
Submitted:
20 November 2024
Posted:
21 November 2024
You are already at the latest version
Abstract
Plastic pollution is one of the biggest current global threats to the environment given that petroleum-based plastic is recalcitrant and can stay in the environment for decades, even centuries, depending on the specific plastic type. Since less than 10% of all the plastic ever made is recycled and other solutions (such as incineration or landfill storage) are pollutant methods, new environmentally friendly solutions are needed. In this regard, the latest biotechnological discovery on this topic is the capability of insect larvae to use plastic polymers as carbon feedstock. This present review describes the most relevant information on insect larvae capable of degrading plastic, mainly from Galleria mellonella (Fabricius, 1798), Tenebrio molitor (Linnaeus, 1758) and Zophobas atratus (Fabricius, 1776) but also adds new information about other less studied “plastivore” insects such as the termites. The review covers from the very first work describing plastic degradation by larvae in 2014 all the way till the very latest research available (till June 2024) focusing on the identification of a wide variety of plastic-degrading microorganisms isolated from larvae’s gut to date and on the understanding of the potential molecular mechanisms present for the degradation to take place. It also describes the latest discoveries which include the identification of novel enzymes from the waxworm’s saliva.
Keywords:
plastic degradation
; plastivore larvae
; waxworms
; mealworms
; superworms
1. Introduction
Plastic pollution represents one of the major global challenges of this era and yet, it has been reported that only the 9% out of the 9 billion tons of plastic ever produced had been recycled [1]. As plastic pollution accumulates in the environment at alarming rates new and more effective solutions to address this problem are needed. A novel biotechnological approach is to degrade this plastic into its original monomers using microorganisms and their enzymes so that these monomers can be potentially later upcycled into new high-value products [2]. An even newer biotechnological trend is the use of insect larvae for the same purpose [3].
Even though, observation of common insect pests such as Rhyzopertha dominica (commonly referred to as the lesser grain borer) and Tenebroides mauritanicus (Cadelle beetle) penetrating packaging materials has been observed since the 1950’s, the main concern at the time was to protect packaged food from these invaders [4,5].
The first scientific report suggesting the revolutionizing idea of using insects to fight plastic pollution did not come until 2014 using the larvae Plodia interpunctella [6,7]. This report was followed by the first report of full PS mineralization into CO2 by T. molitor’s larvae [8] and the first observation of the waxworm (Galleria mellonella’s larvae) degrading PE in 2017 [9]. The same year, this discovery hit the news and was published in National Geographic to reach the general public where Dr. Federica Bertocchini was acknowledged as the discoverer [10]. She and her research group in Spain , later in 2022, identified 4 novel waxworm saliva enzymes responsible for this degradation and named them as Demetra, Cibeles, Ceres and Cora which are the first plastic-degrading enzymes ever isolated from an invertebrate organism [11,12]. Other relevant events also include the introduction of the term “plastivore” to describe insect larvae or any other organism capable of using plastic as carbon feedstock [13] and the report of plastic-degrading yeasts from adult termites gut [14]. All these important events are illustrated in Figure 1 in chronological order.
The number of scientific papers related to plastic-eating larvae is growing every year and yet, the number of papers to date are still scarce. Till June 2024 only 366 papers resulted from the keyword search “insect larvae to degrade plastic” from PubMed and only a couple of them are literature reviews. The first literature review published ever that summarizes insect degradation of plastics was in the year 2021 [15] and the latest is from in 2024 [16] and there are very few reviews in between [3,17]. This review expands the knowledge on plastivore larvae even more and includes the latest research information to date (Till June 2024). Specifically, it is the most comprehensive and thorough literature review about the waxworm (Galleria’s mellonella larvae) published to date, but it also reviews mealworms (Tenebrio molitor) and superworms ( Zophobas atratus), focusing on the identification of plastic-degrading microorganisms that have been identified in these larvae’s gut and on the understanding of the potential molecular mechanisms present on the larvae for the degradation to take place. It also describes the latest discoveries which includes the identification of novel enzymes from the waxworm’s saliva and the first potential adult plastivores: the termites.
2. Plastic Pollution
Around 8300 million metric tons of virgin plastics had been produced by 2017 from non-renewable petrochemical feedstocks and only a small proportion has been recycled or incinerated. It is estimated that if current trends continue, approximately 1200 metric tons of plastic waste will accumulate in landfills or in the natural environment by 2050. Whilst this analysis includes thermoplastics, thermosets, polyurethanes (PURs), elastomers, coatings, and sealants, it mainly focuses on the most abundant resins and fibres: high-density polyethylene (HDPE), low-density and linear low-density PE (LPE), polypropylene (PP), polystyrene (PS), polyvinylchloride (PVC), poly-ethylene terephthalate (PET), and PUR resins; and polyester, poly-amide, and acrylic (PP&A) fibres [18].
Molecular structure of these common plastic resins (81% plastics) along with their density, crystallinity, life span in the environment [19,20], common uses and demand distribution by resin type in the year 2018 in Europe [21] are reported in Figure 2. The rest 19% of resins that are not presented in Figure 2 include PTFE for cable coatings in communications, PMMA for touch screens, PC for roofs and eye glasses, PBT as optical fibre , ABS for keyboards, LEGO toys and others [21]. Half-life of PU is still unknown [22].
The ubiquitous distribution of plastic contamination in both terrestrial and marine environments has identified the phenomenon as a key geological indicator of the Anthropocene [23] which is an epoch of time defined by the domination of humanity over surface geological processes [24]. Plastic pollution is a serious issue that affects animal and human life. In the sea, for example, it has been reported that over 260 of marine species including mammals, seabirds, turtles and invertebrates become entangled or ingest plastic waste which impair their movement, feeding and reproductive capabilities or cause internal lacerations and ulcers ultimately resulting in death [25].
One of the major problems of plastic pollution is that incomplete degradation of plastics in the environment leads to accumulation of microplastics (particles of less than 5 mm) rather than complete mineralization of the material [26,27]. In humans, microplastics enter the human body through inhalation, ingestion and dermal contact and although more studies related to the human health hazard related to microplastics are needed, some potential hazards include metabolic disorder, inflammation, oxidative stress and multisystem adverse effects (respiratory and digestive) [28,29] as well as potential male and female fertility issues [30]. It can also induce DNA damage and oxidative stress which in turn leads to carcinogenesis [31]. Unfortunately, this threat is now imminent as there has been microplastics found in the marine environment, soil, in drinking water and even in commonly consumed food like fish, vegetables, sugar, honey and salt [32]. Studies also show that these microplastics are indeed present in human blood [33], stool, lungs, placenta, internal organs [34] and in the reproductive system [35].
Some solutions for this problem include recycling, incineration or disposal of plastics in designated landfills. Unfortunately, even though, plastic recycling has existed for decades, scientists estimate that only the 9% is recycled globally, some 12% is incinerated and a greater 79% is either in landfills or in the environment [36]. These numbers are surprising because, in principle, most plastics are recyclable, however there are many factors that represent a barrier towards recycling. For example, contamination of the items in the form of labels, food or mixing of the recycling bins with other products may inhibit recycling entirely. Some plastic items are a complex bend of chemical additives which are harmful for human health which makes recycling dangerous for workers and other items are so unique that they cannot be recycled together [36].
Incineration is a method that could permanently degrade and eliminate plastic waste, nevertheless, the residual ashes from municipal incinerators are still a source of microplastics [37] moreover, the process releases toxic volatile organic compounds to the air. These compounds include chlorinated and aromatics such as benzene and chloroform [38]. Consequently, more environmentally friendly solutions are needed.
3. Degradation of Plastics-A General Perspective
Degradation of plastics can take place because of abiotic and biotic factors present in the environment as shown in Figure 3. When the biotic degradation results in fragments or microplastics, the process is considered bio-disintegration of the polymer, whereas if the polymer is entirely assimilated and mineralized inside the cell, it is considered biodegradation [26]. Alternatively, a polymer is considered as being degraded by abiotic factors when there is any change that may cause depolymerization, change in physical properties, alteration of chemical composition, mass loss or complete mineralization into carbon dioxide and water [39,40].
4. Microbial Degradation of Plastics
One of the most relevant events with regards of microbial degradation of plastics was the discovery of the new bacteria species, Ideonella sakaiensis in 2016 outside a bottle-recycling facility in Japan [43]. This bacterium breaks down PET using two novel enzymes. The first one was labelled as PETase (NCBI accession number A0A0K8P6T7.1) and converts PET to Bis(2-Hydroxyethyl) terephthalate (BHET), TPA and mono(2-hydroxyethyl) terephthalic acid (MHET) which in turn is converted to more terephthalic acid (TPA) and ethylene glycol (EG) monomers by the MHETase enzyme [44]. (NCBI accession number A0A0K8P8E7.1) as shown in Figure 4. After this discovery, dozens of other new PETases have also been identified from several other bacteria. Example: Vibrio gazogenes, Oleispira antarctica, Polyangium brachysporum [45], Marinobacter sp.[46], Ketobacter sp. and Thermobifida [47].
The final PET degradation products, terephthalic acid (TPA) and ethylene glycol (EG), can be both either further metabolized by the cells through the Krebs cycle for biomass accumulation or they can be converted into high value products [48]. TPA can be converted, for example, into vanillic acid, muconic acid, catechol, pyrogallol, gallic acid and adipic acid [48] while ethylene glycol can be separated and used mainly to produce polyester fibres and for antifreeze products [49,50]. BHET can also be used in the industry for making resins, coatings, foams and tissue scaffolds [51] or can be further hydrolysed, inside the cell, into more MHET and TPA by esterase enzymes [52].
The number of research publications reporting plastic-degrading microorganisms keeps increasing every day and by 2020, there had been reported approximately 436 different species [53]. These species include bacteria from the classes Actinobacteria, Firmicutes, Cyanobacteria, Proteobacteria y Bacteroidetes [53] while plastic-degrading fungi are found in eleven classes in the fungal phyla Ascomycota (Dothideomycetes, Eurotiomycetes, Leotiomycetes, Saccharomycetes, and Sordariomycetes), Basidiomycota (Agaricomycetes, Microbotryomycetes, Tremellomycetes, Tritirachiomycetes, and Ustilaginomy-cetes), and Mucoromycota (Mucoromycetes) [54].
To name a few, bacteria such as Cupriavidus necator H16 [55] , Pseudomonas putida LS46, and Pseudomonas putida IRN22 have also been discovered to degrade polyethylene [56] while Pseudomonas putida CA-3 can be fed with styrene to accumulate intracellular polyhydroxyalkanoates [57] and a unique Raoultella sp. DY2415 strain from petroleum-contaminated soil can degrade PE and PS film [58] while the fungus Aspergillus fumigatus, and Phanerochaete chrysosporium degrade a wide range of plastics [53]. The countries that have isolated the most strains are Japan (14.1%) and India (13.8%) [53].
In an effort to compile all this new information, in 2022 the database PlaticDB was created (https://plasticdb.org/). To this date the database contains 753 organisms and 219 proteins that include cutinases, esterases, PETases etc. [59]. Cutinases, specifically, are hydrolases that degrade the cutin which is a component of higher plants cuticle and they have been extensively studied to degrade plastics (PET, PE, PU, Poly (butylene succinate(PBS) and Poly (ε-caprolactone) (PCL)) [60]. They are usually isolated from thermophilic actinomycetes such as Thermobifida fusca (KEGG: Tfu_0882)[61].
However, despite the number of microorganisms and enzymes available, most of them has low activity levels and are not thermostable [43,62]. As a result, efforts have been made to engineer these proteins to increase activity and thermostability. Examples of these enhanced proteins are ThermoPETase, HotPETase, [63,64] , DuraPETase[65] and the novel FAST-PETase (FAST-PETase: functional, active, stable and tolerant PETase) [66] from Ideonella sakaiensis. Moreover, for a more environmentally friendly approach, the native I. sakaiensis PETase has also been successfully expressed in the chloroplast of the microalgae Chlamydomonas reinhardtii [67].
5. Insect Plastic Degradation Order: Lepidoptera (Butterflies and Moths)
Lepidoptera is an order of winged insects and it is the 2nd largest order there is with approximately 180,000 described species [68]. Aside from the wings, the more representative features are the presence of scales and the proboscis (tubular sucking organ) [69]. Larvae of the following insects from this order have been reported to have plastic-degrading capabilities: A. grisella, P. interpunctella, C. cephalonica, S. frugiperda. (Refer to Figure 5)
The Waxworm Galleria Mellonella (Fabricius, 1798) [Lepidoptera: Pyralidae] to Degrade Plastic
Commonly referred as the greater wax moth, Galleria mellonella is a natural honeycomb pest that has contributed to the decline of bee populations at global scale due to the larvae’s capability to feed on wax [75]. In science, these larvae’s importance has gradually increased as a model organism for biomedical studies [76]. They are specially used as an infectious-disease model due to the presence of an immune system that is similar to that of vertebrates [77] (See Figure 6).
To date (June 2024) the PubMed entry “Galleria mellonella to degrade plastic” gives 43 entries from which only 28 are related to the larvae’s capacity to degrade plastic and are summarised here.
Degradation of plastics using the larvae from Galleria mellonella (commonly referred as waxworm) is a fairly novel research topic: The first experiment reporting the capability of this insect to degrade polyethylene (PE) was presented in 2017 when Bombelli, Howe and Bertocchini [9] left worms in a polyethylene bag and observed that they were eating it. The plastic degradation capability of Galleria mellonella has also been found true for other petroleum-based plastics such as expanded polystyrene and polypropylene [78] and for the bio-plastic polylactic acid (PLA).[79] Galleria mellonella is naturally capable of decomposing long-chain hydrocarbons from beeswax without the help of intestinal microorganism using specific carboxylesterases, lipases and fatty-acid metabolism related enzymes. It has been hypothesized that a similar metabolic approach is used to degrade plastic by the waxworm. [80] However, plastic is not nutritious enough as studies show that most larvae (≥50%), growing on an exclusive PE diet lose weight and die in between 3 and 15 days, indicating that supplementary diet is necessary [81,82,83] or the use of older larvae (last developmental stage 25-30 mm) [84] for this type of plastic bioremediation to take place. Pre-treating low density polyethylene under solar radiation for 15 days before feeding the larvae with the material is also being suggested as another technique to increase plastic degradation rate and larvae survival [85].
Polyethylene degradation starts with the expression of salivary enzymes after exposing the larvae to the material [86]. Sanluis-Verdes, Colomer-Vidal, Rodríguez-Ventura, Bello-Villarino, Spinola-Amilibia, Ruiz-López, Illanes-Vicioso, Castroviejo, Cigliano, Montoya, Falabella, Pesquera, González-Legarreta, Arias-Palomo, Solà, Torroba, Arias and Bertocchini [11] discovered and published the first report of 2 novel enzymes isolated from the wax worm saliva with the capability of oxidizing and depolymerizing polyethylene (PE) after only few hours of exposure to the material at room temperature and neutral pH. These enzymes, named as Demetra (NCBI accession number: XP_026756396.1) and Ceres (NCBI accession number: XP_026756459.1) are classified as arylphorin and hexamerin respectively. Gas Chromatography-Mass Spectrometry (GC-MS) was used to confirm the presence of degradation products such as small oxidized aliphatic chains in PE treated with saliva. This discovery opened the way to new ground-breaking solutions for plastic waste management. A closely related protein (81% sequence identity with Demetra) is also described in the study and named as Cibeles. (NCBI accession number XP_026756460.1) Cibeles forms a heterocomplex with Demetra but has not proven to degrade PE on its own [11]. A fourth PE-degrading protein was later identified and named Cora (NCBI accession number XP_026749149.2) late in 2023.[12] The protein 3D structure of all these proteins have been elucidated [12].
Microplastic and plastic depolymerization products are then swallowed and further processed in the gut, where the microbiome plays a key role in plastic degradation [17]: Desulfovibrio vulgaris, Enterobacter sp. D1 [78,87], Acinetobacter [13], the fungus Aspergillus flavus PEDX3 [88] and the fungus Cladosporium halotolerans [89] are examples of microorganisms isolated from the waxworm’s gut with reported capability of degrading PE on experiments in vitro. For the case of A. flavus, for example, microplastics of HDPE were degraded into microplastics with lower molecular weight when exposed to the fungi in liquid culture for 30 days. Chemical changes on the microplastic particles such as the appearance of hydroxyl, carbonyl and ether groups also validate the degradation. Two laccase-like multicopper oxidases enzymes are believed to be responsible for this degradation .[88] Highly similar results were observed when the fungus Cladosporium halotolerans was cultivated in a HDPE microparticle suspension [89].
Likewise, the bacteria Lysinibacillus fusiformis, Bacillus aryabhattai, and Microbacterium oxydans, isolated from the whole worm’s body extract are able to degrade and grow using low-density polyethylene LDPE as carbon source [56].
To add up , other microorganisms from the worm have been studied and proven to be capable of acting over other plastics different from polyethylene (PE). Example: Bacillus cereus can degrade polypropylene (PP) in vitro [90] while mastication of expanded polystyrene (EPS) and polypropylene (PP) increase the abundance of Enterococcus sp. in the gut [78]. Also, the genus Bacillus, Serratia and the bacteria identified as Massilia sp. FS1903 have been associated with polystyrene (PS) degradation [83,91] .Some enzymes, pathways and gut microorganisms mentioned in th is section to degrade plastic by waxworms are summarized in Figure 7 and Figure 8.
Despite of all the above studies, plastic degradation in the gut cannot be solely attributed to microbiota presence. Gut RNA sequencing and biochemical approaches showed that polyethylene-fed larvae show enhanced fatty acid metabolism (FAM) [92,93]. Additionally, early this year 2024, an improved version of the whole genome of Galleria mellonella was published (GenBank: JAPDED000000000.1) [94]. In this study various new putative probable PE-degrading enzymes found are highlighted.
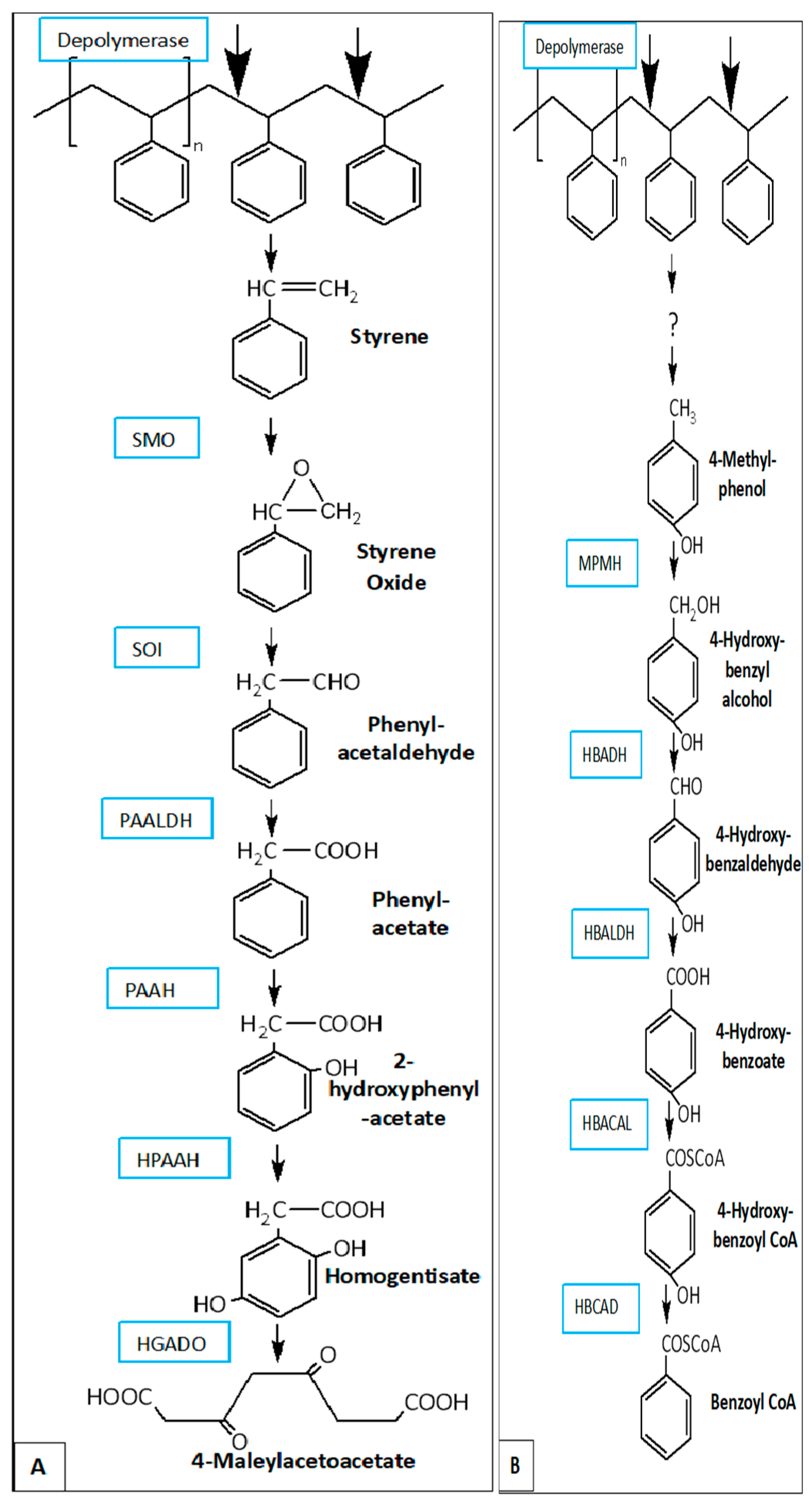
As for the case of polystyrene metabolism a list of possible styrene-degrading enzymes present in the wax worm have been published and two potential metabolic pathways have been proposed [95,96]: the styrene oxide–phenylacetaldehyde [97] pathway that has also expressed in the presence of beeswax [98] (Refer to Figure 8A) and the 4-methylphenol–4- hydroxybenzaldehyde–4-hydroxybenzoate pathway (Figure 8B) which takes place prior to the β-oxidation pathway (main FAM process) in the larvae’s intestines [96].
Figure 8.
Proposed metabolic pathways for PS degradation in G. mellonella. [96] SMO: styrene monooxygenase; SOI: styrene oxide isomerase; PAALDH: phenacetaldehyde dehydrogenase; PAAH: phenylacetate hydroxylase; HPAAH: 2-hydroxyphenylacetate hydroxylase; HGADO: homogentisate 1,2-dioxygenase; MPMH: 4-methylphenol methyl hydroxylase; HBADH: 4-hydroxybenzyl alcohol dehydrogenase; HBALDH: 4-hydroxybenzaldehyde dehydrogenase; HBACAL: 4-hydroxybenzoic acid-CoA ligase; HBCAD: 4-hydroxybenzoyl-CoA reductase[96].
Figure 8.
Proposed metabolic pathways for PS degradation in G. mellonella. [96] SMO: styrene monooxygenase; SOI: styrene oxide isomerase; PAALDH: phenacetaldehyde dehydrogenase; PAAH: phenylacetate hydroxylase; HPAAH: 2-hydroxyphenylacetate hydroxylase; HGADO: homogentisate 1,2-dioxygenase; MPMH: 4-methylphenol methyl hydroxylase; HBADH: 4-hydroxybenzyl alcohol dehydrogenase; HBALDH: 4-hydroxybenzaldehyde dehydrogenase; HBACAL: 4-hydroxybenzoic acid-CoA ligase; HBCAD: 4-hydroxybenzoyl-CoA reductase[96].
6. Insect Plastic Degradation Order: Coleoptera (Beetles and Weevils) [99]
The order Coleoptera represents the largest group of insects with 40% of known insect species. In this group, generally, the wings develop internally but some have no wings [99]. Plastivore Coleoptera larvae include the larvae from the beetles Alphitobius diaperinus, Plesiophthalmus davidis, Tribolium castaneum and Uloma. The type of plastic and microbiome associated with these insects are shown in Figure 9.
The Yellow Mealworm Tenebrio Molitor (Linnaeus, 1758) [Coleoptera: Tenebrionidae] to Degrade Plastic
Commonly referred to as mealworm, Tenebrio molitor’s larvae is commonly used as protein source for domestic animals (monogastric animal feed) [104] , for fish [105] and can also be grown for human consumption [106,107]. In science, it has been studied as a model for cellular and humoral immunity against pathogenic infections [108]. A representation of an adult beetle is shown in Figure 10.
Starting from 2015, to date (June 2024) the entry in PubMed “Tenebrio to degrade plastic” gives 82 entries from which 67 are related to the larvae’s capacity to degrade plastic. Most of those papers were used for this review.
The mealworms, at the moment, are growing a reputation as polystyrene (PS) plastic eaters [8,83,93,110,111,112,113,114,115,116]. This larvae is capable of converting ≈47% of the ingested Styrofoam (a common PS product) into CO2 and the residue (≈49%) is excreted as fecula with a limited fraction incorporated into biomass [8]. Although Galleria mellonella, can degrade PS, [117] the mealworms lose their capacity to degrade PS plastic when the bacteria gut is inhibited [115,118] which suggest a strong dependency on microbiota to degrade PS. Even so, it has also been demonstrated that the mealworm secretes emulsifying factors that increase plastic bioavailability in the gut [113] as well as a wide range of oxidases, cytochrome P450, monooxygenase, superoxidase, and dehydrogenase and other enzymes related to the fatty acid metabolism [119,120].
Mealworms of approximately 3-4 instars (20–25 mm in length) fed only with PS are able to survive and complete their entire life cycle and emerge into adult beetles [8,116,120]. This could partially be explained by the fact that gut microbiome, specially from the genus Klebsiella, is capable of nitrogen fixation, thus the worm is supplied with this element as well [121]. Still, it is recommended to supplement with corn flour (T. obscurus ) or wheat bran (T. molitor), sucrose and hydrate with H2O [122] to increase the PS degradation rate and enable breeding of a second generation with favourable capabilities for PS degradation as well [114,116,123,124]. It has been hypothesized that the mechanism used to degrade PS is similar to the mechanism described for Galleria mellonella in Figure 8A in which PS is degraded into styrene first [93] and after several intermediate steps the benzene ring is destroyed [117] and assimilated through the β-oxidation pathway [96].
Additionally, T. molitor larvae can also biodegrade polyethylene (PE) through the mechanism presented in Figure 11 [112,125,126] . They can also degrade PP, PVC [127,128,129], Nylon 11 Polymers[130,131], Polyethylene terephthalate (PET) [119] , melamine formaldehyde (MF)[132] and the biopolymer PLA but the mechanism for this polymers remains unknown [133]. T. molitor can even chew and ingest polyurethane (PU) but the digestion/degradation of this plastic has not been demonstrated [134,135,136]. During the COVID-19 pandemic, polypropylene (PP) face mask production and contamination increased considerably and it was observed that T. molitor can consume face masks.[137] The capability of biodegradation can be affected by the molecular weight, branching and crystallinity of the material [138].
The mealworm’s gut bacteria Exiguobacterium sp. strain YT2 degrades polystyrene (PS) [115]. Citrobacter sp. and Kosakonia sp. were also found in the gut and strongly relate with PE and PS consumption [112]as well as the bacteria Priestia megaterium S1 [139]. Other strains related to PS degradation include Erwinia olea, Lactococcus lactis, Lactococcus garviae [122] Serratia marcescens, Pseudomonas aeruginosa, Acinetobacter septicus, Agrobacterium tumefaciens, Klebsiella grimontii, Pseudomonas multiresinivorans, Pseudomonas nitroreducens, Pseudomonas plecoglossicida, and Yokenella regensburgei [110,111,113,140]. Bacteria from the families Enterobacteriaceae, such as Enterobacter hormaechei LG3 [141]), the families Spiroplasmataceae, Enterococcaceae, [114,117] Staphylococcus and Rhodococcus [120] also play a role in PS degradation. It is important to note that a study, where mealworms from 3 different regions in China were compared, showed that larvae from different regions have different metabolism [142] which suggests that the gut microbiota can change depending on the environment of the larvae, diet and even depending on the PS molecular weight provided [143], yet PS consumption is ubiquitous to this species. [144]
Additionally, the family Enterobacteriaceae has also been linked with polyurethane (PU) degradation along with the family Hafnia [145] while the genus’s Spiroplasma, Dysgonomonas and Hafnia-Obesumbacterium are associated with PET degradation [119]. Gut bacteria from Tenebrio molitor is even capable of degrading vulcanized poly(cis-1,4-isoprene) rubber (vPR) (strain Acinetobacter sp. BIT-H3) [146]. It is also capable of degrading PVC and PP but the genre of these microorganisms has not been elucidated [127,128] . Tenebrio molitor’s microbiome related to plastic-degradation is illustrated in Figure 12.
Interestingly enough, Although less studied, a comparison between the yellow mealworms (T. molitor) and the dark ones (T. obscurus) show that the latter degrade PS at higher rates [114]. T. obscurus larvae also degrades LDPE using gut bacteria mainly from the genus Spiroplasma and Enterococcus [147,148].
The Superworm Zophobas Atratus (Fabricius, 1776) [Coleoptera: Tenebrionidae] to Degrade Plastic
Zophobas morio (Fabricius, 1776) is a dark beetle [Coleoptera: Tenebrionidae] that is currently considered as the same species as Zophobas atratus. It was also previously identified as Tenebrio morio and/or Helops morio and commonly referred as the giant mealworm beetle which has been a matter of confusion and controversy [149]. (Figure 13) Zophobas larvae is commonly refer as the superworm [149]. This larvae is highly nutritious and it is promising as fish, poultry and pig feed [149].
To date (June 2024) the entry in PubMed “Zophobas atratus to degrade plastic” gives 17 entries from which all 17 are related to the larvae’s capacity to degrade plastic. All 17 studies were used for this review.
Zophobas is also a main polystyrene (PS) consumer. A comparison in a 30 days long experiment, between the larvae of Tenebrio molitor (yellow mealworm), Galleria mellonella (greater wax moth) and Zophobas atratus (superworm) showed that the latter has the strongest polystyrene consumption capacity and the highest survival rate of the three [117] being able to consume 4 times more PS than the yellow mealworm per day [150]. Superworms also outdid the yellow mealworms by 11 folds on PU consumption in another study [145]. But, similarly to Tenebrio, this capability is lost when the gut microbiota is supressed using antibiotics [150].
Moreover, new research indicates that the superworm’s microbiota is also capable of degrading PE, PP, PVC and PET[151,152,153,154] and even polyurethane (PU) [155], melamine formaldehyde (MF)[132], ethylene vinyl acetate (EVA)[156] and polybutylene succinate (PBS)[157].
PE and PS degradability is being attributed to Pseudomonas aeruginosa [152,158] and Enterococcus (also associated with PU degradation)[155], Citrobacter with PE and PVC [153], Brevibacterium [159] and Dysgonomonas and Sphingobacterium with PS and Mangrovibacter with PU degradation [155] (Summarized in Figure 14).
Little is known about the mechanisms these specific larvae use to degrade plastic but PS degradation seems to be partly achieved by the synergistic effect of the generation of reactive oxygen species (ROS) inside the gut and the production of oxidases and other enzymes by the microbiome [160].
PS was achieved by the synergistic effect of ROS and complex functional microbes and enzymes as well as other enhancing factors in the gut of larvae.
both compounds [80].
Insect plastic degradation. Order: Blattodea (cockroaches and termites)[161]
Termites have a physical appearance that is similar to ants as observed in Figure 15. They used to be classified in their own order named Isoptera. However, new studies show that they are actually closely related to cockroaches and should be classified in the same order Blattodea [161]. The most characteristic feature of these insects is that they feed on wood, which is composed mainly of polymers of cellulose, hemicellulose and lignin [162,163]. Lignocellulose and plastic polymers have similar physicochemical features, for example, their carbon chains have similar chemical bonds and hydrophobicity properties which has led to believe that termites could also be plastivore organisms [164]. The relationship between lignocellulose and plastic consumption is further supporter by the fact that, as mentioned before, some enzymes like cutinases which are involved in degrading plants are very well documented plastic degraders [60,61].
In fact, higher adult termites (Nasutitermes nigriceps) have been observed and reported degrading wood-HDPE plastic composites (WPCs) in one study from Yucatan, Mexico [165]. In a later study, a group of previously isolated yeast symbionts from the guts of the adult Coptotermes formosanus (termite) showed low density PE degrading capabilities [14,166]. Termites are the first example of adult insects degrading plastic as all the previously reported degradation has been reported on larvae.
Other wood-eating insects examples include the pest Chrysobothris sp. which is a beetle that attacks cedar trees, [167] the emerald ash borer (Agrilus planipennis) which attacks ash trees, [168] the red bay ambrosia beetle (Xyleborus glabratus) that attacks laurel trees, [169] and the Asian long horned beetle (Anoplophora glabripennis) that attack maple trees. [170] in between others woodboring beetles [171].
Other Organisms from the Class Insecta to Degrade Plastic
The list of plastivores in this review includes all the insects mentioned in previews reviews [3] [16,172] from the orders Lepidoptera and Coleoptera and expands to the order Blattodea that could be further explored in the future for plastic degradation. It was confirmed in 2023 that 3 novel yeasts isolated from termite’s gut (Blattodea) can metabolise polyethylene (PE) into alkanes, aldehydes, ethanol, and fatty acids [14].
Interestingly, similar to Galleria mellonella, the larvae from Achroia grisella and from Uloma feed on the long-chain hydrocarbon beeswax and can degrade the plastics PE and PS (pre-print study) [173]. The larvae from Plodia interpunctella is another larvae that eats both beeswax and PE [6]. The positive relationship between capability for eating beeswax and capability for eating plastic might not be a coincidental one but rather a case of cause and consequence since, as mentioned before in this document, it has been suggested that similar metabolic approaches are used to degrade both compounds [80].
The number of plastivore insects is growing every day and in the latest 2024 review [174] the authors suggested a large variety of insects with potential plastic-degrading capabilities which include insects from the order Diptera (Example: Black soldier fly), Blattodea (several type of cockroaches) and Orthoptera (Ex. The cricket Gryllodes sigillatus) as well as other families within the already studied order Coleoptera (Example: the lesser grain borer or Rhyzopertha dominica ,the rice weevil or Sitophilus oryzae, and cigar beetle or Lasioderma serricorne) and Lepidoptera (The larvae of Hofmannophila pseudospretella or the Brown House moth). However, scientific studies are needed to confirm whether there is actual degradation by these species and which microorganisms/enzymes could be responsible. Another recent review estimated that over 23 species of insects (including the 12 insects described in this review) have been observed consuming plastics [175] and the list could expand even further in the future. If a positive relationship between wax degradation and plastic degradation is confirmed then other wax-eating larvae could be explored. An example of this type of larvae would be the American wax worm Vitula edmandsii which is a honeybee comb pest [176]. Similarly, other woodboring beetles or any other xylophagous (wood diet) larvae or insect could be a good candidate for research.
Analysis of Plastic Degradation After Exposure to Insect Larvae
Several techniques and protocols are available for characterization of degraded polymer after exposure to larvae. The simplest method is to measure plastic weight loss over time. [70] Afterwards, the plastic polymer can be closely inspected for changes in the surface morphology using scanning electron microscopy (SEM)[101]. To check for chemical modifications one of several options is the analysis using X-Ray photoelectron spectroscopy (XPS)[101].
Once the plastic has been digested, frass can be collected and analysed using different techniques. For example, Brandon, Gao, Tian, Ning, Yang, Zhou, Wu and Criddle [112] measured the molecular weight of the polymer using gel permeation chromatography (HT-GPC) followed by characterization of the degradation products through magnetic resonance (1H NMR) and Fourier Transform Infrared Spectroscopy (FTIR). Thermal gravimetric analysis (TGA) can also be used to analyse thermal changes from plastic to frass.
In order to isolate plastic-degrading microorganisms, faecal matter collected has to be diluted and plated in Tryptic soy agar (TSA) and in a defined media as described in literature [131] to obtain isolated colonies. To confirm the capability of these colonies to degrade plastic several tests can be performed such as the clear zone assay in agar plate and turbidity measurements in liquid culture. [139] Gas Chromatography-Mass Spectometry (GC-MS), which is an analytical technique used to identify and quantify compounds, can also be used to confirm the presence of PE degradation products in liquid culture such as small oxidized aliphatic chains. [11]
Challenges and Future Perspectives
Even though plastivore insects are a new exciting avenue for bioremediation of plastic pollution, several challenges need to be overcome before the technology can be industrialized.
One of these challenges is to provide optimal, standardized conditions for larval rearing at the industrial scale to ensure reproducible results in terms of larvae quality (weight, survival rate lipid content etc.) and plastic degradation rate. Environmental conditions such as light exposure, temperature and ventilation greatly affect the development of the larvae. In the case of Galleria mellonella, for example, constant exposure to light significantly reduces its size and delays metamorphosis, so they need to be grown in darkness at a temperature of 28–32°C. Providing ventilation is also highly important not only to provide oxygen but to prevent infections too [177]. Therefore, for the use of live larvae for plastic degradation, we consider essential to have a contained and controlled area (a dark greenhouse for example) with controlled conditions. The other indispensable benefit of the use of an enclosed area is the responsible containment of insects that are recognized as pests.
Another problematic source of variability at industrial scale is the chemical composition and properties of the waste plastic used as feedstock. Evidence shows that the presence of other contaminants present in plastic pollution such as plastic additives, in between other factors, may affect larvae’s digestion [178]. This challenge could be overcome by processing the plastic waste prior to feeding the larvae as it is normally processed for plastic recycling: 1) Sorting and categorizing: In this step several types of plastics need to be separated from each other, 2) Washing: Impurities that can be toxic for the larvae are removed , 3) Shredding: Plastic is broken down into much smaller pieces and 4) Testing: At this point, the plastic pieces are tested for their quality and density [179]. Additionally, other novel steps shall be explored, for example, (as mentioned previously) pre-treating PE under solar radiation for 15 days before feeding the larvae increases plastic degradation rate and larvae survival [85].
There are also concerns raised about the economic feasibility of the technology due to the high cost of breeding the larvae and the lack of sufficient research to obtain high-value end products. It has been calculated that it would cost more than €300 to degrade 1 ton of low LDPE plastic using ≥ 4 tons of waxworms or mealworms in approximately 38 days [180] while recycling 1 ton of LDPE costs less than €250 in less time [181]. In this regard, due to their high fat content, the waxworm, the yellow mealworm and even PE-plastivore larvae from Corcyra cephalonica (up to 60%, 38% and 43.3% respectively) could potentially be used for biodiesel production [182,183]. Another solution suggested is the extraction of chitin from adult plastic-fed Tenebrio molitor’s exoskeleton [184] which can then be processed to use as biomedical materials, food additives, cosmetic ingredients, agricultural materials, analytical reagents and others [185]. Larvae could also, potentially, be used as animal feed as some studies show that there are no microplastic nor nano plastic residues present in the frass as a result of plastic consumption by larvae [186]. However more studies are needed to corroborate safety.
On the other hand, it is important to note that also this same study [180] calculated that the process to degrade 1 ton of plastic using larvae would also release ≥ 4 tons of CO2 to the atmosphere which is a much larger number than the ≈2.9 tons of CO2 that would be produced during plastic incineration [187]. This information is worrying and lead us to wonder if this technology can be used as a sustainable process. In future the process will need a more comprehensive Life Cycle Assessment [188] and the possible co- implementation of a CO2 capture system.
Lastly, scale up standardization and the production of high-value end products could be achieved using other , more advanced, biotechnological approaches such as the recombinant expression of insect-derived novel enzymes or the use of gut-isolated microorganisms to degrade plastics in cell culture. Insect cells specifically, are already successfully used as factories for biomanufacturing of several proteins, vaccines and vectors for gene therapy [189] while the industrial cultivation of microorganisms to obtain biotechnological products has been practiced for thousands of years, starting from the production of wine, beer and bread [190]. Cell culture plastic degradation would allow the recovery of plastic monomers that can be converted into high-value components. For example, as mentioned above, TPA monomers from PET degradation can be transformed into vanillin [191] which is considered the second most important flavouring agent, after saffron, and has a wide variety of applications in the food and beverage industry but also in the pharmaceutical industry and for production of home-use products such as perfumes and deodorants [192].
Author Contributions
Isabel Vital-Vilchis wrote the manuscript, Esther Karunakaran supervised and reviewed the document.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nikiema, J. ;Asiedu, Z. A review of the cost and effectiveness of solutions to address plastic pollution, Environmental Science and Pollution Research, 2022 29, 17: 24547-24573. [CrossRef]
- Bergeson, A. R.;Silvera, A. J.;Alper, H. S. Bottlenecks in biobased approaches to plastic degradation, Nature Communications, 2024 15, 1: 4715. [CrossRef]
- Pivato, A. F. et al. Hydrocarbon-based plastics: Progress and perspectives on consumption and biodegradation by insect larvae, Chemosphere, 2022 293: 133600. [CrossRef]
- Essig, E. O.;Hoskins, W. M.;Linsley, E. G.;Micrelbacher, A. E.;Smith, R. F. A Report on the Penetration of Packaging Materials by Insects, Journal of Economic Entomology, 1943 36, 6: 822-829. [CrossRef]
- Gerhardt, P. D. ;Lindgren, D. L. Penetration of packaging films: Film materials used for food packaging tested for resistance to some common stored-product insects, Hilgardia, 1954 8, 6: 3-4.
- Yang, Y.;Chen, J.;Wu, W.-M.;Zhao, J.;Yang, J. Complete genome sequence of Bacillus sp. YP1, a polyethylene-degrading bacterium from waxworm's gut, Journal of Biotechnology, 2015 200: 77-78. [CrossRef]
- Yang, J.;Yang, Y.;Wu, W.-M.;Zhao, J.;Jiang, L. Evidence of Polyethylene Biodegradation by Bacterial Strains from the Guts of Plastic-Eating Waxworms, Environmental Science & Technology, 2014 48, 23: 13776-13784. [CrossRef]
- Yang, Y. et al. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 1. Chemical and Physical Characterization and Isotopic Tests, Environmental Science & Technology, 2015 49, 20: 12080-12086. [CrossRef]
- Bombelli, P.;Howe, C. J.;Bertocchini, F. Polyethylene bio-degradation by caterpillars of the wax moth <em>Galleria mellonella</em>, Current Biology, 2017 27, 8: R292-R293. [CrossRef]
- Arnold, C. This Bug Can Eat Plastic. But Can It Clean Up Our Mess?, National Geographic, 2017, https://www.nationalgeographic.com/science/article/wax-worms-eat-plastic-polyethylene-trash-pollution-cleanup.
- Sanluis-Verdes, A. et al. Wax worm saliva and the enzymes therein are the key to polyethylene degradation by <em>Galleria mellonella</em>, bioRxiv, 2022: 2022.04.08.487620. [CrossRef]
- Spínola-Amilibia, M. et al. Plastic degradation by insect hexamerins: Near-atomic resolution structures of the polyethylene-degrading proteins from the wax worm saliva, Science Advances, 2023 9, 38: eadi6813. [CrossRef]
- Cassone, B. J.;Grove, H. C.;Elebute, O.;Villanueva, S. M. P.;LeMoine, C. M. R. Role of the intestinal microbiome in low-density polyethylene degradation by caterpillar larvae of the greater wax moth, Galleria mellonella, Proceedings of the Royal Society B: Biological Sciences, 2020 287, 1922: 20200112. [CrossRef]
- Elsamahy, T.;Sun, J.;Elsilk, S. E.;Ali, S. S. Biodegradation of low-density polyethylene plastic waste by a constructed tri-culture yeast consortium from wood-feeding termite: Degradation mechanism and pathway, Journal of Hazardous Materials, 2023 448: 130944. [CrossRef]
- Sanchez-Hernandez, J. C. A toxicological perspective of plastic biodegradation by insect larvae, Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2021 248: 109117. [CrossRef]
- Siddiqui, S. A.;Abdul Manap, A. S.;Kolobe, S. D.;Monnye, M.;Yudhistira, B.;Fernando, I. Insects for plastic biodegradation – A review, Process Safety and Environmental Protection, 2024 186: 833-849. [CrossRef]
- An, R.;Liu, C.;Wang, J.;Jia, P. Recent Advances in Degradation of Polymer Plastics by Insects Inhabiting Microorganisms, Polymers, 2023 15, 5. [CrossRef]
- Geyer, R.;Jambeck, J. R.;Law, K. L. Production, use, and fate of all plastics ever made, Science Advances, 3, 7: e1700782. [CrossRef]
- Mohanan, N.;Montazer, Z.;Sharma, P. K.;Levin, D. B. Microbial and Enzymatic Degradation of Synthetic Plastics, (in English), Frontiers in Microbiology, 2020Review 11. [CrossRef]
- Telmo, O., "Polymers and the Environment," in Polymer Science, Y. Faris Ed. Rijeka: IntechOpen, 2013, p. Ch. 1.
- PlasticsEurope. "Plastics - the Facts 2019: an Analysis of European Plastics Production, Demand and Waste Data." (accessed 28 June, 2024).
- Cregut, M.;Bedas, M.;Durand, M. J.;Thouand, G. New insights into polyurethane biodegradation and realistic prospects for the development of a sustainable waste recycling process, Biotechnology Advances, 2013 31, 8: 1634-1647. [CrossRef]
- Zalasiewicz, J. et al. The geological cycle of plastics and their use as a stratigraphic indicator of the Anthropocene, Anthropocene, 2016 13: 4-17. [CrossRef]
- Crutzen, P. J., "The “Anthropocene”," in Earth System Science in the Anthropocene, E. Ehlers and T. Krafft Eds. Berlin, Heidelberg: Springer Berlin Heidelberg, 2006, pp. 13-18.
- Laist, D. W., "Impacts of Marine Debris: Entanglement of Marine Life in Marine Debris Including a Comprehensive List of Species with Entanglement and Ingestion Records," in Marine Debris: Sources, Impacts, and Solutions, J. M. Coe and D. B. Rogers Eds. New York, NY: Springer New York, 1997, pp. 99-139.
- Zhang, K. et al. Understanding plastic degradation and microplastic formation in the environment: A review, Environmental Pollution, 2021 274: 116554. [CrossRef]
- Wu, P. et al. Environmental occurrences, fate, and impacts of microplastics, Ecotoxicology and Environmental Safety, 2019 184: 109612. [CrossRef]
- Vethaak, A. D. ;Legler, J. Microplastics and human health, Science, 2021 371, 6530: 672-674. [CrossRef]
- Zhao, B.;Rehati, P.;Yang, Z.;Cai, Z.;Guo, C.;Li, Y. The potential toxicity of microplastics on human health, Science of The Total Environment, 2024 912: 168946. [CrossRef]
- Zurub, R. E.;Cariaco, Y.;Wade, M. G.;Bainbridge, S. A. Microplastics exposure: implications for human fertility, pregnancy and child health, (in English), Frontiers in Endocrinology, 2024Review 14. [CrossRef]
- Kumar, R. et al. Micro(nano)plastics pollution and human health: How plastics can induce carcinogenesis to humans?, Chemosphere, 2022 298: 134267. [CrossRef]
- Ziani, K. et al., "Microplastics: A Real Global Threat for Environment and Food Safety: A State of the Art Review," Nutrients, vol. 15, no. 3. [CrossRef]
- Leslie, H. A.;van Velzen, M. J. M.;Brandsma, S. H.;Vethaak, A. D.;Garcia-Vallejo, J. J.;Lamoree, M. H. Discovery and quantification of plastic particle pollution in human blood, Environment International, 2022 163: 107199. [CrossRef]
- Yang, Y. et al. Detection of Various Microplastics in Patients Undergoing Cardiac Surgery, Environmental Science & Technology, 2023 57, 30: 10911-10918. [CrossRef]
- Hong, Y.;Wu, S.;Wei, G. Adverse effects of microplastics and nanoplastics on the reproductive system: A comprehensive review of fertility and potential harmful interactions, Science of The Total Environment, 2023 903: 166258. [CrossRef]
- Programme, U. N. D. "United Nations Development Programme .Why aren't we recycling more plastic?" United Nations Development Programme https://stories.undp.org/why-arent-we-recycling-more-plastic#:~:text=Recycling%20rates%20vary%20by%20location,Some%2012%20percent%20is%20incinerated. (accessed November 28th, 2023).
- Yang, Z. et al. Is incineration the terminator of plastics and microplastics?, Journal of Hazardous Materials, 2021 401: 123429. [CrossRef]
- Jang, M. et al. Analysis of volatile organic compounds produced during incineration of non-degradable and biodegradable plastics, Chemosphere, 2022 303: 134946. [CrossRef]
- Shah, A. A.;Hasan, F.;Hameed, A.;Ahmed, S. Biological degradation of plastics: A comprehensive review, Biotechnology Advances, 2008 26, 3: 246-265. [CrossRef]
- Chamas, A. et al. Degradation Rates of Plastics in the Environment, ACS Sustainable Chemistry & Engineering, 2020 8, 9: 3494-3511. [CrossRef]
- Fairbrother, A. et al. Temperature and light intensity effects on photodegradation of high-density polyethylene, Polymer Degradation and Stability, 2019 165: 153-160. [CrossRef]
- Yousif, E. ;Haddad, R. Photodegradation and photostabilization of polymers, especially polystyrene: review, SpringerPlus, 2013 2, 1: 398. [CrossRef]
- Yoshida, S. et al. A bacterium that degrades and assimilates poly(ethylene terephthalate), Science, 2016 351, 6278: 1196-1199. [CrossRef]
- Austin, H. P. et al. Characterization and engineering of a plastic-degrading aromatic polyesterase, Proceedings of the National Academy of Sciences, 2018 115, 19: E4350-E4357. [CrossRef]
- Danso, D. et al. New Insights into the Function and Global Distribution of Polyethylene Terephthalate (PET)-Degrading Bacteria and Enzymes in Marine and Terrestrial Metagenomes, Applied and Environmental Microbiology, 2018 84, 8: e02773-17. [CrossRef]
- Meyer Cifuentes, I. E. et al. Molecular and Biochemical Differences of the Tandem and Cold-Adapted PET Hydrolases Ple628 and Ple629, Isolated From a Marine Microbial Consortium, (in English), Frontiers in Bioengineering and Biotechnology, 2022Original Research 10. [CrossRef]
- Erickson, E. et al. Sourcing thermotolerant poly(ethylene terephthalate) hydrolase scaffolds from natural diversity, Nature Communications, 2022 13, 1: 7850. [CrossRef]
- Qi, X.;Yan, W.;Cao, Z.;Ding, M.;Yuan, Y., "Current Advances in the Biodegradation and Bioconversion of Polyethylene Terephthalate," Microorganisms, vol. 10, no. 1. [CrossRef]
- Hollis, J. M.;Lovas, F. J.;Jewell, P. R.;Coudert, L. H. Interstellar Antifreeze: Ethylene Glycol, The Astrophysical Journal, 2002 571, 1: L59. [CrossRef]
- Hemmat Esfe, M.;Saedodin, S.;Mahian, O.;Wongwises, S. Efficiency of ferromagnetic nanoparticles suspended in ethylene glycol for applications in energy devices: Effects of particle size, temperature, and concentration, International Communications in Heat and Mass Transfer, 2014 58: 138-146. [CrossRef]
- Westover, C. C. ;Long, T. E., "Envisioning a BHET Economy: Adding Value to PET Waste," Sustainable Chemistry, vol. 4, no. 4, pp. 363-393. [CrossRef]
- Qiu, L.;Yin, X.;Liu, T.;Zhang, H.;Chen, G.;Wu, S. Biodegradation of bis(2-hydroxyethyl) terephthalate by a newly isolated Enterobacter sp. HY1 and characterization of its esterase properties, Journal of Basic Microbiology, 2020 60, 8: 699-711. [CrossRef]
- Gambarini, V.;Pantos, O.;Kingsbury Joanne, M.;Weaver, L.;Handley Kim, M.;Lear, G. Phylogenetic Distribution of Plastic-Degrading Microorganisms, mSystems, 2021 6, 1. [CrossRef]
- Ekanayaka, A. H. et al., "A Review of the Fungi That Degrade Plastic," Journal of Fungi, vol. 8, no. 8. [CrossRef]
- Montazer, Z.;Habibi Najafi, M. B.;Levin, D. B. Microbial degradation of low-density polyethylene and synthesis of polyhydroxyalkanoate polymers, Canadian Journal of Microbiology, 2018 65, 3: 224-234. [CrossRef]
- Montazer, Z.;Habibi Najafi, M. B.;Levin, D. B. In vitro degradation of low-density polyethylene by new bacteria from larvae of the greater wax moth, Galleria mellonella, Canadian Journal of Microbiology, 2020 67, 3: 249-258. [CrossRef]
- O'Leary Niall, D.;O'Connor Kevin, E.;Ward, P.;Goff, M.;Dobson Alan, D. W. Genetic Characterization of Accumulation of Polyhydroxyalkanoate from Styrene in Pseudomonas putida CA-3, Applied and Environmental Microbiology, 2005 71, 8: 4380-4387. [CrossRef]
- Yuan, Y. et al. Unique Raoultella species isolated from petroleum contaminated soil degrades polystyrene and polyethylene, Ecotoxicology and Environmental Safety, 2023 263: 115232. [CrossRef]
- Gambarini, V.;Pantos, O.;Kingsbury, J. M.;Weaver, L.;Handley, K. M.;Lear, G. PlasticDB: a database of microorganisms and proteins linked to plastic biodegradation, Database, 2022 2022: baac008. [CrossRef]
- Sahu, S.;Kaur, A.;Khatri, M.;Singh, G.;Arya, S. K. A review on cutinases enzyme in degradation of microplastics, Journal of Environmental Management, 2023 347: 119193. [CrossRef]
- Oda, M.;Numoto, N.;Bekker, G.-J.;Kamiya, N.;Kawai, F., "Chapter Eight - Cutinases from thermophilic bacteria (actinomycetes): From identification to functional and structural characterization," in Methods in Enzymology, vol. 648, G. Weber, U. T. Bornscheuer, and R. Wei Eds.: Academic Press, 2021, pp. 159-185.
- Müller, R.-J.;Schrader, H.;Profe, J.;Dresler, K.;Deckwer, W.-D. Enzymatic Degradation of Poly(ethylene terephthalate): Rapid Hydrolyse using a Hydrolase from T. fusca, Macromolecular Rapid Communications, 2005 26, 17: 1400-1405. [CrossRef]
- Son, H. F. et al. Rational Protein Engineering of Thermo-Stable PETase from Ideonella sakaiensis for Highly Efficient PET Degradation, ACS Catalysis, 2019 9, 4: 3519-3526. [CrossRef]
- Bell, E. L. et al. Directed evolution of an efficient and thermostable PET depolymerase, Nature Catalysis, 2022 5, 8: 673-681. [CrossRef]
- Cui, Y. et al. Computational Redesign of a PETase for Plastic Biodegradation under Ambient Condition by the GRAPE Strategy, ACS Catalysis, 2021 11, 3: 1340-1350. [CrossRef]
- Lu, H. et al. Machine learning-aided engineering of hydrolases for PET depolymerization, Nature, 2022 604, 7907: 662-667. [CrossRef]
- Di Rocco, G. et al. A PETase enzyme synthesised in the chloroplast of the microalga Chlamydomonas reinhardtii is active against post-consumer plastics, Scientific Reports, 2023 13, 1: 10028. [CrossRef]
- Mallet, J. "Taxonomy of Lepidoptera: the scale of the problem." University College. https://www.ucl.ac.uk/taxome/lepnos.html (accessed.
- Powell, J. A., "Chapter 151 - Lepidoptera: Moths, Butterflies," in Encyclopedia of Insects (Second Edition), V. H. Resh and R. T. Cardé Eds. San Diego: Academic Press, 2009, pp. 559-587.
- Kundungal, H.;Gangarapu, M.;Sarangapani, S.;Patchaiyappan, A.;Devipriya, S. P. Efficient biodegradation of polyethylene (HDPE) waste by the plastic-eating lesser waxworm (Achroia grisella), Environmental Science and Pollution Research, 2019 26, 18: 18509-18519. [CrossRef]
- Ali, S. S.;Elsamahy, T.;Zhu, D.;Sun, J. Biodegradability of polyethylene by efficient bacteria from the guts of plastic-eating waxworms and investigation of its degradation mechanism, Journal of Hazardous Materials, 2023 443: 130287. [CrossRef]
- Lou, H. et al. Biodegradation of polyethylene by Meyerozyma guilliermondii and Serratia marcescens isolated from the gut of waxworms (larvae of Plodia interpunctella), Science of The Total Environment, 2022 853: 158604. [CrossRef]
- Kesti, S. S. ;Thimmappa*, S. C. First report on biodegradation of low density polyethylene by rice moth larvae, Corcyra cephalonica Holistic Approach Environmental 2019 9, 4: 79-83. [CrossRef]
- Zhang, Z.;Peng, H.;Yang, D.;Zhang, G.;Zhang, J.;Ju, F. Polyvinyl chloride degradation by a bacterium isolated from the gut of insect larvae, Nature Communications, 2022 13, 1: 5360. [CrossRef]
- Kwadha, C. A.;Ong’amo, G. O.;Ndegwa, P. N.;Raina, S. K.;Fombong, A. T., "The Biology and Control of the Greater Wax Moth, Galleria mellonella," Insects, vol. 8, no. 2. [CrossRef]
- Mikulak, E.;Gliniewicz, A.;Przygodzka, M.;Solecka, J. Galleria mellonella L. as model organism used in biomedical and other studies, Przegląd Epidemiologiczny - Epidemiological Review, 2018journal article 72, 1: 57-73. [Online]. Available: https://www.przeglepidemiol.pzh.gov.pl/Galleria-mellonella-L-as-model-organism-used-in-biomedical-and-other-studies,180812,0,2.html.
- Asai, M.;Li, Y.;Newton, S. M.;Robertson, B. D.;Langford, P. R. Galleria mellonella–intracellular bacteria pathogen infection models: the ins and outs, FEMS Microbiology Reviews, 2023 47, 2: fuad011. [CrossRef]
- Peydaei, A.;Bagheri, H.;Gurevich, L.;de Jonge, N.;Nielsen, J. L. Mastication of polyolefins alters the microbial composition in Galleria mellonella, Environmental Pollution, 2021 280: 116877. [CrossRef]
- Shah, R. et al. Exposure to polylactic acid induces oxidative stress and reduces the ceramide levels in larvae of greater wax moth (Galleria mellonella), Environmental Research, 2023 220: 115137. [CrossRef]
- Kong, H. G. et al. The <em>Galleria mellonella</em> Hologenome Supports Microbiota-Independent Metabolism of Long-Chain Hydrocarbon Beeswax, Cell Reports, 2019 26, 9: 2451-2464.e5. [CrossRef]
- Réjasse, A.;Waeytens, J.;Deniset-Besseau, A.;Crapart, N.;Nielsen-Leroux, C.;Sandt, C. Plastic biodegradation: Do Galleria mellonella Larvae Bioassimilate Polyethylene? A Spectral Histology Approach Using Isotopic Labeling and Infrared Microspectroscopy, Environmental Science & Technology, 2022 56, 1: 525-534. [CrossRef]
- Cassone, B. J.;Grove, H. C.;Kurchaba, N.;Geronimo, P.;LeMoine, C. M. R. Fat on plastic: Metabolic consequences of an LDPE diet in the fat body of the greater wax moth larvae (Galleria mellonella), Journal of Hazardous Materials, 2022 425: 127862. [CrossRef]
- Lou, Y. et al. Biodegradation of Polyethylene and Polystyrene by Greater Wax Moth Larvae (Galleria mellonella L.) and the Effect of Co-diet Supplementation on the Core Gut Microbiome, Environmental Science & Technology, 2020 54, 5: 2821-2831. [CrossRef]
- Ruiz Barrionuevo, J. M. et al. Consumption of low-density polyethylene, polypropylene, and polystyrene materials by larvae of the greater wax moth, Galleria mellonella L. (Lepidoptera, Pyralidae), impacts on their ontogeny, Environmental Science and Pollution Research, 2022 29, 45: 68132-68142. [CrossRef]
- Kundungal, H.;Gangarapu, M.;Sarangapani, S.;Patchaiyappan, A.;Devipriya, S. P. Role of pretreatment and evidence for the enhanced biodegradation and mineralization of low-density polyethylene films by greater waxworm, Environmental Technology, 2021 42, 5: 717-730. [CrossRef]
- Peydaei, A.;Bagheri, H.;Gurevich, L.;de Jonge, N.;Nielsen, J. L. Impact of polyethylene on salivary glands proteome in Galleria melonella, Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 2020 34: 100678. [CrossRef]
- Ren, L. et al. Biodegradation of Polyethylene by Enterobacter sp. D1 from the Guts of Wax Moth Galleria mellonella, International Journal of Environmental Research and Public Health, 2019 16, 11. [CrossRef]
- Zhang, J. et al. Biodegradation of polyethylene microplastic particles by the fungus Aspergillus flavus from the guts of wax moth Galleria mellonella, Science of The Total Environment, 2020 704: 135931. [CrossRef]
- Di Napoli, M. et al. High density polyethylene (HDPE) biodegradation by the fungus Cladosporium halotolerans, FEMS Microbiology Ecology, 2023 99, 2: fiac148. [CrossRef]
- Nyamjav, I.;Jang, Y.;Park, N.;Lee, Y. E.;Lee, S. Physicochemical and Structural Evidence that Bacillus cereus Isolated from the Gut of Waxworms (Galleria mellonella Larvae) Biodegrades Polypropylene Efficiently In Vitro, Journal of Polymers and the Environment, 2023 31, 10: 4274-4287. [CrossRef]
- Jiang, S.;Su, T.;Zhao, J.;Wang, Z. Isolation, Identification, and Characterization of Polystyrene-Degrading Bacteria From the Gut of Galleria Mellonella (Lepidoptera: Pyralidae) Larvae, (in English), Frontiers in Bioengineering and Biotechnology, 2021Original Research 9. [CrossRef]
- LeMoine, C. M. R.;Grove, H. C.;Smith, C. M.;Cassone, B. J. A Very Hungry Caterpillar: Polyethylene Metabolism and Lipid Homeostasis in Larvae of the Greater Wax Moth (Galleria mellonella), Environmental Science & Technology, 2020 54, 22: 14706-14715. [CrossRef]
- Zhong, Z.;Nong, W.;Xie, Y.;Hui, J. H. L.;Chu, L. M. Long-term effect of plastic feeding on growth and transcriptomic response of mealworms (Tenebrio molitor L.), Chemosphere, 2022 287: 132063. [CrossRef]
- Young, R. et al. Improved reference quality genome sequence of the plastic-degrading greater wax moth, Galleria mellonella, G3 Genes|Genomes|Genetics, 2024: jkae070. [CrossRef]
- Venegas, S. et al., "Biodegradation of Polystyrene by Galleria mellonella: Identification of Potential Enzymes Involved in the Degradative Pathway," Int J Mol Sci, vol. 25, no. 3. [CrossRef]
- Wang, S. et al. Complete digestion/biodegradation of polystyrene microplastics by greater wax moth (Galleria mellonella) larvae: Direct in vivo evidence, gut microbiota independence, and potential metabolic pathways, Journal of Hazardous Materials, 2022 423: 127213. [CrossRef]
- O'Connor, K.;Buckley, C. M.;Hartmans, S.;Dobson, A. D. Possible regulatory role for nonaromatic carbon sources in styrene degradation by Pseudomonas putida CA-3, Applied and Environmental Microbiology, 1995 61, 2: 544-548. [CrossRef]
- Noël, G.;Serteyn, L.;Sare, A. R.;Massart, S.;Delvigne, F.;Francis, F. Co-diet supplementation of low density polyethylene and honeybee wax did not influence the core gut bacteria and associated enzymes of Galleria mellonella larvae (Lepidoptera: Pyralidae), International Microbiology, 2023 26, 2: 397-409. [CrossRef]
- Gressitt, J. L. "Coleopteran." https://www.britannica.com/animal/beetle. (accessed.
- Cucini, C.;Leo, C.;Vitale, M.;Frati, F.;Carapelli, A.;Nardi, F. Bacterial and fungal diversity in the gut of polystyrene-fed Alphitobius diaperinus (Insecta: Coleoptera), Animal Gene, 2020 17-18: 200109. [CrossRef]
- Woo, S.;Song, I.;Cha Hyung, J. Fast and Facile Biodegradation of Polystyrene by the Gut Microbial Flora of Plesiophthalmus davidis Larvae, Applied and Environmental Microbiology, 2020 86, 18: e01361-20. [CrossRef]
- Wang, Z.;Xin, X.;Shi, X.;Zhang, Y. A polystyrene-degrading Acinetobacter bacterium isolated from the larvae of Tribolium castaneum, Science of The Total Environment, 2020 726: 138564. [CrossRef]
- Kundungal, H.;Synshiang, K.;Devipriya, S. P. Biodegradation of polystyrene wastes by a newly reported honey bee pest Uloma sp. larvae: An insight to the ability of polystyrene-fed larvae to complete its life cycle, Environmental Challenges, 2021 4: 100083. [CrossRef]
- Hong, J.;Han, T.;Kim, Y. Y., "Mealworm (Tenebrio molitor Larvae) as an Alternative Protein Source for Monogastric Animal: A Review," Animals, vol. 10, no. 11. [CrossRef]
- Shafique, L. et al., "The Feasibility of Using Yellow Mealworms (Tenebrio molitor): Towards a Sustainable Aquafeed Industry," Animals, vol. 11, no. 3. [CrossRef]
- Seong Hyeon, K.;Wonho, C.;Seong-Jin, H.;Nam-Jeong, K. Nutritional Value of Mealworm, Tenebrio molitor as Food Source, International Journal of Industrial Entomology., 2012 25, 1: 93-98. [CrossRef]
- Zielińska, E. et al. The impact of polystyrene consumption by edible insects Tenebrio molitor and Zophobas morio on their nutritional value, cytotoxicity, and oxidative stress parameters, Food Chemistry, 2021 345: 128846. [CrossRef]
- Jo, Y. H.;Lee, J. H.;Patnaik, B. B.;Keshavarz, M.;Lee, Y. S.;Han, Y. S. Autophagy in Tenebrio molitor Immunity: Conserved Antimicrobial Functions in Insect Defenses, (in English), Frontiers in Immunology, 2021Review 12. [CrossRef]
- McConnell, M. W. ;Judge, K. A. Body size and lifespan are condition dependent in the mealworm beetle, Tenebrio molitor, but not sexually selected traits, Behavioral Ecology and Sociobiology, 2018 72, 3: 32. [CrossRef]
- Machona, O.;Chidzwondo, F.;Mangoyi, R. Tenebrio molitor: possible source of polystyrene-degrading bacteria, BMC Biotechnology, 2022 22, 1: 2. [CrossRef]
- Urbanek, A. K.;Rybak, J.;Wróbel, M.;Leluk, K.;Mirończuk, A. M. A comprehensive assessment of microbiome diversity in Tenebrio molitor fed with polystyrene waste, Environmental Pollution, 2020 262: 114281. [CrossRef]
- Brandon, A. M. et al. Biodegradation of Polyethylene and Plastic Mixtures in Mealworms (Larvae of Tenebrio molitor) and Effects on the Gut Microbiome, Environmental Science & Technology, 2018 52, 11: 6526-6533. [CrossRef]
- Brandon, A. M.;Garcia, A. M.;Khlystov, N. A.;Wu, W.-M.;Criddle, C. S. Enhanced Bioavailability and Microbial Biodegradation of Polystyrene in an Enrichment Derived from the Gut Microbiome of Tenebrio molitor (Mealworm Larvae), Environmental Science & Technology, 2021 55, 3: 2027-2036. [CrossRef]
- Peng, B.-Y. et al. Biodegradation of Polystyrene by Dark (Tenebrio obscurus) and Yellow (Tenebrio molitor) Mealworms (Coleoptera: Tenebrionidae), Environmental Science & Technology, 2019 53, 9: 5256-5265. [CrossRef]
- Yang, Y. et al. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms, Environmental Science & Technology, 2015 49, 20: 12087-12093. [CrossRef]
- Yang, S.-S. et al. Biodegradation of polystyrene wastes in yellow mealworms (larvae of Tenebrio molitor Linnaeus): Factors affecting biodegradation rates and the ability of polystyrene-fed larvae to complete their life cycle, Chemosphere, 2018 191: 979-989. [CrossRef]
- Jiang, S.;Su, T.;Zhao, J.;Wang, Z. Biodegradation of Polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus Larvae and Comparison of Their Degradation Effects, Polymers, 2021 13, 20. [CrossRef]
- Yang, L. et al. Biodegradation of expanded polystyrene and low-density polyethylene foams in larvae of Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae): Broad versus limited extent depolymerization and microbe-dependence versus independence, Chemosphere, 2021 262: 127818. [CrossRef]
- He, L. et al. Responses of gut microbiomes to commercial polyester polymer biodegradation in Tenebrio molitor Larvae, Journal of Hazardous Materials, 2023 457: 131759. [CrossRef]
- Mamtimin, T. et al. Gut microbiome of mealworms (Tenebrio molitor Larvae) show similar responses to polystyrene and corn straw diets, Microbiome, 2023 11, 1: 98. [CrossRef]
- Yang, Y.;Hu, L.;Li, X.;Wang, J.;Jin, G. Nitrogen Fixation and Diazotrophic Community in Plastic-Eating Mealworms Tenebrio molitor L, Microbial Ecology, 2022. [CrossRef]
- Tsochatzis, E.;Berggreen, I. E.;Tedeschi, F.;Ntrallou, K.;Gika, H.;Corredig, M., "Gut Microbiome and Degradation Product Formation during Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies," Molecules, vol. 26, no. 24. [CrossRef]
- Gan, S. K.;Phua, S.-X.;Yeo, J. Y.;Heng, Z. S.;Xing, Z. Method for Zero-Waste Circular Economy Using Worms for Plastic Agriculture: Augmenting Polystyrene Consumption and Plant Growth, Methods and Protocols, 2021 4, 2. [CrossRef]
- Gan, S. K.;Phua, S.-X.;Yeo, J. Y.;Heng, Z. S.;Xing, Z., "Method for Zero-Waste Circular Economy Using Worms for Plastic Agriculture: Augmenting Polystyrene Consumption and Plant Growth," Methods and Protocols, vol. 4, no. 2. [CrossRef]
- Lou, Y. et al. Response of the yellow mealworm (Tenebrio molitor) gut microbiome to diet shifts during polystyrene and polyethylene biodegradation, Journal of Hazardous Materials, 2021 416: 126222. [CrossRef]
- Peng, B.-Y. et al. Unveiling Fragmentation of Plastic Particles during Biodegradation of Polystyrene and Polyethylene Foams in Mealworms: Highly Sensitive Detection and Digestive Modeling Prediction, Environmental Science & Technology, 2023 57, 40: 15099-15111. [CrossRef]
- Xu, Y.;Xian, Z.-N.;Yue, W.;Yin, C.-F.;Zhou, N.-Y. Degradation of polyvinyl chloride by a bacterial consortium enriched from the gut of Tenebrio molitor larvae, Chemosphere, 2023 318: 137944. [CrossRef]
- Xian, Z.-N.;Yin, C.-F.;Zheng, L.;Zhou, N.-Y.;Xu, Y. Biodegradation of additive-free polypropylene by bacterial consortia enriched from the ocean and from the gut of Tenebrio molitor larvae, Science of The Total Environment, 2023 892: 164721. [CrossRef]
- Jin, L.;Feng, P.;Cheng, Z.;Wang, D. Effect of biodegrading polyethylene, polystyrene, and polyvinyl chloride on the growth and development of yellow mealworm (Tenebrio molitor) larvae, Environmental Science and Pollution Research, 2023 30, 13: 37118-37126. [CrossRef]
- Leicht, A. ;Masuda, H. Ingestion of Nylon 11 Polymers by the Mealworm (Tenebrio molitor) Beetle and Subsequent Enrichment of Monomer-Metabolizing Bacteria in Fecal Microbiome, FBE, 2023 15, 2. [CrossRef]
- Leicht, A.;Gatz-Schrupp, J.;Masuda, H. Discovery of Nylon 11 ingestion by mealworm (Tenebrio molitor) larvae and detection of monomer-degrading bacteria in gut microbiota, AIMS Microbiology, 2022 8, 4: 612-623. [CrossRef]
- Li, X. et al. Differences in ingestion and biodegradation of the melamine formaldehyde plastic by yellow mealworms Tenebrio molitor and superworms Zophobas atratus, and the prediction of functional gut microbes, Chemosphere, 2024 352: 141499. [CrossRef]
- Peng, B.-Y. et al. Biodegradation of polyvinyl chloride, polystyrene, and polylactic acid microplastics in Tenebrio molitor larvae: Physiological responses, Journal of Environmental Management, 2023 345: 118818. [CrossRef]
- Liu, J. et al. Biodegradation of polyether-polyurethane foam in yellow mealworms (Tenebrio molitor) and effects on the gut microbiome, Chemosphere, 2022 304: 135263. [CrossRef]
- Orts, J. M. et al., "Polyurethane Foam Residue Biodegradation through the Tenebrio molitor Digestive Tract: Microbial Communities and Enzymatic Activity," Polymers, vol. 15, no. 1. [CrossRef]
- Guo, B.;Yin, J.;Hao, W.;Jiao, M. Polyurethane foam induces epigenetic modification of mitochondrial DNA during different metamorphic stages of Tenebrio molitor, Ecotoxicology and Environmental Safety, 2019 183: 109461. [CrossRef]
- Wang, J. et al. Ingestion and biodegradation of disposable surgical masks by yellow mealworms Tenebrio molitor larvae: Differences in mask layers and effects on the larval gut microbiome, Science of The Total Environment, 2023 904: 166808. [CrossRef]
- Yang, S.-S. et al. Impacts of physical-chemical property of polyethylene on depolymerization and biodegradation in yellow and dark mealworms with high purity microplastics, Science of The Total Environment, 2022 828: 154458. [CrossRef]
- Akash, K.;Parthasarathi, R.;Elango, R.;Bragadeeswaran, S. Characterization of Priestia megaterium S1, a polymer degrading gut microbe isolated from the gut of Tenebrio molitor larvae fed on Styrofoam, Archives of Microbiology, 2023 206, 1: 48. [CrossRef]
- Park, J.-W.;Kim, M.;Kim, S.-Y.;Bae, J.;Kim, T.-J. Biodegradation of polystyrene by intestinal symbiotic bacteria isolated from mealworms, the larvae of <em>Tenebrio molitor</em>, Heliyon, 2023 9, 6. [CrossRef]
- Kang, M.-G.;Kwak, M.-J.;Kim, Y. Polystyrene microplastics biodegradation by gut bacterial Enterobacter hormaechei from mealworms under anaerobic conditions: Anaerobic oxidation and depolymerization, Journal of Hazardous Materials, 2023 459: 132045. [CrossRef]
- Wu, Q.;Tao, H.;Wong, M. H. Feeding and metabolism effects of three common microplastics on Tenebrio molitor L, Environmental Geochemistry and Health, 2019 41, 1: 17-26. [CrossRef]
- Peng, B.-Y. et al. Influence of Polymer Size on Polystyrene Biodegradation in Mealworms (Tenebrio molitor): Responses of Depolymerization Pattern, Gut Microbiome, and Metabolome to Polymers with Low to Ultrahigh Molecular Weight, Environmental Science & Technology, 2022 56, 23: 17310-17320. [CrossRef]
- Yang, S.-S. et al. Ubiquity of polystyrene digestion and biodegradation within yellow mealworms, larvae of Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae), Chemosphere, 2018 212: 262-271. [CrossRef]
- Wang, Y. et al. Different plastics ingestion preferences and efficiencies of superworm (Zophobas atratus Fab.) and yellow mealworm (Tenebrio molitor Linn.) associated with distinct gut microbiome changes, Science of The Total Environment, 2022 837: 155719. [CrossRef]
- Cheng, X.;Xia, M.;Yang, Y. Biodegradation of vulcanized rubber by a gut bacterium from plastic-eating mealworms, (in eng), J Hazard Mater, 2023 448: 130940. [CrossRef]
- Ding, M.-Q. et al. Gut Microbiome Associating with Carbon and Nitrogen Metabolism during Biodegradation of Polyethene in Tenebrio larvae with Crop Residues as Co-Diets, Environmental Science & Technology, 2023 57, 8: 3031-3041. [CrossRef]
- Yang, S.-S. et al. Confirmation of biodegradation of low-density polyethylene in dark- versus yellow- mealworms (larvae of Tenebrio obscurus versus Tenebrio molitor) via. gut microbe-independent depolymerization, Science of The Total Environment, 2021 789: 147915. [CrossRef]
- Rumbos, C. I. ;Athanassiou, C. G. The Superworm, Zophobas morio (Coleoptera:Tenebrionidae): A ‘Sleeping Giant’ in Nutrient Sources, Journal of Insect Science, 2021 21, 2: 13. [CrossRef]
- Yang, Y.;Wang, J.;Xia, M. Biodegradation and mineralization of polystyrene by plastic-eating superworms Zophobas atratus, Science of The Total Environment, 2020 708: 135233. [CrossRef]
- Lu, B. et al. Understanding the Ecological Robustness and Adaptability of the Gut Microbiome in Plastic-Degrading Superworms (Zophobas atratus) in Response to Microplastics and Antibiotics, Environmental Science & Technology, 2024. [CrossRef]
- Zaman, I. et al. Biodegradation of polyethylene and polystyrene by Zophobas atratus larvae from Bangladeshi source and isolation of two plastic-degrading gut bacteria, Environmental Pollution, 2024 345: 123446. [CrossRef]
- Nyamjav, I.;Jang, Y.;Lee, Y. E.;Lee, S. Biodegradation of polyvinyl chloride by Citrobacter koseri isolated from superworms (Zophobas atratus larvae), (in English), Frontiers in Microbiology, 2023Original Research 14. [CrossRef]
- Liu, Y.-N.;Bairoliya, S.;Zaiden, N.;Cao, B. Establishment of plastic-associated microbial community from superworm gut microbiome, Environment International, 2024 183: 108349. [CrossRef]
- Luo, L. et al. Biodegradation of foam plastics by Zophobas atratus larvae (Coleoptera: Tenebrionidae) associated with changes of gut digestive enzymes activities and microbiome, Chemosphere, 2021 282: 131006. [CrossRef]
- Weng, Y.;Han, X.;Sun, H.;Wang, J.;Wang, Y.;Zhao, X. Effects of polymerization types on plastics ingestion and biodegradation by Zophobas atratus larvae, and successions of both gut bacterial and fungal microbiomes, Environmental Research, 2024 251: 118677. [CrossRef]
- Jung, H. et al. Circular waste management: Superworms as a sustainable solution for biodegradable plastic degradation and resource recovery, Waste Management, 2023 171: 568-579. [CrossRef]
- Kim, H. R. et al. Biodegradation of Polystyrene by Pseudomonas sp. Isolated from the Gut of Superworms (Larvae of Zophobas atratus), Environmental Science & Technology, 2020 54, 11: 6987-6996. [CrossRef]
- Arunrattiyakorn, P. et al. Biodegradation of polystyrene by three bacterial strains isolated from the gut of Superworms (Zophobas atratus larvae), Journal of Applied Microbiology, 2022 132, 4: 2823-2831. [CrossRef]
- Chen, Z. et al. Reactive Oxygen Species Triggered Oxidative Degradation of Polystyrene in the Gut of Superworms (Zophobas atratus Larvae), Environmental Science & Technology, 2023 57, 20: 7867-7874. [CrossRef]
- Inward, D.;Beccaloni, G.;Eggleton, P. Death of an order: a comprehensive molecular phylogenetic study confirms that termites are eusocial cockroaches, Biology Letters, 2007 3, 3: 331-335. [CrossRef]
- Kalleshwaraswamy, C. M.;Shanbhag, R. R.;Sundararaj, R., "Wood Degradation by Termites: Ecology, Economics and Protection," in Science of Wood Degradation and its Protection, R. Sundararaj Ed. Singapore: Springer Singapore, 2022, pp. 147-170.
- Côté, W. A., "Chemical Composition of Wood," in Principles of Wood Science and Technology: I Solid Wood, F. F. P. Kollmann and W. A. Côté Eds. Berlin, Heidelberg: Springer Berlin Heidelberg, 1968, pp. 55-78.
- Al-Tohamy, R. et al. Environmental and Human Health Impact of Disposable Face Masks During the COVID-19 Pandemic: Wood-Feeding Termites as a Model for Plastic Biodegradation, Applied Biochemistry and Biotechnology, 2023 195, 3: 2093-2113. [CrossRef]
- López-Naranjo, E. J.;Alzate-Gaviria, L. M.;Hernández-Zárate, G.;Reyes-Trujeque, J.;Cupul-Manzano, C. V.;Cruz-Estrada, R. H. Effect of biological degradation by termites on the flexural properties of pinewood residue/recycled high-density polyethylene composites, Journal of Applied Polymer Science, 2013 128, 5: 2595-2603. [CrossRef]
- Samir Ali, S.;Al-Tohamy, R.;Sun, J.;Wu, J.;Huizi, L. Screening and construction of a novel microbial consortium SSA-6 enriched from the gut symbionts of wood-feeding termite, Coptotermes formosanus and its biomass-based biorefineries, Fuel, 2019 236: 1128-1145. [CrossRef]
- Guzmán, L. F.;Tirado, B.;Cruz-Cárdenas, C. I.;Rojas-Anaya, E.;Aragón-Magadán, M. A., "De Novo Transcriptome Assembly of Cedar (Cedrela odorata L.) and Differential Gene Expression Involved in Herbivore Resistance," Current Issues in Molecular Biology, vol. 46, no. 8, pp. 8794-8806. [CrossRef]
- Baranchikov, Y.;Mozolevskaya, E.;Yurchenko, G.;Kenis, M. Occurrence of the emerald ash borer, Agrilus planipennis in Russia and its potential impact on European forestry, EPPO Bulletin, 2008 38, 2: 233-238. [CrossRef]
- Mayfield Iii, A. E. et al. Suitability of California bay laurel and other species as hosts for the non-native redbay ambrosia beetle and granulate ambrosia beetle, Agricultural and Forest Entomology, 2013 15, 3: 227-235. [CrossRef]
- Meng, P. S.;Hoover, K.;Keena, M. A. Asian Longhorned Beetle (Coleoptera: Cerambycidae), an Introduced Pest of Maple and Other Hardwood Trees in North America and Europe, Journal of Integrated Pest Management, 2015 6, 1: 4. [CrossRef]
- Linnakoski, R. ;Forbes, K. M. Pathogens—The Hidden Face of Forest Invasions by Wood-Boring Insect Pests, (in English), Frontiers in Plant Science, 2019Opinion 10. [CrossRef]
- Sangiorgio, P.;Verardi, A.;Dimatteo, S.;Spagnoletta, A.;Moliterni, S.;Errico, S. Tenebrio molitorin the circular economy: a novel approach for plastic valorisation and PHA biological recovery, Environmental Science and Pollution Research, 2021 28, 38: 52689-52701. [CrossRef]
- Kundungal, H. ;Devipriya, S. P. Nature’s solution to degrade long-chain hydrocarbons: A life cycle study of beeswax and plastic eating insect larvae, Research Square 2024.
- Yang, S.-S. et al. Radical innovation breakthroughs of biodegradation of plastics by insects: history, present and future perspectives, Frontiers of Environmental Science & Engineering, 2024 18, 6: 78. [CrossRef]
- Boctor, J.;Pandey, G.;Xu, W.;Murphy, D. V.;Hoyle, F. C., "Nature’s Plastic Predators: A Comprehensive and Bibliometric Review of Plastivore Insects," Polymers, vol. 16, no. 12. [CrossRef]
- Milum, V. G. Vitula edmandsii as a Pest of Honeybee Combs, Journal of Economic Entomology, 1953 46, 4: 710-711. [CrossRef]
- Pereira, M. F. ;Rossi, C. C. Overview of rearing and testing conditions and a guide for optimizing Galleria mellonella breeding and use in the laboratory for scientific purposes, APMIS, 2020 128, 12: 607-620. [CrossRef]
- Zhu, P.;Shen, Y.;Li, X.;Liu, X.;Qian, G.;Zhou, J. Feeding preference of insect larvae to waste electrical and electronic equipment plastics, Science of The Total Environment, 2022 807: 151037. [CrossRef]
- Guberman, R. "The Complete Plastics Recycling Process." https://www.rts.com/blog/the-complete-plastics-recycling-process-rts/ (accessed September 12, 2024).
- Billen, P.;Khalifa, L.;Van Gerven, F.;Tavernier, S.;Spatari, S. Technological application potential of polyethylene and polystyrene biodegradation by macro-organisms such as mealworms and wax moth larvae, Science of The Total Environment, 2020 735: 139521. [CrossRef]
- Donnelly, J. "Current Plastic Recycling Prices." https://blog.recycleduklimited.com/current-plastic-recycling-prices (accessed 12 September 2024).
- Manzano-Agugliaro, F.;Sanchez-Muros, M. J.;Barroso, F. G.;Martínez-Sánchez, A.;Rojo, S.;Pérez-Bañón, C. Insects for biodiesel production, Renewable and Sustainable Energy Reviews, 2012 16, 6: 3744-3753. [CrossRef]
- Siow, H. S.;Sudesh, K.;Ganesan, S. Insect oil to fuel: Optimizing biodiesel production from mealworm (Tenebrio molitor) oil using response surface methodology, Fuel, 2024 371: 132099. [CrossRef]
- Ilijin, L. et al. Sourcing chitin from exoskeleton of Tenebrio molitor fed with polystyrene or plastic kitchen wrap, International Journal of Biological Macromolecules, 2024 268: 131731. [CrossRef]
- Hirano, S., "Chitin Biotechnology Applications," in Biotechnology Annual Review, vol. 2, M. R. El-Gewely Ed.: Elsevier, 1996, pp. 237-258.
- Peng, B. Y. et al. Unveiling Fragmentation of Plastic Particles during Biodegradation of Polystyrene and Polyethylene Foams in Mealworms: Highly Sensitive Detection and Digestive Modeling Prediction, (in eng), Environ Sci Technol, 2023 57, 40: 15099-15111. [CrossRef]
- "Carbon Emissions and Plastic Waste. QM recylced energy." https://www.qmre.ltd/ (accessed 16 September 2024).
- Finnveden, G. et al. Recent developments in Life Cycle Assessment, Journal of Environmental Management, 2009 91, 1: 1-21. [CrossRef]
- Drugmand, J.-C.;Schneider, Y.-J.;Agathos, S. N. Insect cells as factories for biomanufacturing, Biotechnology Advances, 2012 30, 5: 1140-1157. [CrossRef]
- Buchholz, K. ;Collins, J. The roots—a short history of industrial microbiology and biotechnology, Applied Microbiology and Biotechnology, 2013 97, 9: 3747-3762. [CrossRef]
- Sadler, J. C. ;Wallace, S. Microbial synthesis of vanillin from waste poly(ethylene terephthalate), Green Chemistry, 202110.1039/D1GC00931A 23, 13: 4665-4672. [CrossRef]
- Banerjee, G. ;Chattopadhyay, P. Vanillin biotechnology: the perspectives and future, Journal of the Science of Food and Agriculture, 2019 99, 2: 499-506. [CrossRef]
Figure 1.
Main historic events related to insects degrading petroleum-based plastic.
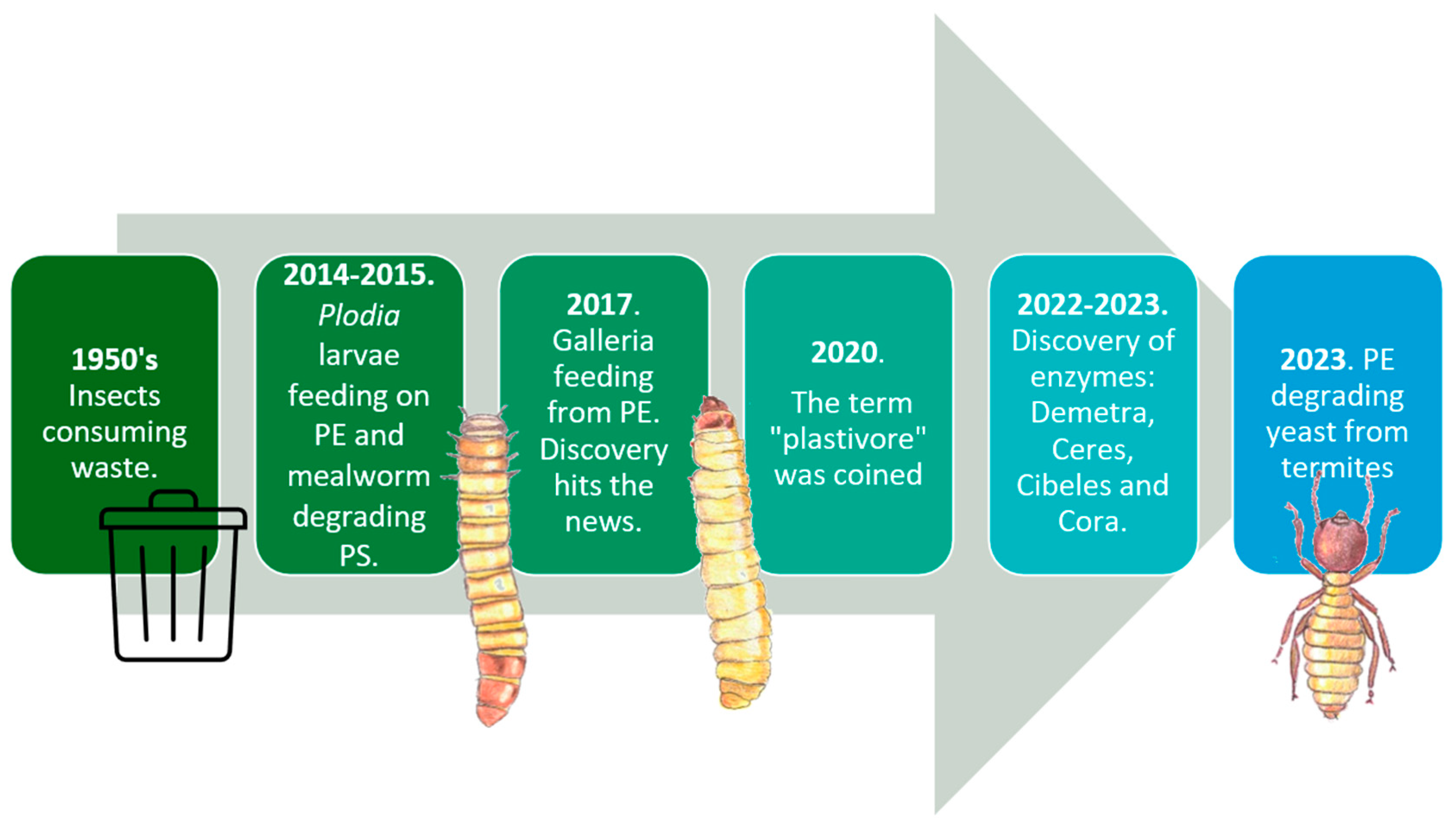
Figure 2.
Structure, density, crystallinity, life span and common uses if the most abundant plastic resins [19,20]. Half-life of PU is still unknown [22].
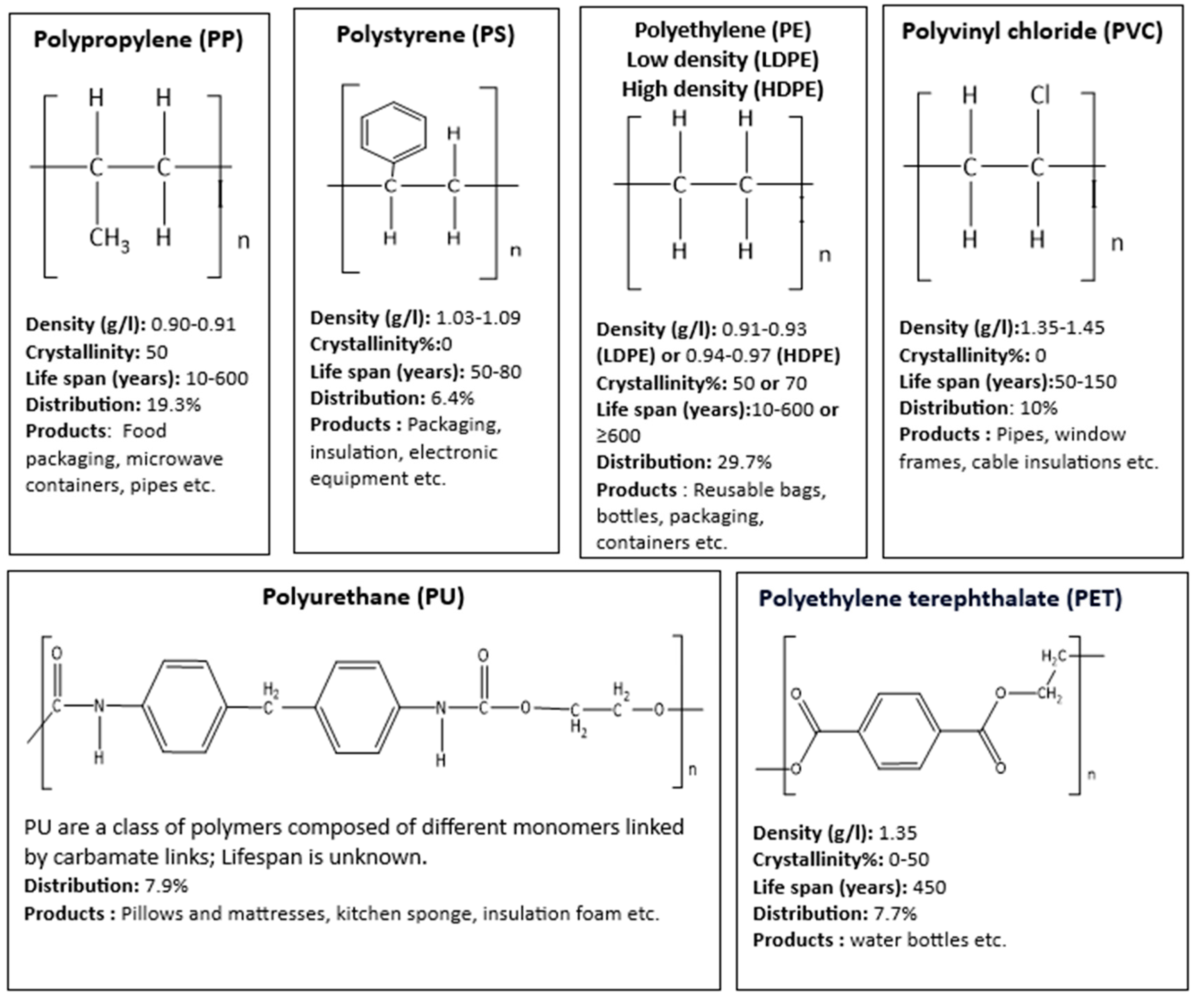
Figure 3.
Types of plastic degradation factors in the environment. [26].
Figure 3.
Types of plastic degradation factors in the environment. [26].
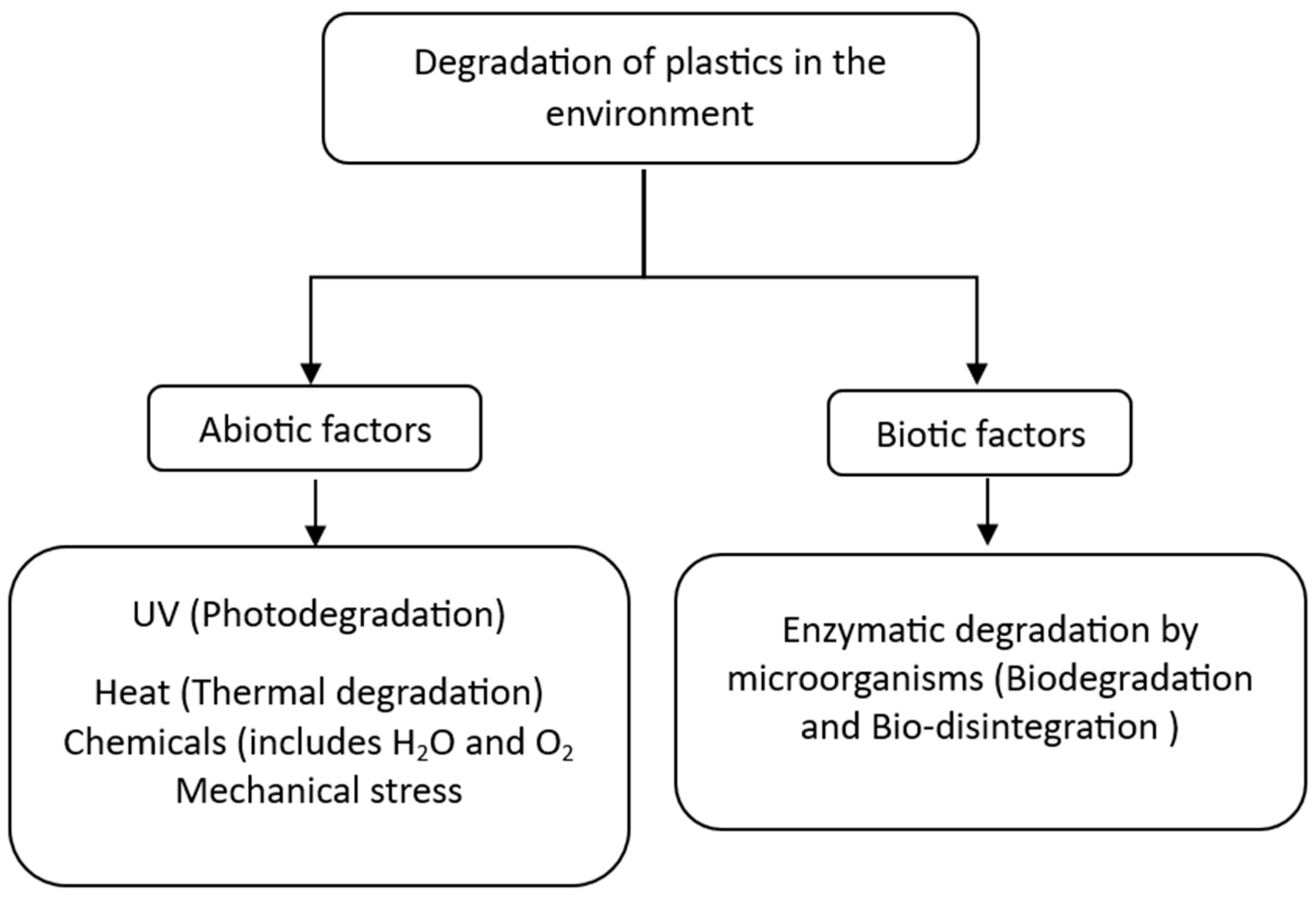
Figure 4.
PETase enzymes degrades PET into Bis(2-Hydroxyethyl) terephthalate (BHET), mono(2-hydroxyethyl) terephthalic acid (MHET) and terephthalic acid (TPA). MHETase enzyme further degrades MHET into more TPA and ethylene glycol (EG).
Figure 4.
PETase enzymes degrades PET into Bis(2-Hydroxyethyl) terephthalate (BHET), mono(2-hydroxyethyl) terephthalic acid (MHET) and terephthalic acid (TPA). MHETase enzyme further degrades MHET into more TPA and ethylene glycol (EG).
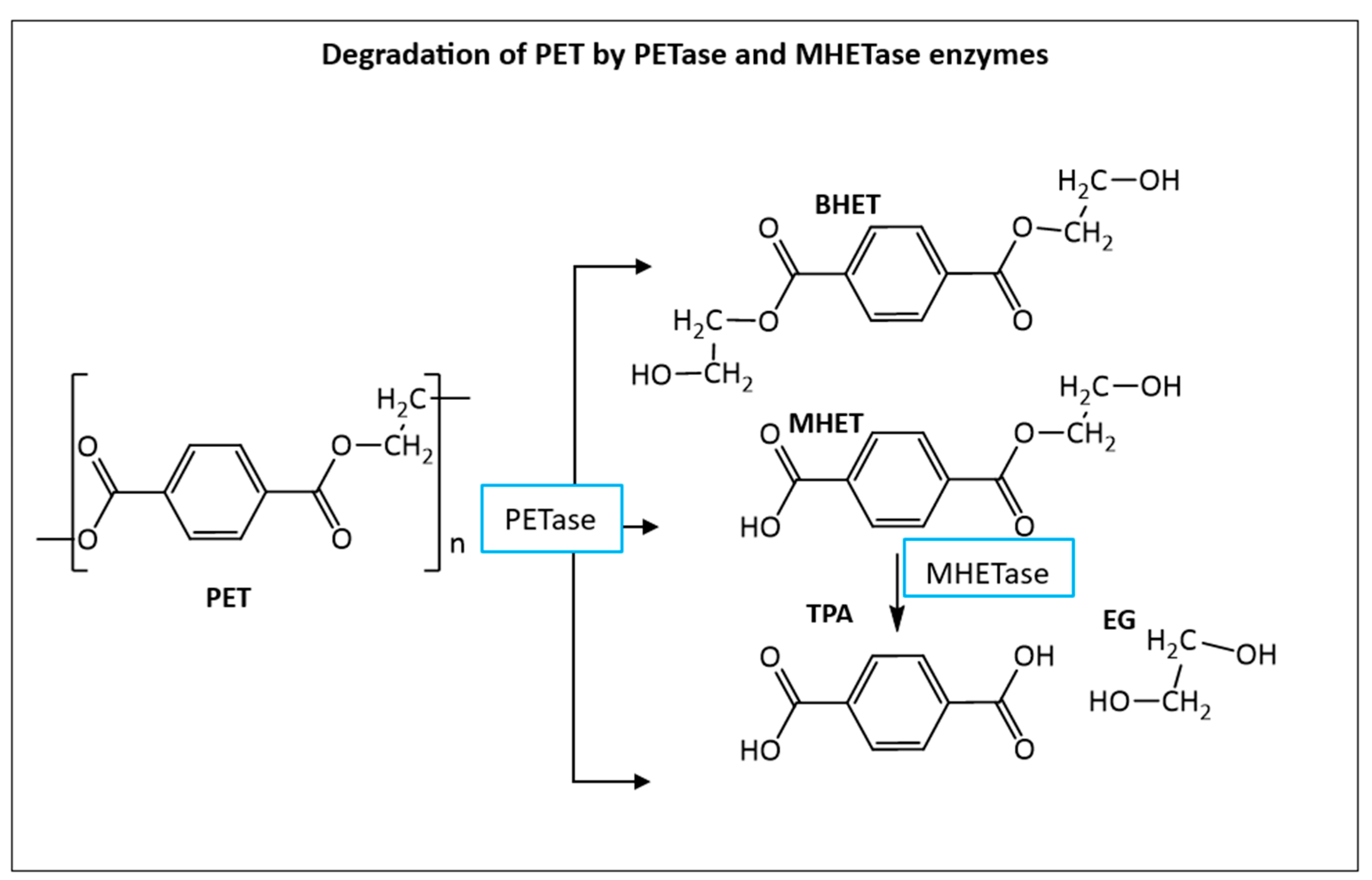
Figure 5.
Insects from the order lepidoptera whose larvae has been reported to have plastic-degrading capabilities. The larvae’s common name is reported as well as the plastic degraded and the microorganism associated. Note: insect figures do not represent real scale. Achroia Grisella [70,71], Plodia interpunctella [6,7,72], Corcyra cephalonica [73], Spodoptera frugiperda [74].
Figure 5.
Insects from the order lepidoptera whose larvae has been reported to have plastic-degrading capabilities. The larvae’s common name is reported as well as the plastic degraded and the microorganism associated. Note: insect figures do not represent real scale. Achroia Grisella [70,71], Plodia interpunctella [6,7,72], Corcyra cephalonica [73], Spodoptera frugiperda [74].
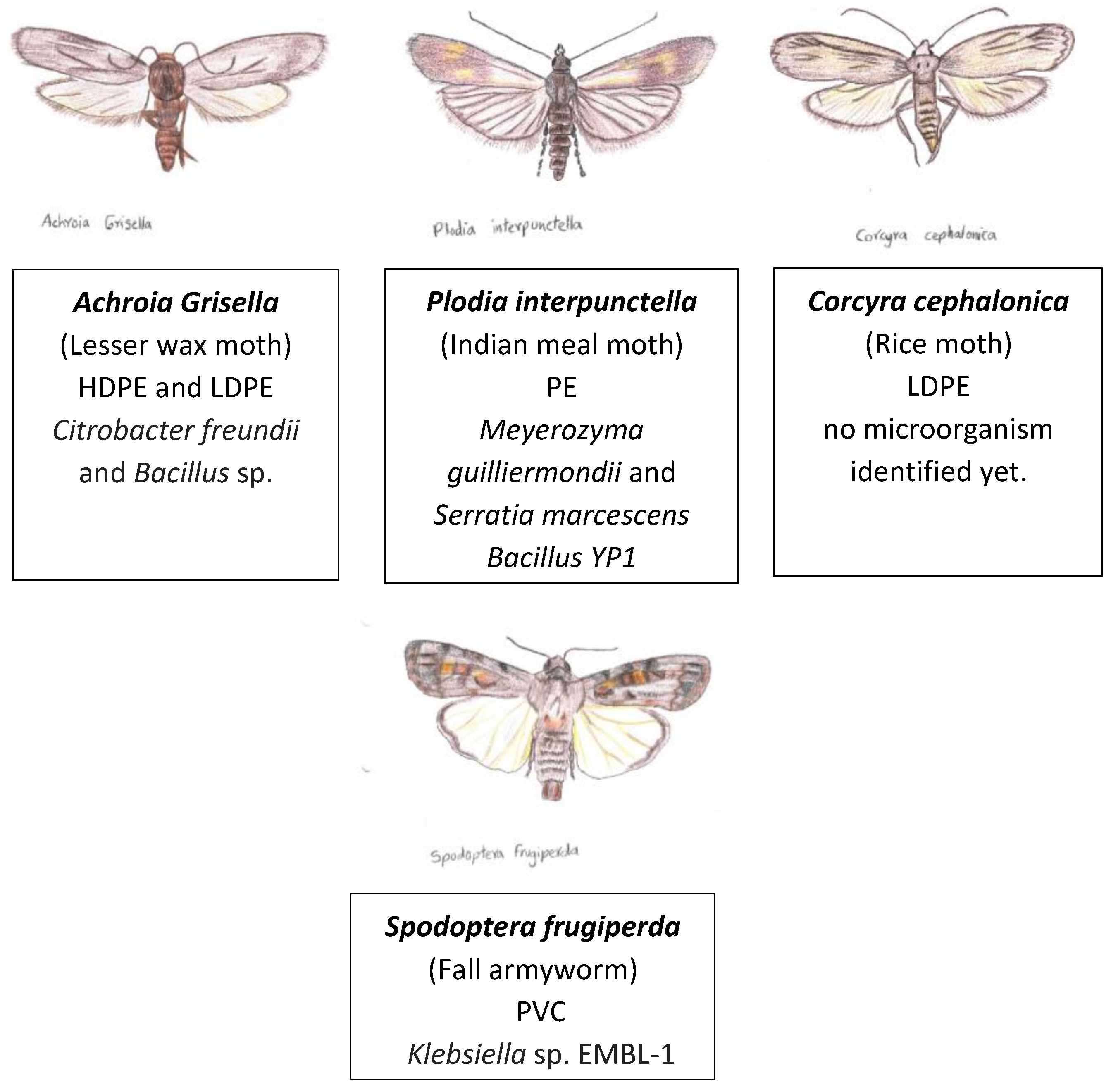
Figure 6.
Galleria mellonella. The female wax moth average body length is 15–20 mm. The male is considerably smaller and darker [75].
Figure 6.
Galleria mellonella. The female wax moth average body length is 15–20 mm. The male is considerably smaller and darker [75].
Figure 7.
The greater wax worm uses different biological tools to degrade plastic: saliva enzymes, metabolic pathways and gut microorganisms.
Figure 7.
The greater wax worm uses different biological tools to degrade plastic: saliva enzymes, metabolic pathways and gut microorganisms.
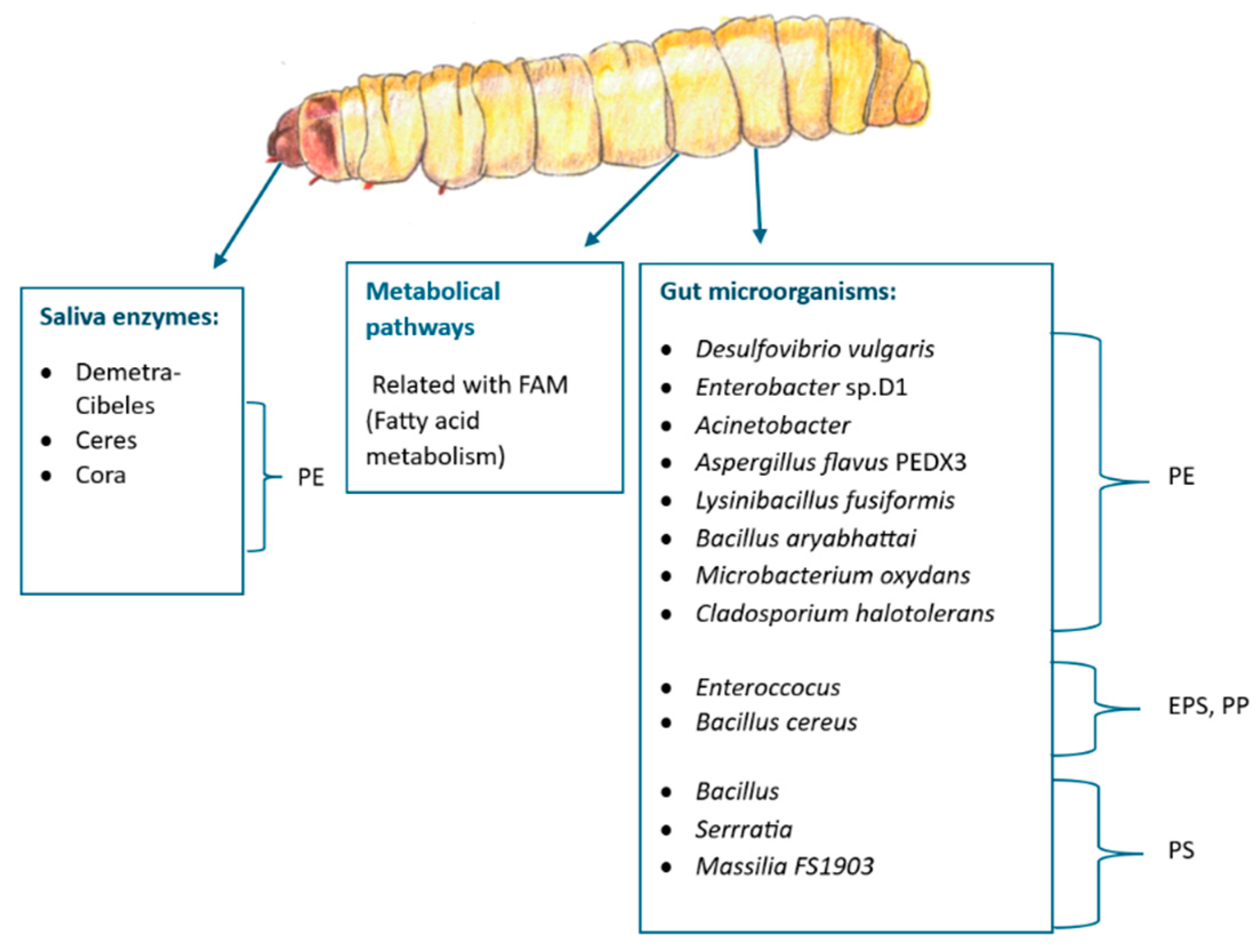
Figure 9.
Insects from the order coleoptera whose larvae has been reported to have plastic-degrading capabilities. The larvae’s common name is reported as well as the plastic degraded and the microorganism associated. Note: insect figures do not represent real scale. Alphitobius diaperinus [100], Plesiophthalmus davidis [101], Tribolium castaneum 1 [102], Uloma sp. (0) (PS)[103].
Figure 9.
Insects from the order coleoptera whose larvae has been reported to have plastic-degrading capabilities. The larvae’s common name is reported as well as the plastic degraded and the microorganism associated. Note: insect figures do not represent real scale. Alphitobius diaperinus [100], Plesiophthalmus davidis [101], Tribolium castaneum 1 [102], Uloma sp. (0) (PS)[103].
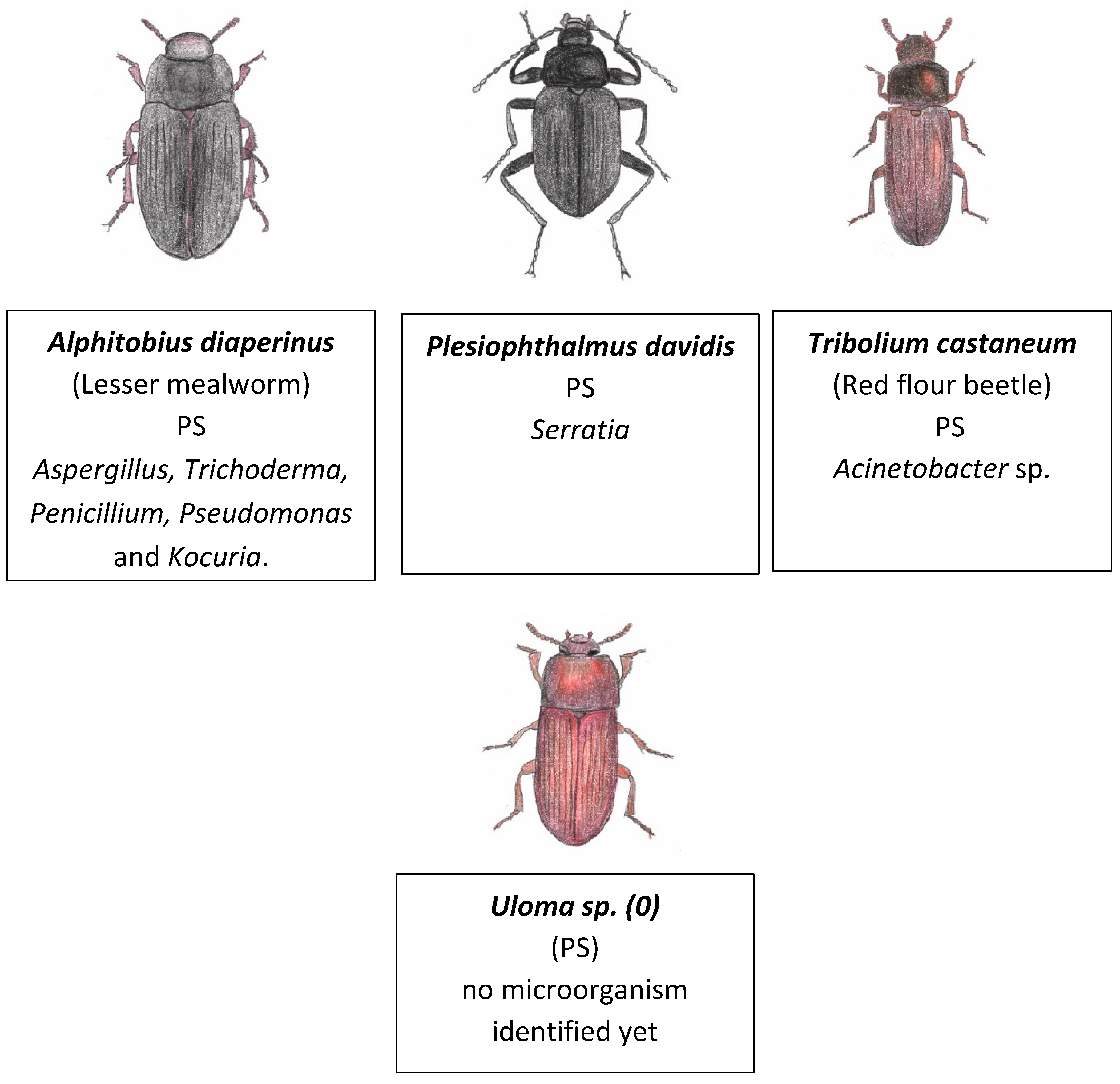
Figure 10.
Tenebrio molitor. Adults are commonly 13 to 18 millimetres long. T. molitor is dark brown or black.[109].
Figure 10.
Tenebrio molitor. Adults are commonly 13 to 18 millimetres long. T. molitor is dark brown or black.[109].
Figure 11.
Proposed mechanism of PE degradation in Tenebrio molitor larvae (mealworms) presented by Zhong, Nong, Xie, Hui and Chu. [93].
Figure 11.
Proposed mechanism of PE degradation in Tenebrio molitor larvae (mealworms) presented by Zhong, Nong, Xie, Hui and Chu. [93].
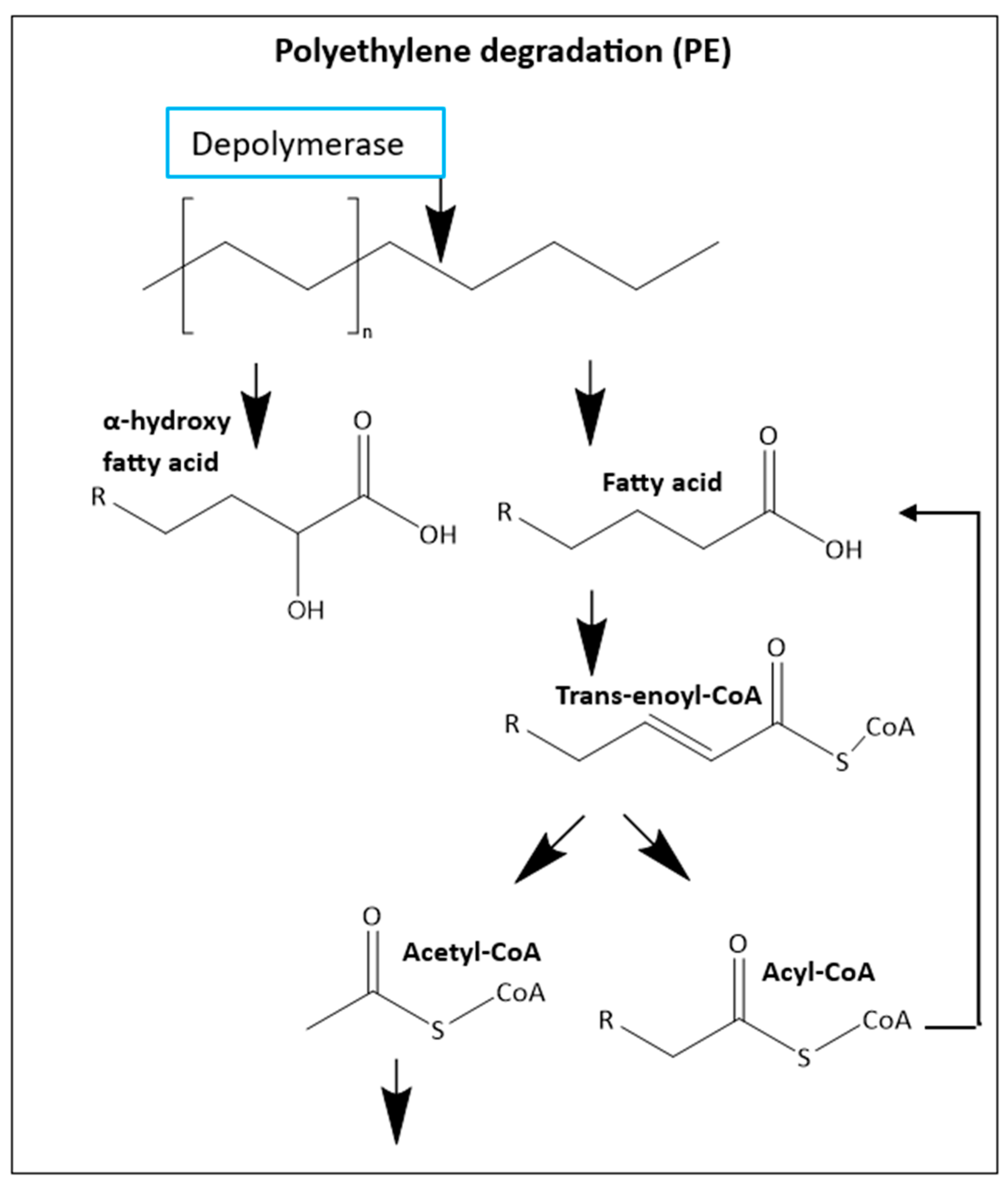
Figure 12.
Tenebrio molitor uses a wide variety of gut microorganisms to degrade plastics.
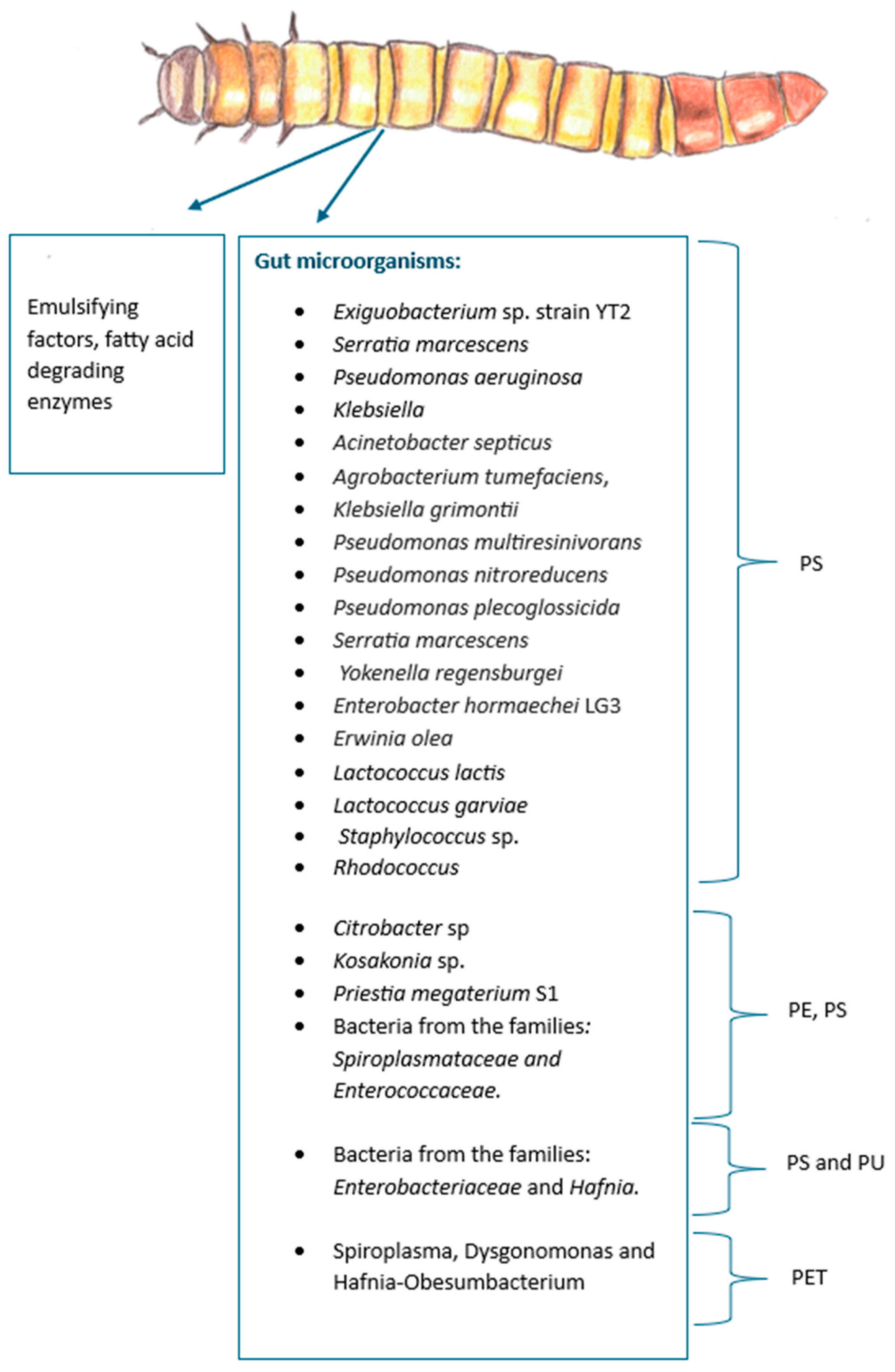
Figure 13.
Adults are dark and 38- to 57mm long.
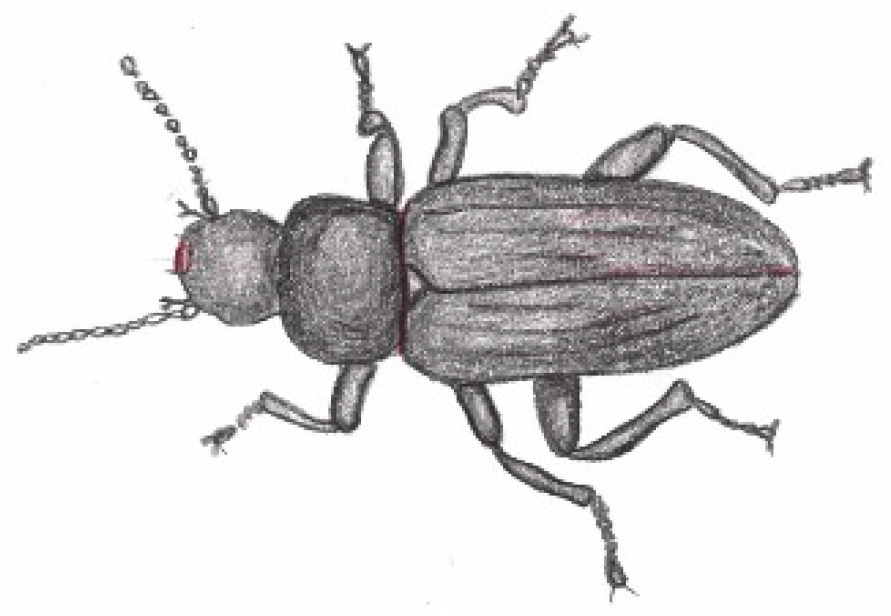
Figure 14.
Zophobas can digest a wide variety of plastics with the aid of gut microbes.
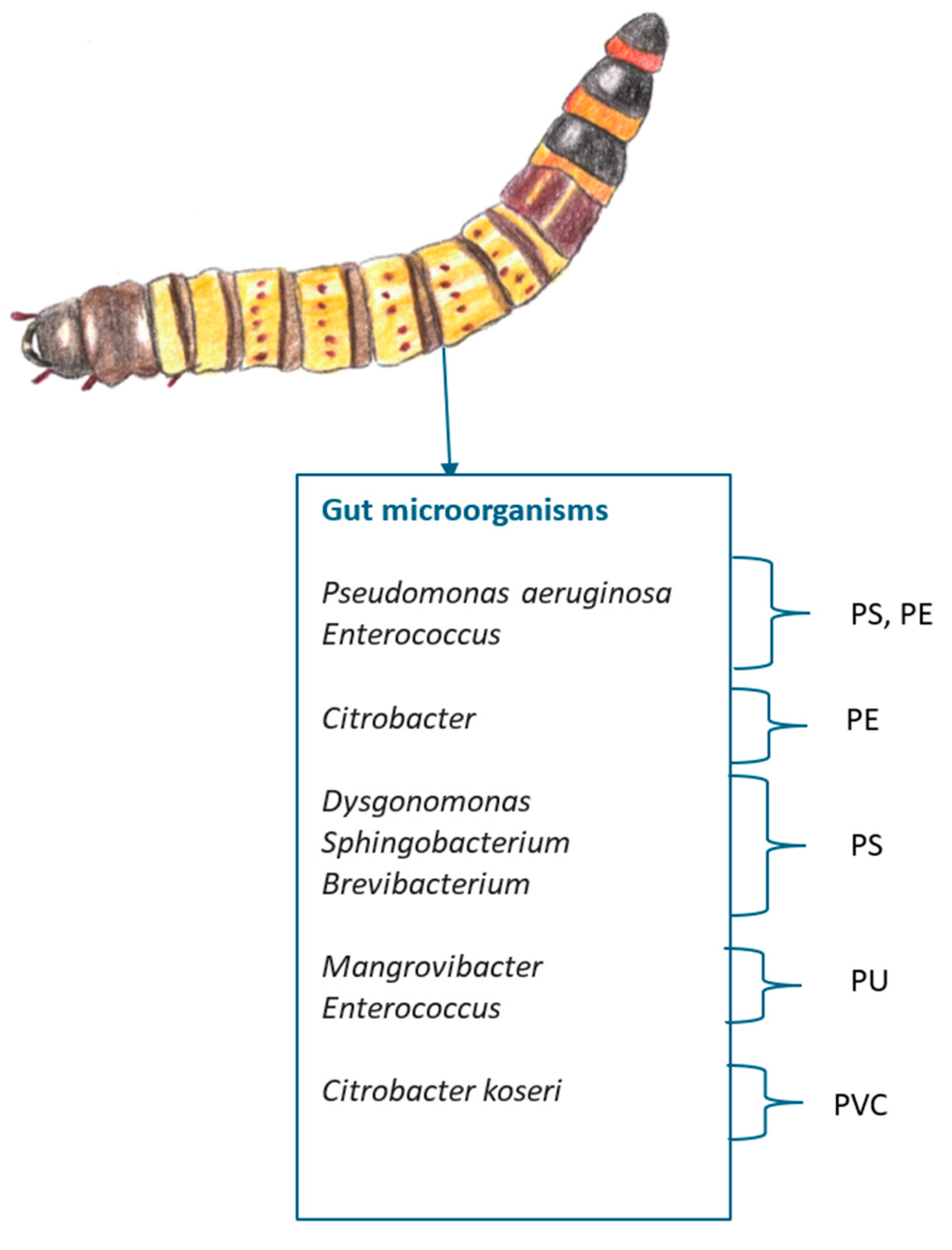
Figure 15.
Termites live in colonies formed by worker, soldier and winged reproductive termites which are represented in this figure. a. [14].
Figure 15.
Termites live in colonies formed by worker, soldier and winged reproductive termites which are represented in this figure. a. [14].
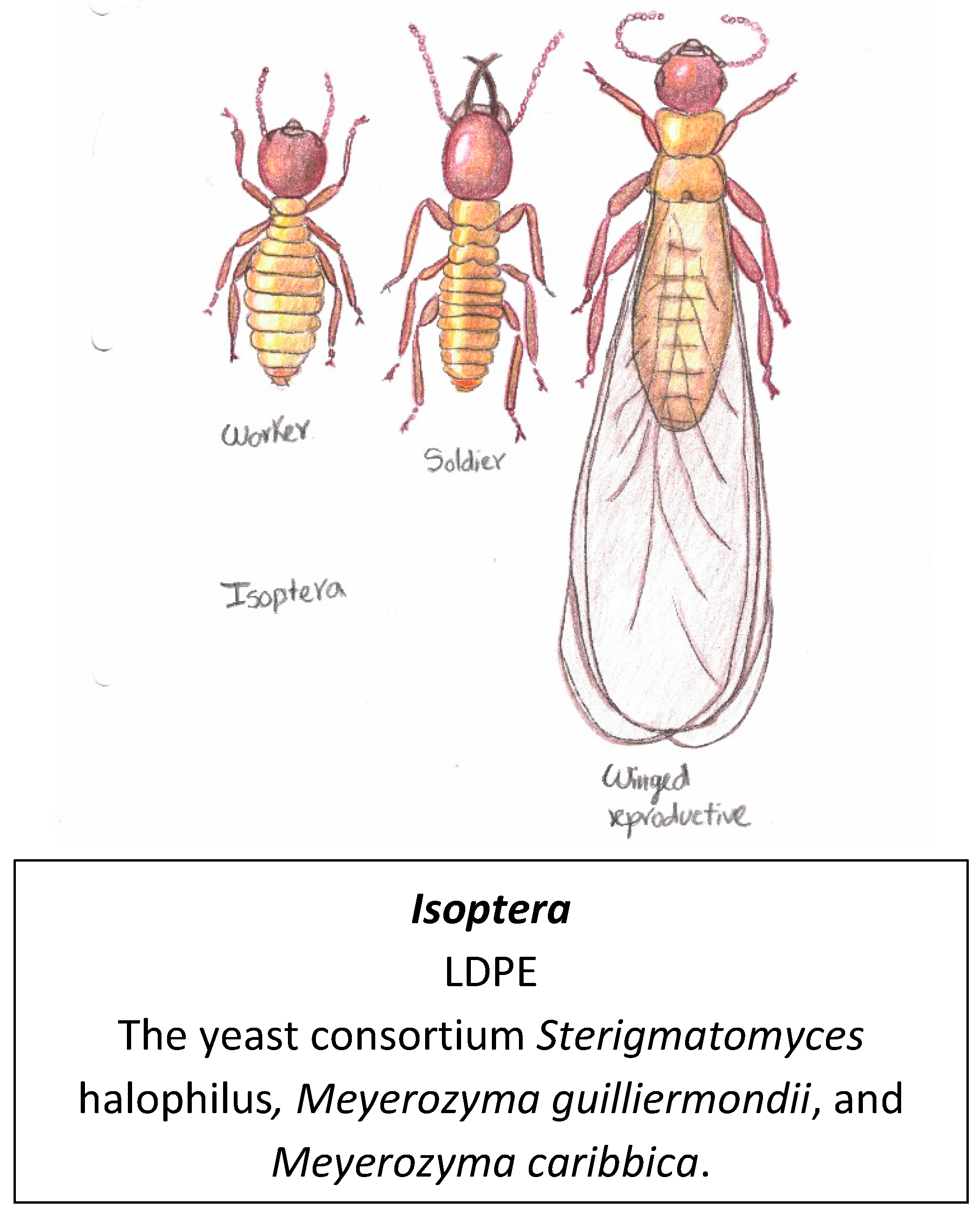
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



