Submitted:
21 November 2024
Posted:
25 November 2024
You are already at the latest version
Abstract
Calcium (Ca) is an essential nutrient for plant growth and development, playing a key role in cell wall stabilization and various physiological processes. Ca fertilizers are widely used in agriculture to meet crop demands and improve yield and quality. However, traditional Ca fertilizers often suffer from low solubility, poor absorption, and mobility issues. Chelated Ca fertilizers offer enhanced efficiency and uptake. In this study, we compared the effects of lignosulfonate-chelated Ca (LS-Ca), EDTA-chelated Ca (EDTA-Ca), Ca(NO3)2, and alcohol sugar-chelated Ca (AS-Ca) on tomato growth. The results showed that LS-Ca increased chlorophyll a and b contents by 26% and 46%, respectively. The application of Ca fertilizers significantly enhanced Ca2+ uptake and transport, with LS-Ca achieving the highest efficiency. Without altering fruit weight, LS-Ca treatment increased the firmness of mature tomato fruits by 29%. Furthermore, LS-Ca improved fruit sweetness by 33%, with total sugar content increasing by 45%, sucrose by 80%, reducing sugars by 64%, and titratable acidity by 18%. This study aims to compare the effects of different chelated Ca fertilizers on tomato cultivation and to explore optimal Ca supplementation strategies, thereby contributing to the improvement of tomato cultivation practices and fruit quality.
Keywords:
Tomato
; calcium
; chelated calcium
; calcium uptake and transport
; fruit quality
1. Introduction
Ca is an essential nutrient for plant growth and development. It enhances the rigidity and stability of cell walls, participates in cell division and elongation, and serves as a key component of cell walls and membranes [1,2]. Acting as a secondary messenger, Ca is involved in signal transduction for various physiological processes, such as hormone regulation and stress responses, and regulates enzyme activities that affect plant metabolism [3].
Ca deficiency can lead to symptoms such as apical necrosis, leaf curling, poor root development, and delayed fruit growth. In agricultural production, Ca fertilizers can neutralize soil acidity, improve soil pH and structure, promote beneficial microbial activity, and enhance fruit firmness, shelf life, and resistance to physiological disorders like fruit cracking and blossom-end rot. Additionally, Ca application improves crop yield and quality and enhances resistance to diseases, drought, and salt stress [4]. Inorganic Ca fertilizers, such as Ca(NO₃)₂, CaCO3, CaSO4, and Ca3(PO4)2, are widely used due to their high purity, fast action, multifunctionality, and affordability. However, challenges remain, including soil acidification, rapid dissolution, uneven crop absorption, and environmental sensitivity [5,6,7].
Chelated Ca fertilizers, which stabilize Ca2+ using chelating agents, offer higher bioavailability and stability. EDTA-chelated Ca (EDTA-Ca), for example, binds Ca2+ to ethylenediaminetetraacetic acid (EDTA) and promotes flowering in tomatoes, although excessive application can cause chlorosis and necrosis in older leaves [8,9]. Polyols, such as xylitol and mannitol, also serve as carriers for Ca2+, improving its mobility and uptake within plants. Foliar application of sorbitol-Ca fertilizer, for instance, enhances Ca2+ absorption and boosts peanut yields [10], while alcohol sugar-chelated Ca (AS-Ca) treatments significantly improve grape berry size and shape index [11].
Lignin, a natural polymer in plant cell walls, exhibits excellent chelating ability. Lignosulfonates, derived from sulfite pulping processes, are low-cost materials containing hydrophilic sulfonate and electroactive methoxy phenol groups, offering potential for improving nutrient bioavailability and reducing production costs [12,13]. Chelated Ca fertilizers, including lignosulfonate-Ca (LS-Ca), have demonstrated superior Ca2+ transport, enhanced crop growth, and improved disease resistance.
Tomatoes, with their increasing yield and quality demands, are particularly sensitive to Ca nutrition. Ca deficiency in tomatoes causes disorders such as blossom-end rot, necrosis of growth points, and abnormal floral structures, severely impacting yield and quality. Ca absorption depends on soil Ca²⁺ availability and xylem transport driven by transpiration. During early fruit development, both xylem and phloem contribute to nutrient flow, but as fruits mature, reduced transpiration and xylem function shift reliance entirely to phloem, leading to Ca²⁺ dilution in tissues [14,15]. Enhancing Ca²⁺ transport in soluble chelated forms can effectively address late-stage deficiencies.
To evaluate the effectiveness of different chelating agents on Ca supplementation in tomatoes, we used Mic-TOM as the experimental material and applied foliar sprays and root irrigation of LS-Ca, EDTA-Ca, Ca(NO₃)₂ and AS-Ca. We systematically analyzed plant growth and fruit quality differences and monitored Ca²⁺ distribution across tissues during different developmental stages. This study aims to identify effective Ca supplementation strategies for tomato cultivation, providing technical guidance for improving tomato quality and productivity.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
Tomato cultivar ‘Micro-TOM’ deeds were germinated and sown in greenhouse soil. The greenhouse conditions were maintained at a 16 h/8 h light/dark cycle with temperatures of 25°C during the day and 20°C at night, a relative humidity of 60%, and a light intensity of 300 μmol m⁻² s⁻¹. Ca fertilizer treatments for tomato plants were applied three times, at 7, 14, and 21 days after transplanting, through both foliar spraying and root irrigation. The Ca concentration for each application was maintained at 15 mg/L.
2.2. Measurement of Plant Fresh and Dry Weight
Fresh tomato plants were cut at the junction between the root and shoot, and surface moisture was removed. The fresh weight of the samples was then measured using an electronic balance. For dry weight determination, the tissues were placed in a forced-air drying oven at 70°C for three days until completely dried, after which the weight was recorded.
2.3. Measurement of Photosynthetic Rate, Chlorophyll, and Carotenoid Content
The net photosynthetic rate of tomato leaves was measured on sunny days between 9:00 AM and 12:00 PM using a CIRAS-3 portable photosynthesis system. For chlorophyll and carotenoid content analysis, 0.1 g of fresh tomato leaves (with veins removed) was ground into powder using liquid nitrogen and extracted with 80% acetone on a shaker at 400 rpm/min in darkness for 4 h. The extract was centrifuged at 8000 rpm/min, and the supernatant was collected. The chlorophyll and carotenoid contents were quantified spectrophotometrically at wavelengths of 663 nm, 647 nm, and 470 nm, following the method described by Niu et al. (2022) [16].
2.4. Measurement of Fruit Yield and Individual Fruit Weight
After the third fertilization, all fruits that had successfully set on each plant were harvested and weighed using a precision balance to calculate the total fruit yield per plant. Mature fruits were collected and weighed individually to determine the average fruit weight.
2.5. Measurement of Fruit Firmness and Brix
Fruit firmness was measured at the mature stage using a penetrometer. The sweetness of the fruit, expressed as °Brix, was determined using a refractometer.
2.6. Measurement of Total Sugar, Sucrose, Reducing Sugar, and Titratable Acid Contents
A 200 mg dry sample was dissolved in 8 ml of 80% ethanol and heated at 85°C for 30 minutes. The reaction mixture was then centrifuged at 3000 rpm for 5 minutes, and the supernatant was collected. This extraction process was repeated three times, and the supernatants were combined and diluted to a final volume of 50 ml.
Total sugar content was determined using the anthrone colorimetric method. To 0.1 ml of the supernatant, 3.5 ml of 0.15% (w/v) anthrone reagent was added and heated at 90°C for 15 minutes. The absorbance was measured at 620 nm using glucose solution as a standard.
For reducing sugar determination, 0.4 ml of the extract was diluted with 1.6 ml of ddH₂O, and then 1 ml of 1% (w/v) 3,5-dinitrosalicylic acid (containing 20% (v/v) 1N NaOH and 30% (w/v) sodium tartrate) was added. The reaction was carried out at 95°C for 5 minutes, and absorbance was measured at 520 nm using glucose solution as a standard.
For sucrose determination, 0.4 ml of the extract was mixed with 0.1 ml of 30% KOH (w/v) and heated at 95°C for 10 minutes, followed by the addition of 3.5 ml of anthrone reagent. The reaction was continued at 95°C for 15 minutes, and absorbance was measured at 620 nm using sucrose solution as a standard.
Titratable acidity was determined by performing an acid-base titration. A 5 g sample of the fruit juice or extract was diluted to a known volume with distilled water. A few drops of phenolphthalein indicator were added to the solution. The sample was then titrated with a standardized 0.1 M NaOH solution until a pale pink endpoint was reached. The volume of NaOH used was recorded, and titratable acidity was calculated using the following formula:
V: volume of NaOH (ml), C: concentration of NaOH (mol/l), Em: equivalent weight of acid, m: weight of sample (g).
2.7. Determination of Flavonoid Content in Fruit
A 0.1g sample of fruit pulp was ground using liquid nitrogen, and 1ml of pre-chilled 65% ethanol was added and mixed thoroughly. The mixture was then extracted at 4°C in the dark for 4 hours, followed by centrifugation at 12,000 rpm for 20 minutes. A 50μl aliquot of the supernatant was collected and sequentially mixed with 0.1mL of 5% NaNO2, 0.1mL of 10% Al(NO3)3, and 0.4mL of 2mol/L NaOH. After standing for 15 minutes, the absorbance was measured at 510 nm. Rutin (Sigma Chemical, St. Louis, USA) was used as the standard, and the flavonoid content was expressed as mg of rutin per gram of fresh weight (mg rutin·g⁻¹ FW).
2.8. Determination of Ca and Mg Content
The various organs of the tomato plants were separated, dried in a forced-air oven at 80°C for 2 days, and then ground into a fine powder. A 5 ml aliquot of H2SO4 was added to the sample and treated overnight to ensure complete carbonization. The sample was then subjected to digestion in a digestion furnace at 300°C, with the addition of 30% H2O2 dropwise during the heating process until the solution became clear. After filtration, the total Ca and Mg concentration was measured using an atomic absorption spectrophotometer (flame atomic absorption method).
2.9. Determination of Exchangeable Ca Content
We weighed 5 g of air-dried soil samples and added 50 mL of 1 mol/L ammonium acetate solution. The mixture was shaken on a shaker at 70 rpm/min for 1 hour to ensure sufficient interaction between the Ca2+ and the exchange reagent. The extract was then filtered through filter paper, and the filtrate was collected. The Ca concentration in the filtrate was determined using an atomic absorption spectrophotometer (flame atomic absorption method).
2.10. Determination of Ca Content in Xylem Sap
The Ca content in xylem sap was determined using the gravimetric method [17]. Tomato plants were cut 0.5 cm above the root, and a plastic film was shaped into a tube and sealed at both ends based on the required thickness. A small amount of defatted cotton was placed inside the plastic tube and tied with fine thread, then weighed. The defatted cotton was placed in contact with the cut surface and secured with string. After 6 hours, the plastic tube was removed and weighed again. The difference between the two measurements was calculated. The absorbed cotton was placed in a centrifuge tube, eluted with 5% HNO₃, centrifuged, and the supernatant was collected. Finally, the volume was adjusted to 1 mL with 5% HNO₃. The Ca concentration was determined using an atomic absorption spectrophotometer (flame atomic absorption method).
2.11. Data Analysis
All data were recorded in Excel. Statistical analysis was performed using one-way ANOVA test plus Duncan's multiple range test in the data processing system (DPS) software. Different letters indicate significant difference with p < 0.05.
3. Results
3.1. Lignosulfonate Maintains Soil Available Ca
Ca in soil is generally abundant, but the availability of Ca²⁺ is influenced by the complex soil environment (Warton and Matthiessen, 2005). To investigate the chelating ability of lignosulfonate on soil Ca²⁺, we treated field soil with 1% CaCl₂ and lignosulfonate (LS, Figure 1A) for 14 days and measured the Ca content in the soil. The results showed that, without the application of CaCl₂, LS treatment did not alter the total Ca content (Figure 1B) or the exchangeable Ca content (Figure 1C) in the soil. However, under CaCl₂ treatment, LS treatment did not significantly affect the total Ca content (Figure 1B), but it significantly increased the replaceable Ca content (Figure 1C). This indicates that LS treatment maintains a higher level of absorbable Ca²⁺.
3.2.LS-Ca Fertilizer Promotes the Growth of Aboveground Parts of Tomato Plants
To investigate the effects of different Ca fertilizers on tomato growth, we conducted pot experiments using ‘Micro-TOM’ tomato seedlings as the model. Four Ca fertilizers—LS-Ca, EDTA-Ca, Ca(NO₃)₂, and SA-Ca—were applied via root irrigation and foliar spraying. The results showed that compared to the control, LS-Ca treatment significantly enhanced the aboveground growth of tomato plants (Figure 2A), with fresh and dry weights of the aboveground parts increasing by 56% and 67%, respectively (Figure 2B, D). In contrast, no significant differences were observed in the fresh and dry weights of the underground parts between LS-Ca treatment, the control, and other Ca fertilizers (Figure 2C, E).
3.3. LS-Ca Fertilizer Improves Light Use Efficiency in Tomatoes
Chlorophyll a and b, along with carotenoids, are essential pigments for photosynthesis. Chlorophyll a and b primarily absorb red and blue-violet light, while carotenoids mainly absorb blue-violet light. To investigate the impact of Ca fertilizers on tomato photosynthesis, we measured the net photosynthetic rate and chlorophyll content in tomato leaves. The results showed no significant differences in the net photosynthetic rate among the Ca fertilizer treatments compared to the control (Figure S1). However, the LS-Ca treatment significantly increased the contents of chlorophyll a and chlorophyll b by 27% and 46%, respectively (Figure 3A, B). Additionally, LS-Ca treatment reduced the carotenoid content in leaves by 29% (Figure 3C). Studies have shown that plants utilize red light more efficiently than blue-violet light. Therefore, the LS-Ca treatment may enhance tomato light use efficiency by optimizing the absorption of light for photosynthesis.
3.4. LS-Ca Fertilizer Enhances Ca²⁺ Uptake and Transport
Plants absorb Ca²⁺ from the soil and transport it through the xylem to vegetative and reproductive organs. To investigate the effects of various chelated Ca fertilizers on Ca uptake and transport in tomatoes, we measured the changes in Ca²⁺ concentrations in roots, stems, and leaves after each of three fertilizer applications. The results revealed that, except for LS-Ca, Ca²⁺ concentrations in roots did not significantly change after the first and third applications for other treatments but increased significantly after the second application (Figure 4A-C). In contrast, LS-Ca treatment significantly increased root Ca²⁺ concentrations across all three applications (Figure 4A), indicating faster root uptake and sustained Ca availability during late tomato growth stages. Foliar application is a rapid and efficient method for Ca supplementation. Following Ca fertilizer treatments, leaf Ca²⁺ concentrations increased significantly until the third application, after which the concentrations plateaued (Figure 4A-C). Additionally, LS-Ca treatment notably enhanced Mg concentrations in roots and stems (Figure S2), suggesting that LS may also facilitate Mg²⁺ uptake and transport.
Ca²⁺ absorbed by roots is transported long distances through the xylem, moving sequentially from roots to stems and other organs. After the first two fertilizer applications, stem Ca²⁺ concentrations increased significantly (Figure 4A, B), indicating enhanced Ca²⁺ transport. Interestingly, following the third application, only the LS-Ca treatment resulted in a significant increase in stem Ca²⁺ concentration compared to the control (Figure 4C), demonstrating its sustained efficacy in promoting Ca²⁺ transport within the plant. Moreover, xylem sap analysis revealed that LS-Ca treatment significantly elevated Ca²⁺ concentrations compared to the control and other treatments (Figure 4D, E). After the third application, xylem sap Ca²⁺ concentrations in EDTA-Ca, Ca(NO₃)₂, and SA-Ca treatments showed no significant difference from the control, whereas LS-Ca concentrations were markedly higher (Figure 4F). These findings further confirm the role of lignosulfonate chelation in enhancing Ca²⁺ transport within the plant.
3.5. LS-Ca Fertilizer Enhances Tomato Fruit Quality
The bioavailability of Ca directly affects tomato fruit quality, growth, development, and the structure of cell membranes and cell walls (Taylor and Locascio, 2004). To investigate the effects of various Ca fertilizers on tomato fruit development and quality, red ripe fruits were collected, and their yield and quality traits were analyzed. Across treatments, while Ca fertilizer applications advanced flowering time and maintained higher flower numbers after the third application (Figure S3), there were no significant differences in total fruit weight per plant (Figure 5B). Compared to the control, EDTA-Ca and Ca(NO₃)₂ treatments slightly increased individual fruit weight (Figure 5A, C). Notably, fruit firmness in the LS-Ca treatment was significantly higher than that of the control and other treatments (Figure 5D). In terms of fruit quality, LS-Ca, Ca(NO₃)₂, and SA-Ca treatments significantly improved fruit sweetness, with LS-Ca leading to a 33% increase, while Ca(NO₃)₂ and SA-Ca resulted in 13% and 14% increases, respectively (Figure 5E). These findings highlight the superior efficacy of LS-Ca in enhancing both the structural integrity and taste quality of tomato fruits.
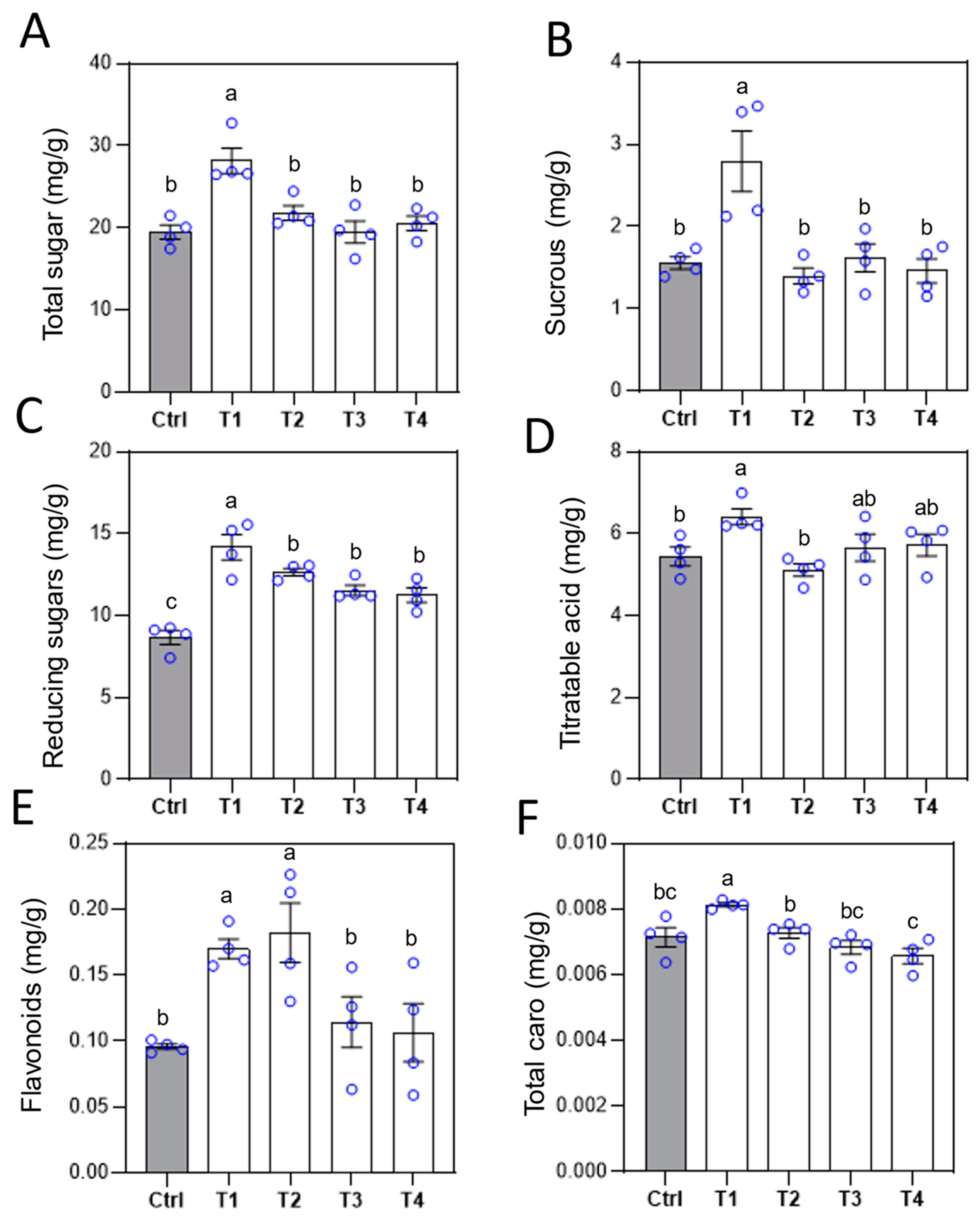
To explore the variations in fruit sweetness, the sugar and acid contents in the fruits were analyzed. Results showed that, compared to the control, LS-Ca treatment increased total sugar, sucrose, and reducing sugar contents by 45%, 79%, and 64%, respectively (Figure 6A-C). The reducing sugar content was also elevated by 46%, 33%, and 30% under EDTA-Ca, Ca(NO₃)₂, and SA-Ca treatments, respectively (Figure 6C). For titratable acid content, LS-Ca treatment resulted in an 18% increase compared to the control, whereas no significant differences were observed in other treatments (Figure 6D). Additionally, LS-Ca treatment significantly enhanced the flavonoid and carotenoid contents of the fruits, with increases of 77% and 14%, respectively (Figure 6E, F). In summary, our findings indicate that LS-Ca treatment enhances fruit firmness and sweetness without affecting fruit weight and yield.
4. Discussion
Lignin, a natural and renewable polyphenolic polymer, is deposited in plant cell walls and tightly cross-linked with cellulose and hemicellulose, providing rigidity to the cell wall [18]. A significant portion of Ca in plants is located in the cell wall, where it binds to carboxyl groups in pectin, particularly pectic acid, forming Ca -pectate complexes essential for cell wall integrity [19,20,21]. This synergy between lignin and Ca within plant cell walls ensures the adaptability and durability of the cell wall under various environmental conditions.
Lignin-derived materials effectively chelate elements such as Fe and Ca, promoting root development in rice [12]. The sulfonic and hydroxyl groups on lignosulfonates (LS) bind to soil particles, enhancing their resistance to wind and water erosion [22]. Additionally, lignin-based slow-release fertilizers for Ca and Fe significantly improve the growth of plants such as holly, poplar, and ginseng [23]. In this study, LS-Ca fertilizer was applied to tomato cultivation and significantly increased the content of chlorophyll a and b in leaves during the growth stages, particularly chlorophyll b (Figure 3). Chloroplasts, known as organelles with a high concentration of Ca²⁺, are closely associated with photosynthesis. Light-dependent Ca²⁺ uptake occurs in isolated chloroplasts, where Ca²⁺ is absorbed from the cytoplasm and stored either in the thylakoid lumen or bound by Ca²⁺-binding proteins under light conditions [24,25,26,27]. With the combined effects of root and foliar applications, Ca²⁺ was highly enriched in leaves. However, despite the significant increase in chlorophyll a and b, there were no observable changes in net photosynthesis (Pn) (Figure S1). Studies have demonstrated that Ca²⁺ optimizes photosynthetic performance and minimizes photooxidative damage. Under low light conditions, the photosynthetic electron transport system captures excitation energy to drive photochemical processes, while under high excitation pressure, Ca²⁺ contributes to the quenching of excess photons to prevent photodamage [27]. Ca has also been shown to regulate enzymatic activity, either catalyzing or inhibiting specific processes. Low resting free [Ca²⁺] in the stroma is essential for activating Calvin-Benson-Bassham (CBB) cycle enzymes, whereas high concentrations of Ca²⁺ can lead to the inactivation of fructose-1,6-bisphosphatase (FBPase) through the release of stored Ca²⁺ following light-to-dark transitions [28,29,30]. Thus, the increased Ca content may enhance leaf structural stability, improve stress resistance, or optimize resource allocation, without necessarily resulting in a measurable increase in Pn.
The availability of Ca in soil is significantly influenced by pH [31]. In alkaline soils, Ca readily precipitates as insoluble compounds like CaCO₃ and Ca₃(PO₄)₂, making it difficult for plants to absorb Ca even when it is abundant. Acidic soils promote the release of Ca²⁺ but also lead to substantial Ca leaching during irrigation. Moreover, microbial activity in acidic soils may be reduced, limiting the release of organically bound Ca [32]. Additionally, other cations present in acidic soils, such as hydrogen ions (H⁺), ferrous ions (Fe²⁺), and manganese ions (Mn²⁺), can adversely affect Ca solubility [33]. Chelated Ca fertilizers provide an effective strategy for Ca supplementation. Lignin, a natural chelating agent extracted from plants, contains abundant hydroxyl and carboxyl functional groups that form stable complexes with metal ions such as iron, zinc, copper, and Ca. These chelates prevent metal ions from precipitating or being fixed in the soil. Compared with other chelated Ca fertilizers, the LS-Ca treatment resulted in a significant increase in Ca content in tomato roots after the first fertilization (Figure 4), indicating that LS-Ca enhances the bioavailability of Ca to plants. This trend persisted throughout the later stages of fertilization. Even when total Ca content in the soil remained constant, LS-Ca treatments mobilized many free Ca²⁺, suggesting that lignosulfonate-chelated Ca provides a slow and sustained release of Ca²⁺. This slow-release characteristic enhances fertilizer efficiency by ensuring a steady nutrient supply and minimizing nutrient loss through leaching or precipitation. Consequently, LS-Ca offers an efficient approach to optimizing Ca availability in various soil conditions, addressing challenges associated with pH-induced Ca fixation or leaching.
Ca also plays a critical role in fruit development. During the early stages, it supports cell division and metabolism, while in the later stages, it contributes to cell-to-cell adhesion [15,34]. Within the cell wall, pectic acids bind to Ca to form Ca pectate, which serves as a structural backbone, preventing gel layer disintegration and enhancing cell wall strength [35]. Typically, Ca is absorbed by roots and transported through the xylem to various plant organs. However, only a small fraction reaches the fruit [36]. Efficient Ca transport within the plant is, therefore, critical for ensuring adequate Ca supplementation in fruits. The LS-Ca treatment not only enhanced Ca uptake but also significantly increased the Ca²⁺ concentration in xylem sap (Figure 4). This indicates that LS-Ca facilitates more efficient transport of Ca to various plant organs, including fruits. Consistent with expectations, Ca content in the fruit significantly increased, contributing to enhanced fruit firmness. Interestingly, the sweetness of tomatoes treated with LS-Ca was 33% higher than that of the control (Figure 5). Additionally, the contents of sugars, acids, flavonoids, and carotenoids in the fruit also showed varying degrees of increase (Figure 6). In the late stages of fruit ripening, the transport of water, sugars, and essential nutrients primarily occurs through the phloem [36,37,38]. However, because Ca²⁺ has limited mobility in the phloem, its supplementation in fruits largely depends on xylem transport [36]. The increased xylem transport of Ca²⁺ likely contributed to timely Ca accumulation in the fruit. This may have optimized the phloem’s capacity for transporting sugars and acids to the fruit. Ca in the cell wall also delayed fruit softening, leading to higher accumulation of sugars, acids, and other metabolites, ultimately improving fruit quality.
Plants naturally produce approximately 150 billion tons of lignin annually. Additionally, the pulp and paper industry generates about 50 million tons of lignin as a byproduct each year [39]. This abundant availability makes lignin a low-cost resource. Transforming lignin into high-value controlled-release fertilizers not only enhances nutrient absorption by plants but also conserves resources and mitigates the environmental pollution caused by lignin incineration. As a chemically modified material, lignin serves as an excellent carrier for nutrients. Its ability to form controlled-release fertilizers stems from chemical interactions between lignin and nutrients, allowing for gradual nutrient release. With advancements in the preparation techniques for controlled-release fertilizers, lignin-based fertilizers hold significant potential for agricultural sustainability. They can improve resource use efficiency, protect agricultural ecosystems, and contribute to sustainable agricultural growth. By utilizing lignin, a renewable and eco-friendly resource, the agricultural sector can address both productivity and environmental challenges, highlighting lignin’s promising role in modern sustainable practices.
5. Conclusions
This study investigated the effects of LS-Ca, EDTA-Ca, Ca(NO3)2, and AS-Ca treatments on tomato growth and development. The results demonstrated that LS-Ca application significantly enhanced Ca absorption and transport, improving the photosynthetic efficiency of tomato leaves. Additionally, LS-Ca treatment increased fruit hardness and sweetness, surpassing the effects of other Ca fertilizers. These findings provide a theoretical foundation for optimizing tomato cultivation practices and improving fruit quality through the application of LS-Ca fertilizers.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
F.M. and Z.B. conceived and designed the experiments. J.Z. and M.D. performed most of experiments, and G.L. provided technical assistance. J.Z., F.M. and Z.B. analyzed the data. J.Z., F.M. and Z.B. wrote the manuscript.
Funding
This research was supported by Taishan Scholar Foundation of Shandong Province (tsqn201909073 to F.M.; tsqn201812034 to Z.B). We thank plant growth facility members in the state key laboratory of crop biology at Shandong Agricultural University.
Data Availability Statement
Data are contained within the article/Supplementary Materials.
Acknowledgments
We are grateful to Chunxiang You’s lab for help with metal element determination experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Naveed Ahmad, I.A., Asif Ali, Muhammad Sajid, Izhar Ullah, Abdur Rab, Syed Tanveer Shah, Fazal-i-Wahid,; Masood Ahmad, A.B., Fareeda Bibi,. 2. Foliar application of Ca improves growth, yield and quality of tomato cultivars. Pure and Applied Biology (PAB) 2020, 10-19%V 19.
- Kadir, S.A. Fruit Quality at Harvest of “Jonathan” Apple Treated with Foliarly-Applied Ca Chloride. J. Plant Nutr. 2005, 27, 1991-2006. [CrossRef]
- McMahon, M.J.; Kofranek, A.; Rubatzky, V.E.; Hartmann, H.T. Plant Science: Growth, Development, and Utilization of Cultivated Plants. BioScience 1983.
- Bangerth, F. Ca-Related Physiological Disorders of Plants. Annu. Rev. Phytopathol. 1979, 17, 97-122. [CrossRef]
- Goulding, K.W.T. Soil acidification and the importance of liming agricultural soils with particular reference to the United Kingdom. Soil Use Manage. 2016, 32, 390-399. [CrossRef]
- Rajan, M.; Shahena, S.; Chandran, V.; Mathew, L. Chapter 3 - Controlled release of fertilizers—concept, reality, and mechanism. In Controlled Release Fertilizers for Sustainable Agriculture, Lewu, F.B., Volova, T., Thomas, S., K.R, R., Eds.; Academic Press: 2021; pp. 41-56.
- Jing, T.; Li, J.; He, Y.; Shankar, A.; Saxena, A.; Tiwari, A.; Maturi, K.C.; Solanki, M.K.; Singh, V.; Eissa, M.A.; et al. Role of Ca nutrition in plant Physiology: Advances in research and insights into acidic soil conditions - A comprehensive review. Plant Physiol. Biochem. 2024, 210, 108602. [CrossRef]
- de Souza Alonso, T.A.; Ferreira Barreto, R.; de Mello Prado, R.; Pereira de Souza, J.; Falleiros Carvalho, R. Silicon spraying alleviates Ca deficiency in tomato plants, but Ca-EDTA is toxic. J. Plant Nutr. Soil Sci. 2020, 183, 659-664. [CrossRef]
- Norvell, W.A. Reactions of Metal Chelates in Soils and Nutrient Solutions. In Micronutrients in Agriculture; 1991; pp. 187-227.
- Li, T.; Wei, Q.; Sun, W.; Tan, H.; Cui, Y.; Han, C.; Zhang, H.; Zeng, F.; Huang, M.; Yan, D. Spraying sorbitol-chelated Ca affected foliar Ca absorption and promoted the yield of peanut (Arachis hypogaea L.). Front. Plant Sci. 2022, 13. [CrossRef]
- Ma, T.; Hui, Y.; Zhang, L.; Su, B.; Wang, R. Foliar application of chelated sugar alcohol Ca fertilizer for regulating the growth and quality of wine grapes. Int. J. Agric. Biol. Eng. 2022, 15, 153-158. [CrossRef]
- Liu, Q.; Kawai, T.; Inukai, Y.; Aoki, D.; Feng, Z.; Xiao, Y.; Fukushima, K.; Lin, X.; Shi, W.; Busch, W.; et al. A lignin-derived material improves plant nutrient bioavailability and growth through its metal chelating capacity. Nat. Commun. 2023, 14, 4866. [CrossRef]
- Kok, A.D.-X.; Wan Abdullah, W.M.A.N.; Tang, C.-N.; Low, L.-Y.; Yuswan, M.H.; Ong-Abdullah, J.; Tan, N.-P.; Lai, K.-S. Sodium lignosulfonate improves shoot growth of Oryza sativa via enhancement of photosynthetic activity and reduced accumulation of reactive oxygen species. Sci. Rep. 2021, 11, 13226. [CrossRef]
- Fischer, A.M. Nutrient Remobilization During Leaf Senescence. In Annual Plant Reviews Volume 26: Senescence Processes in Plants; 2007; pp. 87-107.
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Ca: Transport and Physiology. Front. Plant Sci. 2016, 7. [CrossRef]
- Niu, C.; Wang, G.; Sui, J.; Liu, G.; Ma, F.; Bao, Z. Biostimulants alleviate temperature stress in tomato seedlings. Sci. Hortic. 2022, 293, 110712. [CrossRef]
- Alexou, M.; Peuke, A.D. Methods for Xylem Sap Collection. In Plant Mineral Nutrients: Methods and Protocols, Maathuis, F.J.M., Ed.; Humana Press: Totowa, NJ, 2013; pp. 195-207.
- Rubin, E.M. Genomics of cellulosic biofuels. Nature 2008, 454, 841-845. [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850-861. [CrossRef]
- Hepler, P.K.; Winship, L.J. Ca at the Cell Wall-Cytoplast Interface. J. Integr. Plant Biol. 2010, 52, 147-160. [CrossRef]
- Gilliham, M.; Dayod, M.; Hocking, B.J.; Xu, B.; Conn, S.J.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.D. Ca delivery and storage in plant leaves: exploring the link with water flow. J. Exp. Bot. 2011, 62, 2233-2250. [CrossRef]
- Ahmad, U.M.; Ji, N.; Li, H.; Wu, Q.; Song, C.; Liu, Q.; Ma, D.; Lu, X. Can lignin be transformed into agrochemicals? Recent advances in the agricultural applications of lignin. Ind. Crops Prod. 2021, 170, 113646. [CrossRef]
- Kevers, C.; Soteras, G.; Baccou, J.C.; Gaspar, T. Lignosulfonates: Novel promoting additives for plant tissue cultures. In Vitro Cellular & Developmental Biology - Plant 1999, 35, 413-416. [CrossRef]
- Muto, S.; Izawa, S.; Miyachi, S. Light-induced Ca2+ uptake by intact chloroplasts. FEBS Lett. 1982, 139, 250-254. [CrossRef]
- Kreimer, G.; Melkonian, M.; Holtum, J.A.M.; Latzko, E. Characterization of Ca fluxes across the envelope of intact spinach chloroplasts. Planta 1985, 166, 515-523. [CrossRef]
- Kreimer, G.; Melkonian, M.; Latzko, E. An electrogenic uniport mediates light-dependent Ca2+ influx into intact spinach chloroplasts. FEBS Lett. 1985, 180, 253-258. [CrossRef]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Ca-dependent regulation of photosynthesis. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2015, 1847, 993-1003. [CrossRef]
- Charles, S.A.; Halliwell, B. Action of Ca ions on spinach (Spinacia oleracea) chloroplast fructose bisphosphatase and other enzymes of the Calvin cycle. Biochem. J. 1980, 188, 775-779. [CrossRef]
- Portis, A.R.; Heldt, H.W. Light-dependent changes of the Mg2+ concentration in the stroma in relation to the Mg2+ dependency of CO2 fixation in intact chloroplasts. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1976, 449, 434-446. [CrossRef]
- Hertig, C.; Wolosiuk, R.A. A dual effect of Ca2+ on chloroplast fructose-1,6-bisphosphatase. Biochem. Biophys. Res. Commun. 1980, 97, 325-333. [CrossRef]
- Lawrence, G.; Siemion, J.; Antidormi, M.; Bonville, D.; McHale, M. Have Sustained Acidic Deposition Decreases Led to Increased Ca Availability in Recovering Watersheds of the Adirondack Region of New York, USA? Soil Systems 2021, 5. [CrossRef]
- Feng, C.; Cui, B.; Ge, H.; Huang, Y.; Zhang, W.; Zhu, J. Reinforcement of Recycled Aggregate by Microbial-Induced Mineralization and Deposition of Ca Carbonate—Influencing Factors, Mechanism and Effect of Reinforcement. Crystals 2021, 11. [CrossRef]
- Ng, J.F.; Ahmed, O.H.; Jalloh, M.B.; Omar, L.; Kwan, Y.M.; Musah, A.A.; Poong, K.H. Soil Nutrient Retention and pH Buffering Capacity Are Enhanced by Calciprill and Sodium Silicate. Agronomy 2022, 12. [CrossRef]
- Bemadac, A.; Jean-Baptiste, I.; Bertoni, G.; Morard, P. Changes in Ca contents during melon (Cucumis melo L.) fruit development. Sci. Hortic. 1996, 66, 181-189. [CrossRef]
- Zhang, L.; Wang, J.-w.; Chen, J.-y.; Song, T.; Jiang, Y.-g.; Zhang, Y.-f.; Wang, L.-j.; Li, F.-l. Preharvest spraying Ca ameliorated aroma weakening and kept higher aroma-related genes expression level in postharvest ‘Nanguo’ pears after long-term refrigerated storage. Sci. Hortic. 2019, 247, 287-295. [CrossRef]
- DRAŽETA, L.; LANG, A.; HALL, A.J.; VOLZ, R.K.; JAMESON, P.E. Causes and Effects of Changes in Xylem Functionality in Apple Fruit. Ann. Bot. 2004, 93, 275-282. [CrossRef]
- Rogiers, S.Y.; Greer, D.H.; Hatfield, J.M.; Orchard, B.A.; Keller, M. Solute Transport into Shiraz Berries during Development and Late-Ripening Shrinkage. Am. J. Enol. Vitic. 2006, 57, 73. [CrossRef]
- Choat, B.; Gambetta, G.A.; Shackel, K.A.; Matthews, M.A. Vascular Function in Grape Berries across Development and Its Relevance to Apparent Hydraulic Isolation. Plant Physiol. 2009, 151, 1677-1687. [CrossRef]
- Chen, J.; Fan, X.; Zhang, L.; Chen, X.; Sun, S.; Sun, R.-C. Research Progress in Lignin-Based Slow/Controlled Release Fertilizer. ChemSusChem 2020, 13, 4356-4366. [CrossRef]
Figure 1.
Effects of lignin sulfonates on Ca2+ chelation. (A) Chemical formula of sodium lignin sulfonate (LS). (B) Differences in total soil Ca content. (C) Differences in soil replaceable Ca content. Error bars represent standard deviations (n = 3). Different letters represent significantly different values at P < 0.05.
Figure 1.
Effects of lignin sulfonates on Ca2+ chelation. (A) Chemical formula of sodium lignin sulfonate (LS). (B) Differences in total soil Ca content. (C) Differences in soil replaceable Ca content. Error bars represent standard deviations (n = 3). Different letters represent significantly different values at P < 0.05.
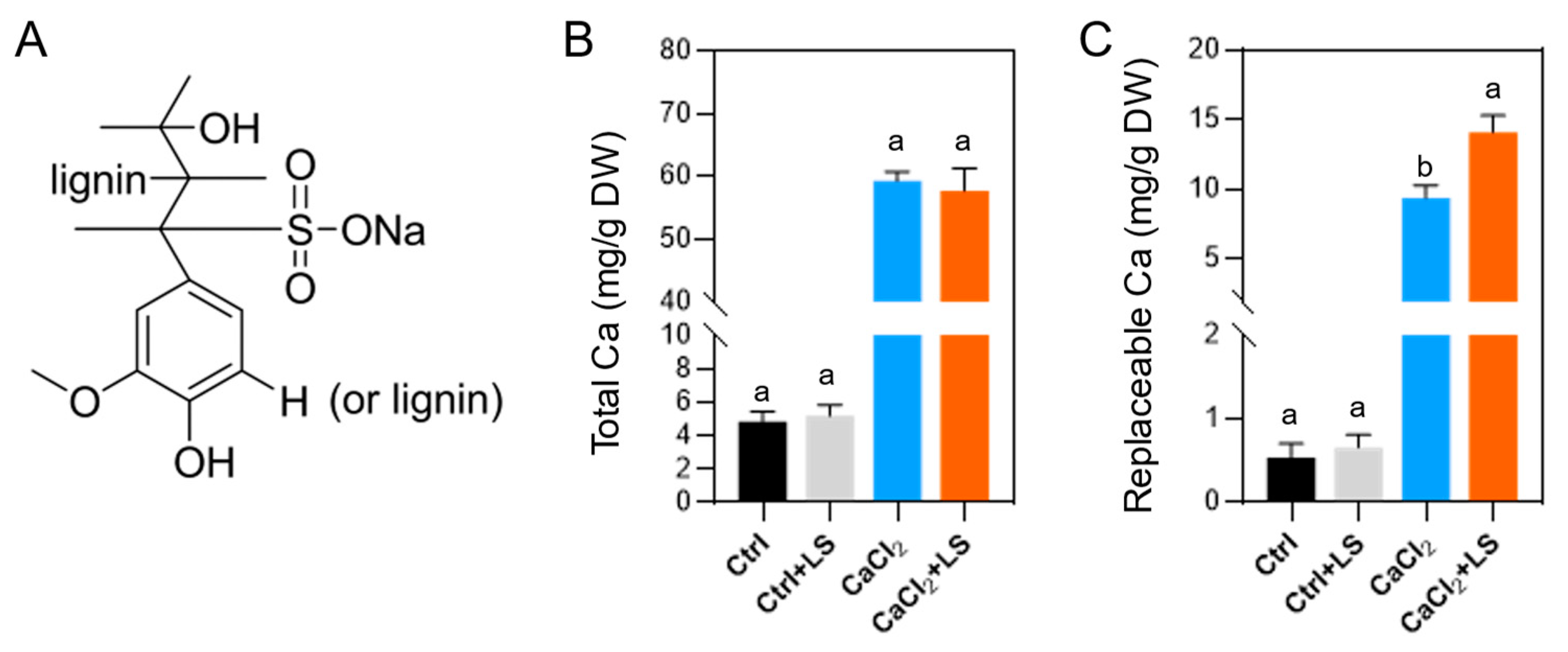
Figure 2.
Effects of different Ca fertilizer treatments on tomato growth. At the end of the third treatment, representative plants were photographed (A) and sampled for fresh weights in aboveground (B) and underground (C). Finally, dry weights in aboveground (D) and underground (E) were measured after the samples were completely dried. Error bars represent standard deviations (n = 5). Different letters represent significantly different values at P < 0.05.
Figure 2.
Effects of different Ca fertilizer treatments on tomato growth. At the end of the third treatment, representative plants were photographed (A) and sampled for fresh weights in aboveground (B) and underground (C). Finally, dry weights in aboveground (D) and underground (E) were measured after the samples were completely dried. Error bars represent standard deviations (n = 5). Different letters represent significantly different values at P < 0.05.
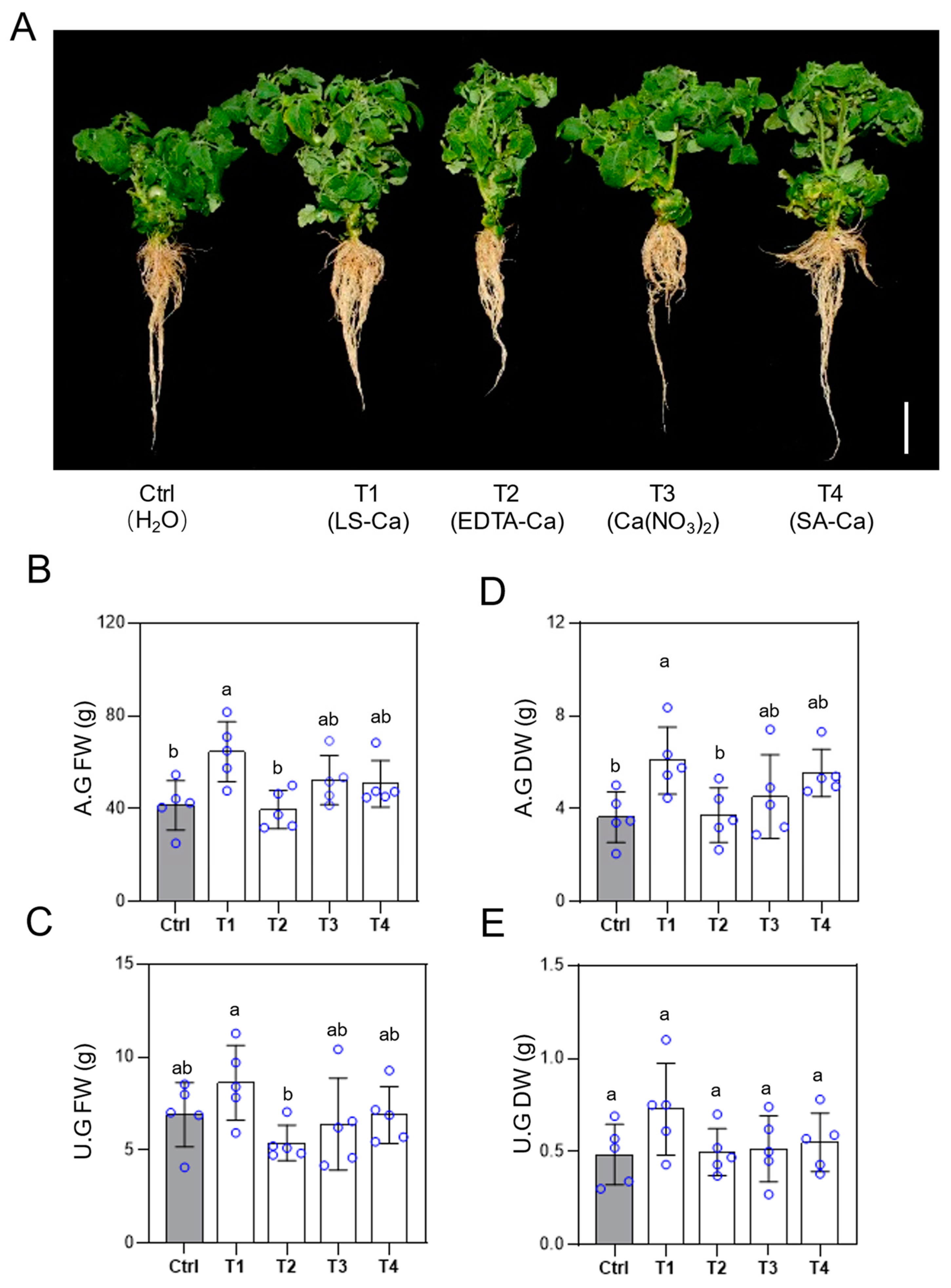
Figure 3.
Effects of different Ca fertilizer treatments on tomato phenotypes. After the second treatment, chlorophyll (A, B) and carotenoid (C) contents in tomato leaves were measured. Error bars represent standard deviations (n = 4). Different letters represent significantly different values at P < 0.05.
Figure 3.
Effects of different Ca fertilizer treatments on tomato phenotypes. After the second treatment, chlorophyll (A, B) and carotenoid (C) contents in tomato leaves were measured. Error bars represent standard deviations (n = 4). Different letters represent significantly different values at P < 0.05.
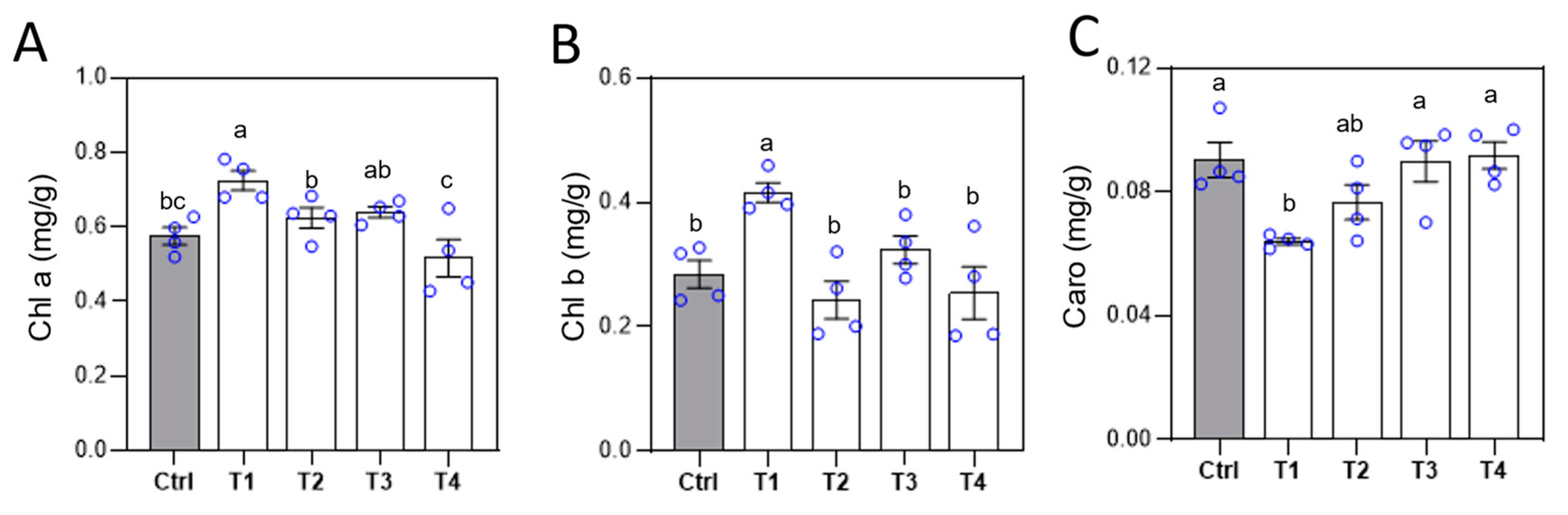
Figure 4.
Effects of different Ca fertilizer treatments on Ca content in tomato. (A) After the first treatment, Ca contents in tomato roots, stems and leaves. (B) After the second treatment, Ca contents in tomato roots, stems and leaves. (C) After the third treatment, Ca contents in tomato roots, stems, leaves, flowers and fruits. (D-F) Ca²⁺ concentrations in xylem sap during three times treatment. Error bars represent standard deviations (n ≥ 3). Different letters represent significantly different values at P < 0.05.
Figure 4.
Effects of different Ca fertilizer treatments on Ca content in tomato. (A) After the first treatment, Ca contents in tomato roots, stems and leaves. (B) After the second treatment, Ca contents in tomato roots, stems and leaves. (C) After the third treatment, Ca contents in tomato roots, stems, leaves, flowers and fruits. (D-F) Ca²⁺ concentrations in xylem sap during three times treatment. Error bars represent standard deviations (n ≥ 3). Different letters represent significantly different values at P < 0.05.
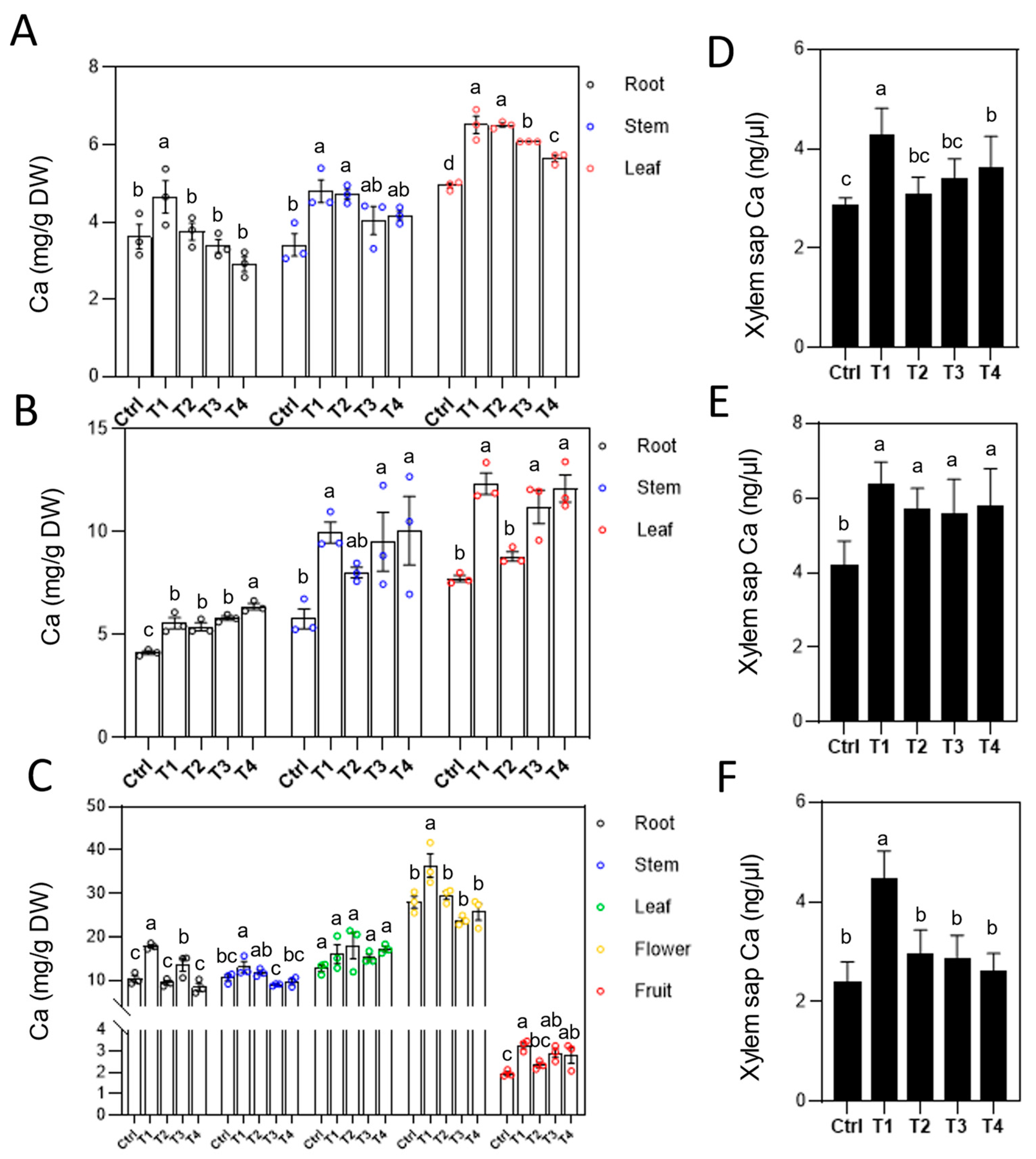
Figure 5.
Effects of different Ca fertilizer treatments on fruit size, firmness and brix of tomato. After the third treatment, (A) representative red ripe fruit were photographed. Scale bar = 1cm. (B) The fruit yield. (C) single fruit weight. (D) fruit firmness. (E) soluble solid contents. Error bars represent standard deviations (n ≥ 5). Different letters represent significantly different values at P < 0.05.
Figure 5.
Effects of different Ca fertilizer treatments on fruit size, firmness and brix of tomato. After the third treatment, (A) representative red ripe fruit were photographed. Scale bar = 1cm. (B) The fruit yield. (C) single fruit weight. (D) fruit firmness. (E) soluble solid contents. Error bars represent standard deviations (n ≥ 5). Different letters represent significantly different values at P < 0.05.
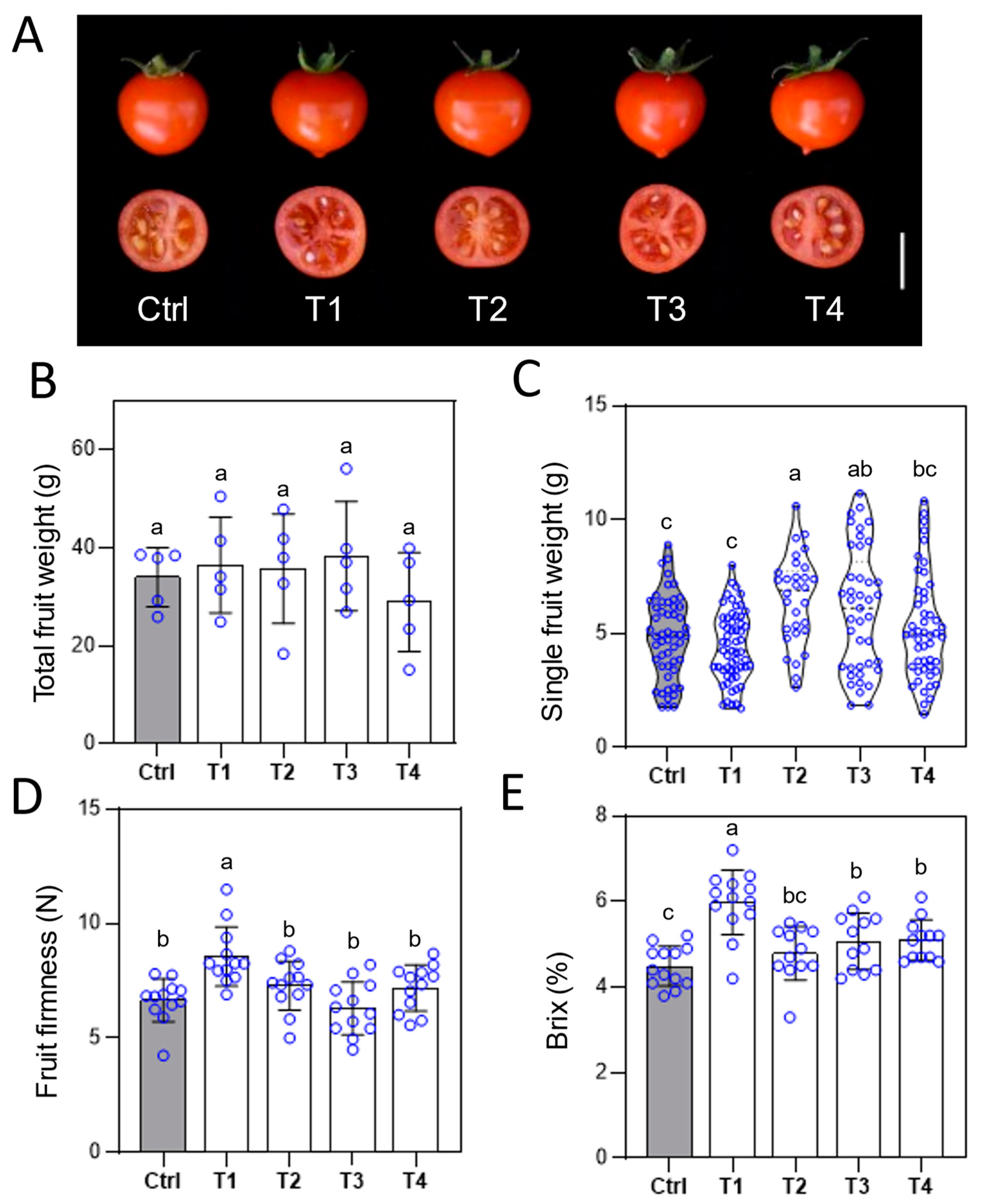
Figure 6.
Effects of different Ca fertilizer treatments on tomato fruit quality. After the third treatment, the contents of total sugar (A), reducing sugar (B), sucrose (C), titratable acid (D), flavonoids (E) and carotenoids (F) in tomato fruits were measured. Error bars represent standard deviations (n = 4). Different letters represent significantly different values at P < 0.05.
Figure 6.
Effects of different Ca fertilizer treatments on tomato fruit quality. After the third treatment, the contents of total sugar (A), reducing sugar (B), sucrose (C), titratable acid (D), flavonoids (E) and carotenoids (F) in tomato fruits were measured. Error bars represent standard deviations (n = 4). Different letters represent significantly different values at P < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



