
Submitted:
27 November 2024
Posted:
28 November 2024
You are already at the latest version
Abstract
Antimicrobial drug resistance has emerged as a significant public health challenge, exacerbated by the rise of multidrug-resistant strains and the emergence of new infections. This has intensified the need for the development of novel therapeutic agents. Antibiotics, originally derived from microorganisms, are compounds that either inhibit or kill other microbes. Historically, most antibiotics used in clinical settings were produced by Actinomycetes and fungi, particularly during the golden age of antibiotic discovery (1940s to 1960s). However, progress in drug development slowed in the following decades due to challenges in cultivation techniques, including isolation difficulties, microbial overgrowth, and discrepancies between natural environments and laboratory conditions. Recently, innovative approaches, such as advancements in cultivation methods, modification of existing compounds, gene induction, exploration of animal microbiota, co-cultivations, and identification of alternative antimicrobial targets, have provided new pathways for drug discovery. Additionally, cutting-edge molecular tools like metagenomics have enabled the exploration of bioactive molecules from uncultured microbes in diverse microenvironments, potentially ushering in a new era in antibiotic development. This review examines both past and current methodologies in antimicrobial drug discovery, highlighting future perspectives for the field.
Keywords:
drug discovery
; antimicrobial agents
; metagenomics
; animals’ microbiota
; co-cultivation
1. Introduction
Antimicrobial agents are a broad term for natural or synthetic chemical substances that inhibit or kill different microorganisms. At the same time, antibiotics offer a more specific definition as the natural substances produced by microorganisms that inhibit or kill other microorganisms’ growth, especially bacteria [1]. Nature contains diverse microorganisms that remain an under-exploitation resource of antimicrobial agents. Seeking novel antimicrobials from natural sources has become one of the most fundamental efforts to encounter and overcome microbial infections [2].
Pathogenic microorganisms always pose vital threats to human and animal health. Indeed, infectious diseases are the second principal cause of death all over the world [3,4]. Humans have been infected by microbes from the past times to the present [5]. Pathogens like Escherichia coli, Klebsiella pneumonia, and Pseudomonas aeruginosa cause various skin and subcutaneous infections, bacteremia, septicemia, as well as lower and upper respiratory tract infections [6,7]. Moreover, E. coli is implicated in the urinary tract and gynecological infections, particularly in young adult and immunosuppressed patients. Staphylococcus aureus causes skin, gastrointestinal, joint, and bone infections, as well as bronchitis of the respiratory system [8]. Candida albicans cause mouth thrush urinary and genital candidiasis [9].
Researchers have been developing synthetic antimicrobial agents that originated from microbial sources. Interestingly, over and improper use of such drugs in humans and other animals, and their rising appearance continually in water, food, and soil habitats, lead to the serious problem of antimicrobial resistance [10,11,12]. It is predicted that resistance to antimicrobials leads to about 700,000 fatalities per year throughout the globe; by 2050, the figure is expected to grow to 10 million, according to current estimates [13]. Nowadays, AMR has become the leading health problem caused by many antibiotics that do not work against pathogenic microbes. After more than fifty years of widespread usage of antibiotics, so-called “miracle drugs,” antibiotics are less effective than before. Thus, there is a fast increase in the quantity of strains of multiple drug-resistant (MDR). Many infectious agents are showing resistance toward antibiotics. In addition, there is a significant decrease in the antibiotics manufacturing of new synthetic.
In light of this growing crisis, researchers have been actively seeking alternative solutions to combat AMR [14]. One promising avenue involves the development of synthetic materials, including nanoparticles, which have shown significant potential as antimicrobial agents [15,16,17]. Nanoparticles, due to their unique size, surface area, and reactivity, can interact with microbial cells in ways that traditional antibiotics cannot. These nanoparticles, especially those biosynthesized from plant, fungal, or microbial sources, offer an eco-friendly and sustainable approach to tackling resistant pathogens. Recent studies have focused on exploring nanoparticles such as silver, copper, and selenium as effective antimicrobial agents, often exhibiting potent activity against both gram-positive and gram-negative bacteria, including MDR strains [7,18] Additionally, the use of synthetic antimicrobial peptides and other novel materials has shown promise in overcoming the limitations of conventional antibiotics. This approach aligns with the growing need for innovative solutions to address the alarming rise of AMR and the shortage of new antibiotics in the pharmaceutical pipeline.
Moreover, synthetic antibiotics on the host are linked with many side effects such as allergic reactions, immunological suppression, and hypersensitivity [4]. Furthermore, strains resistant to nearly all antibiotics have been discovered in clinical trials and have made a generalized impact on human life. Resistance emerges due to microbes‘ mutations. Antibiotic dosage of sub-inhibitory assists step-by-step resistance selection, and the originating clones of resistant such as Methicillin-resistant Staphylococcus aureus, pseudomonas aeruginosa, Klebsiella, and Escherichia coli are rapidly dispersed [19].
The strategies for controlling antimicrobial resistance comprise utilizing antibiotics carefully, specifically on clinical and farms, improving investigation and study areas related to drug discovery, and developing innovative tools and molecular techniques [2,20]. Current improvements in practices of cultivation, analytical skills, and genome-driven techniques have significantly prolonged access to our previously untapped resources of microbes. Such methods allow us to access novel microbe reservoirs and unlock their chemical potential, thus creating new possibilities for developing new antimicrobial medicines [21]. Finally, most critically, we must step up our efforts to find new antimicrobial drugs to fight infections that are resistant to existing antimicrobial treatments.
In this review, we suggest remembering the historical background regarding antimicrobial drug discovery and their structural natures. Moreover, it concentrates on recent and current promising approaches concerning culture-dependent and culture-independent for discovering and identifying novel bioactive compounds with potential antimicrobial activities. Finally, we summarize the future perspectives related to incorporating innovative approaches for finding a new class of antimicrobial drugs.
2. History of Antimicrobial Drug Discovery
Several natural remedies for infection, such as honey, herbs, and even the feces of animals and humans, are effective in the historical record [22]. One of the most effective therapies was the topical utilization of moldy bread, documented in ancient Rome, Greece, Serbia, China, and Egypt as having significant benefits [23]. Several heavy metals, including arsenic, bismuth, and mercury, were tested specifically; tests were administered systemically or locally in mainly provided syringes. The primary possible antibiotic for being applied in human infection treatments was Pyocyanin [24]. Initially, Paul Ehrlich was interested in creating stains for histological inspection, and he noticed that certain stains were poisonous to bacteria [25]. Ehrlich created several antibacterial dyes that are still in use today. In 1909, Paul Ehrlich developed arsphenamine, a derivative of arsenic that was effective against the syphilis-causing bacteria Treponema pallidum. Salvarsan was the brand name of this antibiotic when it was first introduced to the market in 1911. Gerhard Domagk discovered the antibacterial properties of sulphanilamide in 1930, and this antibiotic was later commercialized under the brand name Prontosil [26]. Terrestrial microbes are an abundant source of structurally various biologically active substances. The story of the penicillin discovery by Alexander Fleming (1881–1955) in 1928 is well-known to most people; one year later, Joseph Lister (1827– 1912) experimented with Penicillium glaucium (revealing its activity of antibacterial on tissues of humans) in the laboratory [27]. Howard Florey and Ernst Chain described the purification of penicillin, which led to the drug becoming accessible for restricted use in 1945 [28]. Selman Waksman was pioneered the systematic investigation of the soil bacteria’s antimicrobial action, predominantly from streptomycetes [29]. He invented cultural techniques and tactics to draw attention to the antagonistic relationships between microbial species. As a result, numerous vital antibiotics and antifungals, including clavacin from Aspergillus clavatus, fumigacin from Aspergillus fumigatus, neomycin from Streptomyces fradiae, streptomycin from Streptomyces griseus, and actinomycin from Streptomyces were discovered. Streptomycin, neomycin, and actinomycin are among the antibiotics still in use today [20]. Teicoplanin, discovered from the Actinoplanes teichomyceticus, is considered the first of the novel glycopeptides. Even though certain antibiotics have just been commercialized, the most current groups of antibiotics discovered date back to the 1980s. No recent groups have been discovered in 50 years of research. As a result, new and creative methods are needed [30]. Immediately after developing the culture technique via Waksman’s platform, the pharmaceutical industry shifted its attention to the in vitro synthesis of novel compounds according to knowledge about the well-known antibiotic action mechanism. There have been just a few additions to the antibiotic family throughout the years: in 1953 nitrofuran, in 1960 quinolones, in 1961 sulphonamides, and in 1987 Oxazolidinones, to name a few. In addition, modifications and enhancements of previously discovered compounds have been accomplished. This has been diverted due to the commercialization of Daptomycin in 2001 and linezolid in 2003, despite these compounds being discovered as far back as 1986 and 1955 [31]. The discovery of cephalosporin, a novel cefiderocol that is effective against Gram-negative bacteria of carbapenem-resistant, has just recently been made [32]. Semisynthetic chemicals were also created that originated from natural sources, including ketolides made from metronidazole or macrolides that they produced from Streptomyces spp [31]. More than 1200 antimicrobial peptides (AMPs) have been identified from various sources, including animals and plants, but none have been successfully utilized as antibiotics [33]. Novel classes of antimicrobial agents were discovered from bacterial and fungal species, as shown in (Table 1).
3. Structural Natures of Antimicrobials
A vast range of chemical components characterizes antimicrobial substances. Natural secondary metabolites and bioactive products are often found in the environment. However, they are not required for the microbes‘ development and growth; they provide certain advantages, such as competition and protection [34]. It is possible to classify all antimicrobial agents used in human medicine into six groups based on the chemical nature of molecules; the first group is polyketides (PKs), which are derivatives of malonyl coenzyme A or acetyl coenzyme A. The second is nonribosomal peptides (NRPs), derivatives of amino acids that are not produced by the ribosome. The third kind of drug is a hybrid of PKs and NRPs, while the fourth type is made up of many units of carbohydrate that have been substituted with groups of amine (aminoglycosides). The fifth comprises various chemicals, such as alkaloids and terpenoids (metronidazole) [20]. The sixth category includes antibiotics such as antimicrobial peptides (AMPs) identified in mammals, amphibians, plants, and insects. These are usually categorized as ‘peptides of ribosomally produced and post-translationally modified peptides (RiPPs) [35]. In order to synthesize NRPs and PKs, multi-enzyme complexes known as nonribosomal peptide synthases enzyme complex and polyketides synthetase enzymes, respectively, are required, encoded by biosynthesis gene clusters [36]. PK antibiotics include macrolides, mupirocin, and tetracyclines, while NRP antibiotics include β-lactams, polymyxins, lincomycin, and vancomycin. Rifampicin, Kirromycin, and Rapamycin belong to the hybrid of NRPs and PKs [34]. RiPPs include Lanthipeptides, Thiopeptides, Cyanobactins, and bottromycin [37].
4. Approaches in Antimicrobial Drug Discovery
4.1. Culture Techniques
Most natural products of commercial value, such as antibiotics or other drugs, are derived from Soil microorganisms that have been cultivated [38]. It was in the 1940s that Selman Waksman developed a technique of routinely screening soil microorganisms for antagonisms, and this culture-based approach is still in use today [29]. According to specific estimates, approximately 0.1–1 percent of the total soil population may be cultured [39]. In 1 g of soil, 107 cells were observed in research conducted by Kellenberger (2001) [40], but just 0.1 percent of the cells were culturable, according to the findings. As a result, 99.9 percent of the genetic diversity in this group was lost due to the difficulties encountered during the isolation and enrichment of microorganisms. There are numerous techniques of innovative cultivation available to overcome these difficulties; one recent technique to the rise revival for strains of less culturable is substituting traditional isolation media of nutrient-rich with isolation media of oligotrophic because the application of isolation media of oligotrophic inhibits the overgrowth of microbial species of fast-growing, particular group of microorganisms can be cultured [41]. Another recent technique for the rising revival of strains less culturable is substituting traditional isolation media with methods for the separation of bacteria from complicated settings, including the creation of mimicked natural environments and the encapsulation of cells in gel microdroplets, have been developed [39]. Sixty-seven percent of the cultures generated from microdroplets could grow to densities above 107 cells/ml when the cultures were incubated in a nutrient-rich medium, according to Zengler et al. (2002) [42] findings. In this way, these new strains may be deployed to discovery platforms of natural products with relative simplicity.
There are two chief techniques concerning investigating the antimicrobial activity of environmental isolates. Every approach follows a similar principle: a test strain inhibition on an indicator strain of closely cultivated. The test strain is the strain supposed to produce an antimicrobial aiming the strain used as an indicator.
4.1.1. Streaking and Spreading Agar Plate Technique
This technique involves inoculating the strain of bacterial test vertically on a plate of agar using a needle. The time required for incubation of the plate depends on the time required for the bacterial strain to reach the phase, that is, the point at which secondary metabolites are produced. The time of incubation is measured in minutes. It is later cultured for another 24 hours, after which the indicator strain is injected into a horizontal stripe on the plate. This method is simple and effective for screening; however, it requires that both strains of bacteria be grown in the same conditions of culture (pH, environment, and temperature) [43]. The method of spreading consists of putting a drop of the test strain on a lawn of the indicator strain that has been dispersed. Following the incubation period, an inhibitory zone is seen surrounding the growth of the test strain (Figure 1) [44].
4.1.2. Well Diffusion Agar Technique
According to the antimicrobials diffusion via agar (Figure 1), which limits the growth of sensitive microorganisms. Using aseptic techniques, a plate of agar is inoculated by a lawn of the indicator strain or pooled by the strain of indicator; later, wells are punched in the agar aseptically. There are two main variations to consider. The first technique is the diffusion of agar plug, which involves deleting an agar cylinder in a plate previously inoculated with the test strain and allowing it to diffuse into the plate. It is necessary to insert this cylindrical piece of agar into a corresponding well on the plate of the indicator [45]. The second variation technique is involving the insertion of growth supernatant of the test strain liquid broth into a well on an agar plate on which previously inoculated with the indicator strain [46]. A rest period of optional degrees Celsius is followed by incubation of the agar plate, with the inhibitory growth zone being measured around the well. Many variations have been developed by including iron chelators or stress situations. The most significant limitation of solid culture assays is that they cannot be performed on species of bacteria with different growth circumstances or are fastidious [47]. Approaches based on liquid culture may be applied to address this issue. Because of the existence of the Waksman platform, co-culture of liquid broth has been utilized to cultivate bacteria. In this case, the strain of indicator and the test species are grown together in the same culture while separated with a filter that allows nutrients to disperse but does not enable cell dispersion [29]. The indicator strain’s growth is monitored after incubation and measured by optical density, coloring, or enumeration. Adding the test species growth supernatant, previously concentrated and filtered, to an indicator strain liquid culture provides an additional technique [48].
4.2. Biochemical Modification of Already Known Molecules
Sometimes, the original natural products do not essentially produce the most bioactive and effective drugs. In these cases, alterations of particular structural frameworks are desired to create products with acceptable pharmacologic properties. Primary structures must be influenced biologically or chemically [36]. For instance, altering erythromycin into the new desirable azithromycin involves synthetic biochemical modifications of the primary molecule. Azithromycin would be manufactured in perfect condition by an engineered microbial host [49]. Alteration of cephalosporins led to the creation of cefiderocol that shows tolerability and safety in healthy individuals; clinical trials to treat urinary system infections are ongoing [50]. Another instance is the modification in the sisomicin of aminoglycoside, which led to the creation of plazomicin [51]. It can be expected that synthetic biology may contribute meaningfully to scaffold-based innovation of therapeutic drugs.
4.3. Induction of Silent Genes Encoding Bioactive Molecules
Silent genes encode the synthesis of potential biologically active secondary metabolites while induced or stimulated. These genes are observed in various actinobacterial DNA genomes. They typically do not encode metabolites under standard laboratory growth conditions. Recently, numerous innovative works have been considered for triggering these silent clusters of genes to discover novel secondary metabolites. In this regard, strains of diverse expression are established to enable the recognition of metabolite and expressing gene clusters [52]. For example, the sample microorganism Streptomyces coelicolor is engineered to genetically mark antibiotic production positively through altering ribosome and RNA polymerase [53]. In gene clusters, heterologous expressions are encountered long because cluster measures could gain over 100 kb, making it hard to clone. Currently, much-expanded approaches are accessible, such as the cloning system of transformation-related recombination, which is an approach newly accepted in Saccharomyces cerevisiae which in actinomycetes encourages expressing and cloning huge fragments of DNA [54].
Moreover, the information accessible in analyses of gene clusters could decisively be applied for operating pathway particular regulations such as exchanging the natural promoter with a robust synthetic one, stimulating the transcription of the biosynthesis genes [55]. The stimulation of a silent glycopeptides gene cluster is defined for Amycolatopsis japonicum. Introducing the transcriptional activator specific for a glycopeptide caused the manufacturing of ristomycin, an antibiotic for glycopeptide [56]. Microbial co-culturing, the addition of unusual environmental factors, nutritional signals, stress conditions such as heat stress or ethanol, and application of N-acetylglucosamine of cell wall metabolite are effectively shown to provoke the expressing of the clusters of the silent gene [57]. This permits the exploitation of natural product manufacturers more thoroughly than in previous years and would initiate the identification of several novel secondary metabolites in the near future.
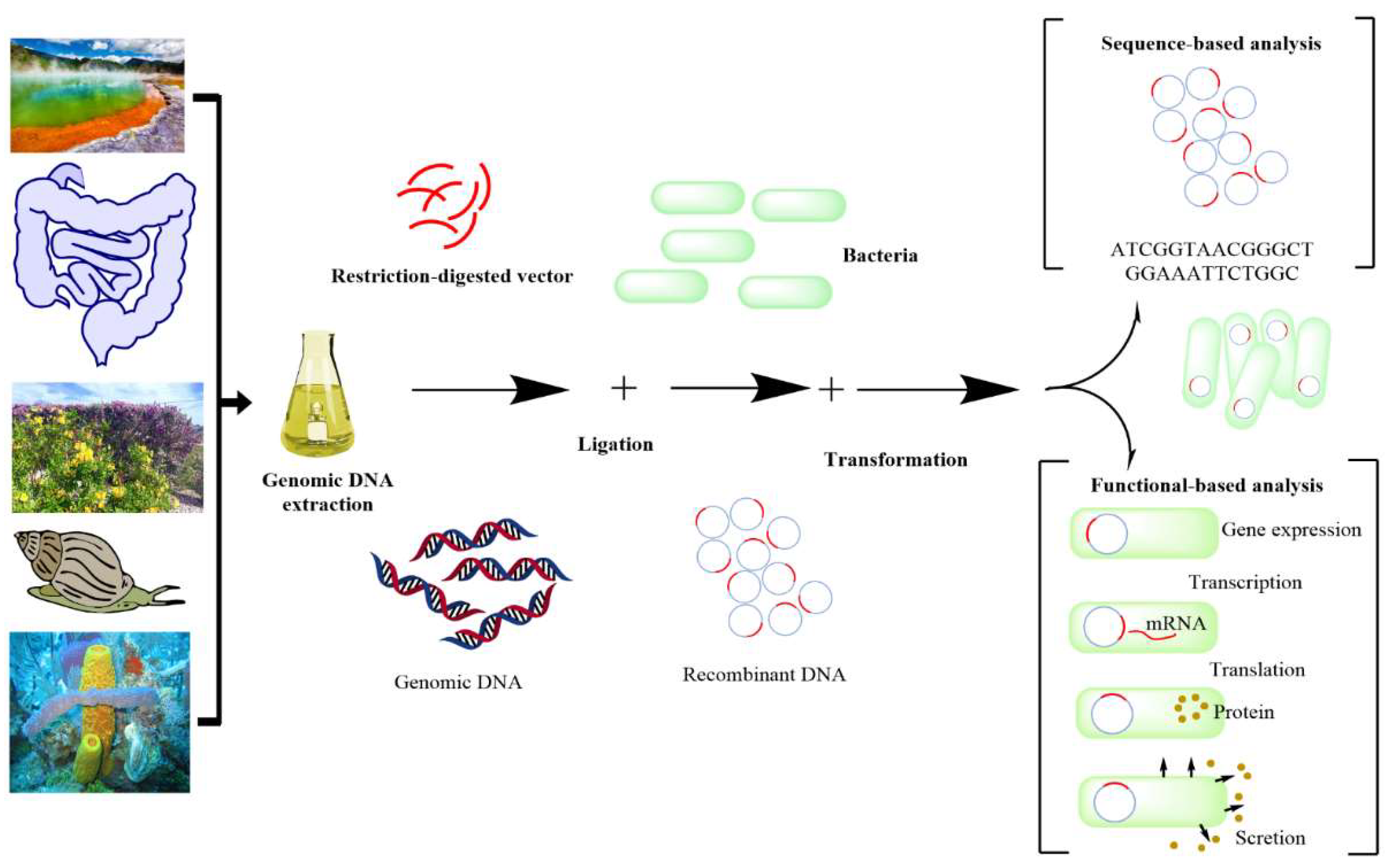
4.4. Metagenomic Tools and Uncultured Microbes
As an approach, Metagenomics (Figure 2) aims to get accessibility to the microbial species‘ biosynthetic capacity, which has not been previously cultivated. Metagenomics is possible to provide an ideal toolkit to bring a variety of biosynthetic in samples of the environment in drug discovery platforms by obtaining DNA from different environments and successively recognizing, separating, and expressing biosynthetic gene clusters in cells of heterologous hosts [58,59] Alternative methods for obtaining the metagenome of soil involve a genome extraction of nucleic acid from environmental samples, amplification of DNA by polymerase chain reaction (PCR), and DNA sample direct cloning [60,61]. These environmental DNA-based methods do not rely on the isolation and cultivation of specific microbes and, as a result, reduce the bias associated with such procedures. Genetic material is collected in samples of the environment and put into vectors, including genomes of plasmids and bacterial artificial chromosomes (BACs), after which it is multiplied in certain strains of bacteria, including Escherichia coli, according to the standard procedure. In theory, the complex libraries of genomics that emerge from this process provide accessibility for the whole content of metagenomics in soil. It is suggested that a library of DNA showing the entire soil sample metagenome will require about 106 BAC clones by inserting almost 100 kb. However, this is not confirmed [62]. The creation of complicated libraries derived from samples of soil, as well as the beneficial genes cloning, is dependent on the high quality of the extracting DNA, as the alterations of enzymatic needed in the cloning phases are responsive to every contamination with various abiotic and biotic elements including humic constituents, which can be harmful. Various methods are available to isolate soil DNA [63]. In order to isolate DNA from soils, methods must be developed that include either the recovery of bacterial cells and cells direct lysis or subsequent lysis in the sample followed by DNA extraction. Freeze-thawing, mechanical disruption, strong detergents, and high temperatures are all standard methods of cell lysis used in direct lysis procedures. Direct lysis methods have been frequently suggested since they are more suitable for various soil types, require less work, and yield more DNA [64,65]. Direct lysis practices are also suggested more frequently than techniques of fractionation. As stated, the fractionation approach is advantageous for soil samples that contain a high concentration of substances that interfere with the purification and separation of DNA, as the microbes’ concentration and recovery prior to lysis could prevent several complications associated with such contaminations. This method is preferred because the isolated DNA measure is more considerable than that typically obtained with the direct lysis technique. This technique is used to get DNA, and the DNA obtained via this process has been used to create large-insert libraries [61].
The screening of metagenomic libraries of soil samples may be done in two ways: by looking at the recombinant strains‘ enzymatic activity or how closely the sequences are related. As opposed to sequence-based screening, activity-driven screening (functional-driven screening) does not require information originating from the previously recognized sequences, so it can discover new genes encoding types and groups of enzymes or recognize the synthesizing active compounds of innovative biological activity. Moreover, it is optional to yield functional and full-length genes, as well as other characteristics. One drawback is that the cloned gene expression is dependent on the gene’s expression with an external host (Gillespie et al. 2002). According to a study [66,67], E. coli was successfully used as a host in the screening of DNA of soil-derived metagenomic in the presence of small biomolecules and new biocatalysts. The advantage of applying E. coli is that it is often used in industry fermentations, which reduces the risk of contamination. Consistently, techniques for downstream processing, separation, and production of streamlining batches are becoming well-established. This indicates that several steps in developing highly valued products are mastered before genes are sequenced and cloned [39].
One advantage of sequence-based screening is that it is independent of cloned gene expression with foreign hosts, which is a disadvantage of traditional screening methods. In addition, this technique may be used in conjunction with other screening approaches, like polymerase chain reaction (PCR) or colony hybridization, to screen for various targets [68]. It is necessary to construct primers and probes of DNA generated from conserved portions of families of genes or proteins discovered before using sequence-based techniques. As a result, solely new variations of well-known functional families of proteins may be discovered [69]. It is possible to get genomes in uncultured soil microorganisms using this technique, and it may also provide information on unidentified species physiology [70]. Studies on small biomolecules revealed that soil monitoring metagenomic libraries may be used as a drug discovery technique. In turbomycin, A and B, the isolation of these antibiotics serves as an example [66]. In a screening effort that included more than 1,000 bacterial genomes, more than 30,000 gene clusters of putative biosynthetics that code for bioactive chemicals were discovered [52]. Since several clusters of the biosynthetic gene-producing antimicrobial compounds and other small biomolecules are between 30-100 kb in size, using large insert libraries is beneficial for drug discovery. It has consistently been shown that variations among the soil metagenome and microorganisms in the soil provide an abundant, highly concentrated source of new natural products [39].
4.5. Microbiota and Bioactive Metabolites
Different microbes populate the gastrointestinal tract of animals, typically referred to as the microbiota of the digestive tract, bacteria are dominant, while Archaea, fungi, yeast, and protozoa also reside inside GIT [71]. However, most parts of these microbes are still unculturable [72]. The intestinal microbiota consisted of 1011 microbes per gram of feces for hundreds of different species [73]. The host and microbiota interactions in healthy people are symbiotic in that the host prepares the microbiota with nutrient materials in a balanced and steady environment. The microbiota assists in the digestion of metabolizable substrates, vitamin production, colonization resistance, mucosal immunity maturation, immune system induction, intestinal transit regulation, and formation of metabolic end products [74].
Because of competition among microbes and the rest of environmental stressors, microbes generate bioactive chemicals to combat these challenges. Such natural compounds are used as therapeutic agents for infections in animals and humans. Following the penicillin discovery, such organic products had a heyday from the 1940s through the 1960s. Although natural products were first discovered via screening of marine, soil, and other varied environments, the emphasis of natural product research has increasingly shifted to extracting these compounds from unculturable microorganisms [75]. Current research focuses on discovering novel bioactive chemicals, such as new antibiotics, via the activation of quiet biosynthetic pathways and the metagenomic study of various ecosystems [76]. It is necessary to find substitute natural compounds to fight pathogenic microorganisms resistant to antibiotic derivatives and familiar antibiotics. The microbiome of animals has a significant amount of untapped potential in the form of microorganisms that may produce peptides and secondary metabolites [55], and it is worth exploring further. In the human and animal microbiome, the identification of small peptides and biomolecules may herald the beginning of a new era in the development of antimicrobial drugs. Macromolecules are encoded via commensal microbes and are responsible for the microbe–microbe interactions and microbe-host relationships. In order to have a better understanding of such relationships and identify possible compounds of antimicrobial, macromolecules were observed on a large scale by the sequencing of the isolate’s genomes in different body places. To research the human microbiome, the collected Metagenomes and metatranscriptomes were examined by bioinformatics to get a better understanding of the presence of such compounds in healthy individuals. [77].
Various metabolic compounds produced by microbiota, these metabolites include oligosaccharides, terpenoids, lipids, glycolipids, ribosomally synthesized modified peptides of post-translationally (RiPPs), nonribosomal peptides, amino acids, and polyketides [78]. Such chemical diversity reveals different properties: antimicrobial, immunomodulatory, cytotoxic, and antioxidant, while the properties of several compounds have not been recognized [47]. In healthy people’s different body places, approximately 3118 biosynthetic gene clusters (BGCs) that encode macromolecules have been recognized from various samples acquired. Three classes of secondary metabolites, lantibiotics, thiopeptides, and thiozole/ oxazole-modified microcins, were discovered to be present in significant quantities at every site. Limited clusters of nonribosomal peptide sequence (NRPS) were discovered solely in gut microorganisms, and this was the first time this had been discovered. On most occasions, these BGCs differed from those found in microorganisms linked to nonhuman hosts. One BGC found in an oral microbiome was comparable to a macrolide found in a microbiome of a marine environment, which was surprising. Interestingly, one BGC from the metagenome encodes for a new thiopeptide that may be used as a therapeutic candidate [79]. Because in the microbiomes of animals and humans, several BGCs along unidentified functions occur, their identification and utilization for our benefit could aid in emerging new therapeutic agents for various infections and diseases [77].
4.6. Microbial Co-Cultivation (Mixed-Culture)
The presence of more than one type of microorganism in the same culture or medium is called mixed cultures, where neighboring microbes cooperate significantly [80]. Because of the significant number of fungal and bacterial species in such consortia, there is specialized communication among the microorganisms in these communities [81]. Natural products (SMs or secondary metabolites) are essential in communication [82]. Mixed cultures have been utilized in the production of beverages and food for millennia, and they have been employed in the production of solvents, industrial enzymes, and wastewater treatment for many years. Such a strategy’s benefits include lower prices, improved efficacy, and a greater variety of products [83]. In addition to interaction with the substrate of fermentable, Microbes interact with one another and the environment around them. It is believed that the interactions among microbes are complicated, but they are necessary to develop desirable product features [84]. Comparing the complex consortia of microbial mixed cultures to pure cultures, it may be concluded that they can perform more complicated activities and withstanding environmental changes and stresses [85].
In mixed culture, product yield and the rate of growth may be higher; one microbe may produce the required growth factors advantageous to the second microbe. Mixed cultures are capable of bringing about multistep biotransformations, allowing better consumption of the substrate. A broad spectrum of enzymes is present, allowing it to assault a wider variety of substances and facilitate the utilization of inexpensive and impure substrates [86]. The actual triggers causing the stimulation of natural product biosynthesis in such communities are different from the products themselves. They range from ecological signals, including pH, carbon, nitrogen, and phosphate sources, to different growth factors and cellular signaling molecules to organisms living in the same environment [87]. Competition for limited natural resources is supposed to be an optional force that stimulates the synthesis of biologically active compounds, which happens in mixed cultures only [88]. The outcomes of mixed cultures involve enhanced antibiotic activity in crude extracts, increased yields of previously defined metabolites, enhanced outcomes of unobserved metabolites, comparing to identify metabolites as a result of combined pathways, and unexpressed pathways stimulation for bioactive constituents [80].
The discovery of new bioactive compounds by unlocking SM genes contained in the microbes’ genomes is a common motivator for co-cultivation investigations. This has been demonstrated for various microbial pairings, including bacterium-bacterium, fungus-fungus, and bacterium-fungus. SMs produced in fungus-fungus co-cultures are the acremostatinsA-C, formed with a culture of Acremonium sp. mixed with Mycogone rosea [89]. In a co-cultivation with the Aspergillus oryzae or fungi Neurospora crassa, Watanabe et al. (1982) [90] explored the formation of the antibacterial polyketide enacyloxin with Gluconobacter sp. A recent antibiotic known as alchivemycin was isolated from the culture broth of the co-cultivation among Streptomyces endus and Tsukamurella pulmonis [91]. The pestalone production as a beneficial antibiotic against vancomycin-resistant Enterococcus faecium and methicillin-resistant Staphylococcus aureus (MRSA) was obtained in the co-culture of a marine-derived Gram-negative bacterium in the Pestalotia as the marine fungus and genus Thalassopia sp. [92]. In addition, communities of marine-based fungal-bacterial are found to be promising sources of new SMs; co-cultivation of the actinomycete Salinispora arenicola and a marine Emericella parvathecia caused a 100-fold production of emericellamides A and B with the fungus, metabolites exhibited antimicrobial activity against MRSA [93]. Co-cultivation of marine a-proteobacterium Thalassopia sp. and Libertella sp. led to the production of libertellenones A-D by the fungus, a direct contact seems to be essential for libertellone production, were neither produced in a Libertella monoculture nor by addition of extract or supernatant of the bacterial culture [88]. Co-cultures of coalmine drainage-derived organisms, an Aspergillus fumigatus strain, and a Sphingomonas strain, led to the discovery of glionitrin, which showed antimicrobial activity against Methicillin-resistant S. aureus [94]. As a result, mixed culture became a new opportunity for isolation and identification of novel antimicrobial drugs in both natural environments and under laboratory cultivations.
4.7. Alternative Targets for Antimicrobials Other Than Cellular Components
Most current antibiotics, even those that have undergone significant modification, target the same cellular processes as their natural or synthetic counterparts. The range of these targets is limited by the constituents of machinery translation, cell wall production, DNA/RNA metabolism, and a few other cellular activities. Due to the wide variety of genomes that have been sequenced, it is now possible to implement the concept of a magic bullet in a more complex manner, with crucial targets being determined with much greater specificity at the molecular level. This desire is complemented by the availability of various chemical compound collections to screen for the target/drug combination [95]. Comparing the metabolic processes of commensal and pathogenic bacteria, as well as medications that target the pathogenic features, may aid in the discovery of novel drug/target combinations in pathogens and, consequently, a novel paradigm of antimicrobial therapy that focuses on virulence [96]. Successful applications of this strategy have already been shown in the inhibition of the QseC-mediated activation of virulence gene expression in several pathogens [97], as well as the suppression of type III secretion system, an essential virulence component [98]. Drugs with a different original intent may end up being used as antimicrobials. For instance, BPH-652, a phosphonosulfonate, which was tested for cholesterol-lowering activity in humans as targeting the enzyme in the cholesterol biosynthesis pathway, squalene synthase, is also inhibiting an essential enzyme involved in Staphylococcus aureus virulence, dehydrosqualene synthase, and may therefore be thought of as a candidate drug to control MRSA [99]. Other areas of bacterial metabolism that may be targeted for modification include fatty acid biosynthesis [100], biosynthesis of aminoacyl-tRNAs [101], cell division [102], quorum sensing mechanism [103], two-component signal transduction system [104], and proton motive force [105].
The effectiveness of antibiotics can also be restored by focusing on the mechanisms of antibiotic resistance, such as efflux pumps or β -lactamases, which are compromised by the expanding resistance issue [106,107]. As it proceed along this path, it will be fundamentally changed from the previously established classical structural divides into antibiotic classes. We are only at the beginning of this, and not many antibacterial medications with novel modes of action have entered clinical trials yet. However, even at this stage, most of them do not belong to the previously identified antibiotic classes [108]. Additionally, developing new antibacterial treatments may be aided by intervention tactics that target biological networks rather than just targets [109]. For instance, combination therapy, which combines antibiotics with an antibiotic-enhancing phage, has shown promise as a viable antimicrobial intervention [110].
Conclusions
The discovery of antibiotics in human medicine has predominantly relied on natural products derived from microorganisms, supplemented by semi-synthetic and synthetic compounds. However, progress in antibiotic discovery declined after the 1960s due to challenges in cultivating microbes under traditional laboratory conditions. Novel approaches must be developed to reactivate natural antibacterial drug discovery. These include harnessing microbial metagenomes from unexplored sources, employing culture techniques mimicking natural microenvironments, and utilizing biosynthetic methods to generate diverse drug candidates. Additionally, exploring the gut microbiota of animals, modifying existing antimicrobial compounds, and identifying alternative drug targets offer promising avenues. These multifaceted strategies, coupled with advancements in purification and screening, hold the potential to accelerate the discovery of novel antibiotics and address the pressing issue of antibiotic resistance.
Author Contributions
Karokh Khdir: Writing original draft. Karzan Qurbani: Writing-original draft. Avin Sidiq: Writing reviewing. Heshu Rahman: Reviewing. Safin Hussein: Writing, review & editing. Sirwan Ahmed: Writing, review & editing. Shokhan Ahmed, Zhyar Hamagharib, Tara Muhamed, Rayan Mahmood, Sanarya Nadir, Gashbin Qadr, Hawnaz Najmaden, Sonia Rahim: Writing.
Funding
This study received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Data Availability Statement
All the data are available on the paper, and the listed references.
Acknowledgements
The authors are thankful to their respective institutions/universities.
Conflicts of Interest
All authors have no conflict of interest.
Code Availability
Not applicable.
Ethical Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
References
- Maartens MMJ, Swart CW, Pohl CH, Kock LJF. 2011. Antimicrobials, chemotherapeutics or antibiotics. Sci Res Essays. 6:3927–9.
- Seal, B.S.; Drider, D.; Oakley, B.B.; Brüssow, H.; Bikard, D.; Rich, J.O.; Miller, S.; Devillard, E.; Kwan, J.; Bertin, G.; et al. Microbial-derived products as potential new antimicrobials. Veter- Res. 2018, 49, 66. [CrossRef]
- Qurbani, K.; Hussein, S.; Ahmed, S.K. Recent meningitis outbreak in Iraq: a looming threat to public health. Ther. Adv. Infect. Dis. 2023, 10. [CrossRef]
- Dar KB, Bhat AH, Amin S, Zargar MA, Masood A, Malik AH, et al. 2017. Evaluation of antibacterial, antifungal and phytochemical screening of Solanum nigrum. Biochem Anal Biochem. 6(309):1009–2161.
- Qurbani, K.; Hussein, S.; Ahmed, S.K.; Darwesh, H.; Ali, S.; Hamzah, H. Biosafety and biosecurity in the Iraqi Kurdistan Region: Challenges and necessities. J. Biosaf. Biosecurity 2024, 6, 65–66. [CrossRef]
- Qurbani, K.; Hussein, S.; Hamzah, H.; Sulaiman, S.; Pirot, R.; Motevaseli, E.; Azizi, Z. Synthesis of Silver Nanoparticles by Raoultella Planticola and Their Potential Antibacterial Activity Against Multidrug-Resistant Isolates. 2022, 20, 75–83. [CrossRef]
- Hussein, S.; Sulaiman, S.; Ali, S.; Pirot, R.; Qurbani, K.; Hamzah, H.; Hassan, O.; Ismail, T.; Ahmed, S.K.; Azizi, Z. Synthesis of Silver Nanoparticles from Aeromonas caviae for Antibacterial Activity and In Vivo Effects in Rats. Biol. Trace Element Res. 2023, 202, 2764–2775. [CrossRef]
- Karch AM. 2013. 2014 Lippincott’s Pocket Drug Guide for Nurses. Lippincott Williams & Wilkins;
- Sherif, R.; Segal, B.H. Pulmonary aspergillosis: clinical presentation, diagnostic tests, management and complications. Curr. Opin. Pulm. Med. 2010, 16, 242–250. [CrossRef]
- Qurbani, K.; Ali, S.; Hussein, S.; Hamzah, H. Antibiotic resistance in Kurdistan, Iraq: A growing concern. New Microbes New Infect. 2024, 57, 101221. [CrossRef]
- Hussein, S.; Ahmed, S.K.; Qurbani, K.; Fareeq, A.; Essa, R.A. Infectious diseases threat amidst the war in Gaza. J. Med. Surg. Public Heal. 2024, 2. [CrossRef]
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Mohamed, M.G. Antimicrobial resistance: Impacts, challenges, and future prospects. J. Med. Surg. Public Heal. 2024, 2. [CrossRef]
- O’Neill J. 2016. Tackling drug-resistant infections globally: final report and recommendations.
- Qurbani KA, Amiri O, Othman GM, Fatah AA, Yunis NJ, Joshaghani M, et al. 2024. Enhanced antibacterial efficacy through piezo memorial effect of CaTiO3/TiO2 Nano-Composite. Inorg Chem Commun. 165:112470. [CrossRef]
- Aziz, D.M.; Hassan, S.A.; Amin, A.A.M.; Abdullah, M.N.; Qurbani, K.; Aziz, S.B. Author response for "A synergistic investigation of azo-thiazole derivatives incorporating thiazole moieties: a comprehensive exploration of their synthesis, characterization, computational insights, solvatochromism, and multimodal biological activity assessment". 2023. [CrossRef]
- Kader, D.A.; Aziz, D.M.; Mohammed, S.J.; Maarof, N.N.; Karim, W.O.; Mhamad, S.A.; Rashid, R.M.; Ayoob, M.M.; Kayani, K.F.; Qurbani, K. Green synthesis of ZnO/catechin nanocomposite: Comprehensive characterization, optical study, computational analysis, biological applications and molecular docking. Mater. Chem. Phys. 2024, 319. [CrossRef]
- Aziz D, Hassan SA, Mamand DM, Qurbani K. 2023. New azo-azomethine derivatives: Synthesis, characterization, computational, solvatochromic UV‒Vis absorption and antibacterial studies. J Mol Struct. 1284:135451.
- Bakhtyar R, Tofiq R, Hamzah H, Qurbani K. 2024. Fabricated Fusarium species-mediated nanoparticles against Gram-negative pathogen. World Acad Sci J. 7(1):1. [CrossRef]
- García-Salinas, S.; Elizondo-Castillo, H.; Arruebo, M.; Mendoza, G.; Irusta, S. Evaluation of the Antimicrobial Activity and Cytotoxicity of Different Components of Natural Origin Present in Essential Oils. Molecules 2018, 23, 1399. [CrossRef]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: history, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [CrossRef]
- Niu, G.; Li, W. Next-Generation Drug Discovery to Combat Antimicrobial Resistance. Trends Biochem. Sci. 2019, 44, 961–972. [CrossRef]
- Keyes K, Lee MD, Maurer JJ. 2003. Antibiotics: Mode of Action, Mechanisms of Resistance and Transfer. Microbial Food Safetry in Animal Agriculture Current Topics. ME Torrence and RE Isaacson, eds. Iowa State Press, Ames, USA;
- Bassett, E.J.; Keith, M.S.; Armelagos, G.J.; Martin, D.L.; Villanueva, A.R.; Keith Tetracycline-Labeled Human Bone from Ancient Sudanese Nubia (A.D. 350). Science 1980, 209, 1532–1534. [CrossRef]
- Playfair J. 2004. Living with Germs: In sickness and in health. Oxford University Press, USA;
- Schwartz, R.S. Paul Ehrlich’s Magic Bullets. New Engl. J. Med. 2004, 350, 1079–1080. [CrossRef]
- Marshall Jr EK. 1939. Bacterial chemotherapy: the pharmacology of sulfanilamide. Physiol Rev. 19(2):240–69.
- Fleming, A. On the Antibacterial Action of Cultures of a Penicillium, with Special Reference to their Use in the Isolation of B. influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236.
- Aminov, R. History of antimicrobial drug discovery: Major classes and health impact. 2016, 133, 4–19. [CrossRef]
- Waksman, S.A. Microbial Antagonisms and Antibiotic Substances. Soil Sci. 1947, 64, 433. [CrossRef]
- Greenwood, D. Microbiological properties of teicoplanin. J. Antimicrob. Chemother. 1988, 21, 1–13. [CrossRef]
- Lewis, K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013, 12, 371–387. [CrossRef]
- Falagas, M.E.; Skalidis, T.; Vardakas, K.Z.; Legakis, N.J.; on behalf of the Hellenic Cefiderocol Study Group Activity of cefiderocol (S-649266) against carbapenem-resistant Gram-negative bacteria collected from inpatients in Greek hospitals. J. Antimicrob. Chemother. 2017, 72, 1704–1708. [CrossRef]
- Boman, H.G. Antibacterial peptides: basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215. [CrossRef]
- Katz, L.; Baltz, R.H. Natural product discovery: past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The Roles of Antimicrobial Peptides in Innate Host Defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [CrossRef]
- Walsh, C.T.; Fischbach, M.A. Natural Products Version 2.0: Connecting Genes to Molecules. J. Am. Chem. Soc. 2010, 132, 2469–2493. [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [CrossRef]
- Strohl, W.R. The role of natural products in a modern drug discovery program. Drug Discov. Today 2000, 5, 39–41. [CrossRef]
- Daniel, R. The soil metagenome – a rich resource for the discovery of novel natural products. Curr. Opin. Biotechnol. 2004, 15, 199–204. [CrossRef]
- Kellenberger E. 2001. Exploring the unknown. EMBO Rep. 2(1):5–7.
- Connon, S.A.; Giovannoni, S.J. High-Throughput Methods for Culturing Microorganisms in Very-Low-Nutrient Media Yield Diverse New Marine Isolates. Appl. Environ. Microbiol. 2002, 68, 3878–85. [CrossRef]
- Zengler, K.; Toledo, G.; Rappé, M.; Elkins, J.; Mathur, E.J.; Short, J.M.; Keller, M. Cultivating the uncultured. Proc. Natl. Acad. Sci. 2002, 99, 15681–15686. [CrossRef]
- Williston, E.H.; Zia-Walrath, P.; Youmans, G.P. Plate Methods for Testing Antibiotic Activity of Actinomycetes against Virulent Human Type Tubercle Bacilli. J. Bacteriol. 1947, 54, 563–568. [CrossRef]
- Oliveira L de C, Silveira AMM, Monteiro A de S, Dos Santos VL, Nicoli JR, Azevedo VA de C, et al. 2017. In silico prediction, in vitro antibacterial spectrum, and physicochemical properties of a putative bacteriocin produced by Lactobacillus rhamnosus strain L156. 4. Front Microbiol. 8:876.
- Dubourg, G.; Elsawi, Z.; Raoult, D. Assessment of the in vitro antimicrobial activity of Lactobacillus species for identifying new potential antibiotics. Int. J. Antimicrob. Agents 2015, 46, 590–593. [CrossRef]
- Adnan, M.; Patel, M.; Hadi, S. Functional and health promoting inherent attributes ofEnterococcus hiraeF2 as a novel probiotic isolated from the digestive tract of the freshwater fishCatla catla. PeerJ 2017, 5, e3085. [CrossRef]
- Zipperer, A.; Konnerth, M.C.; Laux, C.; Berscheid, A.; Janek, D.; Weidenmaier, C.; Burian, M.; Schilling, N.A.; Slavetinsky, C.; Marschal, M.; et al. Human commensals producing a novel antibiotic impair pathogen colonization. Nature 2016, 535, 511–516. [CrossRef]
- Vijayakumar, P.P.; Muriana, P.M. A Microplate Growth Inhibition Assay for Screening Bacteriocins against Listeria monocytogenes to Differentiate Their Mode-of-Action. Biomolecules 2015, 5, 1178–1194. [CrossRef]
- Davies, J. How to discover new antibiotics: harvesting the parvome. Curr. Opin. Chem. Biol. 2011, 15, 5–10. [CrossRef]
- Saisho, Y.; Katsube, T.; White, S.; Fukase, H.; Shimada, J. Pharmacokinetics, Safety, and Tolerability of Cefiderocol, a Novel Siderophore Cephalosporin for Gram-Negative Bacteria, in Healthy Subjects. Antimicrob. Agents Chemother. 2018, 62. [CrossRef]
- Vickers NJ. 2017. Animal communication: when i’m calling you, will you answer too? Curr Biol. 27(14):R713–5.
- Wohlleben, W.; Mast, Y.; Stegmann, E.; Ziemert, N. Antibiotic drug discovery. Microb. Biotechnol. 2016, 9, 541–548. [CrossRef]
- Hosaka, T.; Ohnishi-Kameyama, M.; Muramatsu, H.; Murakami, K.; Tsurumi, Y.; Kodani, S.; Yoshida, M.; Fujie, A.; Ochi, K. Antibacterial discovery in actinomycetes strains with mutations in RNA polymerase or ribosomal protein S12. Nat. Biotechnol. 2009, 27, 462–464. [CrossRef]
- Yamanaka, K.; Reynolds, K.A.; Kersten, R.D.; Ryan, K.S.; Gonzalez, D.J.; Nizet, V.; Dorrestein, P.C.; Moore, B.S. Direct cloning and refactoring of a silent lipopeptide biosynthetic gene cluster yields the antibiotic taromycin A. Proc. Natl. Acad. Sci. 2014, 111, 1957–1962. [CrossRef]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [CrossRef]
- Spohn, M.; Kirchner, N.; Kulik, A.; Jochim, A.; Wolf, F.; Muenzer, P.; Borst, O.; Gross, H.; Wohlleben, W.; Stegmann, E. Overproduction of Ristomycin A by Activation of a Silent Gene Cluster in Amycolatopsis japonicum MG417-CF17. Antimicrob. Agents Chemother. 2014, 58, 6185–6196. [CrossRef]
- Abdelmohsen, U.R.; Grkovic, T.; Balasubramanian, S.; Kamel, M.S.; Quinn, R.J.; Hentschel, U. Elicitation of secondary metabolism in actinomycetes. Biotechnol. Adv. 2015, 33, 798–811. [CrossRef]
- Qurbani K, Hussein S, Ahmed SK. 2024. Molecular Biotechniques for the Isolation and Identification of Microorganisms.
- Charlop-Powers, Z.; Milshteyn, A.; Brady, S.F. Metagenomic small molecule discovery methods. Curr. Opin. Microbiol. 2014, 19, 70–75. [CrossRef]
- Qurbani, K.; Khdir, K.; Sidiq, A.; Hamzah, H.; Hussein, S.; Hamad, Z.; Abdulla, R.; Abdulla, B.; Azizi, Z. Aeromonas sobria as a potential candidate for bioremediation of heavy metal from contaminated environments. Sci. Rep. 2022, 12, 1–8. [CrossRef]
- Courtois, S.; Cappellano, C.M.; Ball, M.; Francou, F.-X.; Normand, P.; Helynck, G.; Martinez, A.; Kolvek, S.J.; Hopke, J.; Osburne, M.S.; et al. Recombinant Environmental Libraries Provide Access to Microbial Diversity for Drug Discovery from Natural Products. Appl. Environ. Microbiol. 2003, 69, 49–55. [CrossRef]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: a new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [CrossRef]
- Lloyd-Jones, G.; Hunter, D. Comparison of rapid DNA extraction methods applied to contrasting New Zealand soils. Soil Biol. Biochem. 2001, 33, 2053–2059. [CrossRef]
- Knietsch, A.; Waschkowitz, T.; Bowien, S.; Henne, A.; Daniel, R. Metagenomes of Complex Microbial Consortia Derived from Different Soils as Sources for Novel Genes Conferring Formation of Carbonyls from Short-Chain Polyols on Escherichia coli. Microb. Physiol. 2003, 5, 46–56. [CrossRef]
- Majernı́k A, Gottschalk G, Daniel R. 2001. Screening of environmental DNA libraries for the presence of genes conferring Na+ (Li+)/H+ antiporter activity on Escherichia coli: characterization of the recovered genes and the corresponding gene products. J Bacteriol. 183(22):6645–53.
- Gillespie, D.E.; Brady, S.F.; Bettermann, A.D.; Cianciotto, N.P.; Liles, M.R.; Rondon, M.R.; Clardy, J.; Goodman, R.M.; Handelsman, J. Isolation of Antibiotics Turbomycin A and B from a Metagenomic Library of Soil Microbial DNA. Appl. Environ. Microbiol. 2002, 68, 4301–4306. [CrossRef]
- Voget, S.; Leggewie, C.; Uesbeck, A.; Raasch, C.; Jaeger, K.-E.; Streit, W.R. Prospecting for Novel Biocatalysts in a Soil Metagenome. Appl. Environ. Microbiol. 2003, 69, 6235–6242. [CrossRef]
- Liles, M.R.; Manske, B.F.; Bintrim, S.B.; Handelsman, J.; Goodman, R.M. A Census of rRNA Genes and Linked Genomic Sequences within a Soil Metagenomic Library. Appl. Environ. Microbiol. 2003, 69, 2684–2691. [CrossRef]
- Knietsch, A.; Bowien, S.; Whited, G.; Gottschalk, G.; Daniel, R. Identification and Characterization of Coenzyme B 12 -Dependent Glycerol Dehydratase- and Diol Dehydratase-Encoding Genes from Metagenomic DNA Libraries Derived from Enrichment Cultures. Appl. Environ. Microbiol. 2003, 69, 3048–3060. [CrossRef]
- Quaiser, A.; Ochsenreiter, T.; Lanz, C.; Schuster, S.C.; Treusch, A.H.; Eck, J.; Schleper, C. Acidobacteria form a coherent but highly diverse group within the bacterial domain: evidence from environmental genomics. Mol. Microbiol. 2003, 50, 563–575. [CrossRef]
- Leser, T.D.; Mølbak, L. Better living through microbial action: the benefits of the mammalian gastrointestinal microbiota on the host. Environ. Microbiol. 2009, 11, 2194–2206. [CrossRef]
- Simpson, J.; Martineau, B.; Jones, W.; Ballam, J.; Mackie, R. Characterization of Fecal Bacterial Populations in Canines: Effects of Age, Breed and Dietary Fiber. Microb. Ecol. 2002, 44, 186–197. [CrossRef]
- Salminen S, von Wright A, Morelli L, Marteau P, Brassart D, de Vos WM, et al. 1998. Demonstration of safety of probiotics—a review. Int J Food Microbiol. 44(1–2):93–106.
- McFarland L V. 2000. Normal flora: diversity and functions. Microb Ecol Health Dis. 12(4):193–207.
- Lam, Y.C.; Crawford, J.M. Discovering antibiotics from the global microbiome. Nat. Microbiol. 2018, 3, 392–393. [CrossRef]
- Garcia-Gutierrez, E.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Gut microbiota as a source of novel antimicrobials. Gut Microbes 2018, 10, 1–21. [CrossRef]
- Suneja G, Nain S, Sharma R. 2019. Microbiome: A source of novel bioactive compounds and antimicrobial peptides. Microb Divers Ecosyst Sustain Biotechnol Appl Vol 1 Microb Divers Norm Extrem Environ. :615–30.
- Mousa, W.K.; Athar, B.; Merwin, N.J.; Magarvey, N.A. Antibiotics and specialized metabolites from the human microbiota. Nat. Prod. Rep. 2017, 34, 1302–1331. [CrossRef]
- Donia, M.S.; Cimermancic, P.; Schulze, C.J.; Wieland Brown, L.C.; Martin, J.; Mitreva, M.; Clardy, J.; Linington, R.G.; Fischbach, M.A. A Systematic Analysis of Biosynthetic Gene Clusters in the Human Microbiome Reveals a Common Family of Antibiotics. Cell 2014, 158, 1402–1414. [CrossRef]
- Pettit, R.K. Mixed fermentation for natural product drug discovery. Appl. Microbiol. Biotechnol. 2009, 83, 19–25. [CrossRef]
- Qurbani, K.; Hamzah, H. Intimate communication between Comamonas aquatica and Fusarium solani in remediation of heavy metal-polluted environments. Arch. Microbiol. 2020, 202, 1397–1406. [CrossRef]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299–299. [CrossRef]
- Salmon I, Bull AT. 1984. Mixed-culture fermentations in industrial microbiology.
- Smid, E.J.; Lacroix, C. Microbe–microbe interactions in mixed culture food fermentations. Curr. Opin. Biotechnol. 2013, 24, 148–154. [CrossRef]
- Brenner, K.; You, L.; Arnold, F.H. Engineering microbial consortia: a new frontier in synthetic biology. Trends Biotechnol. 2008, 26, 483–489. [CrossRef]
- Hesseltine CW. 1992. Mixed-culture fermentations. Appl Biotechnol Tradit fermented foods.
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2012, 11, 21–32. [CrossRef]
- Oh, D.-C.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Libertellenones A–D: Induction of cytotoxic diterpenoid biosynthesis by marine microbial competition. Bioorganic Med. Chem. 2005, 13, 5267–5273. [CrossRef]
- Degenkolb, T.; Heinze, S.; Schlegel, B.; Strobel, G.; Gräfe, U. Formation of New Lipoaminopeptides, Acremostatins A, B, and C, by Co-cultivation ofAcremoniumsp. Tbp-5 andMycogone roseaDSM 12973. Biosci. Biotechnol. Biochem. 2002, 66, 883–886. [CrossRef]
- Watanabe T, Izaki K, Takahashi H. 1982. New Polyenic Antibiotics Active Against Gram-Positive And-Negative Bacteria I. Isolation And Purification Of Antibiotics Produced By Gluconobacter sp. W-315. J Antibiot (Tokyo). 35(9):1141–7.
- Onaka, H.; Mori, Y.; Igarashi, Y.; Furumai, T. Mycolic Acid-Containing Bacteria Induce Natural-Product Biosynthesis in Streptomyces Species. Appl. Environ. Microbiol. 2011, 77, 400–406. [CrossRef]
- Cueto, M.; Jensen, P.R.; Kauffman, C.; Fenical, W.; Lobkovsky, E.; Clardy, J. Pestalone, a New Antibiotic Produced by a Marine Fungus in Response to Bacterial Challenge. J. Nat. Prod. 2001, 64, 1444–1446. [CrossRef]
- Marmann, A.; Aly, A.H.; Lin, W.; Wang, B.; Proksch, P. Co-Cultivation—A Powerful Emerging Tool for Enhancing the Chemical Diversity of Microorganisms. Mar. Drugs 2014, 12, 1043–1065. [CrossRef]
- Park HB, Kwon HC, Lee C-H, Yang HO. 2009. Glionitrin A, an antibiotic− antitumor metabolite derived from competitive interaction between abandoned mine microbes. J Nat Prod. 72(2):248–52.
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for bad bugs: confronting the challenges of antibacterial discovery. Nat. Rev. Drug Discov. 2006, 6, 29–40. [CrossRef]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: a new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [CrossRef]
- Rasko, D.A.; Moreira, C.G.; Li, D.R.; Reading, N.C.; Ritchie, J.M.; Waldor, M.K.; Williams, N.; Taussig, R.; Wei, S.; Roth, M.; et al. Targeting QseC Signaling and Virulence for Antibiotic Development. Science 2008, 321, 1078–1080. [CrossRef]
- Negrea, A.; Bjur, E.; Ygberg, S.E.; Elofsson, M.; Wolf-Watz, H.; Rhen, M. Salicylidene Acylhydrazides That Affect Type III Protein Secretion inSalmonella entericaSerovar Typhimurium. Antimicrob. Agents Chemother. 2007, 51, 2867–2876. [CrossRef]
- Liu, C.-I.; Liu, G.Y.; Song, Y.; Yin, F.; Hensler, M.E.; Jeng, W.-Y.; Nizet, V.; Wang, A.H.-J.; Oldfield, E. A Cholesterol Biosynthesis Inhibitor Blocks Staphylococcus aureus Virulence. Science 2008, 319, 1391–1394. [CrossRef]
- Su Z, Honek JF. 2007. Emerging bacterial enzyme targets. Curr Opin Investig drugs (London, Engl 2000). 8(2):140–9.
- Schimmel, P.; Tao, J.; Hill, J. Aminoacyl tRNA synthetases as targets for new anti-infectives. FASEB J. 1998, 12, 1599–1609. [CrossRef]
- Lock, R.L.; Harry, E.J. Cell-division inhibitors: new insights for future antibiotics. Nat. Rev. Drug Discov. 2008, 7, 324–338. [CrossRef]
- Njoroge, J.; Sperandio, V. Jamming bacterial communication: New approaches for the treatment of infectious diseases. EMBO Mol. Med. 2009, 1, 201–210. [CrossRef]
- Gotoh, Y.; Eguchi, Y.; Watanabe, T.; Okamoto, S.; Doi, A.; Utsumi, R. Two-component signal transduction as potential drug targets in pathogenic bacteria. 2010, 13, 232–239. [CrossRef]
- Diacon, A.H.; Pym, A.; Grobusch, M.; Patientia, R.; Rustomjee, R.; Page-Shipp, L.; Pistorius, C.; Krause, R.; Bogoshi, M.; Churchyard, G.; et al. The Diarylquinoline TMC207 for Multidrug-Resistant Tuberculosis. New Engl. J. Med. 2009, 360, 2397–2405. [CrossRef]
- Lomovskaya, O.; Bostian, K.A. Practical applications and feasibility of efflux pump inhibitors in the clinic—A vision for applied use. Biochem. Pharmacol. 2006, 71, 910–918. [CrossRef]
- Bush K, Macielag MJ. 2010. New β-lactam antibiotics and β-lactamase inhibitors. Expert Opin Ther Pat. 20(10):1277–93.
- Devasahayam, G.; Scheld, W.M.; Hoffman, P.S. Newer antibacterial drugs for a new century. Expert Opin. Investig. Drugs 2010, 19, 215–234. [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: from targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [CrossRef]
- Lu, T.K.; Collins, J.J. Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy. Proc. Natl. Acad. Sci. 2009, 106, 4629–4634. [CrossRef]
Figure 1.
Clarified Streaking and well diffusion methods in culture-dependent approach.
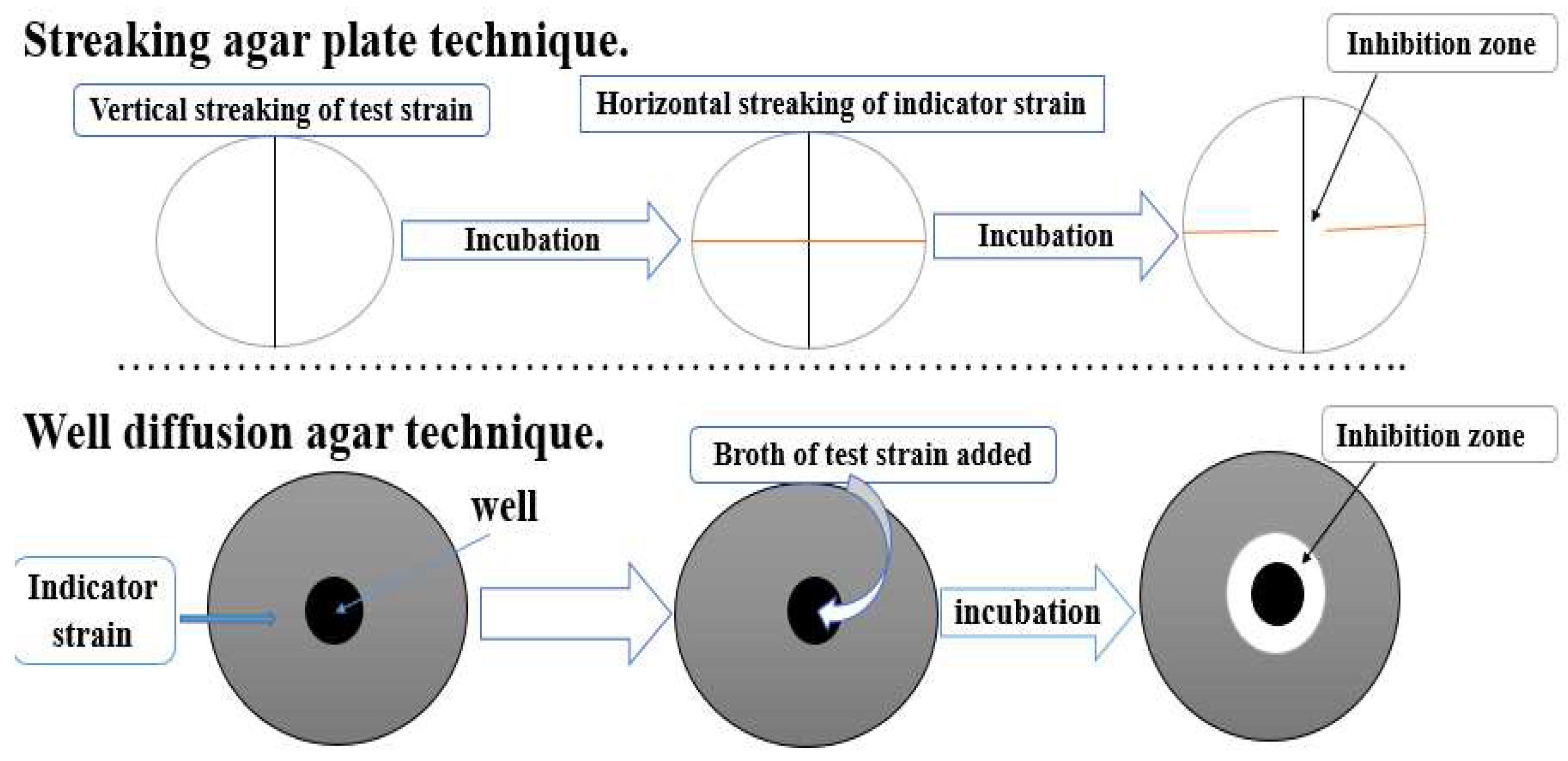
Figure 2.
shows the strategy of Metagenomics as the culture-independent approach to access the environmental DNA from different habitats.
Figure 2.
shows the strategy of Metagenomics as the culture-independent approach to access the environmental DNA from different habitats.

| Classes | Antibiotics | Years of Discovery | Sources |
|---|---|---|---|
| Β-Lactams | Penicillin Carbapenem Cephalosporin |
1928 1976 1948 |
Penicillium notatum, Penicillium chrysogenum Streptomyces cattleya Cephalosporium acremonium |
| Carboxylic acid | Mupirocin | 1971 | Pseudomonas fluorescens |
| Chloramphenicol | Chloramphenicol | 1946 | Streptomyces venezuelae |
|
Glycopeptides |
Teicoplanin Vancomycin |
1978 1953 |
Actinoplanes teichomyceticus Amycolatopsis orientalis |
| Aminoglycosides | Gentamicin Kanamycin Natamycin Neomycin Streptomycin |
1963 1957 1957 1949 1943 |
Micromonospora purpurea Streptomyces kanamyceticus Streptomyces natalensis Streptomyces fradiae Streptomyces griseus |
| Macrolides | Erythromycin Spiramycin Fidaxomicin |
1948 1952 1975 |
Streptomyces erythraeus Streptomyces ambofaciens Dactylosporangium aurantiacum |
| Lipopeptides: | Daptomycin | 1986 | Streptomyces roseosporus |
| Polypeptides: | Polymyxin | 1947 | Paenibacillus polymyxa |
| Rifamycins: | Rifampicin | 1957 | Streptomyces mediterranei |
| Sulfonamides: | Sulfamethoxazole | 1961 | Synthetic |
| Tetracyclines: | Chlortetracycline Tigecycline |
1948 1999 |
Streptomyces aureofaciens Synthetic |
| Nitroimidazoles: | Metronidazole | 1960 | Streptomyces spp. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



