
Submitted:
20 November 2024
Posted:
22 November 2024
You are already at the latest version
Abstract
Background/Objectives: Cold stress poses a significant threat to Asian rice cultivation, disrupting important physiological processes crucial for seedling establishment and overall plant growth. It is thus crucial to elucidate genetic pathways involved in cold stress tolerance response mechanisms. Methods: We mapped OsUBC7, a RAD6 type homolog of rice, to a low-temperature seedling survivability (LTSS) QTL and used genomics, molecular-genetics, and physiological assays to assess its role in plant resilience against low-temperature stress. Results: OsUBC7 is cold responsive and has higher expression levels in cold-tolerant japonica than cold-sensitive indica. Overexpression of OsUBC7 enhances LTSS of indica and freezing tolerance of Arabidopsis, increases levels of soluble sugars and chlorophyll A, and boosts leaf development after cold exposure and increases leaf cell numbers and plants size, but does not affect membrane stability after cold stress exposure. Additionally, OsUBC7 has a positive role for germinability in the presence of salt and for flowering and yield-related traits. The OsUBC7 protein physically interacts with the developmental stage specific and histone modifying E3 ligases OsRFPH2-12 and OsHUB1/2, respectively, and potential target genes such as cell cycle dependent kinases were identified. Conclusions: OsUBC7 might contribute to cold resilience by regulating sugar metabolism to provide energy for promoting cellular homeostasis restoration after cold stress exposure via new cell growth, particularly in leaf cells crucial for photosynthesis and metabolic activity, possibly by interacting with cell cycle regulating proteins. Overall, this study suggests that OsUBC7 may be involved in plant development, reproduction, and stress adaptation, and contributes to a deeper understanding of rice plant cold stress tolerance response mechanisms. OsUBC7 may be a promising candidate for improving crop productivity and resilience to stressful environments.
Keywords:
chilling stress
; LTSS
; low-temperature stress recovery
; Oryza sativa L.
1. Introduction
Seedling development is crucial for plant growth, establishing the foundation for plant maturation and stress resilience [1,2,3]. It involves physiological changes such as root system establishment and leaf emergence, essential for adaptation to the environment [4,5,6]. This phase influences overall plant architecture and productivity [7,8]. The successful establishment of healthy seedlings is important for ensuring optimal yield and agricultural sustainability, particularly in the case of Asian rice [1,8], a staple food crop supporting billions of people worldwide. Asian rice cultivation involves two primary subspecies, japonica and indica, each characterized by unique genetic traits and adaptive responses to environmental conditions [1,2].
A common threat to rice cultivation is low temperature exposure [1,2], which poses a significant challenge to plant growth and development, disrupting various physiological processes essential for seedling establishment and survival [4,9]. Cold stress can induce cellular damage, disrupt membrane integrity, and impair metabolic functions, ultimately compromising plant growth and productivity [1,9,10]. In rice, exposure to low temperatures at early growth stages can lead to reduced germination rates, delayed seedling emergence, and stunted growth, culminating in diminished yields and economic losses for farmers [1,2,11]. Understanding the molecular basis of cold stress tolerance response mechanisms is essential for developing strategies to enhance resilience and improve seedling establishment in rice cultivation.
The genetic diversity within rice subspecies provides valuable insights into their resilience to abiotic stresses [2,11]. Accessions from the two subspecies, japonica and indica, stand out for their distinctive characteristics regarding cold tolerance. Japonica accessions generally have a greater potential for cold tolerance than indica accessions, making them better suited for cultivation in regions with cooler climates [1,4]. This inherent genetic diversity demonstrates the intricate relations between genetic factors and environmental cues in shaping plant growth and offer a rich resource for studying the molecular mechanisms underlying cold stress tolerance responses in rice [1,12].
The selection of OsUBC7 (LOC_Os07g07240) as a central point for this study is rooted in its potential as a regulator of seedling development and stress responses in rice due to its association with the multiple low-temperature stress tolerance trait QTL qMT7-3 [2]. OsUBC7 codes for a RAD6-like protein sharing sequence similarity with genes known to be involved in critical cellular processes such as DNA repair, cell cycle regulation, gene expression regulation, and protein quality control mechanisms [13,14,15,16,17,18]. We hypothesized that OsUBC7 is likely involved in similar growth dynamics, cellular proliferation, and stress response pathways via interactions with various protein partners affecting different downstream signaling pathways for rice plants to cope with low-temperature stress. This is supported by previous mapping studies associating OsUBC7 with yield-related traits and stress tolerance mechanisms, thus justifying yield related investigations besides low-temperature stress studies [19]. Moreover, metadata analyses indicated that OsUBC7 is differentially expressed under various environmental conditions and in response to changing environmental cues, including temperature stress [15,19]. By investigating the potential roles of OsUBC7 in cellular homeostasis, growth regulation, and stress responses, this study aimed to reveal some of the complex molecular mechanisms underlying rice seedling development and stress resilience.
Specifically, we investigated the potential molecular function(s) and regulatory roles of OsUBC7 in key aspects of seedling growth and development and cold stress adaptation to acquire insights for future breeding strategies to enhance crop productivity and resilience in the face of a changing environment. Determining the role of OsUBC7 in seedling development and stress responses not only contributes to fundamental knowledge in characterizing rice genes, but also to developing novel breeding strategies to enhance crop performance and resilience in response to environmental challenges by revealing the complex relationship between genetic factors and environmental cues in crop research and development.
2. Materials and Methods
2.1. Plant Materials and Growth Condition
Rice seedlings were grown hydroponically in an AR-66L growth chamber (Percival Scientific, Perry, IA) under a photon flux of ~165 μE and 12/12 hr light/dark cycles at 28/25°C day/night temperatures. On the tenth day, water was replaced with quarter-strength Murashige and Skoog (MS) liquid medium. Arabidopsis seeds, both wild-type (WT) and transgenic, were surface-sterilized using 10% sodium hypochlorite before germination on agar-solidified half-strength MS medium in an AR-66L growth chamber under standard conditions of 16/8 hr light/dark cycles and 22/20°C day/night temperatures at a photon flux of ~165 μE. The aus subgroup reference accession Kasalath from the indica rice subspecies and Arabidopsis ecotype Columbia-0 (Col-0) were used for the generation of overexpression (OE) transgenic lines, while the AtUBC2 knockout (KO) line SALK_152462C was obtained from the Arabidopsis Biological Resource Center at Ohio State University.
2.2. Cold-Temperature Stress Treatment of Rice Plants
Rice seeds were germinated for 2 days at 37°C and grown as described in 2.1. For most experiments, two-week-old lines overexpressing OsUBC7 from the strong 35S enhancer-mannopine synthase promoter (cloning primers and degree of upregulation shown in Supplementary Tables S1 and S2, respectively) and WT Kasalath seedlings were continuously stressed at 10°C for 2 days, as this treatment led to half plant population survival (LT50= 50% survival) for Kasalath WT plants [4,9]. All physiological assays on rice seedlings were done at the conclusion of the cold-temperature treatment. For survival scoring, cold-treated rice seedlings were returned to standard growth conditions in growth chambers as described in 2.1. for one week of recovery growth.
2.3. Assessment of Percent Low-Temperature Seedling Survivability (LTSS) in Rice
Two-week-old seedlings rice seedlings were subjected for various periods of time to 10°C cold-temperature stress and then permitted to recover for 7 days under 12/12 hr photoperiod conditions at 28/25°C (day/night) with a photon flux of ~165 μE. Mean survivability was determined as percent LTSS after 7 days of recovery, calculated by dividing the number of green and healthy-looking seedlings after recovery by the initial number of healthy-looking seedlings before cold treatment x 100.
2.4. Assessment of Freezing Stress Tolerance in Arabidopsis
Col-0 WT and OsUBC7 OE transgenic lines were exposed to freezing stress using a refrigerated circulating bath (Polyscience, Niles, IL). Two-week-old individual seedlings were carefully removed from agar plates and transferred onto pre-wetted Light-Duty Tissue Wipes (VWR, Radnor, PA) in 50-ml conical tubes, ensuring humidity was maintained by covering them with additional pre-wetted Tissue Wipes. The tubes were then placed in the circulating bath at –2°C for 15 min, followed by the addition of approximately 50 μl of double distilled (dd) H2O ice chips to facilitate ice nucleation in the plant tissues. After incubating at –3°C for 75 min, the seedlings were allowed to thaw inside the tubes at room temperature for 30 min before being transferred to pre-wetted filter papers. Subsequently, seedlings were kept under standard growth conditions in the AR-66L growth chamber (16/8 hr light/dark cycles and 22/20°C day/night temperatures). Percent freezing survival was calculated at 4, 5, 6, and 7 days of recovery growth by dividing the number of green and healthy-looking seedlings by the initial number of healthy-looking seedlings before treatment x 100.
2.5. Gene Expression Analysis
Quantitative reverse-transcription PCR (qPCR) analyses were conducted to assess mRNA abundance in rice plants. Following various treatments, whole seedling or leaf tissues at the flowering stage were collected and promptly frozen in liquid nitrogen. Total RNA was extracted using the Trizol reagent (Invitrogen, Carlsbad, CA). Subsequently, first-strand cDNA was synthesized utilizing M-MLV reverse-transcriptase (Promega, Madison, WI), and qPCR reactions were performed using 2× universal SYBR green supermix (BioRad, Hercules, CA) in a BioRad CFX96 Touch™ Real-Time PCR Detection System under the following conditions: initial denaturation at 95°C for 5 min, followed by 40 cycles of denaturation at 95°C for 15 sec, annealing at 55°C for 30 sec, and extension at 72°C for 1 min. The primer sequences used are shown in Supplementary Table S1.
2.6. Measurement of Percent Electrolyte Leakage (EL)
Following the low-temperature stress treatments, 4 equally sized tissue sections from the middle part of the second leaf were collected from rice seedlings, while whole
plants of Arabidopsis were collected. Tissue samples were washed in ddH2O after which they were transferred into screw-cap glass tubes containing 5 ml of ddH2O and placed on a rotary shaker at 200 rpm for 1 hr to facilitate the release of electrolytes from damaged cells. Electrolyte conductivity of the 4 replicate samples was measured in microsiemens (µS) twice using a hand-held LAQUAtwin B-771 conductivity meter (Horiba Scientific, Japan), with precautions taken to ensure glass tube cleanliness and close to 0 µS conductivity of the ddH2O supply. Subsequently, after the initial measurement, tissue samples were boiled for 10 min to release the total amount cellular electrolytes. Following cooling to room temperature, samples were again shaken at 200 rpm for 1 h, and total conductivity was measured. Electrolyte leakage (EL) for each treatment was then calculated as %EL = [(initial conductivity)/(total conductivity)]×100.
2.7. Measurement of Sugar Content in Two-Week-Old Rice and Arabidopsis Seedlings
Sugar content in plant tissues was assessed by a two-step process involving sugar extraction and colorimetric staining, as previously described in [20] and [21], with minor modifications. Collected tissues were first ground in liquid nitrogen, weighed, and treated on ice with 1 mL of 12:5:3 methanol:chloroform:water (MCW) extraction buffer containing lactose (0.1 μg/μL). After brief mixing and incubation at 50°C for 30 min, samples were centrifuged (12,052 x g) for 5 min at room temperature, and the resulting supernatant transferred to new conical tubes and kept on ice. Subsequently, another round of centrifugation was performed after adding 1 mL of MCW extraction buffer without lactose to the plant tissues, and the supernatant was collected and transferred as before. This process was repeated, with sterile ddH2O added to each tube as the third and final step. After mixing and centrifugation, 1.5 mL of the aqueous solution containing the sugar extract was transferred to microcentrifuge tubes and stored at –20°C overnight. For the colorimetric assay, 40 μL of each sample was combined with 200 μL of 5% phenol and 1 mL of concentrated sulfuric acid, followed by incubation at room temperature for 20 min. Absorbance of the samples was then measured at 510 nm, and a standard curve was generated using a 1 mg/mL sucrose stock solution. The results were normalized by fresh tissue weight (mg). This experiment was conducted three times, with nine rice plant samples collected each time, where every three plants were pooled together. For Arabidopsis, thirty plant samples were collected, with every ten plants pooled together for each reading.
2.8. Measurement of Chlorophyll Content in Rice Flag Leaves
Chlorophyll content of Kasalath flag leaves at the flowering stage was determined as previously described [22], with minor modifications. Flag leaf tissues were submerged in 7 mL of 100% ethanol and kept in the dark at room temperature for 4 days. Subsequently, leaves were removed from ethanol and dried at 60°C for 24 hrs to determine dry weight. The absorbance of 1 mL of the ethanol was measured, using a spectrophotometer, at 665 nm and 649 nm to quantify chlorophyll A and B, respectively, with 1 mL of pure ethanol serving as blank. Chlorophyll content was calculated as follows: chlorophyll A, (13.70×A665nm−5.76×A649nm) μg/mL; chlorophyll B, (−7.60×A665+25.8×A649) μg/mL. All values were multiplied by 7 to account for the initial 7 mL of ethanol used. The amounts of chlorophyll A and B were normalized by the milligrams of dry weight of leaf tissues. This experiment was conducted three times, with at least six plant samples collected each time.
2.9. Measurement of Leaf Length, Leaf Width, and Yield-Related Phenotypes
The flag leaf dimensions were assessed using a 30-cm ruler, with precise measurements recorded to the nearest mm. Half-length of the flag leaf was determined by dividing total length by 2. Quantification of seed yield in both OsUBC7 OE and WT plants done by counting seeds produced by representative plants from each line across multiple transgenic (T) generations (T2, second; T3, third, and T4, fourth). Number of seeds per panicle was calculated by dividing the total seed yield by the corresponding total number of panicles per plant. Percent fertility was calculated by dividing the number of filled grains by the total number of filled and unfilled grains x100.
2.11. Yeast Two-Hybrid (Y2H) Analyses
The coding sequence of OsUBC7 was PCR amplified using cDNA derived from the temperate japonica accession Krasnodarskij 3352 as template with forward primer 5’-AAACATATGATGTCGACGCCGGCGAGGAA-3’ (NdeI site underlined) and reverse primer 5’-AAAGGATCCCTAGTCAGCTGTCCAGCTCT-3’ (BamHI site underlined) and cloned into pGEM-T-easy (Promega). OsUBC7 cDNA was removed from the vector via NdeI & BamHI restriction enzyme digestion and cloned into the pGBKT7 bait vector (TakaraBio, San Jose, CA). The coding sequence of the E3 ubiquitin ligase gene OsRFPH2-12 (LOC_Os01g58780) was amplified from the same cDNA source using forward primer 5’-AAACATATGATGCCAAATAGAGCCACGCAT-3’ (NdeI site underlined) and reverse primer 5’-AAAGGATCCTCAGTAGTCGTAATGCCACTC-3’ (BamHI site underlined) and cloned into pGEM-T-easy. The RING finger domain of the E3 ubiquitin ligase genes OsHUB1/OsHUB2 (LOC_Os04g46450/LOC_Os10g41590) were PCR amplified using the same cDNA source as template with forward primer 5’-AAAGAATTCATGGAGGGTGTTAAAGCAAA-3’ (EcoRI site underlined) and reverse primer 5’-AAAGGATCCTCATATGTAGATAGGTTTCA-3’ (BamHI site underlined) and cloned into pGEM-T-easy. E3 ligase gene sequences were removed from the pGEM-T-easy vector via NdeI & BamHI and EcoRI & BamHI digestions for OsRFPH2-12 and OsHUB1/OsHUB2 cDNA, respectively, and cloned into the pGAKT7 prey vector (TakaraBio). Positive protein interaction control vectors pGBKT7-53 (bait) and pGADT7-T (prey) were obtained from TakaraBio. pGBKT7-OsUBC7 (bait) and pGAKT7 (empty prey), and pGBKT7 (empty bait) and pGAKT7-OsRFPH2-12 (prey) and pGAKT7-OsHUB1/OsHUB2 (prey) were used as negative controls. Each interacting pair of plasmids was transformed into yeast strain Y2HGold (TakaraBio) using the heat shock method. Yeast cultures were grown overnight at 30°C in YPD after which 1 mL of the culture was diluted into 5 mL of fresh YPD and incubated at 30°C for 3–5 hrs until the culture reached an OD600 of 0.2–0.8. Isolated vector DNA (5.0 µL) was added to sterile Eppendorf tubes, and for each sample a master mix consisting of 100 µL One Step Buffer, 10 µL 1M DTT, and 5 µL salmon sperm DNA was prepared. Yeast cultures were centrifuged at 3,000 rpm for 2 min, resuspended in 1 mL ddH2O, and centrifuged again at maximum speed for 30 sec. The pellet was resuspended in 115 µL of master mix, and 115 µL of this mixture was added to the DNA vector containing tubes. After brief mixing, samples were heat shocked at 45°C for 30 min. Cells were plated onto synthetically defined (SD) selective medium with the following “Dropped Out “formulations: SD/–Leu/–Trp Double Dropout (DDO) for selection of the bait and prey vectors, and SD/–Leu/–Trp/–His/–Ade Quadruple Dropout (QDO) for selection of the two vectors and protein-protein interaction between the bait and prey construct. Yeast colonies were obtained after 3 days of incubation at 30°C.
2.10. Assessment of Salt Stress Tolerance in Arabidopsis
Salt germination assays were done as described previously [9,23]. A total of 100 surface-sterilized seeds of the Col-0 WT and one OsUBC7 OE homozygous transgenic Arabidopsis line were arranged in a grid pattern on agar-solidified half-strength MS medium supplemented with various NaCl concentrations (0, 50, and 100 mM). The seeds were allowed to germinate for two weeks under standard growth conditions. Each plate contained ~50 seeds from WT and OE lines, with seed positions randomized across three plates to mitigate potential positional effects. Germination, defined as the emergence of the radicle, was monitored daily during a two-week period. Percent germination for each line was calculated for each replicate plate as % germination = [(number of germinated seeds)/(total number of seeds)] × 100. Mean germination percentages were calculated from replicate experiments.
3. Results
3.1. Identification of LOC_Os07g07240 as a Chilling Tolerance Candidate Gene Associated with qMT7-3
Using the Rice Diversity Panel 1 (RDP1) from the USDA and a genome-wide association study (GWAS) mapping approach, we previously identified numerous Multi-cold tolerance response Trait (MT) quantitative trait loci (QTL) [2]. Since the peak QTL single nucleotide polymorphism (SNP) within the 3.4–3.7 Mb QTL region for qMT7-3 on chromosome 7 was in the transcribed region of LOC_Os07g07240, annotated as a Rad6 type ubiquitin-conjugating (UBC) enzyme encoding gene classified as OsUBC7, we focused on this gene as a candidate for qMT7-3. In silico genomics analyses indicated that OsUBC7 met several criteria for a cold tolerance candidate gene, including differential gene expression between chilling-tolerant temperate japonica Nipponbare and chilling-sensitive indica line 93-11 in response to low-temperature treatment, and the presence of additional 10 SNPs in the coding regions between the rice reference genomes of two temperate japonica lines, Nipponbare and Krasnodarskij 3352, and the two aus lines Carolino 164 and Kasalath. Of the 11 SNPs, one (the peak QTL SNP) was in the 5’ untranslated region (UTR), eight were in introns, and two in the 3’ UTR. Matching of these SNPs in all 354 accessions of the Rice Diversity Panel 1 (RDP1) used for the GWAS study [2] to low-temperature seedling survivability (LTSS) scores showed that accessions with low LTSS scores predominantly had indica type SNPs at the 5’ UTR, at three intron locations, and both 3’ UTR regions, which we defined as an “alternate” haplotype compared to the japonica reference genome haplotype (Figure 1A). Further analyses uncovered eight different haplotypes (I, II, III, IV, V, VI, VII, VIII) among 4,000 accessions analyzed. Among those, three major haplotypes (I, II, III) covered more than 1,000 accessions. Haplotype I (AGCAGGCC) was predominantly found in japonica accessions such as Nipponbare (82.11%), haplotype II (AACAGGCC) mostly in indica accessions such as 93-11 (81.92%) and some japonica accession (16.52%), and haplotype III (GGCTGGTT) predominantly in indica (83.49%) and aus accessions such as Kasalath (14.17%) (Figure 1B). Matching these haplotypes to LTSS scores of the 354 RDP1 accessions showed that accessions with haplotype I had high LTSS scores, those with haplotype II intermediate LTSS scores, and those with haplotype III low LTSS scores (Figure 1C).
Taken together, these data suggested that OsUBC7 is a probable LTSS trait candidate gene in rice, supported by differential expression patterns and SNP differences between chilling-tolerant and chilling-sensitive accessions, pointing to a potential role in mediating cold stress tolerance responses. The significant correlation between OsUBC7 haplotypes and LTSS scores (Figure 1C) further emphasized its potential role in modulating cold stress tolerance in rice, highlighting the importance of studying the gene further for future research strategies to enhance rice crop resilience against cold temperature stress.
3.2. OsUBC7 Is Upregulated by low Temperature Exposure
To validate that OsUBC7 was cold regulated, we measured mRNA abundance in both indica and japonica subspecies under warm control condition and after 12, 24, 36, and 48 hrs of exposure to 10°C (for indica) and 4°C (for japonica) using four different housekeeping genes to normalize expression levels: 18S ribosomal RNA, OsACT1, OsUBC32, and OsUBQ5. Although normalizing using different housekeeping genes gave slightly different results, overall, OsUBC7 mRNA increased progressively in both indica and japonica accessions during cold temperature exposure (Table 1). Compared to warm controls, fold changes of OsUBC7 mRNA levels in the cold were higher in japonica than in indica samples. When averaging values using different housekeeping genes, log2 fold changes of OsUBC7 mRNA abundance at 12 hrs of cold were 0.88±0.81 and 1.01±0.53 in indica and japonica, respectively; at 24 hrs of cold, 1.45±0.84 and 2.39±0.83 in indica and japonica, respectively (paired t-test, p < 0.05); at 36 hrs of cold, 1.97±0.67 and 3.15±0.36 in indica and japonica, respectively (p < 0.05); and at 48 hrs, 2.09±0.75 and 3.39±1.00 fold in indica and japonica, respectively (p < 0.05).
Taken together, these results indicated that OsUBC7 mRNA levels are upregulated by cold temperature exposure in both the indica and japonica subspecies of rice. Across different time points and normalization methods, the log2 fold change in OsUBC7 expression consistently showed upregulation in response to low temperatures, with japonica showing significantly higher mean log2 fold changes than indica after 24-48 hrs of cold exposure. Overall, this progressive upregulation of OsUBC7 in response to cold suggests that the gene might play a role in the cold stress tolerance response mechanisms of rice.
3.3. Overexpression of OsUBC7 Enhances LTSS Scores of Transgenic Rice Lines
To determine whether constitutive overexpression (OE) of OsUBC7 in cold-sensitive rice accessions had a positive effect on LTSS, fourth generation (T4) OE transgenic plants were exposed for 2, 3, 4, 5, and 7 days to 10°C and allowed to recover for one week at warm temperatures. After 2 days of cold, all four OE lines tested had significantly higher LTSS scores, ranging from 75% to 80.6%, compared to segregated wild-type (WT) lines, ranging from 52.1% to 54.2% (p < 0.05). After 3 days, two OE lines had significantly higher LTSS scores, 61.1% and 63.9% (p < 0.05), while the other two had a non-significant trend of 55.6% and 63.9% LTSS compared to 43.8% and 47.9% LTSS of segregated WT plants. After 4 days of cold stress, both transgenic and WT lines had similar LTSS scores, averaging between 33.3% to 38.9% LTSS. After 5 days, LTSS scores remained similar among all lines, ranging from 16.7% to 30.6%. Moreover, after 7 days of cold stress, all lines reached an LTSS score of zero (Figure 2A).
These results indicated that OE of OsUBC7 in a cold sensitive rice accession helped overall seedling survival of plants exposed to 10°C for a short period of 2-3 days, increasing LTSS scores by approximately 50%. After longer exposure times, LTSS scores between OsUBC7 OE transgenic and WT lines were similar, suggesting that OsUBC7 might be an early cold stress tolerance response gene.
3.4. Overexpression of OsUBC7 Increases Freezing Tolerance of Transgenic Arabidopsis Lines
Since OE of OsUBC7 enhanced LTSS scores of transgenic rice lines after cold exposure, we overexpressed OsUBC7 in Arabidopsis to determine whether it had a positive effect on freezing survival in a heterologous system sharing a closely related homolog. The results indicated that over a 7-day recovery period, OE lines consistently had higher freezing survival than WT and AtUBC2 (the Arabidopsis homolog of OsUBC7) knockout (KO) lines after a 75-min exposure to -3°C. That is, OE lines had on average 69.9% survival, while Col-0 WT and AtUBC2 KO lines had 50% and 17.1% survival, respectively (Figure 2B). These results showed a consistent enhancement of LTSS across different plant species, from the monocot rice to the eudicot Arabidopsis, highlighting potentially a general role of Rad6 type ubiquitin conjugating enzyme encoding genes in enhancing resilience of plants against short periods of low temperature stress.
3.5. OsUBC7 OE Promotes Plumule Recovery Growth and Enhances Formation of the Third Leaf After Low Temperature Exposure
As shown previously, rice lines can have relatively good LTSS values but fail to resume growth during the recovery period at higher temperatures [11]. Seedlings can remain green, indicating that the leaves are chilling tolerant, but cell division and elongation activity at the base of leaves or in the shoot apical meristem is compromised after the cold treatment. Because OsUBC7 encodes a potential RAD6 type check-point protein for the cell cycle, we investigated whether OE of this gene had a positive effect on plumule recovery growth as a potential mechanism for why transgenic lines had higher LTSS scores than WT plants after cold stress. OE lines had indeed a faster plumule growth rate than WT plants (p < 0.05), ranging from 1.68 cm/day to 1.79 cm/day (1.74 cm/day average), compared to WT plants ranging from 1.20 cm/day to 1.25 cm/day (1.23 cm/day average) (Figure 2D). Additionally, all four OE lines developed their third leaves earlier than WT plants (p < 0.05), 14.7 days after cold exposure, while the two segregated WT lines developed their third leaves 16.9 days after the cold treatment (Figure 2E).
These results showed that OE in young rice seedlings of the potential cell cycle regulatory protein encoding gene OsUBC7 significantly enhanced plumule recovery growth after cold exposure, promoting a tendency for taller OE than WT seedlings (Figure 2F). Moreover, the third leaf emerged consistently earlier in OE than WT lines, suggesting a potential role of OsUBC7 in promoting faster recovery and growth rates to replace damaged cells in response to cold stress.
3.6. Overexpression of OsUBC7 in Rice and Arabidopsis Increases Soluble Sugar Content Under Warm, Low-Temperature Exposure, and Recovery Conditions
Because OsUBC7 OE transgenic lines had improved recovery after cold stress in rice and freezing stress in Arabidopsis, we determined whether OE lines had a higher amount of soluble sugars than WT plants as a potential energy source for successful recovery. Under warm conditions, OsUBC7 OE rice lines had higher soluble sugar contents than WT plants, ranging from 3.6 to 4.1 µg/mg of tissue, compared to 3.1 to 3.2 µg/mg of tissue for WT plants, with significantly higher levels in OE-1-2 and OE-2 (p < 0.05). After 2 days of cold exposure at 10°C, all three OE lines had significantly higher levels of soluble sugar than WT plants (p < 0.05), ranging from 5.2 to 5.7 µg/mg of tissue compared to less than 4 µg/mg of tissue for WT plants. During recovery at warm temperatures, soluble sugar contents in OE lines were like those in WT plants, ranging from 3.6 to 4.8 and 2.9 to 4.2 µg/mg of tissue after one day and three days of recovery, respectively (Figure 3A).
Two OsUBC7 OE Arabidopsis lines also had significantly (p < 0.05) higher soluble sugar contents, averaging 0.51 µg/mg of tissue, compared to the Col-0 WT plants and the AtUBC2 KO line, having means of 0.43 µg/mg of tissue and 0.40 µg/mg of tissue, respectively. Following freezing treatment, OE lines maintained higher soluble sugar contents (0.64 µg/mg of tissue) than WT (0.46 µg/mg of tissue) and KO (0.52 µg/mg of tissue) lines (p < 0.05). After 2 days of stress recovery, OsUBC7 OE lines retained higher soluble sugar levels, averaging 0.5 µg/mg of tissue, while WT and KO lines had similar levels around 0.41 µg/mg of tissue (Figure 3B).
These results indicated that OsUBC7 OE transgenic lines have higher levels of soluble sugars than WT plants under both normal and low temperature conditions, potentially contributing to their improved recovery after cold stress and freezing stress. Hence, OsUBC7 may have a potential role in sugar metabolism regulation, which could be a key factor in enhancing cold stress tolerance, in both rice and Arabidopsis, and possibly, other plant species.
3.7. Overexpression of OsUBC7 Does Not Improve Membrane Damage After Low Temperature Exposure
Since OsUBC7 OE transgenic lines had better survival rates than WT plants following cold or freezing stress, we investigated whether this improvement correlated with enhanced membrane stability right after stress exposure. To assess membrane stability, electrolyte leakage (EL) from various plant parts was measured, including the second leaf, upper stem, lower stem, and roots for rice, and whole plants for Arabidopsis. In rice, EL levels were similar in OsUBC7 OE lines and WT plants in all plant tissues tested, averaging, respectively, 31.2% and 29.9% for leaves, 31.9% and 31.1% for upper stems, 24.3% and 25.5% for lower stems, and 44.2% and 48.4% for roots (Figure 3C). Similarly, OsUBC7 OE Arabidopsis lines had similar EL levels as Col-0 WT plants after freezing treatment, ranging from 20.5% to 31.9% for OE lines, and 23.5% for Col-0 WT plants (Figure 3E). These results indicated that OE of OsUBC7 does not positively affect membrane integrity in these plants, although it might have a small but non-significant positive effect in rice roots. This indicated that the positive effect on LTSS seen in OsUBC7 OE lines is not due to protection of membrane integrity during cold stress exposure, at least not as measured by the electrolyte leakage assay.
3.8. Overexpression of OsUBC7 Does Not Result in Lower Lipid Peroxidation Levels After Low Temperature Exposure
To evaluate an alternative potential positive effect of OsUBC7 OE on membrane stability, we measured malondialdehyde levels as a final product of polyunsaturated fatty acid peroxidation due to excess of reactive oxygen species (ROS) in cells extracted from different parts of rice plants, including the second leaf, upper stem, lower stem, and root. This showed that malondialdehyde levels were similar in OE and WT lines across different plant parts, with mean levels, respectively, of 3.6 and 3.2 nmol/ml/mg of tissue in leaves, 3.4 and 3.6 nmol/ml/mg of tissue in upper stems, 2.9 and 3.3 nmol/ml/mg of tissue in lower stems, and 4.3 and 4.7 nmol/ml/mg of tissue in roots. Although OsUBC7 OE lines had a trend for slightly lower malondialdehyde levels than WT plants in sections other than leaves, the differences were not significant (Figure 3D). This indicated that OE of OsUBC7 may somewhat help to reduce the degree of lipid peroxidation in roots and lower stem regions after cold stress exposure, although the effect as measured by malondialdehyde levels was not significant.
3.9. Leaves of OsUBC7 Overexpression Lines Have a Higher Cell Density Than Wild-Type Plants
Because OE of OsUBC7 boosted plumule growth rates and showed a trend toward increased plant height at the seedling stage, we investigated whether this resulted from an increase in cell elongation or an increase in cell number. We found that all four OE lines had a higher leaf cell density than segregated WT plants, ranging from 1.33 to 1.44 cells/1,000 µm2 compared to 1.02 to 1.08 cells/1,000 µm2 (p < 0.05) (Figure 4B). By contrast, OsUBC7 OE lines and WT plants had similar leaf cell lengths, ranging from 69.2 to 72.7 µm and from 69.3 to 70.6 µm, respectively (Figure 4C). Interestingly, all four OsUBC7 OE lines showed a trend for narrower leaf cells than WT plants, averaging 3.65 µm in OE lines and 4.14 µm in WT plants (Figure 4D).
3.10. Compared to Other RAD6 Paralogs OsUBC7 Is Highly Expressed at the Flowering Stage
Because public databases indicated that OsUBC7 expression was highest during the flowering stage, we analyzed mRNA levels of OsUBC7 in leaf tissues of 2-week-old seedlings and leaf tissues of plants that were at the flowering stage (3 days after emergence of the first flag leaf). Normalized gene expression levels using two different housekeeping genes confirmed that OsUBC7 mRNA abundance increased at the flowering compared to the seedling stage, with log2 fold changes of 3.24±0.20 and 3.94±0.78 in indica and japonica accessions, respectively (Table 2).
To determine whether OsUBC7 has a distinct expression pattern from other RAD6 type genes, we analyzed expression levels of two closely related paralogs, OsUBC8 and OsUBC9. Normalized gene expression levels using two different housekeeping genes showed that compared to the seedling stage, OsUBC8 mRNA levels were reduced at the flowering stage, with log2 fold changes of -1.10±0.02 and -1.34±0.56 in indica and japonica accessions, respectively (Table 2). Normalized gene expression levels using two different housekeeping genes showed that compared to the seedling stage, OsUBC9 mRNA levels did not change much at the flowering stage, with log2 fold changes of 0.64±0.33 and 0.75±0.03 in indica and japonica accessions, respectively (Table 2). Therefore, OsUBC7 had a distinct temporal gene expression pattern: while it was strongly upregulated at the flowering stage in both indica and japonica rice accessions, the other two RAD6 paralogs, OsUBC8 and OsUBC9, were either downregulated or showed little expression change at the flowering compared to the seedling stage. These findings suggested that compared to its paralogs, OsUBC7 may play a specific regulatory role during the flowering stage and when upregulated in response to cold stress.
3.11. Overexpression of OsUBC7 Tends to Improve Yield-Related Traits in Rice
Because OsUBC7 is upregulated at the flowering stage, we investigated whether OE of the gene might affect plant yield-related traits, such as total seed number, number of seeds per panicle, and fertility. Averaging results from three generations of two OsUBC7 OE lines showed that the OE lines had higher average seed yields of 80.3 and 66.9 seeds/plant for OE-1 and OE-2, respectively, than their segregated WT plants with 30.7 and 36.3 seeds/plant for WT-1 and WT-2, respectively. Similarly, the two OE lines had increased seeds per panicle of 33.3 and 32.2 seeds/panicle, compared to wild-types, 23.5 and 17.0 seeds/panicle. Additionally, OE lines had higher fertility scores of 86.4% and 83.7% compared to WT plants with 79.0% and 62.1%. Due to large standard deviations of some samples, the trend was not significant (Table 3), however, the results showed that both OE lines consistently outperformed their segregated WT plants for those yield-related phenotypes, suggesting that OE of OsUBC7 might boost seed yield traits across at least three generations.
3.12. Overexpression of OsUBC7 Does Not Alter Flowering Time
Because OsUBC7 is upregulated at the flowering stage, we investigated whether OE of OsUBC7 affected flowering time of transgenic rice lines. No significant differences in flowering time between OsUBC7 OE lines and segregated WT plants were detected. On average, all growth chamber-grown OE and WT lines flowered 98–100 days after germination (Table 3). Thus, OsUBC7 is not a new flowering time gene.
3.13. Overexpression of OsUBC7 Increases Flag Leaf Length, Width, and Dry Weight
Because OsUBC7 OE lines were generally taller than segregated WT plants, we compared the architecture of their mature plant tissues by measuring the length, width, and dry weight of the flag leaf subtending the first panicle. OE lines had significantly longer, wider, and heavier flag leaves than WT plants (p < 0.05) (Figure 5A-C). Flag leaves of OE lines had mean half lengths ranging from 18.2 to 20.5 cm, widths ranging from 0.83 to 0.93 cm, and dry weights ranging from 44 to 57 mg, while WT flag leaves had mean half lengths of 13.4 to 13.7 cm, widths of 0.51 to 0.56 cm, and dry weights of 22 to 27 mg. Overall, flag leaves of OE lines were 1.40-fold longer, 1.61-fold wider, and 2.09-fold heavier than those of WT plants. These results indicated that OE of OsUBC7 significantly (p < 0.05) alters plant architecture at the flowering stage, leading to longer, wider, and heavier flag leaves compared to WT plants, suggesting a significant impact of OsUBC7 OE on plant morphology during the flowering stage.
3.14. Overexpression of OsUBC7 Leads to an Increase of Chlorophyll Content in Flag Leaves
Since OE of OsUBC7 affected soluble sugar content and flag leaf architecture, we investigated whether OE also had an effect of the leaf chlorophyll content. OE lines had a significantly higher content of chlorophyll A than segregated WT lines (p < 0.05), ranging from 16.5 to 18.3 µg/mL, while values in WT flag leaves ranged from 10.2 to 11.7 µg/mL (Figure 5D). On the other hand, OE lines had a similar chlorophyll B content as WT plants, with values ranging from 7.6 to 9.4 µg/mL, while WT plants had values ranging from 6.4 to 7.9 µg/mL (Figure 5E). This showed that OE of OsUBC7 significantly increased chlorophyll A but not chlorophyll B content in flag leaves, possibly leading to enhanced photosynthetic capacity at this stage.
3.15. OsUBC7 Interacts with the E3 Protein OsRFPH2-12 and the RING Finger Domain of the Two E3 Proteins OsHUB1 and OsHUB2
Because E2 ubiquitin conjugating enzymes form complexes with E3 ligases and target proteins, we interrogated the String database (https://string-db.org/) to identify potential UBC7 interacting proteins. From this we selected an E3 ligase expressed during the flowering stage, OsRFPH2-12, and two histone E3 ligases potentially involved in cell cycle regulation, OsHUB1 and OsHUB2. To determine whether OsUBC7 physically interacted with those E3 ligases, we performed yeast two-hybrid assays, indicating that OsUBC7 interacted with the full-length OsRFPH2-12 protein and the RING finger domains of OsHUB1/HUB2, as evidenced by robust growth on quadruple selective media lacking adenine, histidine, tryptophan, and leucine (Figure 6). These findings suggested that OsUBC7 may act as an E2 ubiquitin conjugating enzyme in concert with developmental and cell cycle stage specific E3 ligases to regulate protein abundance and/or protein function.
Since the protein sequences of OsUBC7 and its Arabidopsis homolog AtUBC2 are highly conserved by sharing 145/152 identical and 151/152 positive amino acids, we performed an unbiased yeast two-hybrid screen with OsUBC7 as bait and a commercial Arabidopsis cDNA library made from all tissues and developmental stages as prey. This screen yielded E3 ligases, cell cycle dependent kinases, other kinases, transcription factors, enzymes, a hydroxyproline-rich glycoprotein, and a GCN5-type histone modifying acetyltransferase (Supplementary Table S3). These proteins are potential OsUBC7 targets that might have roles in cold stress tolerance response mechanisms, and rice homologs of those proteins merit future investigations.
3.16. Overexpression of OsUBC7 Tends to Increase Germination Rates of Arabidopsis Seeds Under High Salinity Stress
Because abiotic stress response mechanisms often converge through a network of coordinated biochemical and molecular pathways, we investigated whether OsUBC7 could improve high salinity stress tolerance by performing germination assays with seeds from homozygous OsUBC7 OE Arabidopsis lines and WT control plants. We found significant differences in germination rates between OsUBC7 OE and WT lines over the two-week period on 50 and 100 mM NaCl (Supplementary Table S4). This suggested that OsUBC7 may play a role in different abiotic stress tolerance response mechanisms.
4. Discussion
4.1. OsUBC7 May Mediate Cold Stress Tolerance Responses Through Sugar Metabolism and Growth Regulation
The identification of OsUBC7 as a candidate gene associated with a low-temperature seedling survivability (LTSS) trait QTL in rice [2] is supported by its differential expression patterns during cold induction and single-nucleotide polymorphisms (SNPs) between chilling-tolerant and chilling-sensitive accessions (Table 1; Figure 1A). The significant correlation between OsUBC7 haplotypes and LTSS scores (Figure 1C) further emphasizes its potential in modulating cold stress tolerance responses in rice.
OsUBC7 overexpression (OE) lines overall had higher LTSS scores than wild-type (WT) plants after cold treatment, which agrees with the hypothesis that upregulation of this gene during cold helps boost plant resilience to low-temperature stress in both rice and Arabidopsis (Figure 2A–B). However, the similar LTSS scores between OE and WT lines after extended cold exposure suggests that OsUBC7-mediated stress tolerance mechanisms are limited to early timepoints of stress exposure.
Our findings suggest that a potential mechanism involved in boosting cold stress resilience in OsUBC7 OE lines might be the higher growth rate in OE than WT lines before and after cold stress (Figure 2C–D), which may be critical for successful recovery and enhanced survivability. This might occur through the restoration of cellular homeostasis and regulation of new cell growth, particularly in leaf cells (Figure 2E), which are critical for photosynthesis and metabolic activity [6,24,25,26]. The increased growth rate and higher number of cells per unit leaf area (Figure 4A-B) might contribute to the elevated levels of soluble sugars observed in OsUBC7 OE lines before, during, and after cold or freezing stress (Figure 3A-B). Boosting of cold tolerance via growth regulation has been previously observed. For example, RNAi-mediated knockdown of OsOFP6 reduced the number of leaf cells, which reduced the percent of starch-filled pollen grains under cold treatment [27]. OE of OsCYP19-4 promoted an increase in tiller and spike numbers and grain weight, which correlated with enhanced cold tolerance [28]. OE of OsRAN2, an essential protein for mitosis, enhanced cold tolerance by promoting export of intranuclear tubulin and maintaining cell division under cold stress and recovery [29]. And OE of OsMYB3R-2 helped with cell cycle progression during chilling stress [30]. These findings agree with our interpretation that OE of OsUBC7 might boost LTSS scores by increasing the growth rate and soluble sugar content of transgenic plants.
The observed increase in soluble sugar levels before stress may serve as a critical energy source for OsUBC7 OE plants during cold stress and subsequent recovery periods, facilitating essential metabolic processes required for recovery and survival (Figure 2C–D, Figure 3A) [10,31,32]. Interestingly, the increase in soluble sugar content peaked during cold or freezing stress for OsUBC7 OE plants, while it peaked one day after recovery in WT plants (Figure 3A-B), indicating a dynamic regulation of genes, proteins, and metabolites throughout the stress-response period [33,34,35]. In agreement with this, OE of OsSUT1-5 increased total sucrose levels prior to chilling treatment and helped with the plant phloem loading of sucrose [36], and OE of OsTERF2 increased accumulation of soluble sugar and other osmotic substances as well as overall chlorophyll content [37]. Moreover, OE of OsCIPK03 and OsCIPK12 significantly increased soluble sugar contents and improved cold stress tolerance [38], and OE of the japonica allele of OsbZIP73 increased soluble sugar contents under chilling stress [39]. Thus, in agreement with these studies, elevated soluble sugar contents in OsUBC7 OE lines suggests a potential role in regulating cold tolerance by modulating soluble sugar mediated metabolic processes.
Work with the Arabidopsis homolog of OsUBC7 showed that knocking out AtUBC2 significantly reduced histone H2B mono-ubiquitination, leading to gene- and protein-specific changes in the patterns of histone ubiquitination and methylation [40]. AtUBC2 moreover was shown to have roles in regulating protein homeostasis and growth of Arabidopsis plants under high salinity conditions [41]. Furthermore, overexpression of the soybean RAD6 homolog GmUBC9 increased drought resistance and affected flowering time via histone H2B mono-ubiquitination in both Arabidopsis and soybean plants [42]. In agreement, our germination assays showed that OsUBC7 OE Arabidopsis lines maintained nearly 100% germination rates compared to WT under high salinity stress (Supplementary Table S4), indicating that OsUBC7 is involved in other stress tolerances mechanisms.
Interestingly, an increase in soluble sugar content has been previously linked to maintenance of plasma membrane integrity [1,4,9,10], however, our OsUBC7 OE lines did not have lower electrolyte leakage or lipid peroxidation levels than wild-type plants after cold stress (Figure 3C–E). One explanation for this is that the observed increase in soluble sugar in OsUBC7 OE plants provided energy for other metabolic processes including cell cycle progression, leading to higher number of cells per leaf area and better recovery growth and development after cold stress exposure.
4.2. Dynamic Expression of OsUBC7 in Response to Cold Stress and Flowering
We show here that mRNA abundance of OsUBC7 increases in response to prolonged cold treatment, but the extent of upregulation was higher in cold tolerant japonica than cold sensitive indica (Table 2). This differential response may contribute, at least in part, to the higher cold tolerance potential of japonica than indica accessions [1,2,12]. However, both subspecies probably employ the E2 conjugating enzyme function of OsUBC7 as part of a conserved cold stress tolerance response mechanism that might positively affect protein homeostasis [43,44], which involves maintaining a balanced and functional proteome within cells, ensuring proper protein folding, localization, and function [45,46]. Interfering with proteostasis mechanism can have both positive and negative effects [47,48]. For example, OE of the two E3 ligase encoding genes OsPUB2/PUB3 or OsDIRP1 boosted cold stress tolerance and chlorophyll content of transgenic rice [49,50]. Conversely, OE of the E3 ligase encoding gene OsATL38 led to turnover OsGF14, a positive regulator of cold tolerance, thus, negatively affecting cold stress tolerance in rice [51], while, knocking down the stress inducible E3 ligase encoding gene OsSRFP1 improved cold stress tolerance [52]. Taken together, upregulating genes such as OsUBC7 in both indica and japonica accessions may be beneficial for maintaining protein homeostasis during cold stress and during recovery after stress exposure [43,44,45,46].
Based on its expression profile, OsUBC7 may have a predominant function during the flowering stage of rice development, because compared to its Rad6 paralogs OsUBC8 and OsUBC9, it was specifically upregulated at that developmental stage (Table 3). These expression profiles agree with RNA-seq and microarray results deposited into publicly available databases such as RiceXPro, the Rice RNA-seq Database, and the Rice Genome Annotation Project [14,19,53]. To the best of our knowledge, we show here for the first time that the E2 conjugating enzyme encoding gene OsUBC7 has an additional function as a cold stress upregulated gene, which implies that the OsUBC7 protein might interact with different E3 ligase partners and target proteins at the flowering than during cold induction at the young seedling stage.
4.3. A Role for OsUBC7 in Promoting Rice Cell Proliferation
As a RAD6 type protein, OsUBC7 has a predicted role as a checkpoint protein in the cell cycle [14,53], suggesting a role in regulating cell proliferation and expansion. This prediction is supported by the increased growth rates we observed in OsUBC7 OE seedlings (Figure 2C–F; Figure 5A–C). The increased growth rate promoted by OsUBC7 OE was mediated by an increase in cell density per unit leaf area rather than cell elongation (Figure 4B), suggesting that OsUBC7 may primarily regulate cell proliferation rather than cell elongation. From the very well-studied functions of RAD6 in yeast as a protein involved in DNA repair and cell cycle regulation [17,18,54], we can draw further parallels to the potential molecular mechanisms underlying OsUBC7-mediated growth regulation. Because yeast RAD6 is upregulated during the G1/S and S/G2 phase transitions to serve as a checkpoint protein during cell division [16,54], OsUBC7 may similarly play a role in modulating cell cycle progression and growth dynamics. The longer cells stay in G1 the larger they get [55,56], therefore, it is possible that OsUBC7 may shorten the cell cycle by speeding up the transition from G1 to S phase, thus making the cell narrower and increase cell density over time. In support of this, OE of RAD6 in human cells led to a quicker transition from G1 to S phase and to increased cell proliferation [13]. Taken together, OsUBC7 might have a similar role as its yeast homolog RAD6 in regulating cell cycle progression and growth dynamics in rice, possibly by promoting a quicker transition from G1 to S phase, leading to increased cell density and proliferation.
4.4. A Role for OsUBC7 in Yield Enhancement and Flowering Regulation of Rice
Our analysis of yield related traits revealed a strong trend toward increases in seed yield, seeds per panicle, and fertility in OsUBC7 OE compared to segregated WT lines (Table 4). Since OsUBC7 is located on at 3.5 Mb segment of chromosome 7 that overlaps with several yield-related trait QTL such as number of grains per panicle (gp7a), 1,000-grain weight (gw7), yield per plant (yd7a), and number of tillers per plant (tp7b) [57,58], it is possible that OsUBC7 is a candidate gene for those traits, as supported by our observations. However, this trend toward higher yields was not due to changes in flowering time, because OsUBC7 OE and WT lines had similar heading dates (Table 4). Thus, OsUBC7 is not a novel flowering time gene. This result agrees with flowering time studies in Arabidopsis where it was shown that although the OsUBC7 homolog AtUBC2 was involved in histone H2B mono-ubiquitination in the chromatin region of the flowering repressor gene FLOWERING LOCUS C [59,60], OE of AtUBC2 did not affect flowering time [40].
Additionally, OE of OsUBC7 effected flag leaf morphology and physiology, leading to longer, wider, and heavier flag leaves compared to segregated WT plants (Figure 5A–C). This suggests that OsUBC7, annotated as a cell division check-point protein encoding gene [53], may play a role in regulating leaf growth and development during the flowering stage, where it has the highest expression levels. Interestingly, OE of OsUBC7 also resulted in higher chlorophyll A content in the flag leaves compared to WT plants (Figure 5D). Together with a higher cell density, the higher chlorophyll content most likely led to enhanced photosynthetic activity and potentially improved plant vigor in OsUBC7 OE lines [61,62,63].
4.5. Interaction of OsUBC7 with E3 Type RING Finger Domain and Potential Target Proteins
Our yeast two-hybrid assay showed that OsUBC7 physically interacts with OsRFPH2-12, an E3 ligase co-expressed with OsUBC7 during flowering in rice, and with the RING finger domain of the more broadly expressed E3 ligases OsHUB1/2, indicating that OsUBC7 has different E3 partners that may have different roles in rice development and stress tolerance response mechanisms. OsHUB1 and OsHUB2 are involved in mono-ubiquitinating histone H2B [65,66,67], a key modification that regulates chromatin structure and gene expression [68,69]. This histone modification affects expression of genes involved in both normal growth and abiotic stress responses [42,49,68,69], and most likely facilitates upregulation of cold responsive genes such as those coding for CBF/DREB1 transcription factors that are early regulators of the cold stress tolerance responses [70,71] by activating COR genes that help stabilize cell membranes, protect cellular proteins, and maintain osmotic balance under low temperatures stress [72,73].
In addition to regulating gene expression, OsUBC7 likely contributes to protein turnover through its interaction with the E3 ligase OsRFPH2-12, which may target key regulatory proteins for degradation during the flowering stage of rice development [74]. During both developmental switches and cold stress responses, cells must rapidly degrade obsolete or damaged proteins, as well as negative regulators of stress response pathways to enable the activation of protective mechanisms [75,76,77]. Flowering is a highly regulated developmental event, influenced by environmental cues such as day length and temperature [77,78,79], and timely protein degradation of specific inhibitors is required to ensure that flowering signals can proceed [60,75,79]. Perhaps the main function of OsUBC7 during rice development is degradation of proteins that inhibit flowering by interacting with developmental stage specific E3 ligases such as OsRFPH2-12, while it assumes a specialized function during cold stress tolerance responses by complexing with different E3 partners such as OsHUB1 or OsHUB2 to mono-ubiquitinate histones. A better understanding of how OsUBC7 integrates these different functions will provide valuable insights into potential strategies for enhancing cold tolerance and perhaps yield in rice, such as via targeted manipulation of ubiquitination pathways to optimize stress response and growth. Identification of additional developmental stage or cold stress response E3 partners and protein targets will help with this task. We have already obtained value information using an unbiased yeast-two hybrid screen (Supplementary Table S3), and future studies on rice homologs of those OsUBC7 interacting Arabidopsis proteins are merited.
5. Conclusions
This study revealed multiple roles for OsUBC7 in the rice cold temperature stress tolerance response, highlighting its involvement in orchestrating sugar metabolism and growth regulation. Although the main function of OsUBC7 in rice most likely occurs during the flowering stage, we uncovered via genomics, differential gene expression and overexpression analyses an additional function for OsUBC7 as a cold induced gene, indicating that it has a potential for enhancing plant fitness in response to low-temperature and possibly other abiotic stress. We propose that an interaction of OsUBC7 with the histone H2B mono-ubiquitinating E3 ligases OsHUB1 and OsHUB2 has a role in the early cold stress tolerance response mechanisms by activating positive regulators of the stress response and metabolic processes leading to higher amounts of soluble sugars providing resources for growth and development during stress recovery. This study contributes to the recent interest in exploring the involvement of OsUBC genes in low-temperature germination (via ABA signaling; [81]), root development (via auxin signaling; [82]), pathogen response [83,84], and regulation of plant height and yield [84,85].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Sequences of primers used in this study; Table S2: qPCR measurements of mRNA levels in 2-week-old seedlings containing OsUBC7 overexpression (OE) constructs in the Kasalath (aus subgroup) genetic background compared to their respective wild-type (WT) plants under warm growth conditions; Table S3: Arabidopsis genes corresponding to expressed sequence tags (EST) isolated from a yeast two-hybrid screen using OsUBC7 as bait; Table S4: Germination rates of Arabidopsis seeds from Col-0 wild-type (WT) plants and OsUBC7 overexpression (OE) lines under different NaCl concentrations (0, 50, 100 mM).
Author Contributions
Conceptualization, M.S. and H.P.; methodology, M.S. and H.P.; validation, M.S. and H.P.; formal analysis, M.S. and H.P.; investigation, H.P.; resources, M.S.; writing—original draft preparation, H.P.; writing—review and editing, M.S.; supervision, M.S..; project administration, M.S.; funding acquisition, M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by USDA-AFRI-NIFA grant number 2016-67013-24587, and in part by USDA-AFRI-NIFA grant number 2023-67013-39534.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We are grateful to Marquette University undergraduate students Sir Lawrence Tender and Samantha Malinis for helping as part of their BIOL 4956 supervised research experiences with qPCR and soluble sugar extraction experiments, respectively.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schläppi, M. R.; Jessel, A. R.; Jackson, A. K.; Phan, H.; Jia, M. H.; Edwards, J. D.; Eizenga, G. C. Navigating Rice Seedling Cold Resilience: QTL Mapping in Two Inbred Line Populations and the Search for Genes. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, N.; Johnson, M.; Beaumont, A.; Schläppi, M. Multiple Cold Tolerance Trait Phenotyping Reveals Shared Quantitative Trait Loci in Oryza Sativa. Rice 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Dolferus, R. To Grow or Not to Grow: A Stressful Decision for Plants. Plant Science 2014, 229, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Nasim Maghboli Balasjin; Maki, J. S.; Schläppi, M. R. Pseudomonas Mosselii Improves Cold Tolerance of Asian Rice (Oryza Sativa L.) in a Genotype-Dependent Manner by Increasing Proline in Japonica and Reduced Glutathione in Indica Varieties. Canadian Journal of Microbiology 2023, 70, 15–31. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Chong, K.; Xu, Y. Chilling Tolerance in Rice: Past and Present. Journal of Plant Physiology 2022, 268, 153576. [Google Scholar] [CrossRef]
- Gazquez, A.; Abdelgawad, H.; Baggerman, G.; Van Raemdonck, G.; Asard, H.; Maiale, S. J.; Rodríguez, A. A.; Beemster, G. T. S. Redox Homeostasis in the Growth Zone of the Rice Leaf Plays a Key Role in Cold Tolerance. J. Exp. Bot. 2020, 71(3), 1053–1066. [Google Scholar] [CrossRef]
- Nie, L.; Liu, H.; Zhang, L.; Wang, W. Enhancement in Rice Seed Germination via Improved Respiratory Metabolism under Chilling Stress. Food and Energy Security 2020, 9. [Google Scholar] [CrossRef]
- Arshad, M. S.; Farooq, M.; Asch, F.; Krishna, J. S. V.; Prasad, P. V. V.; Siddique, K. H. M. Thermal Stress Impacts Reproductive Development and Grain Yield in Rice. Plant Physiology and Biochemistry 2017, 115, 57–72. [Google Scholar] [CrossRef]
- Shi, Y.; Phan, H.; Liu, Y.; Cao, S.; Zhang, Z.; Chu, C.; Schläppi, M. R. Glycosyltransferase OsUGT90A1 Helps Protect the Plasma Membrane during Chilling Stress in Rice. J. Exp. Bot. 2020, 71(9), 2723–2739. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Gao, G.; Ali, I.; Wu, X.; Tang, M.; Chen, L.; Jiang, L.; Liang, T. Effects of Various Seed Priming on Morphological, Physiological, and Biochemical Traits of Rice under Chilling Stress. 2023, 14. [CrossRef]
- Schläppi, M. R.; Jackson, A. K.; Eizenga, G. C.; Wang, A.; Chu, C.; Shi, Y.; Shimoyama, N.; Boykin, D. L. Assessment of Five Chilling Tolerance Traits and GWAS Mapping in Rice Using the USDA Mini-Core Collection. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Phan, H.; Schläppi, M. Low Temperature Antioxidant Activity QTL Associate with Genomic Regions Involved in Physiological Cold Stress Tolerance Responses in Rice (Oryza Sativa L.). Genes 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Chen, P.; Chen, L.; Biskup, E.; Liu, Y.; Chen, P.-C.; Chang, J.-F.; Jiang, W.; Jing, Y.; Chen, Y.; Jin, H.; Chen, S. Human RAD6 Promotes G1-S Transition and Cell Proliferation through Upregulation of Cyclin D1 Expression. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J. P.; Kanamori, H.; McCombie, W. R.; Ouyang, S.; Schwartz, D. C.; Tanaka, T.; Wu, J.; Zhou, S.; Childs, K. L.; Davidson, R. M.; Lin, H.; Quesada-Ocampo, L.; Vaillancourt, B.; Sakai, H.; Lee, S. S.; Kim, J.; Numa, H.; Itoh, T. Improvement of the Oryza Sativa Nipponbare Reference Genome Using next Generation Sequence and Optical Map Data. Rice 2013, 6. [Google Scholar] [CrossRef]
- Sato, Y. S. ; Hinako Takehisa; Kaori Kamatsuki; Minami, H. ; Nobukazu Namiki; Hiroshi Ikawa; Hajime Ohyanagi; Sugimoto, K.; Antonio, B. A.; Yoshiaki Nagamura. RiceXPro Version 3.0: Expanding the Informatics Resource for Rice Transcriptome. 2012, 41, D1206–D1213. [Google Scholar] [CrossRef]
- Game, J. C.; Williamson, M. S.; Spicakova, T.; Brown, J. M. The RAD6/BRE1 Histone Modification Pathway in Saccharomyces Confers Radiation Resistance through a RAD51-Dependent Process That Is Independent of RAD18. Genetics 2006, 173, 1951–1968. [Google Scholar] [CrossRef]
- Lyakhovich, A.; Shekhar, M. P. V. RAD6B Overexpression Confers Chemoresistance: RAD6 Expression during Cell Cycle and Its Redistribution to Chromatin during DNA Damage-Induced Response. Oncogene 2004, 23, 3097–3106. [Google Scholar] [CrossRef]
- Madura, K.; Prakash, S.; Prakash, L. Expression of theSaccharomyces CerevisiaeDNA Repair GeneRAD6That Encodes a Ubiquitin Conjugating Enzyme, Increases in Response to DNA Damage and in Meiosis but Remains Constant during the Mitotic Cell Cycle. Nucleic Acids Research 1990, 18, 771–778. [Google Scholar] [CrossRef]
- Chao, H.; Zhang, S.; Hu, Y.; Ni, Q.; Xin, S.; Zhao, L.; Ivanisenko, V. A.; Orlov, Y. L.; Chen, M. Integrating Omics Databases for Enhanced Crop Breeding. Journal of integrative bioinformatics 2023, 20, 20230012. [Google Scholar] [CrossRef]
- Kerepesi, I.; Tóth, M.; Boross, L. Water-Soluble Carbohydrates in Dried Plant. Journal of Agricultural and Food Chemistry 1996, 44, 3235–3239. [Google Scholar] [CrossRef]
- Leach, K. A.; Braun, D. M. Soluble Sugar and Starch Extraction and Quantification from Maize (Zea Mays) Leaves. Current Protocols in Plant Biology 2016, 1, 139–161. [Google Scholar] [CrossRef]
- Ritchie, R. J. Consistent Sets of Spectrophotometric Chlorophyll Equations for Acetone, Methanol and Ethanol Solvents. Photosynthesis Research 2006, 89, 27–41. [Google Scholar] [CrossRef]
- Xu, D.; Huang, X.; Xu, Z.-Q.; Schläppi, M. The HyPRP Gene EARLI1 Has an Auxiliary Role for Germinability and Early Seedling Development under Low Temperature and Salt Stress Conditions in Arabidopsis Thaliana. Planta 2011, 234, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xin, W.; Wang, Y.; Zhang, D.; Wang, J.; Zheng, H.; Yang, L.; Nie, S.; Zou, D. An Integrated Analysis of the Rice Transcriptome and Lipidome Reveals Lipid Metabolism Plays a Central Role in Rice Cold Tolerance. BMC Plant Biology 2022, 22. [Google Scholar] [CrossRef] [PubMed]
- Hajihashemi, S.; Noedoost, F.; Geuns, J. M. C.; Djalovic, I.; Siddique, K. H. M. Effect of Cold Stress on Photosynthetic Traits, Carbohydrates, Morphology, and Anatomy in Nine Cultivars of Stevia Rebaudiana. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Gammulla, C. G.; Pascovici, D.; Atwell, B. J.; Haynes, P. A. Differential Proteomic Response of Rice (Oryza Sativa ) Leaves Exposed to High- and Low-Temperature Stress. PROTEOMICS 2011, 11, 2839–2850. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, C.; He, Y.; Tian, Z.; Li, J. Rice OVATE Family Protein 6 Regulates Plant Development and Confers Resistance to Drought and Cold Stresses. Journal of Experimental Botany 2017, 68, 4885–4898. [Google Scholar] [CrossRef]
- Yoon, D. H.; Lee, S. S.; Park, H. J.; Lyu, J. I.; Chong, W. S.; Liu, J. R.; Kim, B.-G.; Ahn, J. C.; Cho, H. S. Overexpression ofOsCYP19-4Increases Tolerance to Cold Stress and Enhances Grain Yield in Rice (Oryza Sativa). Journal of Experimental Botany 2015, 67, 69–82. [Google Scholar] [CrossRef]
- Chen, N.; Xu, Y.; Wang, X.; Du, C.; Du, J.; Yuan, M.; Xu, Z.; Chong, K. OsRAN2, Essential for Mitosis, Enhances Cold Tolerance in Rice by Promoting Export of Intranuclear Tubulin and Maintaining Cell Division under Cold Stress. Plant, Cell & Environment 2010, 34, 52–64. [Google Scholar] [CrossRef]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.-H.; Zhang, X.; Chong, K. Enhanced Tolerance to Chilling Stress in OsMYB3R-2 Transgenic Rice Is Mediated by Alteration in Cell Cycle and Ectopic Expression of Stress Genes. Plant Physiology 2009, 150, 244–256. [Google Scholar] [CrossRef]
- De Freitas, G. M.; Thomas, J.; Liyanage, R.; Lay, J. O.; Basu, S.; Ramegowda, V.; do Amaral, M. N.; Benitez, L. C.; Bolacel Braga, E. J.; Pereira, A. Cold Tolerance Response Mechanisms Revealed through Comparative Analysis of Gene and Protein Expression in Multiple Rice Genotypes. PLOS ONE 2019, 14, e0218019. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, Q.; Wang, S.; Hong, Y.; Wang, Z. Rice and Cold Stress: Methods for Its Evaluation and Summary of Cold Tolerance-Related Quantitative Trait Loci. Rice 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Singh, A. K.; Dhanapal, S.; Yadav, B. S. The Dynamic Responses of Plant Physiology and Metabolism during Environmental Stress Progression. Molecular Biology Reports 2019, 47, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Furtauer, L.; Weiszmann, J.; Weckwerth, W.; Nägele, T. Dynamics of Plant Metabolism during Cold Acclimation. International Journal of Molecular Sciences 2019, 20, 5411. [Google Scholar] [CrossRef]
- Yang, M.; Yang, J.; Su, L.; Sun, K.; Li, D.; Liu, Y.; Wang, H.; Chen, Z.; Guo, T. Metabolic Profile Analysis and Identification of Key Metabolites during Rice Seed Germination under Low-Temperature Stress. Plant Science 2019, 289, 110282. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Meguro-maoka, A.; Yoshida, M. Analysis of Sugar Content and Expression of Sucrose Transporter Genes (OsSUTs) in Rice Tissues in Response to a Chilling Temperature. Japan Agricultural Research Quarterly: JARQ 2017, 51, 137–146. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, H.; Pan, X.; Chen, X.; Zhang, Z.; Lu, X.; Huang, R. Overexpression of Ethylene Response Factor TERF2 Confers Cold Tolerance in Rice Seedlings. Transgenic Research 2010, 20, 857–866. [Google Scholar] [CrossRef]
- Xiang, Y.; Huang, Y.; Xiong, L. Characterization of Stress-Responsive CIPK Genes in Rice for Stress Tolerance Improvement. Plant Physiol. 2007, 144(3), 1416–1428. [Google Scholar] [CrossRef]
- Liu, C.; Schläppi, M.; Mao, B.; Wang, W.; Wang, A.; Chu, C. The BZIP73 Transcription Factor Controls Rice Cold Tolerance at the Reproductive Stage. Plant Biotechnology Journal 2019, 17, 1834–1849. [Google Scholar] [CrossRef]
- Xu, L.; Rozenn Ménard; Berr, A.; Fuchs, J.; Valérie Cognat; Meyer, D.; Shen, W.-H. The E2 Ubiquitin-Conjugating Enzymes, AtUBC1 and AtUBC2, Play Redundant Roles and Are Involved in Activation ofFLCExpression and Repression of Flowering inArabidopsis Thaliana. Plant Journal 2009, 57, 279–288. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, J.; Li, X.; Li, Y. E2 Conjugases UBC1 and UBC2 Regulate MYB42-Mediated SOS Pathway in Response to Salt Stress in Arabidopsis. New Phytologist 2020, 227, 455–472. [Google Scholar] [CrossRef]
- Chen, K.; Tang, W.-S.; Zhou, Y.-B.; Xu, Z.-S.; Chen, J.; Ma, Y.-Z.; Chen, M.; Li, H.-Y. Overexpression of GmUBC9 Gene Enhances Plant Drought Resistance and Affects Flowering Time via Histone H2B Monoubiquitination. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- E, Z.; Zhang, Y.; Li, T.; Wang, L.; Zhao, H. Characterization of the Ubiquitin-Conjugating Enzyme Gene Family in Rice and Evaluation of Expression Profiles under Abiotic Stresses and Hormone Treatments. PLOS ONE 2015, 10, e0122621. [Google Scholar] [CrossRef]
- Bae, H.; Kim, W. T. Classification and Interaction Modes of 40 Rice E2 Ubiquitin-Conjugating Enzymes with 17 Rice ARM-U-Box E3 Ubiquitin Ligases. Biochemical and Biophysical Research Communications 2014, 444, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.; Pang, Y.; Bao, J. Comparative Ubiquitome Analysis Reveals Diverse Functions of Ubiquitination in Rice Seed Development under High-Temperature Stress. Seed Biology 2023, 2. [Google Scholar] [CrossRef]
- Zhu, L.; Cheng, H.; Peng, G.; Wang, S.; Zhang, Z.; Ni, E.; Fu, X.; Zhuang, C.; Liu, Z.; Zhou, H. Ubiquitinome Profiling Reveals the Landscape of Ubiquitination Regulation in Rice Young Panicles. Genomics Proteomics & Bioinformatics 2020, 18, 305–320. [Google Scholar] [CrossRef]
- Melo, F. V.; Oliveira, M. M.; Saibo, N. J. M.; Lourenço, T. F. Modulation of Abiotic Stress Responses in Rice by E3-Ubiquitin Ligases: A Promising Way to Develop Stress-Tolerant Crops. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef]
- Dametto, A.; Buffon, G.; Dos Reis Blasi, É. A.; Sperotto, R. A. Ubiquitination Pathway as a Target to Develop Abiotic Stress Tolerance in Rice. Plant Signaling & Behavior 2015, 10, e1057369. [Google Scholar] [CrossRef]
- Byun, M. Y.; Cui, L. H.; Oh, T. K.; Jung, Y.-J.; Lee, A.; Park, K. Y.; Kang, B. G.; Kim, W. T. Homologous U-Box E3 Ubiquitin Ligases OsPUB2 and OsPUB3 Are Involved in the Positive Regulation of Low Temperature Stress Response in Rice (Oryza Sativa L.). Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Cui, L. H.; Min, H. J.; Byun, M. Y.; Oh, H. G.; Kim, W. T. OsDIRP1, a Putative RING E3 Ligase, Plays an Opposite Role in Drought and Cold Stress Responses as a Negative and Positive Factor, Respectively, in Rice (Oryza Sativa L.). Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef]
- Cui, L. H.; Min, H. J.; Yu, S. G.; Byun, M. Y.; Oh, T. R.; Lee, A.; Yang, H. W.; Kim, W. T. OsATL38 Mediates Mono-Ubiquitination of the 14-3-3 Protein OsGF14d and Negatively Regulates the Cold Stress Response in Rice. Journal of Experimental Botany 2021, 73, 307–323. [Google Scholar] [CrossRef]
- Fang, H.; Meng, Q.; Xu, J.; Tang, H.; Tang, S.; Zhang, H.; Huang, J. Knock-down of Stress Inducible OsSRFP1 Encoding an E3 Ubiquitin Ligase with Transcriptional Activation Activity Confers Abiotic Stress Tolerance through Enhancing Antioxidant Protection in Rice. Plant Molecular Biology 2015, 87, 441–458. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y. S. ; Hinako Takehisa; Kaori Kamatsuki; Minami, H. ; Nobukazu Namiki; Hiroshi Ikawa; Hajime Ohyanagi; Sugimoto, K.; Antonio, B. A.; Yoshiaki Nagamura. RiceXPro Version 3.0: Expanding the Informatics Resource for Rice Transcriptome. 2012, 41, D1206–D1213. [Google Scholar] [CrossRef]
- Verkade, H.; Teli, T.; Laursen, L.; Murray, J.; O’Connell, M. A Homologue of the Rad18 Postreplication Repair Gene Is Required for DNA Damage Responses throughout the Fission Yeast Cell Cycle. Molecular Genetics and Genomics 2001, 265, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, C.; Skotheim, J. M.; de Bruin, R. A. M. Control of Cell Cycle Transcription during G1 and S Phases. Nature Reviews Molecular Cell Biology 2013, 14, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Moeller, S. J.; Sheaff, R. J. G1 Phase: Components, Conundrums, Context. Results and Problems in Cell Differentiation 1–29. [CrossRef]
- Mansueto, L.; Fuentes, R. R.; Borja, F. N.; Detras, J.; Abriol-Santos, J. M.; Chebotarov, D.; Sanciangco, M.; Palis, K.; Copetti, D.; Poliakov, A.; Dubchak, I.; Solovyev, V.; Wing, R. A.; Hamilton, R. S.; Mauleon, R.; McNally, K. L.; Alexandrov, N. Rice SNP-Seek Database Update: New SNPs, Indels, and Queries. Nucleic Acids Research 2016, 45, D1075–D1081. [Google Scholar] [CrossRef]
- Yonemaru, J.; Yamamoto, T.; Fukuoka, S.; Uga, Y.; Hori, K.; Yano, M. Q-TARO: QTL Annotation Rice Online Database. Rice 2010, 3, 194–203. [Google Scholar] [CrossRef]
- Gu, X.; Jiang, D.; Wang, Y.; Bachmair, A.; He, Y. Repression of the Floral Transition via Histone H2B Monoubiquitination. The Plant Journal 2009, 57, 522–533. [Google Scholar] [CrossRef]
- Cao, Y.; Dai, Y.; Cui, S.; Ma, L. Histone H2B Monoubiquitination in the Chromatin of FLOWERING LOCUS C Regulates Flowering Time in Arabidopsis. Plant Cell 2008, 20(10), 2586–2602. [Google Scholar] [CrossRef]
- Alemu, S. T. Photosynthesis Limiting Stresses under Climate Change Scenarios and Role of Chlorophyll Fluorescence: A Review Article. Cogent Food & Agriculture 2020, 6, 1785136. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A. M. Responses of Photosynthesis, Chlorophyll Fluorescence and ROS-Scavenging Systems to Salt Stress during Seedling and Reproductive Stages in Rice. Annals of Botany 2007, 99, 1161–1173. [Google Scholar] [CrossRef]
- Tanaka, A.; Tanaka, R. Chlorophyll Metabolism. Current Opinion in Plant Biology 2006, 9, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Zheng, G.; Hamdani, S.; Jemâa Essemine; Song, Q.; Wang, H.; Chu, C.; Sirault, X.; Zhu, X. Leaf Photosynthetic Parameters Related to Biomass Accumulation in a Global Rice Diversity Survey. Plant Physiology 2017, 175, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Tang, N.; Li, X.; Xie, Y.; Xiang, D.; Fu, J.; Shen, J.; Yang, J.; Tu, H.; Li, X.; Hu, H.; Xiong, L. Reversible Histone H2B Monoubiquitination Fine-Tunes Abscisic Acid Signaling and Drought Response in Rice. 2019, 12 (2), 263–277. [CrossRef]
- Zhou, W.; Zhu, Y.; Dong, A.; Shen, W.-H. Histone H2A/H2B Chaperones: From Molecules to Chromatin-Based Functions in Plant Growth and Development. The Plant Journal 2015, 83, 78–95. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Li, X.; Wang, Z.; Ding, M.; Sun, Y.; Dong, F.; Chen, F.; Liu, L.; Doughty, J.; Li, Y.; Yong Xiu Liu. Histone H2B Monoubiquitination Mediated by HISTONE MONOUBIQUITINATION1 and HISTONE MONOUBIQUITINATION2 Is Involved in Anther Development by Regulating Tapetum Degradation-Related Genes in Rice. Plant Physiology 2015, 168, 1389–1405. [Google Scholar] [CrossRef]
- Min, H. J.; Jung, Y. J.; Kang, B. G.; Kim, W. T. CaPUB1, a Hot Pepper U-Box E3 Ubiquitin Ligase, Confers Enhanced Cold Stress Tolerance and Decreased Drought Stress Tolerance in Transgenic Rice (Oryza Sativa L.). Molecules and Cells 2015, 39, 250–257. [Google Scholar] [CrossRef]
- Lourenço, T.; Sapeta, H.; Figueiredo, D. D.; Rodrigues, M.; Cordeiro, A.; Abreu, I. A.; Saibo, N. J. M.; Oliveira, M. M. Isolation and Characterization of Rice (Oryza Sativa L.) E3-Ubiquitin Ligase OsHOS1 Gene in the Modulation of Cold Stress Response. Plant molecular biology 2013, 83, 351–363. [Google Scholar] [CrossRef]
- Lin, W.; Wu, S.; Wei, M. Ubiquitylome Analysis Reveals the Involvement of Ubiquitination in the Cold Responses of Banana Seedling Leaves. Journal of Proteomics 2023, 288, 104994. [Google Scholar] [CrossRef]
- Hwarari, D.; Guan, Y.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.; Lu, Y.; Chen, J.; Yang, L. ICE-CBF-COR Signaling Cascade and Its Regulation in Plants Responding to Cold Stress. International Journal of Molecular Sciences 2022, 23, 1549. [Google Scholar] [CrossRef]
- Guan, Y.; Delight Hwarari; Harriet Mateko Korboe; Ahmad, B.; Cao, Y.; Movahedi, A.; Yang, L. Low Temperature Stress-Induced Perception and Molecular Signaling Pathways in Plants. Environmental and Experimental Botany 2023, 207, 105190–105190. [Google Scholar] [CrossRef]
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Cold-Stress Responses. Trends in Plant Science 2022. [Google Scholar] [CrossRef]
- Sung Don Lim; Hwang, J.-G.; Chang Gyo Jung; Hwang, S.-G.; Moon, J.-C.; Cheol Seong Jang. Comprehensive Analysis of the Rice RING E3 Ligase Family Reveals Their Functional Diversity in Response to Abiotic Stress. DNA research 2013, 20, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, M.; Jarillo, J. A. Ubiquitination in the Control of Photoperiodic Flowering. Plant Science 2013, 198, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, H.; Estelle, M. Plant Development: Regulation by Protein Degradation. Science 2002, 297, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. F-Box Proteins in Flowering Plants. Chinese Science Bulletin 2002, 47, 1497. [Google Scholar] [CrossRef]
- Shi, Y.; Guo, E.; Wang, L.; Li, T.; Jiang, S.; Xiang, H.; Zhang, T.; Cheng, X.; Zhu, X.; Zhou, L.; Ma, H.; Yang, X. Effects of Chilling at the Booting and Flowering Stages on Rice Phenology and Yield: A Case Study in Northeast China. Journal of Agronomy and Crop Science 2021, 208, 197–208. [Google Scholar] [CrossRef]
- Xu, L.; Rozenn Ménard; Berr, A.; Fuchs, J.; Valérie Cognat; Meyer, D.; Shen, W.-H. The E2 Ubiquitin-Conjugating Enzymes, AtUBC1 and AtUBC2, Play Redundant Roles and Are Involved in Activation of FLC Expression and Repression of Flowering inArabidopsis Thaliana. Plant Journal 2009, 57, 279–288. [Google Scholar] [CrossRef]
- Zhao, H.; Yao, W.; Ouyang, Y.; Yang, W.; Wang, G.; Lian, X.; et al. RiceVarMap: a comprehensive database of rice genomic variations. Nucl. Acid Res. 2015, 43, D1018–D1022. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, H.; Tian, X.; Lin, X.; Han, Y.; Han, Z.; Sha, H.; Liu, J.; Liu, J.; et al. A transposon insertion in the promoter of OsUBC12 enhances cold tolerance during japonica rice germination. Nature Comm. 2024, 15, 2211. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, C.; Sha, H.; Wang, X.; Yu, Y.; Liu, J.; Zhao, G.; Wang, J.; Qiu, G.; Xu, X.; Fang, J. Ubiquitin-Conjugating Enzyme OsUBC11 Affects the Development of Roots via Auxin Pathway. Rice 2023, 16, 9. [Google Scholar] [CrossRef]
- Liu, J.; Nie, B.; Yu, B.; Xu, F.; Zhang, Q.; Wang, Y.; Xu, W. Rice ubiquitin-conjugating enzyme OsUbc13 negatively regulates immunity against pathogens by enhancing the activity of OsSnRK1a. Plant Biotechnol. J. 2023, 21, 1590–1610. [Google Scholar] [CrossRef]
- Wang, Y.; Yue, J.; Yang, N.; Zheng, C.; Zheng, Y.; Wu, X.; Yang, J.; Zhang, L.; Liu, L.; Ning, Y.; Bhadauria, V.; et al. An ERAD-related ubiquitin-conjugating enzyme boosts broad-spectrum disease resistance and yield in rice. Nature Food 2023, 4, 774–787. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, S.; Luo, Y; Li, F.; Tan, J.; Wang, B.; Zhao, Z.; Lin, H.; Zhang, T.; Liu, J., Liu, X., Guo, J.; et al. Rice OsUBR7 modulates plant height by regulating histone H2B monoubiquitination and cell proliferation. Rice OsUBR7 modulates plant height by regulating histone H2B monoubiquitination and cell proliferation. Plant Commun. 2022, 3(6), 100412. [Google Scholar] [CrossRef]
Figure 1.
Single-nucleotide polymorphisms (SNPs) and haplotypes of OsUBC7. (A) 11 SNPs of OsUBC7 in the 354 accessions of the Rice Diversity Panel 1 are aligning with low-temperature seedling survivability (LTSS) scores. Two-week-old seedlings of the 354 accessions were exposed to constant 10°C for 7 days and allowed to recover at warm temperatures for 7 days (28/25°C day/night), after which LTSS was determined. (B) Haplotype analysis of OsUBC7 using RiceVarMap [80] data from a population of 4,402 rice accessions. (C) Haplotype–LTSS correlation analysis based on three major haplotypes of mentioned accessions. p values for Two-Way ANOVA: (**) p < 0.01; (***) p < 0.001.
Figure 1.
Single-nucleotide polymorphisms (SNPs) and haplotypes of OsUBC7. (A) 11 SNPs of OsUBC7 in the 354 accessions of the Rice Diversity Panel 1 are aligning with low-temperature seedling survivability (LTSS) scores. Two-week-old seedlings of the 354 accessions were exposed to constant 10°C for 7 days and allowed to recover at warm temperatures for 7 days (28/25°C day/night), after which LTSS was determined. (B) Haplotype analysis of OsUBC7 using RiceVarMap [80] data from a population of 4,402 rice accessions. (C) Haplotype–LTSS correlation analysis based on three major haplotypes of mentioned accessions. p values for Two-Way ANOVA: (**) p < 0.01; (***) p < 0.001.
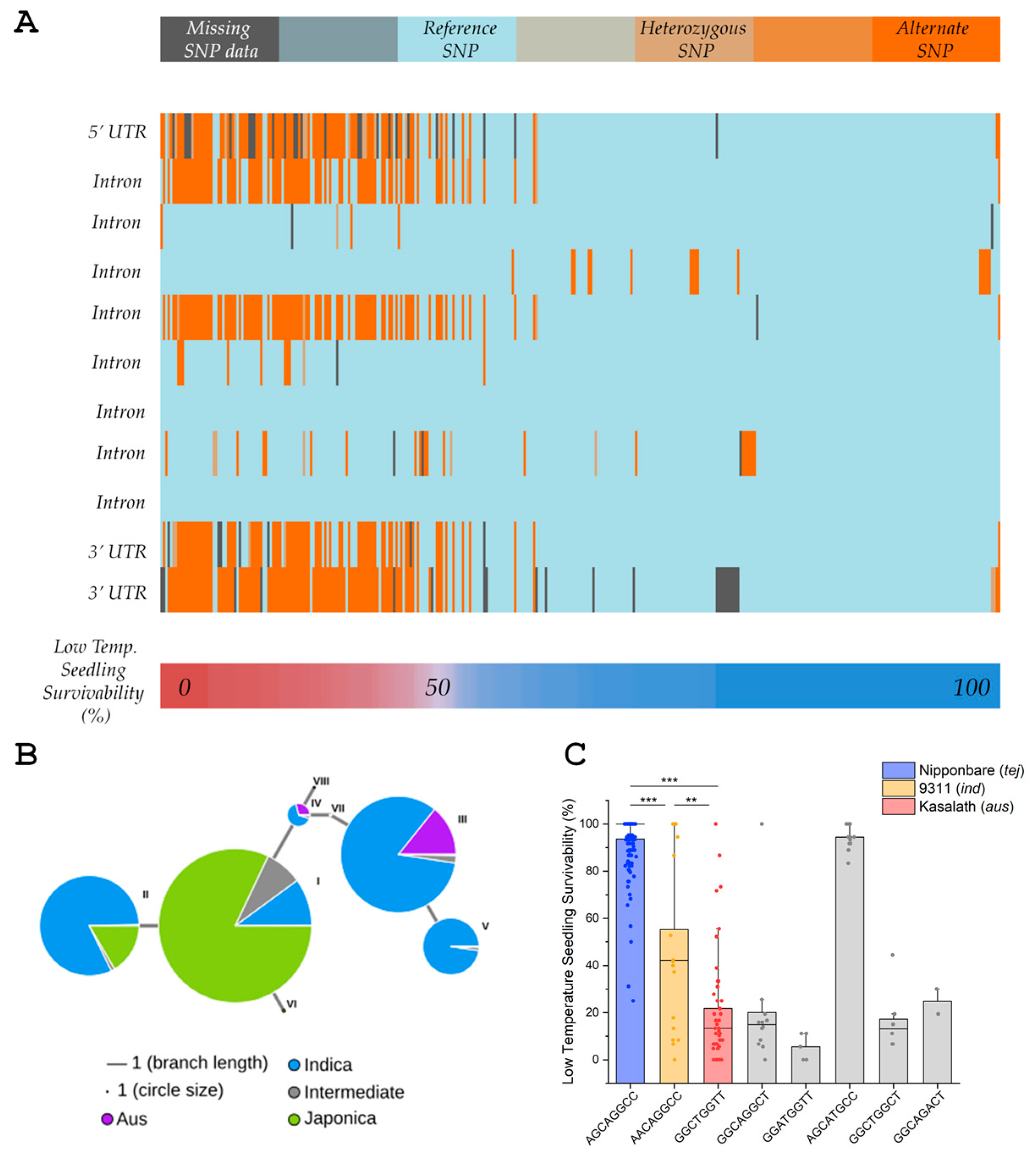
Figure 2.
Cold-stress-related physiological phenotypes of OsUBC7 overexpression (OE) transgenic lines and wild-type (WT) plants. (A) Low temperature seedling survivability (LTSS) of OsUBC7 OE and WT rice plants. (B) LTSS of OsUBC7 OE Arabidopsis, WT, and AtUBC2 (OsUBC7 homolog) knockout (KO) plants exposed to -3°C and recovering over a 7-day period. (C) Growth rate before cold treatment of OsUBC7 OE and WT rice plants. (D) Growth rate after cold treatment of OsUBC7 OE and WT rice plants. (E) Days to third leaf emergence after cold stress. (F) Plant height of 14-day-old OsUBC7 OE and WT seedlings. (α, ß, γ, δ) significant difference in OE or AtUBC2 KO lines compared to their respective WT plants. p values for Two-Way ANOVA: (*) p < 0.05; (**) p < 0.01; (ns) no significance.
Figure 2.
Cold-stress-related physiological phenotypes of OsUBC7 overexpression (OE) transgenic lines and wild-type (WT) plants. (A) Low temperature seedling survivability (LTSS) of OsUBC7 OE and WT rice plants. (B) LTSS of OsUBC7 OE Arabidopsis, WT, and AtUBC2 (OsUBC7 homolog) knockout (KO) plants exposed to -3°C and recovering over a 7-day period. (C) Growth rate before cold treatment of OsUBC7 OE and WT rice plants. (D) Growth rate after cold treatment of OsUBC7 OE and WT rice plants. (E) Days to third leaf emergence after cold stress. (F) Plant height of 14-day-old OsUBC7 OE and WT seedlings. (α, ß, γ, δ) significant difference in OE or AtUBC2 KO lines compared to their respective WT plants. p values for Two-Way ANOVA: (*) p < 0.05; (**) p < 0.01; (ns) no significance.
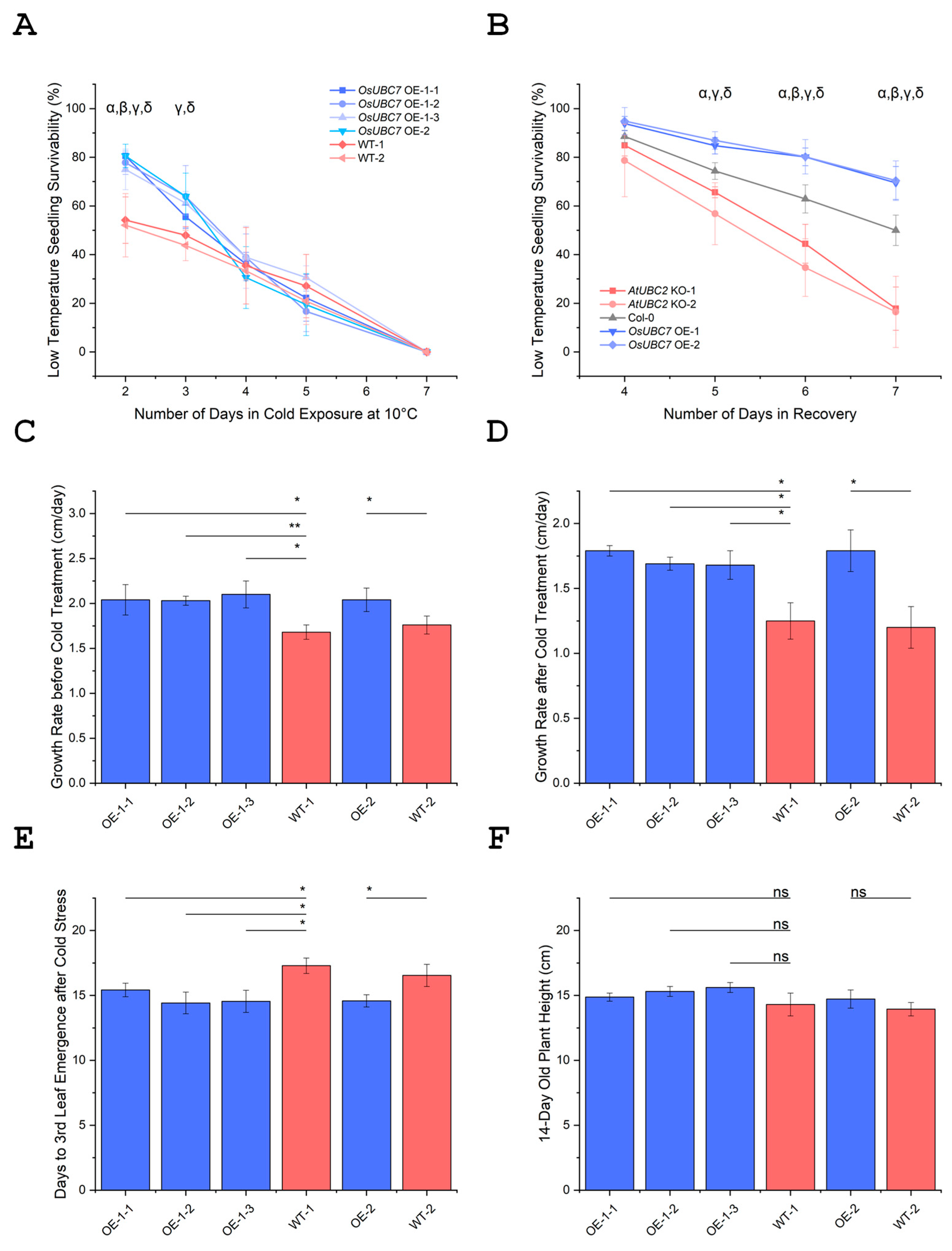
Figure 3.
Effect of OsUBC7 overexpression (OE) in transgenic plants on cold-stress-related metabolites. (A) Soluble sugar content in OsUBC7 OE rice lines and wild-type (WT) plants. (B) Soluble sugar content in OsUBC7 OE Arabidopsis lines, WT Col-0, and AtUBC2 knockout (KO) plants. (C) Mean % electrolyte leakage levels in different tissues of OsUBC7 OE rice lines and WT plants after cold exposure. (D) Malondialdehyde content in different tissues of OsUBC7 OE rice lines and WT plants after cold exposure. (E) Electrolyte leakage in OsUBC7 OE Arabidopsis lines and WT plants. (α, ß, γ, δ) significance detected in OsUBC7 OE transgenic or AtUBC2 KO lines compared to their corresponding WT lines. p < 0.05, Two-Way ANOVA. DAS, day(s) after stress; ns, not significant.
Figure 3.
Effect of OsUBC7 overexpression (OE) in transgenic plants on cold-stress-related metabolites. (A) Soluble sugar content in OsUBC7 OE rice lines and wild-type (WT) plants. (B) Soluble sugar content in OsUBC7 OE Arabidopsis lines, WT Col-0, and AtUBC2 knockout (KO) plants. (C) Mean % electrolyte leakage levels in different tissues of OsUBC7 OE rice lines and WT plants after cold exposure. (D) Malondialdehyde content in different tissues of OsUBC7 OE rice lines and WT plants after cold exposure. (E) Electrolyte leakage in OsUBC7 OE Arabidopsis lines and WT plants. (α, ß, γ, δ) significance detected in OsUBC7 OE transgenic or AtUBC2 KO lines compared to their corresponding WT lines. p < 0.05, Two-Way ANOVA. DAS, day(s) after stress; ns, not significant.
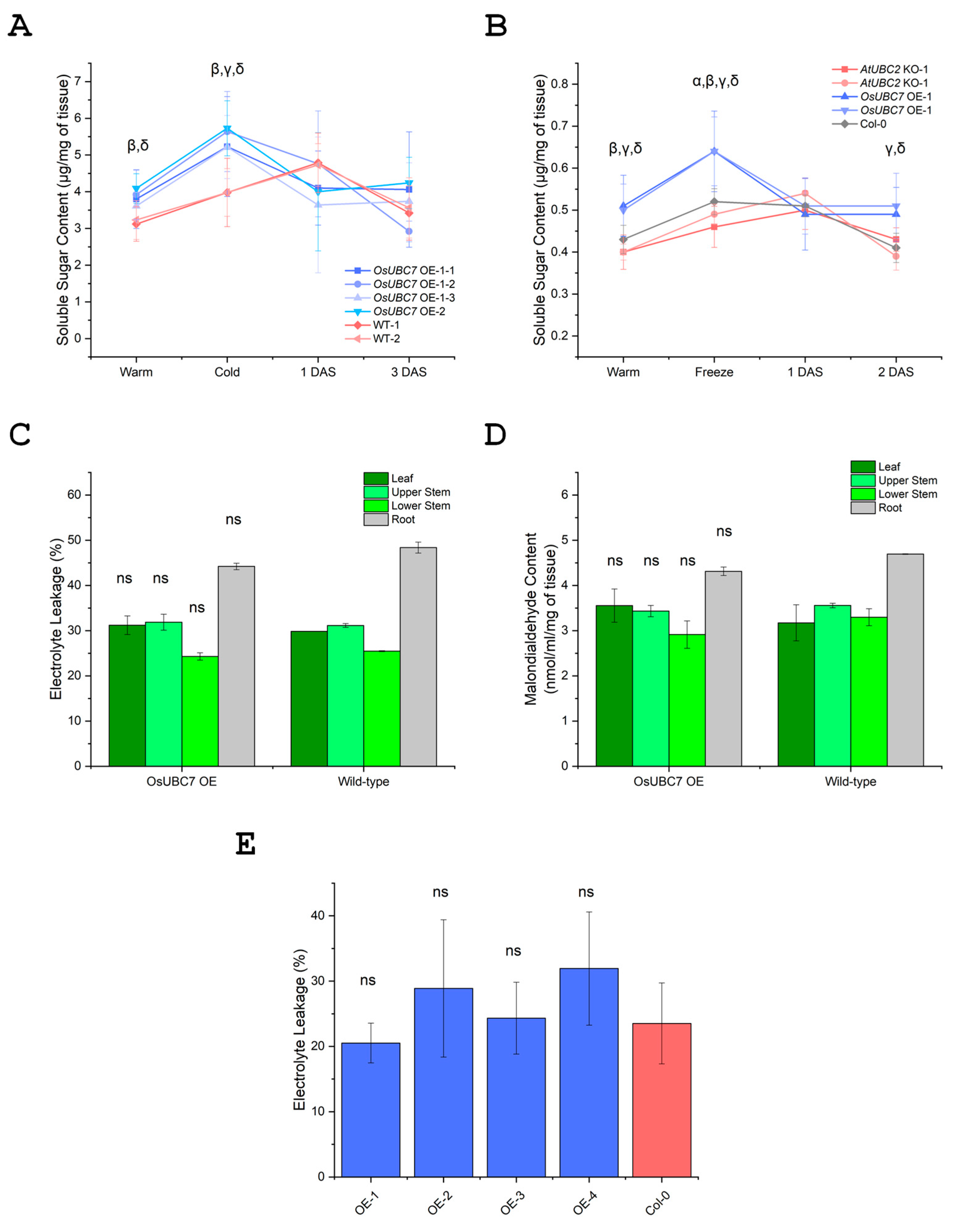
Figure 4.
Effect of OsUBC7 overexpression (OE) on leaf cell dimensions and cell density. (A) Leaf tissue of OsUBC7 OE and segregated wild-type (WT) lines stained with propidium iodide. (B) Cell density of OsUBC7 OE and WT lines. (C) Cell lengths of OsUBC7 OE and WT lines. (D) Cell widths of OsUBC7 OE and WT lines. p-values for Two-Way ANOVA: (*) < 0.05; (**) < 0.01; (ns) no significance.
Figure 4.
Effect of OsUBC7 overexpression (OE) on leaf cell dimensions and cell density. (A) Leaf tissue of OsUBC7 OE and segregated wild-type (WT) lines stained with propidium iodide. (B) Cell density of OsUBC7 OE and WT lines. (C) Cell lengths of OsUBC7 OE and WT lines. (D) Cell widths of OsUBC7 OE and WT lines. p-values for Two-Way ANOVA: (*) < 0.05; (**) < 0.01; (ns) no significance.
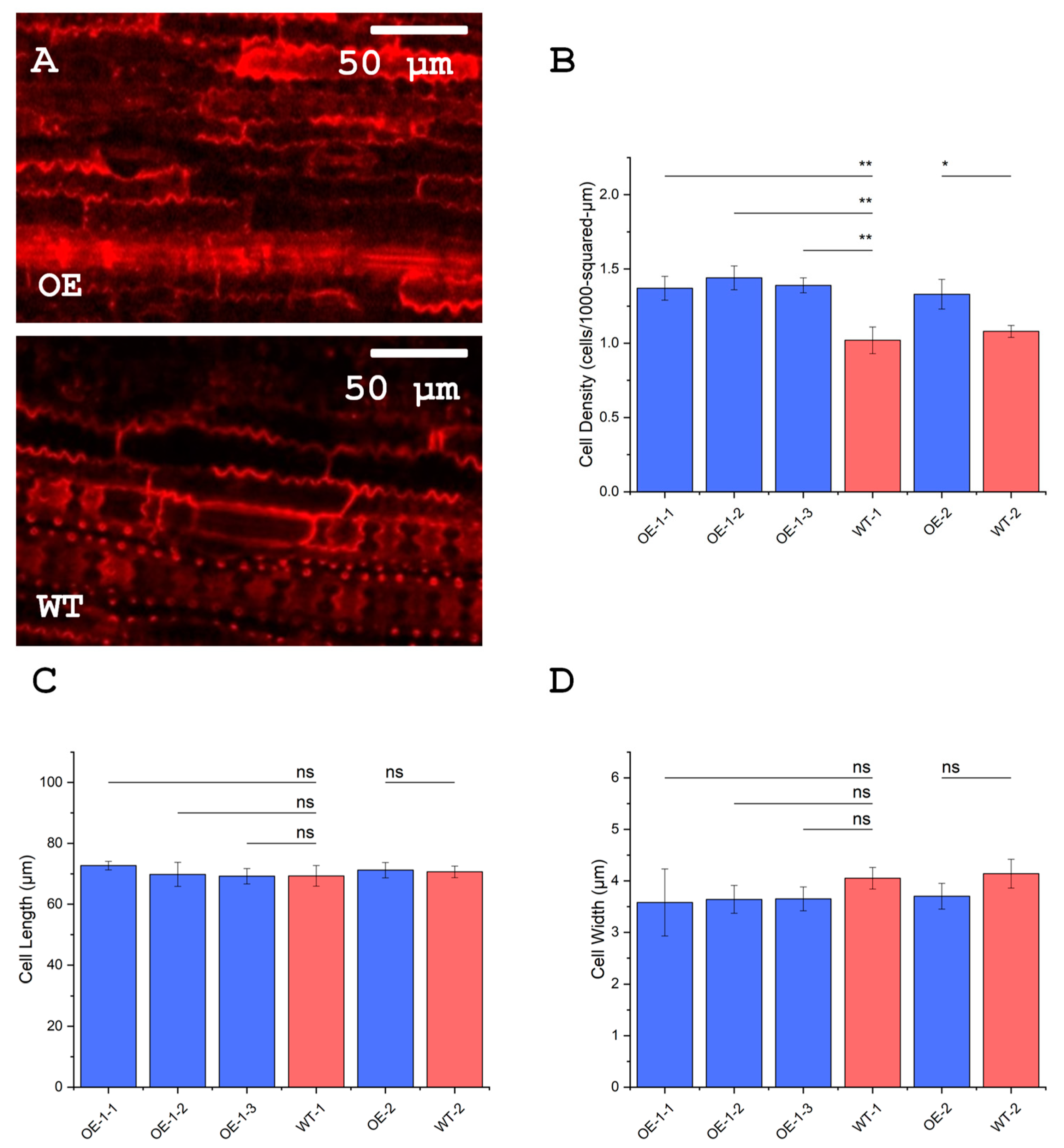
Figure 5.
Architecture of the first flag leaf and chlorophyll content of OsUBC7 overexpressing (OE) and wild-type (WT) lines. (A) Half-length of the first flag leaf, from the leaf tip to the middle. (B) Width of the first flag leaf. (C) Dry weight of the first flag leaf. (D) Total chlorophyll A content extracted from the entire flag leaf. (E) Total chlorophyll B content extracted from the entire flag leaf. p-values for Two-Way ANOVA: (*) < 0.05; (**) < 0.01; (ns) no significance.
Figure 5.
Architecture of the first flag leaf and chlorophyll content of OsUBC7 overexpressing (OE) and wild-type (WT) lines. (A) Half-length of the first flag leaf, from the leaf tip to the middle. (B) Width of the first flag leaf. (C) Dry weight of the first flag leaf. (D) Total chlorophyll A content extracted from the entire flag leaf. (E) Total chlorophyll B content extracted from the entire flag leaf. p-values for Two-Way ANOVA: (*) < 0.05; (**) < 0.01; (ns) no significance.
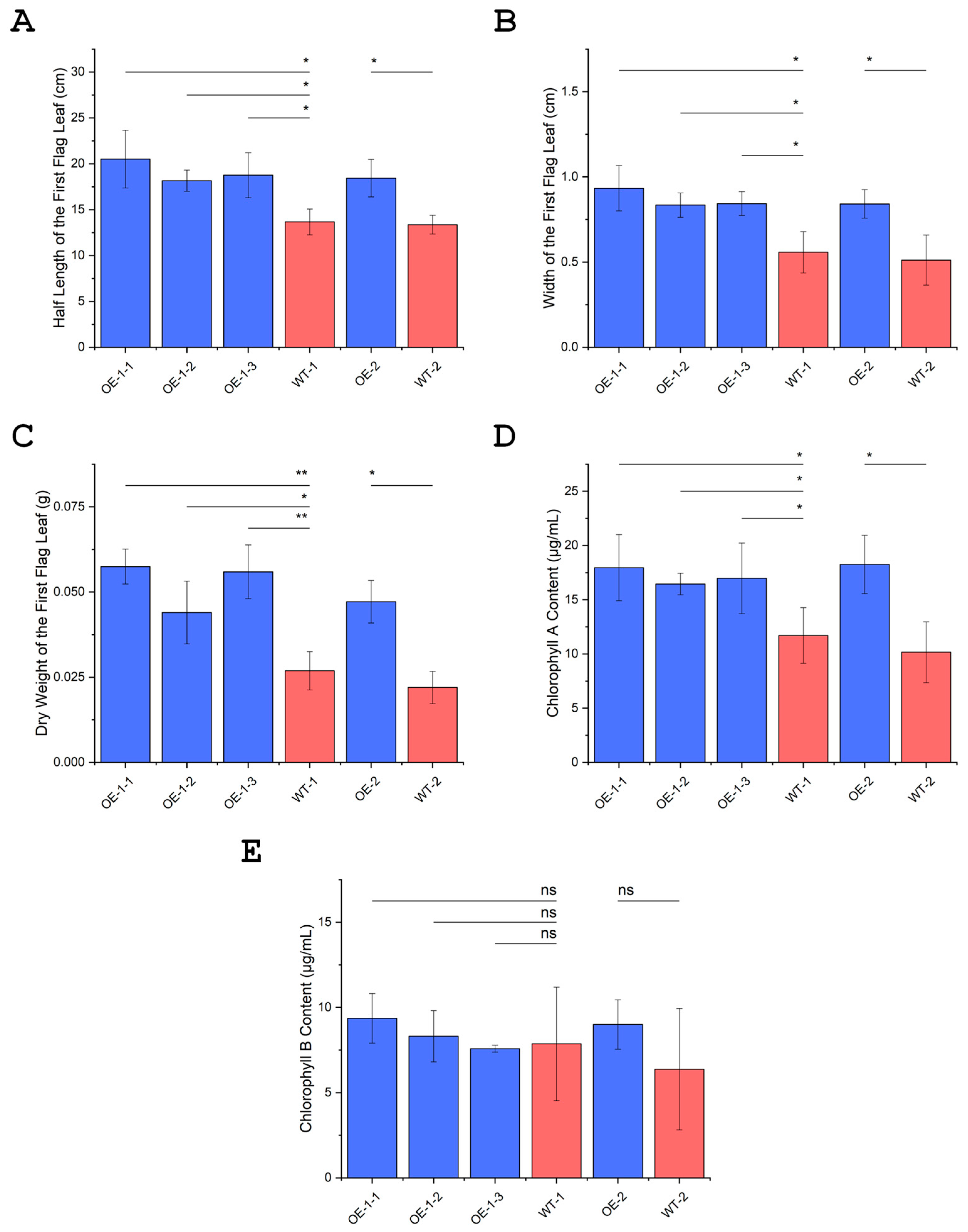
Figure 6.
Yeast two-hybrid assay showing interaction between Gal4-DNA binding domain (BD)-OsUBC7 and Gal4-Activation domain (AD)-E3 ligase fusions of OsRFPH2-12 (Left) and OsHUB1/HUB2 (Right; RING finger domain only). Positive interactions are shown by robust colony growth on quadruple dropout (QDO) selective plates, as seen for OsUBC7::RFPH2-12 and OsUBC7::OsHUB1/HUB2 in rows labeled “P”. A positive control showing interaction between a Gal4-BD-murine P53 fusion (bait) and a Gal4-AD-SV40 large T-antigen fusion (prey) is seen in rows labeled “+”. Negative controls of yeast transformed with the OsUBC7 bait and an empty prey vector (and vice versa) have no growth on QDO selective plates, as shown in rows labelled “–“. Bait and prey plasmids have robust growth on double dropout (DDO) selective plates.
Figure 6.
Yeast two-hybrid assay showing interaction between Gal4-DNA binding domain (BD)-OsUBC7 and Gal4-Activation domain (AD)-E3 ligase fusions of OsRFPH2-12 (Left) and OsHUB1/HUB2 (Right; RING finger domain only). Positive interactions are shown by robust colony growth on quadruple dropout (QDO) selective plates, as seen for OsUBC7::RFPH2-12 and OsUBC7::OsHUB1/HUB2 in rows labeled “P”. A positive control showing interaction between a Gal4-BD-murine P53 fusion (bait) and a Gal4-AD-SV40 large T-antigen fusion (prey) is seen in rows labeled “+”. Negative controls of yeast transformed with the OsUBC7 bait and an empty prey vector (and vice versa) have no growth on QDO selective plates, as shown in rows labelled “–“. Bait and prey plasmids have robust growth on double dropout (DDO) selective plates.
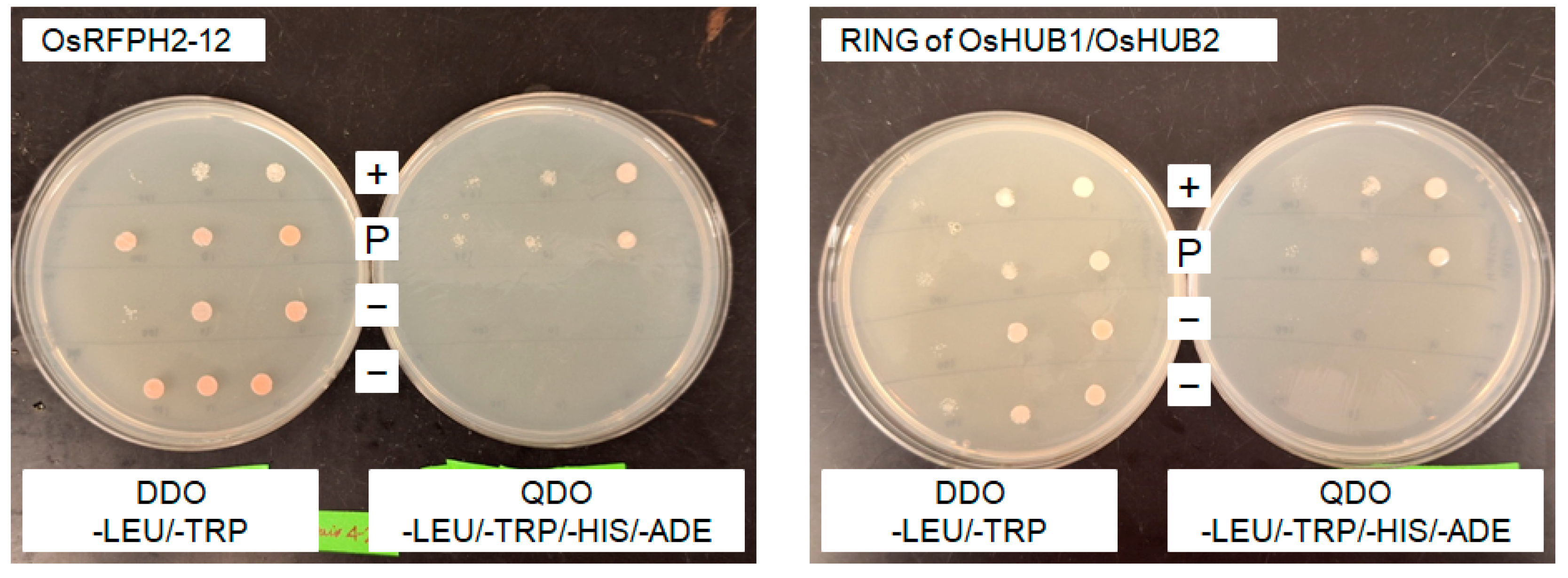

Table 1.
mRNA abundance of OsUBC7 in aus Kasalath and temperate japonica Krasnodarskij 3352 (Krasnodars.) in 2-week-old seedlings under warm condition (0 hr) and after different periods of cold temperature exposure (10°C for Kasalath; 4°C for Krasnodars.). Four house-keeping genes were used to normalize mRNA abundance: 18S ribosomal RNA, OsACT1, OsUBC32, and OsUBQ5.
Table 1.
mRNA abundance of OsUBC7 in aus Kasalath and temperate japonica Krasnodarskij 3352 (Krasnodars.) in 2-week-old seedlings under warm condition (0 hr) and after different periods of cold temperature exposure (10°C for Kasalath; 4°C for Krasnodars.). Four house-keeping genes were used to normalize mRNA abundance: 18S ribosomal RNA, OsACT1, OsUBC32, and OsUBQ5.
| House-Keeping Gene | Hrs of Exposure | Normalized mRNA Abundance | Log2 Fold-Change | ||
| Kasalath | Krasnodars. | Kasalath | Krasnodars. | ||
| 18S ribosomal RNA | 0 | 2.09E-03 ± 1.20E-04 | 1.63E-03 ± 2.90E-04 | 0 | 0 |
| 12 | 3.13E-03 ± 4.06E-03 | 2.30E-03 ± 6.20E-04 | 0.585 | 0.496 | |
| 24 | 3.12E-03 ± 2.80E-04 | 3.86E-03 ± 1.57E-03 | 0.575 | 1.245 | |
| 36 | 4.11E-03 ± 1.25E-03 | 1.10E-02 ± 2.55E-03 | 0.978 | 2.751 | |
| 48 | 4.35E-03 ± 1.26E-03 | 7.51E-03 ± 2.56E-03 | 1.057 | 2.205 | |
| OsACT1 | 0 | 1.13E-01 ± 8.10E-02 | 7.70E-02 ± 1.30E-02 | 0 | 0 |
| 12 | 1.54E-01 ± 4.90E-02 | 1.58E-01 ± 3.60E-02 | 0.444 | 1.029 | |
| 24 | 2.16E-01 ± 9.20E-02 | 3.81E-01 ± 1.92E-01 | 0.941 | 2.305 | |
| 36 | 5.46E-01 ± 1.31E-01 | 6.05E-01 ± 4.67E-01 | 2.275 | 2.973 | |
| 48 | 6.26E-01 ± 2.92E-01 | 7.89E-01 ± 1.38E-01 | 2.470 | 3.356 | |
| OsUBC32 | 0 | 5.15E-02 ± 1.10E-02 | 3.00E-02 ± 1.07E-02 | 0 | 0 |
| 12 | 6.87E-02 ± 2.26E-02 | 5.20E-02 ± 1.48E-02 | 0.411 | 0.791 | |
| 24 | 1.90E-01 ± 3.45E-02 | 2.48E-01 ± 8.90E-02 | 1.880 | 3.046 | |
| 36 | 2.41E-01 ± 3.00E-02 | 3.04E-01 ± 1.50E-01 | 2.227 | 3.338 | |
| 48 | 2.18E-01 ± 9.81E-02 | 3.03E-01 ± 1.48E-01 | 2.084 | 3.333 | |
| OsUBQ5 | 0 | 5.22E-02 ± 1.97E-02 | 4.23E-02 ± 2.55E-02 | 0 | 0 |
| 12 | 2.22E-01 ± 8.00E-03 | 1.40E-01 ± 3.95E-02 | 2.091 | 1.731 | |
| 24 | 2.76E-01 ± 1.11E-01 | 3.28E-01 ± 1.09E-01 | 2.403 | 2.954 | |
| 36 | 2.76E-01 ± 6.04E-02 | 4.96E-01 ± 6.11E-02 | 2.403 | 3.551 | |
| 48 | 3.55E-01 ± 1.31E-01 | 1.07E+00 ± 2.18E-01 | 2.766 | 4.659 | |
Table 2.
Expression levels of OsUBC7, OsUBC8, OsUBC9 in Indica Kasalath (aus) and Japonica Krasnodarskij 3352 (Krasnodars.; temperate japonica) in 2-week-old seedlings and flag leaves at the flowering stage. Two house-keeping genes were used: 18S ribosomal RNA, OsACT1.
Table 2.
Expression levels of OsUBC7, OsUBC8, OsUBC9 in Indica Kasalath (aus) and Japonica Krasnodarskij 3352 (Krasnodars.; temperate japonica) in 2-week-old seedlings and flag leaves at the flowering stage. Two house-keeping genes were used: 18S ribosomal RNA, OsACT1.
| House-Keeping Genes | Target Gene and Development Stage | Relative mRNA Abundance | Log2 Fold-Change | ||
| Kasalath | Krasnodars. | Kasalath | Krasnodars. | ||
| 18S ribosomal RNA | OsUBC7 | ||||
| 2-week-old | 2.09E-03 ± 1.20E-04 | 1.63E-03 ± 2.90E-04 | 0 | 0 | |
| Flowering | 1.78E-02 ± 9.07E-03 | 1.71E-02 ± 4.60E-03 | 3.093 | 3.392 | |
| OsACT1 | 2-week-old | 1.13E-01 ± 8.10E-02 | 7.71E-02 ± 1.34E-02 | 0 | 0 |
| Flowering | 1.78E+00 ± 3.88E-01 | 1.74E+00 ± 4.30E-01 | 3.379 | 4.493 | |
| 18S ribosomal RNA | OsUBC8 | ||||
| 2-week-old | 1.22E-02 ± 4.81E-03 | 1.36E-02 ± 5.92E-03 | 0 | 0 | |
| Flowering | 5.55E-03 ± 1.81E-03 | 4.01E-03 ± 8.80E-04 | -1.120 | -1.737 | |
| OsACT1 | 2-week-old | 1.41E+00 ± 3.49E-01 | 1.20E+00 ± 2.59E-01 | 0 | 0 |
| Flowering | 6.58E-01 ± 1.66E-01 | 6.23E-01 ± 1.00E-01 | -1.089 | -0.943 | |
| 18S ribosomal RNA | OsUBC9 | ||||
| 2-week-old | 5.49E-02 ± 2.12E-02 | 5.44E-02 ± 1.21E-02 | 0 | 0 | |
| Flowering | 1.00E-01 ± 1.96E-02 | 9.04E-02 ± 2.59E-02 | 0.872 | 0.731 | |
| OsACT1 | 2-week-old | 1.69E+01 ± 9.10E+00 | 1.13E+01 ± 1.07E+00 | 0 | 0 |
| Flowering | 2.23E+01 ± 8.68E+00 | 1.94E+01 ± 1.89E+00 | 0.401 | 0.774 | |
Table 3.
Yield-related trait phenotypes of OsUBC7 overexpression (OE) and segregated wild-type (WT) lines in the Kasalath (aus) accession.
Table 3.
Yield-related trait phenotypes of OsUBC7 overexpression (OE) and segregated wild-type (WT) lines in the Kasalath (aus) accession.
| Yield-Related Phenotype | Line | Data from Three Generations | p-value(Two-Way ANOVA) |
| Total Seeds per Plant | OE-1 | 80.3 ± 33.7 | 0.124 |
| WT-1 | 30.7 ± 3.3 | ||
| OE-2 | 66.9 ± 16.5 | 0.072 | |
| WT-2 | 36.3 ± 14.1 | ||
| Number of Seeds per Panicle | OE-1 | 34.4 ± 12.7 | 0.278 |
| WT-1 | 23.5 ± 6.9 | ||
| OE-2 | 32.2 ± 17.5 | 0.274 | |
| WT-2 | 17.0 ± 9.2 | ||
| Fertility (%) | OE-1 | 86.4 ± 1.0 | 0.080 |
| WT-1 | 77.0 ± 4.1 | ||
| OE-2 | 83.7 ± 5.1 | 0.163 | |
| WT-2 | 62.1 ± 17.8 | ||
| Flowering Time (No. of Days to heading) | OE-1-1 | 101.2 ± 1.9 | 0.684 |
| OE-1-2 | 100.3 ± 2.8 | 0.933 | |
| OE-1-3 | 102.2 ± 5.4 | 0.648 | |
| OE-2 | 100.3 ± 1.9 | 0.375 | |
| WT-1 | 100.5 ± 2.2 | ||
| WT-2 | 98.5 ± 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



