
Submitted:
24 November 2024
Posted:
25 November 2024
You are already at the latest version
Abstract
Fasciolosis is a neglected tropical disease caused by helminth parasites of the genus Fasciola spp. including F. hepatica and F. gigantica being a zoonotic major problem of human and animal health. Its control with antihelminthics is becoming ineffective due to the increase of the parasite resistances. To develop new therapeutic protocols is crucial a deeper knowledge about the molecular bases in the host-parasite interactions. The high-throughput omics technologies have dramatically provided unprecedented insights into the complexity of the molecular crosstalk host-parasite. miRNAs are key players as critical regulators in numerous biological processes modifying the gene expression of cells by degradation of mRNA, regulating transcription and translation functions, protein positioning, cell cycle integrity, differentiation and apoptosis. The large-scale exploration of miRNAs including the miRNome has offered great scientific knowledge steps in fasciolosis further scrutinizing the pathogenesis, the growth and development of their strains and their interaction with the host for the survival of the different parasite stages. This review compilate the updated knowledge related to miRNAs involved in fasciolosis and the generated miRNome highlighting the importance of these key molecules in the host-parasite interactions and the pathogenesis of Fasciola spp. directing towards the development of new biotherapeutic protocols for the control of fasciolosis.
Keywords:
miRNAs
; miRNome
; Fasciola hepatica
; Fasciola gigantica
; zoonosis
; pathogenesis
; ncRNAs
; miRBase
; immunity
1. Introduction
Fasciolosis is part of the neglected tropical diseases catalogued by the World Health Organization (WHO) [1]. It is caused by parasites of the genus Fasciola spp., which includes the species of Fasciola hepatica (F. hepatica) and Fasciola gigantica (F. gigantica) and has been described as the most geographically widespread parasitic zoonosis [2,3] with more than 180 million people at risk of infection [4]. Fasciolosis is a major problem in the farming industry as is responsible for low fertility and a reduction in milk and meat production in sheep, goats and cattle, causing economic losses estimated at USD 3.2 billion per year [5].
The disease control system is based on the rotation of paddocks and the use of antihelminths such as triclabendazole (TCBZ) [6]. However, the widespread use of TCBZ has led to the reduction in the efficacy due to the development of parasitic resistance [6,7], adopted by an evolutionary mechanism in the new parasitic strains, starting from the populations that survive the treatment, where a small genomic region was inherited with dominant traits of resistance [8,9].
Infections are recognized for their chronicity due to the successful survival of invaders within the mammalian host [10,11]. F. hepatica survival is managed by a series of interactions between the immune system and the excretory/secretory products (ESPs) the parasite releases, some of them provided through extracellular vesicles (EVs) [12,13]. This ESPs are involved in the dysregulation of host genes associated with metabolism, immune response and tissue repair/regeneration [14].
The use of omics has improved information and understanding of the entire F. hepatica genome [15,16], evidencing the role of non-coding RNAs (ncRNAs) such as micro-RNAs (miRNAs) in modifying the gene expression of cells by degradation of mRNA [17,18], regulating transcription and translation functions, protein positioning, cell cycle integrity, differentiation and apoptosis [19,20]. Small RNAs are a type of non-coding RNAs (sncRNAs) that include miRNAs among others. They are associated with Argonaute family proteins (Ago family proteins) and suppress unwanted genetic materials and transcripts (RNA silencing or RNA interference, RNAi) [21,22,23]. RNAi fragments provide post-transcriptional silencing of proteins resulting in the genetic domain in a cell type directing its phenotype and functionality, being classified as vital in biological development, homeostasis and alteration in a living organism [24,25].
The large-scale exploration of miRNAs including the miRNome has offered great evolutionary steps, revealing the participation of these in processes such as apoptosis, differentiation and cell proliferation, DNA repair, lipid metabolism, enzymes and amino acids, among other functions of cells [26]. This innovation has been key in the interaction of pathogens in a mammalian host, as they are involved in the immune response, which helps to understand the pathogenesis of many metabolic alterations and in the search for new biotherapeutics that help in the control and/or eradication of these [25,27,28].
The miRNA annotation system is based on published experimental investigations of different organisms (animals and plants) that identify sequences of stable loops in the genome. With mapping system, the stable loops that correctly assemble generate a reliable mature miRNA [29]. These sequences are integrated into open-access databases such as miRBase.org (Release 22.1; http://www.mirbase.org/), miRCarta (Version 1.1; https://mircarta.cs.uni-saarland.de), miRGeneDB (Version 2.0; https://mirgenedb.org), RumimiR (http://rumimir.sigenae.org/) as repositories to facilitate prediction, validation and annotation of identified target miRNAs and generate a landscape for searching for new sequences [30,31,32,33].
Since the first comparative characterization of miRNAs from F. gigantica and F. hepatica and the discovery of its implication in the modulation of host immune responses, a better understanding about miRNAs and miRNome in fasciolosis has been postulated as crucial considering that are hypothesized as promising "anchors" in the binding of molecular networks, directing towards new biotherapeutic protocols for the control of this disease [34,35,36,37,38]. Thus, this review present a current version of the miRNAs identified in fasciolosis and the miRNome derived from its analysis, highlighting the importance of these key molecules in the host-parasite interactions and the pathogenesis of Fasciola spp.
2. A Look at the Past in miRNAs: Discovery and Historical Landmarks
The dogma of molecular biology recognizes RNA molecules for their ability to synthetize proteins from the genetic information encoded in the DNA. However, the function of RNA molecules is not limited to being a messenger for protein synthesis and, only about 1–2% of the RNA present within a human cell is protein-coding, the remainder being non-coding RNA (ncRNA) [39].
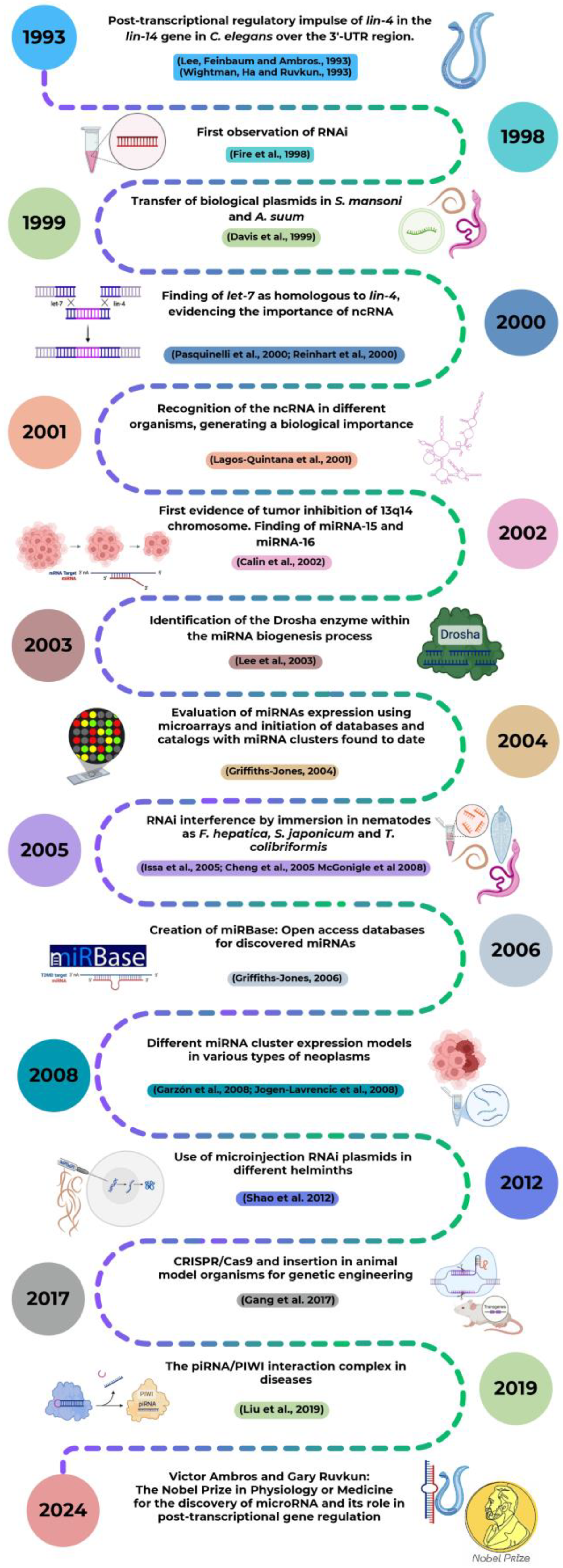
The first evidence that not all RNA molecules code for proteins described that the heterochronic lin-4 gene can interact with the regulatory site of the lin-14 gene, exerting a negative control leading to suppression of its function [40]. Eventually, the implication of lin-4 gene in the regulation of nematode larval development in Caenorhabditis elegans (C. elegans) was demonstrated [41] (Figure 1). Interestengly, lin-4 does not encode a protein but small transcripts of approximately 22 and 61 nucleotides (nt) which contain sequences complementary to a repeated sequence element in the 3’ untranslated region (3’-UTR) of lin-14 mRNA, suggesting that lin-4 regulates lin-14 translation via an antisense RNA-RNA interaction [41]. In this way, Lee et al. 1993 [41] and Wightman et al. 1993 [42] demonstrated that the gene lin-4 in C. elegans exerts a post-transcriptional regulatory impulse on the gene lin-14 specifically on the 3’-UTR region that did not code for proteins, being these the first miRNA discoveries that paved the way to a new gene regulation paradigm where short ntRNAs of 22 nt have the ability to repress mRNA expression by specific binding to the 3’-UTR region [41,42].
Since this first discovery, a serial of crucial findings occurred enriching the knowledge about miRNAs and the role they play as critical regulators in numerous biological processes (Figure 1). Studies related to the effectiveness of double-stranded RNA compared to single strands at producing interference and RNA transfection experiments in helminth parasites to study gene expression were eventually carried out [43,44]. In 2000, the let-7 RNA gene and its implication in the timing of C. elegans development together with lin-4 is reported, reaffirming the manipulation exerted by ncRNAs in gene expression [45,46]. Particularly, miRNAs postranscriptional regulatory mechanisms both in invertebrates and vertebrates seems to play an important role that should be taken into account in future studies [47]. Being up or downregulated, miRNAs were discovered to be implicated in the development of certain disorders, namely B cell chronic lymphocytic leukemia and acute myeloid leukemia [48,49,50]. The identification of Dicer as the enzyme implicated in the cytoplasmic processing of pre-miRNAs into mature miRNAs was previously described, and the discovery of the Drosha enzyme implication in nuclear processing of the pri-miRNAs completed the understanding of the microRNAs biogenesis mechanism [51]. The first sequence and annotation data derived from the discovered miRNAs was published in an accesible database [52] and eventually the miRBase repository was created, aiming to provide integrated interfaces to comprehensive miRNA sequence data, annotation and predicted gene targets [53].
The use of advanced techniques such as immersion RNAi [54,55,56] and RNAi microinjection [57] as gene knock-out strategies that could reduce parasitic infection were implemented. The development of a method to target gene disruptions and mutant phenotypes was studied in parasites, introducing CRISPR-Cas9-mediated gene disruptions to enable future studies of gene function [58]. This, together with, the discovery of sncRNAs that interacts with the PIWI protein (piRNA/PIWI complex) which are expressed in a tissue-specific manner thus participating in various diseases, could become potential targets for therapeutic intervention [59].
The unvaluable potential of miRNAs has been oficially recognised when Victor Ambros and Gary Ruvkun were awarded with the 2024 Nobel Prize in Physiology or Medicine for the discovery of microRNA and ist role in post-transcriptional gene regulation [60].
3. Location and Biogenesis of miRNAs
The understanding of the location of miRNAs is key to determine how miRNAs are transcriptionally regulated. According to their genomic position, they can be classified into intragenic and intergenic miRNAs. Those located within protein-coding or non-coding genes are called intragenic miRNAs and genes in which the miRNAs are embedded are called host genes (intergenic) [63]. The biogenesis and function of miRNAs are tightly regulated and their dysregulation is often associated with diseases and disorders [23]. The biogenesis of miRNAs is divided into two important pathways: canonical and non-canonical [61,64]. The canonical is the dominant pathway by which miRNAs are processed (Figure 2). In this, the primary miRNA (pri-miRNA) is transcribed from its gene and processed into precursor miRNA (pre-miRNA) by the Microprocessor complex, which consist of a Drosha Ribonuclease II (DROSHA) and a RNA binding protein called DiGeorge syndrome critical region 8 (DGCR8) in vertebrates, which are localized in the nucleus [64,65]. DROSHA is responsible for cleaving the DGCR8-recognized pri-miRNA to the 70 nt pre-miRNA at sufficient distance from the base of the hairpin, defining one end of the mature miRNA. Once pre-miRNAs are synthesized, an exportin 5 (XPO5)/RanGTP complex transports them to the cytoplasm and they are processed by the RNase III endonuclease enzyme called Dicer which further cleaves the pre-miRNA to a miRNA duplex and selects the miRNA with the least stable 50-end for loading into the RNA-induced silencing complex (RISC), whose primary constituent is an Argonaute (Ago) protein, which degrades the duplex miRNA to generate a single strand with 3‘ and 5’ ends called mature miRNA or miRNA-RISC (miRISC) (Figure 2). The mature miRNA is then capable of targeting mRNA transcripts for repression through a variety of mechanisms [49,66].
In the non-canonical pathway, diverse small RNAs structurally an functionally like miRNAs are synthesized [28]. It is classified into three groups: Drosha/DGCR8-independent, Dicer-independent and terminal uridylyl transferases (TUT) dependent [23,28,67]. miRNAs resulted from the non-canonical biogenesis are low in abundance and poorly conserved and the vast majority of functional miRNAs follow the canonical pathway for their biogenesis. For this reason, the functional relevance of non-canonical miRNAs should be interpreted with caution [23].
4. miRNAs involved in Fasciolosis and its Pathogenesis
F. hepatica modulates the host immune response since newly excysted juveniles (NEJs) enter the host peritoneal cavity by penetrating the intestinal wall after the excystment of the metacercaries within the duodenum. This NEJs together with its ESPs released promote the development of M2 activated macrophages since 24-48 hours post-infection [68,69] thus imparing the ability of innate immune cells to respond to stimulation and avoiding the development of antigen specific Th1 and Th17 responses and enhancing the differentiation of Th2 and Treg cells. This scenario promotes infection and long-term parasitic survival within the host [70,71,72].
miRNAs identified within these ESPs play a role in predicted targeted sites related to the establishment of an immune response, thus suggesting its implication in the immunomodulation of host’s responses, helping in the progression of parasitic infection [19,73,74]. For all these, miRNAs have been postulated as promising biomarkers in the diagnosis of parasitic diseases and may serve as novel targets for therapeutic intervention [19,75]. In addition, it has been reported that the altered expression of miRNAs may be associated with antihelmintic resistance [76].
The first study focused on F. hepatica miRNAs described a very small subset of the actual miRNAs available. The absence of a F. hepatica reference genome lead to Xu et al. 2012 [34] to utilize the genome of Schistosoma japonicum and use a smallRNAseq and BLAST approach, identifying a total of 16 miRNA candidates from F. hepatica. 11 miRNAs were shared by F. hepatica and F. gigantica, including 8 conserved and 3 novel. The conserved miRNAs all matched perfectly with those from S.japonicum in the miRBase database. A total of 5 F. hepatica-specific miRNA sequences were identified, namely fhe-mmu-miR-1957, fhe-miR-novel-01, fhe-miR-novel-08, fhe-miR-novel-07 and fhe-miR-novel-10, representing the first Small RNAseq data for F. hepatica adults [34]. Whether or not these are important for infection of the mammalian host has yet to be determined [17]. The predicted target genes of F. hepatica miRNAs are proteins related to reproduction, development processes, response to stimuli, immunomodulation, and locomotion [34].
Using the pipeline miRCandRef and available genomic sequence data [16], the miRNA complement was increased from 16 to 55 miRNAs (41 conserved and 13 novel), and the production of miRNA-containing EVs by F. hepatica adults was confirmed [77]. In this new 55 miRNAs list, the 8 conserved miRNAs identified by Xu et al. 2012 [34] were observed. Nevertheless, the 8 novel miRNAs proposed were not found [77], suggesting that they are not genuine miRNAs and thus being removed from the listed miRNAs for F. hepatica [37]. In addition, mir-2a-A, bantam, mir-1175, mir-277 and let-7 were found among the 10 most highly expressed molecules in the cellular fraction. On the other hand, mir-125b, mir-87, mir-2a-B, mir-2b-A and mir-1993 were among the 10 most highly expressed in the EV data. Concretely, fhe-miR-87 and fhe-miR-1993 are absent in mammals and fhe-miR-2b-A and fhe-miR-2a-B are orthologs of miR-27, bta-mir-27a, and bta-mir-27b. On the other hand, fhe-miR-125b represented the highest expression in EVs [77].
miR-125b in Fasciola spp. is closely related to its human homologue hsa-miR-125b suggesting the implication in the control of host immune responses during early infection stages [35]. Previous studies have documented human hsa-miR-125b as a miRNA that regulates the infammatory response of macrophages by controlling the activation of M1 macrophages [78,79] and its overexpression in host macrophages is related to an effective antigen presenting cell stimulation of T cell responses, elevating the responsiveness to IFN-γ, thus suggesting a role for miR-125b in activating macrophages [80]. Concretely, fhe-miR-125b is internalized by host macrophages, mimicking host miR-125b and decreasing inflammatory cytokines by interrupting the MAPK signaling pathway through TRAF6 targeting [35].
All the conserved miRNA families previously reported in the adult stage of F. hepatica (fhe-miR-125b, fhe-miR-bantam, fhe-let-7c, fhe-miR-277, and fhe-miR-71/miR-2 cluster members) [34] were detected and expressed at very high levels in the juvenile stage (NEJs), being miR125b the most abundantly expressed, followed by let-7 and bantam [81]. miR-277 serves as a biomarker for the diagnosis of infestasion as has been recognized in host serum [82,83], and it is related to the regulation of enzymes that play a role in the survival of the parasite under stress or starvation conditions [84]. Let-7 is an ancient class of miRNA that is tipically associated to miR-125 as a polycistronic transcript and both miRNAs can act together as key regulators of development [85]. In humans, there are 10 mature let-7 miRNAs, including let-7c [86]. It has been demonstrated that let-7c regulates bactericidal and phagocytic activities of macrophages and the overexpression of let-7c in Granulocyte Macrophage Colony-Stimulating Factor (GM-CSF)-induced bone marrow-derived macrophages (GM-BMM) diminished M1 phenotype expression while promoting polarization to the M2 phenotype expression [87].
The miRNA bantam has been implicated in the regulation of cell proliferation and apoptosis [88] and represent a direct marker for schistosomiasis in human serum [82]. Interestingly, Xue et al. (2008) [89] showed that three of the miRNAs that we find in serum (sja-bantam, sja-miR-71 and sja-let-7) are expressed during all the stages of parasite development but are enriched in the cercariae, suggesting that they may be important during the initial stages of schistosome infection. In addition, it has been demonstrated that miR-125b and bantam from S. japonicum interfere in host macrophage functions by increasing TNF-α, facilitating parasite development, metabolism and egg laying [90,91]. Interestingly, the miR-10, bantam and let-7 family members have been postulated as important miRNAs implicated in the modulation of the host immune respones, being the most abundant miRNAs in EVs secreted by adult trematodes [38,92]. The metazoan conserved fhe-miR10-P2a (named fhe-miR-125b in miRBase) was the most highly expressed miRNA across all stages and in the EV fraction in the adult stage of F. hepatica [38]. On the other hand, the miR-1 and let-7 family are part of the ES of EV in Trichinella spiralis where they mediate the polarization of macrophages M2 and are related to the low regularity of the IL-10 and the activity of TGF-β in the host [93].
With the available full genome of F. hepatica [16] and the three small RNA-seq libraries, Fromm et al. 2017 [94] found evidence for the presence of 2 new miRNA families in F. hepatica (mir-210 and mir-750), using improved algorithms. Thus, the miRNA complement of F. hepatica consist of 53 members of 32 conserved miRNA families and 5 novel F. hepatica-specific miRNA genes. There was evidence that F. hepatica miRNA profile vary significantly in adults, juveniles and EVs and also the abundance of the expressed miRNAs varies through the F. hepatica lifecycle, indicating stage-specific roles [94]. Concretely, mir-71-P1b is the most expressed miRNA in adult worms, mir-277-p2 the most expressed in EV data and mir-10-p2a the most expressed in juvenile worm-derived sample. Mir-10-p2a, mir-2-p1b, bantam, mir-1175 and mir-190-p2 are the most highly expressed molecules in the cellular fraction and mir-10-p2b, mir87, mir-2-p2b and mir-2-p4 are among the most highly expressed miRNAs in the EV data. These miRNAs have been catalogued with immunoregulatory characteristics [94].
EVs-derived miRNAs perform a key role in communication processes between parasite-parasite and host-parasite [95]. In ruminants infected with F. gigantica, a dysregulation of circulating miRNAs was observed and 4 parasite-derived miRNAs (fgi-miR-87, fgi-miR-71, fgi-miR-124 and fig-miR-novel-1) were detected in serum, indicating that host’s circulating miRNA profile is altered during Fasciola infection [96]. Concretely, miR-71 was evidenced in the exosome content in Brugia malayi and F. gigantica [96,97], mediates in nitric oxide levels on host macrophages [98] and modulates the host immune response together with miR-87 [92]. In addition, miR-71 and miR-87 have been suggested as biomarkers in fasciolosis [96]. Robb et al. 2022 [99] described a strong downregulation of fhe-mir-124-3p in in vivo F. hepatica juveniles, suggesting its relation with neuronal cell differentiation and contributing to the control of numerous biological processes.
Apart from those, miR-126a-3p, miR-150-5p, miR-155-5p, miR-181a-5p and miR-362-3p were found to be dysregulated in the liver of mice treated with ESPs from F. gigantica at 1, 4 and 12 weeks post-exposure, revealing their important roles in the interplay between liver and F. gigantica ESPs. Interestingly, all these dysregulate miRNA participate in cellular apoptosis, migration and proliferation, suggesting that they may play roles in liver pathological changes induced by the liver flukes [100]. In addition, the downregulated expression of miR-466i-5p and miR-423-5p may be implicated in biliary secretion and liver functions in F. gigantica infection, and high expression of miR-466-5p has been found in hepatocytes in response to inflammatory stimuli [100,101]. The expression of miR-155 has been related to chronic liver inflammation and fibrosis and miR-126 plays a role in increasing the inflammatory activity in the liver while inducing the expression of IFN-γ, MAP kinase pathways and differentiation of FoxP3 regulatory cells [102,103].
Ovchinnikov et al. 2020 [104] described an increase in the abundance of let-7-P3_5p, mir-10-p2b_5p, mir-83_3p, mir279_3p and mir-1993_3p in EVs relative to the adult worm both in S. mansoni and F. hepatica and mir-1989_5p was substantially upregulated. In addition, mir-71 and mir-277 families were found to be protostome-specific miRNA, being absent in cow and human. Moreover, a single nucleotide difference between mir-277-P2 mature sequence could be interesting to differentiate the two flukes. A significant part of the predicted targets for S. mansoni and F. hepatica EVs miRNAs are related to the glucose concentration in blood and surrounding tissues, T cell immunity and cytokine signaling. According to this, helminths may modulate level of glucose concentration in host blood and tissue, reduce inflammatory and immune responses and these modulations can contribute to the survival of the parasites inside their hosts. F. hepatica miRNAs may also have an interesting target: LIN28A gene, that may minimize host’s Let-7 miRNAs expression thus helping in Treg differentiation [104,105].
Hu et al. 2021 [106] demonstrated that F. gigantica and F. hepatica showed considerable levels of conservation of miRNAs. A small RNA sequencing of 8 lifecycle stages of F. gigantica was conducted and 56 miRNAs from 33 conserved families were detected, and out of these only 10 miRNAs were previously known in F.gigantica, being these miRNAs from 8 conserved and previously known families (bantam, let-7, two miR-2, miR-10, miR-12, two miR-71, miR-87, and miR124). The top expressed miRNA varied greatly at each stage, evidencing the substantial roles of miRNAs in the parasite’s development. Three of the most highly expressed miRNAs (bantam_3p, miR-71-P1_5p and miR-2-P1a_3p) have been also found in F. hepatica belonging to the top expressed miRNAs from NEJs, adults and EVs [94,104]. In addition, two mature miR-1992 and miR-novel-3 sequences could be of interest to differentiate the two fluke species [106]. This could be of interest for their use as taxonomic markers as F. gigantica and F. hepatica can co-infect the same host, increasing the virulence of the infection mainly due to the development of antihelminthic resistance scenarios [106].
It has been previously reported the capacity of F. gigantica ESPs to dysregulate hepatic miRNAs in mice and to change the expression of miRNAs in goat peripheral blood mononuclear cells PBMCs) [100,107]. Overall, in the study of Wang et al. 2021 [107] a reduction in the expression of miR-211 and miR-204-5p was reported, which may imply a possible involvement of F. gigantica ESPs in promoting the proliferation of PBMCs. The expression of miR-148a-5p was upregulated and this has been described to inhibit pro-inflammatory cytokines and may help in shifting Th1/Th2 towards the Th2 immune response [107].
Fontenla et al. 2022 [38] analyzed the expression of diverse sncRNA in the key stages of invasion and establishment of F. hepatica infection in the definitive host. The results indicated that sncRNAs are present in all stages, and also in secreted vesicles, being miRNAs the most abundant ncRNA followed by tRNA-derived fragments. The comparison with the recent study in F. gigantica [106] confirms the presence of 34 conserved families and a growing set of miRNAs described so far only in Fasciolidae. In addition, 9 novel miRNAs (fhe-miR-NEW-1 to 9) to those already described and recompiled recently [37] are added, extending the Fasciola specific miRNome [38]. The conserved miRNAs most highly expressed accross stages and EVs were fhe-miR-125b fhe-miR-71-P1b and bantam. Moreover, miRNAs were classified into 5 expression clusters. Genes targeted to cluster 1 (representing a set of miRNAs highly expressed in metacercariae while lowly expressed in NEJs) are enriched in processes associated with vesicle organization and transport and membrane fusion. Cluster 2 (miRNAs strongly downregulated in metacercariae while upregulated in NEJs and adults) are related with genes implicated in vesicular cargo, maturation and trafficking in the Golgi, IL-12 signaling and response to inorganic substances/temperature stimuli that could be relevant for succesful excystment and development. Cluster 3 includes miRNAs highly expressed in adults and is related to cell differentiation and tissue morphogenesis of epithelial cells and cell to cell contact genes, suggesting its role in the maturation of juvenile forms during the formation of the syncytial tegument. Cluster 4 include miRNAs highly expressed in NEJs but strongly regulated in adults and functions associated with the production of eggs in the adult stage, like nervous development, transcription regulation and germinal function. Finally, cluster 5 represents miRNAs strongly downregulated in adults but expressed in metacercariae and NEJs, being associated with vacuolar proton transport ATPases, enzymes that are related to the acidification of vacuoles, a relevant requisite in feeding and interaction with the host [38].
The formation and release of vesicles in F. hepatica may be regulated by fhe-mir-71-P1b, fhe-miR-71-P2, fhe-miR-1-P1, fhe-miR-1-P2, fhe-miR-96 and fhe-miR-7-P1 [38]. The secretion of large amounts of different types of vesicles in NEJs has been reported in different developmental stages of F. hepatica. These vesicles were found to be pre-formed in the metacercariae [108].
A redundant total of 186 mature miRNA sequences is obtained from the manual analysis performed by Herron et al. 2022 [109] of all published F. hepatica miRNAs [34,77,81,104]. Naming of miRNA sequences was a difficult task as they were found to be inconsistent between individual papers and with the originally described miRBase namin system [110]. Small RNA sequencing of multiple F. hepatica life stage libraries was obtained and these datasets were combined and then qualitatively analyzed for miRNAs using miRDeep2, yielding 91 mature miRNAs sequences across multiple F. hepatica life stages, 29 of which has been previously reported in F. hepatica [109] and representing the largest single miRNA dataset reported for F. hepatica. Most sequences (61) were newly described miRNAs that lacked matches in miRBase or miRGeneDB searches, extending the F. hepatica miRNome to 150 sequences, providing the first developmental profile of miRNAs in intra-mammalian F. hepatica life stages, providing also computational predictions of miRNA – mRNA functional regulatory networks for both cellular and secreted miRNAs, including neuromuscular transcipts, secreted metabolic modulators and nutrient scavengers, secreted proteases and individual components of exosome and glycan biosynthesis [109]. Fhe-mir-125a-5p, fhe-mir-1899-5p, fhe-mir-277-3p and fhe-mir-71b-5p were present in all six libraries. Overall, the majority of miRNAs were highly expressed in metacercariae. This may be related to the induction of stasis in this life stage, where miRNAs could be involved in pausing transcription until a host is encountered. In addition, Herron et al. 2022 [109] were the first to focus on miRNA targets that are demonstrably downregulated during fasciolosis using transcriptomes from sheep/cattle tissues and 298 miRNA-targeted transcripts were significantly downregulated across fasciolosis transcriptomes from lymph node, PBMCs and liver from sheep and/or cattle.
Ricafrente et al. 2022 [18] described that the genome location of most F. hepatica miRNAs (70%) were found to be intergenic and this could be interpreted as a parasite capacity to transcribe their miRNA independently of worm gene expression. This has been recorded as an adaptation in other platyhelminthes [111,112] to transcribe miRNAs in response to external host signals and regulating host genes without significantly disturbing the parasite’s developmental transcriptome [18]. Although let-7 and miR-125 have been associated with the same polycistronic transcript, these miRNAs appear to be organised separately within different scaffolds of the F. hepatica genome [18,81]. The identification of corresponding gene targets for novel miRNAs discovered across F. hepatica three life stages (RNA isolated from NEJs excysted in vitro and cultured for 6h, 24h and 7 days) indicates that there is differential expression between the life stages that ensure the biological processes required for each developmental stage, and are switched on and off as necessary. Thus, genes associated to the integral component of the membrane appears to be predominant biological processes in the NEJ parasites. As the NEJs develop to immature fluke, overexpression of genes associated with nucleotide synthesis, transcription and translation, energy regulation, cell signalling and proteolysis are found to be overexpressed. Bantam-3p, miR-1b-3p, miR-71a-5p, miR-71c-5p, miR-125b-5p, miR-190a-5p and miR-277-3p were the most abundant miRNAs present in every life stage, changing their relative abundance as the parasite develops, grows and matures. It has been reported that FhCL3 expression is switched off as parasite matures and moves from tissue-feeding to blood-feeding. Interestingly, FhCL3 is the unique target of bantam-3p and its increase during immature and adult stages may influence in the downregulation of FhCL3 expression, suggesting that this protein is specifically required by the NEJs and may play a critical role in the transition from NEJ to the immature worm. Similarly, the SAP30-binding protein regulation is impaired by F. hepatica miRNAs in the immature stage and in the adult worms, thus suggesting a critical role in F. hepatica egg production in these life stages. Other gene highly regulated during the NEJ stage is SACMAD1 suggesting that the miRNA-mediated suppression in NEJs is related to the enhanced proliferation of neoblasts observed during the 24 h after excystement [18].
In NEJs cultured in vivo for 21 days, a downregulation of Catephsin B proteases genes together with an upregulation of Cathepsine L protease genes have been reported, thus suggesting a switch to produce Cathepsine L proteases in F. hepatica juveniles in vivo when compare with the NEJs cultured in vitro, which show lower levels of development and are unable to switch Cathepsine expression, suggesting that this switch depends on host specific signals [99]. In addition, the upregulation of fhe-let-7a-5p miRNA and the downregulation of fhe-mir-124-3p in in vivo juveniles is related with cell differentiation, regulation of stem cells, growth induction and lower levels o neuronal development. Overall, in the study of Robb et al. 2022 [99] it is highligted that, despite the fact that in vitro maintained juveniles are smaller in size and experience a delay in growth and development due to the absence of host signals when compared to the in vivo juveniles, the in vitro maintenance platform covers F. hepatica relevant biological processes as >86% of genes are expressed at similar levels in both culture systems. In addition, Wnt signalling pathway proteins may have key roles in early stage developmental processes as an overall downregulation of Wnt-associated mRNAs in in vivo NEJs has been reported [99].
Similarly to miRNAs, long non-coding RNAs (lncRNAs) have been mainly associated with transcriptional regulation. The first description of the lncRNA complement of F. hepatica was made by McVeigh et al. 2023 [113]. lncRNAs contain miRNA binding sites as demonstrated by the in silico binding prediction between the 150 miRNAs previously reported in F. hepatica [18,109] and the lncRNAs described in F. hepatica [113], resulting in a consensus set of 4104 lncRNA:miRNA pairs. The large amount of miRNA binding sites on lncRNAs may indicate a miRNA-driven post-transcriptional regulation of lncRNA expression. The hypothesis that lncRNAs contain miRNA binding sites and thus acting as “sponges” for miRNAs has been previously exposed [114]. Alternative therapies for the treatment of fasciolosis could be derived from the implementation of silencing these lncRNAs or interrupting the regulatory interaction with its target similarly to miRNAs, which are currently under study in many clinical trials to become an alternative mean of therapy for many conditions [113,115].
The potential of miRNAs present in sera as potential biomarkers for the early diagnosis of fasciolosis in sheep was evaluated by Chowdhury et al. 2024 [20]. Two parasite miRNAs (miR-124-3p and miR-Novel-11-5p) were postulated as potential biomarkers as they were detected at all timepoints during F. hepatica infection. Concretely, miR-124 and miR-71 have been previously described in fasciolosis and postulated as promising biomarkers for this disease [96,106]. The sheep miRNAs postulated as potential biomarkers in pre-hepatic infection were oar-miR-133-5p, oar-miR-232a-3p and oar-miR-1197-3p, being oar-miR-3956-5p consistently elevated both in pre-hepatic and hepatic infections [20]. These miRNAs have been evidenced in other types of diseases and postulated to act as possible biomarkers [116,117].
Given the importance of macrophages during early and chronic immune responses in fasciolosis and implication of miRNAs in its function, Bąska et al. 2024 [118] evaluated the expression of miRNAs in human macrophages THP-1 activated by LPS derived from F. hepatica ESPs using microarray assays but no evidence of internalized miRNAs in these cells was observed. miR-1537p was found to be the highly expressed miRNA but with low significance. The authors attribute a low sensitivity for the detection of miRNAs by the microarray assay together with an insuficient concentration of F. hepatica ESPs. Previously, miRNAs expression has been evaluated in Fasciola spp. infections involving the metabolism and response of mononuclear cells [35,107], where used higher concentrations of ESPs and the analysis techniques were through next-generation sequencing (NGS), which are more used and recommended for assertiveness compared to microarray [119]. Moreover, the expression of miRNAs in macrophages in BALB/c mice infected with F. hepatica was also evaluated [120]. The expression of a total of 8, 22 and 3 miRNAs was registered at 6 hours, 18 hours and 5 days postinfection, respectively. These presented specific target genes related to the inflammatory response and induction of proinflammatory cytokines [121,122]. Expression of fhe-miR-125b was prominent within the parasitic miRNAs analyzed, corroborating previous research demonstrating its role within the molecular machinery of such protective cells [35,123]. Likewise, the total number of miRNAs present at 18 hours post infection was related to the migration of NEJs through the peritoneal cavity, a crucial time for their interaction with host-derived macrophages, suggesting the parasite's need to evade a hostile environment until reaching the hepatic organ at approximately 5 days, where they crucially decrease [120].
5. Unraveling the miRNome in Fasciolosis
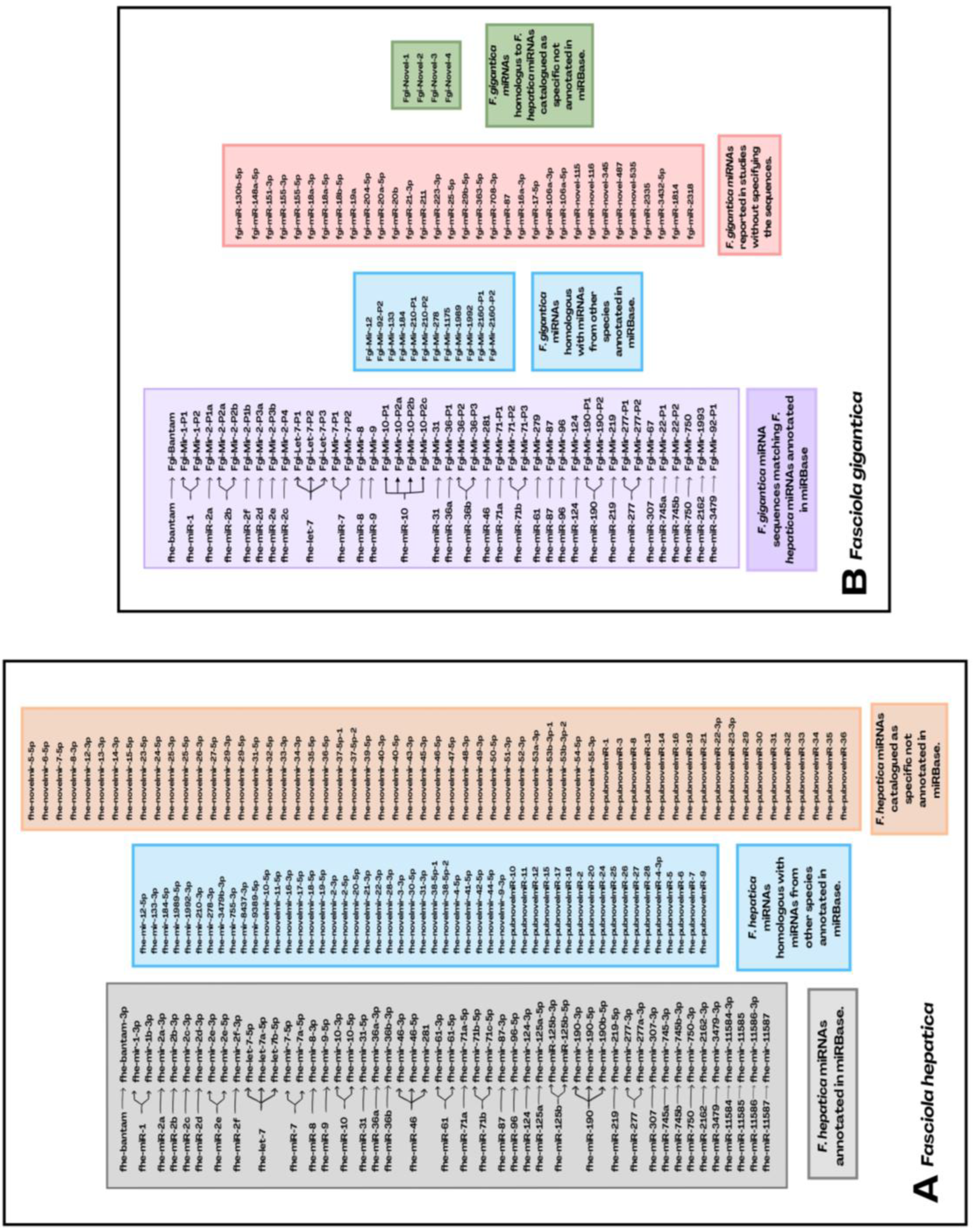
Since the first evidence of miRNAs for Fasciola spp., characterisation of candidate families has been achieved through various methodologies with a greater focus on F. hepatica than F. gigantica. The first five investigations [34,77,81,94,104] gave a total of 85 miRNAs for F. hepatica, but 8 of these [34] were claimed to be genomic repeats [77,81] leaving a total of 77 conserved, where 36 are expressed in the juvenile and adult stages, 15 appear to be exclusive in NEJs and 26 in the adult, considering 17 as novel as they do not match counterparts in other organisms [37].
The number of miRNAs for F. gigantica are less clear. To date, ~92 miRNAs have been published throughout its life cycle where ~47 appear conserved with F. hepatica [34,96,106,107], postulating miR-1992 and miR-Novel-3 which could differentiate the two strains of Fasciolidae, ~12 are homologous with other species [106], and ~33 are suggested to belong to conserved families. However, since their sequences are not presented in the publications or in the supplementary material, it is difficult to confirm this hypothesis.
Ricafrente et al. 2022 [18] expanded the F. hepatica miRNome to 124 miRNA among NEJs, immature (12 dpi) and adult stages of which 72 were previously reported and 52 were discovered with three of these as conserved sequences (fhe-miR-493-5p, fhe-miR-2335-5p and fhe-miR-6613-3p) and 49 were assigned as novel, with 31 miRNAs homologous to F. gigantica and 18 specific to F. hepatica. At the same time, Fontenla et al. 2022 [38] confirmed the miRNAs previously identified in F. hepatica by Ricafrente et al. 2021 [37], validating 4 of them as homologues of the miRNAs described for F. gigantica [106] and adding 9 miRNAs classified as novel (fhe-miR-NEW- 1 to 9) present in all stages of the parasite and its EVs, suggesting that 7 sequences are conserved within F. gigantica.
A manual reset of the miRNAs published for F. hepatica previously [34,77,81,94,104], obtained a set of 89 non-redundant miRNAs conserved for the worm, including 3‘and 5’ and precursor hairpin regions [109]. The sequence alignment performed expands the number of specific miRNAs to 52 (fhe-novelmir-1 to 45), maintaining the expression of 3‘ and 5’ regions of mature miRNA sequences that manifest it, leaving a total of 150 miRNAs present at different stages of their life cycle. The presence of a large number of novel miRNAs suggests the evolution of these ncRNA, inviting the discovery of homologues in F. gigantica that support this hypothesis [109].
Thus, in 2022 three studies were published adding 70 new sequences for F. hepatica [18,38,109] but 2 of these, named fhe-Novel-3 and fhe-Novel-39, were reported by Ricafrente et al. 2022 [18] as identical of 2 miRNAs (fhe-novelmir-44-5p and fhe-novelmir-18-5p) described by Herron et al. 2022 [109], suggesting that the current number of novels is 68 miRNAs. However, Herron et al. 2022 [109] performed sequence alignment taking into account the 3‘ and 5’ regions that a single miRNA might have, which may suggest divergence between mature sequences of the same miRNA [53]. In addition, Herron et al. 2022 [109] gave a consensus name to the 150 miRNAs using the nomenclature for the annotation of miRNAs in databases [110]. This would avoid the redundancy generated by different authors thus allowing miRNAs to be precisely identified. The last two publications [99,113] that integrate miRNome into their study refer to the report by Herron et al. 2022 [109] making it possible for the miRNome to support 150 miRNAs in total.
Finally, Robb et al. 2022 [99] identify 14 new miRNAs for F. hepatica with supported mature sequences classifying them as novel and listing them under the previously stipulated consensus name (fhe-novelmir-45 to 53) and retaining the 3‘ and 5’ regions of the miRNAs that presented it, leaving a total of 164 miRNAs for the worm at present. However, the miRNA sequences accepted to date in miRBase are 38 for F. hepatica and considered to be genuine to 1 sequence, but others have not been added because they do not meet the classification criteria for miRNA formation and maturation, which possibly refers to the type of precursor hairpin assembly and the assertiveness of the analytical tools used [110]. Nevertheless, the criteria in other databases, such as mirGeneDB, consider a wider range for hairpin type and expression of miRNAs according to the variety in their canonical/non-canonical biogenesis, drosha- or dicer-dependent, change at the 3‘ and/or 5’ ends postulating further criteria for the annotation of miRNAs in metazoans [37]. For this reason, it is suggested to take into account within the miRNome the number of miRNAs that researchers describe in their studies, even if they are not yet included in the miRBase (Figure 3).
6. Conclusions
Fasciolosis is considered one of the most pathogenic parasitic zoonosis in the world and its control has been undermine mainly because the increased resistance to the antihelmintic treatments specially in the endemic areas. The deeper knowledge of the molecular bases leading the pathogenesis of fasciolosis is determinant to establish efficient strategies for the control of the disease diminishing its pathogenicity. The development of high-throughput omics technologies has dramatically provided unprecedented insights into the complexity of the molecular crosstalk host-parasite emphasaising the critical role of bioinformatics tools in managing the big data generated. To date, the discovery of miRNAs and the current miRNome have improved the understanding of fasciolosis, further scrutinizing the pathogenesis, the growth and development of their strains and their interaction with the host for the survival of the parasite. This innovative advancement is important for understanding the molecular networks involved, leading to the discovery of key points for achieving disease control for human and animal health. Future directions are focus on integrating multi-omics data to allow researchers gain a holistic view of biological systems, leading to the identification of new biomarkers and therapeutic targets.
Author Contributions
Literature retrieval and information collection: D.M.B.-T., G.H.-T., M.T.C.-R. and V.M.-H.; Manuscript writing—original draft preparation, and designed the figures: D.M.B.-T., G.H.-T., M.T.C.-R. and V.M.-H.; Writing and reviewing of the paper: D.M.B.-T., G.H.-T., M.T.R.-C., J.P., V.M.-H., A.M.-M., F.J.M.-M., R.Z., L.B. and P.J.R.-M; Project management: V.M.-H., M.T.C.-R. and J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Authors thanks the websites BioRender.com and Canva.com for their help in perform part of the figures 1-3.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Available online:. Available online: https://www.who.int/teams/control-of-neglected-tropical-diseases (accessed on 16 September 2024).
- Cabada, M.M.; White Jr., C. A. New developments in epidemiology, diagnosis, and treatment of fascioliasis. Curr. Opin. Infect. Dis. 2012, 25, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Nyindo, M.; Lukambagire, A. Fascioliasis: An Ongoing Zoonotic Trematode Infection. Biomed. Res. Int. 2015, 2015, 786195. [Google Scholar] [CrossRef]
- Rufino-Moya, P.J.; Zafra, R.; Martínez-Moreno, A.; Buffoni, L.; Valderas, E.; Pérez, J.; Molina-Hernández, V.; Ruiz-Campillo, M.T.; Herrera-Torres, G. , Martínez-Moreno, F.J. Advancement in Diagnosis, Treatment, and Vaccines against Fasciola hepatica: A Comprehensive Review. Pathogens 2024, 13, 669. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, K.; Zhang, H.; Sabir, A.J.; Abbas, R.Z.; Ijaz, M.; Durrani, A.Z.; Saleem, M.H.; Rehman, M.U. , Iqbal, M.K.; Wang, Y.; et al. A review on epidemiology, global prevalence and economical losses of fasciolosis in ruminants. Microb. Pathog. [CrossRef]
- Kamaludeen, J; Grahan-Borwn, J; Stephens, N. ; Miller, J.; Howell, A.; Beesley, N.J.; Hodgkinson J.; Learmount, J; Williams. D. Lack of efficacy of triclabendazole against Fasciola hepatica is present on sheep farms in three regions of England, and Wales. Vet. Rec. 2019, 184, 502. [Google Scholar] [CrossRef] [PubMed]
- Kelley, J.M.; Elliot, T.P.; Beddoe, T.; Anderson, G.; Skuce, P.; Spithill, T.W. Current Threat of Triclabendazole Resistance in Fasciola hepatica. Trends Parasitol. 2016, 32, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.N.; Winters, A.; Milic, I.; Devitt, A.; Cookson, A.; Brophy, P.M.; Morphew, R.M. Evidence of sequestration of triclabendazole and associated metabolites by extracellular vesicles of Fasciola hepatica. Sci. Rep. 2020, 10, 13445. [Google Scholar] [CrossRef]
- Beesley, N.J.; Cwiklinski, K.; Allen, K.; Hoyle, R.C.; Spithill, T.W.; La Course, E.J.; Williams, D.J.L.; Paterson, S; Hodgkinson, J. E. A major locus confers triclabendazole resistance in Fasciola hepatica and shows dominant inheritance. PLoS Pathog. 2023, 19, e1011081. [Google Scholar] [CrossRef]
- Lalor, R; Cwiklinski, K; Davies Calvani, N. E.; Dorey, A.; Hamon, S.; López Corrales, J.; Dalton, J.P.; De Marco Verissimo, C. Pathogenicity and virulence of the liver flukes Fasciola hepatica and Fasciola gigantica that cause the zoonosis Fasciolosis. Virulence 2021, 12, 2839–2867. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Human and Animal Fascioliasis: Origins and Worldwide Evolving Scenario. Clin. Microbiol. Rev. 2022, 35, e0008819. [Google Scholar] [CrossRef]
- Cwiklinski, K.; de la Torre-Escudero, E.; Trelis, M.; Bernal, D.; Dufresne, P.J.; Brennan, G.P.; O’Neill, S.; Tort, J.; Paterson, S.; Marcilla, A.; et al. The Extracellular Vesicles of the Helminth Pathogen, Fasciola hepatica: Biogenesis Pathways and Cargo Molecules Involved in Parasite Pathogenesis. Mol. Cell. Proteomics 2015, 14, 3258–3257. [Google Scholar] [CrossRef]
- Durey, C.; Maizels, R.M. Helminth extracellular vesicles: Interactions with the host immune system. Mol. Immunol. 2021, 137, 124–133. [Google Scholar] [CrossRef]
- Álvarez-Rojas, C.A.; Ansell, B.R.E.; Hall, R.S.; Gasser, R.B.; Young, N.D.; Jex, A.R.; Scheerlinck, J.P.Y. Transcriptional analysis identifies key genes involved in metabolism, fibrosis/tissue repair and the immune response against Fasciola hepatica in sheep liver. Parasites & Vectors 2015, 8, 124. [Google Scholar] [CrossRef]
- Young, N.D.; Hall, R.S.; Jex, A.E.; Cantacessi, C.; Gasser, R.B. Elucidating the transcriptome of Fasciola hepatica – A key to fundamental and biotechnological discoveries for a neglected parasite. Biotechnol. Adv. 2010, 28, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Dalton, J.P.; Dufresne, P.J.; La Course, J.; Williams, D.J.; Hodgkinson, J.; Paterson, S. The Fasciola hepatica genome: gene duplication and polymorphism reveals adaptation to the host environment and the capacity for rapid evolution. Genome Biol. [CrossRef]
- Cwiklinski, K.; Dalton, J.P. Advances in Fasciola hepatica research using ‘omics’ technologies. Int. J. Parasitol. 2018, 48, 321–331. [Google Scholar] [CrossRef]
- Ricafrente, A.; Cwiklinski, K.; Nguyen, H.; Dalton, J.P.; Tran, N.; Donnelly, S. Stage-specific miRNAs regulate gene expression associated with growth, development and parasite-host interaction during the intra-mammalian migration of the zoonotic helminth parasite Fasciola hepatica. BMC Genomics, 2022, 23, 419. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Tripathi, S.; Singh, A.K.; Mondal, P.; Mishra, A; Prasad, A. Micromanagement of Immune System: Role of miRNAs in Helminthic Infections. Front. Microbiol. 2017, 8, 586. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Ricafrente, A.; Cwiklinski, K.; Sais, D.; Dalton, J.P.; Tran, N.; Donnelly, S. Exploring the utility of circulating miRNAs as diagnostic biomarkers of fasciolosis. Sci. Rep. 2024, 14, 7431. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: an expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Anglicheau, D.; Muthukumar, T.; Suthanthiran, M. MicroRNAs: small RNAs with big effects. Transplantation 2010, 90, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Vimal, D.; Sharma, D.; Rai, V.; Gupta, S.C.; Chowdhuri, D.K. Role of miRNAs in development and disease: Lessons learnt from small organisms. Life Sci. 2017, 185, 8–14. [Google Scholar] [CrossRef]
- Almeida, M.I.; Reins, R.M.; Calin, G.A. MicroRNA history: Discovery, recent applications, and next frontiers. Mutat. Res. 2011, 717, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Pirela, M; Andrade-Alviárez, D. ; Medina, L.; Castillo, C., Liempi, A., Guerrero-Muñoz, J, Ortega, Y, Maya, J.D, Rojas, V, Quiñones, W., Eds.; et al. MicroRNAs: master regulators in host-parasitic protist intertactions. Open. Biol. 2022, 12, 210395. [Google Scholar] [CrossRef]
- Rojas-Pirela, M.; Andrade-Alviárez, D.; Quiñones, W.; Rojas, M.V.; Castillo, C.; Liempi, A.; Medina, L.; Guerrero-Muñoz, J.; Fernández-Moya, A.; Ortega, Y.A. , et al. microRNAs: Critical Players during Helminth Infections. Microorganisms 2022, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S. The microRNA Registry. Nucleic Acids Res. 2004, 32, 109–111. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, 152–157. [Google Scholar] [CrossRef]
- Backes, C.; Fehlmann, T.; Kern, F.; Kehl, T.; Lenhof, H.; Meese, E.; Keller, A. miRCarta: a central repository for collecting miRNA candidates. Nucleic Acids Res. 2018, 46, D160–D167. [Google Scholar] [CrossRef]
- Bourdon, C.; Bardou, P.; Aujean, E.; Le Guillou, S.; Tosser-Klopp, G.; Le Provost, F. RumimiR: a detailed microRNA database focused on ruminant species. Database (Oxford). 2019, 2019, baz099. [Google Scholar] [CrossRef]
- Fromm, B.; Høye, E.; Domanska, D.; Zhong, X.; Aparicio-Puerta, E.; Ovchinnikov, V.; Umu, S.U.; Chabot, P.J.; Kang, W.; Aslanzadeh, M.; et al. MirGeneDB 2.1: toward a complete sampling of all major animal phyla. Nucleic Acids Res. 2021, 50, D204–D210. [Google Scholar] [CrossRef]
- Xu, M.; Ai, L.; Fu, J.; Nisbet, A.J.; Liu, Q.; Chen, M.; Zhou, D.; Zhu, X. Comparative characterization of microRNAs from the liver flukes Fasciola gigantica and F. hepatica. PLoS One 2012, 7, e53387. [Google Scholar] [CrossRef]
- Tran, N.; Ricafrente, A.; To, J.; Lund, M; Marques, T. M.; Gama-Carvalho, M.; Cwiklinski, K.; Dalton, J.P.; Donnely, S. Fasciola hepatica hijacks host macrophages miRNA machinery to modulate early innate immune responses. Sci. Rep. 2021, 11, 6712. [Google Scholar] [CrossRef] [PubMed]
- Britton, C.; Winter, A.D.; Marks, N.D.; Gu, H.; McNeilly, T.N.; Gillan, V.; Devaney, E. Application of small RNA technology for improved control of parasitic helminths. Vet. Parasitol. 2015, 212, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Ricafrente, A.; Nguyen, H.; Tran, N.; Donnelly, S. An Evaluation of the Fasciola hepatica miRnome Predicts a Targeted Regulation of Mammalian Innate Immune Responses. Front. Immunol. 2021, 11, 608686. [Google Scholar] [CrossRef] [PubMed]
- Fontenla, S.; Langleib, M.; de la Torre-Escudero, E.; Domínguez, M.F.; Robinson, M.W.; Tort, J. Role of Fasciola hepatica Small RNAs in the Interaction With the Mammalian Host. Front. Cell. Infect. Microbiol. 2022, 11, 812141. [Google Scholar] [CrossRef]
- Karapetyan, A.R.; Buiting, C.; Kuiper, R.A.; Coolen, M.W. Regulatory Roles for Long ncRNA and mRNA. Cancers 2013, 5, 462–490. [Google Scholar] [CrossRef]
- Ruvkun, G.; Giusto, J. The Caenorhabditis elegans heterochromic gene lin-14 encodes a nuclear protein that forms a temporal developmental switch. Nature 1989, 338, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementary to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Davis, R.E.; Parra, A.; LoVerde, P.T.; Ribeiro, E.; Glorioso, G.; Hodgson, S. Transient expression of DNA and RNA in parasitic helminths by using particle bombardment. Proc. Natl. Acad. Sci. U S A. 1999, 96, 8687–8692. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Müller, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature, 2000, 403, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci U S A. 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- Garzón, R.; Volinia, S.; Liu, C.; Fernandez-Cymering, C.; Palumbo, T.; Pichiorri, F.; Fabri, M.; Coombes, K.; Alder, H.; Nakamura, T. MicroRNA signatures associated with cytogenetics and prognosis in acute myeloid leukemia. Blood 2008, 111, 3183–3189. [Google Scholar] [CrossRef]
- Jogen-Lavrencic, M.; Sun, S.M.; Dijkstra, M.K.; Valk, P.J.M.; Löwenberg, B. MicroRNA expression profiling in relation to the genetic heterogeneity of acute myeloid leukemia. Blood 2008, 111, 5078–5085. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Radmark, O.; Kim, S.; Kim, V.N. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. The microRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRbase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Issa, Z.; Grant, W.N.; Stasiuk, S.; Shoemaker, C.B. Development of methods for RNA interference in the sheep gastrointestinal parasite, Trichostrongylus colubriformis. Int. J. Parasitol. 2005, 35, 935–940. [Google Scholar] [CrossRef]
- Cheng, G.; Lin, J.; Jin, Y.; Fu, Z.; Jin, Y.; Zhou, Y.; Cai, Y. Dose-dependent inhibition of gynecophoral canal protein gene expression in vitro in the schistosome (Schistosoma japonicum) by RNA interference. Acta Biochim. Biophys. Sin. (Shangai) 2005, 37, 386–390. [Google Scholar] [CrossRef] [PubMed]
- McGonigle, L.; Mousley, A.; Marks, N.J.; Brennan, G.P.; Dalton, J.P.; Spithill, T.W.; Day, T.A.; Maule, A.G. The silencing of cysteine proteases in Fasciola hepatica newly excysted juveniles using RNA interference reduces gut penetration. Int. J. Parasitol. 2008, 38, 149–155. [Google Scholar] [CrossRef]
- Shao, H.; Li, X.; Nolan, T.J.; Massey Jr, H.C.; Pearce, E.J.; Lok, J.B. Transposon-mediated chromosomal integration of transgenes in the parasitic nematode Strongyloides ratti and establishment of stable transgenic lines. PLoS Pathog. 2012, 8, e1002871. [Google Scholar] [CrossRef] [PubMed]
- Gang, S.S.; Castelletto, M.L.; Bryant, A.S.; Yang, E.; Mancuso, N.; Lopez, J.B.; Pellegrini, M.; Hallem, E.A. Targeted mutagenesis in a human-parasitic nematode. PLoS Pathog. 2017, 13, e1006675. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dou, M.; Song, X.; Dong, Y.; Liu, S.; Liu, H.; Tao, J.; Li, W.; Yin, X.; Xu, W. The emerging role of the piRNA/piwi complex in cancer. Mol. Cancer 2019, 18, 123. [Google Scholar] [CrossRef]
- Available online:. Available online: https://www.nobelprize.org/prizes/medicine/2024/press-release/ (accessed on 4 November 2024).
- Weiss, C.N.; Ito, K. A. Macro View of MicroRNAs: The Discovery of MicroRNAs and Their Role in Hematopoiesis and Hematologic Disease. Int. Rev. Cell Mol. Biol. 2017, 334, 99–175. [Google Scholar] [CrossRef]
- Quinzo, M.J.; Perteguer, M.J.; Brindley, P.J.; Loukas, A.; Sotillo, J. Transgenesis in parasitic helminths: a brief history and prospects for the future. Parasit Vectors 2022, 15, 110. [Google Scholar] [CrossRef]
- Liu, B.; Shyr, Y.; Cai, J.; Liu, Q. Interplay between miRNAs and host genes and their role in cancer. Brief Func. Genomics 2019, 18, 155–266. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.J.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature. [CrossRef]
- Saliminejad, K.; Khorshid, H.R.K.; Fard, S.S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Guasconi, L.; Serradell, M.C.; Garro, A.P.; Iaconelli, L.; Masih, D.T. C-type lectins on macrophages participate in the immunomodulatory response to Fasciola hepatica products. Immunology 2011, 133, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Campillo, M.T.; Molina-Hernández, V.; Pérez, J.; Pacheco, I.L.; Pérez, R.; Escamilla, A.; Martínez-Moreno, F.J.; Martínez-Moreno, A.; Zafra, R. Study of peritoneal macrophage immunophenotype in sheep experimentally infected with Fasciola hepatica. Vet. Parasitol. 2018, 257, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.P.; Robinson, M.W.; Mulcahy, G.; O’Neill, S.M.; Donnelly, S. Immunomodulatory molecules of Fasciola hepatica: candidates for both vaccine and immunotherapeutic development. Vet. Parasitol. 2013, 195, 272–285. [Google Scholar] [CrossRef]
- Santiago-Figueroa, O.; Espino, A.M. Fasciola hepatica Fatty Acid Binding Protein Induces the Alternative Activation of Human Macrophages. Infect. Immun. 2014, 82, 5005–5012. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Campillo, M.T.; Barrero-Torres, D.M. , Abril, N.; Pérez, J.; Zafra, R.; Buffoni, L.; Martínez-Moreno, Á.; Martínez-Moreno, F.J.; Molina-Hernández, V. Fasciola hepatica primoinfections and reinfections in sheep drive distinct Th1/Th2/Treg immune responses in liver and hepatic lymph node at early and late stages. Vet. Res. 2023, 54, 2. [Google Scholar] [CrossRef]
- Zheng, Y.; Cai, X.; Bradley, J.E. microRNAs in parasites and parasite infection. RNA Biol. 2013, 10, 371–379. [Google Scholar] [CrossRef]
- Cucher, M.A.; Ancarola, M.E.; Kamenetzky, L. The challenging world of extracellular RNAs of helminth parasites. Mol. Immunol. 2021, 234, 150–160. [Google Scholar] [CrossRef]
- Alizadeh, Z.; Mahami-Oskouei, M.; Spotin, A.; Kazemi, T.; Ahmadpour, E.; Cai, P.; Shanehbandi, D.; Shekari, N. Parasite-derived microRNAs in plasma as novel promising biomarkers for the early detection of hydatid cyst infection and post-surgery follow-up. Acta Trop. 2020, 202, 105255. [Google Scholar] [CrossRef]
- Britton, C.; Laing, R.; Devaney, E. Small RNAs in parasitic nematodes – forms and functions. Parasitology 2020, 147, 855–864. [Google Scholar] [CrossRef]
- Fromm, B.; Trelis, M.; Hackenberg, M.; Cantalapiedra, F.; Bernal, D.; Marcilla, A. The revised microRNA complement of Fasciola hepatica reveals a plethora of overlooked microRNAs and evidence for enrichment of immune-regulatory microRNAs in extracellular vesicles. Int. J. Parasitol. 2015, 45, 697–702. [Google Scholar] [CrossRef]
- Murphy, A.J.; Guyre, P.M.; Pioli, P.A. Estradiol suppresses NF-kappa B activation through coordinated regulation of let-7a and miR-125b in primary human macrophages. J. Immunol. 2010, 184, 5029–5037. [Google Scholar] [CrossRef] [PubMed]
- Duroux-Richard, I.; Roubert, C.; Ammari, M.; Présumey, J.; Grün, J.R.; Häupl, T.; Grützkau, A.; Lecellier, C.; Boitez, V.; Codogno, P.; et al. miR-125b controls monocyte adaptation to inflammation through mitochondrial metabolism and dynamics. Blood 2016, 128, 3125–3136. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.A.; So, A.Y.; Sinha, N.; Gibson, W.S.J.; Taganov, K.D.; O’Connell, R.M.; Baltimore, D. MicroRNA-125b potentiates macrophage activation. J. Immunol. 2011, 187, 5062–5068. [Google Scholar] [CrossRef]
- Fontenla, S.; Dell’Oca, N.; Smircich, P.; Tort, J.F.; Siles-Lucas, M. The miRnome of Fasciola hepatica juveniles endorses the existence of a reduced set of highly divergent micro RNAs in parasitic flatworms. Int. J. Parasitol. 2015, 45, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Hoy, A.M.; Lundie, R.J.; Ivens, A.; Quintana, J.F.; Nausch, N.; Forster, T.; Jones, F.; Kabatereine, N.B. Parasite-Derived MicroRNAs in Host Serum As Novel Biomarkers of Helminth Infection. PLoS Negl. Trop. Dis. 2014, 8, e2701. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Ambros, V.R. Caenorhabditis elegans microRNAs of the let-7 family act in innate immune response circuits and confer robust developmental timing against pathogen stress. Proc. Natl. Acad. Sci. U S A 2015, 112, E2366–E2375. [Google Scholar] [CrossRef]
- Stark, A.; Brennecke, J.; Russell, R.B.; Cohen, S.M. Identification of Drosophila MicroRNA targets. PLoS Biol. 2003, 1, E60. [Google Scholar] [CrossRef]
- Rubio, M.; Belles, X. Subtle roles of microRNAs let-7, miR-100 and miR-125 on wing morphogenesis in hemimetabolan metamorphosis. J. Insect. Physiol. 2013, 59, 1089–1094. [Google Scholar] [CrossRef]
- Sun, X.; Jiao, X.; Pestell, T.G.; Fan, C.; Qin, S.; Mirabelli, E.; Ren, H.; Pestell, R.G. MicroRNAs and cancer stem cells: The sword and the shield. Oncogene 2014, 33, 4967–4977. [Google Scholar] [CrossRef]
- Banerjee, S.; Xie, N.; Cui, H.; Tan, Z.; Yang, S.; Icyuz, M.; Abraham, E.; Liu, G. MicroRNA let-7c regulates macrophage polarization. J. Immunol. 2013, 190, 6542–6549. [Google Scholar] [CrossRef] [PubMed]
- Boulan, L.; Martín, D.; Milán, M. Bantam miRNA promotes systemic growth by connecting insulin signaling and ecdysone production. Curr. Biol. 2013, 23, 473–478. [Google Scholar] [CrossRef]
- Xue, X.; Sun, J.; Zhang, Q. Wanf, Z.; Huang, Y.; Pan, W. Identification and characterization of novel microRNAs from Schistosoma japonicum. PLoS One 2008, 3, e4034. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, L.; Wang, J.; Qiu, L.; Chen, Y.; Davis, R.E.; Cheng, G. Schistosoma japonicum extracellular vesicle miRNA cargo regulates host macrophage functions facilitating parasitism. PLoS Pathog. 2019, 15, e1007817. [Google Scholar] [CrossRef]
- Lopes-Junior, E.; Bertevello, C.; Silveira, G.; Guedes, C.; Rodrigues, G.; Ribeiro, V.; Amaral, M.; Kanamura, C.; Pinto, P.; Krüger, R.; et al. Human tumor necrosis factor alpha affects the egg-laying dynamics and glucose metabolism of Schistosoma mansoni adult worms in vitro. Parasit. Vectors 2022, 15, 176. [Google Scholar] [CrossRef]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat. Commun. 2014, 5, 5488. [Google Scholar] [CrossRef]
- Wu, J.; Liao, Y.; Li, D.; Zhu, Z.; Zhang, L.; Wu, Z.; He, P.; Wang, L. Extracellular vesicles derived from Trichinella Spiralis larvae promote the polarization of macrophages to M2b type and inhibit the activation of fibroblasts. Front. Immunol. 2022, 13, 974322. [Google Scholar] [CrossRef] [PubMed]
- Fromm, B.; Ovchinnikov, V.; Høye, E.; Bernal, D.; Hackenberg, M.; Marcilla, A. On the presence and immunoregulatory functions of extracellular microRNAs in the trematode Fasciola hepatica. Parasite Immunol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Montaner, S.; Galiano, A.; Trelis, M.; Martin-Jaular, L.; del Portillo, H.A.; Bernal, D.; Marcilla, A. The Role of Extracellular Vesicles in Modulating the Host Immune Response during Parasitic Infections. Front. Immunol. 2014, 5, 433. [Google Scholar] [CrossRef]
- Guo, X.; Guo, A. Profiling circulating microRNAs in serum of Fasciola gigantica-infected buffalo. Mol. Biochem. Parasitol. 2019, 232, 111201. [Google Scholar] [CrossRef]
- Ricciardi, A.; Bennuru, S.; Tariq, S.; Kaur, S.; Wu, W.; Elkahloun, A.G.; Arakelyan, A.; Shaik, J.; Dorward, D.W.; Nutman, T.B.; Semnani, R.T. Extracellular vesicles released from the filarial parasite Brugia malayi downregulate the host m TOR pathway. PLoS Negl. Trop. Dis. 2021, 15, e0008884. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Guo, X.; He, W.; Shao, Z.; Zhang, X.; Yang, J.; Shen, Y.; Luo, X.; Cao, J. Effects of Echinococcus multilocularis miR-71 mimics on murine macrophage RAW264.7 cells. Int. Immunopharmacol. 2016, 34, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Robb, E.; McCammick, E.M.; Wells, D.; McVeigh, P.; Gardiner, E.; Armstrong, R.; McCusker, P.; Mousley, A.; Clarke, N.; Marks, N.J.; et al. Transcriptomic analysis supports a role for the nervous system in regulating growth and development of Fasciola hepatica juveniles. PLoS Negl. Trop. Dis. 2022, 16, e0010854. [Google Scholar] [CrossRef]
- Shi, W.; He, J.; Mei, X.; Lu, K.; Zeng, Z.; Zhang, Y.; Sheng, Z.; Elsheikha, H.M.; Huang, W.; Zhu, X. Dysregulation of hepatic microRNA expression in C57BL/6 mice affected by excretory-secretory products of Fasciola gigantica. PLoS Negl. Trop. Dis. 2020, 14, e0008951. [Google Scholar] [CrossRef]
- Saravanan, S.; Thirugnanasambantham, K.; Hanieh, H.; Karikalan, K.; Sekar, D.; Rajagopalan, R.; Islam, V.I.H. miRNA-24 and miRNA-466I-5p controls inflammation in rat hepatocytes. Cell. Mol. Immunol. 2014, 12, 113–115. [Google Scholar] [CrossRef]
- El-Guendy, N.M.; Helwa, R.; El-Halawany, M.S.; Ali, S.A.R.; Aly, M.T.; Alieldin, N.H.; Fouad, S.A.H.; Saeid, H.; Abdel-Wahab, A.A. The Liver MicroRNA Expression Profiles Associated With Chronic Hepatitis C Virus (HCV) Genotype-4 Infection: A Preliminary Study. Hepat. Mon. 2016, 16, e33881. [Google Scholar] [CrossRef] [PubMed]
- Guidi, R.; Wedeles, C.J.; Wilson, M.S. ncRNAs in Type-2 Immunity. Noncoding RNA 2020, 6, 10. [Google Scholar] [CrossRef]
- Ovchinnikov, V.Y. Kashina, E.V.; Mordninov, V.A.; Fromm, B. EV-transported microRNAs of Schistosoma mansoni and Fasciola hepatica: Potential targets in definitive hosts. Infect. Genet. Evol. 2020, 85, 104528. [Google Scholar] [CrossRef]
- Kimura, K.; Hohjoh, H.; Fukuoka, M.; Sato, W.; Oki, S.; Tomi, C.; Yamaguchi, H.; Kondo, Takahashi, R. ; Yamamura, T. Circulating exosomes suppress the induction of regulatory T cells via let-7i in multiple sclerosis. Nat. Commun, 2018, 9, 17. [Google Scholar] [CrossRef]
- Hu, R.; Zhang, X.; Ma, Q.; Elsheikha, H.M.; Ehsan, M.; Zhao, Q.; Fromm, B.; Zhu, X. Differential expression of microRNAs and t RNA fragments mediate the adaptation of the liver fluke Fasciola gigantica to its intermediate snail and definitive mammalian hosts. Int. J. Parasitol. 2021, 51, 405–414. [Google Scholar] [CrossRef]
- Wang, S.; Chen, D.; He, J.; Zheng, W.; Tian, A.; Zhao, G.; Elsheikha, H.M.; Zhu, X. Fasciola gigantica-Derived Excretory-Secretory Products Alter the Expression of mRNAs, miRNAs, lncRNAs, and circRNAs Involved in the Immune Response and Metabolism in Goat Peripheral Blood Mononuclear Cells. Front. Immunol. 2021, 12, 653755. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, C.M.; Trelis, M.; Jara, L.; Cantalapiedra, F.; Marcilla, A.; Bernal, D. Diversity of extracellular vesicles from different developmental stages of Fasciola hepatica. Int. J. Parasitol. 2020, 50, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Herron, C.M.; O’Connor, A.; Robb, E.; McCammick, E.; Hill, C.; Marks, N.; Robinson, M.W.; Maula, A.G.; McVeigh, P. Developmental Regulation and Functional Prediction of microRNAs in an Expanded Fasciola hepatica miRNome. Front. Cell Infect. Microbiol. 2022, 12, 811123. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfus, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A uniform system for microRNA annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef]
- Cai, P.; Hou, N.; Piao, X.; Liu, S.; Liu, H.; Yang, F.; Wang, J.; Jin, Q.; Wang, H.; Chen, Q. Profiles of small non-coding RNAs in Schistosoma japonicum during development, PLoS Negl. Trop. Dis. 2011, 5, e1256. [Google Scholar] [CrossRef]
- Macchiaroli, N.; Cucher, M.; Kamenetzky, L.; Yones, C.; Bugnon, L.; Berriman, M.; Olson, P.D.; Rosenzvit, M.C. Identification and expression profiling of microRNAs in Hymenolepis. Int. J. Parasitol. 2019, 49, 2111–2223. [Google Scholar] [CrossRef]
- McVeigh, P.; McCammick, E.; Robb, E.; Brophy, P.; Morphew, R.M.; Marks, N.J.; Maule, A.G. Discovery of long non-coding RNAs in the liver fluke, Fasciola hepatica. PLoS Negl. Trop. Dis. 2023, 17, e0011663. [Google Scholar] [CrossRef]
- Ulitsky, I. Interactions between short and long noncoding RNAs. FEBS Lett. 2018, 592, 2874–2883. [Google Scholar] [CrossRef]
- Zhang, M.; He, P.; Bian, Z. Long Noncoding RNAs in Neurodegenerative Diseases: Pathogenesis and Potential Implications as Clinical Biomarkers. Front. Mol. Neurosci. 2021, 14, 685143. [Google Scholar] [CrossRef]
- He, M.; Wan, J.; Zeng, H.; Tang, Y. , He, M. miR-133a-5p suppresses gastric cancer through TCF4 down-regulation. J. Gastrointest. Oncol. 2021, 12, 1007–1019. [Google Scholar] [CrossRef]
- Shandilya, U.K.; Sharma, A.; Naylor, D.; Canovas, A.; Mallard, B.; Karrow, N.A. Expression Profile of miRNA from High, Middle and Low Stress-Responding Sheep during Bacterial Endotoxin Challenge. Animals 2023, 13, 508. [Google Scholar] [CrossRef]
- Bąska, P.; Majewska, A.; Zygner, W.; Długosz, E.; Wiśniewski, M. Fasciola hepatica Excretory-Secretory Products (Fh-ES) Either Do Not Affect miRNA Expression Profile in THP-1 Macrophages or the Changes Are Undetectable by a Microarray Technique. Pathogens 2024, 13, 854. [Google Scholar] [CrossRef]
- Batan Pumeda, S. Comparison Between Next-Generation Sequencing and Microarrays for miRNA Expression in Cancer Samples. Natl. Acad. Sci. Lett. 2023. [Google Scholar] [CrossRef]
- Sais, D.; Chowdhury, S.; Dalton, J.P.; Tran, N.; Donnelly, S. Both host and parasite non-coding RNAs co-ordinate the regularion of macrophage gene expression to reduce pro-inflammatory immune responses and promote tissue repair pathways during infection with Fasciola hepatica. RNA Biol. 2024, 21, 62–77. [Google Scholar] [CrossRef]
- Alhallaf, R.; Agha, Z.; Miller, C.M.; et al. The NLRP3 inflammasome suppresses protective immunity to gastrointestinal helminth infection. Cell Rep. 2018, 23, 1085–1098. [Google Scholar] [CrossRef]
- Guan, F.; Jiang, W.; Bai, Y.; et al. Purinergic P2X7 receptor mediates the elimination of Trichinella spiralis by activating nf-κB/NLRP3/IL-1β pathway in macrophages. Infect Immun. 2021, 89, e00683–20. [Google Scholar] [CrossRef]
- De la Torre-Escudero, E.; Gerlach, J.Q.; Bennet, A.P.S.; Cwiklinski, K.; Jewhurst, H.L.; Huson, K.M.; Joshi, L.; Kilcoyne, M.; O’Neill, S.; Dalton, J.P.; Robinson, M.W. Surface molecules of extracellular vesicles secreted by the helminth pathogen Fasciola hepatica direct their internalisation by host cells. PLoS Negl. Trop. Dis. 2019, 13, e0007087. [Google Scholar] [CrossRef]
Figure 2.
Representation of miRNA biogenesis in Fasciola hepatica through the canonical pathway.
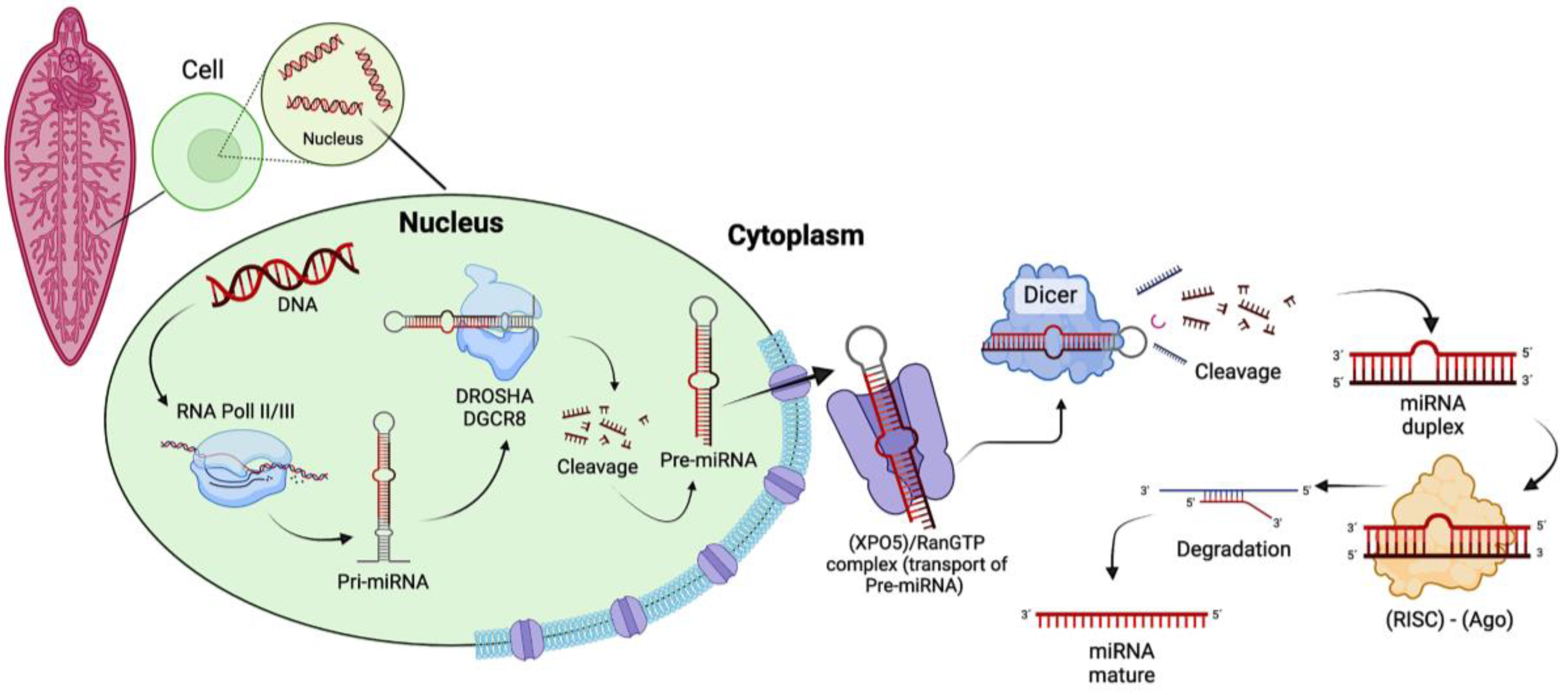
Figure 3.
Overview of the miRNome in Fasciola spp. based on the latest miRNome reports for F. hepatica [99,109] and F. gigantica [106] displaying the nomenclature based on the annotation of miRNAs in databases [110]. A. Represents the 38 miRNAs annotated in miRBase for F. hepatica with the 3p/5p sequences reported to date (grey), the homologous sequences with other species/organisms annotated in miRBase that are not annotated for F. hepatica (blue), and miRNAs without homologous sequences to any of the miRNAs from other species/organisms annotated in miRBase, considered as specific for F. hepatica without annotation in miRBase (orange). B. Represents F. gigantica miRNAs homologous with miRNAs annotated for F. hepatica in miRBase (purple), miRNAs from F. gigantica that have homology with miRNAs from other species/organisms annotated in miRBase (blue), miRNAs reported within the F. gigantica studies without sequences to help identify them (red), and and F. gigantica miRNAs with homologous sequences in miRNAs specific for F. hepatica without being annotated in miRBase (green).
Figure 3.
Overview of the miRNome in Fasciola spp. based on the latest miRNome reports for F. hepatica [99,109] and F. gigantica [106] displaying the nomenclature based on the annotation of miRNAs in databases [110]. A. Represents the 38 miRNAs annotated in miRBase for F. hepatica with the 3p/5p sequences reported to date (grey), the homologous sequences with other species/organisms annotated in miRBase that are not annotated for F. hepatica (blue), and miRNAs without homologous sequences to any of the miRNAs from other species/organisms annotated in miRBase, considered as specific for F. hepatica without annotation in miRBase (orange). B. Represents F. gigantica miRNAs homologous with miRNAs annotated for F. hepatica in miRBase (purple), miRNAs from F. gigantica that have homology with miRNAs from other species/organisms annotated in miRBase (blue), miRNAs reported within the F. gigantica studies without sequences to help identify them (red), and and F. gigantica miRNAs with homologous sequences in miRNAs specific for F. hepatica without being annotated in miRBase (green).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



