
Submitted:
20 November 2024
Posted:
21 November 2024
You are already at the latest version
Abstract
Flea-borne typhus (FBT), due to Rickettsia typhi and R. felis, is an infection typically causing fever, headache, rash, hepatitis, and thrombocytopenia. About one-quarter of patients suffer pulmonary, neurologic, hematologic, renal, hepatic, cardiac, ocular or other complications. In the 21st century, the incidence of FBT has increased in both Texas and California compared to the 1990s. In this paper, county-level epidemiological data for the number of cases of FBT occurring in Texas for two decades, 1990-1999 and 2010-2019, were compared with respect to county of residence, urbanization, and climatic region. Human population growth in Texas has promoted FBT by increased urbanization and abundance of pet dogs and cats, stray/feral dogs and cats, and opossums. Increasing temperatures in Texas in the new millennium has increased the flea-borne transmission of FBT by promoting host infestation and flea feeding and defecation, accelerating the flea life cycle, and increasing rickettsial replication within the flea. Increased opossums and stray cats and dogs in the urban/suburban landscape has increased the risk of flea transfer to humans and their pets.
Keywords:
flea-borne typhus
; rickettsiae
; opossum
; cat
; dog
; Ctenocephalides felis
; climate change
; urbanization
1. Introduction
Flea-borne typhus (FBT) is an infection caused by the bacteria Rickettsia typhi and R. felis. It is typically an acute undifferentiated febrile illness but about one-quarter of patients suffer respiratory, neurologic, renal, hepatic, cardiac, ocular, or other complications [1,2]. The infection is transmitted to humans by a flea bite or by the inoculation of a bite site, a skin abrasion, or mucous membranes with feces from fleas infected with these rickettsiae [3,4]. In the new millenium, the incidence of FBT has increased in both Texas (TX) and California (CA) [5,6,7]. During 1990-1999, 307 cases of FBT were reported in TX [8]; in the decade 2010 to 2019, there were 3750 cases [5], a 12-fold increase.
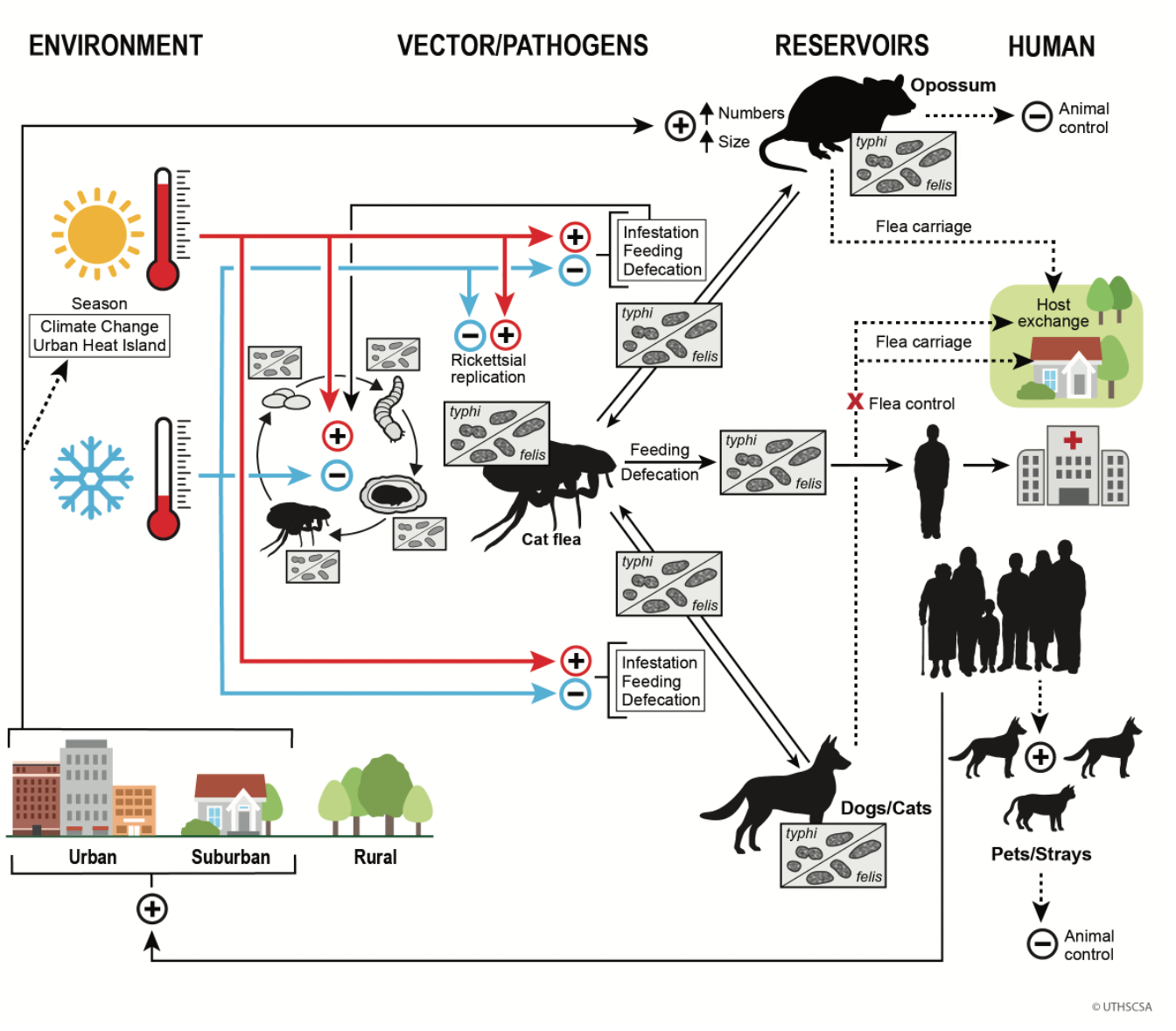
The One Health concept focuses on issues at the intersection of human and animal health and the environment [9]. This manuscript is the first part of a two-part series that integrates the current knowledge of the characteristics of the pathogens and vectors of FBT, the flea host mammals, the environment, and human population trends to derive a One Health model to explain the increase in the number of FBT cases in TX that has occurred in the new millennium. In this schema, the principal driver of the increased number of FBT cases in TX is ecosystem disruption, due to human population growth/urbanization and climate change. The current epidemiologic trends of FBT in TX are dependent on two versatile opportunists, the cat flea Ctenocephalides felis and the Virginia opossum Didelphis virginiana. Furthermore, increasing populations of cats (Felis catus) and dogs (Canis familiaris), including stray and feral animals, may also be affecting the epidemiologic trends [10]. The increasing populations of these mammals are being driven by human population growth. Greater mammalian host abundance may increase cat flea populations, which may also be increased by climate change. In the second part of the series, the natural histories of the host mammals and their relationship to the epidemiology of FBT will be discussed.
2. Methods
Since 1946, the Annual Summary of Notifiable Diseases published by the Texas Department of Health has included data on FBT. In this paper, county-level epidemiological data for the number of cases of FBT occurring in TX for two decades, 1990-1999 [8] and 2010-2019 [5], were compared with respect to county of residence, urbanization, and climatic region. County-level data was obtained from the Texas Department of State Health Services (TDSHS). Temperature data was acquired using the Prism Climate Group database [11]. Populations of TX counties for the indicated years were obtained from US Census data. Historically, most diagnoses of FBT were based on serologic testing of serum of symptomatic patients [12]. More recently, the next generation sequencing technique of whole blood for the detection of the presence of rickettsiae has become commercially available [13,14]. Texas law requires that healthcare providers, hospitals, and laboratories inform the TDSHS if they suspect that someone has a notifiable condition, which includes FBT [15].
3. Historical Epidemiology of Flea-Borne Typhus in Texas, 1923-1999
An episystem is defined as “the set of biological and environmental elements, as well as the epidemiological aspects of these diseases in defined geographic and temporal scales” [16]. In this paper, the geographic space is the state of TX and the time scale that will be discussed is 1930 to The abiotic, biotic, and anthropogenic components of this episystem are listed in Table 1.
In TX in 1923, only five cases of FBT were reported. By 1933, the number of recorded cases had increased to 399 [17]. In the 1930s, many TX farmers made a transition from cotton cultivation to that of peanuts. By 1940, much of the land previously planted in cotton was now devoted to peanuts [18]. The advent of World War II and the Japanese capture of coconut-producing islands of the Pacific necessitated greater US production of peanut oil because it replaced coconut oil for the production of glycerin, which was required in the manufacture of munitions. Spurred by the promotion of peanut production by the US Department of Agriculture, TX farmers tripled their peanut production during the war [19]. Texas ranked second among the states in acres planted in peanuts for the decade 1938–1948, with an average of 645,200 harvested acres and an average yearly yield of 284 million pounds [18].
During the 1930s and 1940s, the dominant reservoir animals for FBT were the brown and black rats, Rattus norvegicus and R. rattus, respectively, and the main vector was the Oriental Rat Flea, Xenopsylla cheopis [20,21]. Peanut cultivation in the American South in the 1930s and 1940s promoted rat proliferation, which likely contributed to an increase the number of cases of FBT [22]. Rat proliferation was not restricted only to areas of peanut cultivation because there were peanut processing facilities located throughout TX in the 1940s [18]. The annual number of cases of FBT peaked in Texas in 1945 at 1844 cases [23]. By 1946, 180 of the 254 Texas counties had reported cases [24] (Figure 1 and Figure 2). The overall incidence of FBT in Texas in 1945 was 27/100,000 persons. Other factors in the rise of FBT in Texas and the American South in the late 1930s and early 1940s included: the poor condition of rural and urban housing; poor environmental sanitation in urban areas; the inability to import insecticides and rodenticides that previously originated in military theaters of operation; and decreased civilian availability of pesticides due to wartime demands [22]. In 1944, Lavaca Co. (TX) had the highest incidence of FBT in the USA, at 438 cases per 100,000 persons [25]. FBT became a notifiable disease in TX in 1946 [26].
Starting in 1945, there was a massive public health campaign to eradicate rats and their fleas in the American South. The incidence of FBT in TX in a single decade reached its nadir in the 1960s, with only 246 cases reported from 1960 through However, by the late 1960s in Texas, FBT was undergoing a transition from the Oriental rat flea as the major vector to the cat flea [6,27].
From 1980 to 1984, there were 200 cases of FBT reported in TX, with four south Texas counties dominating, two adjacent counties along the Gulf Coast (Nueces Co. with 57 cases, and Kleberg Co. with 9 cases) and two counties in the Rio Grande Valley (Hidaldo Co., 39 cases, and Cameron Co., to the east, 22 cases). In Nueces and Hidalgo Counties, the incidence was 4.2 and 2.7 per 100,000 residents, respectively (see Figure 3) [28].
From 1990 to 1999, there were 307 cases of FBT in Texas [8] (Figure 4), occurring in 28 counties. Officially recorded cases of FBT in TX reached a nadir of 9 cases in 1994 [8]. The majority of FBT cases in TX in the 1990s occurred south of the 66 ºF isotherm (the isotherm is based on weather from 1981 to 2010) [29], the area climatically most favorable for FBT transmission. I refer to this area of higher FBT endemicity as the Lower Rio Grande-Texas Gulf Coast FBT Refugium; of the 307 total cases in this decade, 88.9% of cases occurred in the Refugium’s 13 counties and only 34 cases were seen outside the Refugium (11.1%). Another 7 cases occurred in San Patricio County, adjacent to the 66 ºF isotherm. If data from 1980-1984 is re-examined, 145 of the total 200 cases (72.5%) were reported from the Refugium counties (Figure 3) [28].
4. Typhus in Texas in the New Millenium: the 2003-2013 Texas DSHS Study
During 2003–2013, a total of 1,762 Typhus Group Rickettsioses (TGR) cases were reported to the TDSHS. During this period, the number of cases per year varied from 27 in 2003 to 222 in an average of 102 cases were reported annually during 2003–2007, less than half of the average number of annual cases (209) reported during the latter half of the period (2008–2013). The researchers were unable to discern the reasons for the increasing number of cases of TGR in TX in the last decade; they doubted that greater case ascertainment and reporting was the cause. Overall, TGR cases peaked in June and July; however, in south TX (< 28° N latitude), peaks occurred both in June/July and the other in December/January. The reason for this bimodal distribution of cases in south Texas was uncertain. Geographic expansion of FBT within TX also occurred during the 2003-2013 period. In 2003, cases were reported from nine counties in south Texas, but by 2013, cases had been reported from 41 counties, primarily in south and central TX [30].
The increasing number of cases of TGR in TX is even more apparent if we look back to the 1990s and compare to more recent data. There were 307 cases in TX in the decade 1990 to 1999, an average of 30.7 per year, with 26 counties reporting cases; 82% of the cases occurred in two Lower Rio Grande Valley and one Gulf Coast counties (Hidalgo, Cameron, and Nueces counties; 113, 21, and 117 cases, respectively) [8]. The TDSHS has released county-level FBT data from 2010 to 2019; in this manuscript, this data was analyzed for geographic patterns. In this ten-year period, there were 3750 cases (an average of 375 cases per year) reported from 91 TX counties. The purpose of the current manuscript is to derive a One Health Model to account for this increase in FBT cases in TX in the new millenium.
5. Building a One Health Model. The Epidemiology of Typhus in Texas, 2010-2019
The human population of TX is increasing at a rapid rate. In 1990, the population of the Lone Star State was 17 million; in 2019, the population stood at 29 million (US Census), a 70.7% increase. The urban population of TX grew even more in this period, at 83%; thus, the proportion of Texans living in a metropolitan area increased from 83.4 to 89.4% (Table 2).
The populations of the six counties with the highest number of FBT cases (Bexar, Cameron, Harris, Hidalgo, Nueces, and Travis) in the decade 2010-2019 increased 67.1, 62.7, 66.3, 126, 24.2, and 121%, from 1990 to 2019, respectively (Table 2). The rate of population growth of Hidalgo, Cameron, and Travis counties exceeded that of TX as a whole. Human population growth results in further urbanization/suburbanization, with effects on FBT reservoir/flea host populations and local climate (which will be described below).
A TX county is defined as urban if the population is greater than 50,000 [32]. From 2010-2019, 3394 out of 3750 total cases of FBT (90.5%) occurred in counties classified as urban (Figure 5). Compared to 1990-1999, cases of FBT have increased markedly in Bexar, Cameron, Harris, Hidalgo, Nueces, and Travis Counties in the period 2010-2019 (Table 3). These six urban counties had 2824 cases during 2010-2019, which comprised 75% of all cases in TX for that period. The increase in FBT cases in these growing urban counties far exceeded the number expected from human population growth alone. Likewise, in Orange Co., CA, cases of FBT were found in highly populated urban areas in the northern half of the county, with no cases recorded in the less urbanized south [33].
Humans are accompanied by their companion animals, which may act as reservoirs for FBT and blood meal and transport hosts for fleas. In 2014, it was estimated that there 7,163,000 pet dogs and 5,565,000 pet cats in TX [34]. At that time, the human population of TX was 27 million. Thus, there were 0.27 dogs and 0.21 cats per Texan. Assuming the popularity of dog and cat ownership has not significantly changed from 1990 to 2019, the estimated number of pet cats and dogs can be compared for the two time periods 1990-1999 and 2010-2019 (Table 4 and Table 5). There are no reliable estimates of the number of feral dogs in the U.S., but they are considered to be ubiquitous in the USA [35]. In 2009, Bexar County (TX) was estimated to have 100,000 stray dogs [36]; at that time, the human population of the county was 1.65 million (US census), giving a 17:1 human to stray dog ratio. If other Texas urban counties are similar, the stray and total dog populations can be estimated for the six Texas counties with the highest number of FBT cases (see Table 4).
Likewise, in 2014, it was estimated that there were 5,565,000 pet cats in TX [34], or 0.21 cats per Texan. The number of stray and feral cats in the USA is also not known. Estimates are one-third to about equal of the owned cat population [37,38]. In warmer climates, there may be higher numbers of free-roaming cats, because in areas with mild winters, females may produce up to three litters per year [37]. If a conservative estimate of 40% of the pet cat population is used, this would be approximately 2.4 million free-roaming cats in TX in Table 5 provides an estimated number of stray and feral cats in TX and the six counties with highest number of FBT cases.
Thus, the total dog and cat populations of the top six Texas counties for FBT cases are estimated to be 3.06 million and 2.74 million, respectively for the years 2014/2015 (mid-point of the decade), as compared to 1.99 million and 1.78 million, respectively, for the years 1994/1995 (mid-point of that decade). Thus, there were about 1 million additional dogs and 1 million additional cats in the six high incidence counties in the 2010s compared to the 1990s. This would have increased the number of potential reservoirs for FBT and sources of blood meals for adult and larval flea survival, and provided an additional armada of transport hosts for fleas to infiltrate human yards and households.
In addition to its urban predominance, FBT also has a distinct climatic distribution in the state of TX. Climatic conditions may affect: the rapidity of the flea’s life cycle, the survival of each of the stages of the life cycle; flea mating, feeding, and defecation rates; the extent of flea colonization with endosymbiotic organisms; the percent of reservoir animals infested; and the flea burden (these factors will be discussed in detail in this paper and the subsequent paper in the series). In the 1940s in the USA, FBT was most prevalent in areas where the average January temperature was above 40 °F and the average relative humidity in July at noon is above 37% [39]; these were the conditions most favorable for the survival of the Oriental rat flea (Xenopsylla cheopis), the most significant vector at that time [40]. Seasonally, the lowest numbers of FBT cases were reported in the fall and winter and the greatest in July through September. Except for 1940, the number of cases of FBT reported in the USA increased each year since recognition of the disease in the early 1920s until 1945 [22]. The decrease in cases in 1940 may have been due to an unusually cold January in that year. There was no association between precipitation and the incidence of FBT [41]. The main vector in TX is now the cat flea, but climatic factors will also likely affect the current distribution of FBT in the state.
Texas has a south-to-north gradient of decreasing mean annual temperature and increasing seasonal temperature variation and an east-to-west gradient of decreasing precipitation [29,42]. Based on the temperature, precipitation, humidity, vegetational characteristics, and seasonal weather changes, the National Climatic Data Center divides TX into 10 climate zones (Figure 6; Table 6) [29]. Flea-borne typhus cases in TX in the period 2010 to 2019 show a very distinct distribution in these climate zones. In the northern “cooler” half of TX, proceeding west to east from areas of lower precipitation to higher, are the divisions: (1) High Plains; (2) Low Rolling Plains; (3) Cross Timbers; and (4) Piney Woods. This northern set of climate divisions had only 229 reported cases of FBT during 2010-2019, only 6.1% of the total TX cases (see Table 6 and Figure 6). The divisions comprising the central belt across TX are, from west to east: (5) Trans Pecos; (6) Edwards Plateau; (7) Post-Oak Savannah; and (8) Gulf Coastal Plains. These four divisions had 1, 56, 1465, and 408 cases of FBT in the period 2010 to 2019, respectively, amounting to 51.4% of the total. The two most southerly climatic divisions, from west to east, are the South Texas Plains and Lower Rio Grande Valley, with 227 and 1364 cases, respectively (42.4% of the total). Thus, TX FBT cases are focused in the Post-Oak Savannah and Lower Rio Grande Valley climate divisions (1465 and 1364 cases, respectively), which account for 75.5% of the total cases (Figure 6). The Gulf Coastal Plains and South Texas Plains account for another 16.9% of the total cases. (For average temperature and rainfall data for each climate region, see [29]). Compared to 1990-1999, in 2010-2019, FBT increased in eight of the ten climate regions, especially in the Gulf Coastal Plains and Cross-Timbers (136- and 49.5-fold, respectively).
From the 1960s through the 1990s, FBT mostly retreated to its subtropical refugium deep in South TX (see Figure 3 for 1980-1984 and Figure 4 for 1990-1999), with the majority of cases occurring south of the 28th-parallel, approximately the northern border of Nueces, Jim Wells, Duval, and Webb counties (Figure 4), which roughly corresponds to the 66 °F isotherm (based on the average annual temperature for 1981 to 2010) [29]. I refer to this area of TX south of the 66 °F isotherm as the Rio Grande Valley-Texas Gulf Coast FBT Refugium (Refugium for short). Presumably, this is the area of TX that was most climatically favorable for flea development and infection transmission prior to the 2000s [43]. In the 1990s, 273 out of the 307 (88.9%) of the FBT cases in TX occurred in the Refugium’s 13 counties, which comprise only 6.8% of the area of Texas; only 34 cases (11.1%) were reported outside of the Refugium. By contrast, in the decade 2010-2019, 2238 cases (60%) occurred in the Refugium; 1512 cases (40%) were reported outside the Refugium (45-X higher than the 1990s). Thus, in 2010s, the areas favorable to flea proliferation and infection transmission significantly expanded outside the previous most favorable zone. A graph of the number of cases of FBT cases occurring in TX from 1944 to 2019 is shown in Figure 7, it is apparent that there was a steep drop in the number of cases when rodent and flea control programs were implemented in the mid-1940s and then a steady low level of cases from 1960-2003 and then a rise in the number of cases in the new millennium.
In the Rio Grande Valley, it is likely that both climatic factors and socioeconomic conditions contribute to the high incidence of FBT. In 2019, the McAllen-Edinburgh-Mission Metropolitan Statistical Area (MSA) was the poorest in the country, with a 27% poverty rate. The Brownsville-Harlingen MSA ranked as the fourth poorest, with a 25.6% poverty rate [44]. Yao et al. analyzed the spatial distribution of 555 cases of FBT reported from 1996 to 2006 in 18 counties in South Texas [45]. Most FBT cases were seen in urban areas with relatively high human population densities and low household incomes and home values, i.e., areas of lower socioeconomic status (SES). Thus, Yao and coworkers proposed several demographic factors that contribute to a higher prevalence of FBT in these areas: urban areas provide increased harborage and the availability of pet food and water sources for opossums; higher numbers of companion cats and dogs and strays in urban areas may act as reservoirs of FBT, blood meal sources for flea proliferation, and mechanical carriers of fleas; and there may be less attention to the flea control of pets is areas of lower socioeconomic level [45]. Environmental hygiene is likely be lower in low SES areas, resulting in increased harborage and food sources for opossums and stray/feral dogs and cats. The epidemiology of FBT in TX in the period 1990-2019 presents an example of infection re-emergence, the reappearance of a known infection after a decline in incidence. Infection re-emergence is often dependent on human influence on ecological systems, causing habitat alteration and changes in species assemblage and contact rates between vectors/hosts and humans. The spread and persistence of re-emerging pathogenscan be precipitated by land-use changes, such as urbanization, and by proliferation of reservoir and vector populations [46]. The re-emergence of a vector-borne infectious disease in a particular geographic area is the result of the responsible pathogen responding to changes in the vector and/or host ecologies [47].
Urbanization and suburbanization may have multiple effects on the epidemiology of FBT. First, the pet dogs and cats of the populace act as an ongoing source of stray and feral animals that may act as flea hosts and infection reservoirs. Urbanization/suburbanization also alters wildlife communities, generating low biodiversity with increases in the abundance of generalist urban-adapted species, such as opossums, skunks, and raccoons [46,48,49], which are cat flea hosts and reservoirs for flea-transmitted pathogens facilitating parasite spillover into natural areas [50]. A close proximity between natural and urban/suburban habitats increases exposure of wildlife to feral and domestic animals which facilitates C. felis infestation of wildlife.
6. The Rickettsiae
Flea-borne typhus is caused by R. typhi and R. felis, obligate intracellular Gram-negative bacteria in the Class Alphaproteobacteria, Order Rickettsiales [51]. Rickettsia felis was not definitively identified until 1990, when it was observed in the cytoplasm of midgut cells of cat fleas collected from feral cats and differentiated from R. typhi by the polymerase chain reaction (PCR) [52]. In humans, infection with either R. felis or R. typhi produces mutual serologic cross-reactivity with routine indirect fluorescent antibody testing [53]. Because of this cross-reactivity, it is uncertain if the causative organism for all the prior clinical and epidemiologic studies on FBT was R. typhi or R. felis or both [54].
Although other pathogenic rickettsiae, such as R. rickettsii and R. prowazekii, adversely affect the fitness of their arthropod hosts (ticks and the human body louse, respectively), R. felis and R. typhi are not thought to influence flea longevity and fecundity [55,56,57]. This is due to the ability of these rickettsiae to evade the flea immune response [58] and indicates a long evolutionary association of these bacteria with the flea [59]. Rickettsia felis is known to undergo transovarial transmission based on its detection in freshly deposited cat flea eggs [60] and trans-stadial transmission based on observing R. felis in newly emerged unfed adult cat fleas [61]. A population of cat fleas is able vertically maintain R. felis for at least 12 generations without feeding on an infected host, although the infection rates decline from 63% to 2.5% [57]. The host blood source may also influence the sustainability of R. felis in cat fleas [57]. The prevalence of R. felis within cat fleas also varies between colonies [62]. Other investigators have postulated that the high variability of the vertical transmission of R. felis in C. felis indicates that this organism does adversely affect flea fitness; thus, some degree of horizontal transmission is necessary for long-term pathogen maintenance [58,63,64,65]. In horizontal transmission, an uninfected flea acquires infection by taking a bloodmeal from a rickettsemic host [66]. For R. typhi, ingestion of only a few rickettsial organisms is sufficient to result in active infection of the flea [67]. The ingested rickettsiae then enter the midgut epithelium, initiating a process of extensive replication [68]. After replication within the midgut cells, rickettsiae are released into the gut lumen and excreted in the feces [3,69]. Traditionally the route of transmission of R. typhi from flea to host was thought to be by the posterior station or stercorarian transmission (the inoculation of infected flea feces into cutaneous lesions or mucus membranes [3,64]. Flea feces become infectious approximately 10 days after the fleas acquire the infection and the fleas remain infectious for the remainder of their lives [69]. Unlike R. typhi, infection of blood-feeding fleas by R. felis requires a higher infectious dose [63]. The transmission of R. felis in the absence of flea feces (i.e., oral transmission) has been demonstrated by the induction of an antibody response in cats briefly fed upon by R. felis-infected cat fleas in which no fecal contamination of the cat’s skin was evident [70]. However, it is uncertain if R. felis is transmitted by the posterior station route, although R. felis can be found in the feces of infected fleas [4,63,71].
Another mode of transmission is the inhalation of infected flea feces, which may remain infective for up to nine years [66]. More recently, the observation of R. felis in the salivary gland of C. felis by electron microscopy, PCR, and immunofluorescence suggests the potential for infection of vertebrates through blood feeding [72,73]. Simple mechanical transmission of rickettsiae may also occur because the organism is released from contaminated mouthparts into the skin of the host during probing by the flea [64,74]. The transmission of R. felis in the absence of flea feces (i.e., oral transmission) has been demonstrated by the induction of an antibody response in cats briefly fed upon by R. felis-infected cat fleas in which no fecal contamination of the cat’s skin was evident [70]. Similarly, Azad and Traub showed R. typhi could be transmitted to rats by X. cheopis by the oral route [75]. Flea-to-flea transmission of rickettsiae may occur by the process of cofeeding; i.e., when there is simultaneous feeding of multiple fleas on one host, pathogens are transferred from an infected flea to an uninfected flea that is feeding nearby due to the regurgitation of rickettsiae into the bite siteby the former. Thus, host rickettsemia is not necessary for flea-to-flea transmission [4]. Flea larvae feeding on R. felis-infected feces, flea eggs, or other larvae did not acquire the infection [57].
Multiple studies have documented the simultaneous presence of R. typhi and R. felis in the cat flea and the Oriental Rat Flea X. cheopis [33,56,76,77]. Cat fleas can also be experimentally infected with both species [78]. However, it is not clearly known if either pathogen has any advantage for acquisition, persistence, or transmission by fleas [33].
In ticks, interference occurs, in which the establishment of one rickettsial species inhibits the transovarial transmission of a second species [79]. However, if R. felis-infected cat fleas were fed blood containing R. typhi for nine days, cat fleas contained both pathogens; however, infection rates occurred at a lower prevalence than with either single infection, indicating that flea infection with R. felis may inhibit R. typhi infection [78]. It is unknown if R. felis inhibits vertical transmission of R. typhi or transmission to a susceptible vertebrate host [71].
In studies from different locales, the prevalence of R. felis infection of cat fleas ranges from 5% to 45.8% [80,81]. Based on PCR studies conducted on fleas, the global distribution of R. felis does not necessarily coincide with that of R. typhi; the latter is less commonly observed [33,56,71,82,83,84]. For example, in a study conducted in Orange and Los Angeles Counties (CA) in 2007-2008, about 48% of cat fleas tested positive for R. felis but less than 2% were infected with R. typhi [77].
Both species of rickettsiae display “pathogen flexibility” or polyhostality, the ability of a pathogen to infect more than one host and more than one order of hosts; data will be presented in part 2 of this series that these rickettsiae can infect dogs, cats, and opossums. Polyhostality increases the likelihood that a pathogen will be emergent or re-emergent [49,85,86]. Pathogens that spillover to the human population typically involve a network of species, which is the case for R. typhi and R. felis. The phylogenetic and ecological host breath of a pathogen increases its zoonotic potential, especially if both domestic and wild hosts are infected [47,87]. Multi-host pathogens, such as R. typhi and R. felis, are typically associated with multi-host vectors, such as the cat flea [88].
However, some researchers have questioned the pathogenicity and epidemiologic significance of R. felis [89,90,91]. In 2016, Billeter and Metzger collected cat fleas from cats in Los Angeles Co. (an endemic area for human FBT) and in Sacramento and Contra Costa Counties (non-endemic areas). PCR confirmed the presence of R. felis in cat fleas from both the endemic and non-endemic areas; R. typhi was not detected. Because R. felis was widespread in cat flea populations in both FBT endemic and non-endemic areas, the investigators concluded that it is unlikely that R. felis is a major cause of human FBT in California [89]. Although R. felis has been reported as a cause of fever in Africa, it has also been detected from skin swabs from afebrile patients [92], suggesting this organism is a commensal [90]. Nevertheless, over 100 cases of FBT (sometimes designated as flea-borne spotted fever (FBSF) to distinguish it from the disease caused by R. typhi) have been specifically ascribed to R. felis, based on PCR and serologic tests that are able to differentiate R. felis and R. typhi infections [54,90]. These cases have been documented in the Americas, Asia, Africa, the Pacific, and Europe [93]. Based on limited studies, R. typhi is more common in humans in TX [53,94].
6.1. Rickettsia typhi
Rickettsia typhi belongs to the Typhus Group of the Family Rickettsiaceae. After the entry of the rickettsiae through the skin, respiratory tract, or mucous membranes, they disseminate hematogenously or by the lymphatics and localize to the vascular epithelium of multiple organs. Endothelial injury is the principal mechanism of rickettsial pathogenesis. Rickettsiae adhere to endothelial cells with their outer membrane proteins (Omp); OmpA and OmpB are found in the Spotted Fever and the Transitional Groups, whereas Typhus Group rickettsiae lack OmpA [95]. Eight different strains of R. typhi have been characterized [96]. Rickettsia typhi has been detected in at least twelve species of fleas of nine genera [97]. Rickettsia typhi can be ingested by fleas during a blood meal and transmitted vertically to progeny and horizontally to other fleas by cofeeding [3].
6.2. Rickettsia felis
Rickettsia felis is now placed in the Transitional Group of the Rickettsiaceae, based on the discovery that it possesses a conjugative plasmid [98]. The first reported human case of R. felis infection was reported from TX in 1994 [99]. Although R. felis has been detected in over 40 species of hematophagous arthropods globally, the only known competent vector is the cat flea [4,71,100]. The cat flea can acquire and horizontally transmit multiple genotypes of R. felis [65,80,101,102], but how the genotype affects virulence and transmissibility is unknown. Also, the ability of R. felis to infect C. felis depends on the specific clade of the cat flea [103]. The clinical presentation of R. felis infection in humans is similar to that of R. typhi [71], but may be milder [93].
7. The Cat Flea and Its Many Hosts
The cat flea was discovered as a vector of FBT to humans in 1942 when four persons in Austin, TX, became ill with typhus after exposure to cat fleas on a kitten [104]. However, at that time in the United States the cat flea’s significance as a vector was overshadowed by the Oriental rat flea. In 1969, the cat flea was incriminated in additional cases of FBT in TX [27] and in 1970 Adams and coworkers implicated the cat flea in the epidemiology of FBT in the Los Angeles (CA) area [105].
Most species of fleas infest only a single order or family of mammals [106]. Of the 2500 species of fleas [107], only nine species are considered indiscriminate feeders; in North America, these are C. felis, Pulex simulans, P. irritans, Echidnophaga gallinacea, and Nosophyllus fasciatus [106]. The former four species are known to harbor both R. typhi and R. felis [3] and N. fasciatus is a vector of R. typhi [69]. The cat flea possesses genal (head) spines and pronotal (thoracic) combs to facilitate adherence to the fur of its host but it is not specialized to the pelage of a specific host [106]. Cat fleas feed on a wide diversity of mammals; 138 species have been found to harbor cat fleas. The principal host groups for cat flea infestation are: felines (Felidae); foxes and dogs (Canidae); possums and opossums (Phalangeridae, Didelphidae); skunks (Mephitidae); porcupines (Hystricidae); mice and rats (Muridae); shrews (Soricidae); hedgehogs (Erinaceidae); and weasels (Mustelidae). Thus, they among the most host generalist of all ectoparasites, a characteristic that facilitates host switching at the human-wildlife interface [50]. Those host species that frequently enter anthropogenic habitats are at highest risk to acquire cat fleas. However, all hosts are not equally advantageous for flea survival. For example, cat fleas fed on rats consumed more blood, produced more eggs, and had higher offspring female-to-male sex ratios than those fed on mice [108]. Blakey and coworkers compared dog blood to cat blood and found that cat fleas consume higher quantitites of dog blood and produce greater numbers of eggs [109].
In the USA, multiple studies have found that C. felis is the most prevalent flea species on dogs, cats, and opossums [110,111]. Other North American wildlife species that harbor C. felis include coyotes, red and gray foxes, bobcats, skunks, raccoons, ringtails, ferrets, pumas, and several species of rodents [96,110,111,112]. Disease transmission within an ecological community is more likely when the vector parasitizes a diversity of host species [113]. Habitat encroachment by urbanization and suburbanization increases the potential contact between wildlife and flea-infested human-associated hosts. Thus, anthropogenic habitat use by wild and feral domestic mammals is a strong positive predictor of C. felis infestation. The odds of cat flea infestation for anthropogenic habitat-using species is increased by 256% compared to species that do not use anthropogenic habitats [50].
The cat flea serves as a bridging vector between sylvatic hosts (opossums, racoons, skunks), or hosts with intermittent human contact (free-roaming dogs and cats), and domestic dogs and cats [68,69]. The broad host range of the cat flea and the ability of a developing flea to remain in its cocoon until a host is present are both adaptations to survive conditions of irregular host availability [114]. The domestication of cats and dogs by humans has also contributed to cat flea survival by providing favorable locations within human dwellings to complete its life cycle year-round [115].
The proclivity of the cat flea to attack both cats and dogs has led to its widespread geographic distribution [116]. It has been called “the most pervasive flea species on Earth [50].” The cat flea, being a generalist, is less susceptible to environmental disruptions that may affect the populations of a single host. Host-opportunistic fleas generally display a greater tolerance to climate and environmental variability and thus are more geographically widespread [117,118] and attain higher local densities by exploiting the ability to parasitize multiple hosts [107]. Increased abundance of generalist vectors is strongly associated with increased pathogen transmission [113,119].
In a multi-host pathogen system, the dynamics of the zoonotic agent involves two phases: (i) transmission between maintenance and/or non-maintenance host species (wildlife and/or domestic) and (ii) spillover transmission to humans from the maintenance community. The most important host species communities for FBT in the United States in this case are opossums, and both domestic and free-roaming dogs and cats. For FBT, the force of infection from flea to human depends on the prevalence of infection in the flea vector and the rate of contact between humans and the vector (which will depend on the abundance of the vector) [46].
The cat flea meets several criteria of a generalist vector able to successfully exploit the environmental changes wrought by humanity in the 21st century USA: wide geographical distribution; capacity to feed on a range of hosts; capacity for zoophily and anthropophily; ability to exploit peri-domestic and peri-urban settings; and ability of the larva to utilize indoor and outdoor habitats [120]. The adult flea’s requirement for multiple feedings per day potentially increases its ability to infect multiple hosts [121]. However, the transmission efficiency from flea to host is low (rates of 10–30% for R. felis [74]) and transovarial transmission is likely the primary mechanism of rickettsial persistence in the flea population. The cat flea also avidly bites humans [122,123]. Thus, the cat flea has emerged as the most important vector of FBT in recent decades in TX and CA [68,112,124].
8. Flea-Borne Typhus and Climate Change: General Considerations
Climate change refers to long-term shifts in weather conditions resulting from human activities [125]. Temperatures in TX have increased almost 1.5 ˚F (0.83 ˚C) since the beginning of the 20th century. Although there is no overall trend in the number of hot days, the number of warm nights was particularly high during the 2010s. The urban heat island effect has amplified this problem in TX cities [126,127].
Temperature, precipitation, and humidity may affect the reproduction, lifecycle, behavior (infestation and biting rates) and population dynamics of arthropod vectors [128]. Thus, climate change has been implicated in affecting the epidemiology of multiple vector-borne infections, including malaria, leishmaniasis, plague, and dengue [129]. Increasing temperatures may increase pathogen transmission by decreasing vector generation time, increasing vector population growth rate, decreasing vector winter mortality, decreasing the pathogen extrinsic incubation period, and lengthening the seasonal duration of vector activity. However, there may also be decreased vector longevity at higher temperatures [16,130]. Ctenocephalides felis is already globally widespread; climate change will probably not affect its geographic distribution. However, climate change effects on the cat flea life cycle may increase its density within its established range [116,131].
At many locations throughout its global distribution, FBT tends to be seasonal, with the highest incidence rates observed in the warmer months and fewer cases seen in the winter [12,132,133,134]. These results are in accordance with the seasonality of flea infestation, which tends to be higher during warmer months [135,136,137]. For example, in Laos, FBT is more common during the hot, dry years following El Niño [138]. In Taiwan, from 1992-2009, a significant correlation was found between average monthly temperature and number of cases of FBT [139].
Because fleas are poikilothermic, temperature will affect their life histories [140,141]. Weather and climate determine the temperature and relative humidity of the host’s den, which is affected by the ambient temperature, precipitation, and vegetation cover. These conditions will determine the developmental durations and survival of both pre-imaginal and adult fleas [107]. Temperature also greatly affects rickettsial growth within cat fleas; R. typhi grows at levels > 1000-fold higher at 24 and 30 ºC compared to 18 ºC [142].
The maintenance cycle of FBT in the environment depends on the thermal tolerance of the bacteria, the flea hosts (opossums, cats, and dogs), and the fleas. If the thermal tolerance for any component of the system is exceeded, the maintenance cycle will be disrupted, and thus the number of cases of human FBT is expected to decrease. However, even during the extreme TX drought and heat wave year of 2011, the number of cases of human FBT increased, indicating that the limits of thermal tolerance of the components of the maintenance cycle were not exceeded, even under the extreme weather conditions of Up to that time, in that year TX had the highest average June-to-August temperatures and the least rainfall since data was first collected in 1895 [143]. Despite these sweltering conditions, TX cases of FBT more than doubled in 2011, to 286 compared to the 135 cases in the previous year [5]. In 2023, TX once again endured record high temperatures and drought conditions. Although the data on FBT cases are not yet available, it will be interesting to see how these extreme conditions affected the epidemiology of FBT for that year.
High levels of rainfall decrease flea infestation, either because the eggs and larvae wash away from the hosts or that rain promotes microbial growth that is harmful to the pre-imago stages of the flea lifecycle [144]. Conversely, cycles of drought may restrict the distribution of the cat flea in certain areas [145].
All six high FBT prevalence counties showed increases in the overall minimum temperature (Tmin), the mean temperature (Tm), and the maximum temperature (Tmax) for the decade 2010-2019 compared to 1990-1999, except Travis Co. had a slight decrease in the Tmin (Table 7). Although a less than 1 °F temperature increase may not seem sizeable, this effect was, on average, operative over an entire decade and temperature affects multiple events in the FBT transmission cycle: rickettsial replication in the flea, infestation rates, biting frequency, flea mating frequency, and the duration of the off-host flea life cycle, as will be described in subsequent sections of this paper. Since climate change effects are often greater at higher latitudes, the temperatures were also obtained for Tarrant Co. (major city Fort Worth) in North Texas, a county in which the number of cases of FBT increased from one in the 1990s to 93 in 2010-As predicted, Tarrant Co. had a larger increase in the average Tmin, Tmean, and Tmax compared to the other six more southerly high FBT incidence counties.
9. The Urban Heat Island Effect in Texas
In the continental USA, urban areas are on average 2.9 °C warmer than surrounding areas due to replacement of the natural soil and vegetation with artificial surfaces [146,147]. There are reduced daily temperature fluctuations due to higher nocturnal temperatures in cities compared to the countryside [148]. The urban heat island effect is complex and depends on vegetation cover, seasonal effects, and proximity to bodies of water. In Dallas from 2001 to 2011, the July average was 2.4 °C higher than the surrounding rural area [149]. In San Antonio, from June 1 to September 30 during the years 2002 to 2008, the nocturnal temperature was 6-7 °C higher than the surrounding area [127]. Urban sprawl amplifies the heat island effect. For example, the Austin metro area population increased from 655,000 in 1993 to 1.43 million in 2011 (US census data); in that period, the average surface temperature of Austin rose by 4.7 °C [150]. The urban heat island effect has definite biological consequences; for example, there is increase in the vegetation growing season by about 15 days in urban areas relative to adjacent rural areas [151]. Thus, the urban heat island effect may prolong the period of flea activity during cooler months and at night.
10. The Cat Flea Life History and the Effects of Temperature and Humidity: General Aspects
To understand how climate change may affect the incidence of FBT in TX, it is necessary to consider the effects of temperature on each stage of the cat flea life history: host-seeking and infestation, feeding, mating, oviposition, hatching of the eggs into larvae; feeding by the larvae; pupation; and eclosion. In terms of its life history, the cat flea can be described as a fur flea (versus a nest or fur/nest flea). Fur fleas spend more time on the host and thus are potentially exposed to higher fluctuations in ambient temperature and humidity compared to nest fleas [118]. Gracia and coworkers have proposed that C. felis has a wider environmental tolerance than C. canis or P. irritans [152].
10.1. Host Seeking and Infestation
After emergence from its cocoon, the imperative for a newly emerged adult flea is to locate a host. Higher temperatures promote jumping activity in fleas, which is necessary for host acquisition [107]. Although the direct transfer of fleas from one host to another does occur, the more common route of infestaton is the acquisition of newly emerged fleas from the environment [110]. Data from various locales indicate that cat flea infestation of cats, dogs, and opossums is more common during warmer months [76,110,135,136,152,153,154,155,156,157,158,159,160,161,162,163,164]. A specific example of the effect of temperature on flea infestation was seen in Germany during a heat wave in 2003 in which the prevalence of flea infestations of dogs and cats doubled [136]. In a Spanish study from 2002-2004, a positive relationship was found between C. felis abundance on dogs and the mean annual temperature, without effects of rainfall [152]. Thus, warmer weather due to climate change may increase the prevalence and intensity of flea infestation [116].
Female and male fleas typically take their first blood meal within two hours of emerging from their cocoons and then feed 4–10 times per day thereafter. There is a specific temperature threshold that must be met to stimulate the initial feeding [165]. For males, a bloodmeal is essential to initiate mating behavior [166]. Furthermore, the epididymis of the newly emerged unfed male cat flea is obstructed by folded columnar epithelium, but the initial blood meal unblocks the epididymis, permitting sperm transfer [167].
The feces of adult fleas constitute the major component of the larval diet. The maggot-like larvae also consume flea eggs and injured flea larvae [168]. At higher temperatures, there is a higher rate of flea feeding and defecation [107]. Also, the rate of digestion is accelerated at higher temperatures and is associated with lower energy expenditure for digestion, allowing fleas to allocate more metabolic resources to other activities, such as mating or egg production [169].
Increased frequency and duration of feeding may increase the probability that a flea will ingest infected blood during a period of host bacteremia [170] or increase the likelihood of acquiring infection from another flea by cofeeding. Farhang and coworkers found higher fecal output from cat fleas that fed ad libitum on rats at 24 °C and 30 °C versus 18 °C [142]. Increased defecation increases the amount of flea feces in the flea nursery, providing nutriture for larval growth and development. Larger larvae are less prone to desiccation due a smaller surface to volume ratio [171]. Thus, if larvae are better fed because of a more bountiful fecal food supply, they may be more likely to survive. Furthermore, a well-fed larva produces an adult with higher fat reserves better able to withstand a period without access to a blood meal [107].
10.2. Mating and Oviposition
Feeding by adult fleas, induced by warmer temperatures, is a stimulus for mating [172]. Temperature also determines mating frequency in cat fleas. Under experimental conditions, at 27 °C, no mating occurred. Between 34 and 42 °C, mating commenced, with most of copulation occurring at 38 °C. However, there were no mating attempts at 44 °C [166]. Male cat fleas may inseminate multiple females within 24 hours after a blood meal. Multiply-mated female cat fleas have higher fecundity (total eggs) and fertility (egg viability) than singly mated females because with each mating, the female may acquire nutrients, oviposition stimulants, and viable sperm, all of which enhance their reproductive success [172]. Because R. felis may be transmitted sexually between fleas [58], increased copulation frequency and copulation between different pairs of fleas may also promote the transmission of rickettsiae between fleas. Virgin female cat fleas lay nonviable eggs after a blood meal; however, within 24 hours after male fleas are introduced, they began ovipositing viable eggs and quadruple their egg output [173].
Female fleas decrease egg production during colder weather and resume egg laying when environmental conditions become favorable for larval development [174]. The number of eggs produced by the adult female flea correlates with volume of blood consumed [166]. Larger females (presumably from feeding more frequently) have higher lifetime fecundity [59,175]. It has been proposed that male fleas with larger body size have increased mating success [169]. Adult flea longevity likely increases with larger body size [176], allowing fleas to perform a greater number of matings during their lifetimes [172]. A female cat flea produces about one egg an hour while on the host. The eggs readily fall from the host’s fur, accumulating in areas where the host spends the most time [110,177].
10.3. Egg Hatching and Larval Development
The duration of the developmental period and the survival of preimaginal fleas are highly dependent on microclimatic conditions [178]. Thus, flea abundance is strongly influenced by off-host environmental conditions [107], because only 1-5% of the flea population (all stages) live on their host at a given time, with the remainder dispersed around the resting or feeding areas of the host [179].
The eggs of C. felis hatch within 1.5 to 6 days following oviposition. Temperature and humidity determine egg survival and the incubation period. At 35 °C and 70% relative humidity, most of eggs hatch within 36 hours; at 13 °C, egg hatching requires six days [180]. Optimally, the flea larvae development occurs in protected microenvironments that combine moderate to warm temperatures, high humidity, and an ample supply of protein-rich flea feces and infertile eggs for larval nutriture [181]. The development of the eggs and larvae is restricted by temperatures outside a range of 4-35 °C and by a relative humidity <50% [182]. The high FBT-incidence cities of TX all have sufficient humidity for optimal flea larval development. The average relative humidity levels of Brownsville, Houston, Corpus Christi, San Antonio, and Austin are 90, 89, 89, 83 and 83%, respectively [183].
An underground burrow is ideal as a flea nursery because it is warmer in the winter and cooler and more humid during hot weather. The larvae develop through three stadia and the survival and rate of development of each stage depends on the temperature and humidity. The duration of the larval stage is 5-11 days, depending on the temperature, humidity, and food availability [180]. The highest rate of emergence (82%) of adult cat fleas occurs when the larvae have access to feces and nonviable eggs compared to feces alone (47%); feeding on nonviable eggs also shortens the duration of larval development [172]. The number of non-viable eggs depends on mating frequency, which is temperature-dependent [166]. The larval stage is highly sensitive to low humidity due to the inability of flea larvae to absorb atmospheric water at low humidity levels and to withstand respiratory water loss [180,184].
10.4. The Temperature-Driven Infestation--> Feeding--> Reproduction--> Larval Trophic Cascade
Thus, higher environmental temperatures for newly emerged adult fleas promote an Infestation → Feeding → Reproduction → Larval Trophic Cascade. After eclosion, increased temperature promotes active host questing by the newly emerged adult flea. Once a host is acquired, higher temperatures stimulate feeding. Feeding and higher temperatures stimulate mating, which leads to the production of both viable and nonviable eggs, with the latter serving as a food source for larvae. Increased feeding leads to larger adult body size, which is associated with higher fecundity. The more frequent feeding due to higher temperatures leads to greater adult flea feces production. Greater availability of feces and nonviable eggs in the flea nursery promotes accelerated larval development and increased fitness.
10.5. Pupation and Pupa to Adult Eclosion
The pupal stage is less sensitive to temperature than the larval stage, with its duration equal from 16 to 27 oC. However, a temperature of 8 oC for 20-days is lethal as well as > 35 oC [185]. During unfavorable conditions (too cold, hot, or dry), cocoons enter diapause until environmenta circumstances improve [117]. The ability to survive for extended periods in the cocoon is important for species such as C. felis, which infest mobile hosts that may not frequently return to the same nest or burrow. Increasing temperatures and mechanical pressure stimulate emergence of the adult [186]. The pupae are the most environmentally resistant stage of the life cycle and can survive for up to six months awaiting a host [165].
10.6. Overall Temperature Effects on the Cat Flea Life Cycle
Thus, at higher temperatures, fleas progress through their life cycles at a faster rate, i.e., eggs hatch faster and the duration of the larval stage of fleas is reduced. In one study conducted in north central Florida (a latitude comparable to South TX), fleas survived all year outdoors. In June and July eggs developed into adults in 20–24 days whereas in the winter it took 36–50 days. The immature stages survive frosts in protected microhabitats. From September to November, larval survival was up to 85% [171].
11. Endosymbionts of the Cat Flea
Steinina ctenocephali is a temperature-dependent endosymbiotic protozoan of the cat flea alimentary canal. Flea larvae infected with S. ctenocephali develop faster than an uninfected control group [187]. Fleas were more likely to be infected with this protozoan during warmer seasons [188].
Bacteria of the genus Wolbachia are endosymbionts of insects that may affect their reproduction, longevity, and vector competence and efficiency [103]. Infection of insects by Wolbachia may also affect their thermotolerance and thermal preferences [189], which may be relevant to rickettsial replication in the flea and flea behavior. Three Wolbachia strains have been isolated from C. felis [103]. It is uncertain how Wolbachia may affect R. typhi/R. felis acquisition, maintenance, and/or transmission in C. felis, so this is a question that needs to be investigated in areas of high FBT incidence [103,190]. In a study of cat fleas from Malaysia, Tay found that R. felis was found only in Wolbachia-positive fleas [191]. Wolbachia endosymbiosis of insects displays a complex temperature dependence [192].
12. The One Health Model of Flea-borne Typhus Epidemiology in Texas in the Early 21st Century
Figure 8 summarizes the One Health Model of FBT epidemiology in TX in the early 21st century. In the lower right corner, human population growth leads to increasing urbanization and suburbalization. Human population growth causes a warming climate, along with an urban heat island effect. Warmer weather promotes flea infestation of host mammals, feeding, and defecation. It also accelerates the flea life cycle and increases rickettsial replication with the flea. Increased rickettsial replication facilitates transmission of rickettsiae to host animals and humans. Human population growth also indirectly increases the population of dogs and cats, both domestic and free-roaming, that can act as flea hosts and possibly as reservoirs. Opossum populations also increase in an urban environment [26]. Opossums will enter yards and pass fleas to dogs and cats, which, in turn, may spread infected fleas into human domiciles. Control of opossum and stray dog and cat populations and the use of flea control products on pets can serve to suppress the transmission of FBT to humans.
13. Limitations of the Data
The reliance on serologic testing for the diagnosis of FBT has a number of pitfalls that likely result in an underestimation of the true number of cases of FBT in TX. Anti-rickettsial antibodies are present in less than 20% of patients at seven days after the onset of illness [91]; it may take up to 14 days for antibodies to develop, and confirmation of infection requires a 4-fold rise of antibody titers between acute and convalescent periods [193]. If patients are lost to follow-up evaluation after their acute illness, a confirmatory convalescent serologic test will not be obtained. Also, convalescent testing may not be performed if the medical practitioner seen at a follow-up appointment deems that the test is no longer important or if the patient does not have the means to afford the test. Cases of FBT are identified only after a patient seeks health care, a serologic diagnosis is made, and the data is captured by the reporting of serologic tests; thus, mild cases that did not come to medical attention or cases treated empirically without serologic testing being performed will be missed. Also, lack of health insurance and healthcare access likely reduces the recording of cases among medically underserved populations [12]. Furthermore, there are gaps in the passive surveillance system in which clinical cases are appropriately recorded at the state level. A 2017 study of pediatric patients with FBT in Houston found that only 48% of the 31 confirmed or probable cases were reported to the TDSHS [194].
14. Research Questions About FBT in Texas
This schema proposed in Figure 8 facilitates the generation of research questions to further understand the current epidemiology of FBT in TX. Understanding the dynamic relationships of each component to each other will allow the generation of predictive models for the future epidemiology of FBT. Specific research questions are listed below:
- #1 What is the magnitude of the stray and feral cat and dogs problem in the high FBT incidence areas of TX and their degree of flea infestation over the course of a year?
- #2 How do the numbers of opossums, dogs, and cats and strays relate to increases in human cases in individual counties?
- #3 What is the net effect of climate change on the flea life cycle (is the decrease in life cycle duration countervailed by the decrease in adult flea life span)?
- #4 Are changes in temperature, rainfall, and relative humidity associated with increased human cases in specific locales?
- #5 What endosymbiotic organisms (Wolbachia pipientis and Steinina ctenocephali) occur within the flea vectors in Texas and how they affect flea survival and infection with R. felis and R. typhi?
- #6 What is the impact of co-infection of R. felis/R. typhi and Bartonella sp. on flea survival and vectorial capacity?
- #7 Are R. felis and R. typhi still co-circulating in TX? Do they vary geographically and by season?
15. Conclusions
Flea-borne typhus is an infection, caused by generalist bacteria, spread by generalist fleas, with flea hosts that are also generalists. This increases the risk of spread of FBT to pets and humans. It is difficult to control from a public health perspective, because its transmission is closely associated with human behavior, domestic pets, stray/feral dogs and cats, and urban wildlife [54]. Human population growth in TX promotes FBT by increased urbanization and abundance of pet dogs and cats, stray/feral dogs and cats, and opossums. Increasing temperatures in Texas in the new millennium increases the flea-borne transmission of typhus by promoting host infestation and flea feeding and defecation, accelerating the flea life cycle, and increasing rickettsial replication within the flea. Increased opossums and stray cats and dogs in the urban/suburban landscape increase the risk of flea transfer to humans and their pets.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the article.
Acknowledgments
I thank Adam Lynch and Bonny Mayes from the Texas Department of State Health Services for providing data on FBT in Texas. I also thank Sue Simon and Susan Bolden of the University of Texas Health San Antonio for the figures.
Dedication
This paper is dedicated to my scientific mentor, Professor John A. Katzenellenbogen of the University of Illinois at Urbana-Champaign, on the occasion of his 80th birthday.
Conflicts of Interest
The author declares no conflict of interest.
References
- Tsioutis, C.; Zafeiri, M.; Avramopoulos, A.; Prousali, E.; Miligkos, M.; Karageorgos, S.A. Clinical and laboratory characteristics, epidemiology, and outcomes of murine typhus: a systematic review. Acta Trop. 2017, 166, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Chueng, T.; Koch, K.; Anstead, G.M.; Dayton, C. Case report: early doxycycline therapy for potential rickettsiosis in critically ill patients in flea-borne typhus endemic areas. Am. J. Trop. Med. Hyg. 2019, 101, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.F. Epidemiology of murine typhus. Annu. Rev. Entomol. 1990, 35, 553–569. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.D.; Macaluso, K.R. Rickettsia felis, an emerging flea-borne rickettsiosis, Curr. Trop. Med. Rep. 2016, 3, 27–39. [Google Scholar] [CrossRef]
- Texas Department of Health Services. Flea-borne Typhus Cases in Texas by County Reported, 2008-Available online:. Available online: https://www.dshs.texas.gov/IDCU/disease/typhus/Typhus-2008-2019.pdf (accessed on 30 April 2022).
- Anstead, G.M. History, rats, fleas, and opossums. II. The decline and resurgence of flea-borne typhus in the United States, 1945. Trop. Med. Infect. Dis. 2020, 6, 2. [Google Scholar] [CrossRef]
- California Dept of Public Health, Dept of Public Health Vector-Borne Disease Section. “Human Flea-Borne Typhus Cases in California (2001-2019),” Sacramento, California, USA, Available online:. Available online: https://www.CDPH.ca.gov/Programs/CID/DCDC/CDPH%20Document%20Library/Flea-borneTyphusCaseCounts.pdf (accessed on 18 March 2019).
- Texas Department of Health. Murine Typhus. Infectious Diseases Epidemiology and Surveillance Division, Austin, TX, USA, May 2001.
- Sleeman, J.M.; DeLiberto, T.; Nguyen, N. Optimization of human, animal, and environmental health by using the One Health approach. J. Vet. Sci. 2017, 18, 263–268. [Google Scholar] [CrossRef]
- Maina, A.N.; Fogarty, C.; Krueger, L.; Macaluso, K.R.; Odhiambo, A.; Nguyen, K.; Farris, C.M.; Luce-Fedrow, A.; Bennett, S.; Jiang, J.; et al. Rickettsial infections among Ctenocephalides felis and host animals during a flea-borne rickettsioses outbreak in Orange County, California. PLoS One 2016, 11, e0160604. [Google Scholar] [CrossRef]
- Prism Climate Group. Northwest Alliance for Computational Science and Engineering. Available online: https://prism.oregonstate.edu/ (accessed on 1 September 2022).
- Yomogida, K.; Kjemtrup, A.; Martínez-López, B.; Ibrahim, M.; Contreras, Z.; Ngo, V.; Halai, U.A.; Balter, S.; Feaster, M.; Zahn, M.; et al. Surveillance of flea-borne typhus in California, 2011-Am. J. Trop. Med. Hyg. 2023, 110, 142–149. [Google Scholar] [CrossRef]
- Centeno, F.H.; Lasco, T.; Ahmed, A.A.; Al Mohajer, M. Characteristics of Rickettsia typhi infections detected with next-generation sequencing of microbial cell-free deoxyribonucleic acid in a tertiary care hospital. Open Forum Infect. Dis. 2021, 8, ofab147. [Google Scholar] [CrossRef]
- Alarcón, J.; Sanosyan, A.; Contreras, Z.A.; Ngo, V.P.; Carpenter, A.; Hacker, J.K.; Probert, W.S.; Terashita, D.; Balter, S.; Halai, U.A. Fleaborne Typhus-Associated Deaths—Los Angeles County, California, MMWR Morb. Mortal. Wkly Rep. 2023, 72, 838–843. [Google Scholar] [CrossRef]
- Texas Department of State Health Services. Flea-borne Typhus. Available online: https://www.dshs.texas.gov/notifiable-conditions/zoonosis-control/zoonosis-control-diseases-and-conditions/flea-borne-typhus (accessed on 5 November 2024).
- Tabachnick, W.J. Challenges in predicting climate and environmental effects on vector-borne disease episystems in a changing world. J. Exp. Biol. 2010, 213, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Bohls, S.W. Typhus fever in Texas. South. Med. J. 1935, 28, 1162–1165. [Google Scholar] [CrossRef]
- Langley, B.C. Peanut culture. Texas State Historical Association. Available online: https://www.tshaonline.org/home/ (accessed on 15 January 2022).
- Wooster, R. East Texas in World War II. East Texas Hist. J. 2007, 45, Iss–2. Available online: https://scholarworks.sfasu.edu/ethj/vol45/iss2/9 (accessed on 20 February 2022).
- Reece, C.D. Typhus fever in Texas. Texas State J. Med., 1934, 30, 192–195. [Google Scholar]
- Davis, D.E.; Pollard, M. The distribution of murine typhus in rats and humans in San Antonio. Am. J. Trop. Med. Hyg. 1946, 26, 619–624. [Google Scholar] [CrossRef]
- Anstead, G.M. History, rats, fleas, and opossums: the ascendency of flea-borne typhus in the United States, 1910. Trop. Med. Infect. Dis. 2020, 5, 37. [Google Scholar] [CrossRef]
- Waldrop, R.H.; Ogden, L.J. A survey to determine the prevalence and distribution of typhus in rats in Texas. CDC Bull. 1951, 10, 53. Available online: https://stacks.cdc.gov/view/cdc/55659 (accessed on 18 July 2024).
- Irons, J.V.; Cox, G.W. An epidemiological investigation of typhus fever in Texas, 1943. Texas State J. Med. 1946, 42, 332–336. [Google Scholar]
- Bradley, G.H.; Wiley, J.S. The control of murine typhus in the United States. In Rickettsial Diseases of Man; Moulton, F.R., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1948; pp. 229–240. [Google Scholar]
- Boostrom, A.; Beier, M.S.; Macaluso, J.A.; Macaluso, K.R.; Sprenger, D.; Hayes, J.; Radulovic, S.; Azad, A.F. Geographic association of Rickettsia felis-infected opossums with human murine typhus, Texas. Emerg. Infect. Dis. 2002, 8, 549–554. [Google Scholar] [CrossRef]
- Older, J.J. The epidemiology of murine typhus in Texas. JAMA 1970, 14, 2011–2017. [Google Scholar] [CrossRef]
- Taylor, J.P.; Betz, T.G.; Rawlings, J.A. Epidemiology of murine typhus in Texas, 1980. JAMA 1986, 255, 2173–2176. [Google Scholar] [CrossRef] [PubMed]
- Texas Water Development Board. Water for Texas Jan Austin, TX. Available online: https://www.twdb.texas.gov/publications/state_water_plan/2012/04.pdf (accessed on 27 October 2024).
- Murray, K.O.; Evert, N.; Mayes, B.; Fonken, E.; Erickson, T.; Garcia, M.N.; Sidwa, T. Typhus group rickettsiosis, Texas, USA, 2003-Emerg. Infect. Dis. 2017, 23, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Rural Health Information Hub. Texas. Available online: https://www.ruralhealthinfo.org/states/texas (accessed on 1 September 2022).
- Research Division of the Texas Legislative Council. Definitions of “Rural” in Texas Statutes and the Texas Administrative Code as of April Texas Legislative Council P.O. Box 12128 Austin, Texas 78711. Available online: https://tlc.texas.gov/docs/policy/Def_Rural_Statutes.pdf (accessed on 3 October 2021).
- Eremeeva, M.E.; Warachina, W.R.; Sturgeon, M.M.; Buchholz, A.E.; Olmsted, G.K.; Park, S.Y.; Effler, P.V.; Karpathy, S.E. Rickettsia typhi and R. felis in rat fleas (Xenopsylla cheopis), Oahu, Hawaii. Emerg. Infect. Dis. 2008, 14, 1613–1615. [Google Scholar] [CrossRef] [PubMed]
- Ferdman, R.A.; Ingraham, C. Where cats are more popular than dogs in the U.S.—and all over the world. Washington Post, July 28, 2014.
- Bergman, D.L.; Breck, S.W.; Bender, S.C. Dogs gone wild: feral dog damage in the United States. Proc. 13th Wildlife Damage Management Conf. (J.R. Boulanger, Ed.) 2009, 13, 1–9. [Google Scholar]
- Chasnoff, B. Collaring the problem of stray dogs. San Antonio Express-News Sep 19. Available online: https://www.mysanantonio.com/news/local_news/article/Collaring-the-problem-of-stray-dogs-845675.php (accessed on 18 July 2024).
- Slater, M.R. The welfare of feral cats. In: Rochlitz, I. (ed), The Welfare of Cats. Dordrecht: Springer, pp. 141–176, 2005.
- Levy, J.K.; Crawford, P.C. Humane strategies for controlling feral cat populations, J. Am. Vet. Med. Assoc. 2004, 225, 1354–1360. [Google Scholar] [CrossRef]
- Mohr, C.O.; Good, N.E.; Schubert, J.H. Status of murine typhus infection in domestic rats in the United States, 1952, and relation to infestation by Oriental rat fleas. Am. J. Public Health, 1953, 43, 1514–1522. [Google Scholar] [CrossRef]
- Mohr, C.O. Entomological background of the distribution of murine typhus and murine plague in the United States. Am. J. Trop. Med. Hyg. 1951, 31, 355–372. [Google Scholar] [CrossRef]
- Eskey, C.R.; Hemphill, F.M. Relation of reported cases of typhus fever to location, temperature, and precipitation. Public Health Rep. 1948, 63, 941–948. [Google Scholar] [CrossRef]
- Davis, W.B.; Schmidly, D.J. The mammals of Texas, University of Texas Press, Austin, TX, USA, 1994.
- Buttery, C.M.; Magnuson, L.W.; McLerran, G.; Villarreal, T. Endemic (murine) typhus in Corpus Christi. Tex. Med. 1984, 80, 53–54. [Google Scholar]
- Statista. Available online: https://www.statista.com/statistics/432924/us-metropolitan-areas-with-the-highest-poverty-rate/ (accessed on 18 July 2024).
- Yao, Z.; Tang, J.; Zhan, F.B. Detection of arbitrarily-shaped clusters using a neighbor-expanding approach: a case study on murine typhus in south Texas. Int. J. Health Geogr. 2011, 10, 23. [Google Scholar] [CrossRef]
- Hassell, J.M.; Begon, M.; Ward, M.K.; Fèvre, E.M. Urbanization and disease emergence: dynamics at the wildlife–livestock–human interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J. Population biology of emerging and re-emerging pathogens. Trends Microbiol. 2002, 10, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA, 2003, 100, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett 2006, 9, 485–498. [Google Scholar] [CrossRef]
- Clark, N.J.; Seddon, J.M.; Šlapeta, J.; Wells, K. Parasite spread at the domestic animal—wildlife interface: anthropogenic habitat use, phylogeny and body mass drive risk of cat and dog flea (Ctenocephalides spp.) infestation in wild mammals. Parasit. Vectors, 2018, 11, 8. [Google Scholar] [CrossRef]
- Salje, J. Cells within cells: Rickettsiales and the obligate intracellular bacterial lifestyle. Nature Rev. Microbiol. 2021, 19, 375–390. [Google Scholar] [CrossRef]
- Adams, J.R.; Schmidtmann, E.T.; Azad, A.F. Infection of colonized cat fleas, Ctenocephalides felis (Bouché), with a Rickettsia-like microorganism. Am. J. Trop. Med. Hyg. 1990, 43, 400–409. [Google Scholar] [CrossRef]
- Wiggers, R.J.; Martin, M.C.; Bouyer, D. Rickettsia felis infection rates in an East Texas population. Tex. Med. 2005, 101, 56–58. [Google Scholar]
- Cummings, R.; Krueger, L.; Nguyen, K.; Rangel, D.; Penicks, A.; Sims, J. Challenges of responding to flea-borne typhus cases: lessons learned after 15 years of investigations in Orange County. Proc. Mosquito Vector Control Assoc. Calif. 2022, 90, 33–42. [Google Scholar]
- Hurd, H. Manipulation of medically important insect vectors by their parasites. Annu. Rev. Entomol. 2003, 48, 141–161. [Google Scholar] [CrossRef]
- Abramowicz, K.F.; Rood, M.P.; Krueger, L.; Eremeeva, M.E. Urban focus of Rickettsia typhi and Rickettsia felis in Los Angeles, California. Vector-Borne Zoonot. Dis. 2011, 11, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Wedincamp, J., Jr.; Foil, L.D. Vertical transmission of Rickettsia felis in the cat flea (Ctenocephalides felis Bouche). J. Vector Ecol. 2002, 27, 96–101. [Google Scholar] [PubMed]
- Hirunkanokpun, S.; Thepparit, C.; Foil, L.D.; Macaluso, K.R. Horizontal transmission of Rickettsia felis between cat fleas, Ctenocephalides felis. Molec. Ecol. 2011, 20, 4577–4586. [Google Scholar] [CrossRef] [PubMed]
- Morick, D.; Krasnov, B.R.; Khoklova, I.S.; Gutierrez, R.; Fielden, L.J.; Gottlieb, Y.; Harrus, S. Effects of Bartonella spp. on flea feeding and reproductive performance. Appl. Environ. Microbiol. 2013, 79, 3438–3443. [Google Scholar] [CrossRef]
- Azad, A.F.; Sacci, J.B., Jr.; Nelson, W.M.; Dasch, G.A.; Schmidtmann, E.T.; Carl, M. Genetic characterization and transovarial transmission of a typhus-like rickettsia found in cat fleas. Proc. Natl. Acad. Sci. U.S.A. 1992, 89, 43–46. [Google Scholar] [CrossRef]
- Higgins, J.A.; Sacci, J.B.; Schriefer, M.E.; Endris, R.G.; Azad, A.F. Molecular identification of Rickettsia-like microorganisms associated with colonized cat fleas (Ctenocephalides felis). Insect Mol. Biol. 1994, 3, 27–33. [Google Scholar] [CrossRef]
- Yang, W.-H.; Hsu, M.-S.; Shu, P.-Y.; Tsai, K.-H.; Fang, C.-T. Neglected human Rickettsia felis infection in Taiwan: a retrospective seroepidemiological survey of patients with suspected rickettsiosis. PLoS Negl. Trop. Dis. 2021, 15, e0009355. [Google Scholar] [CrossRef]
- Reif, K.E.; Kearney, M.T.; Foil, L.D.; Macaluso, K.R. Acquisition of Rickettsia felis by cat fleas during feeding. Vector Borne Zoonotic Dis. 2011, 11, 963–968. [Google Scholar] [CrossRef]
- Legendre, K.P.; Macaluso, K.R. Rickettsia felis: a review of transmission mechanisms of an emerging pathogen. Trop. Med. Infect. Dis. 2017, 2, 64. [Google Scholar] [CrossRef]
- Healy, S.P.; Brown, L.D.; Hagstrom, M.R.; Foil, L.D.; Macaluso, K.R. Transmission effect of Rickettsia felis strain variation on infection, transmission, and fitness in the cat flea (Siphonaptera: Pulicidae). J. Med. Entomol. 2017, 54, 1037–1043. [Google Scholar] [CrossRef]
- Traub, R.; Wisseman, C.L.; Fahrang-Azad, A. The ecology of murine typhus, a critical review. Trop. Dis. Bull. 1978, 75, 237–317. [Google Scholar] [PubMed]
- Vaughan, J.A.; Azad, A.F. Acquisition of murine typhus rickettsiae by fleas. Ann. N. Y. Acad. Sci. 1990, 590, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.F.; Radulovic, S.; Higgins, J.A.; Noden, B.H.; Troyer, J.M. Flea-borne rickettsiosis: ecologic considerations. Emerg. Infect. Dis. 1997, 3, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Gage, K.L. Transmission of flea-borne zoonotic agents. Annu. Rev. Entomol. 2012, 57, 61–82. [Google Scholar] [CrossRef]
- Wedincamp, J., Jr.; Foil, L.D. Infection and seroconversion of cats exposed to cat fleas (Ctenocephalides felis Bouche) infected with Rickettsia felis. J. Vector Ecol. 2000, 25, 123–126. [Google Scholar]
- Reif, K.E.; Macaluso, K.B. Ecology of Rickettsia felis: a review. J. Med. Entomol. 2009, 46, 723–735. [Google Scholar] [CrossRef]
- Macaluso, K.R.; Pornwiroon, W.; Popov, V.L.; Foil, L.D. Identification of Rickettsia felis in the salivary glands of cat fleas. Vector-Borne Zoonotic Dis. 2008, 8, 391–396. [Google Scholar] [CrossRef]
- Thepparit, C.; Hirunkanokpun, S.; Popov, V.L.; Foil, L.D.; Macaluso, K.R. Dissemination of bloodmeal acquired Rickettsia felis in cat fleas. Parasit. Vectors 2013, 24, 149. [Google Scholar] [CrossRef]
- Brown, L.D.; Banajee, K.H.; Foil, L.D.; Macaluso, K.R. Transmission mechanisms of an emerging insect-borne rickettsial pathogen. Parasit. Vectors 2016, 9, 237. [Google Scholar] [CrossRef]
- Azad, A.F.; Traub, R. Experimental transmission of murine typhus by Xenopsylla cheopis flea bites. Med. Vet. Entomol. 1989, 3, 429–433. [Google Scholar] [CrossRef]
- Nogueras, M.M.; Pons, I.; Ortuño, A.; Miret, J.; Pla, J.; Castellá, J.; Segura, F. Molecular detection of Rickettsia typhi in cats and fleas. PLoS ONE 2013, 8, e71386. [Google Scholar] [CrossRef] [PubMed]
- Eremeeva, M.E.; Karpathy, S.E.; Krueger, L.; Hayes, E.K.; Williams, A.M.; Zaldivar, Y.; Bennett, S.; Cummings, R.; Tilzer, A.; Velten, R.K.; et al. Two pathogens and one disease: detection and identification of flea-borne Rickettsiae in areas endemic for murine typhus in California. J. Med. Entomol. 2012, 49, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Noden, B.H.; Radulovic, S.; Higgins, J.A.; Azad, A.F. Molecular identification of Rickettsia typhi and R. felis in co-infected Ctenocephalides felis (Siphonaptera: Pulicidae). J. Med. Entomol. 1998, 35, 410–414. [Google Scholar] [CrossRef]
- Macaluso, K.R.; Sonenshine, D.E.; Cerual, S.M.; Azad, A.F. Rickettsial infection in Dermacentor variabilis (Acari: Ixodidae) inhibits transovarial transmission of a second Rickettsia. J. Med. Entomol. 2002, 39, 809–813. [Google Scholar] [CrossRef]
- Bitam, I.; Dittmar, K.; Parola, P.; Whiting, M.F. Fleas and flea-borne diseases, Internat. J. Infect. Dis. 2010, 14, e667–e676. [Google Scholar]
- De Sousa, R.; Edouard-Fournier, P.; Santos-Silva, M.; Amaro, F.; Bacellar, F.; Raoult, D. Molecular detection of Rickettsia felis, Rickettsia typhi and two genotypes closely related to Bartonella elizabethae. Am. J. Trop. Med. Hyg. 2006, 75, 727–731. [Google Scholar] [CrossRef]
- Pérez-Osorio, C.E.; Zavala-Velázquez, J.E.; Arias León, J.; Zavala-Castro, J.E. Rickettsia felis as emergent global threat for humans. Emerg. Infect. Dis. 2008, 14, 1019–1023. [Google Scholar] [CrossRef]
- Richards, A.L.; Jiang, J.; Omulo, S.; Dare, R.; Abdirahman, K.; Ali, A.; Sharif, S.K.; Feikin, D.R.; Breiman, R.F.; Njenda, M.K. Human infection with Rickettsia felis, Kenya. Emerg. Infect. Dis. 2010, 16, 1081–1086. [Google Scholar] [CrossRef]
- Fogarty, C.L.; Krueger, L.; Nguyen, K.; Velten, R.; Bennett, S.; Sun, S.; Cummings, R. Development of an ELISA for determining the presence of rickettsial antibodies in the Virginia opossum, Didelphis virginiana. Proc. Mosquito Vect. Control Assoc. Calif. 2013, 81, 96–100. [Google Scholar]
- Cleaveland, S.; Laurenson, M.K.; Taylor, L.H. Diseases of humans and their domestic mammals: pathogen characteristics, host range and the risk of emergence. Philos. Trans. Roy. Soc. B Biol. Sci. 2001, 356, 991–999. [Google Scholar] [CrossRef]
- Brisson, D.; Brinkley, C.; Humphrey, P.T.; Kemps, B.D.; Ostfeld, R.S. It takes a community to raise the prevalence of a zoonotic pathogen. Interdiscip. Perspect. Infect. Dis. 2011, 2011, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Han, B.A.; Kramer, A.M.; Drake, J.M. Global patterns of zoonotic disease in mammals. Trends Parasitol. 2016, 32, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Taylor, L.H.; Haydon, D.T. Population biology of multi-host pathogens. Science 2001, 292, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Billeter, S.A.; Metzger, M.E. Limited evidence for Rickettsia felis as a cause of zoonotic flea-borne rickettsiosis in Southern California. J. Med. Entomol. 2017, 54, 4–7. [Google Scholar] [CrossRef]
- Angelakis, E.; Mediannikov, O.; Parola, P.; Raoult, D. Rickettsia felis: the complex journey of a emergent human pathogen. Trends Parasitol. 2016, 32, 554–564. [Google Scholar] [CrossRef]
- Blanton, L.S.; Walker, D.H. Flea-borne rickettsioses and Rickettsiae. Am. J. Trop. Med. Hyg. 2017, 96, 53–56. [Google Scholar] [CrossRef]
- Mediannikov, O.; Fenollar, F.; Bassene, H.; Tall, A.; Sokhna, C.; Trape, J.F.; Raoult, D. Description of “yaaf”, the vesicular fever caused by acute Rickettsia felis infection in Senegal. J. Infect. 2013, 66, 536–540. [Google Scholar] [CrossRef]
- Parola, P. Rickettsia felis: from a rare disease in the USA to a common cause of fever in sub-Saharan Africa. Clin. Microbiol. Infect. 2011, 17, 996–1000. [Google Scholar] [CrossRef]
- Schriefer, M.E.; Sacci, J.B., Jr.; Dumler, J.S.; Bullen, M.G.; Azad, A.F. Identification of a novel rickettsial infection in a patient diagnosed with murine typhus. J. Clin. Microbiol. 1994, 32, 949–54. [Google Scholar] [CrossRef]
- Peniche Lara, G.; Dzul-Rosado, K.R.; Zavala Velazquez, J.E.; Zavala-Castro, J. Murine typhus: clinical and epidemiological aspects. Colomb. Med. (Cali) 2012, 43, 175–180. [Google Scholar] [CrossRef]
- Santoyo-Colín, V.; Sánchez-Montes, S.; Salceda-Sánchez, B.; Huerta-Jiménez, H.; Alcántara-Rodríguez, V.; Becker, I.; Gual-Sill, F.; López-Pérez, A.M. Urban foci of murine typhus involving cat fleas (Ctenocephalides felis felis) collected from opossums in Mexico City. Zoonoses Public Health 2021, 68, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Merhej, V.; Angelakis, E.; Socolovschi, C.; Raoult, D. Genotyping, evolution and epidemiological findings of Rickettsia species. Infect. Genet. Evol. 2014, 25, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Renesto, P.; Audic, S.; Robert, C.; Blanc, G.; Fournier, P.E.; Parinello, H.; Claverie, J.M.; Raoult, D. The genome sequence of Rickettsia felis identifies the first putative conjugative plasmid in an obligate intracellular parasite. PLoS Biol. 2005, 3, e248. [Google Scholar] [CrossRef] [PubMed]
- Schriefer, M.E.; Sacci, J.B. Jr; Taylor, J.P.; Higgins, J.A.; Azad, A.F. Murine typhus: updated roles of multiple urban components and a second typhus like rickettsia. J. Med. Entomol. 1994, 31, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.F.; Sacci, J.B., Jr.; Nelson, W.M.; Dasch, G.A.; Schmidtmann, E.T.; Carl, M. Genetic characterization and transovarial transmission of a typhus-like rickettsia found in cat fleas. Proc. Natl. Acad. Sci. U.S.A. 1992, 89, 43–46. [Google Scholar] [CrossRef]
- Abramowicz, K.F.; Wekesa, J.W.; Nwadike, C.N.; Zambrano, M.L.; Karpathy, S.E.; Cecil, D.; Burns, J.; Hu, R.; Eremeeva, E. Rickettsia felis in cat fleas, Ctenocephalides felis parasitizing opossums, San Bernardino County, California. Med. Vet. Entomol. 2012, 26, 458–462. [Google Scholar] [CrossRef]
- Laukaitis, H.J.; Macaluso, K.R. Unpacking the intricacies of Rickettsia-vector interactions. Trends Parasitol. 2021, 37, 734–746. [Google Scholar] [CrossRef]
- Moore, C.; Breitschwerdt, E.B.; Kim, L.; Li, Y.; Ferris, K.; Maggi, R.; Lashnits, E. The association of host and vector characteristics with Ctenocephalides felis pathogen and endosymbiont infection. Front. Microbiol. 2023, 14, 1137059. [Google Scholar] [CrossRef]
- Irons, J.V.; Bohls, S.W.; Thurman, D.C.; McGregor, T. Probable role of the cat flea, Ctenocephalides felis, in transmission of murine typhus. Am. J. Trop. Med. 1944, 24, 359–362. [Google Scholar] [CrossRef]
- Adams, W.H.; Emmons, R.W.; Brooks, J.E. The changing ecology of murine (endemic) typhus in southern California. Am. J. Trop Med. Hyg. 1970, 19, 311–318. [Google Scholar] [CrossRef]
- Traub, R. The coevolution of fleas and mammals. Pp. 295-437 in Kim, K.C. (ed). Coevolution of parasitic arthropods and mammals, Wiley, NY, USA, 1985.
- Krasnov, B.R. Functional and evolutionary ecology of fleas: a model for ecological parasitology, Cambridge University Press, New York, NY, USA, 2008.
- Yiguan, W.; Jie, T.; Qiyong, L.; Cannan, S.; Wenlong, K.; Henglu, D.; Cheng, X.; Wenzhu, Z.; Fajun, C.; Fengxia, M. Influence of bloodmeal host on blood feeding, egg production, and offspring sex ratio of Ctenocephalides felis felis (Siphonaptera: Pulicidae). J. Med. Entomol. 2016, 53, 888–893. [Google Scholar] [CrossRef] [PubMed]
- Blakely, BN.; Agnew, J.; Gard, C.; Romero, A. Effects of blood meal source on blood consumption and reproductive success of cat fleas, Ctenocephalides felis. PLoS Negl. Trop. Dis. 2023, 17, E0011233. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K.; Dryden, M.W. The biology, ecology, and management of the cat flea. Annu. Rev. Entomol. 1997, 42, 451–473. [Google Scholar] [CrossRef] [PubMed]
- Penicks, A.; Krueger, L.; Campbell, J.; Fogarty, C.; Rangel, D.; Nguyen, K.; Cummings, R. Flea abundance, species composition, and prevalence of rickettsiosis, from urban wildlife in Orange County, California, 2015-Proc. Vertebr. Pest Conf. 2020, 29, e7. [Google Scholar]
- Penicks, A.; Krueger, L.; Morgan, T.; Nguyen, K.; Campbell, J.; Fogarty, C.; Bennett, S.; Cummings, R. Jumping into the future: an analysis of 50 years of flea data from mammalian wildlife collected during three flea-borne rickettsioses surveys in Orange County, 1967-Proc. Calif. Mosquito Vector Control Assoc. 2019, 87, 203–207. [Google Scholar]
- Gage, K.; Kosoy, M. Natural history of plague: perspectives from more than a century of research. Annu. Rev. Entomol. 2005, 50, 505–528. [Google Scholar] [CrossRef]
- Marshall, A.G. The sex ratio in ectoparasitic insects. Ecol. Entomol. 1981, 6, 155–174. [Google Scholar] [CrossRef]
- Dryden, M.W.; Gaafar, S.M. Blood consumption by the cat flea, Ctenocephalides felis (Siphonaptera: Pulicidae). J. Med. Entomol. 1991, 28, 394–400. [Google Scholar] [CrossRef]
- Rust, M.K. The biology and ecology of cat fleas and advancements in their pest management: a review. Insects 2017, 8, 118. [Google Scholar] [CrossRef]
- Gálvez, R.; Musella, V.; Descalzo, M.A.; Montoya, A.; Checa, T.R.; Marino, V.; Martín, O.; Cringoli, G.; Rinaldi, L.; Miró, G. Modelling the current distribution and predicted spread of the flea species Ctenocephalides felis infesting outdoor dogs in Spain. Parasit. Vectors, 2017, 10, 428. [Google Scholar] [CrossRef]
- Van der Mescht, L.; le Roux, P.C.; Matthee, C.A.; Raath, M.J.; Mattee, S. The influence of life history characteristics on flea (Siphonaptera) species distribution models. Parasit. Vectors 2016, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Gettinger, D.; Ernest, K.A. Small-mammal community structure and the specificity of ectoparasite associations in central Brazil. Rev. Bras. Biol. 1995, 55, 331–341. [Google Scholar]
- Molyneux, D.H. Climate change and tropical disease: common themes in changing vector-borne disease scenarios. Trans. R. Soc. Trop. Med. Hyg. 2003, 97, 129–132. [Google Scholar] [CrossRef]
- Azad, A.F.; Beard, C.B. Rickettsial pathogens and their arthropod vectors. Emerg. Infect. Dis. 1998, 4, 179–186. [Google Scholar] [CrossRef]
- Chin, H.C.; Ahmad, N.W.; Lim, L.H.; Jeffrey, J.; Hadi, A.A.; Othman, H.; Omar, B. Infestation with the cat flea, Ctenocephalides felis felis (Siphonaptera: Pulicidae) among students in Kuala Lumper, Malaysia. Southeast Asian, J. Trop. Med. Public Health 2010, 41, 1331–1334. [Google Scholar]
- Leelavathi, M.; Norhayati, M.; Lee, Y.Y. Cat flea infestation in a hospital: a case report. Korean J. Parasitol. 2012, 50, 79–82. [Google Scholar] [CrossRef]
- Adjemian, J.; Parks, S.; McElroy, K.; Campbell, J.; Eremeeva, M.E.; Nicholson, W.L.; McQuiston, J.; Taylor, J. Murine typhus in Austin, Texas, USA, Emerg. Infect. Dis. 2010, 16, 412–417. [Google Scholar] [CrossRef]
- Chala, B.; Hamde, F. Emerging and re-emerging vector-borne infectious diseases and the challenges for control: a review. Front. Public Health 2021, 9, 715759. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration. National Centers for Environmental Information. State climate summaries Texas. Available online: https://statesummaries.ncics.org/chapter/tx/ (accessed on 4 April 2022).
- Xie, H.; Chang, N.-B.; Daranpob, A.; Prado, D. Assessing the long-term urban heat island in San Antonio, Texas based on moderate resolution imaging spectroradiometer/Aqua data. J. Appl. Remote Sens. 2010, 4, 043508–10.1117. [Google Scholar]
- Gage, K.L.; Burkot, T.R.; Eisen, R.J.; Hayes, E.B. Climate and vectorborne diseases. Am. J. Prevent. Med. 2008, 35, 436–450. [Google Scholar] [CrossRef]
- Ostfeld, R.S. Climate change and the distribution and intensity of infectious diseases. Ecology 2009, 90, 903–905. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Lindsay, L.R. Effects of climate and climate change on vectors and vector-borne diseases: ticks are different. Trends Parasitol. 2016, 32, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Roy, H.E.; Beckmann, B.C.; Comont, R.F.; Hails, R.S.; Harrington, R.; Medlock, J.; Purse, B.; Shortall, C.R. Nuisance insects and climate change. Department Environ, Food Rural Affairs; London, UK. Available online: https://nora.nerc.ac.uk./id/eprint/8332/ (accessed on 6 June 2022).
- Bernabeu-Wittel, M.; Pachón, J.; Alarcón, A.; López-Cortés, L.F.; Viciana, P.; Jiménez-Mejías, M.E.; Villanueva, J.L.; Torronteras, R.; Caballero-Granado, F.J. Murine typhus as a common cause of fever of intermediate duration: a 17-year study in the south of Spain. Arch. Intern. Med. 1999, 159, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.G.; Sacci, J.B., Jr.; Schriefer, M.E.; Andersen, E.M.; Fujioka, K.K.; Sorvillo, F.J.; Barr, A.R.; Azad, A.F. Typhus and typhus like rickettsiae associated with opossums and their fleas in Los Angeles County, California. J. Clin. Microbiol. 1992, 30, 1758–1762. [Google Scholar] [CrossRef]
- Hsueh, Y.L.; Chen, H.F.; Chang, M.C.; Yen, T.Y.; Su, C.L.; Chiu, H.C.; Hu, H.C.; Chung, Y.T.; Shu, P.Y.; Yang, S.L. Epidemiology of murine typhus in Taiwan from 2013 to Am. J. Trop. Med. Hyg. 2024, 110, 768–778. [Google Scholar] [CrossRef]
- Farrell, S.; McGarry, J.; Noble, P.M.; Pinchbeck, G.J.; Cantwell, S.; Radford, A.D.; Singleton, D.A. Seasonality and other risk factors for fleas infestations in domestic dogs and cats. Med. Vet. Entomol. 2023, 37, 1–12. [Google Scholar] [CrossRef]
- Beck, W.; Boch, K.; Mackensen, H.; Wiegand, B.; Pfister, K. Qualitative and quantitative observations on the flea population dynamics of dogs and cats in several areas of Germany. Vet. Parasitol. 2006, 137, 130–136. [Google Scholar] [CrossRef]
- Mohr, C.O.; Morlan, H.B. The nature of parasitism of the opossum by fleas in southwestern Georgia. J. Parasitol. 1959, 45, 233–237. [Google Scholar] [CrossRef]
- Roberts, T.; Parker, D.M.; Bulterys, P.L.; Rattanavong, S.; Elliott, I.; Phommasone, K.; Mayxay, M.; Chansamouth, V.; Robinson, M.T.; Blacksell, S.D.; et al. A spatio-temporal analysis of scrub typhus and murine typhus in Laos; implications from changing landscapes and climate. PLoS Negl. Trop. Dis. 2021, 15, e0009685. [Google Scholar] [CrossRef]
- Chang, K.; Chen, Y.H.; Lee, N.Y.; Lee, H.C.; Lin, C.Y.; Tsai, J.J.; Lu, P.L.; Chen, T.C.; Hsieh, H.C.; Lin, W.R.; et al. Murine typhus in southern Taiwan during 1992-Am. J. Trop. Med. Hyg. 2012, 87, 141–147. [Google Scholar] [CrossRef]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-dependent effects on the replication and transmission of arthropod-borne viruses in their insect hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the environmental temperature on Aedes aegypti and Aedes albopictus mosquitoes: a review. Insects 2018, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Farhang-Azad, A.; Traub, R.; Sofi, M.; Wisseman, C.L., Jr. Experimental murine typhus infection in the cat flea, Ctenocephalides felis (Siphonaptera: Pulicidae). J. Med. Entomol. 1984, 21, 675–680. [Google Scholar] [CrossRef] [PubMed]
- U.S. Global Change Research Program. Available online: https://www.globalchange.gov/our-work/third-national-climate-assessment (accessed on 12 November 2024).
- Pham, H.V.; Dang, D.T.; Tran Minh, N.N.; Nguyen, N.D.; Nguyen, T.V. Correlates of environmental factors and human plague: an ecological study in Vietnam. Internat. J. Epidemiol. 2009, 38, 1634–1641. [Google Scholar] [CrossRef] [PubMed]
- Crkvencic, N.; Šlapeta, J. Climate change models predict southerly shift of the cat flea (Ctenocephalides felis) distribution in Australia. Parasit. Vectors 2019, 12, 137. [Google Scholar] [CrossRef]
- Oke, T.R. The energetic basis of the urban heat-island. Quart. J. Roy. Meteorol. Soc. 1982, 108, 1–24. [Google Scholar] [CrossRef]
- Imhoff, M.; Zhang, P.; Wolfe, R.E.; Bounoua, L. Remote sensing of the urban heat island effect across biomes in the continental USA. Remote Sens. Environ. 2010, 114, 504–513. [Google Scholar] [CrossRef]
- Kalnay, E.; Cai, M. Impact of urbanization and land-use change on climate. Nature 2003, 423, 528–531. [Google Scholar] [CrossRef]
- Winguth, A.M.E.; Kelp, B. The urban heat island of the North-Central Texas region and its relation to the 2011 severe Texas drought. J. Appl. Meteorol. Climatol. 2013, 52, 2418–2433. [Google Scholar] [CrossRef]
- Richardson, S.M. A geospatial analysis of the urban heat island effect in Austin, TX. Honors Thesis, Texas State University, San Marcos, TX. Available online: https://digital.library.txst.edu/items/d91efa55-c340-4007-865a-59085cb1c82d (accessed on 27 October 2024).
- Zhang, X. The footprint of urban climates on vegetation phenology. Geophys. Res. Lett. 2004, 31, L12209. [Google Scholar] [CrossRef]
- Gracia, M.J.; Calvete, C.; Estrada, R.; Castillo, J.A.; Peribanez, M.A.; Lucientes, L. Fleas parasitizing domestic dogs in Spain. Vet. Parasitol. 2008, 151, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Xhaxhiu, D.; Kusi, I.; Rapti, D.; Visser, M.; Knaus, M.; Lindner, T.; Rehbein, S.; Rehbein, S. Ectoparasites of dogs and cats in Albania. Parasitol. Res. 2009, 105, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Durden, L.A.; Judy, T.N.; Martin, J.E.; Spedding, L.S. Fleas parasitizing domestic dogs in Georgia, USA: species composition and seasonal abundance. Vet. Parasitol. 2005, 130, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, L.; Spera, G.; Musella, V.; Carbone, S.; Veneziano, V.; Iori, A.; Cringoli, G. A survey of fleas on dogs in southern Italy. Vet. Parasitol. 2007, 148, 375–378. [Google Scholar] [CrossRef]
- Hopkins, D. Some aspects of the ecology of the Virginia Opossum (Didelphis virginiana virginiana Kerr 1792) in an urban environment. Masters Thesis. Portland State University, Portland, OR, USA. [CrossRef]
- Hernandez-Valdivia, E.; Cruz-Vazquez, C.; Ortiz-Martínez, R.; Valdivia-Flores, A.; Quintero-Martínez, M.T. Presence of Ctenocephalides canis (Curtis) and Ctenocephalides felis (Bouché) infesting dogs in the city of Aguascalientes, México. J. Parasitol. 2011, 97, 1017–1019. [Google Scholar] [CrossRef]
- Amin, O.M. The fleas (Siphonoptera) of Egypt: distribution and seasonal dynamics of fleas infesting dogs in the Nile Valley. J. Med. Entomol. 1966, 3, 293–298. [Google Scholar] [CrossRef]
- Cruz-Vazquez, C.; Castro Gamez, E.; Parada Fernandez, M.; Ramos Parra, M. Seasonal occurrence of Ctenocephalides felis felis and Ctenocephalides canis (Siphonaptera: Pulicidae) infesting dogs and cats in an urban area in Cuernavaca, Mexico. J. Med. Entomol. 2001, 38, 111–113. [Google Scholar] [CrossRef]
- Tavassoli, M.; Ahmadi, A.; Imani, A.; Ahmadiara, E.; Javadi, S.; Hadian, M. Survey of flea infestation in dogs in different geographical regions of Iran. Korean J. Parasitol. 2010, 48, 145–149. [Google Scholar] [CrossRef]
- Osbrink, W.L.A.; Rust, M.K. Fecundity and longevity of the adult cat flea, Ctenocephalides felis felis (Siphonaptera: Pulicidae). J. Med. Entomol. 1984, 21, 727–731. [Google Scholar] [CrossRef]
- Akucewich, L.H.; Philman, K.; Clark, A.; Gillespie, J.; Kunkle, G.; Nicklin, C.F.; Greiner, E.C. Prevalence of ectoparasites in a population of feral cats from north central Florida during the summer. Vet. Parasitol. 2002, 109, 129–139. [Google Scholar] [CrossRef]
- Šlapeta, J.; King, J.; McDonell, D.; Malik, R.; Homer, D.; Hannan, P.; Emery, D. The cat flea (Ctenocephalides f. felis) is the dominant flea on domestic dogs and cats in Australian veterinary practices. Vet. Parasitol. 2011, 180, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Farkas, R.; Gyurkovszky, M.; Solymosi, N.; Beugnet, F. Prevalence of flea infestation in dogs and cats in Hungary combined with a survey of owner awareness. Med. Vet. Entomol. 2009, 23, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Franc, M.; Bouhsira, É.; Beugnet, F. Direct transmission of the cat flea (Ctenocephalides felis) between cats exhibiting social behaviour. Parasite 2013, 20, 49. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.-H.; Wu, W.-J. Off-host observations of mating and postmating behaviors in the cat flea (Siphonaptera: Pulicidae). J. Med. Entomol. 2001, 38, 352–360. [Google Scholar] [CrossRef]
- Dean, S.R.; Meola, R.W. Factors influencing sperm transfer and insemination in cat fleas (Siphonaptera: Pulicidae) fed on an artificial membrane system. J. Med. Entomol. 2002, 39, 475–479. [Google Scholar] [CrossRef]
- Moser, B.A.; Koehler, P.G.; Patterson, R.S. Effect of larval diet on cat flea (Siphonaptera: Pulicidae) developmental times and adult emergence. J. Econ. Entomol. 1991, 84, 1257–1261. [Google Scholar] [CrossRef]
- Khokhlova, I.S.; Serobyan, V.; Degen, AA.; Krasnov, B.R. Host gender and offspring quality in a flea parasitic on a rodent. J. Exp. Biol. 2010, 213, 3299–3304. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Ammerman, N.C.; Beier-Sexton, M.; Sobral, B.S.; Azad, A.F. Louse- and flea-borne rickettsioses: biological and genomic analyses. Vet. Res. 2009, 40, 12. [Google Scholar] [CrossRef]
- Kern, W.H., Jr.; Richman, D.L.; Koehler, P.G.; Brenner, R.J. Outdoor survival and development of immature cat fleas (Siphonaptera: Pulicidae) in Florida. J. Med. Entomol. 1999, 36, 207–211. [Google Scholar]
- Hsu, M.H.; Wu, W.J. Effects of multiple mating on female reproductive output in the cat flea (Siphonaptera: Pulicidae). J. Med. Entomol. 2000, 37, 828–834. [Google Scholar] [CrossRef]
- Zakson-Aiken, M.; Gregory, L.M.; Shoop, W.L. Reproductive strategies of the cat flea (Siphonaptera: Pulicidae): parthenogenesis and autogeny? J. Med. Entomol. 1996, 33, 395–397. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.E.; Rust, M.K. Egg production and emergence of adult cat fleas (Siphonaptera: Pulicidae) exposed to different photoperiods. J. Med. Entomol. 1996, 33, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, D.; Warburton, E.M.; Khokhlova, I.S.; Krasnov, B.R. Reproductive consequences of female size in haematophagous ectoparasites. J. Exp. Biol. 2016, 219, 2368–2376. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Mackinnon, M.J.; Read, A.F. Virulence of mixed-clone and single-clone infections of rodent malaria Plasmodium chabaudi. Evolution 1998, 52, 583–591. [Google Scholar] [CrossRef]
- Hinkle, N.C. “Fleas.” World Health Organization: Public Health Significance of Urban Pests. (Eds. X. Bonnefoy, H. Kampen, and K. Sweeney). WHO European Centre for Environment and Health, Bonn, Germany, 2008.
- Krasnov, B.R.; Khokhlova, I.S.; Fielden, L.J.; Burdelova, N.V. Development rates of two Xenopsylla flea species in relation to air temperature and humidity. Med. Vet. Entomol. 2001, 15, 249–258. [Google Scholar] [CrossRef]
- Beck, W.; Stickel, M. Interhost migration behavior of Ctenocephalides felis on cats and in their resting sites. Wien Klin. Wochenschr. 2008, 120, 40–44. [Google Scholar] [CrossRef]
- Silverman, J.; Rust, M.K.; Reierson, D.A. Influence of temperature and humidity on survival and development of the cat flea, Ctenocephalides felis (Siphonaptera: Pulicidae). J. Med. Entomol. 1981, 18, 78–83. [Google Scholar] [CrossRef]
- Dryden, M.W.; Rust, M.K. The cat flea: biology, ecology and control. Vet. Parasitol. 1994, 52, 1–19. [Google Scholar] [CrossRef]
- Silverman, J.; Rust, M.K. Some abiotic factors affecting the survival of the cat flea, Ctenocephalides felis (Siphonaptera: Pulicidae). Environ. Entomol. 1983, 12, 490–495. [Google Scholar] [CrossRef]
- Wasson, S.; Novak, L. The most and least humid cities in the U.S. (Aug 14, 2023). Available online: https://todayshomeowner.com/moving/guides/most-and-least-humid-cities/ (accessed on 1 September 2023).
- Metzger, M.E.; Rust, M.K. Effect of temperature on cat flea (Siphonaptera: Pulicidae) development and overwintering. J. Med. Entomol. 1997, 34, 173–178. [Google Scholar] [CrossRef]
- Krämer, F.; Mencke, N. Flea Biology and Control, Springer-Verlag, Heidelberg, Germany, 2001.
- Silverman, J.; Rust, M.K. Extended longevity of the pre-emerged adult cat flea (Siphonaptera: Pulicidae) and factors stimulating emergence from the pupal cocoon. Ann. Entomol. Soc. Am. 1985, 78, 763–768. [Google Scholar] [CrossRef]
- Alarcón, M.E.; Jara-F.A., *!!! REPLACE !!!*; Briones, R.C.; Dubey, A.K.; Slamovits, C.H. Gregarine infection accelerates larval development of the cat flea Ctenocephalides felis (Bouché). Parasitology.
- Alarcón, M.E.; Huang, C.G.; Dubey, A.K.; Benítez, H. A gregarine from the gut of cat flea, Ctenocephalides felis (Bouché) (Siphonaptera: Pulicidae) in Taiwan: dynamic of infection patterns. Vet. Parasitol. 2013, 192, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Hague, M.T.J.; Caldwell, C.N.; Cooper, B.S. Pervasive effects of Wolbachia on host temperature preference. mBio 2020, 11, 10.1128–mbio.0176820. [Google Scholar] [CrossRef] [PubMed]
- Pornwiroon, W.; Kearney, M.T.; Husseneder, C.; Foil, L.D.; Macaluso, K.R. Comparative microbiota of Rickettsia felis-uninfected and -infected colonized cat fleas, Ctenocephalides felis. ISME J. 2007, 1, 394–402. [Google Scholar] [CrossRef]
- Tay, S.T. Wolbachia endosymbionts, Rickettsia felis and Bartonella species, in Ctenocephalides felis fleas in a tropical region. J. Vector Ecol. 2013, 38, 200–202. [Google Scholar] [CrossRef]
- Hague, M.T.J.; Shropshire, J.D.; Caldwell, C.N.; Statz, J.P.; Stanek, K.A.; Conner, W.R.; Cooper, B.S. Temperature effects on cellular host-microbe interactions explain continent-wide endosymbiont prevalence. Curr. Biol. 2022, 32, 878–888. [Google Scholar] [CrossRef]
- Paris, D.H.; Dumler, J.S. State of the art of diagnosis of rickettsial diseases: the use of blood specimens for diagnosis of scrub typhus, spotted fever group rickettsiosis, and murine typhus. Curr. Opin. Infect. Dis. 2016, 29, 433–439. [Google Scholar] [CrossRef]
- Erickson, T.; Da Silva, J.; Nolan, M.S.; Marquez, L.; Munoz, F.M.; Murray, K.O. Newly recognized pediatric cases of Typhus Group Rickettsiosis, Houston, Texas, USA. Emerg. Infect. Dis. 2017, 23, 2068–2071. [Google Scholar] [CrossRef]
Figure 1.
Map of the 254 counties of Texas.
Figure 2.
Cumulative incidence of flea-borne in the counties of Texas, 1941-1945 [24]. The 16 highest incidence counties were Nueces, Yoakum, Martin, Howard, Runnels, Jones, Shackelford, Bee, Gonzales, Dewitt, Lavaca, Comal, Matagordo, Waller, Liberty, and Madison (see Figure 1).

Figure 3.
Geodistribution of Flea-Borne Typhus in Texas, 1980-1984 [28].
Figure 3.
Geodistribution of Flea-Borne Typhus in Texas, 1980-1984 [28].
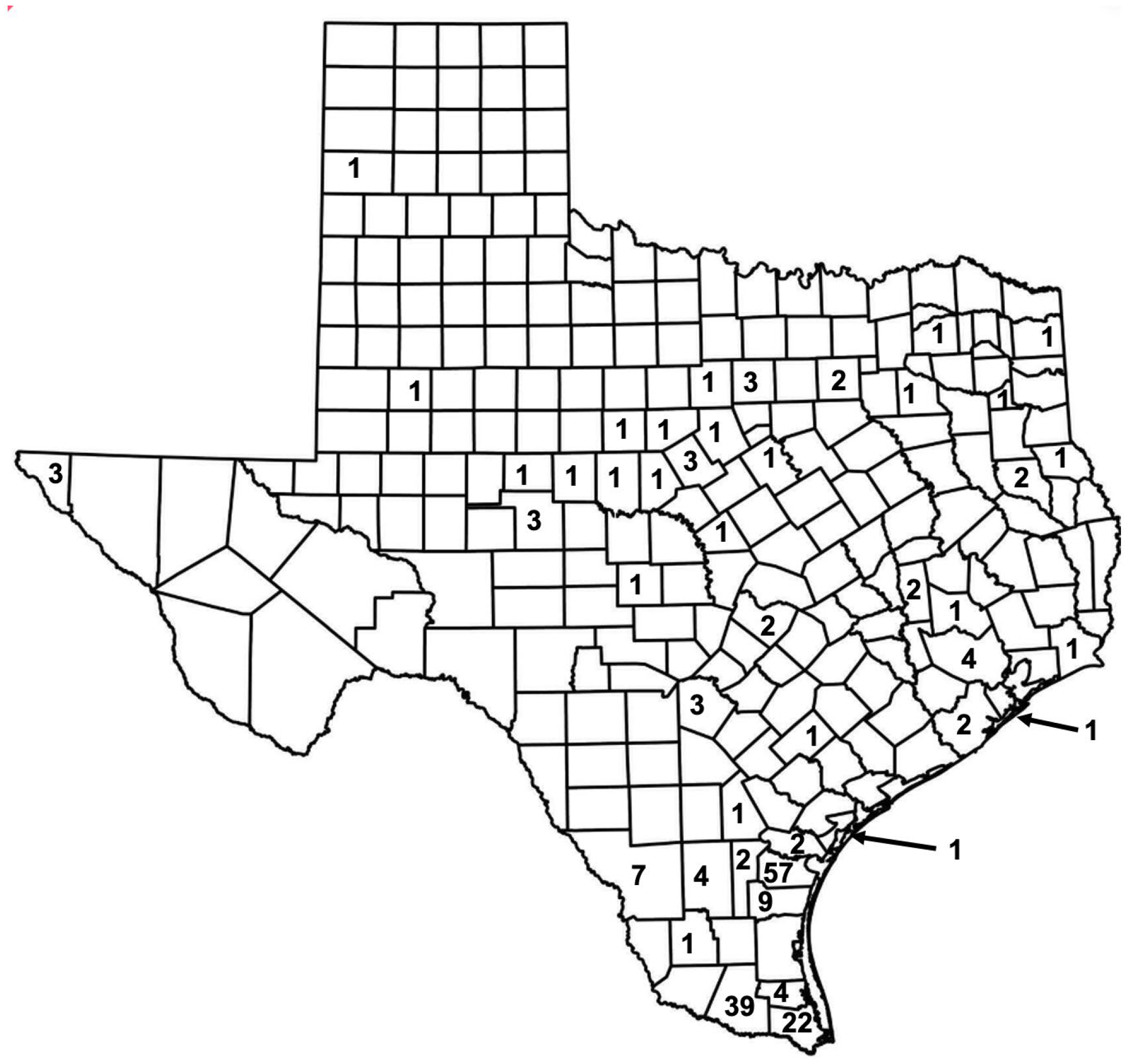
Figure 4.
Geodistribution of the 307 cases of flea-borne typhus by county in Texas 1990-1999 [8]. There were 28 counties with cases; 88.9% of cases occurred in the Refugium’s 13 counties, with only 34 cases outside the refugium (11.1%). Another seven cases occurred in San Patricio Co., a county bordering the Refugium, leaving only 27 cases (8.8%) outside the Refugium and a Refugium bordering county.
Figure 4.
Geodistribution of the 307 cases of flea-borne typhus by county in Texas 1990-1999 [8]. There were 28 counties with cases; 88.9% of cases occurred in the Refugium’s 13 counties, with only 34 cases outside the refugium (11.1%). Another seven cases occurred in San Patricio Co., a county bordering the Refugium, leaving only 27 cases (8.8%) outside the Refugium and a Refugium bordering county.
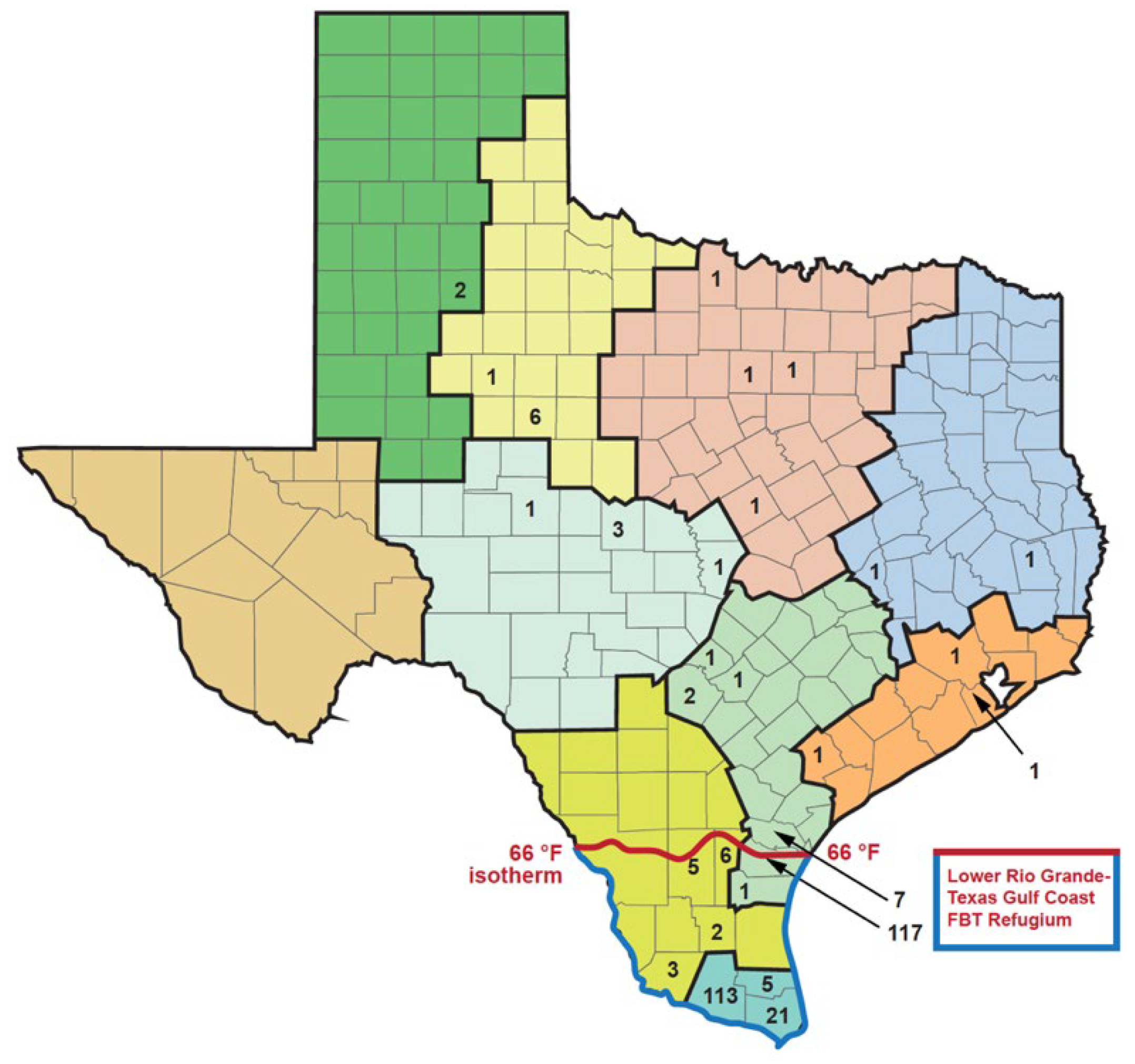
Figure 5.
Geodistribution of cases of flea-borne typhus in Texas in 2010-2019 based on the urban or rural classification of each county, highlighting the six urban counties with the highest number of FBT cases.
Figure 5.
Geodistribution of cases of flea-borne typhus in Texas in 2010-2019 based on the urban or rural classification of each county, highlighting the six urban counties with the highest number of FBT cases.
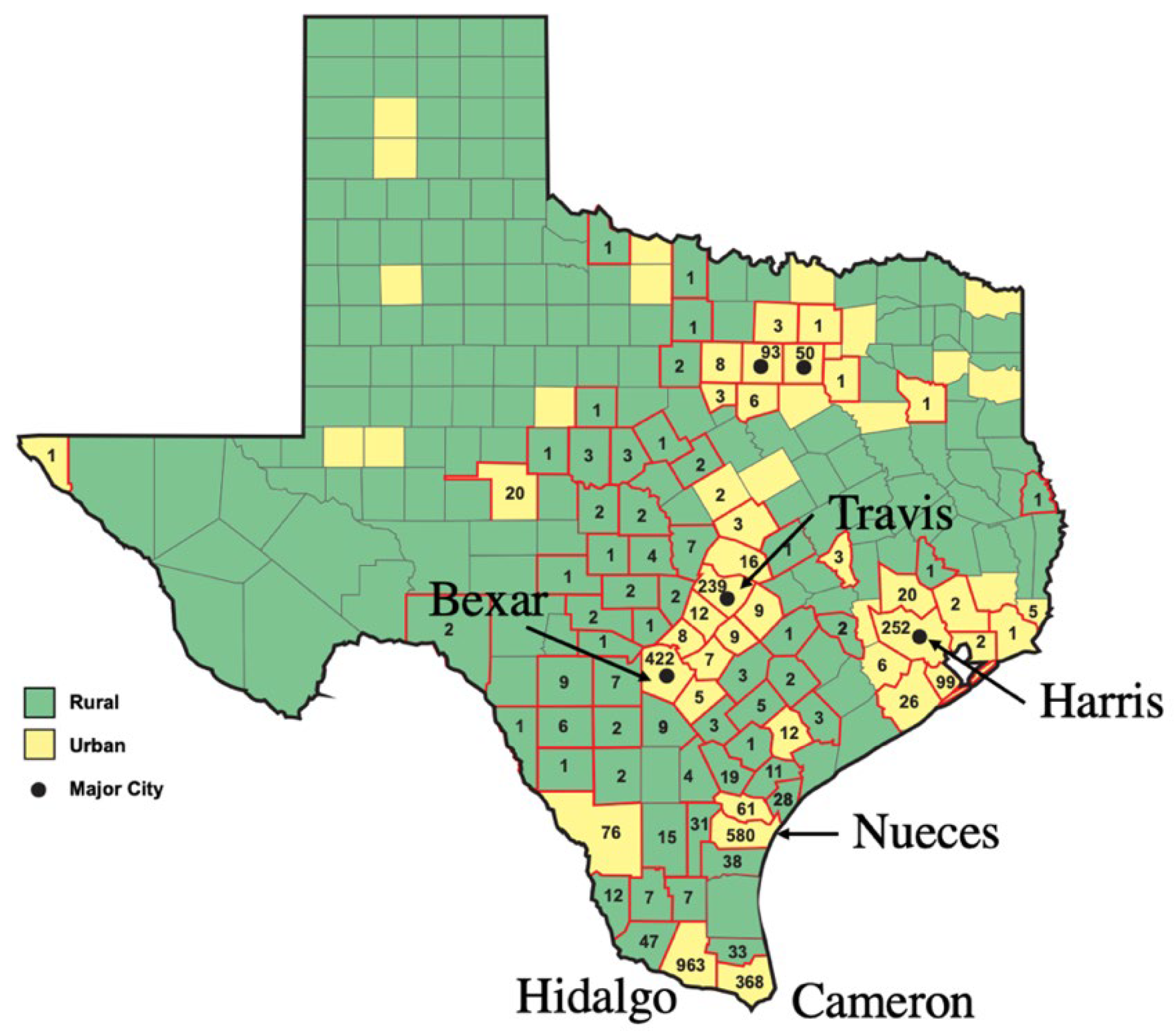
Figure 6.
The geodistribution of FBT cases in Texas with respect to National Climatic Data Center climate zones [29], 2010-2019.
Figure 6.
The geodistribution of FBT cases in Texas with respect to National Climatic Data Center climate zones [29], 2010-2019.
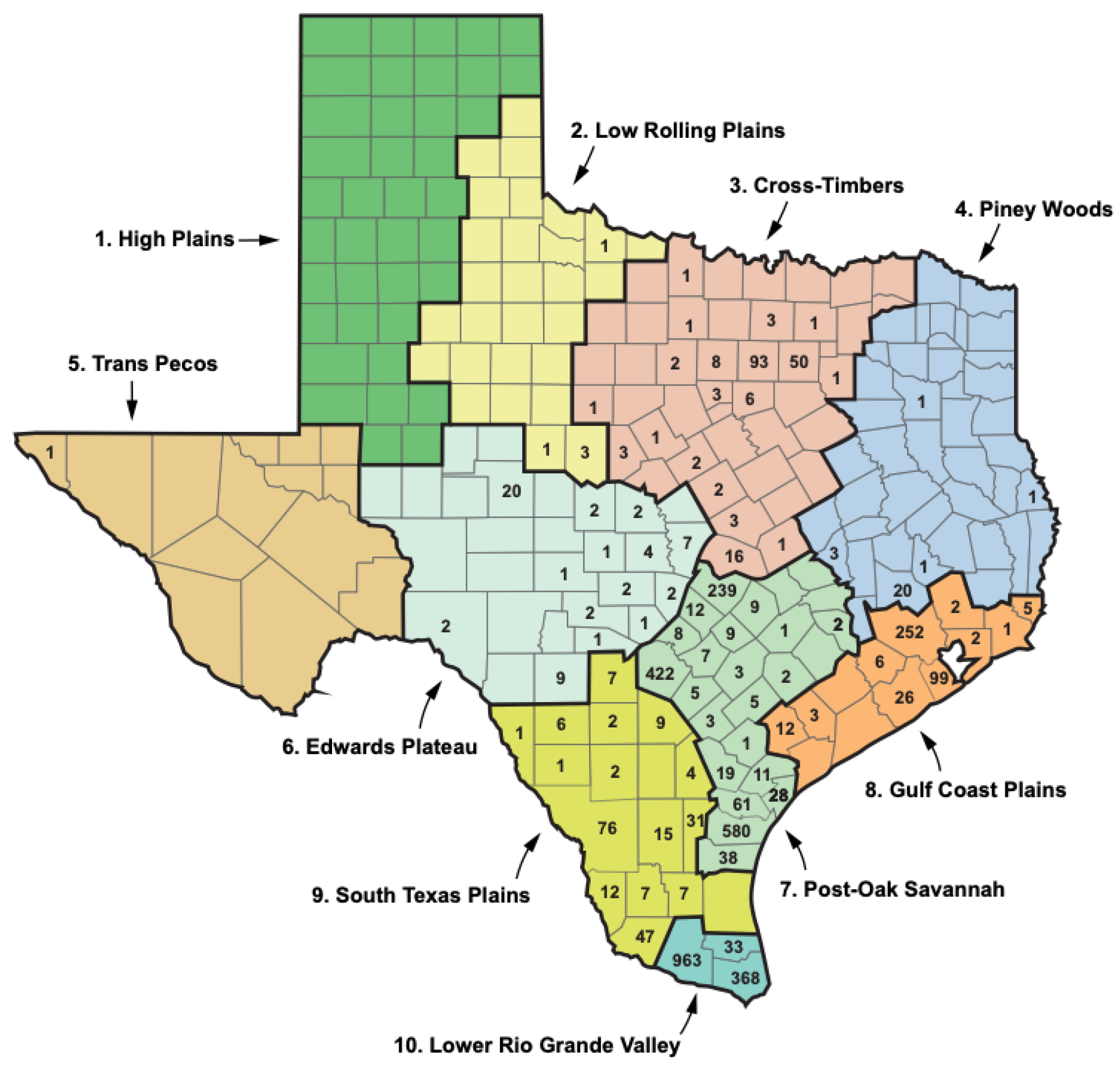
Figure 7.
Epidemiologic curve of flea-borne typhus in Texas, 1942–2019 [6,8,15]. There are superimposed time points for the introduction of technologies and programs for typhus control, the recognition of opossums as typhus reservoirs, the nadir of cases in the time period, and the growth of Texas population (Abbreviations: ANTU, alpha-napthylthiourea (rodenticide); CDC, Centers for Disease Control; DDT, dichlorodiphenyltricholoroethane (insecticide), 1080 (rodenticide)).
Figure 7.
Epidemiologic curve of flea-borne typhus in Texas, 1942–2019 [6,8,15]. There are superimposed time points for the introduction of technologies and programs for typhus control, the recognition of opossums as typhus reservoirs, the nadir of cases in the time period, and the growth of Texas population (Abbreviations: ANTU, alpha-napthylthiourea (rodenticide); CDC, Centers for Disease Control; DDT, dichlorodiphenyltricholoroethane (insecticide), 1080 (rodenticide)).
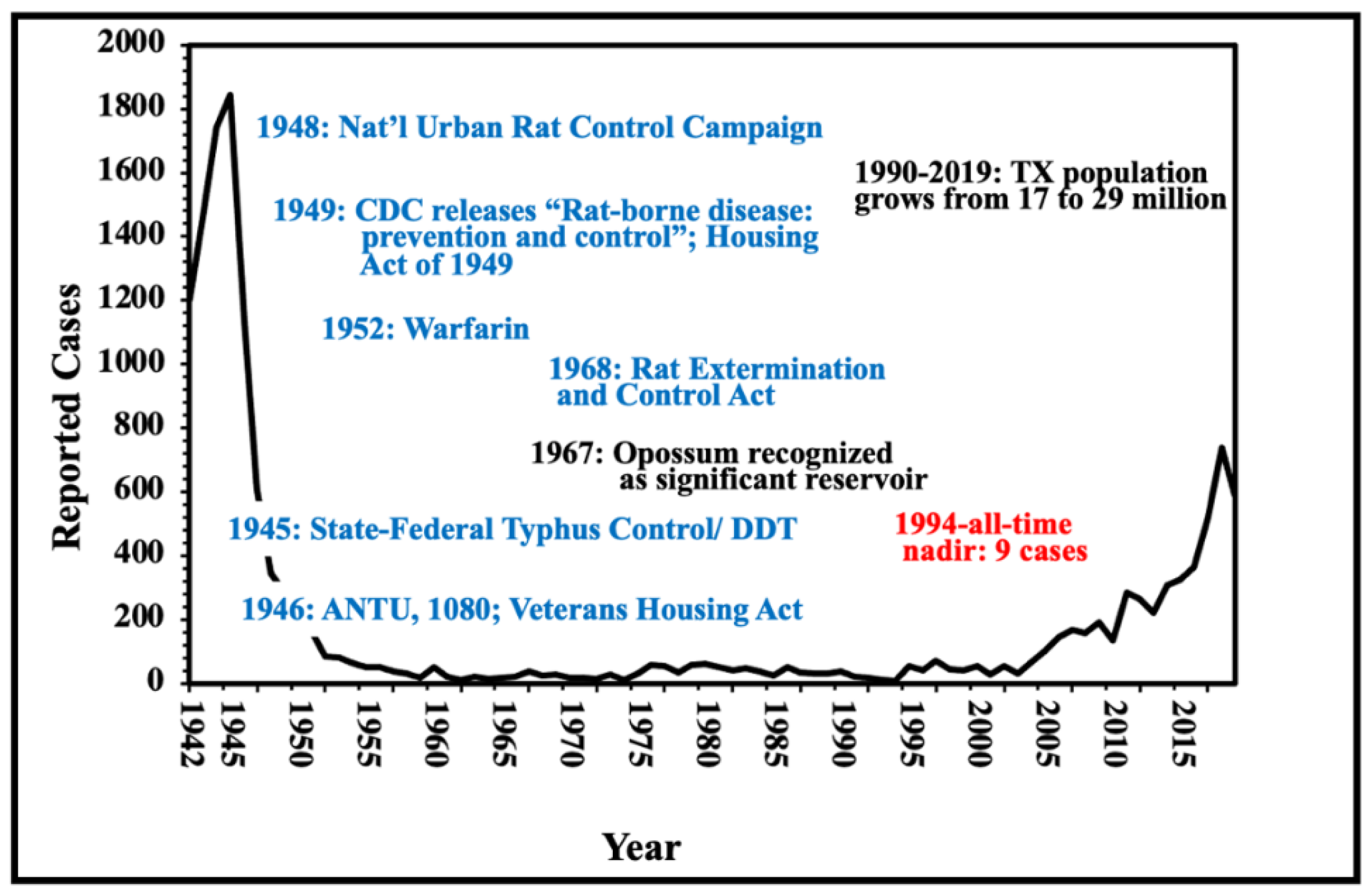
Figure 8.
One Health Model of Flea-borne Typhus Epidemiology in Texas in the Early 21st Century.
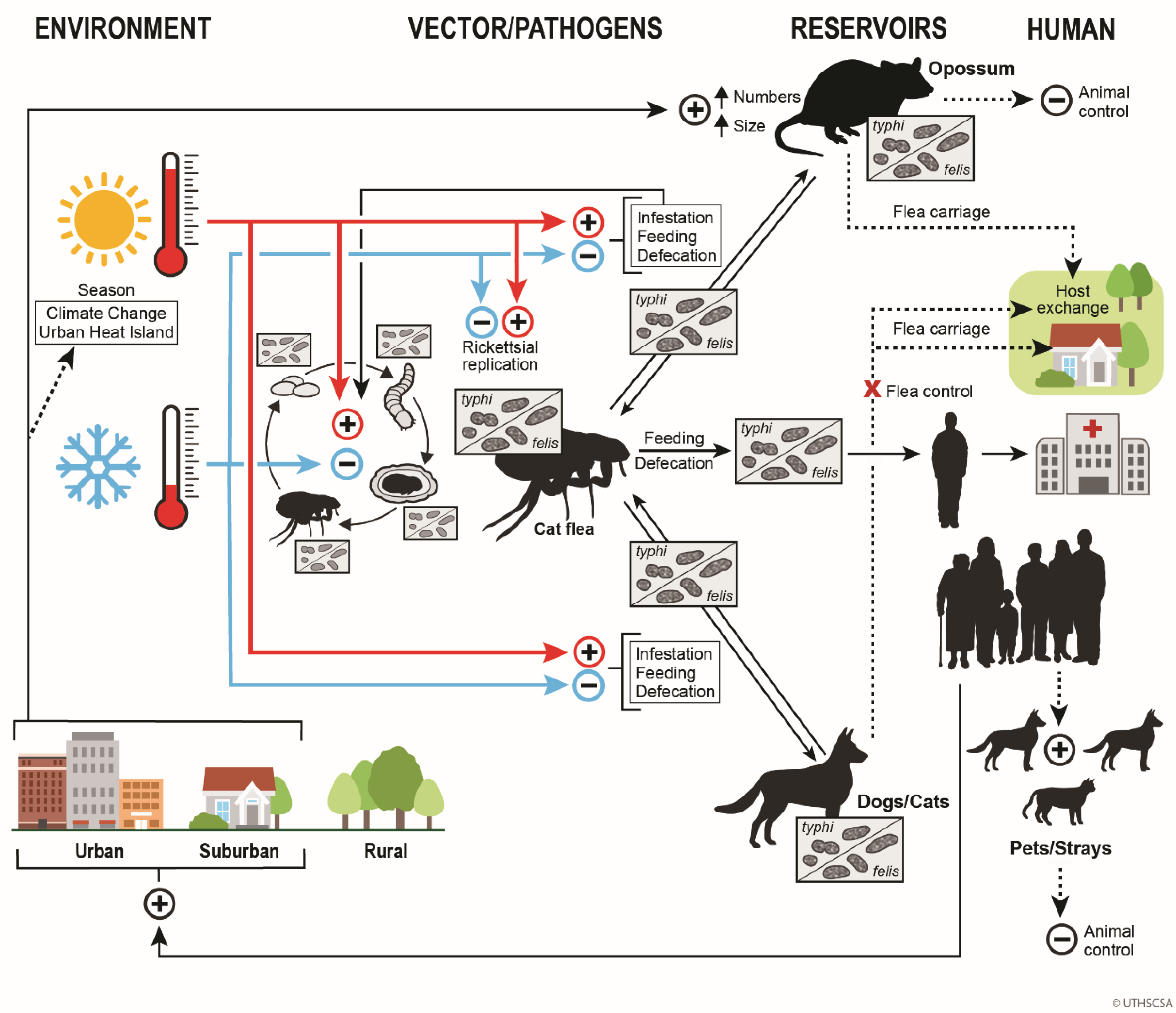

Table 1.
Abiotic, Biotic, and Anthropogenic Factors of the Episystem of Flea-Borne Typhus in Texas in the 20th and 21st Centuries.
Table 1.
Abiotic, Biotic, and Anthropogenic Factors of the Episystem of Flea-Borne Typhus in Texas in the 20th and 21st Centuries.
| Abiotic | Biotic | Anthropogenic |
|---|---|---|
| Climate | Abundance of urban exploiter hostsa | Population growth |
| Temperature | Abundance of companion animals | Urbanization/suburbanization |
| Rainfall | Abundance of other wildlife hosts | Agricultural practices |
| Relative humidity | Prevalence of flea endosymbiontsb | Human effects on climate |
| Cat flea clade-Rickettsia compatability | Urban heat island effect | |
| Pet ownership | ||
| Control of urban exploiter hosts | ||
| Flea control: pets/environmental | ||
| Environmental sanitation | ||
| Housing conditions |
a Opossums, rats, and stray/feral dogs and cats. bWolbachia pipientis, Steinina ctenocephali.
Table 2.
Population Growth of Texas and Texas Counties with Highest Flea-borne Typhus Cases, 1990 versus 2019 a.
Table 2.
Population Growth of Texas and Texas Counties with Highest Flea-borne Typhus Cases, 1990 versus 2019 a.
| 1990a | 2019 | Percent increase, 1990 to 2019 | |
|---|---|---|---|
| Texas | 16,986,510 | 28,995,881 | 70.7 |
|
Metropolitan (% of total) |
14,165,650 (83.4) |
25,920,625 (89.4) |
83.0 |
| non-metro | 2,820,852 | 3,075,261b | 9.0 |
| Bexar | 1,185,394 | 2,004,000 | 67.1 |
| Cameron | 260,120 | 423,163 | 62.7 |
| Harris | 2,833,000 | 4,713,000 | 66.3 |
| Hidalgo | 383,545 | 868,707 | 126 |
| Nueces | 291,145 | 362,294 | 24.4 |
| Travis | 576,407 | 1,274,000 | 121 |
a From US Census data, except where indicated. b [31].
Table 3.
Increase in average annual number of cases of FBT in Texas and the six highest incidence counties, 1990-1999 versus 2010-2019.
Table 3.
Increase in average annual number of cases of FBT in Texas and the six highest incidence counties, 1990-1999 versus 2010-2019.
| State or County |
Largest City | total number of cases 2010-2019a |
Avg number of cases/yr, 1990-1999b |
Avg number of cases/yr, 2010-2019a |
Fold increase in avg # of cases, 2010-2019 vs 1990-1999 |
|---|---|---|---|---|---|
| Texas | ----- | 3750 | 30.7 | 375.0 | 12.2 |
| Bexar | San Antonio | 422 | 0.2 | 42.2 | 211 |
| Cameron | Brownsville | 368 | 2.1 | 36.8 | 17.5 |
| Harris | Houston | 252 | 0.1 | 25.2 | 252 |
| Hidalgo | McAllen | 963 | 11.3 | 96.3 | 8.5 |
| Nueces | Corpus Christi | 580 | 11.7 | 58.0 | 5.0 |
| Travis | Austin | 239 | 0 | 23.9 | --- |
Table 4.
Estimated pet and stray dog populations in the six Texas counties with highest prevalence of flea-borne typhus, 1990-1999 and 2010-2019.
Table 4.
Estimated pet and stray dog populations in the six Texas counties with highest prevalence of flea-borne typhus, 1990-1999 and 2010-2019.
| County | Avg. Human Pop, 1994/ 1995a |
Est. Pet Dog Pop., 1994/ 1995b |
Est. Stray Dog Pop., 1994/ 1995c |
Est. Total Dog Pop., 1994/ 1995 |
Avg. Human Pop. 2014/ 2015a |
Est. pet dog pop. 2014/ 2015 |
Est. Stray Dog Pop, 2014/ 2015c |
Est. Total Dog Pop, 2014/ 2015 |
Increase in Total Dog Pop. 2014/2015 vs 1994/1995 |
|---|---|---|---|---|---|---|---|---|---|
| Bexar | 1,280,000 | 345,600 | 75,294 | 420,894 | 1,876,500 | 506,655 | 110,382 | 617,037 | 196,143 |
| Cameron | 287,204 | 77,545 | 16,894 | 94,439 | 420,705 | 113,590 | 24747 | 138,337 | 43,898 |
| Harris | 3.050,000 | 823,500 | 179,412 | 1,002,912 | 4,506,000 | 1,216,620 | 265059 | 1,481,679 | 478,767 |
| Hidalgo | 468,443 | 126,480 | 27,555 | 154,035 | 868,707 | 234,551 | 51,100 | 285,651 | 131,616 |
| Nueces | 310,079 | 83,721 | 18,240 | 101,961 | 362,294 | 97,819 | 21,311 | 119,130 | 17,169 |
| Travis | 656,303 | 177,202 | 38606 | 215,808 | 1,274,000 | 343,980 | 74,941 | 418,921 | 203,113 |
Table 5.
Estimated pet and stray cat populations in the six Texas counties with highest prevalence of flea-borne typhus, 1990-1999 and 2010-2019.
Table 5.
Estimated pet and stray cat populations in the six Texas counties with highest prevalence of flea-borne typhus, 1990-1999 and 2010-2019.
| County | Avg. Human Pop, 1994/ 1995a |
Est. Pet Cat Pop., 1994/ 1995b |
Est. Stray Cat Pop., 1994/ 1995c |
Est. Total Cat Pop., 1994/ 1995 |
Avg. Human Pop. 2014/ 2015a |
Est. Pet Cat pop. 2014/ 2015 |
Est. Stray Cat Pop, 2014/ 2015c |
Est. Total Cat Pop, 2014/ 2015 |
Increase in Total Cat Pop. 2014/2015 vs 1994/1995 |
|---|---|---|---|---|---|---|---|---|---|
| Bexar | 1,280,000 | 268,800 | 107,520 | 376,320 | 1,876,500 | 394,065 | 157,626 | 551,691 | 175,371 |
| Cameron | 287,204 | 60,313 | 24,125 | 84,438 | 420,705 | 88,348 | 35339 | 123,687 | 39,249 |
| Harris | 3.050,000 | 640,500 | 256,000 | 896,500 | 4,506,000 | 946,260 | 378,504 | 1,324,764 | 455,264 |
| Hidalgo | 468,443 | 98,373 | 39,349 | 137,722 | 868,707 | 182,428 | 72,971 | 255,399 | 117,677 |
| Nueces | 310,079 | 65,117 | 26,046 | 91,163 | 362,294 | 76,082 | 30,432 | 106,515 | 15,352 |
| Travis | 656,303 | 137,823 | 55,129 | 192.953 | 1,274,000 | 267,540 | 107,016 | 374556 | 181,603 |
Table 6.
The Number of Cases of Flea-Borne Typhus in Each of the Climatic Regions of Texas, 1990-1999 and 2010-2019 a,b.
Table 6.
The Number of Cases of Flea-Borne Typhus in Each of the Climatic Regions of Texas, 1990-1999 and 2010-2019 a,b.
| Region Numbera | Region Name/ Alternate Namea | Descriptionc | Number of FBT Cases, 1990sb | % of Total |
Number of FBT Cases, 2010sc |
% of Total |
Fold-increase |
|---|---|---|---|---|---|---|---|
| 1 | High Plains | Continental steppe or semi-arid savanna | 2 | 0.7 | 0 | 0 | Decreased |
| 2 | Low Rolling Plains | Sub-tropical steppe or semi-arid savanna | 7 | 2.3 | 5 | 0.13 | Decreased 0.7 |
| 3 | Cross Timbers/ North Central |
Sub-tropical subhumid mixed savanna, woodlands | 4 | 1.3 | 198 | 5.28 | 49.5 |
| 4 | Piney Woods/ East Texas |
Sub-tropical humid mixed evergreen-deciduous forestland | 2 | 0.7 | 26 | 0.69 | 13 |
| 5 | Trans Pecos | subtropical arid desert | 0 | 0 | 1 | 0.027 | ------ |
| 6 | Edwards Plateau | Sub-tropical steppe or semi-arid brushland, savanna | 5 | 1.6 | 56 | 1.49 | 11.2 |
| 7 | Post-Oak Savannah/ South Central |
Sub-tropical subhumid mixed prairie, savanna, woodlands | 129 | 41.3 | 1465 | 39.1 |
11.5 |
| 8 | Gulf Coastal Plains/ Upper Coast |
Sub-tropical humid marine prairies and marshes | 3 | 1.0 | 408 | 10.88 | 136 |
| 9 | South Texas Plains/ Southern | Sub-tropical steppe or semi-arid brushland | 16 | 5.2 | 227 | 6.05 | 14.2 |
| 10 | Lower Rio Grande Valley/ Lower Valley | Subtropical sub-humid marine | 139 | 45.2 | 1364 | 36.37 | 9.8 |
| Totals |
-------- |
------- |
307 |
86.5% in regions 7 and 10 | 3750 | 75.5% in regions 7 and 10 | 12.2 |
| Refugiumd | ------ | ------- | 273 (88.9%) |
2238 (59.7%) | 8.2-X | ||
| Outside Refugium | ------- | -------- | 34 (11.1%) |
1512 (40.3%) |
44.5-X |
Table 7.
Minimum, Mean, and Maximum Temperatures (ºF) in Texas Counties with the Highest Prevalence of Flea-borne Typhus Cases, 1990-1999 vs 2010-2019 a.
Table 7.
Minimum, Mean, and Maximum Temperatures (ºF) in Texas Counties with the Highest Prevalence of Flea-borne Typhus Cases, 1990-1999 vs 2010-2019 a.
| County | Tmin, °F | Tmean, °F | Tmax, °F | ||||||
| 1990s | 2010s | Diff | 1990s | 2010s | Diff | 1990s | 2010s | Diff | |
| Bexar | 58.93 | 59.43 | +0.5 | 69.60 | 70.18 | +0.58 | 80.27 | 80.93 | +0.66 |
| Cameron | 65.25 | 66.2 | +0.95 | 74.23 | 75.18 | +0.95 | 83.19 | 84.13 | +0.94 |
| Harris | 60.59 | 61.9 | +1.31 | 69.80 | 70.86 | +1.06 | 79.04 | 79.8 | +0.76 |
| Hidalgo | 63.98 | 64.61 | +0.63 | 74.73 | 75.51 | +0.78 | 85.46 | 86.37 | +0.91 |
| Nueces | 63.89 | 64.65 | +0.76 | 72.33 | 72.91 | +0.58 | 80.78 | 81.18 | +0.40 |
| Travis | 58.51 | 58.33 | -0.18 | 68.84 | 69.47 | +0.63 | 79.83 | 80.62 | +0.79 |
| Tarrant | 55.67 | 57.72 | + 2.05 | 65.90 | 67.64 | +1.74 | 76.60 | 77.57 | +0.97 |
aData from Prism Climate Group [11].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



