
Submitted:
20 November 2024
Posted:
22 November 2024
You are already at the latest version
Abstract
Pediococcus pentosaceus is a lactic acid bacterium used inter alia for fermentation of milk, meat, vegetables, fruits, and even beer. The resulting food products are tasty and safe. Furthermore, health-promoting effects such as anti-inflammation, anti-cancer, and detoxification have been attributed. Beside refining foods during the fermentation process, P. pentosaceus might be added to foods as a protective culture to improve food safety and extension of shelf-life, leaving the organoleptic properties of the food untouched. We have addressed the search for novel protective P. pediococcus strains by investigation of 38 isolates from milk samples and teat canal biofilms from dairy farms located in the vicinity of Muenster, Germany. P. pentosaceus strains were unequivocally identified by DNA sequencing of the gene encoding 16S rRNA. Binary matrices obtained from random amplification of polymorphic DNA experiments showed that all isolates differed and thus represent subspecies. The antibacterial profiles of 13 selected isolates showed that seven food pathogens and food spoilage bacteria could efficiently combat, although to various extends, gram-negative bacteria such as Pseudomonas aeruginosa or Salmonella enterica, and gram-positive bacteria such as Staphylococci and Listeria monocytogenes. Interestingly, acid production was dependent on the presence of the challenged pathogen but did not significantly correlate with the extend of inhibition. The genomes of three top-ranked isolates were sequenced. We found that the genome size and the harbored plasmids differed significantly, confirming the subtle genomic plasticity. One to two genes encoding bacteriocins could be detected in each of the three genomes. Our approach for protective culture screening demonstrates that it is worth to examine many isolates from a specific species to select the best performing strain for application in food and care products.
Keywords:
Pediococcus pentosaceus
; protective cultures
; antibacterial
; genome
; lactic acid bacteria
; food safety
; bacteriocins
1. Introduction
A whole variety of microorganisms is used since thousands of years to produce fermented beverages and food, which are very tasty and resilient to food spoilage [1,2,3,4]. In the early 20th century, Illia Iljitsch Metschnikoff postulated that the yogurt-producing lactic acid bacteria (LAB) Lactobacillus bulgaricus and Streptococcus thermophilus exert health-promoting effects and help humans live longer by acting as intestinal residents [5]. Since then, LAB and bifidobacteria have been thoroughly investigated regarding their biology and their probiotic efficacy to uncover and prove beneficial activities [4,5,6]. The result is a huge market that offers to the customer probiotic foods, nutraceuticals, and care products for human and animal health[4,7]. Furthermore, traditional spontaneous (wild) fermentation of food has undergone a renaissance in the past years as a food trend that has been, among other things, strongly acclaimed by chefs at Michelin-starred restaurants to present dishes with novel flavor profiles [8,9].
From a taxonomic point of view, LAB are grouped in the order Lactobacillales comprising the families Aerococcaceae, Carnobacteriaceae, Enterococcaceae, Lactobacillaceae, and Streptococcaceae [10]. The latter two are the most relevant families, of which Lactobacillaceae stand out [11]. They were recently re-classified based on a molecular phylogenetic approach at whole genome level. The authors included additionally the Leuconocaceae family giving a total of 261 described species that comprise 25 genera[11].
The ten species of the genus Pediococcus receive increasingly attention as food fermenting and protecting cultures [12,13]. One most studied member is Pediococcus pentosaceus, which became originally famous for spoiling beer and at the same time is appreciated for its buttery and sour taste in special beer types [14,15]. Strains of P. pentosaceus have become attractive for the fermentation of vegetables, sausages, vegetables, fruits, and wine [16]. Numerous health-promoting effects have been associated including microphage stimulation, anti-inflammation, anti-cancer, antioxidant, detoxification, and cholesterol-lowering [16]. As other LAB, Pediococci contribute to food safety as they combat pathogenic bacteria by producing bacteriocins or other antibacterial compounds such as lactic acid, small peptides, hydrogen peroxide, or diacetyl [12,17,18].
Here, we present the isolation of P. pentosaceus strains from one specific natural environment, the udder of milk cattle to study their potential application as protective culture in foods. We show that P. pentosaceus isolates exhibit a high biodiversity at the subspecies level. We show that each isolate exerts antibacterial activities to various extends against representative pathogens. We provide genomic data from the best performing isolates to provide a molecular view into the genome plasticity. The implication of our approach for the discovery of protective cultures in foods to enhance food safety is discussed.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
P. pentosaceus were cultivated on modified DeMan-Rogosa Sharp agar plates (MRS) [19]. If appropriate, MRS plates were supplemented with cysteine (mMRS) to promote growth and with bromophenol blue (mMRS-bpb) [20] to better distinguish colony morphologies by blueish coloration. Strains were freshly propagated in MRS broth for 48 hours at 30°C. Seven bacterial species were employed as indicators and surrogates for foodborne pathogens and spoilage organisms. These strains, except Listeria monocytogenes, were cultivated in Lysogeny broth (LB) after Lennox at 37°C for 24 hours. Brain Heart Infusion (BHI) Bouillon was utilized for the cultivation of L. monocytogenes.

Table 1.
List of strains.
| species | origin | comment |
|---|---|---|
| Bacillus subtilis | laboratory strain collection | surrogate food spoilage |
| Citrobacter koseri | laboratory strain collection | fish-borne pathogen |
| Listeria monocytogenes | DSM 20600 | food pathogen |
| Pseudomonas aeruginosa | ATCC 15442 | food spoilage & pathogen |
| Salmonella enterica | LT-2 | food pathogen |
| Staphylococcus haemolyticus | laboratory strain collection | surrogate food pathogen |
| Staphylococcus warneri | laboratory strain collection | surrogate food pathogen |
2.2. Species Identification Through 16S rRNA Gene Sequencing
Genomic DNA extraction was performed by resuspending a single colony in 50 µl of 0.05 M NaOH. The mixture was incubated at 100°C for 1 minute. The lysate was precipitated by centrifugation. The supernatant with the genomic DNA was 1:10 diluted in sterile DNAse-free demineralized water. Polymerase chain reactions (PCR) were carried out in a volume of 25 µl, consisting of 12.5 µl oneTaq Polymerase Hot Start mastermix (New England Bio Labs, England), 0.5 µl (10 pmol/μl) F08 primer (5’-AGAGTTTGATCATGGCTCAG-3’), 0.5 µl (10 pmol/μl) 535R primer (5’-TATTACCGCGGCTGCTGGCA-3’) , 9 µl DNase and nuclease-free water, and 2.5 µl (0.1 to 10 ng) DNA template. The resulting amplicon encompassed bases 8 to 535 of the gene encoding 16s rDNA representing the variable regions V1-V3 of the 16S rDNA gene. PCRs were carried out with an initial denaturation step at 95°C for 10 minutes, followed by 40 cycles at 94°C for 30 seconds, 60°C for 30 seconds, 72°C for 30 seconds. PCR amplicons were sequenced at Microsynth (Switzerland). The Basic Logical Alignment Tool (BLAST) service provided by the National Center for Biotechnology Information (NCBI, USA) was used for species identification.
2.3. Random-Amplified-Polymorphic-DNA Analysis (RAPD)
RAPD-PCR fingerprinting was carried out by applying the M13 minisatellite core primer (5'-GAGGGTGGCGGTTCT-3') [23]. The PCR reaction mixture contained equal amounts of 10 ng genomic DNA, 2 µl primer (10 pmol) and 10 µl of a 2x concentrated OneTaq polymerase master mix (New England Bio Labs, England). The thermal cycling protocol involved an initial denaturation step at 94°C for 7 minutes, followed by 40 cycles of denaturation at 94°C for 30 seconds, annealing at 42°C for 30 seconds, and extension at 68°C for 4 minutes. A final extension cycle was performed at 68°C for 10 minutes (modified after Sirichoat et al. [24]). PCR products were subjected to electrophoresis on a 2% agarose gel (AppliChem, Germany).
2.4. Hydrogen Peroxide Production
Hydrogen peroxide production was conducted following the method described by Tomás et al. [25]. Fresh cultures of each isolate were plated on mMRS agar plates supplemented with 1 mM 3,3',5,5'-Tetramethylbenzidin (TMB) and 2 U/ml of type II horseradish peroxidase (Carl Roth, Germany). Agar plates were incubated anaerobically for 48 hours at 30°C. Subsequently, the Petri dish lid was removed, exposing the cultures to fresh air for 10 minutes. Hydrogen production was visible when the yellowisch colonies turned black. Data were reproduced in triplicate.
2.5. In Vivo Detection of Antimicrobial Activity
Antimicrobial activities of P. pentosaceus isolates against foodborne pathogens and food spoilage organisms were evaluated with lawn-on-spot assays modified as described [26,27]. Isolates were inoculated from a liquid overnight culture by positioning 2 µl into the center of an MRS agar plate. Agar plates were incubated under anaerobic conditions at 30°C for 48 hours. The freshly grown colonies were overlaid with 10 ml of LB-soft agar (8%) or BHI-soft agar (8%) in the case of L. monocytogenes. Before pouring, indicator strains were applied by adding 0,2 ml of a cell suspension of 1,0x107 CFU/ml. Following a 20- to 30-hour co-cultivation at 37°C, the diameter of the zone of growth inhibition, from which the colony diameter was subtracted, was measured in millimeter. Acid secretion of the P. pediococcus isolates was determined with pH stripes. In this way, it was possible to measure changes of pH by comparing the pH in the inhibition zone with the pH of the indicator strain grown on the same media in the absence of a P. pediococcus isolate.
2.5. Whole Genome Sequencing
Genomic DNA was extracted using the Wizard HMW DNA Extraction Kit (Promega, USA) and quantified using a DeNovix QFC Fluorometer (DeNovix, United States). Sequencing libraries were prepared with the Rapid Sequencing Kit (SQK-RAD004) (ONT, GB). After that, samples of 30 µl were subjected to a Flongle Flow Cell R9.4.1 (FLO-FLG001) (ONT, GB) on a MinION Mk1B device (ONT, GB) with MinKNOW 22.08.9 [28].
2.6. Data Analyses and Statistics
Data analyses comprised subspecies identification, antimicrobial activity in vivo, and the relationship between antimicrobial compounds and acid secretion. To do so, the statistical and computing package R (v4.2.1) and RStudio (v2023.03.0+386) with packages tidyverse (v2.0.0), readxl (v1.4.2), ggplot2 (v3.4.2), vegan (2.6-4) and ggdendroplot (v0.1.0) [29,30,31,32,33,34]. Subspecies identification was based on the analysis of the data derived from RAPD-PCR DNA band patterns. Therefore, the unweighted pair group method with arithmetic mean (UPGMA) was used by using the hclust algorithm together with the Dice dissimilarity coefficient matrix [35,36] from the R base package. Phylogenetic trees were generated using a threshold of ≤ 5% dissimilarity to distinguish clonal isolates [35,37,38,39,40]. Strains were analyzed for in vivo antimicrobial activity via heatmapping and cluster analysis, by normalizing inhibition zone diameters across strains by z-score calculation. Hierarchical clustering of strains and indicators was performed using Euclidean distances and complete linkage in the hclust function, visualized through heatmap and dendrograms in ggplot2 and ggdendroplot. The above-mentioned statistical methods were also used to evaluate the data obtained from acidification experiments. Pearson correlation and scatterplot visualisation performed on the raw replicate values assessed the contributions of antimicrobial compounds and acid secretion at a 95% confidence level.
2.7. Bioinformatics Pipeline and Genome Assembly
The bioinformatics pipeline, managed by Snakemake [41], began with high-accuracy base-calling of raw ONT sequencing data via Guppy (v6.3.8) [42]. Porechop was used for adapter trimming [43], and Filtlong filtered out reads shorter than 1 kbp and the lowest 10% by quality [44]. Trycycler was then used for assembly, combining Flye, Raven, and Miniasm+MiniPolish to generate a consensus genome [45,46,47,48,49], with the help of MASH and MUSCLE [50,51,52], and final polishing by Medaka [53].
2.8. Genome Analyses
Gene annotations, chromosomal organization und circular maps were obtained using the bioinformatic tools Bakta, Prokka, and FastANI as implemented into the Proksee server [54,55,56,57]. A Venn-Diagram showing the core genome, the pan genomes and singleton genes was computed with EDGAR 3.0 on the basis of the gene annotation list created with Bakta [54,58]. Genes encoding bacteriocins were screened by uploading fasta files of chromosomes and plasmid DNA sequences into the BAGEL4 server [59].
3. Results
3.1. Isolation of Pediococcus Pentosaceus Strains from the Cattle Udder
In the course of a screening for novel food protective strains, we isolated several hundred microbial strains from the udder of milk cows from 15 farms located in Münsterland, Germany. Among those, we found 38 strains of P. pentosaceus, as demonstrated by complete DNA sequence identity of a 483 base pair fragment of the 16S rRNA-encoding gene (data not shown). One isolate, that produced the antibacterial compound hydrogen peroxide (data not shown), was excluded since such strains can alter the taste of food products by oxidation processes, an undesired characteristic for the use of protective cultures in foods [60,61].
3.2. Identification of Subspecies
We then raised the question about the genetic diversity of the obtained strains. Therefore, chromosomal DNA samples of all 37 P. pentosaceus isolates were subjected to RAPD-PCR. Representative band patterns of 21 isolates from farms 4 and 13 are shown in Figure 1.
A matrix of the fragment size distribution was generated and used for UPGMA cluster analyses to obtain phylogenetic trees of isolates within each farm (Figure 2). Across the four dairy farms, from which P. pentosaceus strains were obtained, the relative similarity ranged from 20% to 85%.
While the spatial proximity of isolates within the phylogenetic tree suggested a higher degree of genetic similarity and potential relatedness among individuals, none of the isolates within a farm exhibited a phylogenetic dissimilarity of less than 5%, the threshold to discriminate subspecies. Hence, all isolates were classified as a distinct subspecies within one farm.
3.3. Antimicrobial Profiles
To uncover the most important prerequisite for a protective culture, the antimicrobial potential, we took thirteen diverse isolates for further investigations. They were tested against a set of indicator strains comprising food pathogenic and spoilage bacteria and representative surrogates. A heatmap that was derived from inhibition zone sizes of conducted spot-on-lawn assays is presented in Figure 3. The data were normalized to ensure comparability. Based on the calculated z-values, indicator strains could be clustered according to their antibacterial profiles. Most susceptible were Pseudomonas aeruginosa and Salmonella enterica situated in the first subcluster on the left with inhibition zones of 34 and 56 millimeter.
A second cluster of indicators comprised the gram-negative, enteric bacterium Citrobacter koseri and the two gram-positive staphylococci, ranging from inhibition 17 to 24 millimeter. While C. koseri was generally more sensitive to P. pentosaceus isolates, three of the thirteen isolates were more effective against the Staphylococcus strains. The antimicrobial effect of the third cluster, consisting of the gram-positives L. monocytogenes and B. subtilis, was below average. It is noteworthy that L. monocytogenes was generally susceptible to most isolates, especially to isolate 13.7.8A-4. The zones of inhibition for L. monocytogenes ranged from eight to 17 millimeter. The only indicator that showed very weak or in three cases even zero antibacterial activity was the food spoilage surrogate B. subtilis.
3.4. Acidification
After the antimicrobial profiles were uncovered, it was crucial to know which isolates have a low capacity to produce acids, in order to avoid unwanted changes in the taste of food products. To monitor this, we examined the extent of acidification of P. pentosaceus isolates exposed to all indicator strains (Figure 4).
Acidification of the medium in response to the indicator strains was considerably variable. Notably, the presence of S. enterica induced the strongest acidification by an average decrease in pH from 0.79 (± 0.1). By contrast, the presence of C. koseri yielded the mildest acidification with a pH drop ranging around 0.1 (± 0.08) units. All isolates behaved quite similar in acidification with an average of pH reduction of 0.26 (±0.04) units.
To elucidate the influence of acid secretion on the measured antimicrobial activity, a Pearson correlation analysis was performed, and non-linear correlations were visualized through scatterplot analysis. Figure 5A shows that acid secretion had a slight but significant effect on the measured antimicrobial activity for Listeria monocytogenes, with a correlation coefficient of -0.33 and a p-value of 0.042. No other indicator organism demonstrated sensitivity solely through acid secretion.
Out of the thirteen isolates, five revealed a significant moderate to strong correlation between acid secretion and measured antimicrobial activity across all indicators. These were the isolates 02.3 3A-1, 13.7 10A-6, 13.7 17-3, 13.7 9A-11, and 13.7 8A-4 (Figure 5B).
Assuming that a protective culture should effectively inhibit a broad spectrum of food pathogens and food spoilage microorganisms without affecting the flavor and texture of the food products, isolates were ranked as shown in Table 2. The criteria included above-average inhibition of the number of indicator species combined with below-average acidification.
The best performing strain was isolate 13.7 13A-5, which inhibited five out of seven indicator bacteria above average, followed by isolates that inhibited three indicator species above average. Any isolate could be selected from this group, considering the specific properties of the food in question and the most critical species to be controlled.
3.6. Genome Analyses
Since, and that was somehow surprising, all isolates were according to our RAPD analyses greater than 5% dissimilar, they were classified as subspecies [39]. We sequenced the genomes of the best three performing candidates to proof and substantiate this at the molecular level (Table 2). The chromosomes of the isolates differed already in size, namely 1.771.607 bp, 1.731.150 bp, and 1.835.763 bp for isolates 13.7 13A5, 05.5 8-1, and 13.7 2A-1, respectively. The isolates harbored unequal numbers of three, five, and six plasmids (Table 3). Another measure was comparing the relative locations of homologous genes present in all three genomes. Therefore, we determined the distance from base one of the circular maps, the first nucleotide of the dnaA gene, by taking three genes positioned at around three, six, and nine o’clock of the chromosomal map (Table 2; Figure 6A-C).
Beside the genome sizes and different cellular plasmid numbers and types, the genomic plasticity became apparent if the distance of homologous genes was reviewed. From the circular genome maps, we could infer that the gene order was overall the same, which can be easily seen by the locations of the five 23S ribosomal DNA gene cluster labeled in red (Figure 6A-C) and the relative distance in base pairs from chromosome base one to uvrA, polC, and secY, respectively (Table 2).
It is a quite common event in evolution that the genetic plasticity between subspecies is caused by horizontal gene transfer and transposable elements giving insertions of genes from other species that can be identified by regions of higher or lower G+C-content, that differ from the chromosomal G+C content of the investigated genome. This was observed as shown by the black areas in Figure 6E. Two areas of distinct lower G+C content occur in the genome on the left, one area in the middle genome, and none in the genome on the right, while the patterns are very similar further downstream. A Venn-diagram was computed showing the core set of 1352 genes present in all three genomes (Figure 6F). Strain 13.7.2A-1 had the largest genome with six inclusive plasmids. It possesses an astonishing number of 656 genes that are not present in the other two. Hence, the genome data substantiate the RAPD experiments und demonstrate the genomic plasticity of the three here in depth examined pediococcal isolates.
3.7. Bacteriocin Encoding Genes
The chromosomal and plasmidal DNA sequences were screened for the presence of bacteriocin-encoding genes using the BAGEL4-server, which, to our knowledge, emphasizes the most comprehensive bacteriocin database. As shown in Figure 7, gene regions were detected within the chromosomes of 13.7.13A-5 and 13.7.2A-1 that harbor a putative gene for the bacteriocin penocin A (light green). Directly upstream on the left is a gene associated that might confer immunity to penocin A (red). Two open reading frames are situated downstream. They encode a putative two-component system with a histidine kinase and a response regulator gene. The four genes upstream encode a putative PEP:dependent phosphotransferase system (PTS) for the consumption of ß-glucosides and a gene encoding a IIAGlc homolog of the PTS, which serves in many bacteria in glucose transport and down-regulation of second choice carbon sources when glucose is available [6].
A second bacteriocin gene for pediocin A was found on a plasmid of isolate 13.7.2A-1, showing that this isolate has two bacteriocins at its disposal. Isolate 05.5. 8-1, however, harbors a chromosomal gene locus for enterolysin A, a class III bacteriocin, and on one plasmid a locus with putative genes for pediocin A, an immunity protein, and genes that could encode an exporter for the secretion of the bacteriocin. The gene products of the plasmid gene loci were almost identical regarding pediocins A and the immunity proteins sharing 95% and 98% amino acid identities, respectively. The surrounding genes, including some for bacteriocin export (pink), were quite dissimilar. When all four bacteriocin proteins were aligned, they exhibited an overall protein sequence identity of 45% harboring the conserved motif “YGNGV(L)” and two conserved cysteins that form disulfite-bridges to determine the three-dimensional structures of these class II bacteriocins.
4. Discussion
The incorporation of protective cultures in food is increasing to meet the challenges of food safety and enhance food security [3,62]. Due to the diverse metabolic performance of LAB regarding the inhibition of pathogens, species from the order lactobacillales are of particular interest [3,18]. It has been reported that the LAB P. pentosaceus can serve in food protection and that isolates from different environments behave different, since they adapt genetically to the respective biotope from which they have been isolated [16]. To uncover novel P. pentosaceus strains for food protection, we retrieved from a typical LAB ecosystem, the udder of milk cows. LAB from milk have a long tradition to serve as the prime starter cultures for food fermentation giving taste and furthermore providing food preservation over months. Our data on 37 isolates of P. pentosaceus strains revealed that they effectively could combat pathogenic bacteria. However, there were differences as to the extent and to the ability regarding the inhibition of the challenged pathogens.
Pediococci have been isolated from milk, dairy products, fermented vegetables, sausages, or even aquatic products [12,16,63]. The data of the isolates derived from RAPD experiments showed throughout genetic variations suggesting that all isolates represent subspecies. In a previous study, although in less detail, comparable results from pediococci were reported for plant-derived LAB from traditional Ethiopian foods such as tef dough and kocho [64]. Quite similar results were reported when L. plantarum strains were compared [65]. Rossetti and Giraffa analyzed more than thousand LAB strains from raw cheese [23]. They established a databank of RAPD-profiles which can be used to identify an unknown isolate just by the gene fragment pattern of RAPD. However, P. pentosaceus strains were not included. The here presented data add to this. It is to our knowledge, the first of a set of P. pentosaceus isolates from one environment.
We found that all isolates showed antibacterial activity against a series of food pathogens and surrogates. The isolate P. pentosaceus 13.7 13A-5 was the best promising one. It was able to inhibit five out of indicators above average. The second-best performing isolates were a group of six that inhibited three out of seven above averages. The usage of the antimicrobial potential of P. pentosaceus in foods has been addressed recently on one P. pentosaceus and one Pediococcus acidilactici isolate from silage [66]. It was reported that they could combat 74 Listeria monocytogenes isolates and 27 different enterococci resistant to vancomycin. Furthermore, it was shown that these isolates inhibited many fungal species, most likely by several anti-fungal metabolites produced by this strain [66]. In another study, addition of a mixture of P. pentosaceus, P. acidilactici, and L. plantarum to alfalfa sprouts reduced the presence of L. monocytogenes by 4.5 log and of S. enterica by 1.0 log [67]. In accordance with our observation that isolates of the same species may have different antimicrobial profiles, Santini et. al. published that different isolates of strains of LAB and bifidobacteria exhibited different efficacies against three Campylobacter species [68]. Numerous reports focused on the role of purified bacteriocins produced by Pediococci. Especially, the presence of pediocin and pediocin-like bacteriocins from P. pentosaceus in meat products has been thoroughly examined [69].
The production of acid by a protective culture is an important issue to combat unwanted microorganisms. However, too much acidification may change the organoleptic characteristics of the food product [70,71]. We like to highlight that acid production not only depended on the respective P. pentosaceus isolate, but also on the presence of the competing strain, in this case the applied indicator strain. Our approach to measure the acidification capacity of each isolate against each indicator allowed us to select for weak acid producers while maintaining antimicrobial efficacy.
Finally, the genomes of the three best performing isolates were elucidated. The bioinformatic analyses corroborated the genetic plasticity of the P. pentosaceus isolates, and thus support the data obtained from RAPD analyses on all isolates. The analysis of the core genome shows this in striking detail. Only 1352 common genes occurred in the three genomes, while 178, 286, and 656, respectively, were singletons [58]. These findings were supported when we counter-checked available P. pediococcus genomes in the databases [63]. The number of plasmids of the sequenced isolates was three, five and six ranging from 12 kb to 68 kb. Plasmids carry often operons encoding genes for the metabolization of unusual carbon sources, bacteriocin-encoding genes, and transposable elements [72]. This can be explained by the adaptation to the various environments in which Pediococci occur [63]. Genome analysis for bacteriocin genes revealed that the strains harbor chromosomal and/or plasmidial genes for the bacteriocins pediocin A and penocin A together with genes for immunity to and export of the bacteriocin. The respective genes, genetic organization and functions have been characterized in molecular detail . The genetic surroundings, i.e. the presence of a carbohydrate:phosphotransferase system (PTS) for metabolism of the ß-glucosides and the PTS factor IIAGlc that controls the utilization of carbon sources is commonly associated with the pen operon [63,73,74]. The operons found in our sequenced genomes are conventionally present in many isolates of P. pentosaceus from diverse environments [75,76,77,78].
Although it is appealing to suggest that the action of bacteriocins is a major factor to push back unwanted microorganisms in food products, it should be noted that they might not always be expressed [79,80]. Expression greatly depends on growth conditions and is influenced by pH, temperature, quorum sensing, and the availability of carbon sources, essential amino acids, vitamins and energy [81]. In such cases other compounds such as diacetyl or chemicals like phenyllactate that lower the rH-Value could account for the protective action [3,18]. The answer to this question must be given by a proof of principle for each foodstuff containing an effective protective culture.
5. Conclusions
We showed that the isolation of P. pentosaceus strains from different milk cattle of different dairy farms revealed a high biodiversity. All isolates exhibited inhibitory activity against diverse pathogenic or food spoiling bacteria making them applicable in food products as protective cultures. Hence, the derived data can be used to select the matching isolate for a specific food product that needs a robust food safety along its shelf-life.
Our study reveals that it is worthwhile to set up a screening that includes several isolates of one species for a specific application, in this case for food protection. Our approach might be used as a blueprint for similar purposes such as the choice of the best starter culture as food fermentater and/or as a probiotic health-promoter.
Author Contributions
Conceptualization, SW.F. and F.T.; methodology, SW.F., A.E: and N.MC.; software, SW.F.; validation, SW.F; formal analysis, SW.F.; data curation, SW.F.; writing—original draft preparation, SW.F..; writing—review and editing SW.F. and F.T.; visualization, SW.F. and F.T..; supervision, F.T.; funding acquisition, SW.F. and F.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financed by the Federal Ministry of Economic Affairs and Climate Action through a research grant of the “Zentrales Innovationsprogramm Mittelstand (ZF4679802AJ2)”. Further Support was received by the Open Access Publication Fund of the University of Bonn.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Jan Kok and Antonio Cruz for expert assistance and many helpful discussions. We are grateful to Knut Jahreis for providing strains used in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ozen, M.; Dinleyici, E.C. The History of Probiotics: The Untold Story. Beneficial Microbes 2015, 6, 159–166. [CrossRef]
- Bamforth, C.W.; Cook, D.J. Food, Fermentation and Microorganisms; 2nd ed.; Wiley Blackwell: Hoboken, 2019; ISBN 978-1-119-55743-2.
- Fischer, S.W.; Titgemeyer, F. Protective Cultures in Food Products: From Science to Market. Foods 2023, 12, 1541. [CrossRef]
- Mendonça, A.A.; Pinto-Neto, W. de P.; da Paixão, G.A.; Santos, D. da S.; De Morais, M.A.; De Souza, R.B. Journey of the Probiotic Bacteria: Survival of the Fittest. Microorganisms 2023, 11, 95. [CrossRef]
- Vasiljevic, T.; Shah, N.P. Probiotics—From Metchnikoff to Bioactives. International Dairy Journal 2008, 18, 714–728. [CrossRef]
- Parche, S.; Amon, J.; Jankovic, I.; Rezzonico, E.; Beleut, M.; Barutçu, H.; Schendel, I.; Eddy, M.P.; Burkovski, A.; Arigoni, F.; et al. Sugar Transport Systems of Bifidobacterium Longum NCC2705. Journal of Molecular Microbiology and Biotechnology 2006, 12, 9–19. [CrossRef]
- Foligné, B.; Daniel, C.; Pot, B. Probiotics from Research to Market: The Possibilities, Risks and Challenges. Current Opinion in Microbiology 2013, 16, 284–292. [CrossRef]
- Redzepi, R.; Zilber, D.; Sung, E.; Troxler, P. The Noma Guide to Fermentation: Foundations of Flavour; Artisan: New York, 2018; ISBN 978-1-57965-718-5.
- Katz, S.E. Wild Fermentation: The Flavor, Nutrition, and Craft of Live-Culture Foods; Revised and updated edition.; Chelsea Green Publishing: White River Junction, Vermont, 2016; ISBN 978-1-60358-628-3.
- Bergey’s Manual of Systematic Bacteriology; Boone, D.R., Castenholz, R.W., Garrity, G.M., Eds.; 2nd ed.; Springer: New York, 2009; ISBN 978-0-387-98771-2.
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. International Journal of Systematic and Evolutionary Microbiology 2020, 70, 2782–2858. [CrossRef]
- Jiang, S.; Cai, L.; Lv, L.; Li, L. Pediococcus Pentosaceus, a Future Additive or Probiotic Candidate. Microbial Cell Factories 2021, 20, 45. [CrossRef]
- Raccach, M. Pediococcus. In Encyclopedia of Food Microbiology (Second Edition); Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, 2014; pp. 1–5 ISBN 978-0-12-384733-1.
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.-M.; Landschoot, A.V.; Vuyst, L.D.; Vandamme, P. The Microbial Diversity of Traditional Spontaneously Fermented Lambic Beer. PLOS ONE 2014, 9, e95384. [CrossRef]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The Microbial Diversity of an Industrially Produced Lambic Beer Shares Members of a Traditionally Produced One and Reveals a Core Microbiota for Lambic Beer Fermentation. Food Microbiology 2015, 49, 23–32. [CrossRef]
- Qi, Y.; Huang, L.; Zeng, Y.; Li, W.; Zhou, D.; Xie, J.; Xie, J.; Tu, Q.; Deng, D.; Yin, J. Pediococcus Pentosaceus: Screening and Application as Probiotics in Food Processing. Front. Microbiol. 2021, 12. [CrossRef]
- Porto, M.C.W.; Kuniyoshi, T.M.; Azevedo, P.O.S.; Vitolo, M.; Oliveira, R.P.S. Pediococcus Spp.: An Important Genus of Lactic Acid Bacteria and Pediocin Producers. Biotechnology Advances 2017, 35, 361–374. [CrossRef]
- Vandenbergh, P.A. Lactic Acid Bacteria, Their Metabolic Products and Interference with Microbial Growth. FEMS Microbiology Reviews 1993, 12, 221–237. [CrossRef]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A MEDIUM FOR THE CULTIVATION OF LACTOBACILLI. Journal of Applied Bacteriology 1960, 23, 130–135. [CrossRef]
- Lee, H.M.; Lee, Y. A Differential Medium for Lactic Acid-Producing Bacteria in a Mixed Culture. Lett Appl Microbiol 2008, 46, 676–681. [CrossRef]
- Ames, B.N.; McCann, J.; Yamasaki, E. Methods for Detecting Carcinogens and Mutagens with the Salmonella/Mammalian-Microsome Mutagenicity Test. Mutation Research/Environmental Mutagenesis and Related Subjects 1975, 31, 347–363. [CrossRef]
- Judicial Commission of the International Committee on Systematics of Prokaryotes The Type Species of the Genus Salmonella Lignieres 1900 Is Salmonella Enterica (Ex Kauffmann and Edwards 1952) Le Minor and Popoff 1987, with the Type Strain LT2T, and Conservation of the Epithet Enterica in Salmonella Enterica over All Earlier Epithets That May Be Applied to This Species. Opinion 80. International Journal of Systematic and Evolutionary Microbiology 2005, 55, 519–520. [CrossRef]
- Rossetti, L.; Giraffa, G. Rapid Identification of Dairy Lactic Acid Bacteria by M13-Generated, RAPD-PCR Fingerprint Databases. Journal of Microbiological Methods 2005, 63, 135–144. [CrossRef]
- Sirichoat, A.; Flórez, A.B.; Vázquez, L.; Buppasiri, P.; Panya, M.; Lulitanond, V.; Mayo, B. Antibiotic Susceptibility Profiles of Lactic Acid Bacteria from the Human Vagina and Genetic Basis of Acquired Resistances. IJMS 2020, 21, 2594. [CrossRef]
- Tomás, M.S.J.; Claudia Otero, M.; Ocaña, V.; Elena Nader-Macías, M. Production of Antimicrobial Substances by Lactic Acid Bacteria I: Determination of Hydrogen Peroxide. In Methods in Molecular Biology; Spencer, J.F.T., Ed.; Public Health Microbiology: Methods and Protocols; Humana Press: New Jersey, 2004; Vol. 268, pp. 337–346 ISBN 978-1-59259-766-6. [CrossRef]
- Adeniyi, B.A.; Adetoye, A.; Ayeni, F.A. Antibacterial Activities of Lactic Acid Bacteria Isolated from Cow Faeces against Potential Enteric Pathogens. Afr Health Sci 2015, 15, 888–895. [CrossRef]
- Yadav, M.K.; Singh, B.; Tiwari, S.K. Comparative Analysis of Inhibition-Based and Indicator-Independent Colorimetric Assay for Screening of Bacteriocin-Producing Lactic Acid Bacteria. 2021, 687–695. [CrossRef]
- Nanopore Technologies MinKNOW.
- R Core Team R: A Language and Environment for Statistical Computing 2021.
- RStudio Team RStudio: Integrated Development Environment for R 2022.
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. Journal of Open Source Software 2019, 4, 1686. [CrossRef]
- Wickham, H.; Bryan, J. Readxl: Read Excel Files 2022.
- Huber, N. Ggdendroplot: Create Dendrograms for Ggplot2 2023.
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; M. Henry H. Stevens; Szoecs, E.; et al. Vegan: Community Ecology Package 2022.
- Ghazi, F.; Kihal, M.; Altay, N.; Gürakan, G.C. Comparison of RAPD-PCR and PFGE Analysis for the Typing of Streptococcus Thermophilus Strains Isolated from Traditional Turkish Yogurts. Ann Microbiol 2016, 66, 1013–1026. [CrossRef]
- Hilton, A.C.; Mortiboy, D.; Banks, J.G.; Penn, C.W. RAPD Analysis of Environmental, Food and Clinical Isolates of Campylobacter Spp. FEMS Immunology & Medical Microbiology 1997, 18, 119–124. [CrossRef]
- Szaluś-Jordanow, O.; Krysztopa-Grzybowska, K.; Czopowicz, M.; Moroz, A.; Mickiewicz, M.; Lutyńska, A.; Kaba, J.; Nalbert, T.; Frymus, T. MLST and RAPD Molecular Analysis of Staphylococcus Aureus Subsp. Anaerobius Isolated from Goats in Poland. Arch Microbiol 2018, 200, 1407–1410. [CrossRef]
- Ruiz, P.; Seseña, S.; Palop, M.L. A Comparative Study of Different PCR-Based DNA Fingerprinting Techniques for Typing of Lactic Acid Bacteria. Eur Food Res Technol 2014, 239, 87–98. [CrossRef]
- Nielsen, K.L.; Godfrey, P.A.; Stegger, M.; Andersen, P.S.; Feldgarden, M.; Frimodt-Møller, N. Selection of Unique Escherichia Coli Clones by Random Amplified Polymorphic DNA (RAPD): Evaluation by Whole Genome Sequencing. Journal of Microbiological Methods 2014, 103, 101–103. [CrossRef]
- Birch, M.; Denning, D.W.; Law, D. Rapid Genotyping ofEscherichia Coli O157 Isolates by Random Amplification of Polymorphic DNA. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 297–302. [CrossRef]
- Mölder, F.; Jablonski, K.P.; Letcher, B.; Hall, M.B.; Tomkins-Tinch, C.H.; Sochat, V.; Forster, J.; Lee, S.; Twardziok, S.O.; Kanitz, A.; et al. Sustainable Data Analysis with Snakemake 2021.
- Nanopore Technologies Guppy.
- Wick, R.; Volkening, J. Porechop 2018.
- Wick, R. Filtlong 2024.
- Wick, R.R.; Judd, L.M.; Cerdeira, L.T.; Hawkey, J.; Méric, G.; Vezina, B.; Wyres, K.L.; Holt, K.E. Trycycler: Consensus Long-Read Assemblies for Bacterial Genomes. Genome Biology 2021, 22, 266. [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat Biotechnol 2019, 37, 540–546. [CrossRef]
- Raven Team Raven 2024.
- Miniasm: Ultrafast de Novo Assembly for Long Noisy Reads.
- Wick, R.R.; Holt, K.E. Benchmarking of Long-Read Assemblers for Prokaryote Whole Genome Sequencing 2021. [CrossRef]
- Ondov, B.D.; Starrett, G.J.; Sappington, A.; Kostic, A.; Koren, S.; Buck, C.B.; Phillippy, A.M. Mash Screen: High-Throughput Sequence Containment Estimation for Genome Discovery. Genome Biol 2019, 20, 232. [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast Genome and Metagenome Distance Estimation Using MinHash. Genome Biol 2016, 17, 132. [CrossRef]
- Edgar, R.C. High-Accuracy Alignment Ensembles Enable Unbiased Assessments of Sequence Homology and Phylogeny 2022, 2021.06.20.449169. [CrossRef]
- Nanopore Technologies Medaka 2024.
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and Standardized Annotation of Bacterial Genomes via Alignment-Free Sequence Identification: Find out More about Bakta, the Motivation, Challenges and Applications, Here. Microbial Genomics 2021, 7. [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat Commun 2018, 9, 5114. [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-Depth Characterization and Visualization of Bacterial Genomes. Nucleic Acids Research 2023, 51, W484–W492. [CrossRef]
- Dieckmann, M.A.; Beyvers, S.; Nkouamedjo-Fankep, R.C.; Hanel, P.H.G.; Jelonek, L.; Blom, J.; Goesmann, A. EDGAR3.0: Comparative Genomics and Phylogenomics on a Scalable Infrastructure. Nucleic Acids Research 2021, 49, W185–W192. [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A User-Friendly Web Server to Thoroughly Mine RiPPs and Bacteriocins. Nucleic Acids Research 2018, 46, W278–W281. [CrossRef]
- Albano, H.; Oliveira, M.; Aroso, R.; Cubero, N.; Hogg, T.; Teixeira, P. Antilisterial Activity of Lactic Acid Bacteria Isolated from “Alheiras” (Traditional Portuguese Fermented Sausages): In Situ Assays. Meat Science 2007, 76, 796–800. [CrossRef]
- Banwo, K.; Sanni, A.; Tan, H. Functional Properties of Pediococcus Species Isolated from Traditional Fermented Cereal Gruel and Milk in Nigeria. Food Biotechnology 2013, 27, 14–38. [CrossRef]
- Garin-Murguialday, N.; Espina, L.; Virto, R.; Pagán, R. Lactic Acid Bacteria and Bacillus Subtilis as Potential Protective Cultures for Biopreservation in the Food Industry. Applied Sciences 2024, 14, 4016. [CrossRef]
- Jiang, J.; Yang, B.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Chen, W. Comparative Genomics of Pediococcus Pentosaceus Isolated From Different Niches Reveals Genetic Diversity in Carbohydrate Metabolism and Immune System. Frontiers in Microbiology 2020, 11. [CrossRef]
- Nigatu; Ahrné; Gashe; Molin Randomly Amplified Polymorphic DNA (RAPD) for Discrimination of Pediococcus Pentosaceus and Ped. Acidilactici and Rapid Grouping of Pediococcus Isolates. Letters in Applied Microbiology 1998, 26, 412–416. [CrossRef]
- Johansson, M.-L.; Quednau, M.; Molin, G.; Ahrné, S. Randomly Amplified Polymorphic DNA (RAPD) for Rapid Typing of Lactobacillus Plantarum Strains. Lett Appl Microbiol 1995, 21, 155–159. [CrossRef]
- Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Park, Y.J.; Suh, D.H.; Jung, E.S.; Franco, B.D.G.D.M.; Ivanova, I.V.; Holzapfel, W.H.; Todorov, S.D. Antimicrobial Properties of Pediococcus Acidilactici and Pediococcus Pentosaceus Isolated from Silage. J of Applied Microbiology 2022, 132, 311–330. [CrossRef]
- Rossi, F.; Lathrop, A. Effects of Lactobacillus Plantarum, Pediococcus Acidilactici, and Pediococcus Pentosaceus on the Growth of Listeria Monocytogenes and Salmonella on Alfalfa Sprouts. Journal of Food Protection 2019, 82, 522–527. [CrossRef]
- Santini, C.; Baffoni, L.; Gaggia, F.; Granata, M.; Gasbarri, R.; Di Gioia, D.; Biavati, B. Characterization of Probiotic Strains: An Application as Feed Additives in Poultry against Campylobacter Jejuni. International Journal of Food Microbiology 2010, 141, S98–S108. [CrossRef]
- Khorshidian, N.; Khanniri, E.; Mohammadi, M.; Mortazavian, A.M.; Yousefi, M. Antibacterial Activity of Pediocin and Pediocin-Producing Bacteria Against Listeria Monocytogenes in Meat Products. Front. Microbiol. 2021, 12, 709959. [CrossRef]
- Deshwal, G.K.; Tiwari, S.; Kumar, A.; Raman, R.K.; Kadyan, S. Review on Factors Affecting and Control of Post-Acidification in Yoghurt and Related Products. Trends in Food Science & Technology 2021, 109, 499–512. [CrossRef]
- Ghosh, D.; Chattopadhyay, P. Application of Principal Component Analysis (PCA) as a Sensory Assessment Tool for Fermented Food Products. J Food Sci Technol 2012, 49, 328–334. [CrossRef]
- Shintani, M.; Vestergaard, G.; Milaković, M.; Kublik, S.; Smalla, K.; Schloter, M.; Udiković-Kolić, N. Integrons, Transposons and IS Elements Promote Diversification of Multidrug Resistance Plasmids and Adaptation of Their Hosts to Antibiotic Pollutants from Pharmaceutical Companies. Environmental Microbiology 2023, 25, 3035–3051. [CrossRef]
- Martino, M.E.; Maifreni, M.; Marino, M.; Bartolomeoli, I.; Carraro, L.; Fasolato, L.; Cardazzo, B. Genotypic and Phenotypic Diversity of Pediococcus Pentosaceus Strains Isolated from Food Matrices and Characterisation of the Penocin Operon. Antonie van Leeuwenhoek 2013, 103, 1149–1163. [CrossRef]
- Lengeler, J.W.; Titgemeyer, F.; Vogler, A.P.; Wohrl, B.M.; Kornberg, H.L.; Henderson, P.J.F. Structures and Homologies of Carbohydrate: Phosphotransferase System (PTS) Proteins. Philosophical Transactions of the Royal Society of London. B, Biological Sciences 1997, 326, 489–504. [CrossRef]
- Rodrigues Blanco, I.; José Luduverio Pizauro, L.; Victor Dos Anjos Almeida, J.; Miguel Nóbrega Mendonça, C.; De Mello Varani, A.; Pinheiro De Souza Oliveira, R. Pan-Genomic and Comparative Analysis of Pediococcus Pentosaceus Focused on the in Silico Assessment of Pediocin-like Bacteriocins. Computational and Structural Biotechnology Journal 2022, 20, 5595–5606. [CrossRef]
- Papagianni, M.; Anastasiadou, S. Pediocins: The Bacteriocins of Pediococci. Sources, Production, Properties and Applications. Microbial Cell Factories 2009, 8, 3. [CrossRef]
- Rodríguez, J.M.; Martínez, M.I.; Kok, J. Pediocin PA-1, a Wide-Spectrum Bacteriocin from Lactic Acid Bacteria. Critical Reviews in Food Science and Nutrition 2002, 42, 91–121. [CrossRef]
- Venema, K.; Kok, J.; Marugg, J.D.; Toonen, M.Y.; Ledeboer, A.M.; Venema, G.; Chikindas, M.L. Functional Analysis of the Pediocin Operon of Pediococcus Acidilactici PAC1.0: PedB Is the Immunity Protein and PedD Is the Precursor Processing Enzyme. Molecular Microbiology 1995, 17, 515–522. [CrossRef]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of Culture Media, pH and Temperature on Growth and Bacteriocin Production of Bacteriocinogenic Lactic Acid Bacteria. AMB Express 2018, 8, 10. [CrossRef]
- Mataragas, M.; Metaxopoulos, J.; Galiotou, M.; Drosinos, E.H. Influence of pH and Temperature on Growth and Bacteriocin Production by Leuconostoc Mesenteroides L124 and Lactobacillus Curvatus L442. Meat Science 2003, 64, 265–271. [CrossRef]
- Aasen, I.M.; Møretrø, T.; Katla, T.; Axelsson, L.; Storrø, I. Influence of Complex Nutrients, Temperature and pH on Bacteriocin Production by Lactobacillus Sakei CCUG 42687. Applied Microbiology and Biotechnology 2000, 53, 159–166. [CrossRef]
Figure 1.
RAPD analyses. Shown are 2% agarose gels showing gene fragment pattern from RAPD analyses. Isolates from farm 4 are shown on the left, those from farm 13 are shown on the right.
Figure 1.
RAPD analyses. Shown are 2% agarose gels showing gene fragment pattern from RAPD analyses. Isolates from farm 4 are shown on the left, those from farm 13 are shown on the right.
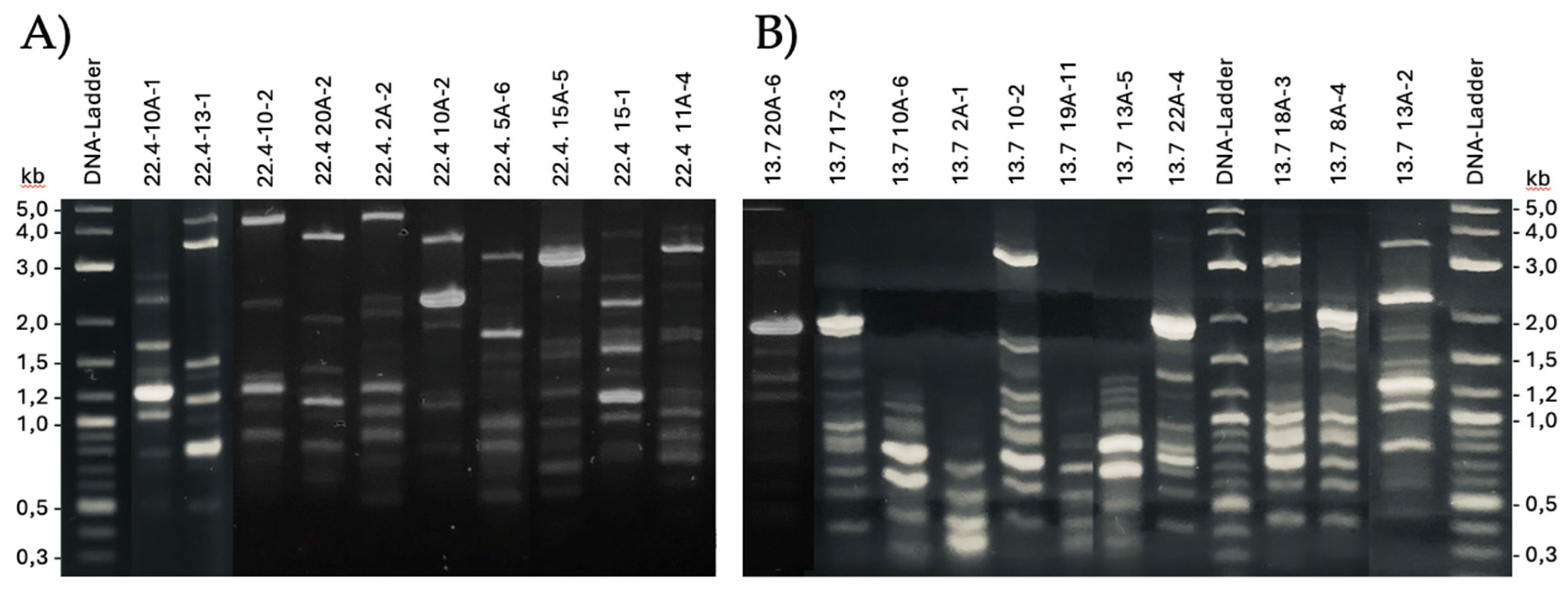
Figure 2.
Depiction of the phylogenetic relationships within isolates from one farm. Relative phylogenetic distances are indicated by scale. The red line depicts the 95% similarity criterium to define a subspecies. A) farm 1 (n=5); B) farm 4 (n=16); C) farm 5 (n=5); D) farm 13 (n=11).
Figure 2.
Depiction of the phylogenetic relationships within isolates from one farm. Relative phylogenetic distances are indicated by scale. The red line depicts the 95% similarity criterium to define a subspecies. A) farm 1 (n=5); B) farm 4 (n=16); C) farm 5 (n=5); D) farm 13 (n=11).
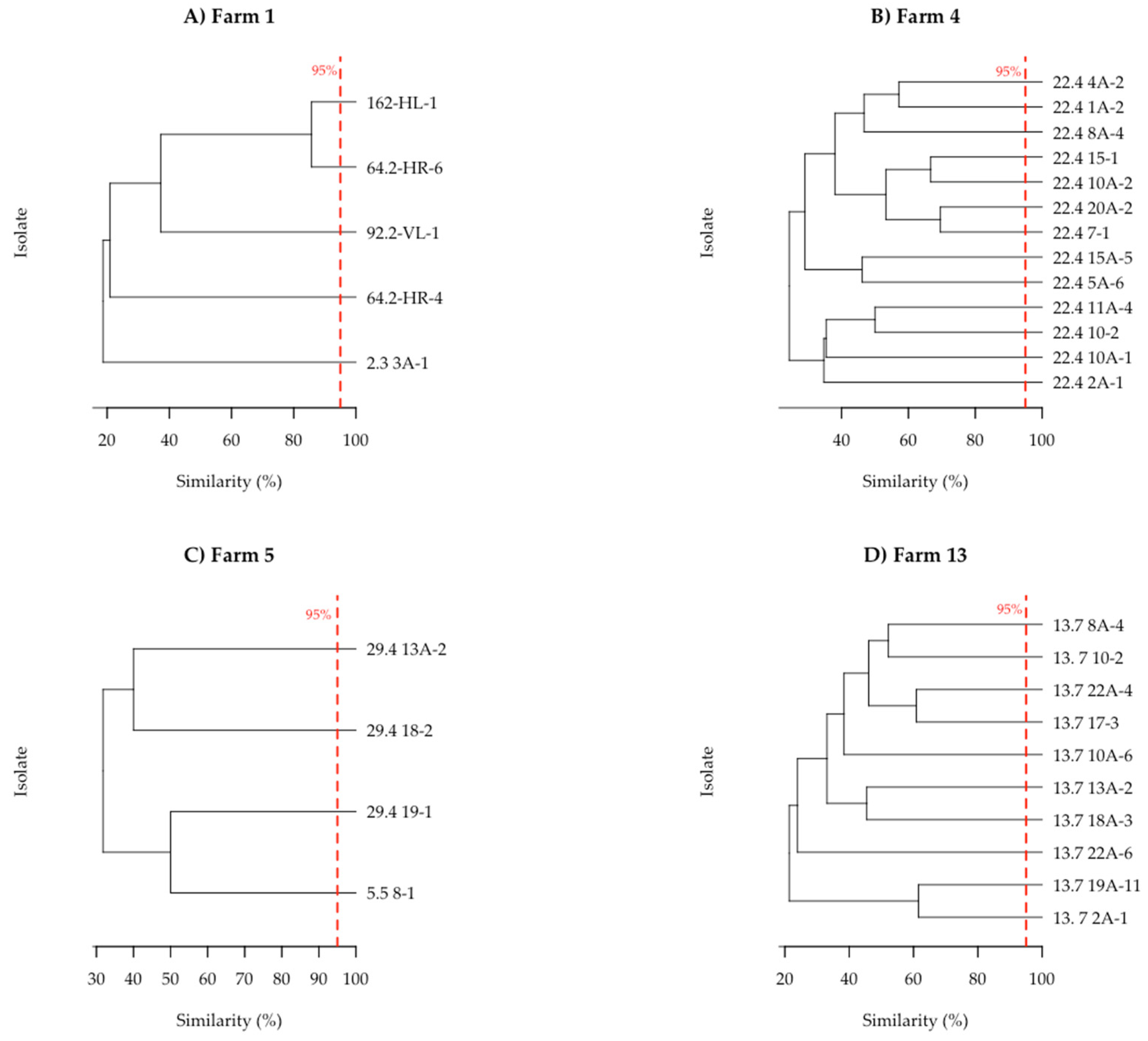
Figure 3.
Heatmap of antibacterial activities. The color scale shows the normalized z-values. Indicators are ordered by falling susceptibility from left to right. The clustering on the right shows the subspecies of P. pentosaceus according to the similarity of their antibacterial profiles. Experiments were carried out in triplicate.
Figure 3.
Heatmap of antibacterial activities. The color scale shows the normalized z-values. Indicators are ordered by falling susceptibility from left to right. The clustering on the right shows the subspecies of P. pentosaceus according to the similarity of their antibacterial profiles. Experiments were carried out in triplicate.
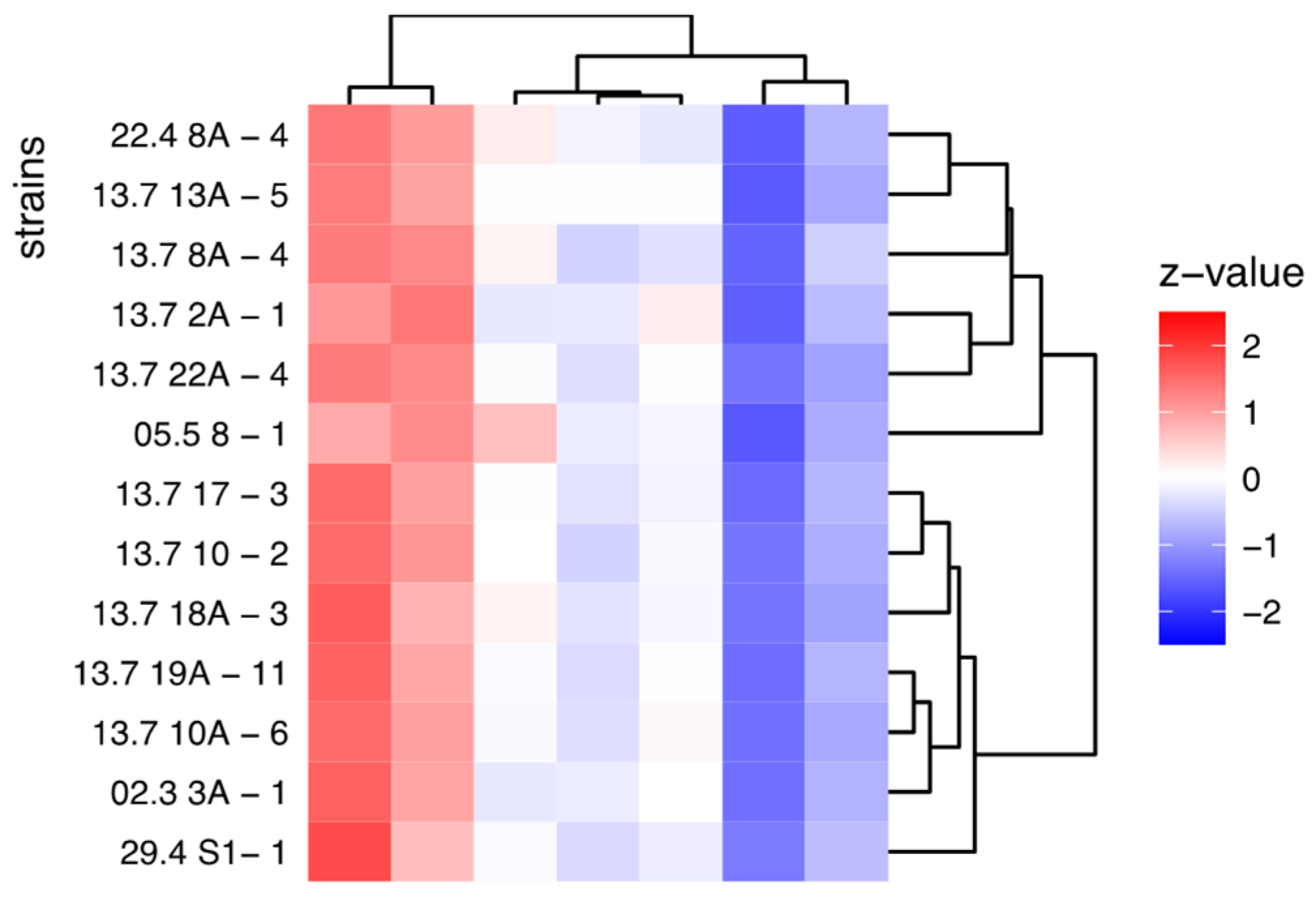
Figure 4.
Heatmap of pH drop of P. pentosaceus isolates against indicator strains. The columns show the normalized z-values of acidification ranging from blue (strong) ) to white (fair) to red (low). Indicator strains clustering is shown on the top, while clustering of the isolates is shown on the right. Data represent the average of triplicates.
Figure 4.
Heatmap of pH drop of P. pentosaceus isolates against indicator strains. The columns show the normalized z-values of acidification ranging from blue (strong) ) to white (fair) to red (low). Indicator strains clustering is shown on the top, while clustering of the isolates is shown on the right. Data represent the average of triplicates.
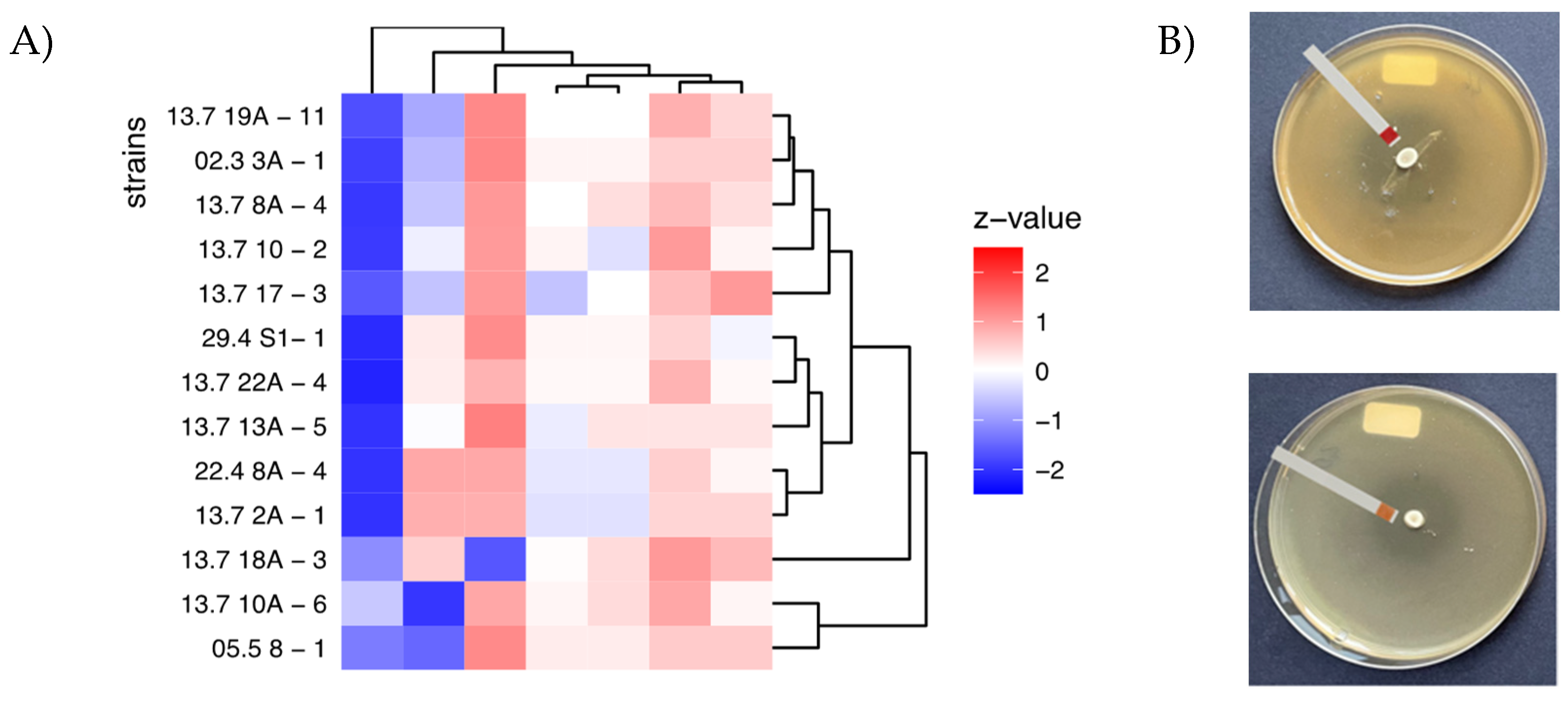
Figure 5.
Pearson correlation analysis. Shown is the drop in pH versus the inhibition zone for A) indicators and B) P. pentosaceus isolates. r, correlation coefficient; p, p-value significance of <0,05.
Figure 5.
Pearson correlation analysis. Shown is the drop in pH versus the inhibition zone for A) indicators and B) P. pentosaceus isolates. r, correlation coefficient; p, p-value significance of <0,05.
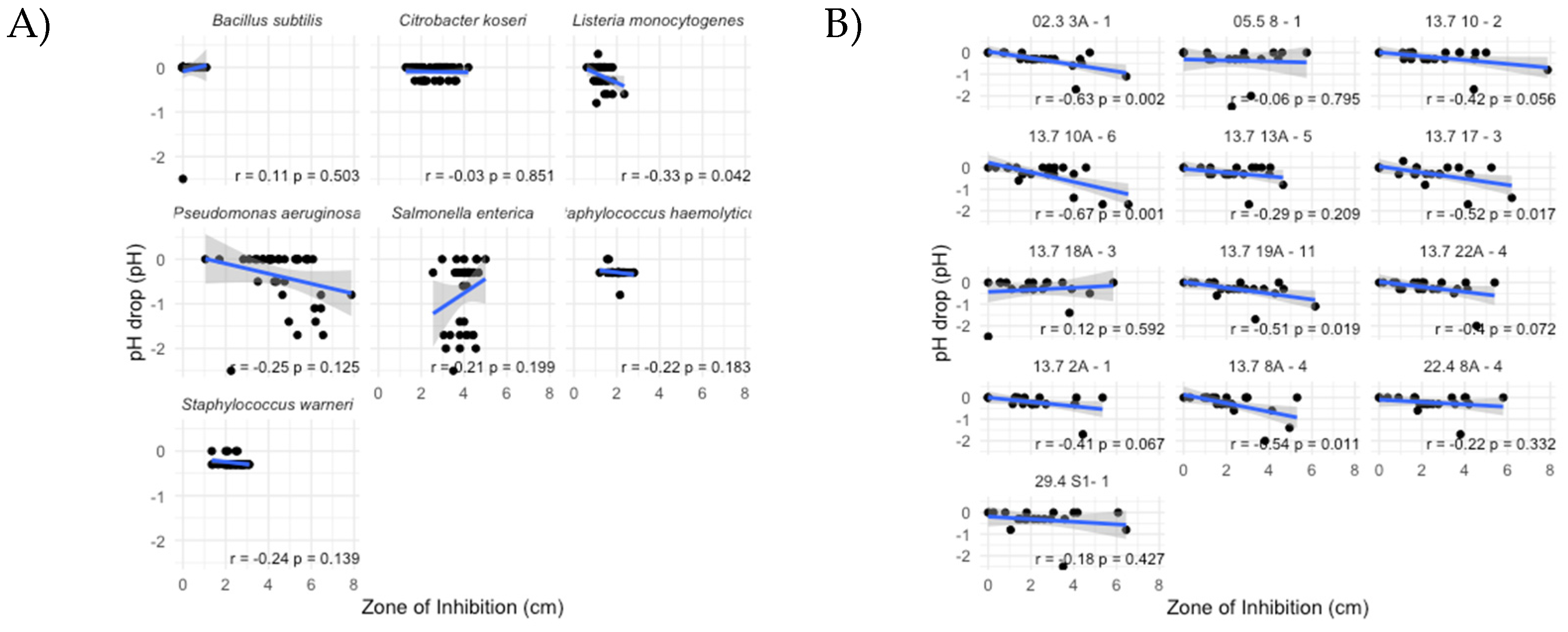
Figure 6.
Genomic landscape. (A-C) Shown are the chromosomal maps of the three indicated P.pentosaceus genomes. Genes are in blue, the five 23S rRNA gene cluster are labeled red. (D) FastANI diagrams showing shifts of gene segments or insertion or deletions of binary genome comparisons. (E) Sections from each genome map around the first rRNA region (red) show gene insertions visible by the G+C content graph (black). (F) Venn diagram showing the numbers of common genes between the genomes.
Figure 6.
Genomic landscape. (A-C) Shown are the chromosomal maps of the three indicated P.pentosaceus genomes. Genes are in blue, the five 23S rRNA gene cluster are labeled red. (D) FastANI diagrams showing shifts of gene segments or insertion or deletions of binary genome comparisons. (E) Sections from each genome map around the first rRNA region (red) show gene insertions visible by the G+C content graph (black). (F) Venn diagram showing the numbers of common genes between the genomes.
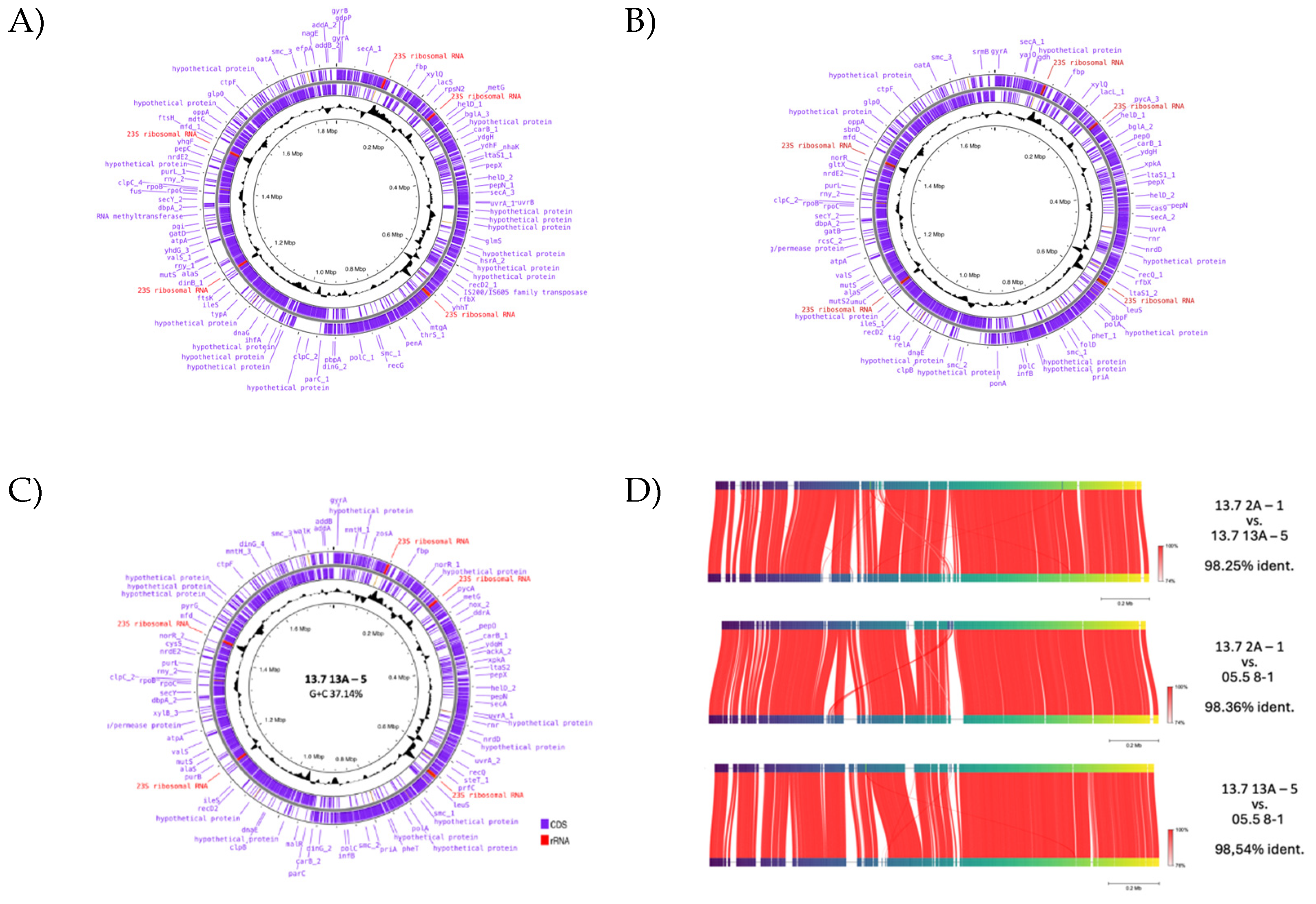
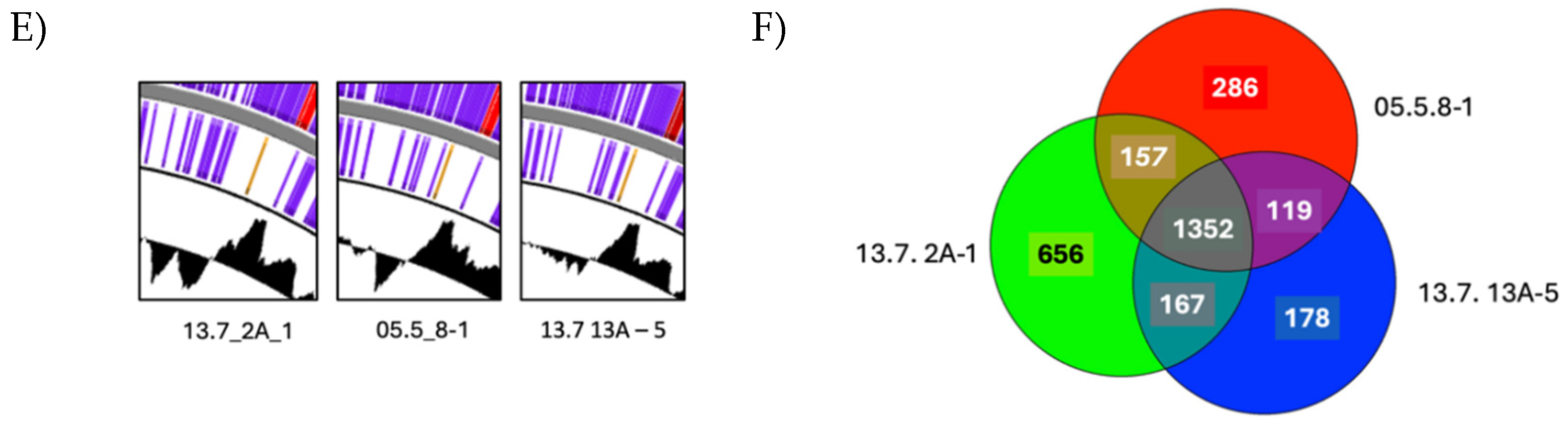
Figure 7.
A) Genetic regions of genes involved in bacteriocin metabolism. B) Multiple alignment of bacteriocin protein sequences. The conserved sequence motifs “YGNGV(L) including the two functional cysteines are shown in red. *chromosomal; **plasmidial. The consensus sequence below shows identical (*) and conserved (:, .) amino acid positions.
Figure 7.
A) Genetic regions of genes involved in bacteriocin metabolism. B) Multiple alignment of bacteriocin protein sequences. The conserved sequence motifs “YGNGV(L) including the two functional cysteines are shown in red. *chromosomal; **plasmidial. The consensus sequence below shows identical (*) and conserved (:, .) amino acid positions.
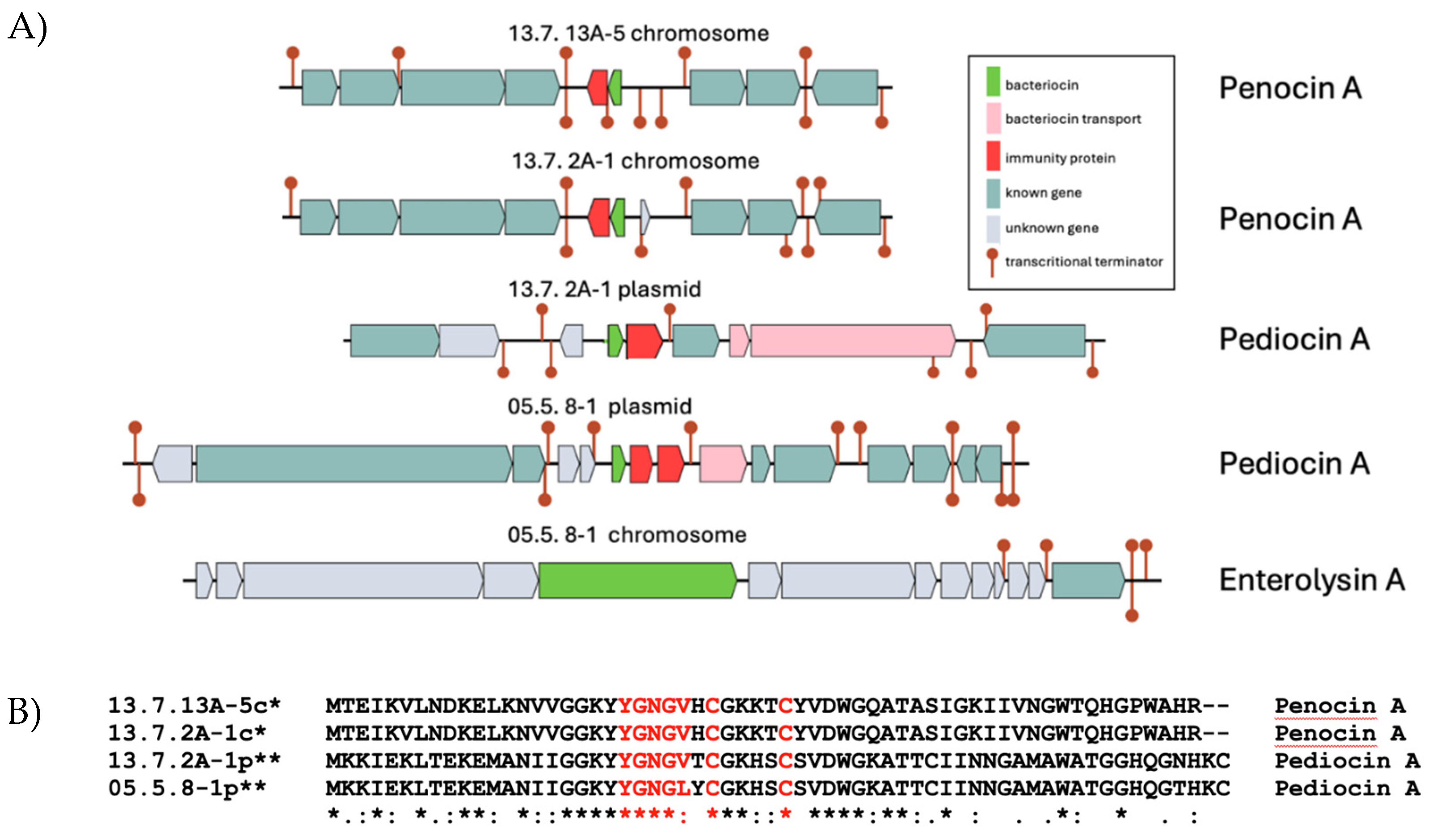
Table 2.
Ranking of isolates regarding antibacterial activity.
| ranking | strain | No. inhibited pathogens above average | sum | mean | p_value |
|---|---|---|---|---|---|
| 1 | 13.7 13A - 5 | 5 | 2.465265 | 0.3521807 | 0.20896 |
| 2 | 05.5 8 - 1 | 3 | 2.728224 | 0.3897463 | 0.79546 |
| 3 | 13.7 2A - 1 | 3 | 2.654947 | 0.3792781 | 0.06718 |
| 4 | 13.7 10 - 2 | 3 | 2.612546 | 0.3732209 | 0.05576 |
| 5 | 22.4 8A - 4 | 3 | 2.607411 | 0.3724873 | 0.33180 |
| 6 | 13.7 22A - 4 | 3 | 2.606640 | 0.3723771 | 0.07161 |
| 7 | 13.7 18A - 3 | 3 | 2.577932 | 0.3682761 | 0.59202 |
| 8 | 29.4 S1- 1 | 2 | 2.506960 | 0.3581371 | 0.42734 |
Table 3.
Key data of the genomes.
| strain | 13.7 13A–5 | 05.5 8-1 | 13.7 2A-1 |
|---|---|---|---|
| genome (bp) | 1771607 | 1731150 | 1835763 |
| predicted genes | 1,816 | 1,914 | 2,332 |
| plasmids (bp) | 12,144; 12,513; 29,159 | 13,332; 20,517; 21,974; 24,254; 68,459 | 12,153; 16,367; 22,192; 36,437; 42,648; 44,253 |
| G+C content (%) | 37.14 | 37.41 | 37.11 |
| uvrA1 | 490,671 | 464,679 | 459,969 |
| polC1 | 856,007 | 806,61 | 867,588 |
| secY1 | 1,320,181 | 1,287,919 | 1,381,749 |
1 position gene start.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



