
Submitted:
26 November 2024
Posted:
27 November 2024
Read the latest preprint version here
Abstract
This study examines the relationship between obesity, oxidative stress, and reproductive dysfunction. It focuses on the effects of sleeve gastrectomy on gene expression and hormone profiles in 29 women with extreme obesity (BMI ≥ 40 kg/m²). Pre- and post-surgical investigations revealed significant differences in major gene expressions and hormonal markers. CART expression reduced significantly from 0.27 ± 4.43 to -3.42 ± 1.14 (p < 0.001), while leptin expression decreased from -1.87 ± 1.75 to -0.13 ± 1.55 (p < 0.001), indicating better metabolic regulation. In contrast, eNOS expression increased considerably from -4.87 ± 1.70 to 1.18 ± 2.31 (p = 0.003), indicating improved endothelial function and nitric oxide bioavailability, which is critical for vascular health and reproduction.Correlation research before to surgery indicated no significant relationships between eNOS, CART, or leptin and clinical indicators, implying that these genes function independently in pre-surgical metabolism. While most associations remained negligible after surgery, a significant negative connection between eNOS expression and SHBG levels appeared (r = -0.365, p = 0.049), indicating potential interactions in hormonal regulation pathways following metabolic improvements.These findings emphasize the importance of bariatric surgery in reducing the negative effects of obesity on reproductive health by altering critical cellular pathways. Significant increases in CART, leptin, and eNOS expression indicate reduced oxidative stress, improved vascular tone, and hormonal balance, all of which contribute to increased reproductive capacity. This study sheds light on the molecular processes that link obesity, metabolic health, and fertility, underlining bariatric surgery's therapeutic potential for women experiencing obesity-related infertility.
Keywords:
infertility
; bariatric surgery
; CART peptide
; leptin
; ENOS
; oxidate stress
1. Introduction
Overweight and obesity have become more common in recent years as a result of dietary and lifestyle changes. In the United States, more than 30% of the population is currently considered obese [1,2]. The frequency varies worldwide, but in industrialized countries, a sizable proportion of the population is afflicted. In America, around one-third of people have a BMI exceeding 30 kg/m² [3]. Obesity is considered the second most important cause of death after smoking. According to Roland Sturm, healthcare costs associated to obesity outweigh those related to smoking [4]. Obesity is observed not only in adults but also in younger ages, because of excessive sugary beverage intake and prolonged screen time [5,6].
Obesity is a complicated metabolic condition that has a substantial influence on reproductive health, especially in women. The disorder is linked to endocrine disruption, chronic inflammation, and vascular dysfunction, which impairs ovulation and endometrial receptivity [7].
Although elevated leptin levels in obesity are supposed to lower food intake and increase energy expenditure, „leptin resistance” confuses this relationship. This resistance results from reduced leptin transport across the blood-brain barrier and altered intracellular signaling pathways, which are frequently exacerbated by chronic inflammation associated with obesity, emphasizing leptin’s complicated function in reproductive physiology [8].In obesity, leptin dysregulation impairs not only gonadotropin-releasing hormone (GnRH) secretion but also the production of fertility-related hormones, potentially leading to infertility [9]. Furthermore, leptin receptors found in ovarian tissue directly influence follicular growth [10]. At the molecular level, leptin has been found to activate the JAK-STAT and PI3K-Akt signaling pathways, which affect downstream targets such as the cocaine- and amphetamine-regulated transcript (CART) neuropeptide in the central nervous system. This connection points to a considerable interplay between leptin and CART in the control of energy balance and reproductive function [10].Obese individuals’ granulosa cells (GCs) have also been found to have elevated CART levels. Leptin signaling induces CART expression, which inhibits important enzymes such as aromatase via transcriptional repression mediated by intracellular signaling cascades such as the CREB and MAPK pathways. This inhibition decreases steroidogenesis, which further impairs ovarian function [11]. These molecular interactions highlight the complex role of leptin-CART crosstalk in linking metabolic disorders and reproductive health.
Endothelial nitric oxide synthase (eNOS) has a major role in obesity-induced reproductive difficulties because it produces nitric oxide (NO), which is a fundamental regulator of vascular tone and cellular signaling. NO is a cellular messenger and effector molecule known for its simple structure, fast diffusion, and short biological half-life. As an inorganic free radical, NO acts as a unique biological signal molecule, impacting a variety of physiological and pathological processes [12,13].
In the reproductive system, NO is necessary for follicular development, oocyte maturation, ovulation, and embryo implantation[14]. In men, it has an effect on spermatogenesis and sperm capacitation[15]. However, with obesity, increasing oxidative stress depletes the necessary cofactor tetrahydrobiopterin (BH4), causing eNOS”uncoupling,” resulting in the formation of superoxide (O2-) rather than NO [16]. This shift produces damaging oxidative byproducts such as peroxynitrite (ONOO-), which causes cellular damage and reduces NO bioavailability[17].
Elevated NO2 (nitrite) and NO3 (nitrate) levels indicate abnormal NO metabolism and decreased vascular function, affecting blood flow and cellular communication in the ovaries and uterus [18]. These molecular alterations have a negative impact on folliculogenesis, oocyte quality, and embryo implantation, and are associated with disorders such as polycystic ovarian syndrome (PCOS), which contributes to infertility [19].Bariatric surgery increases NO bioavailability by lowering oxidative stress and restoring eNOS activity, with benefits for vascular health and reproductive outcomes.
The present study investigates the relationship between obesity, oxidative stress and infertility through the expression of CART, Leptin and eNOS genes focusing on the therapeutic potential of bariatric surgery in restoring reproductive health.
2. Materials and Methods
The study included 29 women under 40 with a BMI of 40 kg/m² or above, according to the Helsinki Declaration ethical norms. The Medical School of Athens (Approval No. 48859, Date: 30 October 2020) and Alexandra General Hospital (Approval No. 4345, Date: 1 April 2023) provided the ethical approval. All participants gave their informed consent for this prospective study, which was realized from March 2022 to November 2024 at the First Department of Obstetrics and Gynecology of ‘Alexandra’ General Hospital. Infertility was defined as the failure to produce clinical pregnancy following a year of regular, unprotected sexual intercourse.
Participants were chosen based on their BMI and infertility diagnosis, and to ensure homogeneity, those with serious medical disorders, those on fertility-affecting drugs, and those over 40 were excluded. Consequentlyconfounding factors which might affect gene expression or reproductive outcome were excluded. Following a sleeve gastrectomy, patients were generally hospitalized for 1-2 days before beginning a gradual nutritional transition from liquids to solids. The rigorous inclusion criteria, ethical concerns, and comprehensive follow-up ensure that the study findings are legitimate. Figure 1 shows the study’s design and participant flow.
The flowchart in Figure 1. depicts the study’s systematic approach to participant recruiting, screening, and data collecting. Initially, a power analysis yielded a desired sample size of 32 individuals. During recruiting, 31 patients were recognized as eligible, however one chose not to proceed. As a result, 29 individuals were included in the trial, with two patients rejected due to particular criteria. The recruiting step includes a rigorous screening to verify that all participants satisfied the study’s eligibility criteria. Although the initial power analysis suggested 32 participants, the final sample size of 29 was considered enough to proceed with data collection and analysis. Following recruiting, the study collected and evaluated data from the remaining 29 individuals, leading to the study’s conclusion. The flowchart successfully demonstrates how the study handled participant exclusions and ensured adherence to criteria, while still maintaining a sample size close to the recommended target.
Hormone Measurements
Blood samples have been obtained from participants before and six months after sleeve gastrectomy. Hormonal measurements have been realized including follicle-stimulating hormone (FSH), luteinizing hormone (LH), estradiol (E2), sex hormone-binding globulin (SHBG), anti-Müllerian hormone (AMH), and free testosterone.
3. Detection of Leptin and Oxidative Stress Parameters Gene Expression
Total RNA was extracted from blood samples using New England Biolabs Monarch Total RNA Miniprep Kit (Ipswich, MA, USA). After extracting RNA, 1 μg was utilized to synthesize complementary DNA (cDNA) using New England BiolabsLunaScript RT SuperMix. To measure gene expression, 5 μL of produced cDNA was used in RT-PCR experiments.
All RT-PCR reactions were done using the LightCycler 480 Instrument II from Roche Life Sciences, using the Luna Universal qPCR Master Mix (New England Biolabs) at a final concentration of 1×. The RT-PCR technique consisted of an initial denaturation phase at 94 °C for one minute, followed by 40 cycles of denaturation at 95 °C for 15 seconds and annealing/extension at 60 °C for 30 seconds. To confirm data specificity, a melting curve study was conducted. To standardize gene expression levels, the internal housekeeping gene G6PD was used. Each experiment was duplicated to verify data accuracy and dependability. In addition, a negative control was used to account for any potential contamination or background signal. The relative mRNA expression levels of all genes were determined using the 2−ΔΔCT technique.
Statistical Analysis
Data were systematically recorded in Microsoft Excel (version 2401, 2016) spreadsheets (Microsoft Corporation, Redmond, Washington, DC, USA), with each row representing a single patient. Statistical analyses were performed with the SAS software platform for Windows (version 9.4, SAS Institute Inc., Cary, NC, USA). Descriptive data were reported as mean values with standard deviations (SD). To examine hormone level changes, the difference was computed as the post-surgical value minus the pre-surgical value, with a positive value indicating a rise and a negative value indicating a drop. The nonparametric Wilcoxon signed-rank test was used to compare the matched data.Additionally, the number and percentage of patients who had a rise or reduction in hormone levels were calculated. To investigate the association between changes in hormone levels, changes in body mass index (BMI), and gene expression levels, the Spearman correlation coefficient (rs) was obtained. A positive rs value near to 1 implies a high positive connection, whereas a negative rs value closer to -1 indicates a strong negative link. The significance level was set at p < 0.05, with all statistical tests being two-sided.
4. Sample Size Determination
A power analysis was performed to determine the optimum sample size for this investigation, guaranteeing adequate statistical power to detect significant differences in the important outcomes. To calculate the effect size for our research, we thoroughly evaluated the current literature and pertinent studies, as well as sought expert input. The two key outcome variables, ENOS, CART and leptin gene expression, were anticipated to have a modest impact size. This impact size was regarded as a conservative estimate that would allow for the detection of significant changes in gene expression.
We used a statistical power objective of 80% (1 - β = 0.80) to increase the likelihood of detecting actual effects. The significance threshold (α) was set at 0.05, representing a 5% chance of making a type I mistake. Paired t-tests were performed to assess the average expression levels of ENOS, CART and leptin genes before and six months after bariatric surgery. For non-parametric analysis of paired data, the Wilcoxon signed-rank test was used. In addition, based on comparable research, we assumed a modest correlation value of 0.5 to account for the link between pre- and post-surgery parameters.
The statistical program G*Power (version 3.1.9.7) was used to perform the power analysis with these parameters. The results showed that a total sample size of 29 individuals was necessary to obtain the appropriate statistical power. To account for probable attrition, an estimated sample size of 32 participants was advised, assuming a 10% loss to follow-up. Practical concerns such as patient availability and budget restrictions were also assessed to ensure the study’s viability in a clinical environment.
5. Results
Table 1 shows acorrelation analysis between eNOS and several clinical indicators was examined prior to bariatric surgery. Mean age was 33.03 years (SD = 4.08), with a Pearson correlation coefficient of 0.111 (p = 0.565). For hormonal parameters, FSH had a mean of 6.03 mIU/mL (SD = 1.09) and a correlation of 0.188 (p = 0.328), whereas LH had a mean of 6.87 mIU/mL (SD = 1.01) and an insignificant correlation of 0 (p = 0.997). Estradiol (E2) levels averaged 30.75 pg/mL (SD = 3.80), with a minor correlation of 0.249 (p = 0.193), whereas SHBG levels averaged 36.24 nmol/L (SD = 7.58), with a slight negative correlation of -0.076 (p = 0.696).Free testosterone exhibited the greatest variability (mean = 28.72 ng/dL, SD = 9.35) and a slight positive correlation of 0.285 (p = 0.133). AMH levels averaged 2.14 ng/mL (SD = 0.24), with a correlation coefficient of 0.115 (p = 0.552). For ovarian reserve markers, AFC in the right ovary (mean = 10.31, SD = 2.51) and left ovary (mean = 9.28, SD = 2.59) had associations of 0.095 (p = 0.623) and 0.063 (p = 0.746), respectively. Finally, the mean BMI before surgery was 41.12 (SD = 3.28), with a negative correlation of -0.266 (p = 0.162). Overall, none of these relationships was statistically significant (all p > 0.05), indicating that eNOS gene expression is not linearly linked with these clinical indicators prior to bariatric surgery.
Table 2 indicates the association of eNOS gene expression and several clinical indicators following bariatric surgery. The mean AFC in the left ovary after surgery was 6.76 (SD = 1.24), with a Pearson correlation of -0.149 (p = 0.439), and in the right ovary, 7.38 (SD = 1.64), with a correlation of 0.037 (p = 0.849). AMH levels after surgery averaged 2.96 ng/mL (SD = 0.15), with a correlation coefficient of 0.102 (p = 0.597). Six months after surgery, BMI averaged 25.98 (SD = 1.38) with a negative correlation of -0.248 (p = 0.194).Estradiol (E2) levels averaged 49.23 pg/mL (SD = 4.54) with a correlation of -0.156 (p = 0.42), whereas FSH levels post-surgery averaged 8.88 mIU/mL (SD = 0.08) with a weak correlation of 0.08 (p = 0.677). Free testosterone levels averaged 9.42 ng/dL (SD = 2.52) and showed a weak negative correlation of -0.151 (p = 0.436). LH levels after surgery averaged 8.83 mIU/mL (SD = 1.03), with a -0.121 correlation (p = 0.532). SHBG had the strongest connection (-0.365, p = 0.049), demonstrating a small but statistically significant negative link with Cpt eNOS. Overall, only SHBG shows a possible association with gene expression post surgery.
Table 3 shows the descriptive statistics and the correlation analysis of gene expression parameters prior to surgery. The mean expression of eNOS was -4.85 (SD = 1.75), with values ranging from -7.52 to -0.23, whereas CART had a mean of 0.27 (SD = 4.43) and ranged from -4.1 to 10.72, indicating greater variability. Leptin had a mean of -1.87 (SD = 1.75) with a range of -5.5 to 1.09.
Regarding Table 4, the connection between eNOS and CART (r = 0.038, p = 0.848), indicate no significant link. Similarly, the connection between Cpt eNOS A and Cpt0 Leptin was negative and modest (r = -0.301), but not statistically significant (p = 0.12).The correlation between Cpt0 CART and Cpt0 Leptin was equally minimal (r = 0.077, p = 0.697). Overall, none of the relationships were statistically significant (all p-values more than 0.05), indicating that these gene expressions are not linearly related prior to surgery.
Following surgery in Table 5, descriptive statistics and correlation analysis of gene expression characteristics reveal modest relationships between Cpt1eNOS, Cpt1 CART, and Cpt1 Leptin. The average expression of Cpt1eNOS was 1.18 (SD = 2.31), with values ranging from -2.07 to 6.77. Cpt1 CART had a mean of -3.4 (SD = 1.12) and ranged from -5.78 to -0.74, whereas Cpt1 Leptin had a mean of -0.07 (SD = 1.55) and ranged from -3.34 to 2.64.
Table 6 indicates the relationship between eNOS and CART was minimal (r = 0.16) and not statistically significant (p = 0.40), indicating no relevant relationship. Similarly, the correlation between Cpt1eNOS and Cpt1 Leptin was modest (r = 0.039) and had a high p-value (p = 0.838), indicating no meaningful relationship. Cpt1 CART and Cpt1 Leptin showed a weak positive connection (r = 0.282), however it was not statistically significant (p = 0.136). Overall, none of the correlations were statistically significant (all p-values > 0.05), indicating that there are no substantial linear connections between these gene expressions following surgery.
Table 7 summarizes the changes in gene expression for CART, Leptin, and eNOS before and after bariatric surgery, including mean values, standard deviations, correlations, and statistical significance. The mean expression of CART fell considerably from 0.273 (SD = 4.43) before surgery to -3.42 (SD = 1.14) after surgery, with a t-statistic of 4.52 and a highly significant p-value of < 0.001. Leptin expression decreased significantly from -1.87 (SD = 1.75) before surgery to -0.130 (SD = 1.55) after surgery, with a t-statistic of -7.35 and a p-value of < 0.001.Following surgery, eNOS expression increased from a mean of -4.87 (SD = 1.7) to 1.18 (SD = 2.31), with a moderate positive correlation (r = 0.529) and a statistically significant p-value (p = 0.003), indicating a substantial relationship in expression changes before and after surgery.
6. Discussion
Obesity treatment options include both nonsurgical and surgical procedures. Among surgical techniques, bariatric surgery stands out for its ability to improve metabolic health, lower cardiovascular risks, and resolve obesity-induced infertility[20,21,22]. This study focused on sleeve gastrectomy, a bariatric surgical technique that involves removing a major section of the stomach, leaving behind a smaller, tube-shaped stomach. This modification not only limits food intake, but it also causes hormonal changes that play an important role in hunger, metabolism, and energy balance. Sleeve gastrectomy has been found to contribute to significant weight loss and metabolic benefits by reducing the synthesis of hunger-stimulating hormones such as ghrelin and increasing the release of satiety-related hormones, making it a common treatment for obesity and associated comorbidities[23,24].Despite advances in ART, the impact of obesity on IVF outcomeremains a major source of concern.
Nitric oxide (NO) is involved in several reproductive processes in female animals, including follicular development, ovulation, oocyte maturation, and luteal function[14]. The enzyme nitric oxide synthase (NOS) produces NO, and its expression in ovarian follicles varies by species and stage of development[25].
This study focused on the connection between BMI changes after bariatric surgery, with hormone profiles, and genetic features in infertile women. Our findings showed considerable changes in gene expression following surgery, as well as poor correlations between gene expression and clinical findings.
Prior to surgery, correlation analysis of eNOS gene expression and clinical parameters revealed no statistically significant linear associations, as evidenced by non-significant p-values for all clinical variables, including age, BMI, and hormonal levels such as FSH, LH, and estradiol. This lack of association implies that, prior to surgical intervention, eNOS expression may function independently of these clinical parameters.After surgery, the correlation analysis of eNOS gene expression with clinical measurements revealed a similar trend, with the majority of associations being non-significant. However, sex hormone-binding globulin (SHBG), had a statistically significant negative connection with eNOS expression (r = -0.365, p = 0.049) showing that the corresponding protein may be associated with hormonal regulatory mechanisms involving SHBG following metabolic changes caused by bariatric surgery. Consequently,more research onto how SHBG and eNOS interactions affect the hormonal milieu following surgery could be of major interest.
Further analysis of gene expression levels of eNOS , CART, and Leptin prior to surgery revealed no significant relationships, with all p-values greater than 0.05 suggesting that the corresponding proteins may function independently.A similar trend emerged in the post-surgery correlations of the studied genes, with no significant linear associations. The lack of significant post-surgical connections among these genes shows that, while each of them may respond to physiological changes following bariatric surgery, there are no strong interdependencies in their expression patterns.
Conversely, eNOS expression increased significantly after surgery, from a mean of -4.87 (SD = 1.7) to 1.18 (SD = 2.31), with a moderate positive correlation between pre- and post-surgery levels (r = 0.529, p = 0.003). This rise in eNOS expression may indicate an improvement in endothelial function following surgery, which could be connected to improved vascular health and metabolic regulation. The increase of eNOS is noteworthy because it indicates a favorable adaptation in response to the lowered oxidative stress and inflammation that are commonly associated with obesity.The most surprising findings were from comparing CART, Leptin, and eNOS gene expression levels before and after surgery. Bariatric surgery significantly reduced the expression of CART and Leptin, with t-statistics of 4.52 and -7.35, respectively, and very significant p-values (both < 0.001). This significant drop shows that bariatric surgery may inhibit pathways involving CART and Leptin, most likely due to the significant changes in metabolic and inflammatory states associated with weight loss. Given that CART regulates hunger and Leptin regulates energy homeostasis, the observed decreases may be a response to the lower caloric intake and changed energy balance following surgery.
Adipose tissue is no longer thought of as a passive storage location for free fatty acids (FFA), but rather as a dynamic endocrine organ that secretes a variety of mediators known as adipokines. These proteins include TNF-α, resistin, IL-6, ASP, AGT, PAI-1, leptin, and adiponectin. They are involved in several metabolic processes, including energy balance, food intake, fat metabolism, vascular tone modulation, and insulin sensitivity [26,27]. Although leptin and adiponectin help regulate energy and metabolism, other variables such as TNF-α, IL-6, and resistin can worsen insulin resistance. Angiotensinogen and PAI-1 are linked to vascular problems associated with obesity [28].
Leptin, a 16-kDa hormone largely generated in adipose tissue, is critical in hunger control. Obesity has been associated to mutations in the leptin and leptin receptor genes [28,29]. In obesity, oxidative stress can be caused by a variety of factors, including hyperglycemia, hyperleptinemia, insufficient antioxidant defenses, increased lipid levels in muscle, increased muscle activity, elevated free radical production rates, mitochondrial dysfunction, endothelial impairment, and chronic inflammation[30,31].
CART expression is important in morbid obesity and is closely linked to hormonal pathways, as evidenced by animal research. Asnicar et al. studied CART-deficient mice and discovered that they were more prone to obesity when fed a high-fat diet [23]. Similarly, Lee et al. found that injecting CART into the nucleus tractus solitarii reduced food consumption in obese rats [32]. Kristensen et al. demonstrated that CART peptides inhibited feeding behavior even in the presence of neuropeptide Y (NPY), a powerful appetite stimulant in the central nervous system[10].Furthermore, Okumura et al. and Asakawa et al. found that CART injection into the fourth ventricle delayed stomach emptying, similar to the effects of corticotropin-releasing factor (CRF) and cholecystokinin [33,34]. Patkar et al. investigated the link between bariatric surgery and CART expression in a mouse model of morbid obesity. Mice were separated into four groups based on the intervention: Roux-en-Y gastric bypass (RYGB), sleeve gastrectomy, calorie restriction, and a control group receiving no intervention. The study found no statistically significant differences in CART mRNA expression among the groups. Furthermore, the findings indicated that RYGB had distinct effects on hypothalamic gene regulation in comparison to calorie restriction or weight loss alone [35].Grayson et al. and Cavin et al. found no significant changes in CART expression following bariatric surgery in obese rats, which supports these findings [36,37]. Patkar et al. confirmed these findings, revealing no quantifiable changes in CART levels before and after RYGB, nor any variations in CART mRNA expression between chow-fed and RYGB-treated mice [35]. Muñoz-Rodríguez et al. found that obese patients had higher CART levels before bariatric surgery than those with normal weight, but there was no significant difference one year later [38]. In their 2021 assessment, Singh et al. suggested that the CART peptide has potential as a treatment agent for morbid obesity [39].
In cattle, eNOS is present in granulosa cells at all stages of follicular development[40].In porcine, eNOS is expressed in big follicles (7-10 mm), but not in medium-sized follicles (3-6 mm)[41]. These findings highlight the species-specific expression patterns of NOS and point to a distinct functional role of NO in ovarian physiology.
NO also regulates the growth and development of preantral follicles. NO levels vary throughout follicular development, indicating that it influences follicular expansion and atresia [42]. In preantral follicle investigations, enhanced eNOS expression was detected in response to gonadotropins such as human chorionic gonadotropin (hCG), emphasizing its role in follicular development and differentiation [43,44]. The dual role of NO in follicular development may be due to its vasodilatory impact, which influences follicular blood flow and nutrientdelivery[45].
Experiments involving mice treated with NOS inhibitors or eNOS mutant animals demonstrate that NO plays a function in oocyte maturation [46]. These oocyte meiosis defects indicate that the cumulus-oocyte complex development is dependent on the NO/NOS system [47]. Furthermore, NO has a concentration-dependent effect, with low levels encouraging oocyte maturation and high levels inhibiting it via decreasing estradiol (E2) production [48]. This underscores the need of precisely regulating NO levels in order to ensure optimum oocyte maturation.
During ovulation, NO is involved in vascular remodeling and follicular rupture. Non-specific NOS inhibitors, such as aminoguanidine (AG) and L-NAME, have been demonstrated in studies to drastically lower ovulation rates, which may be reversed by NO donors such sodium nitroprusside (SNP) [49]. The control of NO during ovulation involves complicated hormonal interactions, such as increasing luteinizing hormone (LH) production to stimulate preovulatory processes [50]. Furthermore, NO has been demonstrated to influence the synthesis of E2, which governs ovulation and follicular development.
NO also regulates follicular atresia by inhibiting granulosa cell death. It has been shown that iNOS-derived NO inhibits apoptosis in rat granulosa cells via autocrine and paracrine signaling, preventing early follicular atresia [40,51]. High NO levels, on the other hand, can induce apoptosis in well-differentiated granulosa cells, demonstrating a dual regulatory role dependent on concentration and follicular stage [52]. NO regulates apoptosis through mechanisms such as the Fas/FasL system and the modulation of pro- and anti-apoptotic gene expression[53].
In fertilization, NO regulates oocyte activation by affecting intracellular calcium (Ca2+) signaling and sperm capacitation [54]. NO’s role in these activities is critical for proper fertilization and early embryo development. Furthermore, NO regulates embryo implantation and pregnancy maintenance by creating a favorable uterine environment, as well as modulating uterine contractility and placental blood flow [55,56]. NO also helps to maintain vascular resistance and promote nutrient exchange, both of which are necessary for fetal growth and development [57].
Although an association between eNOS, CART and Leptin are not established, they should be considered as putative biomarkers for fertility outcome in obese women. Additionally it could be of great interest, to further examine a possible parallel action of miRNAs, regulating the expression of the aforementioned genes. Both regulation of oxidative stress and of metabolic disorders through the creation of a related to them genetic profile may be of great value to fertility reestablishment after bariatric surgery.
7. Conclusions
Bariatric surgery improves reproductive health significantly by restoring nitric oxide (NO) generation, decreasing oxidative stress, and increasing egg quality and endometrial receptivity. The nitrate-to-nitrite-to-nitric oxide (NO3-NO2-NO) pathway is critical for maintaining NO bioavailability, especially during oxidative stress and low oxygen levels. Bariatric surgery improves reproductive results for women with severe obesity by boosting NO generation, supporting mitochondrial function, and restoring hormonal balance. It also regulates hormone output, which promotes normal ovulation and menstrual cycles. Further research is needed to examine the long-term reproductive implications of bariatric surgery and to further understand the particular biochemical processes by which NO contributes in fertility restoration.
Author Contributions
We confirm that each author made a written contribution to the work. Data curation, investigation, methodology, and writing review and editing: C.V., Resources and project administration: D.M. and R.S. Software: K.B. and M.D. Formal analysis: A.K. and A.P., Investigations: V.T. and A.V., Resources: R.M. and P.K., Analysis: D.A. and D.M., Project administration: P.A., Software: C.V. and D.L., Supervision and validation: G.D. and D.L. All authors have readand agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was ethically approved by the scientific boards of the Medical School of Athens (48859, October 30, 2020) and Alexandra General Hospital (4345, March 1, 2023), following the Declaration of Helsinki guidelines.
Informed Consent Statement
All participants in the research provided informed consent.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to all of the women who took part in this study, believing in usand enabling us to finish this project in this manner.
Conflicts of Interest
The authors have declared no conflicts of interest.
References
- Leeners B, Geary N, Tobler PN, Asarian L. Ovarian hormones and obesity. Human Reproduction Update [Internet]. 2017 May 1 [cited 2024 Oct 23];23(3):300–21. Available from: https://academic.oup.com/humupd/article/23/3/300/3058798.
- Pearce EN. Thyroid hormone and obesity. Current Opinion in Endocrinology, Diabetes & Obesity [Internet]. 2012 Oct [cited 2024 Oct 23];19(5):408–13. Available from: https://journals.lww.com/01266029-201210000-00014.
- Zhang Y, Guo X, Guo L, Chang H, Shu J, Leung PCK. Outcomes comparison of IVF/ICSI among different trigger methods for final oocyte maturation: A systematic review and meta-analysis. The FASEB Journal [Internet]. 2021 Jul [cited 2023 Jul 6];35(7). [CrossRef]
- Sturm R, An R, Maroba J, Patel D. The effects of obesity, smoking, and excessive alcohol intake on healthcare expenditure in a comprehensive medical scheme. S Afr Med J [Internet]. 2013 Sep 30 [cited 2024 Oct 23];103(11):840. Available from: http://www.samj.org.za/index.php/samj/article/view/7260.
- Ludwig DS, Peterson KE, Gortmaker SL. Relation between consumption of sugar-sweetened drinks and childhood obesity: a prospective, observational analysis. The Lancet [Internet]. 2001 Feb [cited 2024 Oct 23];357(9255):505–8. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0140673600040411.
- Robinson TN. TELEVISION VIEWING AND CHILDHOOD OBESITY. Pediatric Clinics of North America [Internet]. 2001 Aug [cited 2024 Oct 23];48(4):1017–25. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0031395505703540.
- Moran LJ, Dodd J, Nisenblat V, Norman RJ. Obesity and Reproductive Dysfunction in Women. Endocrinology and Metabolism Clinics of North America [Internet]. 2011 Dec [cited 2023 Jul 9];40(4):895–906. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0889852911000715.
- Zhang Y, Scarpace PJ. The role of leptin in leptin resistance and obesity. Physiology & Behavior [Internet]. 2006 Jun [cited 2023 Sep 14];88(3):249–56. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0031938406002447. [CrossRef]
- Anifandis G, Koutselini E, Louridas K, Liakopoulos V, Leivaditis K, Mantzavinos T, et al. Estradiol and leptin as conditional prognostic IVF markers. Reproduction [Internet]. 2005 Apr [cited 2023 Jun 19];129(4):531–4. Available from: https://rep.bioscientifica.com/view/journals/rep/129/4/1290531.xml. [CrossRef]
- Kristensen P, Judge ME, Thim L, Ribel U, ChristjansenKN, Wulff BS, et al. Hypothalamic CART is a new anorectic peptide regulated by leptin. Nature [Internet]. 1998 May [cited 2023 Jun 20];393(6680):72–6. Available from: https://www.nature.com/articles/29993. [CrossRef]
- Ma X, Hayes E, Prizant H, Srivastava RK, Hammes SR, Sen A. Leptin-Induced CART (Cocaine- and Amphetamine-Regulated Transcript) Is a Novel Intraovarian Mediator of Obesity-Related Infertility in Females. Endocrinology [Internet]. 2016 Mar 1 [cited 2023 Jun 20];157(3):1248–57. Available from: https://academic.oup.com/endo/article/157/3/1248/2422918.
- Tran N, Garcia T, Aniqa M, Ali S, Ally A, Nauli SM. Endothelial Nitric Oxide Synthase (eNOS) and the Cardiovascular System: in Physiology and in Disease States. Am J Biomed Sci Res. 2022;15(2):153–77.
- Man AWC, Zhou Y, Xia N, Li H. Endothelial Nitric Oxide Synthase in the Perivascular Adipose Tissue. Biomedicines. 2022 Jul 21;10(7):1754.
- Luo Y, Zhu Y, Basang W, Wang X, Li C, Zhou X. Roles of Nitric Oxide in the Regulation of Reproduction: A Review. Front Endocrinol [Internet]. 2021 Nov 19 [cited 2024 Sep 23];12:752410. [CrossRef]
- Amiri I, Sheikh N, Najafi R. Nitric oxide level in seminal plasma and its relation with sperm DNA damages. Iran Biomed J. 2007 Oct;11(4):259–64.
- Le Lay S, Simard G, Martinez MC, Andriantsitohaina R. Oxidative Stress and Metabolic Pathologies: From an Adipocentric Point of View. Oxidative Medicine and Cellular Longevity [Internet]. 2014 [cited 2024 Nov 1];2014:1–18. Available from: http://www.hindawi.com/journals/omcl/2014/908539/.
- Jomova K, Raptova R, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, et al. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol [Internet]. 2023 Oct [cited 2024 Nov 1];97(10):2499–574. [CrossRef]
- Barbosa PO, Tanus-Santos JE, Cavalli RDC, Bengtsson T, Montenegro MF, Sandrim VC. The Nitrate-Nitrite-Nitric Oxide Pathway: Potential Role in Mitigating Oxidative Stress in Hypertensive Disorders of Pregnancy. Nutrients [Internet]. 2024 May 14 [cited 2024 Nov 1];16(10):1475. Available from: https://www.mdpi.com/2072-6643/16/10/1475. [CrossRef]
- Kicińska AM, Maksym RB, Zabielska-Kaczorowska MA, Stachowska A, Babińska A. Immunological and Metabolic Causes of Infertility in Polycystic Ovary Syndrome. Biomedicines [Internet]. 2023 May 28 [cited 2024 Nov 1];11(6):1567. Available from: https://www.mdpi.com/2227-9059/11/6/1567. [CrossRef]
- Courcoulas AP. Weight Change and Health Outcomes at 3 Years After Bariatric Surgery Among Individuals With Severe Obesity. JAMA [Internet]. 2013 Nov 4 [cited 2024 Apr 28]. [CrossRef]
- Leitner DR, Frühbeck G, Yumuk V, Schindler K, Micic D, Woodward E, et al. Obesity and Type 2 Diabetes: Two Diseases with a Need for Combined Treatment Strategies - EASO Can Lead the Way. Obes Facts. 2017;10(5):483–92.
- Hannah WN, Harrison SA. Effect of Weight Loss, Diet, Exercise, and Bariatric Surgery on Nonalcoholic Fatty Liver Disease. Clin Liver Dis. 2016 Dec;20(2):339–50.
- Asnicar MA, Smith DP, Yang DD, Heiman ML, Fox N, Chen YF, et al. Absence of cocaine- and amphetamine-regulated transcript results in obesity in mice fed a high caloric diet. Endocrinology. 2001 Oct;142(10):4394–400. [CrossRef]
- Gumbs AA, Gagner M, Dakin G, Pomp A. Sleeve Gastrectomy for Morbid Obesity. OBES SURG [Internet]. 2007 Oct 30 [cited 2024 Jan 17];17(7):962–9. [CrossRef]
- Zhang W, Wei Q wei, Wang Z chao, Ding W, Wang W, Shi F xiong. Cell-specific expression and immunolocalization of nitric oxide synthase isoforms and the related nitric oxide/cyclic GMP signaling pathway in the ovaries of neonatal and immature rats. J Zhejiang Univ Sci B [Internet]. 2011 Jan [cited 2024 Nov 1];12(1):55–64. [CrossRef]
- Rajala MW, Scherer PE. Minireview: The Adipocyte—At the Crossroads of Energy Homeostasis, Inflammation, and Atherosclerosis. Endocrinology [Internet]. 2003 Sep 1 [cited 2024 Oct 23];144(9):3765–73. Available from: https://academic.oup.com/endo/article/144/9/3765/2502070.
- Ferroni P, Basili S, Falco A, Davì G. Oxidant Stress and Platelet Activation in Hypercholesterolemia. Antioxidants & Redox Signaling [Internet]. 2004 Aug [cited 2024 Oct 23];6(4):747–56. [CrossRef]
- Leibel RL. The Role of Leptin in the Control of Body Weight. Nutrition Reviews [Internet]. 2002 Oct 1 [cited 2024 Oct 23];60(suppl_10):S15–9. Available from: https://academic.oup.com/nutritionreviews/article/60/suppl_10/S15/1897536.
- ElmquistJK. Hypothalamic pathways underlying the endocrine, autonomic, and behavioral effects of leptin. Physiology & Behavior [Internet]. 2001 Nov [cited 2024 Oct 23];74(4–5):703–8. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0031938401006138.
- Manna P, Jain SK. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metabolic Syndrome and Related Disorders [Internet]. 2015 Dec [cited 2024 Nov 1];13(10):423–44. [CrossRef]
- Di Domenico M, Pinto F, Quagliuolo L, Contaldo M, Settembre G, Romano A, et al. The Role of Oxidative Stress and Hormones in Controlling Obesity. Front Endocrinol [Internet]. 2019 Aug 13 [cited 2024 Nov 1];10:540. [CrossRef]
- Lee SJ, Krieger JP, Vergara M, Quinn D, McDougle M, De Araujo A, et al. Blunted Vagal Cocaine- and Amphetamine-Regulated Transcript Promotes Hyperphagia and Weight Gain. Cell Reports [Internet]. 2020 Feb [cited 2023 Dec 9];30(6):2028-2039.e4. Available from: https://linkinghub.elsevier.com/retrieve/pii/S2211124720300693. [CrossRef]
- Okumura T, Yamada H, Motomura W, Kohgo Y. Cocaine-amphetamine-regulated transcript (CART) acts in the central nervous system to inhibit gastric acid secretion via brain corticotropin-releasing factor system. Endocrinology. 2000 Aug;141(8):2854–60.
- Asakawa A, Inui A, Yuzuriha H, Nagata T, Kaga T, Ueno N, et al. Cocaine-Amphetamine-Regulated Transcript Influences Energy Metabolism, Anxiety and Gastric Emptying in Mice. HormMetab Res [Internet]. 2001 Sep [cited 2023 Jun 20];33(9):554–8. [CrossRef]
- Patkar PP, Hao Z, Mumphrey MB, Townsend RL, Berthoud HR, Shin AC. Unlike calorie restriction, Roux-en-Y gastric bypass surgery does not increase hypothalamic AgRP and NPY in mice on a high-fat diet. Int J Obes (Lond). 2019 Nov;43(11):2143–50.
- Grayson BE, Hakala-Finch AP, Kekulawala M, Laub H, Egan AE, Ressler IB, et al. Weight loss by calorie restriction versus bariatric surgery differentially regulates the hypothalamo-pituitary-adrenocortical axis in male rats. Stress. 2014 Dec;17(6):484–93. [CrossRef]
- Cavin JB, Voitellier E, Cluzeaud F, Kapel N, Marmuse JP, Chevallier JM, et al. Malabsorption and intestinal adaptation after one anastomosis gastric bypass compared with Roux-en-Y gastric bypass in rats. Am J PhysiolGastrointest Liver Physiol. 2016 Sep 1;311(3):G492-500. [CrossRef]
- Muñoz-Rodríguez JR, Agarrado A, Martín-Fernández J, Salas E, González-Martín C, Alguacil LF. Cocaine and amphetamine regulated transcript and brain-derived neurotrophic factor in morbid obesity. One-year follow-up after gastric bypass. Surg ObesRelat Dis. 2018 Nov;14(11):1732–9.
- Singh A, De Araujo AM, Krieger JP, Vergara M, Ip CK, De Lartigue G. Demystifying functional role of cocaine- and amphetamine-related transcript (CART) peptide in control of energy homeostasis: A twenty-five year expedition. Peptides [Internet]. 2021 Jun [cited 2023 Dec 9];140:170534. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0196978121000425.
- Matsumi H, Yano T, Osuga Y, Kugu K, Tang X, Xu JP, et al. Regulation of Nitric Oxide Synthase to Promote Cytostasis in Ovarian Follicular Development1. Biology of Reproduction [Internet]. 2000 Jul 1 [cited 2024 Oct 23];63(1):141–6. [CrossRef]
- Takesue K, Tabata S, Sato F, Hattori M aki. Expression of Nitric Oxide Synthase-3 in Porcine Oocytes Obtained at Different Follicular Development. J Reprod Dev [Internet]. 2003 [cited 2024 Nov 1];49(2):135–40. Available from: http://www.jstage.jst.go.jp/article/jrd/49/2/49_2_135/_article. [CrossRef]
- Rosselli M. Role of nitric oxide in the biology, physiology and pathophysiology of reproduction. Human Reproduction Update [Internet]. 1998 Jan 1 [cited 2024 Oct 23];4(1):3–24. [CrossRef]
- Pires PRL, Santos NP, Adona PR, Natori MM, Schwarz KRL, De Bem THC, et al. Endothelial and inducible nitric oxide synthases in oocytes of cattle. Animal Reproduction Science [Internet]. 2009 Dec [cited 2024 Oct 23];116(3–4):233–43. Available from: https://linkinghub.elsevier.com/retrieve/pii/S037843200900058X.
- Mitchell LM. Expression of nitric oxide synthase and effect of substrate manipulation of the nitric oxide pathway in mouse ovarian follicles. Human Reproduction [Internet]. 2004 Jan 1 [cited 2024 Oct 23];19(1):30–40. [CrossRef]
- Ferrari AU, Radaelli A, Mori I, Mircoli L, Perlini S, Meregalli P, et al. Nitric Oxide-Dependent Vasodilation and the Regulation of Arterial Blood Pressure: Journal of Cardiovascular Pharmacology [Internet]. 2001 Nov [cited 2024 Oct 23];38:S19–22. Available from: http://journals.lww.com/00005344-200111002-00006.
- Jablonka-Shariff A, Olson LM. Hormonal Regulation of Nitric Oxide Synthases and Their Cell-Specific Expression during Follicular Development in the Rat Ovary 1. Endocrinology [Internet]. 1997 Jan [cited 2024 Oct 23];138(1):460–8. [CrossRef]
- Sengoku K, Takuma N, Horikawa M, Tsuchiya K, Komori H, Sharifa D, et al. Requirement of nitric oxide for murine oocyte maturation, embryo development, and trophoblast outgrowth in vitro. Mol Reprod Dev [Internet]. 2001 Mar [cited 2024 Oct 23];58(3):262–8. [CrossRef]
- Tao Y, Xie H, Hong H, Chen X, Jang J, Xia G. Effects of nitric oxide synthase inhibitors on porcine oocyte meiotic maturation. Zygote [Internet]. 2005 Feb [cited 2024 Oct 23];13(1):1–9. Available from: https://www.cambridge.org/core/product/identifier/S0967199404002953/type/journal_article. [CrossRef]
- Preti SC, Da Cunha V, Vassallo DV, Stefanon I. The Superoxide Dismutase Mimetic, Tempol, Reduces the Bioavailability of Nitric Oxide and does not Alter L-NAME-Induced Hypertension in Rats. Basic Clin Pharma Tox [Internet]. 2005 Jul [cited 2024 Oct 23];97(1):29–34. [CrossRef]
- McCosh RB, Lopez JA, Szeligo BM, Bedenbaugh MN, Hileman SM, Coolen LM, et al. Evidence that Nitric Oxide Is Critical for LH Surge Generation in Female Sheep. Endocrinology [Internet]. 2020 Mar 1 [cited 2024 Oct 23];161(3):bqaa010. [CrossRef]
- Jiang X, Dias JA, He X. Structural biology of glycoprotein hormones and their receptors: Insights to signaling. Molecular and Cellular Endocrinology [Internet]. 2014 Jan [cited 2024 Apr 28];382(1):424–51. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0303720713003584.
- Basini G, Grasselli F, Ponderato N, Bussolati S, Tamanini C. Lipid hydroperoxide and cGMP are not involved in nitric oxide inhibition of steroidogenesis in bovine granulosa cells. ReprodFertil Dev [Internet]. 2000 [cited 2024 Oct 23];12(6):289. Available from: http://www.publish.csiro.au/?paper=RD00089.
- Chen Q, Yano T, Matsumi H, Osuga Y, Yano N, Xu J, et al. Cross-Talk between Fas/Fas Ligand System and Nitric Oxide in the Pathway Subserving Granulosa Cell Apoptosis: A Possible Regulatory Mechanism for Ovarian Follicle Atresia. Endocrinology [Internet]. 2005 Feb [cited 2024 Oct 23];146(2):808–15. [CrossRef]
- Francavilla F, Santucci R, Macerola B, Ruvolo G, Romano R. Nitric Oxide Synthase Inhibition in Human Sperm Affects Sperm-Oocyte Fusion but Not Zona Pellucida Binding1. Biology of Reproduction [Internet]. 2000 Aug 1 [cited 2024 Oct 23];63(2):425–9. [CrossRef]
- Gouge RC, Marshburn P, Gordon BE, Nunley W, Huet-Hudson YM. Nitric Oxide as a Regulator of Embryonic Development. Biology of Reproduction [Internet]. 1998 Apr 1 [cited 2024 Oct 23];58(4):875–9. Available from: https://academic.oup.com/biolreprod/article/2761074/Nitric. [CrossRef]
- Novaro V, González E, Jawerbaum A, Rettori V, Canteros G, Gimeno MF. Nitric oxide synthase regulation during embryonic implantation. ReprodFertil Dev [Internet]. 1997 [cited 2024 Oct 23];9(5):557. Available from: http://www.publish.csiro.au/?paper=R97005. [CrossRef]
- Zullino S, Buzzella F, Simoncini T. Nitric oxide and the biology of pregnancy. Vascular Pharmacology [Internet]. 2018 Nov [cited 2024 Oct 23];110:71–4. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1537189118302763. [CrossRef]
Figure 1.
Flow chart of patients’ elimination and inclusion in the study.
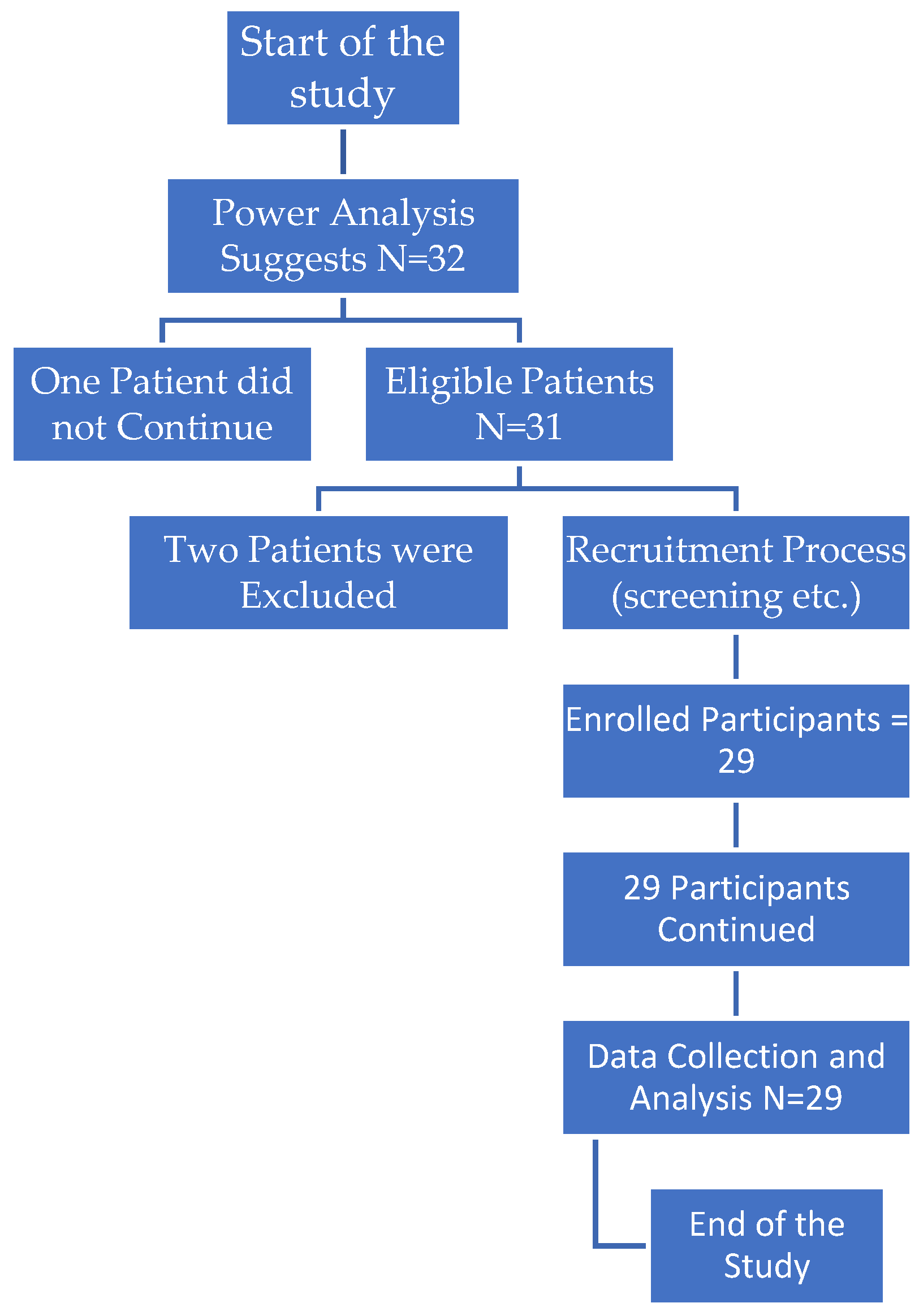

Table 1.
Correlation Analysis of eNOS expression with Clinical Parameters Before Bariatric Surgery.
| Variable | Mean | Standard Deviation | PearsonCorrelation | P-value |
|---|---|---|---|---|
| Age | 33.034 | 4.075 | 0.111 | 0.565 |
| FSH (mIU/mL) | 6.028 | 1.091 | 0.188 | 0.328 |
| LH (mIU/mL) | 6.869 | 1.014 | 0 | 0.997 |
| E2 (pg/mL) | 30.752 | 3.795 | 0.249 | 0.193 |
| SHBG (nmol/l) | 36.241 | 7.579 | -0,076 | 0.696 |
| Free Testosterone (ng/dL) | 28.717 | 9.354 | 0.285 | 0.133 |
| AMH (ng/mL) | 2.141 | 0.244 | 0.115 | 0.552 |
| AFC - RightOvary | 10.31 | 2.509 | 0.095 | 0.623 |
| AFC - LeftOvary | 9.276 | 2.59 | 0.063 | 0.746 |
| BMI (beforesurgery) | 41.117 | 3.281 | -0.266 | 0.162 |
Table 1: Correlation analysis of eNOS expression and other clinical indicators prior to bariatric surgery. The table shows the mean values, standard deviations, Pearson correlation coefficients, and p-values for each parameter.
Table 2.
Correlation Analysis of Cpt eNOS and Clinical Parameters Following Bariatric Surgery.
| Variable | Mean | Standard Deviation | PearsonCorrelation | P-value |
|---|---|---|---|---|
| AFC - Left Ovary - Post Surgery | 6.759 | 1.244 | -0.149 | 0.439 |
| AFC - Right Ovary - Post Surgery | 7.379 | 1.635 | 0.037 | 0.849 |
| AMH (ng/mL) - Post Surgery | 2.955 | 0.153 | 0.102 | 0.597 |
| BMI (After 6 Months) | 25.9 | 1.377 | -0.248 | 0.194 |
| E2 (pg/mL) - Post Surgery | 49.2 | 4.538 | -0.156 | 0.42 |
| FSH (mIU/mL) - Post Surgery | 8.87 | 0.08 | 0,08 | 0.677 |
| Free Testosterone (ng/dL) - Post Surgery | 9.421 | 2.524 | -0.151 | 0.436 |
| LH (mIU/mL) - Post Surgery | 8.831 | 1.032 | -0.121 | 0.532 |
| SHBG (nmol/l) - Post Surgery | 64.188 | 12.408 | -0.365 | 0.049 |
Table 2: Correlation analysis between Cpt eNOS and various clinical parameters after bariatric surgery. The table presents mean values, standard deviations, Pearson correlation coefficients, and p-values for each parameter.
Table 3.
Descriptive Statistics and Correlation Analysis of Gene Expression Parameters Prior to Surgery.
Table 3.
Descriptive Statistics and Correlation Analysis of Gene Expression Parameters Prior to Surgery.
| -Statistic | eNOS | CART | Leptin |
|---|---|---|---|
| Count | 29 | 29 | 29 |
| Mean | -4.85 | 0.27 | -1.87 |
| Standard Deviation | 1.75 | 4.43 | 1.75 |
| Minimum | -7.52 | -4.1 | -5.5 |
| 25th Percentile | -6.19 | -2.2 | -2.72 |
| Median | -5.01 | -1.41 | -2.02 |
| 75th Percentile | -3.38 | 0.08 | -0.67 |
| Maximum | -0.23 | 10.72 | 1.09 |
Table 3: Descriptive statistics and correlation analysis ofeNOS A, CART, and Leptin gene expression levels prior to surgery.
Table 4.
Correlation Analysis of eNOS, CART, and Leptin Gene Expression.
| Gene | Correlationwith Cpt0 CART | p-value with Cpt0 CART | Correlationwith Cpt0 Leptin | p-value with Cpt0 Leptin |
|---|---|---|---|---|
| eNOS | 0.038 | 0.848 | -0.301 | 0.120 |
| CART | - | - | 0.077 | 0.697 |
| Leptin | - | - | - | - |
Table 4: This table presents the correlation analysis between the gene expression levels of eNOS, CART, and Leptin before surgery.
Table 5.
Descriptive Statistics and Correlation Analysis of Gene Expression Parameters Following Surgery.
Table 5.
Descriptive Statistics and Correlation Analysis of Gene Expression Parameters Following Surgery.
| Statistic | Cpt1eNOS | Cpt1 CART | Cpt1 Leptin |
|---|---|---|---|
| Count | 29.0 | 29.0 | 29.0 |
| Mean | 1.18 | -3.4 | -0.07 |
| Standard Deviation | 2.31 | 1.12 | 1.55 |
| Minimum | -2.07 | -5.78 | -3.34 |
| 25th Percentile | -0.78 | -4.1 | -1.08 |
| Median | 0.94 | -3.7 | 0.0 |
| 75th Percentile | 2.38 | -2.77 | 1.05 |
| Maximum | 6.77 | -0.74 | 2.64 |
Table 5: Descriptive statistics of Cpt1eNOS, Cpt1 CART, and Cpt1 Leptin gene expression levels following surgery.
Table 6.
Correlation Analysis of Gene Expression Parameters Following Surgery.
| Gene | Correlationwith Cpt1 CART | p-value with Cpt1 CART | Correlationwith Cpt1 Leptin | p-value with Cpt1 Leptin |
|---|---|---|---|---|
| Cpt1 eNOS | 0,16 | 0,4 | 0,039 | 0,838 |
| Cpt1 CART | - | - | 0,282 | 0,136 |
| Cpt1 Leptin | - | - | - | - |
Table 6: This table summarizes the correlations between Cpt1 eNOS, Cpt1 CART, and Cpt1 Leptin with one another after surgery.
Table 7.
The mean gene expression values and standard deviations for CART, Leptin, and eNOS before and after bariatric surgery.
Table 7.
The mean gene expression values and standard deviations for CART, Leptin, and eNOS before and after bariatric surgery.
| Gene | MeanBeforeSurgery | MeanAfterSurgery | St. DeviationBeforeSurgery | St. Deviation Post Surgery | Correlation | p-value |
|---|---|---|---|---|---|---|
| CART | 0.273 | -3.42 | 4.43 | 1.14 | 4.52 | < 0.001 |
| Leptin | -1.87 | -0.130 | 1.75 | 1.55 | -7.35 | < 0.001 |
| ENOS | -4.87 | 1.18 | 1.7 | 2.31 | 0.529 | 0.003 |
Table 7: Gene Expression Changes and Correlation Analysis Prior to and Following Bariatric Surgery.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



