
Submitted:
21 November 2024
Posted:
22 November 2024
You are already at the latest version
Abstract
Petroleum-based plastics are recalcitrant world-wide used materials that severely pollute the environment, thus biodegradable bioplastics are emerging as a viable alternative. From this group, the study of polyhydroxyalkanoates (PHAs) has stood out for their potential in diverse applications including medicine, packaging and agriculture. The enzyme responsible for PHAs synthesis inside the microbial cell is the PHA synthase (PhaC). PhaCs are present in a wide variety of microorganisms and are classified according to their substrate specificity and subunit composition into 4 classes. Class I, class III and class IV use the acyl-CoA as a precursor to synthesize short-chain-length PHAs while Class II enzymes use an intermediate of the β-oxidation pathways to synthesize medium-chain-length PHAs. Enzymes from this pathway that have been upregulated and downregulated to optimize PHAs production are described in this review. Another important enzyme is the PHA depolymerase (PhaZ) which is responsible for all PHA degradation inside and outside the cell. This review describes both enzymes in detail, including classification, structure, substrate specificity and proven protein engineering techniques for enzymatic rate enhancement and modified substrate specificity of the proteins. It also includes a mutation map for the class II PhaC sequence of Pseudomonas putida that suggest point mutations for future protein engineering work.
Keywords:
Polyhydroxyalkanoates (PHAs)
; bioplastics
; PHA synthase (PhaC)
; PHA depolymerase (Z)
; plastic pollution
; protein engineering
Introduction
Polyhydroxyalkanoates (PHAs) are biopolymers classified as polyesters which many bacteria and archaea accumulate in the form of intracellular granules as a reservoir of energy and carbon in response to stress conditions, especially when nitrogen is limited [1,2,3,4]. Maurice Lemoigne was the first scientist to isolate and characterize poly-3-hydroxybutyrate (P3HB), the simplest type of PHA, as a storage compound from Bacillus megaterium in 1926 [2,5]. P3HB is a polymer composed of 4 carbon-chain-length repeating monomers, and it is the most common and abundant PHA in bacteria [6,7]. PHAs are a good source of chemicals, polymers and biofuels [8].
Since this discovery, almost 100 years ago, thousands of papers have been published and some good reviews have been written describing the generalities of PHAs [9], their structure and composition diversity [10], applications in several fields such as: medicine [11], agriculture, biofuels and packaging [12]. There are also reviews describing potential suitable hosts for industrial production (such as Cupriavidus necator, Pseudomonas, recombinant Escherichia coli and yeasts) and their metabolic pathways [13].
This review re-capitulates some of the information mentioned in these reviews, [14,15] for example, main advantages and disadvantages of producing PHAs and the main metabolic pathway (β-oxidation pathway) is shown again [16] and expands to the specific genes that have been knockout and/or overexpressed from this pathway to enhance PHAs accumulation. It also compiles different methods for protein engineering and enzymatic evaluation as well as classification, structure and substrate specificity of both PhaC and PhaZ enzymes.
The PHA depolymerase (PhaZ) enzyme is responsible of depolymerizing the bioplastic and can be a useful tool for bioremediation of PHAs. Although, there are a volume of studies on PhaZ, for example the PHA Depolymerase Engineering Database published by Knoll, et al. [17], this volume is by far less than the volume of papers published on PhaC and review papers are greatly lacking. The reason PhaZ has been less studied than PhaC is because there isn’t sufficient need for PHAs bioremediation as PHAs accumulation in the environment is still not a problem [18].
Another novelty of this work is the construction of a mutation map for the amino acid sequence of the PHA synthase (PhaC) from Pseudomonas putida. The purpose of the map is to show several potential amino acid change mutations that exist already in nature and that could be used to change the specificity/activity of a target sequence so that polymers with different monomer compositions could be created. The map is also accompanied by a table of single amino acid mutations that have already been studied and their effect over the enzyme.
Bioplastics
Bioplastics are a growing class of materials that can be used as an alternative to petroleum-based plastic [19]. All bioplastics are produced by living organisms from renewable feedstock. However, not all bioplastics are biodegradable [19]. The main types of bioplastics are illustrated in Figure 1. Bioplastics have been studied for more than a century and yet, their industrialization is still embryonic [20]. According to the report of the European Bioplastics, bioplastics currently (2023) represent roughly 0.5% percent of the over 400 million tonnes of plastic produced annually [21].
The main advantage of bioplastics is that they have a faster biodegradation rate than petroleum-based plastics in a wide range of environments. Common petroleum-based plastics such as polyethylene, (PE) polystyrene (PS) and polyethylene terephthalate (PET) have a lifespan of 10-600, 50-80 and up to 450 years depending on the conditions [40,41] while a PHA bottle is estimated to degrade in only 1.5-3.5 years in the marine environment [42]. In soil, with the appropriate temperature and moisture levels, PHA can degrade in as little as 7 weeks [43]. PHAs are also inexhaustible type of material which is an advantage over petroleum given that petroleum reserves are expected to run out by the year 2070 [44].
On the other hand, one of the main roadblocks to produce bioplastics is their higher costs. It has been calculated that the minimum selling price of PHAs in 2022 was 4.83 $/kg [45] while the global cost of, for example, high density polyethylene (HDPE) and of polyethylene terephthalate (PET) is ≈ 1 $/kg [46]. The high costs of PHA production comes mainly from the carbon feedstock used for the fermentation (≈38% of the production costs) [47]. Common industrial feedstocks include sugar beet, sugar cane bioproduct, canola oil, Corn or cassava and sucrose [48,49,50,51,52,53,54,55]. Other high costs come from the fermentation itself and from downstream processing [56]
Additional disadvantages include the use of land to grow these feedstocks, and thermophysical properties of some types of these plastics such as thermal instability, brittleness, low melt strength etcetera [57].
Classification, Thermophysical Characteristics and Advantages of PHAs
PHA polymers are classified by their monomer length as short chain length (scl), which normally contain 5 or less than 5 carbons (≤C5), for example poly-3-hydroxyvalerate (P3HV) (P3HB falls in this category as well) and as medium chain length (mcl) which contain in between 6 and 14 carbons (≥C6, ≤C14), for example: poly-3-hydroxyoctanoate (P3HO) and poly-3-hydroxyhexanoate (P3HHX) [58,59,60]. PHA polymers are composed of a backbone and a lateral chain (R) which is normally an alkane chain (Refer Figure 2) [15].
Aside from alkanes, PHAs can have lateral chains containing double or triple bonds and /or include different functional groups such as halogen atoms, methoxy, benzoyl, ethoxy, cyanophenoxy, acetoxy, phenoxy, hydroxyl, nitrophenyl, epoxy, carbonyl, cyano and others [62].
There are PHAs in which the length of the backbone is longer, so the hydroxyl group to be esterified is not in carbon C3. Other PHAs have a thioester group in the place of the oxoester linkage. Some of these PHAs are uncommon and can only be obtained by chemical or physical modification of naturally occurring ones [7,62,63].
PHAs can also be classified as homopolymers and copolymers, for example P(3HB-co-3HV) is a copolymer made up from 3-hydroxybutyrate and 3-hydroxyvalerate monomers. The monomers are usually incorporated randomly across the polymer, thus they are also referred as random co-polymers [64]. This process depends on the carbon substrate, metabolic pathways available and on the specificity of the enzymes involved [65]. Cupriavidus and other organisms with PhaC type I use acyl-CoA from the metabolism of sugars to polymerize scl-PHA [14,66,67,68,69] through the 3 enzymes from the operon phaCAB [65,70] while organisms with PhaC type II, type III and type IV [14,66,67,68,69] use the intermediate 3-hydroxyacyl-CoA from the metabolism of fatty acids (β-oxidation pathway) to produce mcl-PHA [65,70].
Nevertheless, It is also important to mention that, while the natural substrate of PhaC in C. necator is 3HB-CoA, it was able to accept 3-hydroxyoctanoate-CoA and 3-hydroxydodecanoate-CoA to produce medium chain PHA when expressed in E. coli suggesting that the substrate specificity of the different types of PhaCs is less strict than originally thought [71]. This leads us to believe that the type of PHA (scl or mcl) is not only dependent on the substrate specificity of the PhaC but also on the specificity of the other enzymes involved and the metabolic pathway used.
Carbon feedstock is also a major factor defining the polymer composition. For example, Klebsiella spp. can produce P(3HB-co-3HV) with a higher portion of 3HV using soy waste and with a lower portion of 3HV using malt wastes as carbon feedstock, while Alcaligenes latus and Staphylococcus spp. can only produce homopolymers of hydroxybutyrate using these same substrates [72]. In contrast, Cupriavidus necator (Alcaligenes eutrophus) is capable of increasing the proportion of HV inside P(3HB-co-3HV) when NH4OH is present in the medium [73]. Commercial copolymers include P(3HB-co-4HB) produced using Cupriavidus necator by Metabolix, USA, P(3HB-co-3HHx) produced also using C. necator by Kaneka, Japan and using Aeromonas hydrophila by P&G, China and by Shandong, Lukang [55].
In addition to the above classifications, PHAs can be classified as block copolymers. Block copolymers are macromolecules composed of sequences, or blocks, of chemically distinct repeat units. Hence, PHAs block copolymers are composed of different sets of PHA homopolymers linked together. Possible block copolymer structures include A-B diblock, A-B-A, A-B-C triblock, or (AB)n repeating multiblock [74]. It has been demonstrated that alternating between different carbon feedstocks leads to the formation of block copolymers; For instance Cupriavidus necator can synthetize poly(3-hydroxybutyrate)-block-poly(3-hydroxybutyrate-co-3-hydroxyvalerate) P(P3HB-b-P3HBV) when valeric acid containing substrate is alternated with non-valeric acid substrate [75].
This changes on monomer composition affect the overall characteristics of the polymer. Comparison of the thermophysical characteristics between short-chain-length and medium-chain-length polyhydroxyalkanoates are illustrated in Table 1.
PHAs, in general, can be used in medical devices such as suture threads, patches, meshes, implants, tissue engineering scaffolds and controlled drug delivery systems. In particular, due to their elasticity, medium-chain- length PHAs have been reported to be used for soft tissue implants [79]. Moreover, there is evidence showing that the cytotoxicity of polyhydroxyalkanoates is inversely proportional with the length of the side chain of the monomer (longer side chains are less cytotoxic) making medium-chain- length PHAs better candidates for medical applications [80].
Mcl-PHAs are specially utilized for cardiac tissue engineering applications and heart valves because elastic materials are more suitable to be used as leaflets inside the tri-leaflet valve [80]. They also mimic better the mechanical properties of soft nerve tissue [81]. Scaffolds of the co-polymer P(3HB-co-3HHx) compared against scaffolds with scl-PHAs monomers showed stronger potential to promote differentiation of neural stem cells into neurons and therefore showed biomedical potential for repairing the central nervous system [82]. However, studies remain limited mainly to P3HO and the co-polymer P(3HB-co-3HHx) which are the only mcl-PHAs in large quantities [81].
One of the main advantages of PHAs is its high biodegradability compared to petroleum-based plastic. For example, a PHA-made water bottle would be expected to completely degrade in 1.5 to 3.5 years in marine environment [42] while a high-density polyethylene bottle has a marine half-life of 58 years [83]. However they also present some disadvantages such as the high cost of production [84]. Other advantages and disadvantages are listed below:
Advantages
- Less CO2 emissions and sustainability [18].
- The industrial production has a low safety risk compared to petroleum-based plastic production which includes flammable and toxic by-products [18].
- Waste water is non-toxic [18].
Disadvantages
Metabolic Pathways and Metabolic Engineering to Optimize Medium-Chain-Length (mcl) PHAs Cell Production
The monomer carbon chain length composition of mcl-PHAs is directly influenced by the carbon length of the fatty acid provided [85]. In each complete β-oxidation cycle, 2 carbons are removed from the fatty acid chain and cycles will continue to take place in the cell until the fatty acid is completely metabolized, as a result, monomers of different sizes will integrate into the PHA polymer to create a heteropolymer. The difficulty of controlling the polymer’s monomer composition is one of the main challenges of mcl-PHAs scale up production. This difficulty is even greater in Pseudomonas because it has several gene copies for each of these β-oxidation pathway enzymes.
Nevertheless, weakening the β-oxidation pathway, by knocking out one or more copies of one gene, has proven to increase the carbon flux towards enoyl CoA (mcl-PHAs precursor) and avoid the degradation of the fatty acid substrate, which will enhance PHAs accumulation inside the cell [86,87] and result in more homogenous PHAs in terms of monomer composition [87,88].
The most common method to weaken the β-oxidation pathway is through gene knockout of fadB and fadA genes (See Figure 3, Table 2). However, similar results have been achieved when using the β-oxidation pathway inhibitor acrylic acid (See Figure 3, Table 2) [89]. On the other hand, mcl-PHAs accumulation can also be enhanced by overexpressing genes that are related to the β-oxidation pathway such as phaJ, phaG, fabG and alKk (See Figure 3, Table 2) [90,91]. No change in monomer composition is expected to take place nor has been observed when overexpressing these enzymes [90]. However, more research is needed to observe if substrate specificity of these enzymes can also play a role in monomer composition.
More than 5000 species of bacteria distributed in more than 14 bacterial groups including Firmicutes, Bacilli, Clostridia, Actinobacteria, Micrococcal, Streptomycetales, Corynebacterial, Cyanobacteria/Melainabacteria group, Proteobacteria, Alpha proteobacteria, Gamma proteobacteria, Betaproteobacteria, Delta/Epsilon subdivisions and other groups have the capacity of synthetizing PHA [65,69]. Even some and halophilic archaea can accumulate it [105,106].
However, for use in pilot or industrial scale production, reasonably high productivities of PHAs using fed-batch fermentation should exist in a range of (1 g L−1 h−1 to 2 g L−1 h−1) for the process to be economically feasible according to Blunt, et al. [107]. The study of Jiang, et al. [89] using P. putida does fit this criteria and achieves a production of 1.8 g L−1 h−1 but most of the studies presented on the table above (Table 2) have a much lower productivity. For example, PHAs production in the study in which Ralstonia eutropha (Now Cupriavidus necator) accumulated 71% of its dry weight of the copolymer P(HB-co-17%-HHx) equivalates to a final ≈2 g/l after 72 hours of cultivation [93] while in the study of Bhatia, et al. [95] using Cupriavidus necator as well fed with coffee waste achieved a productivity of approximately only 0.7 g/l in 72 hours. This information leads us to believe that some of the studies that already exist are still not good candidates for scale up and industrial production and more optimization work is needed.
Additionally, there are other strains that have potential as industrial mcl-PHA producers and other genetical modifications, different than β-oxidation modifications, are efficient [108]. For example, genes for the consumption of xylose (xylA, xylB and tktA) were overexpressed along with the PhaC from Aeromonas inside the strain Burkholderia sacchari to produce 55.5% CDW of P(HB-co-HHx) [109].
PHA synthase (PhaC)
Classification
PHA synthases are divided into four classes based on their substrate specificity and on their subunit composition. Classes I, III and IV prefer to synthesize scl-PHAs while class II PhaC synthesize mcl-PHAs [14,110]. (see Table 3) Yet, new studies show a volume of exceptions to this classification. For example, PhaC2 from Pseudomonas stutzeri 1317, (type II PHA synthase) can incorporate several short-chain-length and medium-chain-length monomers into PHAs [111]. Chromobacterium sp. USM2, can also utilize 3-hydroxybutyrate (3HB), 3-hydroxyvalerate (3HV), and 3-hydroxyhexanoate (3HHx) monomers [112] while Aeromonas caviae can synthesize the random co-polymer P(3HB-co-3HHx) [1].
Interestingly, the PhaC from Rhodococcus pyridinivorans BSRT1-1 can synthesize the scl-mcl co-polymer P(3HB-co-3HHx) when expressed recombinantly in a C. necator PHA negative strain [113]. The PhaC from the scl-PHA industrial strain C. necator can also broaden the range of monomers that is can take and incorporate mcl monomers into the polymer when expressed recombinantly in E. coli.
These important findings suggest that the PhaC specificity is not as strict as previously thought and that the monomer composition is not only dependent on the PhaC’s substrate specificity but also on the specificity of the other enzymes from the pathway.
Structure and Sequence
PHA synthases belong to the α/β-hydrolase superfamily and have only 8 strictly conserved amino acid residues. This so-called α/β-hydrolase fold consists of a characteristic succession of alpha helices and beta strands, usually found in lipases, where the catalytic residues aspartate, histidine and cysteine are. Out of the catalytic triad, the conserved cysteine residue is used as catalytic nucleophile [14,114].
This cysteine is embedded in the PhaC box sequence (GS)-X-C-X-(GA)-G (X is an arbitrary amino acid), which is similar to the lipase box G-X-S-X-G proper of lipases, in the active site of the catalytic domain of the protein [114,115]. In a posterior amino acid replacement study using PhaC from Cupriavidus necator the PhaC box sequence was proven to be still active when expanded to ([GAST]-X-C-X-[GASV]-[GA]). This study also shows the low mutational robustness of the last glycine residue as well as that of the central cysteine [110,115].
In C. necator, the PhaC class I active enzyme is a homodimer with only one active catalytic site [114,116]. Each monomer is composed of a single polypeptide chain of 65 kDa which contains an N-terminal domain of unknown function (residues 1–200) and a C-terminal catalytic domain where the polymerization takes place (residues 201–589) (Figure 4A) (5T6O Protein Data Bank elucidated using x-ray diffraction). The active site of each monomer is separated from the other by 33 Å across an extensive dimer interface. The opening in the substrate access channel of the enzyme is near two arginine residues (one from each chain of the dimer), from which Arg398 is strictly conserved in class I PhaCs. This arginine along with the His481 are believed to be important for substrate HB-CoA binding and stabilizing while Arg421 may be involved in chain termination [114].
Figure 4B shows the structure of 3, class II, aligned PhaCs (PmePhaC, PpuPhaC and PstzPhaC) from P. mendocina NK-01, P. putida KT2440 and P. stutzeri 1317 respectively. The catalytic triad Cys, Asp and His are coloured.
The catalytic domain of PHA synthase from Cupriavidus necator shares 29% sequence identity with the class III PhaC from Allochromatium vinosum, which suggests structural similarity with class III synthases. The catalytic domains of class II (Pseudomonas, Figure 4B)) and IV PhaCs, despite of have being a lot less characterized, are also likely to be homologous, sharing around 40% and 30% identity with C. necator [114].
Substrate Specificity and Kinetics
Comparing Km of different substrates gives some clues to the specificity of an enzyme. In the study of Zhang, et al. [116]. the much larger Km value for 4HB-CoA compared with 3HB-CoA and 3HV-CoA indicates the importance of the distance between the hydroxyl group and the CoA moiety for substrate binding in the PhaC of C. necator, however, a longer side chain does not seem to strongly affect the substrate binding to PHA synthase of C. necator. (for comparison of more substrates and its Km read Table II from Yuan, et al. [118].) It is also important to mention that, while the natural substrate of PhaC in C. necator is 3HB-CoA, it has been able to accept 3-hydroxyoctanoate-CoA and 3-hydroxydodecanoate-CoA to produce medium chain PHAs when expressed in E. coli suggesting that it might have a broader substrate specificity than previously found [71].
Point Mutations that Might Affect Substrate Specificity
Proven PHA producing Pseudomonas strains include: Pseudomonas mosselii, [119,120] Pseudomonas corrugata, [121,122,123] Pseudomonas mediterranea, [121,122] Pseudomonas putida, [123,124,125]Pseudomonas mendocina, [126,127] Pseudomonas chlororaphis, [128,129] Pseudomonas stutzeri 1317 [85,130],Pseudomonas entomophila. [64,101], Pseudomonas oleovorans [131] and others.
For this literature review, an amino acid mutation map for the industrially PHA outstanding strain P. putida [132,133,134] was built following the following procedure: The amino acid sequence from the strain (NCBI: WP_010955566.1) was aligned against the NCBI database using the online tool BLAST [135]. The first 50 strains from the search were selected and using all the mutations observed, a mutation map was built and shown in Figure 5 (Mutations shown in red). The occurrence for each mutation (how many times the amino acid was substituted in our pool of sequences) was also recorded and added on the SUPPLEMENTARY Material.
Additionally, a literature search was conducted to identify point mutations that have already been tested in the laboratory. These mutations are shown in blue in Figure 5 and described in Table 4.
Substrate specificity can rely in as much as one amino acid., the substitution of Leu484 in PhaC from Pseudomonas putida for valine shifts the substrate specificity from 8C to 4C. This amino acid (484 amino acid) is adjacent to the catalytic triad and it is conserved as leucine in class II synthases, as valine in class III and as valine or isoleucine in class I (C. necator) and is critical in determining substrate specificity [137]. On the other hand, amino acids Ser326 and Gln482 (also near the catalytic triad) in Pseudomonas spp. are conserved residues, that when substituted for other residues (thr326, cys326 and lys482, arg482 respectively) have a contrary effect and increase the enzyme preference for producing PHB (C4) over mcl-polymers [130]. The reason why different amino acids can change the enzyme properties in different ways is probably because every amino acid has different physicochemical properties such as hydrophobicity, charge, isoelectric point etc. In this regard Chuah, et al. [145] noticed that substitutions with uncharged residues resulted in enhanced PHA production.
Conservative mutations occur when an amino acid is substituted for another with similar physicochemical characteristics while non-conservative mutations occur when the amino acid change for another with completely different characteristics [146]. Non-conservative mutations are expected to have a bigger impact over the protein structure/function.
Another important characteristic of amino acids is their size and structure. In a study using the class I PhaC from Chromobacterium (short-chain length preference), the mutation A479S resulted in a 4-fold increase in 3HHx monomer incorporation and a 1.6-fold increase in PHA biosynthesis. Further analysis suggested that a change in size and the geometry of the substrate-binding pocket due to this mutation was determinant for PhaC yield and specificity [145].
Interestingly enough, the amino acid A479 is also present in PhaC from Cupriavidus necator H16 which is also a class I PhaC. This suggests that this mutation would most likely change the substrate specificity of this enzyme as well but experimental work is needed to confirm this hypothesis.
Protein Engineering for Catalytic Enhancement
PHA synthase engineering is being performed for a variety of purposes that include improved PHAs accumulation yield, substrate specificity modification and/or higher molecular weight [147,148].
The engineering strategies for these purposes include random mutagenesis, error-prone PCR mutagenesis, site-specific saturation mutagenesis, localized semi-random mutagenesis, gene shuffling, recombination of beneficial mutations and engineering of chimeric PHA synthases between others [148,149]. Examples of enhanced PhaCs by enzyme evolution are shown in Table 5.
In addition to the above, PHA yield can be enhanced indirectly by modifying metabolic routes, for instance, the β-oxidation pathway to produce mcl-PHAs [65,88], by recycling the CoA released from the transformation of 3-hydroxyacyl-CoA into PHAs (CoA inhibits PhaC) [159] or by the co-expression of molecular chaperones, which results in the synthesis of larger quantities of enzyme [160]. Another approach would be to use thermo-tolerant PHA synthases in order to enhance the bioconversion process and decrease the energy costs related with managing the exothermic fermentation process [148].
Methods to Evaluate the Enzymatic Activity
The activity of PHA synthase can be determined by measuring the amount of CoA released from thioester-CoA during polymerization. The CoA in the medium is detected spectroscopically at 412 nm by reduction of 5,5-dithio-bis (2 nitrobenzoic acid) (DTNB), a compound which specifically reacts with thiol groups. One unit of enzyme activity is defined as the amount of enzyme that catalyses the release of 1 mol CoA/min. This technique, originally developed for short chain PHA synthases was then modified for medium chain synthases [161,162]. It is the most common technique and it is quick to perform, unfortunately, hydrolysis reactions can lead to the release of CoA independently of polymerisation which can give false results [70].
Another way to measure PHA synthase activity is by measuring, spectrophotometrically at 236 nm, the hydrolysis of thioesters in substrates as described by Fukui, et al. [163]. In this technique one unit of enzyme activity is defined as the amount of enzyme necessary to convert 1 µmol of substrate to PHAs in one minute. It is a convenient technique but less accurate than measuring CoA release [70]. Later on, Gerngross, et al. [164] modified the technique and used a labelled substrate ([3-3H]-hydroxyacyl-CoAs) [70,164].
In the other hand, the enzyme activity essay developed by Kraak, et al. [131] is based on the analysis of substrate depletion by HPLC and production formation by gas chromatography and has proven to be highly accurate to measure the enzyme activity of medium size chain PHA synthases, but, the method is very time consuming.
The reliance on spectrophotometric assays has the drawback of hindering the simultaneous measurement of more than one enzyme activity [70]. Burns, et al. [165] developed a HPLC methodology to measure fluctuations in the concentration of CoA, acetyl-CoA, acetoacetyl-CoA, and β-hydroxybutyryl-CoA metabolites and to associate these fluctuations to the activity of an enzyme on the overall system.
A summary of the methods discussed in this section is shown in Table 6.
PHA Depolymerase (PhaZ)
Classification
PHA depolymerases are carboxylesterases classified in 3 superfamilies according to the place where they are found in the cell (intracellular, extracellular, periplasmic) which at the same time are divided according to their substrate specificity (short or medium chain length) and according to their sequence similarities (presence of the lipase box and catalytic domain type).
Structure and Sequence
PHA depolymerases have a catalytic triad (serine – histidine – aspartic acid) as active site. Similarly to PhaC, the catalytic serine is located in a G-X-S-X-G lipase box that is present in other α/β-hydrolases. Apart from this, a conserved non-catalytic histidine near the oxyanion hole, also present in lipases, can be found in PhaZ. It is important to note that there are also non-lipase box depolymerases [6,17,166].
The best studied PHA depolymerases are dPHAscl depolymerases which contain a short signal peptide, a catalytic domain (containing the oxyanion hole and the lipase box), a short linker domain, and a substrate binding domain [17,167]. In dPHAscl with catalytic domain 1 the oxyanion hole can be found in the N-terminal to the lipase box, like lipases, whereas in dPHAscl with catalytic domain 2 the oxyanion hole is found C-terminal to the catalytic triad. Example of microorganisms harbouring dPHAscl with catalytic domain 1 are Sorangium cellulosum, Paracoccus denitrificans, Alcaligenes faecalis and Burkholderia mallei; whereas examples of microorganisms with catalytic domain type 2 are Alteromonas macleodii, Pseudoalteromonas atlantica, Cupriavidus pinatubonensis and others [6,17].
Similarly, there are three types of short linker domain: fibronectin type (i.e. Alcaligenes faecalis [168]), cadherin-like domain (i.e. Pseudomonas stutzeri [169]) and 2 types of substrate binding domain (i.e. Pseudomonas stutzeri has both of these domains [169] ) as shown in Figure 7 [168,170].
The dPHAscl catalytic domain Type II from Penicillium funiculosum, shown in Figure 8 (2D80 from Protein Data Bank elucidated using x-ray diffraction).is comprised of a single domain. The catalytic triad residues are Ser39, Asp121 and His155 and the amide groups of Ser40 and Cys250 form the oxyanion hole. A mutant revealed that Trp307 plays a role in the recognition of the ester group adjacent to the scissile group [171].
Substrate Specificity and Kinetics
PHA depolymerase catalyzes the reaction in Figure 9.
PHA depolymerization occurs in two steps: adsorption by the substrate binding domain and then hydrolysis by the catalytic domain [175,176]. The C-terminal amino acids of dPHAscl depolymerases represent the PHA-specific binding domain to the substrate (SBD) [167]. SBD has an interaction of 100 pN with the substrate [177] and such interaction consists of hydrogen bonds between hydrophilic residues in the enzyme and ester bonds in the polymer and between the hydrophobic residues in the enzyme and the methyl groups in the polymer. Check Figure 10 [178].
Using Alcaligenes faecalis, in a study with 5 substrates the SBD was capable of adsorbing on the surface of all films (PHB, PHP, P4HB, P(2HP) and P(6Hx)); However, the enzyme, only hydrolysed 3 films (PHB, PHP and P(4HB) out of the 5, demonstrating that the binding domain works independently of the catalytic domain and that it is less specific [176].
In the other hand, dPHAmcl depolymerases do not contain a substrate binding domain. In these enzymes, the N-terminal region of the catalytic domain is thought to function as substrate binding site [6,17].
As mentioned before, there are two types of catalytic domains in PHA depolymerase depending on the position of the lipase box and oxyanion hole. Nevertheless, comparisons of substrate specificities between Alcaligenes faecalis (domain type 1), Pseudomonas stutzeri (type 1), and Comamonas acidovorans (type 2) using 12 different aliphatic polyesters showed that active sites of PHA depolymerases have a similar conformational structure, independently of a difference in the sequence structure of the catalytic domains given that they were able to degrade the same 5 substrates (PHB, PHP, P(4HB), poly(ethylene-succinate) and poly(ethylene-adipate)) and unable to degrade the other seven [179].
The maximum rate of PHA depolymerase catalysed reaction (Vmax) as well as the Km constant for the degradation of PHB at optimal conditions have been measured for several organisms as shown in Table 7. A low Km represents a very big affinity for the substrate.
Protein Engineering for Catalytic Enhancement
Engineering of PHA depolymerases can be achieved using the same techniques described for PHA synthase engineering but a lot less research has been made over PhaZ.
The degradation rate of a piece of PHB is usually on the order of a few months (in anaerobic sewage) or years (in seawater) [184] but genetical engineering can be applied to make the process more efficient, for example, Hiraishi, et al. [185] made an amino acid substitution of the residue Tyr443 for a more highly hydrophobic amino acid (Phe) in PhaZ from Ralstonia pickettii which resulted in a higher PHB degradation activity while Tan, et al. [186] performed directed evolution over the catalytic domain through error- prone PCR to achieve a tenfold increase of the PHB depolymerase activity.
Methods to Evaluate the Enzymatic Activity
As mentioned before, one of the most common and simple techniques to identify PHA-producing colonies is staining them with the hydrophobic dye Sudan Black and observing under the microscope. When PHAs consumption conditions are present, colonies expressing intracellular PHA depolymerase will look less stained than before, [187] whereas, a simple way to observe extracellular depolymerase activity is using an agar plate assay (halo formation) as indicated by Schirmer, et al. [188]. The diameter of the resulting clear zone semi-quantitatively indicates the activity of the enzyme. The technique can also be used to screen microorganisms that utilize extracellular PHAs [6,189].
In the other hand, quantitative methods include the measurement of the 3HB released by, either using 3HB dehydrogenase and hydrazine and measuring spectrophotometrically [190,191] or by directly monitoring absorption at 210 nm of carbonyl groups of 3HB monomer and dimers [192,193]. Another approach to determine PHA depolymerase activity is to take advantage of the esterase activity of the enzyme and use para-nitrophenylalkanoate (pNPA) as substrate. The concentrated p-nitrophenol released by the hydrolysis of pNPB is measured spectrophotometrically at 410 nm [170,188].
Alternatively, there are weight loss measurements in which PHAs lose weight, over time, as consequence of depolymerization. However, this technique is not practical for rapid routine assays [194,195].
Finally, non-common methods for recording degradation of polyesters include electron microscopy [196], 1H-NMR (hydrogen-nuclear magnetic resonance) imaging [197], 13C-NMR (carbon nuclear magnetic resonance) [198] and tracking radiolabelled polymers (this technique can be used to measure both polymerization and depolymerization of PHAs) [199,200].
Summary of the methods discussed in this section are shown in Table 8.
Lipases Capable of Degrading PHAs
It has been proven that several lipases can degrade PHAs [166,201,202]. A comparison study of PhaZ from P. lemoignei and T1 lipase (isolated from the palm Geobacillus zalihae, capable of degrading amorphous P3HB) showed that their active site residues are very well aligned. The enzymes have 21.4% sequence identity and 53.8% sequence similarity only and, yet, their structures are very similar and they both contain the oxyanion pocket and the lipase box pentapeptide (GXSXG) [203], which can explain this phenomenon.
Most bacterial lipases are capable of hydrolyzing polyesters consisting of an ω-hydroxyalkanoic acid such as poly(6-hydroxyhexanoate) or poly(4-hydroxybutyrate), whereas polyesters containing side chains in the polymer backbone such as poly(3-hydroxybutyrate) and other poly(3-hydroxyalkanoates) are not or only somewhat hydrolyzed [166,202]. Great diversity of commercial lipases going from yeast (Candida antartica and C. rugosa), bacteria (Pseudomonas cepacia, P. fluorescens, Chromobacterium viscosum), fungus (Rhizopus arrhizus and R. oryzae), insect (Myagrus javanicus), shrub (Rubus niveus) and animals (porcine pancreas) are capable of degrading P(3HB-co-4HB) [204,205,206].
Conclusions and Future Perspectives
As indicated before in this review, (Figure 3) the substrate used by PhaC to produce mcl-PHAs is the intermediate of the fatty acid β-oxidation pathway, 3-hydroxyacyl-CoA. Fatty acids of different carbon lengths when fed to the microbial cell are metabolized by this pathway, and each time the cycle is complete the chain loses 2 carbons [209]. As a result, hydroxyacyl-CoA of different chain lengths in the cell can be incorporated to the PHA polymer making the monomer composition very difficult to control.
It has been proven that the deletion of all the copies of certain β-oxidation genes (fadA, fadB and PhaG for example, shown in Figure 3) to interrupt the pathway can lead to the accumulation of predicted homopolymers [210]. In the near future, efforts should be made to synthesize and test homopolymers of different chain lengths and to scale up the process for industrial production. Complementary, the upregulation of the genes fabG, fadE and PhaJ from this same pathway increases the metabolic flux to the production of the PHA substrate 3-hydroxyacyl-CoA thus increasing yield.
Other methods to increase yield and modify PhaC’s substrate preference is through protein engineering. In this review we show a list of point mutations that have been proven to change substrate preference and suggest some others that have not been proven yet for future research.
Moreover, in this review we move one step ahead and report the main characteristics, structure, classification and protein engineering methods to modify PhaZ which will be needed in the future to accelerate the degradation of PHA products.
Supplementary Materials
Information is available at Preprints.org.
Acknowledgments
The author Isabel Vital-Vilchis thanks CONACyT (Consejo Nacional de Ciencia y Tecnología) from Mexico for the scholarship granted for this project.
Statements and Declarations
The authors declare no conflicts of interest.
References
- Hazenberg W and Witholt B (1997) Efficient production of medium-chain-length poly(3-hydroxyalkanoates) from octane by Pseudomonas oleovorans: economic considerations, Applied Microbiology and Biotechnology, 48, 5: 588-596. [CrossRef]
- Lenz R W and Marchessault R H (2005) Bacterial Polyesters: Biosynthesis, Biodegradable Plastics and Biotechnology, Biomacromolecules, 6, 1: 1-8. [CrossRef]
- Stainer W R, Kunisawa R, and Contopoulou B (1959) The role of organic substrates in bacterial photosynthesis, Proceedings Natl. Acad. Science, 45: 1246-1260. [CrossRef]
- Poli A, Di Donato P, Abbamondi G R, and Nicolaus B (2011) Synthesis, Production, and Biotechnological Applications of Exopolysaccharides and Polyhydroxyalkanoates by Archaea, Archaea, 2011, 1: 693253. [CrossRef]
- Lemoigne M (1926) Produits de deshydration et de polymerisation de l’acide B-oxybutyric, The Bulletin de la Société Chimique de France 8: 770-778.
- Jendrossek D and Handrick R (2002) Microbial Degradation of Polyhydroxyalkanoates, Annual Review of Microbiology, 56, 1: 403-432. [CrossRef]
- Steinbüchel A and Valentin H E (1995) Diversity of bacterial polyhydroxyalkanoic acids, FEMS Microbiology Letters, 128, 3: 219-228. [CrossRef]
- Gao X, Chen J-C, Wu Q, and Chen G-Q (2011) Polyhydroxyalkanoates as a source of chemicals, polymers, and biofuels, Current Opinion in Biotechnology, 22, 6: 768-774. [CrossRef]
- Reddy C S K, Ghai R, Rashmi, and Kalia V C (2003) Polyhydroxyalkanoates: an overview, Bioresource Technology, 87, 2: 137-146. [CrossRef]
- Samui A B and Kanai T (2019) Polyhydroxyalkanoates based copolymers, International Journal of Biological Macromolecules, 140: 522-537. [CrossRef]
- Grigore M E, Grigorescu R M, Iancu L, Ion R-M, Zaharia C, and Andrei E R (2019) Methods of synthesis, properties and biomedical applications of polyhydroxyalkanoates: a review, Journal of Biomaterials Science, Polymer Edition, 30, 9: 695-712. [CrossRef]
- Pandey A, Adama N, Adjallé K, and Blais J-F (2022) Sustainable applications of polyhydroxyalkanoates in various fields: A critical review, International Journal of Biological Macromolecules, 221: 1184-1201. [CrossRef]
- Nagarajan D, Aristya Ganies R, Lin Y-J, Chang J-J, Yen H-W, and Chang J-S (2021) Microbial cell factories for the production of polyhydroxyalkanoates, Essays in Biochemistry, 65, 2: 337-353. [CrossRef]
- Rehm B H A (2003) Polyester synthases: natural catalysts for plastics, Biochemical Journal, 376, 1: 15-33. [CrossRef]
- Lee S Y (1996) Bacterial polyhydroxyalkanoates, Biotechnology and Bioengineering, 49, 1: 1-14. [CrossRef]
- Steinbüchel A and Lütke-Eversloh T (2003) Metabolic engineering and pathway construction for biotechnological production of relevant polyhydroxyalkanoates in microorganisms, Biochemical Engineering Journal, 16, 2: 81-96. [CrossRef]
- Knoll M, Hamm T M, Wagner F, Martinez V, and Pleiss J (2009) The PHA Depolymerase Engineering Database: A systematic analysis tool for the diverse family of polyhydroxyalkanoate (PHA) depolymerases, BMC Bioinformatics, 10, 1: 89. [CrossRef]
- Wang Y, Yin J, and Chen G-Q (2014) Polyhydroxyalkanoates, challenges and opportunities, Current Opinion in Biotechnology, 30: 59-65. [CrossRef]
- Atiwesh G, Mikhael A, Parrish C C, Banoub J, and Le T-A T (2021) Environmental impact of bioplastic use: A review, Heliyon, 7, 9: e07918. [CrossRef]
- Fredi G and Dorigato A (2021) Recycling of bioplastic waste: A review, Advanced Industrial and Engineering Polymer Research, 4, 3: 159-177. [CrossRef]
- Bioplastics E. “European Bioplastics e.V.” https://www.european-bioplastics.org/bioplastics/ (accessed 11th March, 2024, 2024).
- Bhagwat G et al. (2020) Benchmarking Bioplastics: A Natural Step Towards a Sustainable Future, Journal of Polymers and the Environment, 28, 12: 3055-3075. [CrossRef]
- Das M, Manda B, and Katiyar V, Sustainable routes for synthesis of poly (ε-caprolactone): Prospects in chemical industries. Singapore: Springer, 2020.
- Patni N, Yadava P, Agarwal A, and Maroo V (2014) An overview on the role of wheat gluten as a viable substitute for biodegradable plastics, 30, 4: 421-430. [CrossRef]
- Hardin T. “Plastic: It’s Not All the Same.” Copyright © Plastic Oceans International. https://plasticoceans.org/7-types-of-plastic/ (accessed February, 2024).
- Zhou Y, He Y, Lin X, Feng Y, and Liu M (2022) Sustainable, High-Performance, and Biodegradable Plastics Made from Chitin, ACS Applied Materials & Interfaces, 14, 41: 46980-46993. [CrossRef]
- Chen G-Q, Chen X-Y, Wu F-Q, and Chen J-C (2020) Polyhydroxyalkanoates (PHA) toward cost competitiveness and functionality, Advanced Industrial and Engineering Polymer Research, 3, 1: 1-7. [CrossRef]
- Jian J, Xiangbin Z, and Xianbo H (2020) An overview on synthesis, properties and applications of poly(butylene-adipate-co-terephthalate)–PBAT, Advanced Industrial and Engineering Polymer Research, 3, 1: 19-26. [CrossRef]
- Rishi V, Sandhu A K, Kaur A, Kaur J, Sharma S, and Soni S K (2020) Utilization of kitchen waste for production of pullulan to develop biodegradable plastic, Applied Microbiology and Biotechnology, 104, 3: 1307-1317. [CrossRef]
- Wang J, Wang L, Gardner D J, Shaler S M, and Cai Z (2021) Towards a cellulose-based society: opportunities and challenges, Cellulose, 28, 8: 4511-4543. [CrossRef]
- Gheorghita R, Gutt G, and Amariei S, “The Use of Edible Films Based on Sodium Alginate in Meat Product Packaging: An Eco-Friendly Alternative to Conventional Plastic Materials,” Coatings, vol. 10, no. 2. [CrossRef]
- Guilbert S, Morel M-H, Gontard N, and Cuq B, “Protein-Based Plastics and Composites as Smart Green Materials,” in Feedstocks for the Future, vol. 921, (ACS Symposium Series, no. 921): American Chemical Society, 2006, ch. 24, pp. 334-350. [CrossRef]
- Merino D, Paul U C, and Athanassiou A (2021) Bio-based plastic films prepared from potato peels using mild acid hydrolysis followed by plasticization with a polyglycerol, Food Packaging and Shelf Life, 29: 100707. [CrossRef]
- Baker M I, Walsh S P, Schwartz Z, and Boyan B D (2012) A review of polyvinyl alcohol and its uses in cartilage and orthopedic applications, Journal of Biomedical Materials Research Part B: Applied Biomaterials, 100B, 5: 1451-1457. [CrossRef]
- Jalabert M, Fraschini C, and Prud’homme R E (2007) Synthesis and characterization of poly(L-lactide)s and poly(D-lactide)s of controlled molecular weight, Journal of Polymer Science Part A: Polymer Chemistry, 45, 10: 1944-1955. [CrossRef]
- Patnaik S, Panda A K, and Kumar S (2020) Thermal degradation of corn starch based biodegradable plastic plates and determination of kinetic parameters by isoconversional methods using thermogravimetric analyzer, Journal of the Energy Institute, 93, 4: 1449-1459. [CrossRef]
- Li G et al., “Synthesis and Biological Application of Polylactic Acid,” Molecules, vol. 25, no. 21. [CrossRef]
- Siracusa V and Blanco I, “Bio-Polyethylene (Bio-PE), Bio-Polypropylene (Bio-PP) and Bio-Poly(ethylene terephthalate) (Bio-PET): Recent Developments in Bio-Based Polymers Analogous to Petroleum-Derived Ones for Packaging and Engineering Applications,” Polymers, vol. 12, no. 8. [CrossRef]
- Barletta M et al. (2022) Poly(butylene succinate) (PBS): Materials, processing, and industrial applications, Progress in Polymer Science, 132: 101579. [CrossRef]
- Mohanan N, Montazer Z, Sharma P K, and Levin D B (2020) Microbial and Enzymatic Degradation of Synthetic Plastics, (in English), Frontiers in Microbiology, Review 11. [CrossRef]
- Telmo O, “Polymers and the Environment,” in Polymer Science, Y. Faris Ed. Rijeka: IntechOpen, 2013, p. Ch. 1.
- Dilkes-Hoffman L S, Lant P A, Laycock B, and Pratt S (2019) The rate of biodegradation of PHA bioplastics in the marine environment: A meta-study, Marine Pollution Bulletin, 142: 15-24. [CrossRef]
- Rudnik E, “11 - Biodegradability Testing of Compostable Polymer Materials,” in Handbook of Biopolymers and Biodegradable Plastics, S. Ebnesajjad Ed. Boston: William Andrew Publishing, 2013, pp. 213-263.
- worldometer. “Oil left in the world.” https://www.worldometers.info/oil/#:~:text=The%20world%20has%20proven%20reserves,(at%20current%20consumption%20levels) (accessed October 29, 2024, 2024).
- Rajendran N and Han J (2022) Techno-economic analysis of food waste valorization for integrated production of polyhydroxyalkanoates and biofuels, Bioresource Technology, 348: 126796. [CrossRef]
- Statita. “Statista. Price of high-density polyethylene worldwide from 2017 to 2023.” https://www.statista.com/statistics/1171074/price-high-density-polyethylene-forecast-globally/ (accessed.
- Choi J and Lee S Y (1999) Factors affecting the economics of polyhydroxyalkanoate production by bacterial fermentation, Applied Microbiology and Biotechnology, 51, 1: 13-21. [CrossRef]
- Technavio, “Top 6 vendors in the polyhydroxyalkanoate market from 2017 to 2021:Technavio,” 2017. [Online]. Available: https://www.businesswire.com/news/home/20170824005079/en/Top-6-Vendors-Polyhydroxyalkanoate-Market-2017-2021.
- Kourmentza C et al. (2017) Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production, Bioengineering, 4, 2. [CrossRef]
- Jiang G et al. (2016) Carbon Sources for Polyhydroxyalkanoates and an Integrated Biorefinery, (in eng), Int J Mol Sci, 17, 7: 1157. [CrossRef]
- Bio-on. “Bio-on.” http://www.bio-on.it/production.php#p9 (accessed october 17, 2019).
- Danimerscientific. “Danimer scientific.” https://danimerscientific.com/pha-the-future-of-biopolymers/pha-culture-manufacturing/ (accessed october 17, 2019).
- TianAn. “TianAn Biopolymer.” http://www.tianan-enmat.com/index.html# (accessed october 17, 2019).
- TianjinGreenBio. “Tiajin GreenBio Materials.” http://www.tjgreenbio.com/en/about.aspx?title=About%20GreenBio&cid=25 (accessed october 17, 2019).
- Chen G-Q (2009) A microbial polyhydroxyalkanoates (PHA) based bio- and materials industry, Chemical Society Reviews. 8: 2434-2446. [CrossRef]
- Ray S, “Mass Multiplication, Production Cost Analysis and Marketing of Polyhydroxyalkanoates (PHAs),” in Industrial Microbiology Based Entrepreneurship: Making Money from Microbes, N. Amaresan, D. Dharumadurai, and D. R. Cundell Eds. Singapore: Springer Nature Singapore, 2022, pp. 117-125. [CrossRef]
- Jabeen N, Majid I, and Nayik G A (2015) Bioplastics and food packaging: A review, Cogent Food & Agriculture, 1, 1: 1117749. [CrossRef]
- Noda I, Green P R, Satkowski M M, and Schechtman L A (2005) Preparation and Properties of a Novel Class of Polyhydroxyalkanoate Copolymers, Biomacromolecules, 6, 2: 580-586. [CrossRef]
- Samrot A V, Avinesh R B, Sukeetha S D, and Senthilkumar P (2011) Accumulation of Poly[(R)-3-hydroxyalkanoates] in Enterobacter cloacae SU-1 During Growth with Two Different Carbon Sources in Batch Culture, Applied Biochemistry and Biotechnology, 163, 1: 195-203. [CrossRef]
- Kessler B, Weusthius R, Witholt B, and Eggink G (2001) Production of microbial polyesters: fermentation and downstream processes, Advances in Biochemical Engineering/ Biotechnology, 71: 159-82. [CrossRef]
- Evans D A (2014) History of the Harvard ChemDraw Project, Angewandte Chemie International Edition. 53, 42: 11140-11145. [CrossRef]
- Olivera E R, Arcos M, Naharro G, and Luengo J M, “Unusual PHA Biosynthesis,” in Plastics from Bacteria: Natural Functions and Applications, G. G.-Q. Chen Ed. Berlin, Heidelberg: Springer Berlin Heidelberg, 2010, pp. 133-186. [CrossRef]
- Chen G-Q and Wu Q (2005) Microbial production and applications of chiral hydroxyalkanoates, Applied Microbiology and Biotechnology, 67, 5: 592-599. [CrossRef]
- Wang Y, Chung A, and Chen G-Q (2017) Synthesis of Medium-Chain-Length Polyhydroxyalkanoate Homopolymers, Random Copolymers, and Block Copolymers by an Engineered Strain of Pseudomonas entomophila, Advanced Healthcare Materials, 6, 7: 1601017. [CrossRef]
- Lu J, Tappel R, and Nomura C (2009) Mini-Review: Biosynthesis of Poly(hydroxyalkanoates), Journal of Macromolecular Science®, Part C: Polymer Reviews: 226-248. [CrossRef]
- Matsusaki H, Abe H, and Doi Y (2000) Biosynthesis and Properties of Poly(3-hydroxybutyrate-co-3-hydroxyalkanoates) by Recombinant Strains of Pseudomonas sp. 61-3, Biomacromolecules, 1, 1: 17-22. [CrossRef]
- Tsuge T, Hyakutake M, and Mizuno K (2015) Class IV polyhydroxyalkanoate (PHA) synthases and PHA-producing Bacillus, Applied Microbiology and Biotechnology, 99, 15: 6231-6240. [CrossRef]
- Altschul S F et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs, Nucleic Acids Research, 25, 17: 3389-3402. [CrossRef]
- Liu Z et al. (2019) Domain-centric dissection and classification of prokaryotic poly(3-hydroxyalkanoate) synthases, bioRxiv: 693432. [CrossRef]
- Thomson N, Roy I, Summers D, and Sivaniah E (2010) In vitro production of polyhydroxyalkanoates: achievements and applications, Journal of Chemical Technology & Biotechnology, 85, 6: 760-767. [CrossRef]
- Antonio R V, Steinbüchel A, and Rehm B H A (2000) Analysis of in vivo substrate specificity of the PHA synthase from Ralstonia eutropha: formation of novel copolyesters in recombinant Escherichia coli, FEMS Microbiology Letters, 182, 1: 111-117. [CrossRef]
- Wong P A L, Chua H, Lo W, Lawford H G, and Yu P H (2002) Production of specific copolymers of polyhydroxyalkanoates from industrial waste, Applied Biochemistry and Biotechnology, journal article 98, 1: 655-662. [CrossRef]
- Aragao G M F, Lindley N D, Uribelarrea J L, and Pareilleux A (1996) Maintaining a controlled residual growth capacity increases the production of polyhydroxyalkanoate copolymers by Alcaligenes eutrophus, Biotechnology Letters, 18, 8: 937-942. [CrossRef]
- Bates F S and Fredrickson G H (1990) Block Copolymer Thermodynamics: Theory and Experiment, Annual Review of Physical Chemistry, 41, 1: 525-557. [CrossRef]
- Mantzaris N V, Kelley A S, Daoutidis P, and Srienc F (2002) A population balance model describing the dynamics of molecular weight distributions and the structure of PHA copolymer chains, Chemical Engineering Science, 57, 21: 4643-4663. [CrossRef]
- Muthuraj R, Valerio O, and Mekonnen T H (2021) Recent developments in short- and medium-chain- length Polyhydroxyalkanoates: Production, properties, and applications, International Journal of Biological Macromolecules, 187: 422-440. [CrossRef]
- Pereira J R et al. (2019) Demonstration of the adhesive properties of the medium-chain-length polyhydroxyalkanoate produced by Pseudomonas chlororaphis subsp. aurantiaca from glycerol, International Journal of Biological Macromolecules, 122: 1144-1151. [CrossRef]
- Panaitescu D M et al. (2017) Medium Chain-Length Polyhydroxyalkanoate Copolymer Modified by Bacterial Cellulose for Medical Devices, Biomacromolecules, 18, 10: 3222-3232. [CrossRef]
- Ansari S, Sami N, Yasin D, Ahmad N, and Fatma T (2021) Biomedical applications of environmental friendly poly-hydroxyalkanoates, International Journal of Biological Macromolecules, 183: 549-563. [CrossRef]
- Sun J, Dai Z, Zhao Y, and Chen G-Q (2007) In vitro effect of oligo-hydroxyalkanoates on the growth of mouse fibroblast cell line L929, Biomaterials, 28, 27: 3896-3903. [CrossRef]
- Rai R, Keshavarz T, Roether J A, Boccaccini A R, and Roy I (2011) Medium chain length polyhydroxyalkanoates, promising new biomedical materials for the future, Materials Science and Engineering: R: Reports, 72, 3: 29-47. [CrossRef]
- Xu X-Y et al. (2010) The behaviour of neural stem cells on polyhydroxyalkanoate nanofiber scaffolds, Biomaterials, 31, 14: 3967-3975. [CrossRef]
- Chamas A et al. (2020) Degradation Rates of Plastics in the Environment, ACS Sustainable Chemistry & Engineering, 8, 9: 3494-3511. [CrossRef]
- Kumar M et al. (2020) Bacterial polyhydroxyalkanoates: Opportunities, challenges, and prospects, Journal of Cleaner Production, 263: 121500. [CrossRef]
- Chen J-Y, Song G, and Chen G-Q (2006) A lower specificity PhaC2 synthase from Pseudomonas stutzeri catalyses the production of copolyesters consisting of short-chain-length and medium-chain-length 3-hydroxyalkanoates, Antonie van Leeuwenhoek, 89, 1: 157-167. [CrossRef]
- Lu X, Zhang J, Wu Q, and Chen G-Q (2003) Enhanced production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) via manipulating the fatty acid β-oxidation pathway in E. coli, FEMS Microbiology Letters, 221, 1: 97-101. [CrossRef]
- Liu Q, Luo G, Zhou X R, and Chen G-Q (2011) Biosynthesis of poly(3-hydroxydecanoate) and 3-hydroxydodecanoate dominating polyhydroxyalkanoates by β-oxidation pathway inhibited Pseudomonas putida, Metabolic Engineering, 13, 1: 11-17. [CrossRef]
- Meng D-C and Chen G-Q, “Synthetic Biology of Polyhydroxyalkanoates (PHA),” in Synthetic Biology – Metabolic Engineering, H. Zhao and A.-P. Zeng Eds. Cham: Springer International Publishing, 2018, pp. 147-174. [CrossRef]
- Jiang X J, Sun Z, Ramsay J A, and Ramsay B A (2013) Fed-batch production of MCL-PHA with elevated 3-hydroxynonanoate content, AMB Express, 3, 1: 50. [CrossRef]
- Vo M T, Lee K-W, Jung Y-M, and Lee Y-H (2008) Comparative effect of overexpressed phaJ and fabG genes supplementing (R)-3-hydroxyalkanoate monomer units on biosynthesis of mcl-polyhydroxyalkanoate in Pseudomonas putida KCTC1639, Journal of Bioscience and Bioengineering, 106, 1: 95-98. [CrossRef]
- Flores-Sánchez A, Rathinasabapathy A, López-Cuellar M d R, Vergara-Porras B, and Pérez-Guevara F (2020) Biosynthesis of polyhydroxyalkanoates from vegetable oil under the co-expression of fadE and phaJ genes in Cupriavidus necator, International Journal of Biological Macromolecules, 164: 1600-1607. [CrossRef]
- Budde C F, Riedel S L, Willis L B, Rha C, and Sinskey A J (2011) Production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from plant oil by engineered Ralstonia eutropha strains, (in eng), Applied and environmental microbiology, 77, 9: 2847-2854. [CrossRef]
- Wong Y-M, Brigham C J, Rha C, Sinskey A J, and Sudesh K (2012) Biosynthesis and characterization of polyhydroxyalkanoate containing high 3-hydroxyhexanoate monomer fraction from crude palm kernel oil by recombinant Cupriavidus necator, Bioresource Technology, 121: 320-327. [CrossRef]
- Fukui T, Abe H, and Doi Y (2002) Engineering of Ralstonia eutropha for Production of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from Fructose and Solid-State Properties of the Copolymer, Biomacromolecules, 3, 3: 618-624. [CrossRef]
- Bhatia S K et al. (2018) Production of (3-hydroxybutyrate-co-3-hydroxyhexanoate) copolymer from coffee waste oil using engineered Ralstonia eutropha, Bioprocess and Biosystems Engineering, 41, 2: 229-235. [CrossRef]
- Agnew D E, Stevermer A K, Youngquist J T, and Pfleger B F (2012) Engineering Escherichia coli for production of C12–C14 polyhydroxyalkanoate from glucose, Metabolic Engineering, 14, 6: 705-713. [CrossRef]
- Park S J, Park J P, and Lee S Y (2002) Metabolic engineering of Escherichia coli for the production of medium-chain-length polyhydroxyalkanoates rich in specific monomers, FEMS Microbiology Letters, 214, 2: 217-222. [CrossRef]
- Tan H T, Chek M F, Lakshmanan M, Foong C P, Hakoshima T, and Sudesh K (2020) Evaluation of BP-M-CPF4 polyhydroxyalkanoate (PHA) synthase on the production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from plant oil using Cupriavidus necator transformants, International Journal of Biological Macromolecules, 159: 250-257. [CrossRef]
- Salvachúa D et al. (2020) Metabolic engineering of Pseudomonas putida for increased polyhydroxyalkanoate production from lignin, Microbial Biotechnology, 13, 1: 290-298. [CrossRef]
- Tripathi L, Wu L-P, Chen J, and Chen G-Q (2012) Synthesis of Diblock copolymer poly-3-hydroxybutyrate -block-poly-3-hydroxyhexanoate [PHB-b-PHHx] by a β-oxidation weakened Pseudomonas putida KT2442, (in eng), Microbial cell factories, 11: 44-44. [CrossRef]
- Li M et al. (2019) Engineering Pseudomonas entomophila for synthesis of copolymers with defined fractions of 3-hydroxybutyrate and medium-chain-length 3-hydroxyalkanoates, Metabolic Engineering, 52: 253-262. [CrossRef]
- Ouyang S-P et al. (2007) Production of Polyhydroxyalkanoates with High 3-Hydroxydodecanoate Monomer Content by fadB and fadA Knockout Mutant of Pseudomonas putida KT2442, Biomacromolecules, 8, 8: 2504-2511. [CrossRef]
- Zhao F et al. (2020) Metabolic engineering of Pseudomonas mendocina NK-01 for enhanced production of medium-chain-length polyhydroxyalkanoates with enriched content of the dominant monomer, International Journal of Biological Macromolecules, 154: 1596-1605. [CrossRef]
- Liu W and Chen G-Q (2007) Production and characterization of medium-chain-length polyhydroxyalkanoate with high 3-hydroxytetradecanoate monomer content by fadB and fadA knockout mutant of Pseudomonas putida KT2442, Applied Microbiology and Biotechnology, 76, 5: 1153-1159. [CrossRef]
- Wang L et al. (2019) Bioinformatics Analysis of Metabolism Pathways of Archaeal Energy Reserves, Scientific Reports, 9, 1: 1034. [CrossRef]
- Lane C E and Benton M G (2015) Detection of the enzymatically-active polyhydroxyalkanoate synthase subunit gene, phaC, in cyanobacteria via colony PCR, Molecular and Cellular Probes, 29, 6: 454-460. [CrossRef]
- Blunt W, Levin D B, and Cicek N, “Bioreactor Operating Strategies for Improved Polyhydroxyalkanoate (PHA) Productivity,” Polymers, vol. 10, no. 11. [CrossRef]
- Oliveira-Filho E R, Gomez J G C, Taciro M K, and Silva L F (2021) Burkholderia sacchari (synonym Paraburkholderia sacchari): An industrial and versatile bacterial chassis for sustainable biosynthesis of polyhydroxyalkanoates and other bioproducts, Bioresource Technology, 337: 125472. [CrossRef]
- Oliveira-Filho E R et al. (2022) Engineering Burkholderia sacchari to enhance poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) [P(3HB-co-3HHx)] production from xylose and hexanoate, International Journal of Biological Macromolecules, 213: 902-914. [CrossRef]
- Mezzolla V, D’Urso F O, and Poltronieri P (2018) Role of PhaC Type I and Type II Enzymes during PHA Biosynthesis, Polymers, 10, 8. [CrossRef]
- Chen J-Y, Liu T, Zheng Z, Chen J-C, and Chen G-Q (2006) Polyhydroxyalkanoate synthases PhaC1 and PhaC2 from Pseudomonas stutzeri 1317 had different substrate specificities, FEMS Microbiology Letters, 234, 2: 231-237. [CrossRef]
- Bhubalan K et al. (2011) Characterization of the Highly Active Polyhydroxyalkanoate Synthase of <span class="named-content genus-species" id="named-content-1">Chromobacterium</span> sp. Strain USM2, Applied and Environmental Microbiology, 77, 9: 2926. [CrossRef]
- Trakunjae C et al., “Biosynthesis of P(3HB-co-3HHx) Copolymers by a Newly Engineered Strain of Cupriavidus necator PHB−4/pBBR_CnPro-phaCRp for Skin Tissue Engineering Application,” Polymers, vol. 14, no. 19. [CrossRef]
- Wittenborn E C, Jost M, Wei Y, Stubbe J, and Drennan C L (2016) Structure of the Catalytic Domain of the Class I Polyhydroxybutyrate Synthase from Cupriavidus necator, (in eng), J Biol Chem, 291, 48: 25264-25277. [CrossRef]
- Nambu Y, Ishii-Hyakutake M, Harada K, Mizuno S, and Tsuge T (2019) Expanded Amino Acid Sequence of the PhaC Box in the Active Center of Polyhydroxyalkanoate Synthases, FEBS Letters, n/a, n/a. [CrossRef]
- Zhang S, Yasuo T, Lenz R W, and Goodwin S (2000) Kinetic and Mechanistic Characterization of the Polyhydroxybutyrate Synthase from Ralstonia eutropha, Biomacromolecules, 1, 2: 244-251. [CrossRef]
- PyMOL. (2020). [Online]. Available: http://www.pymol.org/pymol.
- Yuan W et al. (2001) Class I and III Polyhydroxyalkanoate Synthases from Ralstonia eutropha and Allochromatium vinosum: Characterization and Substrate Specificity Studies, Archives of Biochemistry and Biophysics, 394, 1: 87-98. [CrossRef]
- Liu M-H, Chen Y, Jr., and Lee C-Y (2018) Characterization of medium-chain-length polyhydroxyalkanoate biosynthesis by Pseudomonas mosselii TO7 using crude glycerol, Bioscience, Biotechnology, and Biochemistry, 82, 3: 532-539. [CrossRef]
- Chen Y, Jr., Huang Y-C, and Lee C-Y (2014) Production and characterization of medium-chain-length polyhydroxyalkanoates by Pseudomonas mosselii TO7, Journal of Bioscience and Bioengineering, 118, 2: 145-152. [CrossRef]
- Licciardello G, Catara A F, and Catara V, “Production of Polyhydroxyalkanoates and Extracellular Products Using Pseudomonas Corrugata and P. Mediterranea: A Review,” Bioengineering, vol. 6, no. 4. [CrossRef]
- Licciardello G et al. (2017) Transcriptome analysis of Pseudomonas mediterranea and P. corrugata plant pathogens during accumulation of medium-chain-length PHAs by glycerol bioconversion, New Biotechnology, 37: 39-47. [CrossRef]
- Solaiman D K Y, Ashby R D, and Foglia T A (2002) Physiological Characterization and Genetic Engineering of Pseudomonas corrugata for Medium-Chain-Length Polyhydroxyalkanoates Synthesis from Triacylglycerols, Current Microbiology, 44, 3: 189-195. [CrossRef]
- Sharma P K, Fu J, Zhang X, Fristensky B, Sparling R, and Levin D B (2014) Genome features of Pseudomonas putida LS46, a novel polyhydroxyalkanoate producer and its comparison with other P. putida strains, AMB Express, 4, 1: 37. [CrossRef]
- Cerrone F et al. (2014) Medium chain length polyhydroxyalkanoate (mcl-PHA) production from volatile fatty acids derived from the anaerobic digestion of grass, Applied Microbiology and Biotechnology, 98, 2: 611-620. [CrossRef]
- Zhao F et al. (2019) Screening of endogenous strong promoters for enhanced production of medium-chain-length polyhydroxyalkanoates in Pseudomonas mendocina NK-01, Scientific Reports, 9, 1: 1798. [CrossRef]
- Guo W, Duan J, Geng W, Feng J, Wang S, and Song C (2013) Comparison of medium-chain-length polyhydroxyalkanoates synthases from Pseudomonas mendocina NK-01 with the same substrate specificity, Microbiological Research, 168, 4: 231-237. [CrossRef]
- Pereira J R et al. (2021) Production of medium-chain-length polyhydroxyalkanoates by Pseudomonas chlororaphis subsp. aurantiaca: Cultivation on fruit pulp waste and polymer characterization, International Journal of Biological Macromolecules, 167: 85-92. [CrossRef]
- Meneses L, Craveiro R, Jesus A R, Reis M A M, Freitas F, and Paiva A, “Supercritical CO2 Assisted Impregnation of Ibuprofen on Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHA),” Molecules, vol. 26, no. 16. [CrossRef]
- Shen X-W, Shi Z-Y, Song G, Li Z-J, and Chen G-Q (2011) Engineering of polyhydroxyalkanoate (PHA) synthase PhaC2Ps of Pseudomonas stutzeri via site-specific mutation for efficient production of PHA copolymers, Applied Microbiology and Biotechnology, 91, 3: 655-665. [CrossRef]
- Kraak M N, Kessler B, and Witholt B (1997) In vitro Activities of Granule-Bound Poly[(R)-3-Hydroxyalkanoate] Polymerase C1 of Pseudomonas oleovorans, European Journal of Biochemistry, 250, 2: 432-439. [CrossRef]
- Weimer A, Kohlstedt M, Volke D C, Nikel P I, and Wittmann C (2020) Industrial biotechnology of Pseudomonas putida: advances and prospects, Applied Microbiology and Biotechnology, 104, 18: 7745-7766. [CrossRef]
- Fontaine P, Mosrati R, and Corroler D (2017) Medium chain length polyhydroxyalkanoates biosynthesis in Pseudomonas putida mt-2 is enhanced by co-metabolism of glycerol/octanoate or fatty acids mixtures, International Journal of Biological Macromolecules, 98: 430-435. [CrossRef]
- Le Meur S, Zinn M, Egli T, Thöny-Meyer L, and Ren Q (2012) Production of medium-chain-length polyhydroxyalkanoates by sequential feeding of xylose and octanoic acid in engineered Pseudomonas putida KT2440, BMC Biotechnology, 12, 1: 53. [CrossRef]
- Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos,, J. B, and K. a M, T.L. (2009) BLAST+: architecture and applications., BMC Bioinformatics, 10, 421.: 421. [CrossRef]
- Tsuge T et al. (2005) Biosynthesis of Polyhydroxyalkanoate (PHA) Copolymer from Fructose Using Wild-Type and Laboratory-Evolved PHA Synthases, Macromolecular Bioscience, 5, 2: 112-117. [CrossRef]
- Chen Y, Jr., Tsai P-C, Hsu C-H, and Lee C-Y (2014) Critical residues of class II PHA synthase for expanding the substrate specificity and enhancing the biosynthesis of polyhydroxyalkanoate, Enzyme and Microbial Technology, 56: 60-66. [CrossRef]
- Hiroe A, Watanabe S, Kobayashi M, Nomura C T, and Tsuge T (2018) Increased synthesis of poly(3-hydroxydodecanoate) by random mutagenesis of polyhydroxyalkanoate synthase, Applied Microbiology and Biotechnology, 102, 18: 7927-7934. [CrossRef]
- Takase K, Taguchi S, and Doi Y (2003) Enhanced Synthesis of Poly(3-hydroxybutyrate) in Recombinant Escherichia coli by Means of Error-Prone PCR Mutagenesis, Saturation Mutagenesis, and In Vitro Recombination of the Type II Polyhydroxyalkanoate Synthase Gene, The Journal of Biochemistry, 133, 1: 139-145. [CrossRef]
- Matsumoto K i, Takase K, Yamamoto Y, Doi Y, and Taguchi S (2009) Chimeric Enzyme Composed of Polyhydroxyalkanoate (PHA) Synthases from Ralstonia eutropha and Aeromonas caviae Enhances Production of PHAs in Recombinant Escherichia coli, Biomacromolecules, 10, 4: 682-685. [CrossRef]
- Yang T H, Jung Y K, Kang H O, Kim T W, Park S J, and Lee S Y (2011) Tailor-made type II Pseudomonas PHA synthases and their use for the biosynthesis of polylactic acid and its copolymer in recombinant Escherichia coli, Applied Microbiology and Biotechnology, 90, 2: 603-614. [CrossRef]
- Yang T H et al. (2010) Biosynthesis of polylactic acid and its copolymers using evolved propionate CoA transferase and PHA synthase, Biotechnology and Bioengineering, 105, 1: 150-160. [CrossRef]
- Ren Y, Meng D, Wu L, Chen J, Wu Q, and Chen G-Q (2017) Microbial synthesis of a novel terpolyester P(LA-co-3HB-co-3HP) from low-cost substrates, Microbial Biotechnology, 10, 2: 371-380. [CrossRef]
- Shozui F et al. (2010) A New Beneficial Mutation in Pseudomonas sp. 61-3 Polyhydroxyalkanoate (PHA) Synthase for Enhanced Cellular Content of 3-Hydroxybutyrate-Based PHA Explored Using Its Enzyme Homolog as a Mutation Template, Bioscience, Biotechnology, and Biochemistry, 74, 8: 1710-1712. [CrossRef]
- Chuah J-A et al. (2013) Characterization of Site-Specific Mutations in a Short-Chain-Length/Medium-Chain-Length Polyhydroxyalkanoate Synthase: In Vivo and In Vitro Studies of Enzymatic Activity and Substrate Specificity, Applied and Environmental Microbiology, 79, 12: 3813-3821. [CrossRef]
- Seng Wong T and Lan Tee K, Springer, Ed. A Practical Guide to Protein Engineering. 2020.
- Tsuge T (2016) Fundamental factors determining the molecular weight of polyhydroxyalkanoate during biosynthesis, Polymer Journal, 48, 11: 1051-1057. [CrossRef]
- Zou H, Shi M, Zhang T, Li L, Li L, and Xian M (2017) Natural and engineered polyhydroxyalkanoate (PHA) synthase: key enzyme in biopolyester production, Applied Microbiology and Biotechnology, 101, 20: 7417-7426. [CrossRef]
- Nomura C T and Taguchi S (2007) PHA synthase engineering toward superbiocatalysts for custom-made biopolymers, Applied Microbiology and Biotechnology, 73, 5: 969-979. [CrossRef]
- Tajima K et al. (2012) In vitro synthesis of polyhydroxyalkanoate (PHA) incorporating lactate (LA) with a block sequence by using a newly engineered thermostable PHA synthase from Pseudomonas sp. SG4502 with acquired LA-polymerizing activity, Applied Microbiology and Biotechnology, 94, 2: 365-376. [CrossRef]
- Sheu D-S and Lee C-Y (2004) Altering the Substrate Specificity of Polyhydroxyalkanoate Synthase 1 Derived from <em>Pseudomonas putida</em> GPo1 by Localized Semirandom Mutagenesis, Journal of Bacteriology, 186, 13: 4177. [CrossRef]
- Rehm B H A, Antonio R V, Spiekermann P, Amara A A, and Steinbüchel A (2002) Molecular characterization of the poly(3-hydroxybutyrate) (PHB) synthase from Ralstonia eutropha: in vitro evolution, site-specific mutagenesis and development of a PHB synthase protein model, Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology, 1594, 1: 178-190. [CrossRef]
- Taguchi S, Nakamura H, Hiraishi T, Yamato I, and Doi Y (2002) In Vitro Evolution of a Polyhydroxybutyrate Synthase by Intragenic Suppression-Type Mutagenesis1, The Journal of Biochemistry, 131, 6: 801-806. [CrossRef]
- Niamsiri N, Delamarre S C, Kim Y-R, and Batt C A (2004) Engineering of Chimeric Class II Polyhydroxyalkanoate Synthases, Applied and Environmental Microbiology, 70, 11: 6789. [CrossRef]
- Tsuge T, Watanabe S, Shimada D, Abe H, Doi Y, and Taguchi S (2007) Combination of N149S and D171G mutations in Aeromonas caviae polyhydroxyalkanoate synthase and impact on polyhydroxyalkanoate biosynthesis, FEMS Microbiology Letters, 277, 2: 217-222. [CrossRef]
- Sun J, Shozui F, Yamada M, Matsumoto K i, Takase K, and Taguchi S (2010) Production of P(3-hydroxybutyrate-co-3-hydroxyhexanoate-co-3-hydroxyoctanoate) Terpolymers Using a Chimeric PHA Synthase in Recombinant Ralstonia eutropha and Pseudomonas putida, Bioscience, Biotechnology, and Biochemistry, 74, 8: 1716-1718. [CrossRef]
- Chen Y, Jr. (2014) Critical residues of class II PHA synthase for expanding the substrate specificity and enhancing the biosynthesis of polyhydroxyalkanoate, Enzyme and microbial technology, v. 56: pp. 7-66-2014 v.56. [CrossRef]
- Choi S Y et al. (2016) One-step fermentative production of poly(lactate-co-glycolate) from carbohydrates in Escherichia coli, Nature Biotechnology, 34, 4: 435-440. [CrossRef]
- Han X, Satoh Y, Tajima K, Matsushima T, and Munekata M (2009) Chemo-enzymatic synthesis of polyhydroxyalkanoate by an improved two-phase reaction system (TPRS), Journal of Bioscience and Bioengineering, 108, 6: 517-523. [CrossRef]
- Thomson N M et al. (2013) Efficient Production of Active Polyhydroxyalkanoate Synthase in <span class="named-content genus-species" id="named-content-1">Escherichia coli</span> by Coexpression of Molecular Chaperones, Applied and Environmental Microbiology, 79, 6: 1948. [CrossRef]
- Valentin H E and Steinbüchel A (1994) Application of enzymatically synthesized short-chain-length hydroxy fatty acid coenzyme A thioesters for assay of polyhydroxyalkanoic acid synthases, Applied Microbiology and Biotechnology, 40, 5: 699-709. [CrossRef]
- de Roo G, Ren Q, Witholt B, and Kessler B (2000) Development of an improved in vitro activity assay for medium chain length PHA polymerases based on CoenzymeA release measurements, Journal of Microbiological Methods, 41, 1: 1-8. [CrossRef]
- Fukui T et al. (1976) Enzymatic synthesis of poly-β-hydroxybutyrate inZoogloea ramigera, Archives of Microbiology, 110, 2: 149-156. [CrossRef]
- Gerngross T U et al. (1994) Overexpression and Purification of the Soluble Polyhydroxyalkanoate Synthase from Alcaligenes eutrophus: Evidence for a Required Posttranslational Modification for Catalytic Activity, Biochemistry, 33, 31: 9311-9320. [CrossRef]
- Burns K L, Oldham C D, Thompson J R, Lubarsky M, and May S W (2007) Analysis of the in vitro biocatalytic production of poly-(β)-hydroxybutyric acid, Enzyme and Microbial Technology, 41, 5: 591-599. [CrossRef]
- Jaeger K E, Steinbüchel A, and Jendrossek D (1995) Substrate specificities of bacterial polyhydroxyalkanoate depolymerases and lipases: bacterial lipases hydrolyze poly(omega-hydroxyalkanoates), (in eng), Applied and environmental microbiology, 61, 8: 3113-3118. [Online]. Available: https://www.ncbi.nlm.nih.gov/pubmed/7487042. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC167586/. [CrossRef]
- Behrends A, Klingbeil B, and Jendrossek D (1996) Poly(3-hydroxybutyrate) depolymerases bind to their substrate by a C-terminal located substrate binding site, FEMS Microbiology Letters, 143, 2-3: 191-194. [CrossRef]
- Saito T, Iwata A, and Watanabe T (1993) Molecular structure of extracellular poly(3-hydroxybutyrate) depolymerase fromAlcaligenes faecalis T1, Journal of environmental polymer degradation, 1, 2: 99-105. [CrossRef]
- Ohura T, Kasuya K-I, and Doi Y (1999) Cloning and Characterization of the Polyhydroxybutyrate Depolymerase Gene of <em>Pseudomonas stutzeri</em> and Analysis of the Function of Substrate-Binding Domains, Applied and Environmental Microbiology, 65, 1: 189. [Online]. Available: http://aem.asm.org/content/65/1/189.abstract. [CrossRef]
- Papaneophytou C P, Pantazaki A A, and Kyriakidis D A (2009) An extracellular polyhydroxybutyrate depolymerase in Thermus thermophilus HB8, Applied Microbiology and Biotechnology, 83, 4: 659-668. [CrossRef]
- Hisano T et al. (2006) The Crystal Structure of Polyhydroxybutyrate Depolymerase from Penicillium funiculosum Provides Insights into the Recognition and Degradation of Biopolyesters, Journal of Molecular Biology, 356, 4: 993-1004. [CrossRef]
- Wakadkar S, Hermawan S, Jendrossek D, and Papageorgiou A C (2010) The structure of PhaZ7 at atomic (1.2 A) resolution reveals details of the active site and suggests a substrate-binding mode, (in eng), Acta Crystallogr Sect F Struct Biol Cryst Commun, 66, Pt 6: 648-654. [CrossRef]
- Kellici T F, Mavromoustakos T, Jendrossek D, and Papageorgiou A C (2017) Crystal structure analysis, covalent docking, and molecular dynamics calculations reveal a conformational switch in PhaZ7 PHB depolymerase, Proteins: Structure, Function, and Bioinformatics, 85, 7: 1351-1361. [CrossRef]
- Wang Y-L, Lin Y-T, Chen C-L, Shaw G-C, and Liaw S-H (2014) Crystallization and preliminary crystallographic analysis of poly(3-hydroxybutyrate) depolymerase from Bacillus thuringiensis, Structural biology communications, 70, 10: 1421-1423. [CrossRef]
- Mukai K, Yamada K, and Doi Y (1993) Kinetics and mechanism of heterogeneous hydrolysis of poly[(R)-3-hydroxybutyrate] film by PHA depolymerases, International Journal of Biological Macromolecules, 15, 6: 361-366. [CrossRef]
- Kasuya K-i, Inoue Y, and Doi Y (1996) Adsorption kinetics of bacterial PHB depolymerase on the surface of polyhydroxyalkanoate films, International Journal of Biological Macromolecules, 19, 1: 35-40. [CrossRef]
- Fujita M et al. (2005) Interaction between Poly[(R)-3-hydroxybutyrate] Depolymerase and Biodegradable Polyesters Evaluated by Atomic Force Microscopy, Langmuir, 21, 25: 11829-11835. [CrossRef]
- Hiraishi T, Hirahara Y, Doi Y, Maeda M, and Taguchi S (2006) Effects of Mutations in the Substrate-Binding Domain of Poly[(<em>R</em>)-3-Hydroxybutyrate] (PHB) Depolymerase from <em>Ralstonia pickettii</em> T1 on PHB Degradation, Applied and Environmental Microbiology, 72, 11: 7331. [CrossRef]
- Kasuya K-i, Ohura T, Masuda K, and Doi Y (1999) Substrate and binding specificities of bacterial polyhydroxybutyrate depolymerases, International Journal of Biological Macromolecules, 24, 4: 329-336. [CrossRef]
- Gowda U. S V and Shivakumar S (2015) Poly(-β-hydroxybutyrate) (PHB) depolymerase PHAZPenfrom Penicillium expansum: purification, characterization and kinetic studies, 3 Biotech, 5, 6: 901-909. [CrossRef]
- Shivakumar S (2013) Polyhydroxybutyrate (PHB) Depolymerase from Fusarium solani Thom, Journal of Chemistry, 2013: 9, Art no. 406386. [CrossRef]
- Shivakumar S, Jagadish S J, Zatakia H, and Dutta J (2011) Purification, Characterization and Kinetic Studies of a Novel Poly(β) Hydroxybutyrate (PHB) Depolymerase PhaZPenfrom Penicillium citrinum S2, Applied Biochemistry and Biotechnology, 164, 8: 1225-1236. [CrossRef]
- Tanio T et al. (1982) An Extracellular Poly(3-Hydroxybutyrate) Depolymerase from Alcaligenes faecalis, European Journal of Biochemistry, 124, 1: 71-77. [CrossRef]
- Madison L L and Huisman G W (1999) Metabolic Engineering of Poly(3-Hydroxyalkanoates): From DNA to Plastic, Microbiology and Molecular Biology Reviews, 63, 1: 21. [Online]. Available: http://mmbr.asm.org/content/63/1/21.abstract. [CrossRef]
- Hiraishi T, Komiya N, and Maeda M (2010) Y443F mutation in the substrate-binding domain of extracellular PHB depolymerase enhances its PHB adsorption and disruption abilities, Polymer Degradation and Stability, 95, 8: 1370-1374. [CrossRef]
- Tan L-T, Hiraishi T, Sudesh K, and Maeda M (2014) Effects of mutation at position 285 of Ralstonia pickettii T1 poly[(R)-3-hydroxybutyrate] depolymerase on its activities, Applied Microbiology and Biotechnology, 98, 16: 7061-7068. [CrossRef]
- Huisman G W, Wonink E, Meima R, Kazemier B, Terpstra P, and Witholt B (1991) Metabolism of poly(3-hydroxyalkanoates) (PHAs) by Pseudomonas oleovorans. Identification and sequences of genes and function of the encoded proteins in the synthesis and degradation of PHA, Journal of Biological Chemistry, 266, 4: 2191-2198. [Online]. Available: http://www.jbc.org/content/266/4/2191.abstract. [CrossRef]
- Schirmer A, Jendrossek D, and Schlegel H G (1993) Degradation of poly(3-hydroxyoctanoic acid) [P(3HO)] by bacteria: purification and properties of a P(3HO) depolymerase from Pseudomonas fluorescens GK13, Applied and Environmental Microbiology, 59, 4: 1220. [Online]. Available: http://aem.asm.org/content/59/4/1220.abstract. [CrossRef]
- Ramsay B A, Saracovan I, Ramsay J A, and Marchessault R H (1994) A method for the isolation of microorganisms producing extracellular long-side-chain poly (β-hydroxyalkanoate) depolymerase, Journal of environmental polymer degradation, 2, 1: 1-7. [CrossRef]
- Williamson D H, Mellanby J, and Krebs H A (1962) Enzymic determination of d(−)-β-hydroxybutyric acid and acetoacetic acid in blood, Biochemical Journal, 82, 1: 90-96. [CrossRef]
- Kobayashi T and Saito T (2003) Catalytic triad of intracellular poly(3-hydroxybutyrate) depolymerase (PhaZ1) in Ralstonia eutropha H16, Journal of Bioscience and Bioengineering, 96, 5: 487-492. [CrossRef]
- Kasuya K-i, Inoue Y, Yamada K, and Doi Y (1995) Kinetics of surface hydrolysis of poly[(R)-3-hydroxybutyrate] film by PHB depolymerase from Alcaligenes faecalis T1, Polymer Degradation and Stability, 48, 1: 167-174. [CrossRef]
- Kasuya K et al. (1997) Biochemical and molecular characterization of the polyhydroxybutyrate depolymerase of Comamonas acidovorans YM1609, isolated from freshwater, Applied and Environmental Microbiology, 63, 12: 4844. [Online]. Available: http://aem.asm.org/content/63/12/4844.abstract. [CrossRef]
- Handrick R et al. (2001) A New Type of Thermoalkalophilic hydrolase of Paucimonas lemoignei with high specificity for amorphous polyesters of short chain-length hydroxyalkanoic acids., The Journal of Biological Chemistry 276, September 28, 39: 36215-36224. [CrossRef]
- Jendrossek D (2007) Peculiarities of PHA granules preparation and PHA depolymerase activity determination, Applied Microbiology and Biotechnology, 74, 6: 1186. [CrossRef]
- Molitoris H P, Moss S T, de Koning G J M, and Jendrossek D (1996) Scanning electron microscopy of polyhydroxyalkanoate degradation by bacteria, Applied Microbiology and Biotechnology, 46, 5: 570-579. [CrossRef]
- Spyros A, Kimmich R, Briese B H, and Jendrossek D (1997) H NMR Imaging Study of Enzymatic Degradation in Poly(3-hydroxybutyrate) and Poly(3-hydroxybutyrate-co-3-hydroxyvalerate). Evidence for Preferential Degradation of the Amorphous Phase by PHB Depolymerase B from Pseudomonas lemoignei, Macromolecules, 30, 26: 8218-8225. [CrossRef]
- Oh J S, Choi M H, and Yoon S (2005) In vivo 13C-NMR spectroscopic study of polyhydroxyalkanoic acid degradation kinetics in bacteria, Journal of Microbiology and Biotechnology, 15: 1330-1336.
- Taidi B, Mansfield D A, and Anderson A J (1995) Turnover of poly(3-hydroxybutyrate) (PHB) and its influence on the molecular mass of the polymer accumulated by Alcaligenes eutrophus during batch culture, FEMS Microbiology Letters, 129, 2-3: 201-205. [CrossRef]
- De Eugenio L I et al. (2006) Biochemical evidence that phaZ gene encodes a specific intracellular medium chain length polyhydroxyalkanoate depolymerase in Pseudomonas putida KT2442: characterization of a paradigmatic enzyme, Journal of Biological Chemistry. [CrossRef]
- Tokiwa Y and Calabia B P (2004) Review Degradation of microbial polyesters, Biotechnology Letters, 26, 15: 1181-1189. [CrossRef]
- Sharma P K, Mohanan N, Sidhu R, and Levin D B (2019) Colonization and degradation of polyhydroxyalkanoates by lipase-producing bacteria, Canadian Journal of Microbiology, 65, 6: 461-475. [CrossRef]
- Mohamed R A, Salleh A B, Leow A T C, Yahaya N M, and Abdul Rahman M B (2017) Ability of T1 Lipase to Degrade Amorphous P(3HB): Structural and Functional Study, Molecular Biotechnology, 59, 7: 284-293. [CrossRef]
- Mok P-S, Ch’ng D H-E, Ong S-P, Numata K, and Sudesh K (2016) Characterization of the depolymerizing activity of commercial lipases and detection of lipase-like activities in animal organ extracts using poly(3-hydroxybutyrate-co-4-hydroxybutyrate) thin film, AMB Express, 6, 1: 97. [CrossRef]
- Ch’ng D H-E and Sudesh K (2013) Densitometry based microassay for the determination of lipase depolymerizing activity on polyhydroxyalkanoate, AMB Express, 3, 1: 22. [CrossRef]
- Rodríguez-Contreras A, Calafell-Monfort M, and Marqués-Calvo M S (2012) Enzymatic degradation of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) by commercial lipases, Polymer Degradation and Stability, 97, 4: 597-604. [CrossRef]
- Mendes A A, Oliveira P C, Vélez A M, Giordano R C, Giordano R d L C, and de Castro H F (2012) Evaluation of immobilized lipases on poly-hydroxybutyrate beads to catalyze biodiesel synthesis, International Journal of Biological Macromolecules, 50, 3: 503-511. [CrossRef]
- Yang T H, Kwon M-A, Lee J Y, Choi J-E, Oh J Y, and Song J K (2015) In Situ Immobilized Lipase on the Surface of Intracellular Polyhydroxybutyrate Granules: Preparation, Characterization, and its Promising Use for the Synthesis of Fatty Acid Alkyl Esters, Applied Biochemistry and Biotechnology, 177, 7: 1553-1564. [CrossRef]
- Houten S M and Wanders R J A (2010) A general introduction to the biochemistry of mitochondrial fatty acid β-oxidation, Journal of Inherited Metabolic Disease, 33, 5: 469-477. [CrossRef]
- Chung A-L et al. (2011) Biosynthesis and Characterization of Poly(3-hydroxydodecanoate) by β-Oxidation Inhibited Mutant of Pseudomonas entomophila L48, Biomacromolecules, 12, 10: 3559-3566. [CrossRef]
Figure 1.
Types of bioplastics modified from [22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39].
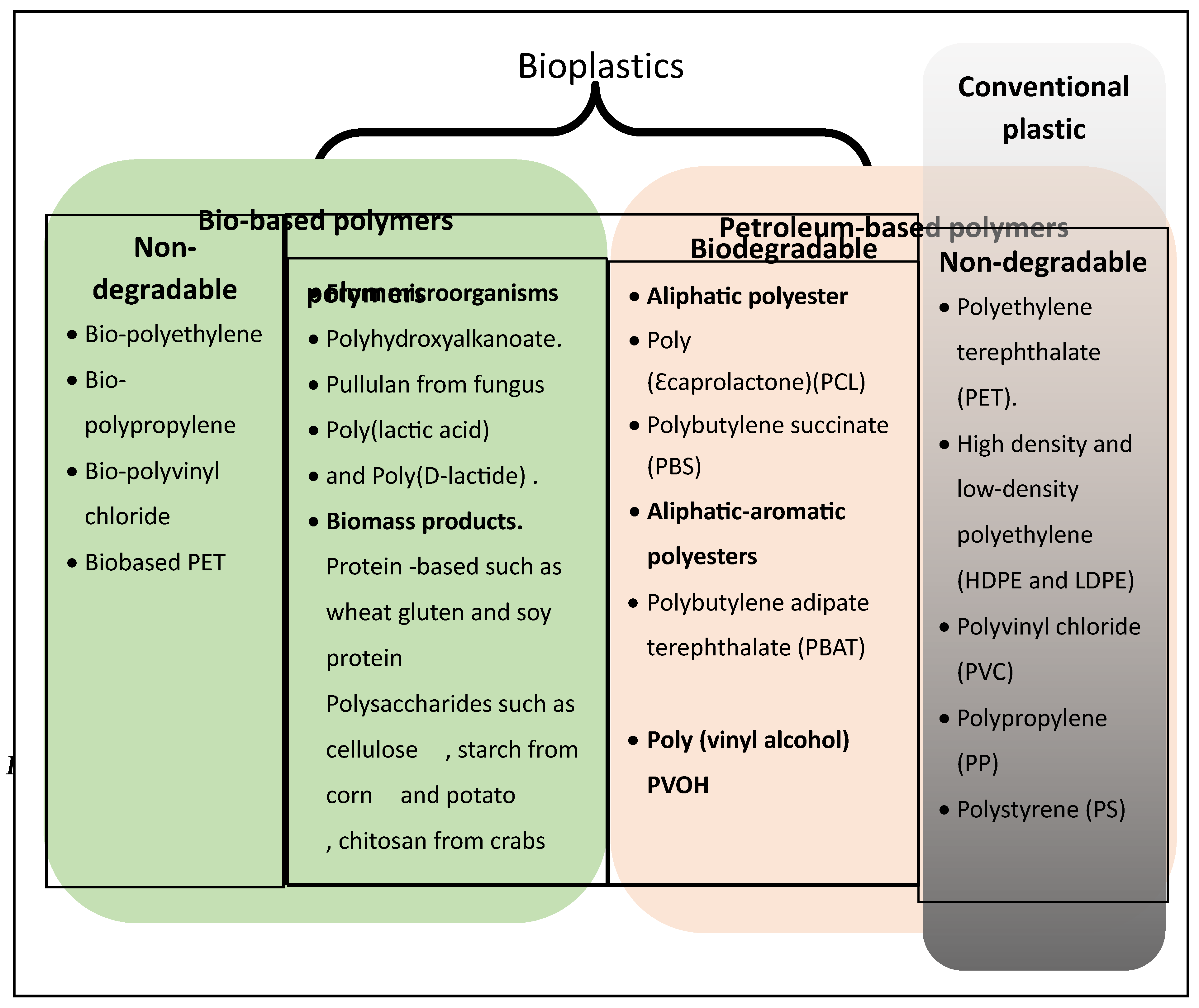
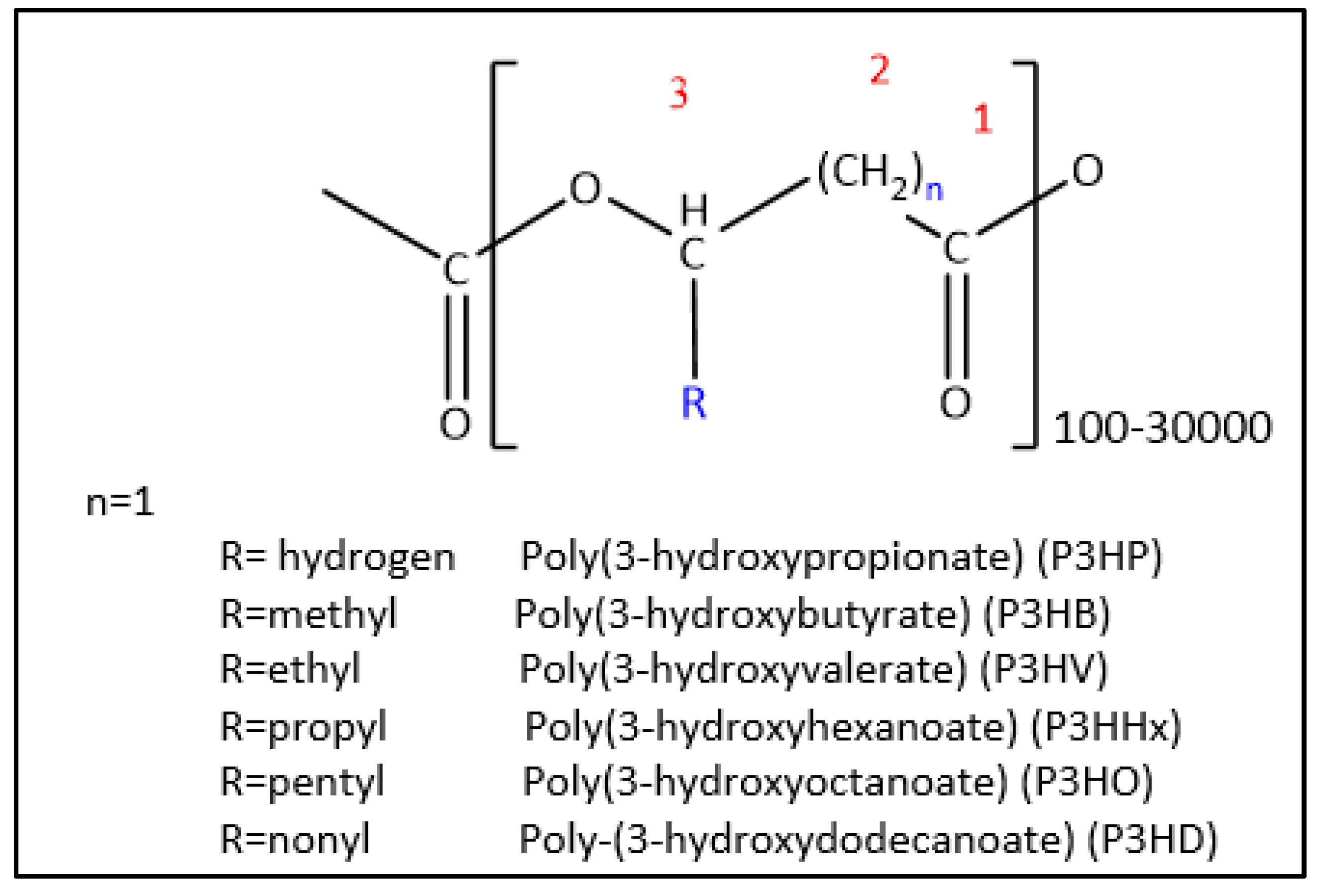
Figure 3.
Representation of the β-oxidation pathway and β-oxidation pathway-related genes. Genes circled in red have promoted PHAs accumulation when inactivated in several studies (Refer to Table 2). Genes circled in green have promoted PHAs accumulation when overexpressed (Refer to Table 2).
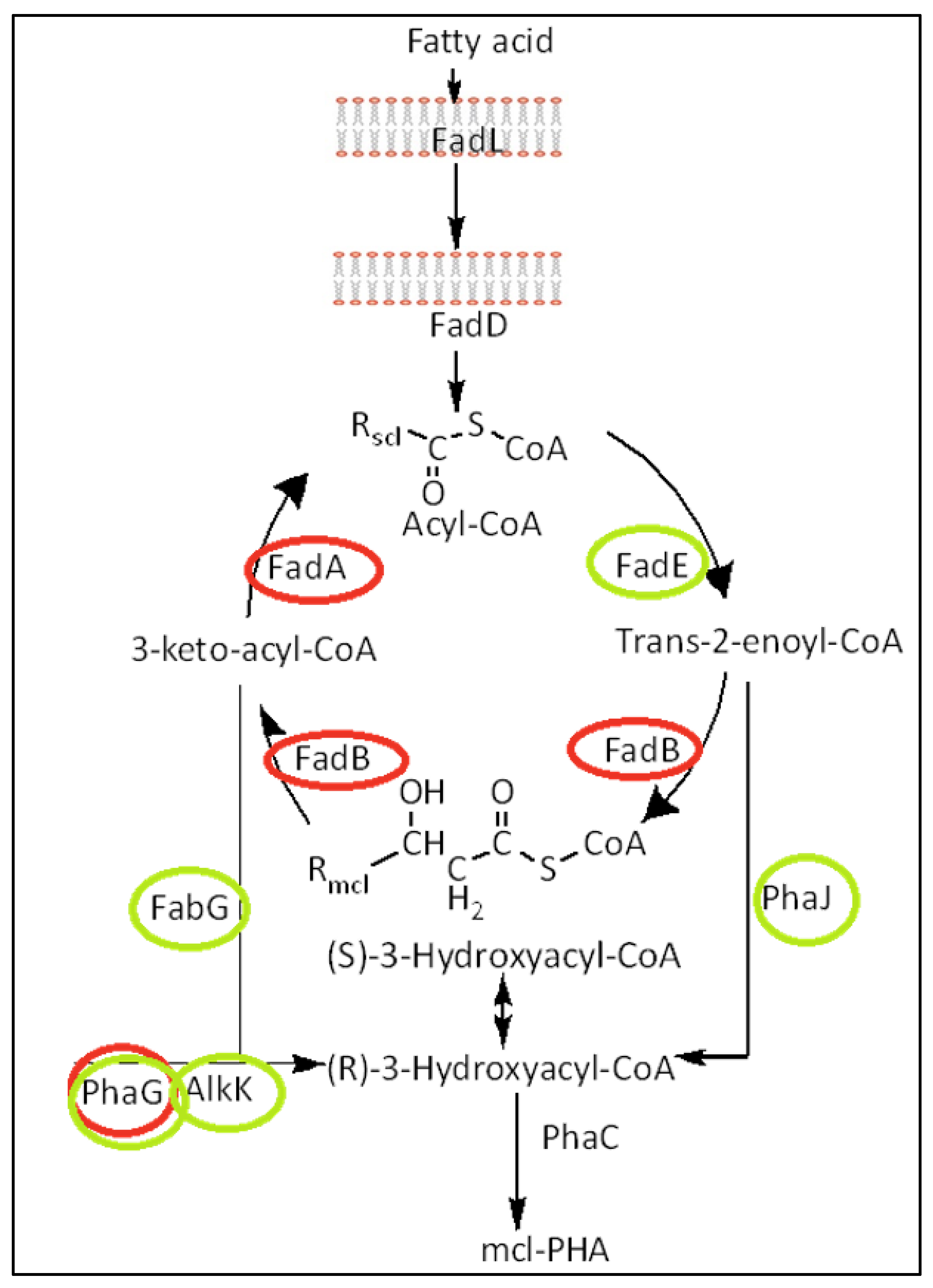
Figure 4.
A) The catalytic domain of PHA synthase from C. necator. Catalytic triad is presented in green. [114] Molecule modelled in PyMOL for this work [117]. (5T6O Protein Data Bank elucidated using x-ray diffraction) B) Pymol structure of 3 aligned PhaCs (PmePhaC, PpuPhaC and PstzPhaC) from P. mendocina NK-01, P. putida KT2440 and P. stutzeri 1317 respectively. The catalytic triad Cys, Asp and His are coloured.
Figure 4.
A) The catalytic domain of PHA synthase from C. necator. Catalytic triad is presented in green. [114] Molecule modelled in PyMOL for this work [117]. (5T6O Protein Data Bank elucidated using x-ray diffraction) B) Pymol structure of 3 aligned PhaCs (PmePhaC, PpuPhaC and PstzPhaC) from P. mendocina NK-01, P. putida KT2440 and P. stutzeri 1317 respectively. The catalytic triad Cys, Asp and His are coloured.
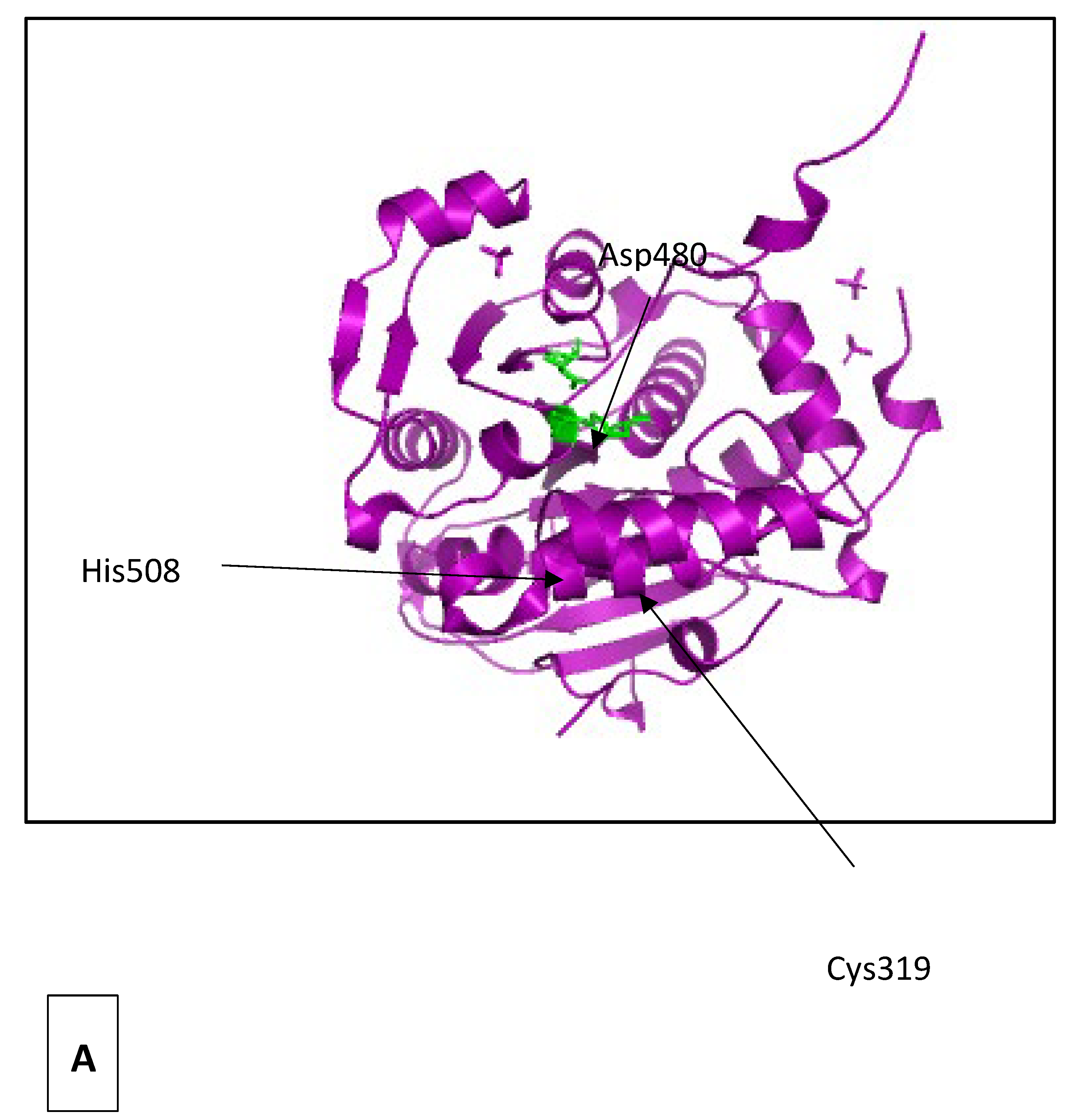
Figure 5.
Mutation map of Pseudomonas putida. The catalytic triad is squared in blue. Lab tested mutations are represented in blue while blast search mutations are written in red. If more than one amino acid is possible in that position then the position is represented by the + + symbol.
Figure 5.
Mutation map of Pseudomonas putida. The catalytic triad is squared in blue. Lab tested mutations are represented in blue while blast search mutations are written in red. If more than one amino acid is possible in that position then the position is represented by the + + symbol.
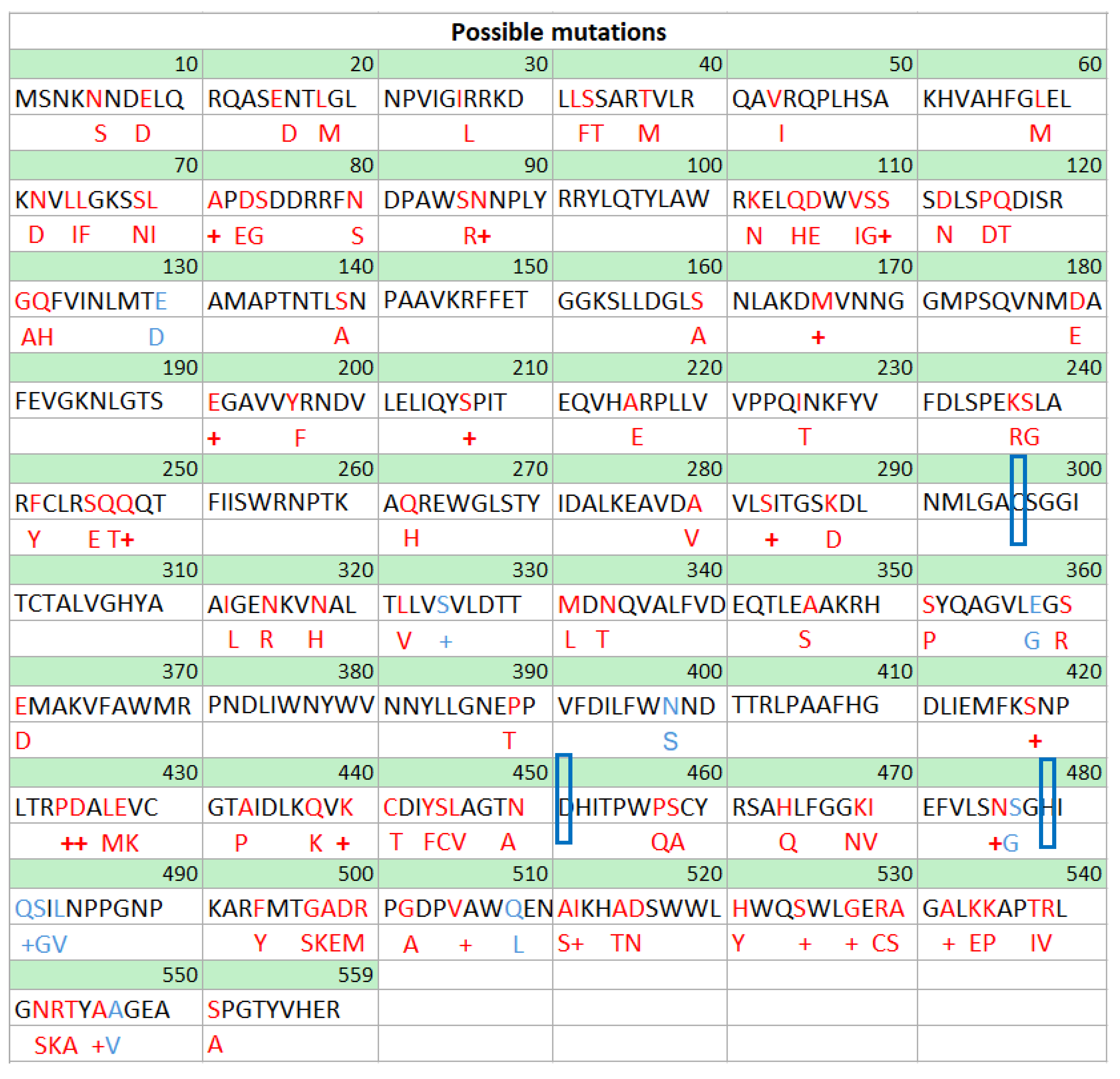
Figure 6.
Classification of PHA depolymerases.
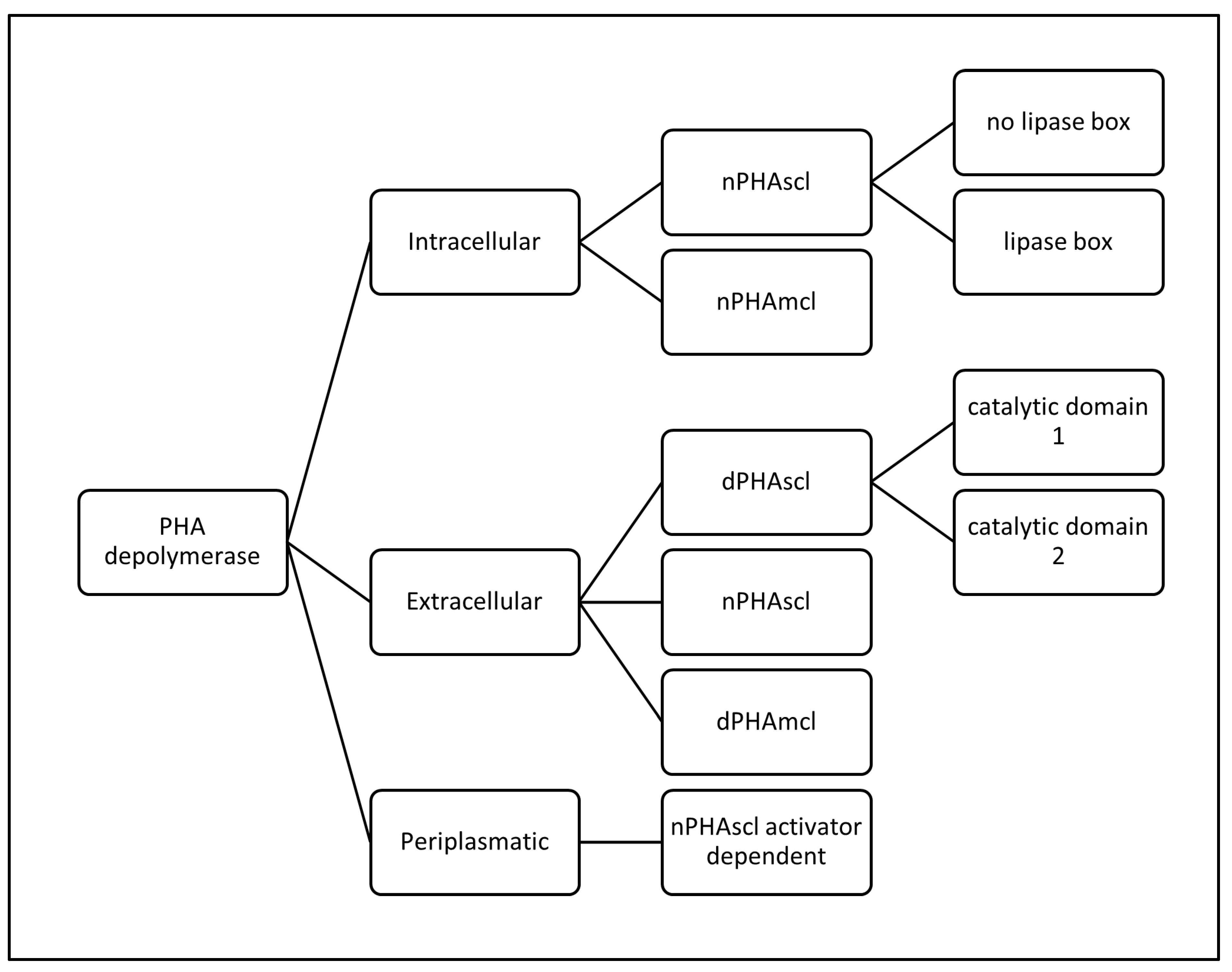
Figure 7.
Regions of a PHA depolymerase and their classifications.
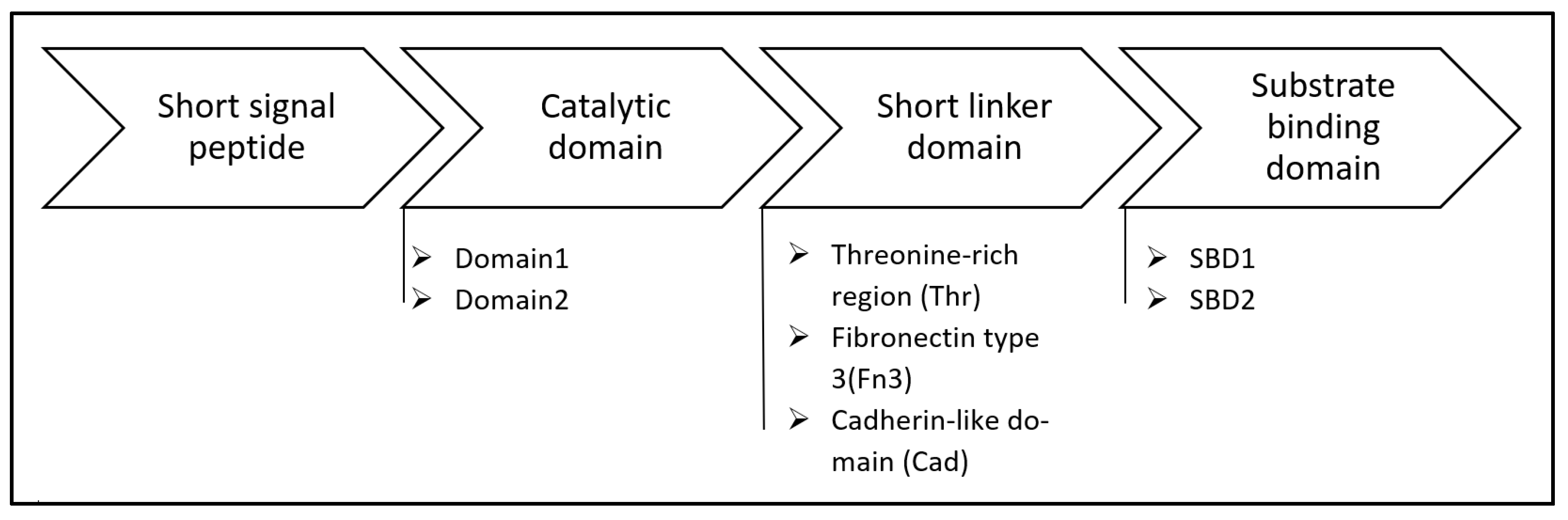
Figure 8.
dPHAscl type II from Penicillium funiculosum. The enzyme has 319 amino acid residues with a molecular mass of 33569 Da. The enzyme has a α/β hydrolase fold where the catalytic residues are (marked in pink). [171] Molecule modelled in PyMOL for this work [117]. (2D80 from Protein Data Bank elucidated using x-ray diffraction).
Figure 8.
dPHAscl type II from Penicillium funiculosum. The enzyme has 319 amino acid residues with a molecular mass of 33569 Da. The enzyme has a α/β hydrolase fold where the catalytic residues are (marked in pink). [171] Molecule modelled in PyMOL for this work [117]. (2D80 from Protein Data Bank elucidated using x-ray diffraction).
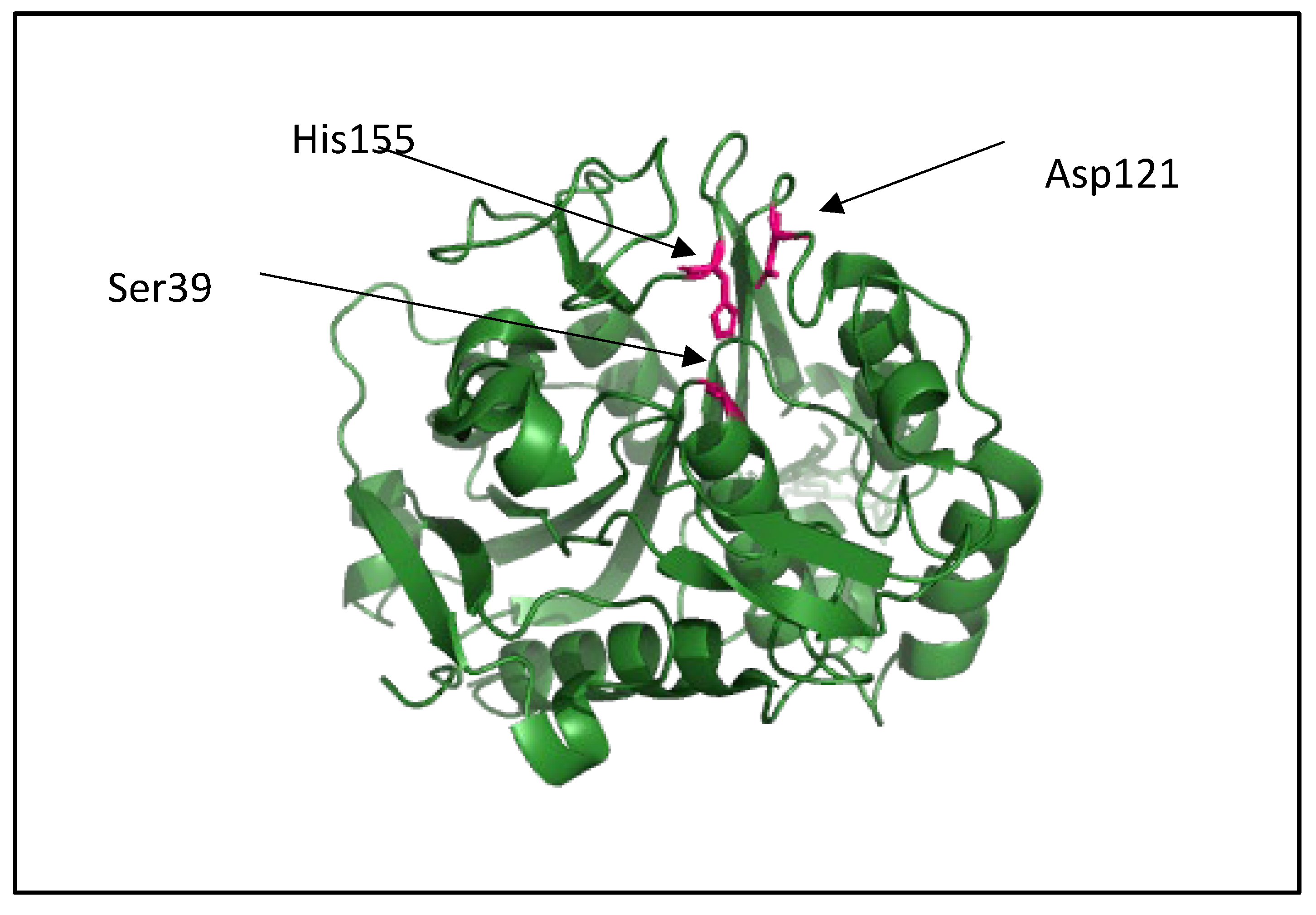
Figure 9.
Polyhydroxyalkanoate polymers are degraded to hydroxyalkanoic acid monomers. Modelled using ChemDraw [61].
Figure 9.
Polyhydroxyalkanoate polymers are degraded to hydroxyalkanoic acid monomers. Modelled using ChemDraw [61].
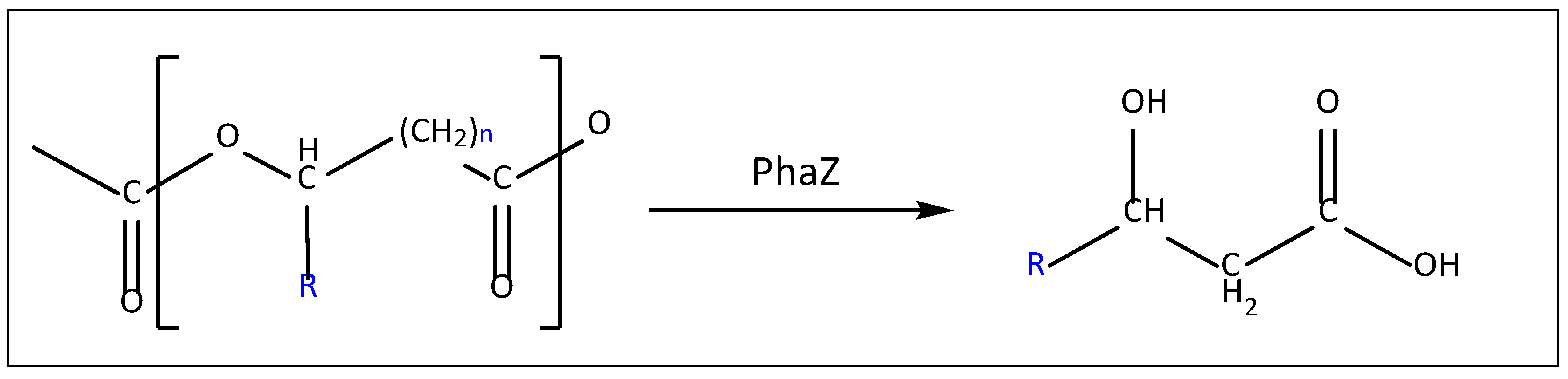
Figure 10.
Schematic model of the interaction of the substrate-binding domain with the PHB (Taken from Hiraishi, 2006 [178]). Dashed bonds are used to represent hydrogen bonds whereas hydrophobic interactions are circled.
Figure 10.
Schematic model of the interaction of the substrate-binding domain with the PHB (Taken from Hiraishi, 2006 [178]). Dashed bonds are used to represent hydrogen bonds whereas hydrophobic interactions are circled.
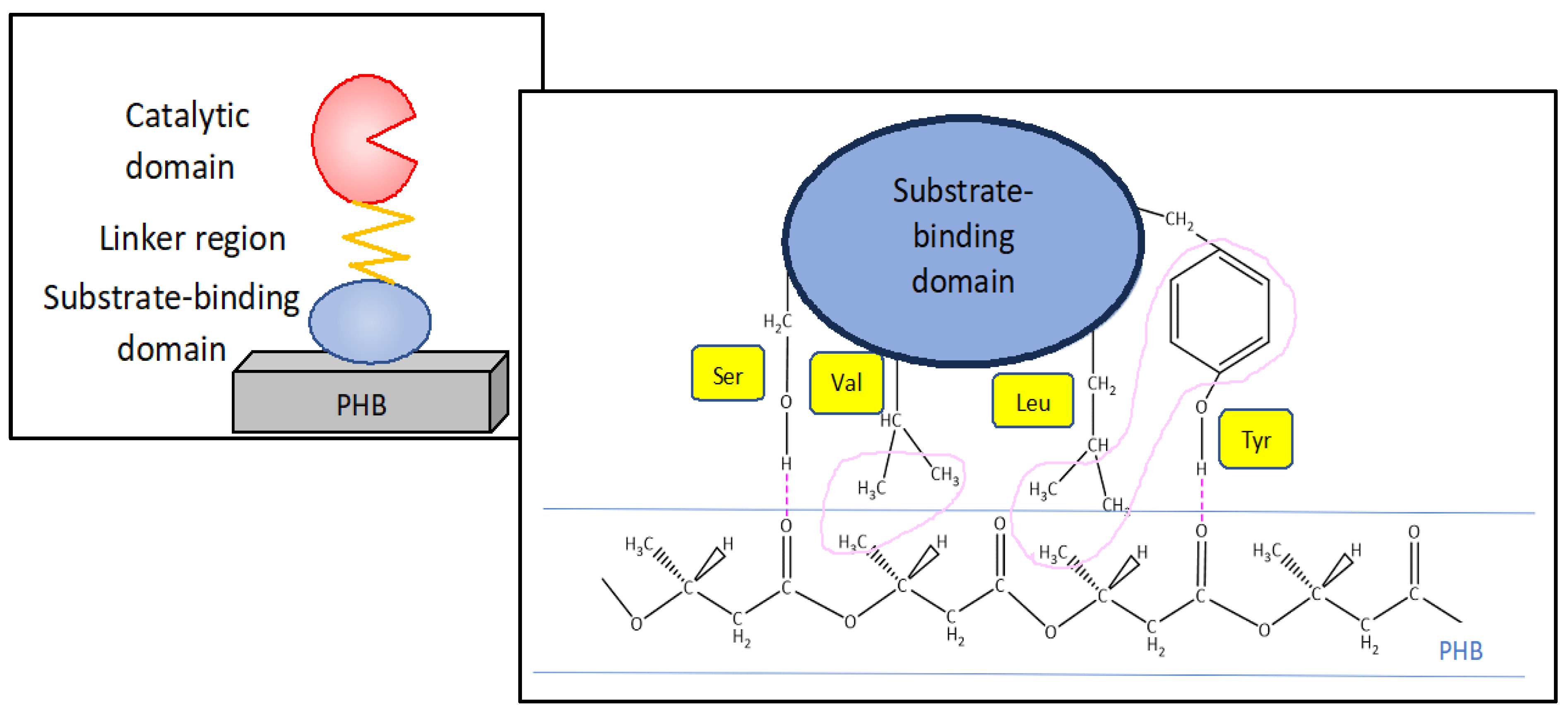

Table 1.
Comparison of the thermophysical characteristics between short-chain-length and medium-chain-length PHAs.
Table 1.
Comparison of the thermophysical characteristics between short-chain-length and medium-chain-length PHAs.
| Short-chain-length PHAs | Medium-chain-length PHAs |
|---|---|
| Highly crystalline [76,77] | Low crystallinity [76,77] |
| Hard [76,77] | Soft [76,77] |
| Brittle [76,77] | Flexible [76,77] |
| Thermo-elastomeric polyesters [76,77] | |
| Low glass transition and melting temperature [76,78] | |
| Low tensile strength and modulus [76,78] | |
| Higher elongation at break [76,78] |
Table 2.
mcl-PHAs accumulation enhanced by metabolic engineering of the β-oxidation pathway and the β-oxidation pathway-related enzymes. Inactivated genes are presented in red, overexpressed genes are presented in green.
Table 2.
mcl-PHAs accumulation enhanced by metabolic engineering of the β-oxidation pathway and the β-oxidation pathway-related enzymes. Inactivated genes are presented in red, overexpressed genes are presented in green.
| Organism used | Β-oxidation pathway modification | PHAs yield | % of mcl-monomer | Substrate used | |
|---|---|---|---|---|---|
| Ralstonia eutropha | Heterologous expression of phaC from, Rhodococcus aetherivorans I24, Expression of phaJ from P. aeruginosa to increase PHAs accumulation, Modification of the phaB activity |
71 wt% 66 wt% |
P(HB-co-17%-HHx) P(HB-co-30%-HHx) |
Palm oil | [92] |
| Cupriavidus necator (Ralstonia eutropha) | phaC from R. aetherivorans I24 and phaJfrom P. aeruginosa expressed in plasmid pCB113 | 45 wt% 1.3 g/l PHAs |
P(HB-co-70%HHx) | Crude palm kernel oil | [93] |
| Ralstonia eutropha | Introduction of crotonyl-CoA reductase from Streptomyces cinnamonensis, phaC and phaJ from A. caviae | 48 wt% 1.48 g/l CDW |
P(HB-co-1.5%HHx) | Fructose | [94] |
| Ralstonia eutropha | Overexpression of enoyl coenzyme-A hydratase (phaJ) and phaC2. Deletion of acetoacetyl Co-A reductases (phaB1, phaB2 and phaB3) | 69 wt% | P(78%HB-co-22%HHx) | Coffee waste oil | [95] |
| Pseudomonas putida KCTC1639 | Overexpression of phaJ Overexpression offabG |
27wt% 0.51 g/l PHAs (FabG overexpression depressed PHAs production) |
NI | Octanoic acid | [90] |
| Cupriavidus necator | Expression of fadE from E. coli and phaJ1 from P. putida KT2440, in plasmid pMPJAS03 | 46.1wt%, 4.1 g/l CDW 38.3 wt%, 3.28 g/l CDW |
P(99%HB-co-0.37%HV-co-0.27%HHx-co-0.21%HO-co-0.08%HD) P(99.39%HB-co-0.33%HV-co-0.18%HHx-co-0.10%HO) |
Canola oil Avocado oil |
[91] |
| DH5α Escherichia coli | Deletion of key genes in β-oxidation pathway and overexpression of acyl-ACP thioesterase (BTE), phaJ3 and phaC2 from P. aeruginosa PAO1 and PP_0763 from P. putida KT2440 strain (fadRABIJ) | 0.93 g/l CDW 0.75 wt% |
P(100%HDD) | Decanoic acid | [96] |
|
Escherichia coli W3110 Escherichia coli WA101(fadA mutant) |
Expression offabGfrom E. coli and phaC2 from Pseudomonas sp 61-3 Expression of rhlG from P. aeruginosa and PhaC2 from Pseudomonas. sp 61-3 Expression of fabG from E. coli and phaC2 from Pseudomonas. sp 61-3 |
4.8 wt%, 1.73g/l CDW, 0.08g/l PHAs 3.2wt%, 1.20g/l CDW, 0.04g/l PHAs 22.1wt%, 0.98g/l CDW, 0.22g/l PHAs |
P(11%HHx-co-39%HO-co-50%HD) P(47%HO-co-53%HD) P(7%HO-co-93%HD) |
Sodium decanoate |
[97] |
| Cupriavidus necator | 5 different transformants harboring phaC BP-M-CPF4 gene. (phaJ from P. aeruginosa expression increased HHx proportion). (phaB and phaA genes expression modify composition) |
(48.9-83.7)wt% (3.6-6.2)CDW (2.1-1.4)g/l PHAs |
HHx(1-18%) | Palm olein Palm kernel oil |
[98] |
| Pseudomonas putida KT2440 | Deletion of phaZ, fadBA1andfadBA2 and overexpression of phaG, alkK, phaC1 and phaC2 (strain AG2162) | 1.758 g/l CDW 54 wt% 0.657g/l CDW, 17.7 wt% |
No information | P-coumaric acid Lignin(corn stover) |
[99] |
| Pseudomonas putida KT2440 | No (acrylic acid inhibits β-oxidation pathway) | 75.5 wt% 1.8 g/lh PHAs |
P(89%HHp-co-11%HN) | nonanoic acid: glucose: acrylic acid (1.25:1:0.05) | [89] |
| P. putida KT2442 | Deletion of: fadB2x, fadAx, fadB, fadA, 3-hydroxyacyl-CoA deshydrogenase, acyl-CoA dehydrogenase and phaG. | 9.19 wt% 1.03 g/l CDW |
P(3HD-co-84%-3HDD) | Decanoic acid Dodecanoic acid |
[87] |
| P. putida KT2442 | Deletion of fadB and fadA, Deletion of phaC and replacement with phaPCJAC operon (KTOYO6ΔC (phaPCJAC) strain) | 5.82 g/l CDW 57.80 wt% |
58%PHB-block-42%PHHx | sodium butyrate, sodium hexanoate (1:2) alternating times | [100] |
| Pseudomonas entomophila LAC32 | Weakening of β-oxidation pathway (Deletion of fadA(x), fadB(x) and phaG) Insertion of phaA and phaB from C. necator. PhaC mutated from P. putida 61-3. Block of some genes in de novo fatty acid synthesis pathway |
Not reported | different rations with mcl from 0 to 100% | Glucose and related fatty acids | [101] |
| Pseudomonas putida KT2442 | fadA and fadB knockout mutant | 84 wt% | P(41%HDD-co-59%HA) | Dodecanoate | [102] |
| Pseudomonas mendocina NK-01 |
fadA and fadB knockout mutant (NKU-∆β5) fadA, fadB, phaG, phaZ knockout mutant (NKU-∆8) NKU-∆8. |
38 wt%, 1.7 g/l CDW 44 wt%, 2.3 g/l CDW 32 wt%, 1.3 g/l CDW |
P(5.57%HHx-co-93.3%HO-co-1.05%HD) P(5.29%HO-co-94%HD) P(2.99%HO-co-28%HD-co-68%HDD) |
Sodium octanoate Sodium decanoate Dodecanoic acid |
[103] |
| Pseudomonas putida KT2442 | Deletion of fadA, fadB (P. putida KTOY06) | 2.86 g/l CDW 45.99 wt% |
P(2.2%HHx-co-11%HO-co-21.6%HD-co-16.1%HDD-co-49%HTD) | Tetradecanoic acid | [104] |
Scl: short-chain-length, mcl: medium-chain- length, PHA: poly(3-hydroxyalkanoate), PLA: polylactate, PGA: polyglycolate, PHB: poly(3-hydroxybutyrate), PHP: poly(3-hydroxypentanoate), PHHx: poly(3-hydroxyhexanoate), PHO: poly(3-hydroxyoctanoate), PHD: poly(3-hydroxydecanoate), PHDD: poly(3-hydroxydodecanoate).
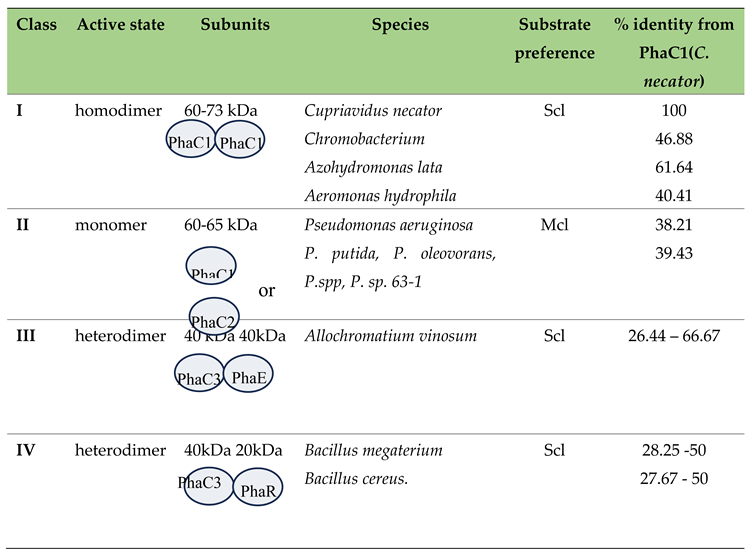
Table 4.
Point mutations in Pseudomonas that affect substrate specificity.
| Source PhaC | Mutation | Host | Carbon source | Effect | |
|---|---|---|---|---|---|
| P. stutzeri 1317 | Ser325Thr Gln481Lys |
Ralstonia eutropha PHB¯4 | Gluconate and Octanoate | Higher PHAs content and more affinity for PHB. | [130] |
| Pseudomonas 61-3 | Ser325Cys Ser325Thr Glu481Lys Glu481Met Glu481Arg |
Ralstonia eutropha PHB¯4 | Fructose | Higher activity. The combination Ser325Cys with Glu481Met has higher activity but all combination worked. Mutation in position 481 produces higher molecular weight. (98-99% of short-chain-length) | [136] |
| P. putida GPo1 | Leu484Val Ala547Val Gln481Met Ser482Gly |
P. putida GPp104 PHAs¯ | Sodium octanoate | Leu484Val increase PHB monomer content Ala547Val increase PHAs content. Gln481Met increases HHx monomer and PHAs content Ser482Gly increases HHx monomer and PHAs content. |
[137] |
| Pseudomonas putida KT2440 | Glu358Gly Asn398Ser |
Β-oxidation deficient E. coli LSBJ | Sodium dodecanoate | Higher PHDD production with higher molecular weight. Amino acids with hydrophobic and smaller residues either retained or increased PHDD. | [138] |
| Pseudomonas sp. 61-3 | Ser325Cys ---TGC Ser325Thr---ACC Gln481Met Gln481Lys--- AAG Gln481Arg---CGG |
E. coli | Glucose | Double mutants increase PHB content. Codon picking is very important to enhance PHB content. The mutation Ser325Thr, which exists in PhaC type I, increases affinity for PHB. | [139] |
|
Cupriavidus necator Aeromonas caviae |
26% of the N-terminal of PhaCAC and 74% of the C-terminal of PhaCRe |
E. coli LSS218 | Sodium dodecanoate | Using sodium dodecanoate, they obtained higher scl-mcl PHAs than the parental enzymes. | [140] |
|
P. chlororaphis P. sp. 61-3 P. putida KT2440 P. resinovorans P. aeruginosa PAO1 |
Glu130Asp Ser325Thr Ser477Gly Gln481Lys |
E. coli XL1-Blue | Glucose Lactate |
PhaC having mutations in these 4 sites were able to accept lactyl-CoA as substrate and produce PLA while wild types did not accumulate polymers. | [141] |
| P. sp. MBEL 6-19 | Glu130Asp Ser325Thr Gln481Met |
E. coli XL1-Blue | Glucose | Increase the substrate specificity towards scl. Production pf P(HB-co-LA) | [142] |
| Pseudomonas stutzeri | Glu130Asp Ser325Thr Ser477Gly Gln481Lys |
E. coli | Lactic acid | Terpolyesters (LA-co-3HB-3HP) Change of substrate specificity. |
[143] |
| Pseudomonas sp.61-3 | Gln508Leu Gln481Lys |
E. coli | Mutation Gln508Leu is present in C. necator. Double mutation enhances PHB content. |
[144] |
Table 5.
Studies of PhaC enhancement through synthetic PHA synthase evolution.
| Method | PhaC source | Changed enzyme property | Polyester | |
|---|---|---|---|---|
| Diversify PCR random mutagenesis | Pseudomonas putida KT2440 | Higher yield with higher molecular weight | P3HDD | [138] |
| Error-prone PCR mutagenesis, saturation mutagenesis, in vitro recombination | Pseudomonas sp. 61-3 | Higher yield | P3HB | [139] |
| Site-specific mutagenesis | Pseudomonas sp. SG4502 | Substrate specificity | P(LA-co-3HB) | [150] |
| Localized semi-random mutagenesis | Pseudomonas oleovorans GPo1 | Substrate specificity | P3HB, P3HA | [151] |
| Gene shuffling | C. necator | Higher yield, substrate specificity | P3HA | [152] |
| PCR-mediated random mutagenesis, intragenic suppression-mutagenesis | Ralstonia eutropha | Yield | P3HB | [153] |
| Chimeric PHA synthase |
P. oleovorans P. fluorescens P. aureofaciens |
Higher yield, substrate specificity | scl/mcl | [154] |
| Recombination of beneficial mutations | Aeromonas caviae | Molecular weight, fraction composition | P(3HB-co-3HHx) | [155] |
| Chimeric PHA synthase |
Aeromonas caviae C. necator |
Substrate specificity | P(3HB-co-3HHx-co-3HO) | [156] |
| Chimeric PHA synthase |
A. caviae C. necator |
Substrate specificity | 3HHx / 3HB | [140] |
| Site specific mutagenesis | Pseudomonas sp. S64502 | Substrate specificity | P(LA-co-3HB) | [150] |
| Site specific mutagenesis |
P. chlororaphis P.sp.61-3 P. putida KT2440 P. resinovorans P. aeruginosa PAC1 |
Substrate specificity | P(3HB-co-LA) | [141] |
| Site specific mutagenesis Saturation mutagenesis |
Pseudomonas sp. MBEL 6-19 | Substrate specificity | P(3HB-co-LA) | [142] |
| Site specific mutagenesis, saturation mutagenesis | Pseudomonas putida GPo | Yield, substrate specificity | Copolymers with 3HB, 3HHx, 3HO, 3HD | [157] |
| None (natural mutation) | Pseudomonas MBEL 6-19 | Substrate specificity | P(LA-co-GA-co-3HB) P(LA-co-GA-co-4HB), P(LA-co-GA-2HB) | [158] |
| Site specific mutagenesis | Pseudomonas stutzeri | Substrate specificity | P(LA-co-3HB-co-3HP) | [143] |
| PCR- mediated random mutagenesis Recombination of beneficial mutations. |
Pseudomonas sp.61-3 | Yield | P3HB | [144] |
Scl: short-chain-length, mcl: medium-chain- length, P3HA: poly(3-hydroxyalkanoate), PLA: polylactate, PGA: polyglycolate, P3HB: poly(3-hydroxybutyrate), P3HP: poly(3-hydroxypentanoate), P3HHx: poly(3-hydroxyhexanoate), P3HO: poly(3-hydroxyoctanoate), P3HD: poly(3-hydroxydecanoate), P3HDD: poly(3-hydroxydodecanoate).
Table 6.
Different methods to analyse PhaC activity.
| Method | Advantages | Disadvantages | Reference |
|---|---|---|---|
| CoA detection 412 nm | Quick | False positives due to other hydrolysis reactions that release CoA. | [70,161,162] |
| Thioesters hydrolysis detection 236 nm | Quick | Less accurate than CoA detection. | [70,163,164] |
| Substrate depletion and product formation analysis with HPLC and GC | Highly accurate | More time consuming | [131] |
| Co-A, acetyl-CoA and β-hydroxybutyryl-CoA analysis with HPLC | It is accurate and measures fluctuation in the overall system | Costly and time consuming | [165] |
Table 7.
PHA depolymerases and their kinetic parameters using PHB as a substrate.
| Organism | Type | Apparent Km(µg/ml) | Apparent Vmax(µg/min) | |
|---|---|---|---|---|
| Penicillium expansum | ePHAscl | 1.04 | 4.5 | [180] |
| Thermus thermophilus | edPHAscl-typeI | 53 | N/A | [170] |
| Fusarium solani | ePHAscl | 100 | 50 | [181] |
| Penicillium citrinum | ePHAscl | 1250 | 12.5 | [182] |
| Alcaligenes faecalis | edPHAscl-typeI | 13.3 | N/A | [183] |
Table 8.
Comparison of common methods to evaluate PhaZ activity.
| Method | Advantages | Disadvantages | Reference |
|---|---|---|---|
| Sudan Black observation | Easy and quick | Not quantitative result | [187] |
| Halo formation in plate assay | Easy and quick, for extracellular PhaZ only. | Semi-quantitative result | [6,188,189] |
| Measurement of 3HB release | Quantitative method | No information | [190,191,192,193] |
| Measure esterase activity at 410 nm | Quantitative method | No information | [170,188] |
| Measurement of PHAs weight lost | Easy | Not practical for rapid routine assays | [194,195] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



