Submitted:
28 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
The honeydew moth, Cryptoblabes gnidiella Millière (Lepidoptera: Pyralidae), is native to the Mediterranean Basin. However, it has recently been reported as an emerging grapevine pest in southern European Union countries and in the Middle East, North Africa and South America. This may be attributed to the global warming trends. In this brief communication, we present the first documented cases of honeydew moth infestation of grapevine in Greece (Northeastern Greece 2024). We discuss the potential implications and concerns regarding the occurrence and management of this pest in Greece. Integrated pest management practices may contribute to reducing reliance on chemical pesticides and developing more sustainable approaches.
Keywords:
honeydew moth
; Mediterranean Basin
; pest management strategies
; Greece
; wine-producing areas
1. Introduction
The subfamily Phycitinae (Pyraloidea, Pyralidae) comprises significant pests, with their larvae exhibiting considerable variation in their feeding habits. For instance, some of the most well-known species of this subfamily infest stored products such as Ephestia elutella (Hübn.), Anagasta kuehniella (Zell.), Cadra cautella (Wlk.) and Plodia interpunctella (Hübn.) [1,2]. Other equally notorious pests include the locust bean moth Apomyelois ceratoniae (Zeller) feeding on carob and pomegranate [3], the pulse pod borer moth Etiella zinckenella (Treitschke), that is a pest of soybean [4] and the quince moth Euzophera bigella (Zeller), that infests olives [5]. In total, Powell [6] suggests that approximately two thirds of the 6,000 described pyralid moth species (Pyralidae) are classified within the Phycitinae subfamily.
To date, the most important grapevine pests reported in Greece include two species of Lepidoptera, Lobesia botrana (D. & Schiff.) (Tortricidae, Olethreutinae) [7,8] and Eupoecilia ambiguella (Hübn.) (Tortricidae, Tortricinae), several hemipteran species, Planococcus ficus (Signoret) (Pseudococcidae) [9,10]; Arboridia adanae (Dlabola); Asymmetrasca decedens (Paoli); Hebata decipiens (Paoli); Hebata vitis (Göthe); Jacobiasca lybica (Bergevin) and Zygina rhamni Ferrari (Ciccadellidae, Typhlocybinae) [11], and one thrips species, namely Frankliniella occidentalis (Pergande) (Thripidae) [12,13].
Among members of the Phycitinae subfamily, the honeydew moth Cryptoblabes gnidiella Millière is the most prevalent and economically damaging species pest of grapevine in the coastal regions of Italy, France, and Israel [14,15,16,17]. In European vineyards, C. gnidiella was never considered a primary pest of the grapevine, likely due to its typically low population density and the fact that its presence in grape clusters, and thus the associated damage, was primarily attributed to other pests such as L. botrana [15,18].
The honeydew moth is native to the Mediterranean Basin, southern Europe, North Africa and Southwest Asia. In Europe, the species is predominantly found in Portugal, Spain, Italy, Greece, Ukraine and Austria [19,20,21]. The distribution of C. gnidiella in Europe is outlined by Karsholt and Razowski [22], who document its natural distribution in Norway, Denmark, Sweden, Finland, the Netherlands, the United Kingdom, France (including Corsica), Spain, Portugal, Italy (including Sardinia and Sicily), Malta, Austria and Greece. On the contrary, records from the United Kingdom, the Netherlands, Scandinavian countries and Poland are likely the result of interceptions and the accidental introduction of the species with imported fruits [15,19,21]. Finally, C. gnidiella has also been reported in Malaysia, New Zealand, Hawaii, several countries in Africa and Asia, as well as numerous tropical and subtropical regions of North and South America [15,21].
The honeydew moth develops on a variety of host plants, including citrus, grapes, plum, peach, apple, pear, medlar and loquat, fig, pomegranate, kiwi, mango, avocado, fejoia, blueberry, cotton, maize, carrot, persimmon, etc. [14,15,23,24,25]. Nevertheless, its larvae show a profound preference for grapes and citrus fruits [18]; in fact, larvae that feed on grapes develop faster than the ones that feed on citrus. Over its natural range, honeydew moth is multivoltine, with six to seven generations in Israel [18,26], three to four generations in Italy and France [14], four generations in Antalya, Turkey [27] and three generations in Uruguay [28]. While the first generation is hardly causing any damage as grapes are still unripe, the second generation causes the majority of damage since it coincides with the period of grape ripening [28]. Consequently, the impact of C. gnidiella is far more intense late-ripening than early-ripening cultivars [17]. To that, the notable rise in captures observed from the onset of grape ripening in Tuscan (Italy) is related to the outset of the second generation as well as to the movement of moths from adjacent vineyards driven by volatile compounds present in ripe and rotten grapes. The last two generations mostly overlap leading to an increase in population density [14].
It has been demonstrated that the larvae of honeydew moth are attracted to the juice of grape bunches previously affected by the grape berry moth, L. botrana [18]. To the same direction, Bagnoli and Lucchi [14] suggest that the irregular infestation in a vineyard is quite likely associated with factors that determine the availability of food resources, such as L. botrana infestation, P. ficus outbreaks (due to the presence of sugars and honeydew that young larvae consume) and disease-related deterioration of ripening grapes. However, tt should also be noted that C. gnidiella larvae have the potential to cause damage to clusters that have not previously been affected by other pests, even prior to veraison [24]. This is attributed to superficial erosions on the rachis and peduncles of grapes that disrupt their vascular system [15]. The honeydew moth overwinters in the vineyard as non-diapausing larvae (1st-5th instar) concealed within the dried grape cluster remnants on the plant or on the ground [15,28]; yet, in late spring-early/mid-summer time (April to July), adults migrate from the vineyard to alternative woody and herbaceous hosts for oviposition and offspring development [15]. This might well explain the inconsistence between the scarcity of the larval population from May to July against the high abundance of adults captured in July [15].
In the wine-producing area of the Regional Unit of Drama (Eastern Macedonia, Northeastern Greece), late infestation of grapes by lepidopteran larvae has frequently been attributed to L. botrana. In light of recent publications on the spread of C. gnidiella in Mediterranean viticultural areas, accompanied by reports of damage, a preliminary investigation has been conducted to identify the potential occurrence of C. gnidiella in wine grapevines in Drama. This brief communication presents the first observation of grape infestation by the honeydew moth in Greece.
2. Materials and Methods
2.1. Population Density Monitoring and Sampling Locations
One pheromone delta trap equipped with sticky plastic delta liners (Suterra®) was installed on the 20th of August 2024 in an organic vineyard located in the Kali Vrysi rural area (Drama, Greece) (41° 9.80’ N, 23° 53.03’ S) for the purpose of monitoring C. gnidiellaon late-harvesting varieties. Adults’ captures were recorded on a weekly basis. The mean, minimum and maximum air temperature was recorded on a daily basis from the meteorological station owned by the National Observatory of Athens (www.noa.gr) that is located in Mikrokampos (Drama, Greece) (41° 7.78’ N, 24° 0.73’ E) and situated 8 km away from the sampling point. Historical temperature data for the period between 2020 and 2023 have also been obtained from the same source. In the temperature data obtained, the average temperature for the period May-September 2024 was compared with the corresponding average temperature for the years 2020-2023 considered for the same period (May-September). For this purpose, the t-test was applied using the statistical programme Jasp 0.19.1 [29]. Additionally, specimens of infested grape bunches from the aforementioned plot and other adjacent vineyards were collected and transferred to the laboratory of the Department of Quality and Phytosanitary Inspections (Rural Economy and Veterinary Directorate, Drama, Greece) for further examination.
2.2. Species Identification
The process of species identification was conducted using both morphological traits and molecular tools. Morphological identification was based on adults collected from both sticky traps and infested grape bunches usingspecies-specific taxonomic identification keys [15,23]. DNA barcoding was performed with larvae collected from infested bunches which were immediately stored in 95% alcohol. In total, DNA was extracted from the body of three (3) larvae using PureLine® Genomic DNA kit (Invitrogen, Waltham, MA, USA), following the manufacturer’s protocol. Polymerase chain reaction was run in 25μl volume with LCO-HCO primers [30] that amplify a 658bp long locus of the mitochondrial cytochrome oxidase subunit I (COI) gene. Concentrations and conditions of thermocycling are provided in Avtzis et al. [31]. PCR products were cleaned up enzymatically using the ExoSAP-IT TM PCR Product Cleanup Reagent (ThermoFisher Scientific), and then shipped to Cemia Company (Larissa, Greece) where they sequenced in an ABI3730XL automated sequencer using the same primers as in PCR. Sequences were initially visualized with Chromas Lite version 2.6.6 software and then blasted in the NCBI GenBank database.
2.3. Assessment of the Damage Extent
The level of damage was assessed immediately after the harvest, in early September, and particularly on the late ripening vines, especially the ‘Xinomavro’ cultivar, in early October. At least five hundred bunches per field were visually examined for symptoms of feeding damage, such as holes or hollow grapes, and evidence of the presence of larvae, as well as the extent of deterioration of the grapes. Bunches deemed to be adversely affected by these factors were then segregated, and the estimated percentage of the total affected was calculated.
3. Results
3.1. Species Identification
Based on the comprehensive morphological keys of Neunzig [23] and Lucchi et al. [15], adults were readily identified as Cryptoblabes gnidiella. This finding was also validated by DNA barcoding, as the obtained sequences matched perfectly (100% resemblance) with MG895658 and by 99.85% with OQ564198 that both correspond to C. gnidiella.
Adult exhibits a wing length of 5.0–6.5 mm [23]. The fore wings are dark grey, punctuated by tiny black spots, veiled in white and dotted with reddish scales, characterized by indistinct lighter bands. The hind wings are shiny white and streaked with terminal grey lines [24]. The proboscis and labial palps of the adults are notably elongated, a trait shared by the majority of Phycitinae. The antennae are characterized by a simple structure and a fine covering of cilia. In males, a distinctive horn-shaped projection is present on the third antennal segment, which serves as a key taxonomic character [15] (Figure 1). Larvae exhibit a prominent, dark brown to black, pinaculum ring in conjunction with the SD1 setae on the mesothorax, in addition to a less pronounced pinaculum ring surrounding the SD1 setae on the eighth adnominal segment [23]. Fifth instar larva is 10–12 mm. The dorsal side of the body is characterised by a yellow to light brown colouration, with two narrow, longitudinal darker bands [24] (Figure 2).
3.2. Monitoring of Flight in Pheromone Traps
Pheromones performed were highly species-specific, with adult captures peaking on the 1 September (105 specimens/trap/week) and remaining high (76 specimens/trap/week) until mid-September.
The mean daily temperature recorded locally in the aforementioned sampling region from May to September 2024 (23.9°C ± 0.3) was statistically significantly higher (t-test, p < 0.001) compared to the same months in 2020–2023 (22.6°C ± 0.1). A similar trend was evinced in the mean minimum (16.1°C ± 0.3 vs 15.5°C ± 0.2; p = 0.001) and maximum (32.0°C ± 0.4 vs. 30.4°C ± 0.3; p < 0.001) temperature for the same period. Accordingly, the phenological stages of ‘Xinomavro’ cultivar began with the onset of bud development on the 20 March (BBCH 05), the end of flowering on the 17 May (BBCH 69) and fruit set on the 24 May (BBCH 71). Grape touching stage (BBCH 77) was observed on the 16 June, followed by the beginning of ripening (BBCH 81) on the 24 July. The harvesting process in ‘Xinomavro’ was initiated on the 1 October.
3.3. Description of the Damage
The presence of frass and excrement deposits from the larvae was observed on infested grape bunches. The larvae’s feeding activity resulted in damage to the grapes, with visible signs of infestation including hollow grapes, desiccation and shriveling. Infested bunches were deteriorated due to the development of gray mold, rendering them unsuitable for vinification. Larval feeding damage compromised the overall quality of grapes, negatively impacting the desired key parameters for winemaking and grape bunches were therefore deemed unsuitable and were discarded.
3.4. Estimation of the Damage Level
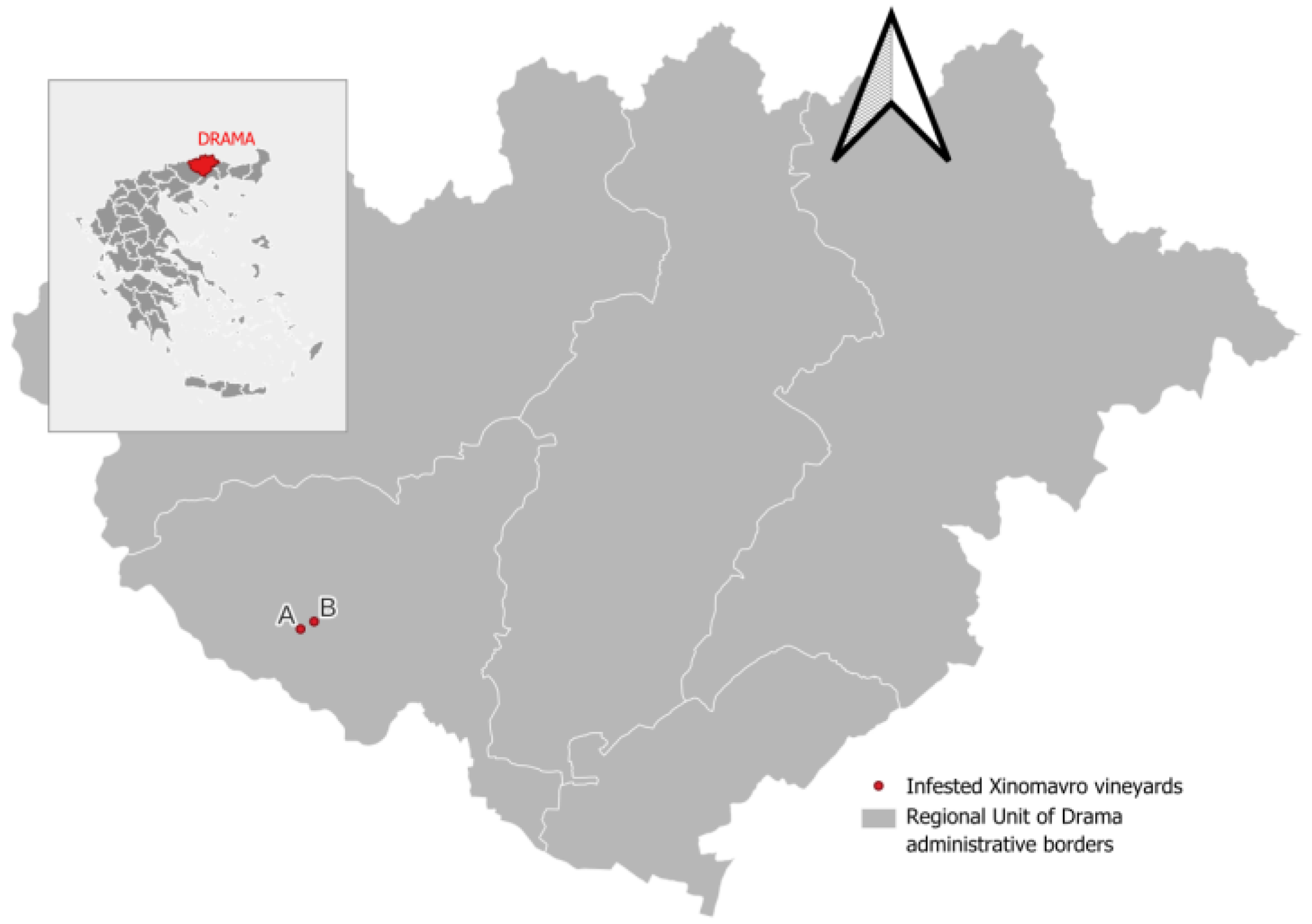
During the harvesting process, it was determined that the late-harvested grape cultivar ‘Xinomavro’ was the sole cultivar affected by the infestation, in two vineyards located at Kali Vrysi (41° 9.80’N, 23° 53.00’E and 41° 10.10’N, 23° 53.80’E) (Figure 3). The level of infestation was estimated to be between 5% and 10% of the total number of bunches harvested from the vineyards of this particular variety. Significant labor costs were expended for the sorting of grapes during the harvesting process and immediately prior to initiating the vinification process. Prior to the beginning of the harvest, an additional sorting operation was conducted to remove infested bunches. It has been estimated that the harvesting rate was approximately 40% of the usual quantity harvested per hour per worker, due to the on-site sorting in the vineyard and the removal of infested bunches.
4. Discussion
This is the first report of Cryptoblabes gnidiella feeding on grapevines in Greece. Specifically, infestations were observed in two organic vineyards, situated in a wine-producing area within the Regional Unit of Drama in Northeastern Greece. This finding triggers further research that is required to assess the potential impact of C. gnidiella on grapevine productivity in Greece, particularly in terms of yield, quality, and economic losses. Lucchi et al. [15] suggest that C. gnidiella has not been regarded a significant pest of grapevines in European vineyards, likely due to its generally low population density coupled with the fact that damage in grape clusters was commonly (and likely erroneously) attributed to other more abundant pests, mainly L. botrana. As a consequence, it seems probable that the actual impact of honeydew moth in Greece is underrated as previous late summer infestations of lepidopteran larvae in vineyards were collectively considered to be L. botrana infestations.
The temperature conditions of the 2024 growing season has been observed to exert a distinctive influence on the phenological stages of the crop. Given that early ripening of grapevines in response to warm conditions is more likely to be caused by shifts in the onset of ripening [32], it is worth noting that the onset of ripening (BBCH 81) was reached in ‘Xinomavro’ on 24 July 2024, whereas it was reached on 20 August the previous year. The prevailing meteorological conditions resulted in an early completion of the harvest, with most of the region’s cultivars, including ‘Assyrtiko’, ‘Malagousia’, ‘Cabernet sauvignon’, ‘Sauvignon blanc’, ‘Limniona’, ‘Chardonnay’, ‘Agiorgitiko’, being harvested at the end of August and beginning of September, several days earlier than the typical harvest period. Harvest in ‘Xinomavro’ initiated on the 1 October, approximately one week before the anticipated date.
The impact of temperature on the progression and spread of infestations appears to be considerable. The mean, minimum and maximum daily temperatures from May to September in 2024 were, respectively, 1.3°C, 0.6°C and 1.6°C higher than those observed over the preceding four years (i.e., 2020 to 2023). Zumbado-Ulate et al. [33] demonstrate that the predicted climatic adaptability for C. gnidiella increases with elevated temperatures along the west coast of the United States.
The late-ripening grape cultivar ‘Xinomavro’ was the only cultivar affected, which is in consistence with previous studies. For example clusters of the late-ripening ‘Gewürztraminer’ cultivar in Uruguay are severely affected at harvest time, whereas ‘Pinot noir’ cultivar, which is harvested earlier, has demonstrated resilience to economic damage, even during high population density years of honeydew moth [28].
A range of organic viticultural practices and biological control methods are used to manage infestations in vineyards. The latter includes mating disruption on a large scale mainly for the management of L. botrana and P. ficus populations [8,34,35]. Biological insecticides and other chemical substances permitted in organic farming are employed only in the event of population outbreak. In 2024, neither L. botrana nor P. ficus were observed in the vineyards of Kali Vrysi, and mating disruption method was implemented exclusively for the former pest. In several Mediterranean coastal areas, the honeydew moth has recently demonstrated a markedly elevated level of harmful activity, characterised by an increased frequency and intensity of infestations. While the underlying causes of this surge remain underexplored, Lucchi et al. [15] postulate that global warming trends and the implementation of mating disruption for the management of L. botrana, with a consequent reduction in the use of pesticides, may be contributing factors.
As the majority of carpophagous insects that infest grapevines, feeding of honeydew moth larvae increase the probability of affected bunches becoming infected by pathogenic rotting agents and other saprophagous insects (Drosophilidae and Nitidulidae). However, the actual extent of this phenomenon varies with microclimate and the phenological stage of grapes [15,24,25]. At present, within the Mediterranean zone, only L. botrana is associated with ochratoxin A (OTA) accumulation in grapes [36], without any concrete evidence that involve C. gnidiella in OTA contamination. Nevertheless, Mondani et al. [36] suggested that given the analogous characteristics of their feeding behavior, it is likely that they could also exert a comparable influence on the enhancement of OTA contamination.
Several studes have conducted to model and describe the phenological stages and to predict the damage cause by the pest. Ringenberg et al. [37] determined the development thresholds and thermal constants for larvae fed on artificial diet. Their findings indicated that C. gnidiella requires a minimum temperature of 12.26°C for its development and a total of 570 degree-days (°C) to complete one generation (egg to adult). Using these parameters, Öztürk [38] assessed the thermal requirements for egg hatching of this species in pomegranates in Mersin (Turkey), and concluded that the first, second, third, fourth and fifth generations require 250, 800, 1375, 1930 and 2500 degree-days (°C), respectively. It has also been established by Wysoki et al. [39] that the preovipositional period lasts for 24 hours following mating. Subsequently, over 50% of the eggs are laid in the first four nights of the female’s lifespan, and the mean fecundity was 105 eggs per female. Finally, using the above-mentioned thermal parameters, Vidart et al. [28] studied the correlation between male catches and damage in Uruguay. Even though pheromone traps detect moths and predict larval feeding on clusters, they proved that damage is more closely related to the cultivar rather than adult population monitoring. It is therefore essential that monitoring efforts are intensified in order to accurately assess the extent of damage caused by C. gnidiella. This should be based on both pheromone traps and visual inspections [15].
As previously proposed by Bagnoli and Lucchi [14], the effective protection of grapes from C. gnidiella necessitates the implementation of effective control measures for both L. botrana and P. ficus. When executed effectively, these measures can drastically reduce the need for pesticide applications targeting at C. gnidiella. Furthermore, application of Bacillus thuringiensis formulations can be additionally beneficial, particularly in organic viticulture when there is an asynchronous outbreak of different pests [14]. However, results have not always been consistent due to various reasons (e.g., insecticidal activity by ingestion, low rain resistance and short residual activity) [15]. In the same direction, mating disruption stands as a promising strategy that needs to be further explored [16,26,40].
The impact of climate change on global viticulture is a complex and multifaceted challenge that requires a comprehensive approach to crop protection strategies. The impact of climate change on grapevine pest insects and their antagonists is becoming increasingly evident. Growers must adapt their plant protection practices to mitigate the risks posed by climate warming. One critical consideration is the effect of rising temperatures and altered precipitation patterns on grapevines, pests, and their natural enemies. The potential for certain pests to expand their geographic ranges or become more abundant in warmer conditions [41] and the possibility of asynchrony between the larvae-resistant growth stages of grapevine and pest [42] represent new threats to growers. Adaptations must, therefore, take these changing dynamics into account. Grower adaptations may include enhanced monitoring of pest populations and adjusting the timing of pesticide applications accordingly. Integrated pest management practices, which combine various control methods, can help growers reduce their reliance on chemical pesticides and develop more sustainable approaches. Additionally, collaboration between researchers, growers, and policymakers is crucial to develop effective and sustainable solutions for crop protection in the face of climate change.
5. Conclusions
The emergence of C. gnidiella as a pest in Greek vineyards highlights the need for increased awareness and research regarding its impact on grapevine productivity. The observed shifts in phenological stages due to rising temperatures underscore the complex interplay between climate change and pest dynamics. Effective management strategies, including integrated pest management and biological control methods, are essential to mitigate the risks posed by C. gnidiella and other pests. As climate change continues to alter the viticultural landscape, growers must adapt their practices to ensure sustainable grape production, emphasizing the importance of collaboration among researchers, growers, and policymakers to develop resilient crop protection strategies.
Author Contributions
Investigation, I.T. and K.B.S.; visualization, K.B.S.; species identification, D.N.A., A.K. and K.B.S; writing—original draft, K.B.S.; writing—review and editing E.R., D.N.A. and A.K.; supervision, E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Data Availability Statement
All data are included in the manuscript.
Acknowledgments
The authors would like to thank the agronomist Mr Hadjitheodorou Theologos for his technical assistance in the field and Mrs. Paraskevi Skarpa for her assistance in the preparation of the mauscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Aitken, A.D. A Key to the Larvae of Some Species of Phycitinae (Lepidoptera, Pyralidae) Associated with Stored Products, and of Some Related Species. Bull. Entomol. Res. 1963, 54, 175–188. [CrossRef]
- Triplehorn, C.A.; Johnson, N.F.; Borror, D.J. Borror and DeLong’s Introduction to the Study of Insects; 7. ed., [3. Nachdr.].; Thompson Brooks/Cole: Belmont, CA, 2006; ISBN 978-0-03-096835-8.
- Liu, C.; Yang, M.; Li, M.; Jin, Z.; Yang, N.; Yu, H.; Liu, W. Climate Change Facilitates the Potentially Suitable Habitats of the Invasive Crop Insect Ectomyelois ceratoniae (Zeller). Atmosphere 2024, 15, 119. [CrossRef]
- Pachkin, A.; Kremneva, O.; Leptyagin, D.; Ponomarev, A.; Danilov, R. Light Traps to Study Insect Species Diversity in Soybean Crops. Agronomy 2022, 12, 2337. [CrossRef]
- Simoglou, K.B.; Karataraki, A.; Roditakis, N.E.; Roditakis, E. Euzophera bigella (Zeller) (Lepidoptera: Pyralidae) and Dasineura Oleae (F. Low) (Diptera: Cecidomyiidae): Emerging Olive Crop Pests in the Mediterranean? J Pest Sci 2012, 85, 169–177. [CrossRef]
- Powell, J.A., 2009. Lepidoptera (Moths, Butterflies). In: Cardι, R.T., Resh, V.H., 2009. Encyclopedia of insects, 2nd ed. ed. Elsevier/Academic Press, Amsterdam London.
- Benelli, G.; Lucchi, A.; Anfora, G.; Bagnoli, B.; Botton, M.; Campos-Herrera, R.; Carlos, C.; Daugherty, M.P.; Gemeno, C.; Harari, A.R.; et al. European Grapevine Moth, Lobesia botrana Part I: Biology and Ecology. Entomologia 2023a, 43, 261–280. [CrossRef]
- Benelli, G.; Lucchi, A.; Anfora, G.; Bagnoli, B.; Botton, M.; Campos-Herrera, R.; Carlos, C.; Daugherty, M.P.; Gemeno, C.; Harari, A.R.; et al. European Grapevine Moth, Lobesia botrana Part II: Prevention and Management. Entomologia 2023b, 43, 281–304. [CrossRef]
- Varikou, K.; Birouraki, A.; Bagis, N.; Kontodimas, D.C. Effect of Temperature on the Development and Longevity of Planococcus ficus (Hemiptera: Pseudococcidae). Annals of the Entomological Society of America 2010, 103, 943–948. [CrossRef]
- Karamaouna, F.; Menounou, G.; Stathas, G.J.; D.N. Avtzis. First record and molecular identifi cation of the parasitoid Anagyrus sp. near pseudococci Girault (Hymenoptera: Encyrtidae) in Greece - Host size preference for the vine mealybug Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). Hellenic Plant Protection Journal 2011, 4, 45–52, Available at: https://www.hppj.gr/wp-content/uploads/2015/02/Volume-4-Issue-2-July-2011.pdf.
- Evangelou, V.; Lytra, I.; Krokida, A.; Antonatos, S.; Georgopoulou, I.; Milonas, P.; Papachristos, D.P. Insights into the Diversity and Population Structure of Predominant Typhlocybinae Species Existing in Vineyards in Greece. Insects 2023, 14, 894. [CrossRef]
- Roditakis, N.E. First Record of Frαnkliniellα occidentαlis in Greece. Entomologia Hellenica 1991, 9, 77. [CrossRef]
- Roditakis, E.; Roditakis, N.E. Assessment of the Damage Potential of Three Thrips Species on White Variety Table Grapes—In Vitro Experiments. Crop Protection 2007, 26, 476–483. [CrossRef]
- Bagnoli, B.; Lucchi, A. Bionomics of Cryptoblabes gnidiella (Milliθre) (Pyralidae Phycitinae) in Tuscan vineyards. IOBC/WPRS Bulletin 2001, 24(7), 79–84. Available at: https://iobc-wprs.org/product/iobc-wprs-bulletin-vol-24-7-2001/.
- Lucchi, A.; Ricciardi, R.; Benelli, G.; Bagnoli, B. What Do We Really Know on the Harmfulness of Cryptoblabes gnidiella (Milliθre) to Grapevine? From Ecology to Pest Management. Phytoparasitica 2019, 47, 1–15. [CrossRef]
- Ricciardi, R.; Di Giovanni, F.; Cosci, F.; Ladurner, E.; Savino, F.; Iodice, A.; Benelli, G.; Lucchi, A. Mating Disruption for Managing the Honeydew Moth, Cryptoblabes gnidiella (Milliθre), in Mediterranean Vineyards. Insects 2021, 12, 390. [CrossRef]
- French Institute of Vine and Wine. The economic cost of Cryptoblabes gniediella. Institut Franηais de la Vigne et du Vin, 2024. Accessed on 26-10-2024. Available at: https://www.vignevin.com/article/le-cout-economique-de-cryptoblabes-gniediella/.
- Avidov, Z.; Gothilf, S. Observations on the honeydew moth (Cryptoblabes gnidiella Milliere) in Israel. Ktavim 1960, 10(3–4), 109–124. Available at: https://download.ceris.purdue.edu/file/2088.
- Dawidowicz, Ł.; Rozwałka, R. Honeydew Moth Cryptoblabes gnidiella (MILLIΘRE, 1867) (Lepidoptera: Pyralidae): An Adventive Species Frequently Imported with Fruit to Poland. Polish Journal of Entomology 2016, 85, 181–189. [CrossRef]
- Elnagar, H. Population Dynamic of Honeydew Moth, Cryptoblabes gnidiella Miller in Vineyards Orchards. Egyptian Academic Journal of Biological Sciences. A, Entomology 2018, 11, 73–78. [CrossRef]
- Velez-Gavilan, J. Cryptoblabes gnidiella (honeydew moth). CABI, 2022, 16381. Accessed on 13-10-2024. Available at: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.16381.
- Karsholt, O.; Razowski, J. The Lepidoptera of Europe: A Distributional Checklist; Apollo books: Stenstrup, 1996; ISBN 978-87-88757-01-9.
- Neunzig, H.H. The Moths of America, North of Mexico. Fasc. 15,2: Fasc. 15. Pyraloidea / H. H. Neunzig Pyralidae (Pt.), Phycitinae (Pt; Neunzig, H.H., Ed.; Classey: London, 1986; ISBN 978-0-933003-01-9.
- Ioriatti, C.; Lucchi, A.; Varela, L.G. Grape Berry Moths in Western European Vineyards and Their Recent Movement into the New World. In Arthropod Management in Vineyards:; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer Netherlands: Dordrecht, 2012; pp. 339–359 ISBN 978-94-007-4031-0.
- Elekcioglu, N.Z.; Olculu, M. Pest, predator and parasitoid species in persimmon orchards in the eastern Mediterranean region of Turkey, with new records. Fresenius Environmental Bulletin 2017, 26, 5170-5176.
- Harari, A.R.; Zahavi, T.; Gordon, D.; Anshelevich, L.; Harel, M.; Ovadia, S.; Dunkelblum, E. Pest Management Programmes in Vineyards Using Male Mating Disruption. Pest Management Science 2007, 63, 769–775. [CrossRef]
- Keçeci̇, M. The Population Dynamic of Honeydew Moth [Cryptoblabes Gnidiella Mill. (Lepidoptera: Pyralidae)] in Vineyard in Antalya. Mediterranean Agricultural Sciences 2021, 34, 169–173. [CrossRef]
- Vidart, M.V.; Mujica, M.V.; Calvo, M.V.; Duarte, F.; Bentancourt, C.M.; Franco, J.; Scatoni, I.B. Relationship between Male Moths of Cryptoblabes gnidiella (Milliθre) (Lepidoptera: Pyralidae) Caught in Sex Pheromone Traps and Cumulative Degree-Days in Vineyards in Southern Uruguay. SpringerPlus 2013, 2, 258. [CrossRef]
- JASP Team. JASP (Version 0.19.3) -Computer software. Available online: https://jasp-stats.org (accessed on 22 November 2024).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I 273 from diverse Metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299.
- Avtzis, D.N.; Markoudi, V.; Mizerakis, V.; Devalez, J.; Nakas, G.; Poulakakis, N.; Petanidou, T. The Aegean Archipelago as Cradle: Divergence of the Glaphyrid Genus Pygopleurus and Phylogeography of P. Foina. Systematics and Biodiversity 2021, 19, 346–358. [CrossRef]
- Sadras, V.O.; Petrie, P.R. Climate Shifts in South-Eastern Australia: Early Maturity of Chardonnay, Shiraz and Cabernet Sauvignon Is Associated with Early Onset Rather than Faster Ripening: Climate Shifts and Grapevine Ripening. Australian Journal of Grape and Wine Research 2011, 17, 199–205. [CrossRef]
- Zumbado-Ulate, H.; Schartel, T.E.; Simmons, G.S.; Daugherty, M.P. Assessing the Risk of Invasion by a Vineyard Moth Pest Guild. NB 2023, 86, 169–191. [CrossRef]
- Moschos, T.; Souliotis, C.; Broumas, T.; Kapothanassi, V. Control of the European Grapevine moth Lobesia botrana in Greece by the Mating Disruption Technique: A Three-Year Survey. Phytoparasitica 2004, 32, 83–96. [CrossRef]
- Daane, K.M.; Yokota, G.Y.; Walton, V.M.; Hogg, B.N.; Cooper, M.L.; Bentley, W.J.; Millar, J.G. Development of a Mating Disruption Program for a Mealybug, Planococcus ficus, in Vineyards. Insects 2020, 11, 635. [CrossRef]
- Mondani, L.; Palumbo, R.; Tsitsigiannis, D.; Perdikis, D.; Mazzoni, E.; Battilani, P. Pest Management and Ochratoxin A Contamination in Grapes: A Review. Toxins 2020, 12, 303. [CrossRef]
- Ringenberg, R.; Botton, M.; Garcia, M.S.; Nondillo, A. Biologia Comparada e Exigκncias Tιrmicas de Cryptoblabes gnidiella Em Dieta Artificial. Pesq. agropec. bras. 2005, 40, 1059–1065. [CrossRef]
- Öztürk, N. Creating a Degree-Day Model of Honeydew Moth [Cryptoblabes Gnidiella (Mill., 1867) (Lepidoptera: Pyralidae)] in Pomegranate Orchards. Turkish Journal of Entomology 2018, 53–62. [CrossRef]
- Wysoki, M.; Ben Yehuda, S.; Rosen, D. Reproductive Behavior of the Honeydew Moth, Cryptoblabes gnidiella. Invertebrate Reproduction & Development 1993, 24, 217–223. [CrossRef]
- Acin, P. Management of the honeydew moth by mating disruption in vineyard. IOBC/WPRS Bulletin 2019, 146, 28–31. Available at: https://iobc-wprs.org/product/iobc-wprs-bulletin-vol-146-2019-copy/.
- Reineke, A.; Thiιry, D. Grapevine Insect Pests and Their Natural Enemies in the Age of Global Warming. J Pest Sci 2016, 89, 313–328. [CrossRef]
- Caffarra, A.; Rinaldi, M.; Eccel, E.; Rossi, V.; Pertot, I. Modelling the Impact of Climate Change on the Interaction between Grapevine and Its Pests and Pathogens: European Grapevine Moth and Powdery Mildew. Agriculture, Ecosystems & Environment 2012, 148, 89–101. [CrossRef]
Figure 1.
(a) Grape ‘Xinomavro’ bunch infested by Cryptoblabes gnidiella larvae; (b) Cryptoblabes gnidiella larva on grape; (c) Dark pinacula in conjunction with the SD1 setae on the larval mesothorax; (d) Dark pinacula surrounding the SD1 setae on the eighth adnominal segment of the larva.
Figure 1.
(a) Grape ‘Xinomavro’ bunch infested by Cryptoblabes gnidiella larvae; (b) Cryptoblabes gnidiella larva on grape; (c) Dark pinacula in conjunction with the SD1 setae on the larval mesothorax; (d) Dark pinacula surrounding the SD1 setae on the eighth adnominal segment of the larva.

Figure 2.
(a) Male individual of Cryptoblabes gnidiella caught on a pheromone sticky trap; (b) A horn-shaped projection is present on the males’ third antennal segment; (c) Detail of the horn-shaped projection on the males’ third antennal segment.
Figure 2.
(a) Male individual of Cryptoblabes gnidiella caught on a pheromone sticky trap; (b) A horn-shaped projection is present on the males’ third antennal segment; (c) Detail of the horn-shaped projection on the males’ third antennal segment.

Figure 3.
Locations of vineyards infested by Crypboblabes gnidiella; Kali Vrysi, Drama, Greece.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



