear zone of pectinase hydrolysis were chosen and preserved under 4 °C in the refrigerator on pectin agar and later used in the next step.
2.8. Quantitative (Secondary) Screening for Pectinolytic Fungal Species
This experiment utilized the protocol established by Kaur and Kaur [37] with some modifications. The four isolates with higher degrees of pectinolytic activity showing larger zones of pectin hydrolysis obtained in the previous experiment were used in a submerged fermentation process. A total of 50 mL of the fermentation medium (without the 3% agar component) prepared earlier underwent inoculation with 1 mL of the spore suspension at a concentration of 1.2 × 106 spores/mL of each of the culture isolates. It was incubated at 30 °C for 7ays on a rotary shaker rotating at 120 revolutions per minute (rpm). Throughout the experiment, triplicate Erlenmeyer flasks (250 mL) were used for each species of fungus. Once the incubation period elapsed, the crude pectinolytic enzyme underwent extraction. The extraction was achieved by mixing 50 mL of sodium acetate buffer with the fermented broth cultures. The mixture was centrifuged for 15 min at 4000 rpm to remove the debris and cells. The supernatants were labeled as crude enzymes. The pectinolytic activities of the enzyme and reducing sugar concentration were measured by estimating the reducing sugars generated from the pectin using the 3, 5-dinitrosalicylic acid (DNS) technique [34]. Glucose was employed as the benchmark. Each sample’s pectinase activity was defined as the quantity of enzyme required to produce 1 μmol reducing groups per minute.
3. Results and Discussion
3.1. Isolation and Sub-Culturing of Fungal Species
In all, five fungal isolates were obtained from the pooled soil samples collected from various locations. The microbes were designated as FS1, FS2, FS3, FS4 and FS5 based on the Petri dishes they were cultured on. This result is clearly in line with recent developments that have involved screening a significant number of microbial species from diverse soil resources, agrowastes and environments. Such developments are aimed at finding microbial species capable of degrading polysaccharides present in plant biomass that would illicit the synthesis of pectinases [5,38,39,40]. These microbial creatures play crucial roles in the maintenance of nutrient cycles, biodiversity and degradation of organic matter. Microbes have also been shown to be very useful for industrial processes. The fact that, all five isolates are fungal species perfectly agrees with Meyer et al., [41]. According to them, fungal species are the most dominant microbial consortia present in the environment. The dominance is a result of their robustness in their lifecycle and activities [31].
The wide range of fungal species in the natural environment, particularly in plant food processing locations, markets and organic waste dumpsites, is also in strong alignment with Thakur et al., [5]. There is no doubt that soil serves as a reservoir for plant-degrading microorganisms that can be employed to carry out many processes. Previous investigations established by Ezenwelu et al., [42] demonstrated the existence of Aspergillus niger as well as Rhizopus sp. within the environments composed of decaying plant materials. Ametefe et al., [43] recorded Aspergillus niger and Penicillium sp. as part of the fungi they isolated from the soil. Aside the soil, decaying fruits have also been implicated in harboring many microbial consortia, especially fungal species for instance. Okonji et al., [31] isolated Aspergillus fumigatus from decaying citrus. Mat Jalil et al., [44] also isolated Aspergillus niger (LFP-1 strain) from decomposed citrus fruits, and Budu et al., [40], showed the abundance of fungal spp. in decaying cassava peels. Plant-decaying soils (especially that of fruits and vegetables) are attractive to the growth and survival of fungi. This is because, they serve as a rich source of nutrients, and the soil creates a conducive environment that is highly favorable to fungal growth and survival [41].
As a result, such soils have the potential to draw fungi to colonize and flourish in them. Secondly, the physical structure of fruits and vegetables, which is usually characterized by their fibrous and porous nature, provides a generous surface area and soil environment for fungal spores to land, establish their growth and thrive successfully. The physical structure, in addition to texture, pH, oxygen content, cation exchange capacity and redox reactions of the soil, provide synergistic conditions for microbe wellbeing. As such, their presence in organic matter in the soil, as a natural consequence, underscores the roles fungi play as the agents involved in breaking down and recycling organic components and nutrients.
3.2. Identification of Fungal Isolates
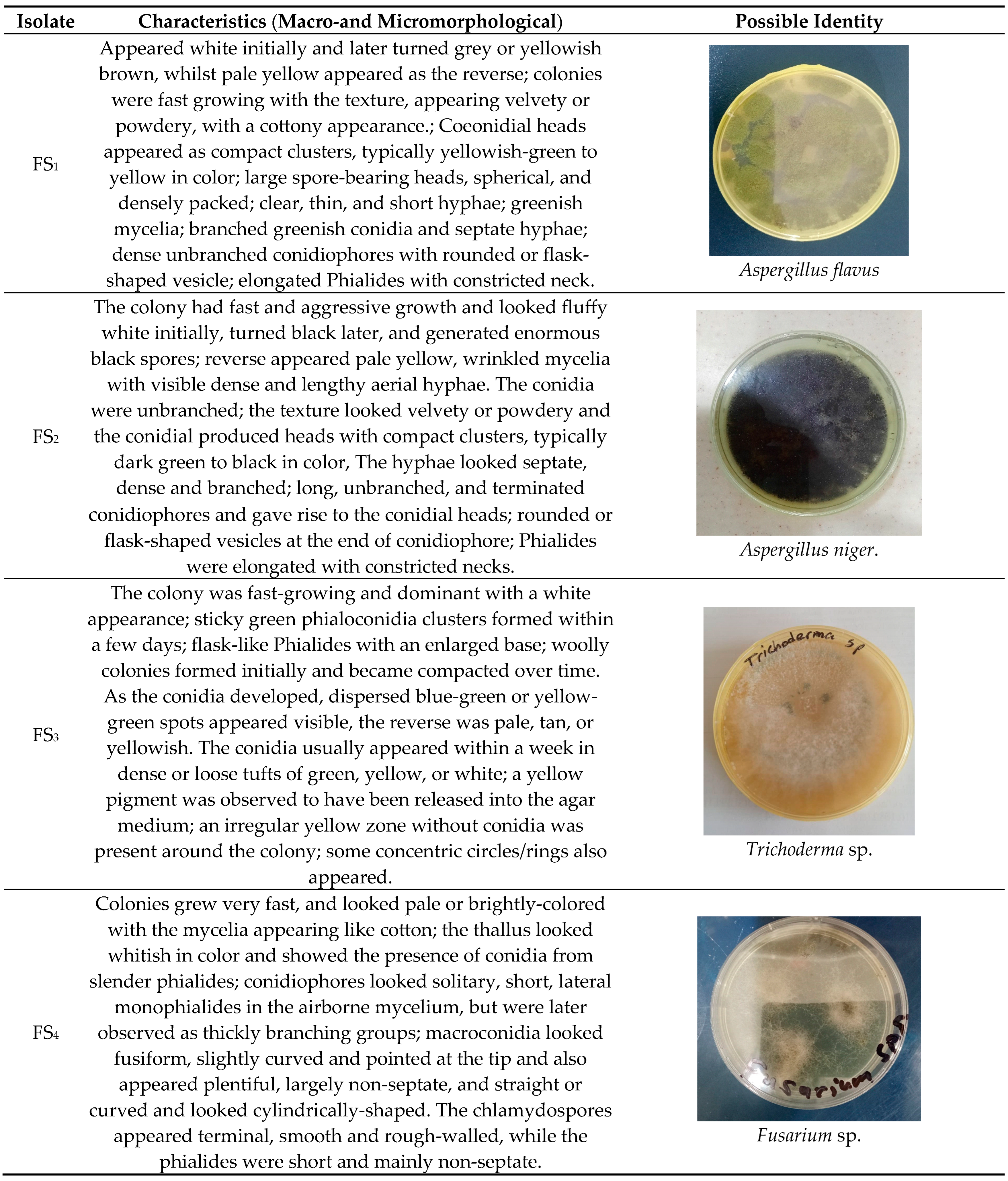
Fungal isolates FS1, FS2, FS3, FS4 and FS5 were thoroughly identified in accordance with their macromorphological and micromorphological features [34,35]. At the end of this section, Aspergillus flavus, Fusarium sp., Aspergillus niger, Trichoderma sp. and Saccharomyces sp. were realized. Table 1 demonstrates a summary of the various fungal species successfully obtained and identified as well as their characteristics.
3.3. Qualitative (Primary) Screening for Pectinolytic Fungal Species
One of the numerous ways to show pectinase’s production ability of microorganisms, especially fungal species, is the method popularly known as pectin/carbohydrate hydrolysis or zone of clearance. Such procedures have been known to be very effective in determining and selecting the best fungal species needed to carry out pectinase production [20,31,38]. By adopting similar paths, this current study conducted a protocol showing carbohydrate hydrolysis. The aim was to select the isolates with the highest pectin degradation potential, after 4 days of incubation. After subjecting the isolates to pectin hydrolysis, the result showed that, all the fungal species possessed various degrees of pectinolytic activity (Table 2 and Figure 2). Aspergillus niger (FS2) emerged slightly on top as the most efficient microbial species (25.0 mm) among the tested species. Fusarium species (FS4) followed second with 23.0 mm, then Trichoderma species (FS3) and Aspergillus flavus (FS1) produced 20.0 mm each. Saccharomyces species (FS5) gave 7. 0 mm, hence, it was left out in subsequent experiments.
It is well noted that, the strains of Aspergillus spp. have been known to dominate pectinase enzyme manufacturing in various studies and industrial applications [31]. The fact that, fungal species dominated the synthesis of pectinase in terms of the zone of clearance in the current study is favorably supported by El-Ghomary et al., [39]. In their work, they opined that, the majority of commercial pectinase enzymes employed in industries are generated from species of fungal sources, including Rhizopus stolonifer, Aspergillus and Penicillium sp. Oumer and Abate, [20] and Adedayo et al., [38], also asserted that, commercial pectinase enzyme production has recognized Aspergillus and Penicillium sp. as the main sources of pectinase enzymes. El-Ghomary et al., [39] isolated four strains of Aspergillus spp., which showed clear zones of pectin hydrolysis of 3.62 mm, 3.70 mm, 3.91 mm, and 4.75 mm for the respective strains, with Aspergillus flavus emerging as the species with the highest zone of clearance. The current result is also, in agreement with Okonji et al., [31] when a fungal isolate identified as Aspergillus fumigatus showed the highest pectin hydrolysis. Other numerous earlier investigations have also indicated the vital role played by Aspergillus sp. [13,20,29,44,45].
Furthermore, the relative dominance of Aspergillus niger, in the current investigation, after exhibiting the highest activity of pectinase in relation to the zone of clearance (25 mm), is no fluke at all. This assertion is true as its ability to hydrolyze pectin-rich media has also been agreeably demonstrated in a series of previous studies cited in Satapathy et al., [21]. In another closely related study conducted by Adedayo et al., [38], Aspergillus niger showed a better zone of clearance of 7.0 mm, whereas Aspergillus flavus produced 5.5 mm. In addition, Kamalambigeswari et al., [46], examined three distinct strains of Aspergillus niger (F-3, F-4, and F-P), which revealed isolate F-4 as one that exhibited the largest region of pectin breakdown (9 mm).
3.4. Quantitative Screening of Potent Pectinolytic Fungal Species (Secondary Screening)
Due to their promising nature, all four isolates with comparatively higher zones of pectin hydrolysis were selected for further work on pectinase production (secondary screening). The four isolates that were deemed the best at their pectinolytic ability exhibiting a clear zone of clearance were scrutinized for quantitative evaluation of pectinase synthesis utilizing a selective enrichment technique to generate comparative assessments. The secondary pectin hydrolysis screening exercise was then performed to identify the greatest amount of reducing sugars generated by the isolates with their respective enzyme activity via the enzymatic breakdown of pectin. The galacturonic acid released was measured spectrophotometrically. The result (Table 3) showed that among the four (4) fungal isolates, FS2, which was identified as Aspergillus niger, once again, emerged as the most potent fungal species for the production of pectinase after recording the highest values for reducing sugar formation (3.92 mg/mL) and pectinase activity (36.23 U/mL) under submerged fermentation after seven (7) days of incubation.
This finding fits in with El-Ghomary et al., [39], who found that, fungal isolates tagged as F9 showed a relatively highest titre of pectinase (35.83 U/mL) than that of all the other isolates. Okonji et al., [31] also discovered that Aspergillus fumigatus yielded 42 U/mL of pectinase enzyme activity in submerged fermentation conditions. The results achieved in this study and subsequent selection of fungal strains are also in line with Sudeep et al., [29], who, in their research, isolated four strains of fungal species with high pectinolytic activities from the soil. After thorough primary and secondary screening procedures, Aspergillus sp. was observed as the most potent fungus for pectinase enzyme production.
4. Conclusions
Soils obtained from the selected markets in Kumasi were found to be a great source of a plethora of diverse pectin-degrading microorganisms, especially fungal species. A. niger has proven to be very dominant, probably as a result of its ubiquitous nature and its ability to colonize diverse environments and substrates as it thrives well in many growth media or substrates, as proven by this and other studies. A successful collection of these industrially-relevant strains of microorganisms from the soil could very well contribute to the increasing demand and adequate supply of pectinase enzymes in commercial quantities by related industries. Pectinase enzymes from microbes would also be very useful in the remediation of areas polluted with plant waste. Further researches should be directed towards the mechanisms involved in the molecular regulation of pectinase enzymes production among fungal species, especially A. niger. With such knowledge, scientists would be able to optimize the production of pectinase for commercial purposes. Also, subsequent studies could be invested in screening of extreme environments with the aim of finding fungal species with enhanced capabilities that can synthesize pectinase with novel properties.
Author Contributions
The conceptualisation and design of this study were done by all authors. Material preparation, data collection and analysis, as well as the first draft of the manuscript, were performed by S.O.B., while H.D.Z. reviewed edited and provided valuable comments and guidance, M.B. supervised the laboratory experiments. A.B.D. reviewed and provided valuable comments. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
All authors have given their approval for the article to be published.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shilpa, M.K.; Jason, Y. Isolation and screening of pectinase-producing bacteria from soil sample. CGC Int. J. Contemp. Res. Eng. Technol. 2021, 3, 166–170. [Google Scholar]
- Belkheiri, A.; Forouhar, A.; Ursu, A.V.; Dubessay, P.; Pierre, G.; Delattre, C.; Djelveh, G.; Abdelkafi, S.; Hamdami, N.; Michaud, P. Extraction, Characterization, and Applications of Pectins from Plant By-Products. Appl. Sci. 2021, 11, 6596. [Google Scholar] [CrossRef]
- Rokade, D.D.; Vaidya, S.L.; Naziya, M.A.; Dixit, P.P. Screening of Pectinase Producing Bacteria, Isolated from Osmanabad fruit market soil. Int. J. Interdiscip. Multidiscip. 2015, 2, 141–145. [Google Scholar]
- Tabssum, F.; Ali, S.S. Screening of pectinase producing Gram-positive bacteria: Isolation and characterization. Punjab Univ. J. Zool. 2018, 33, 11–15. [Google Scholar] [CrossRef]
- Thakur, P.; Kumar, A.S.; Mukherjee, G. Isolation and Characterization of Alkaline Pectinase Productive Bacillus tropicus from Fruit and Vegetable Waste Dump Soil. Braz. Arch. Biol. Technol. 2021, 64, e21200319. [Google Scholar] [CrossRef]
- Rakitikul, W.; Tammasat, T.; Udjai, J.; Nimmanpipug, P. Experimental and DFT study of gelling factor of pectin. Suranaree J. Sci. Technol. 2016, 23, 421–428. [Google Scholar]
- Carlos Álvarez García, Chapter 11 - Application of Enzymes for Fruit Juice Processing, Editor(s): Gaurav Rajauria, Brijesh K. 2018. [CrossRef]
- Freitas, C.M.P.; Coimbra, J.S.R.; Souza, V.G.L.; Sousa, R.C.S. Structure and Applications of Pectin in Food, Biomedical, and Pharmaceutical Industry: A Review. Coatings 2021, 11, 922. [Google Scholar] [CrossRef]
- Lucarini, M.; Durazzo, A. Bernini, R.; Campo, M.; Vita, C.; Souto, E.B.; Lombardi-Boccia, G, Ramadan, M.F.; Santini, A.; Romani, A. Fruit Wastes as a Valuable Source of Value-Added Compounds: A Collaborative Perspective. Molecules 2021, 26, 6338. [Google Scholar] [CrossRef] [PubMed]
- Vinay, C.; Biswas, D.; Roy, S.; Vaidya, D.; Verma, A.; Gupta, A. Current Advancements in Pectin: Extraction, Properties and Multifunctional Applications. Foods 2022, 11, 2683. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Hua, X.; Liu, J.R.; Wang, M.M.; Liu, Y.X.; Yang, R.J.; Cao, Y.P. Extraction of sunflower head pectin with superfine grinding pretreatment. Food Chem. 2020, 320. [Google Scholar] [CrossRef] [PubMed]
- Picot-Allain, M.C.N.; Ramasawmy, B.; Emmambux, M.N. Extraction, characterisation, and application of pectin from tropical and sub-tropical fruits: A review. Food Rev. Int. 2022, 38, 282–312. [Google Scholar] [CrossRef]
- Roman-Benn, A.; Contador, C.A.; Li, M.-W.; Lam, H.-M.; Ah-Hen, K.; Ulloa, P.E.; Ravanal, M.C. Pectin: An overview of sources, extraction and applications in food products, biomedical, pharmaceutical and environmental issues. Food Chem. Adv. 2023, 2, 100192. [Google Scholar] [CrossRef]
- Suckhoequyhonvang.vn (2022). Health benefits of pectin. https://www.suckhoequyhonvang.vn/tin-tuc/nhung-tuc-dung-cua-pectin-doi-voi-suc-khoe-92.html. (Accessed on 5 June 2023).
- Hassan, S. Development of Novel Pectinase and Xylanase Juice Clarifcation Enzymes via a Combined Biorefnery and Immobilization Approach. Doctoral Thesis, Technological University Dublin, Dublin, Ireland, 2020. [Google Scholar] [CrossRef]
- Kauldhar, B.S.; Kaur, H.; Meda, V.; Sooch, B.S. Chapter 12—Insights into Upstreaming and Downstreaming Processes of Microbial Extremozymes. Extremozymes and Their Industrial Applications; Academic Press: Cambridge, MA, USA, 2022; pp. 321–352. ISBN 9780323902748. [Google Scholar] [CrossRef]
- Shet, A.R.; Desai, S.V.; Achappa, S. Pectinolytic enzymes: Classification, production, purification and applications. Res. J. Life Sci. Bioinform. Pharm Chem. Sci. 2018, 4, 337–348. [Google Scholar]
- Basheer, S.M.; Chellappan, S.; Sabu, A. Chapter 8—Enzymes in Fruit and Vegetable Processing; Kuddus, M., Aguilar, C.N., Eds.; Value-Addition in Food Products and Processing through Enzyme Technology; Academic Press: Cambridge, MA, USA, 2022; pp. 101–110. ISBN 9780323899291. [Google Scholar]
- Garg, G.; Singh, A.; Kaur, A.; Singh, R.; Kaur, J.; Mahajan, R. Microbial pectinases: An ecofriendly tool of nature for industries. 3 Biotech 2016, 6, 47. [Google Scholar] [CrossRef]
- Oumer, O.J.; Abate, D. Screening and molecular identification of pectinase producing microbes from coffee pulp. BioMed Res. Int. 2018, 2961767. [Google Scholar] [CrossRef] [PubMed]
- Satapathy, S.; Behera, P.M.; Tanty, D.K.; Srivastava, S.; Thatoi, H.; Dixit, A.; Sahoo, S.L. Isolation and molecular identification of pectinase producing Aspergillus species from different soil samples of Bhubaneswar regions. BioRxiv 2019, 2019 1, 1–22. [Google Scholar]
- Shrestha, S.; Chio, C.; Khatiwada, J.R.; Kognou, A.L.M.; Chen, X.; Qin, W. Optimization of Cultural Conditions for Pectinase Production by Streptomyces sp. and Characterization of Partially Purified Enzymes. Microb. Physiol. 2023, 33, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kokare, C. Chapter 17—Microbial Enzymes of Use in Industry; Brahmachari, G., Ed.; Biotechnology of Microbial Enzymes (Second Edition); Academic Press: Cambridge, MA, USA, 2023; pp. 405–444. ISBN 9780443190599. [Google Scholar]
- Osete-Alcaraz, A.; Gómez-Plaza, E.; Pérez-Porras, P.; Bautista-Ortín, A.B. Revisiting the use of pectinases in enology: A role beyond facilitating phenolic grape extraction. Food Chem. 2022, 372, 131282. [Google Scholar] [CrossRef]
- Haile, S.; Ayele, A. Pectinase from Microorganisms and Its Industrial Applications. Sci. World J. Vol. 2022, 2022, 1881305. [Google Scholar] [CrossRef]
- Inamdar, N.; Dike, A.; Jadhav, A. Isolation, Screening, and Identification of Pectin Degrading Bacteria from Soil. J. Chem. Health Risks JCHR 2024, 14, 930–938. [Google Scholar]
- Ivarsson, M.; Drake, H.; Bengtson, S.; Rasmussen, B. A Cryptic Alternative for the Evolution of Hyphae. BioEssays 2020, 42, 1900183. [Google Scholar] [CrossRef] [PubMed]
- José Carlos, D.L.-M.; Leonardo, S.; Jesús, M.-C.; Paola, M.-R.; Alejandro, Z.-C.; Juan, A.-V.; Cristóbal Noé, A. Solid-State Fermentation with Aspergillus niger GH1 to enhance Polyphenolic content and antioxidative activity of Castilla Rose Purshia plicata. Plants 2020, 9, 1518. [Google Scholar] [CrossRef] [PubMed]
- Sudeep, K.C.; Jitendra, U.; Dev, R.J.; Binod, L.; Dhiraj, K.C.; Bhoj, R.P.; Tirtha, R.B.; Rajiv, D.; Santosh, K.; Niranjan, K.; Vijaya, R. Production, Characterization, and Industrial Application of Pectinase Enzyme Isolated from Fungal Strains. Fermentation 2020, 6, 59. [Google Scholar] [CrossRef]
- Singh, A.; Varghese, L.M.; Yadav, R.D.; et al. , A pollution reducing enzymatic deinking approach for recycling of mixed office waste paper. Environ. Sci. Pollut. Res. 2020, 27, 45814–45823. [Google Scholar] [CrossRef]
- Okonji, R.E.; Itakorode, B.O.; Ovumedia, J.O.; Adedeji, O.S. Purification and biochemical characterization of pectinase produced by Aspergillus fumigatus isolated from soil of decomposing plant materials. J. Appl. Biol. Biotechnol. 2019, 7, 1–8. [Google Scholar]
- Ghosh, M.; Pulicherla, K.K. Psychrophiles as the source for potential industrial psychrozymes. Recent Developments in Microbial Technologies. Recent Dev. Microb. Technol. 2021, 355–366. [Google Scholar] [CrossRef]
- Prasad, R.; Kumar, V.; Singh, J.; Upadhyaya, C.P. Recent Developments in Microbial Technologies. Environmental and Microbial Biotechnology. 2021, 2662–169X. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Intro. Mycology; Wiley: New York, NY, USA, 1996; Volume 4. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 3rd ed.; Burgess Publishing Co.: Minneapolis, MN, USA, 1972; p. 241. [Google Scholar]
- Kaur, H.P.; Kaur, G. Optimization of cultural conditions for pectinase produced by fruit spoilage fungi. Int. J. Adv. Res. 2014, 3, 851–859. [Google Scholar]
- Adedayo, M.R.; Mohammed, M.T.; Ajiboye AE and abdulmumini, S.A. Pectinolytic activity of Aspergillus niger and Aspergillus flavus grown on citrus parasidis peel in solid state fermentation. Glob. J. Pure Appl. Sci. 2021, 27, 93–105. [Google Scholar] [CrossRef]
- El-Ghomary, A.E.; Shoukry, A.A.; EL-Kotkat, M.B. Productivity of pectinase enzymes by Aspergillus sp. isolated from Egyptian soil. Al-Azhar J. Agric. Res. 2021, 46, 79–87. [Google Scholar] [CrossRef]
- Budu, M.; Boakye, P.; Bentil, J.A. Microbial diversity, enzyme profile and substrate concentration for bioconversion of cassava peels to bioethanol. Preprints.org, 2023, 1, 1-13. [CrossRef]
- Timothy, C. Cairns, Corrado Nai and Vera Meyer Aspergillus niger: A Hundred Years of Contribution to the Natural Products Chemistry. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar]
- Ezenwelu, C.O.; Afeez, O.A.; Anthony, O.U.; Promise, O.A.; Mmesoma, U.-E.C.; Henry, O.E. Studies on Properties of Lipase Produced from Aspergillus sp. Isolated from Compost Soil. Adv. Enzym. Res. 2022, 10, 49–60. [Google Scholar] [CrossRef]
- Ametefe, G.D.; Oluwadamilare, L.A.; Ifeoma, C.J.; Olubunmi, I.I.; Ofoegbu, V.O.; Folake, F.; Orji, F.A.; Iweala, E.E.J.; Chinedu, S.N. Optimization of pectinase activity from locally isolated fungi and agrowastes. Res. Sq., 2021, 1, 1–21. [Google Scholar] [CrossRef]
- Jalil, M.T.M.; Ibrahim, D. Partial purification and characterisation of pectinase produced by Aspergillus niger LFP-1 grown on pomelo peels as a substrate. Trop. Life Sci. Res. 2021, 32, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Oumer, O.J. Pectinase: Substrate, production and their biotechnological applications. Int. J. Environ. Agric. Biotechnol. 2017, 2, 238761. [Google Scholar] [CrossRef]
- Kamalambigeswari, R.; Sangilimuthu, A.Y.; Narender, S.; Ushani, U. Isolation, identification, screening and optimization of pectinase producing soil fungi (Aspergillus niger). Int. J. Res. Pharm. Sci. 2018, 9, 762–768. [Google Scholar]
Figure 1.
The structure and powdered form of pectin [14].
Figure 1.
The structure and powdered form of pectin [14].
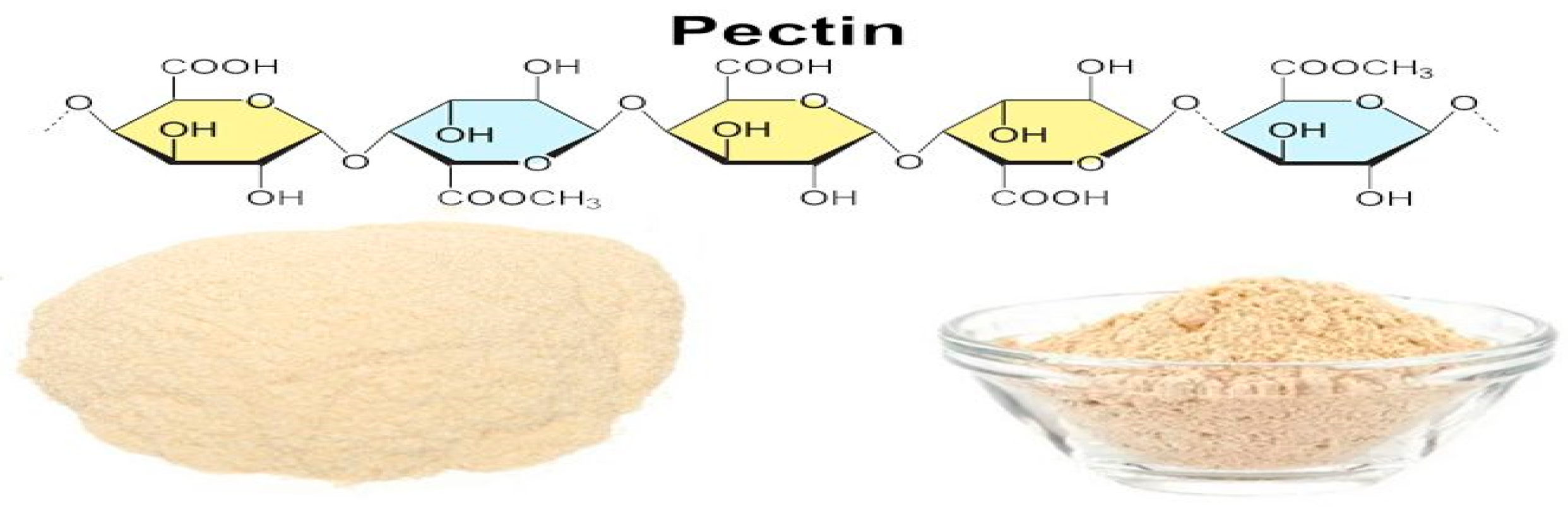
Figure 2.
Qualitative analysis of fungal species based on the zone of pectin hydrolysis: (A) Fusarium sp.; (B) Aspergillus niger; (C) Aspergillus flavus; (D) Trichoderma sp.; and (E) Saccharomyces sp.
Figure 2.
Qualitative analysis of fungal species based on the zone of pectin hydrolysis: (A) Fusarium sp.; (B) Aspergillus niger; (C) Aspergillus flavus; (D) Trichoderma sp.; and (E) Saccharomyces sp.
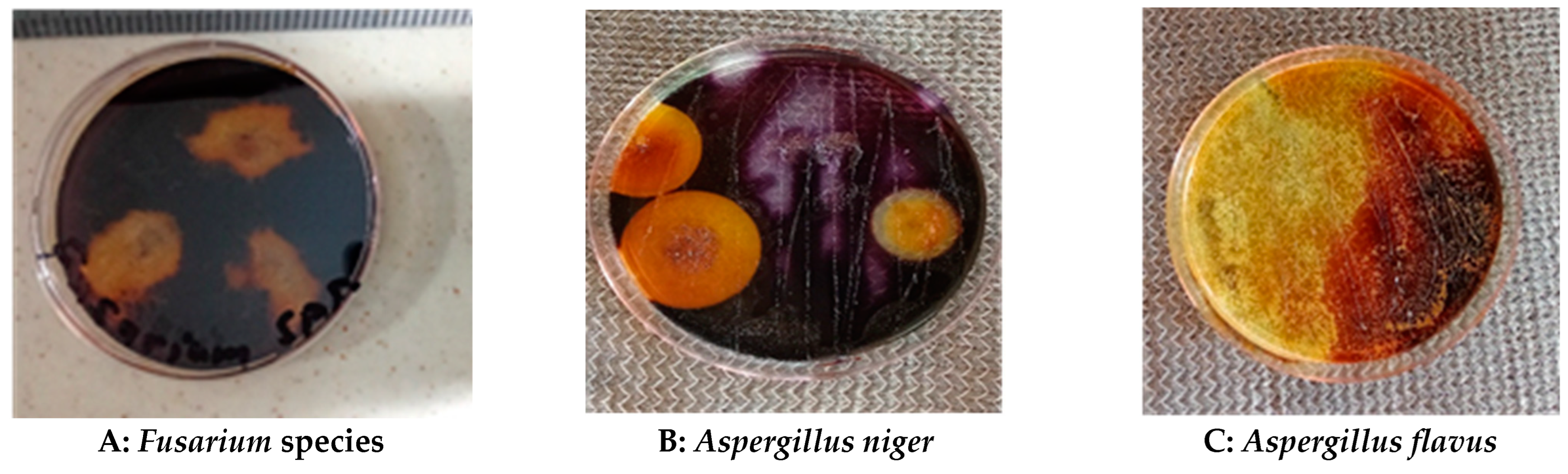


Table 1.
Characteristics of the fungal isolates.
|
Table 2.
Qualitative analysis of fungal species based on the zone of pectin hydrolysis achieved.
| Microorganism | Zone of Pectinolytic Activity/mm |
|---|---|
| Aspergillus niger | 25.00 |
| Fusarium sp. | 23.00 |
| Trichoderma sp. | 20.00 |
| Aspergillus flavus | 20.00 |
| Saccharomyces sp. | 7.00 |
p-value = 0.0511.
Table 3.
The figures obtained during the secondary screening.
| Microorganism | Reducing Sugar (mg/mL) | Enzyme Activity (U/mL) |
|---|---|---|
| Aspergillus niger (FS2) | 3.92 | 36.23 |
| Aspergillus flavus (FS1) | 3.68 | 32.52 |
| Trichoderma sp. (FS3) | 3.23 | 29.74 |
| Fusarium sp. (FS4) | 3.16 | 28.66 |
p-value = 0.0260.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



