
Submitted:
18 November 2024
Posted:
19 November 2024
You are already at the latest version
Abstract
Olive (Olea europaea L.) trees produce many more flowers than fruit. In an “on” year, an adult olive tree may produce as many as 500,000 flowers, but 98% of them will drop before ripening. This waste of resources, better invested in fruit reaching maturation, needs an explanation. Different not mutually exclusive hypotheses elucidating the possible significance of heavy flowering followed by massive and premature fruit abscission are analyzed and confronted based on previously published works as well as on more recent own observations on olive reproductive biology. The results suggest that olive trees selectively abort fruit to increase the quality of the seed in the surviving fruit. A sizable portion of the flowers seems also to enhance the total fitness of the plant by pollen export. On the contrary, the hypotheses alluding to resource limitation, pollination deficits, pollinator attraction or extra flowers constituting an ovary reserve, as the ultimate functions of massive flower production must be rejected in olive. Consequences on olive orchard management are discussed.
Keywords:
Olea europaea
; massive flowering
; fruit abortion
; pollinator attraction
; pollination deficits
; male function of flowers
; resource limitation
; bet hedging
; sexual selection
; selective abortion
1. Introduction
Many plant species abort regularly a large portion of their flowers and juvenile fruits that can be developed up to maturity. The overproduction of flowers that not reach maturation is an extended feature of many Angiosperm, but it is especially common and particularly high in woody outcrossed hermaphrodite plants [1,2], including among them many fruit crops. In these species, surplus flowers and fruit are early abscised. This drop of juvenile fruit is known in pomology as “June drop” because, in the Northern Hemisphere, fruitlet abortion in apple takes mostly place in this month of June. This waste of energy is surprising because it appears that the resources invested in surplus flowers and aborting fruitlets could be better allocated in increasing the number and size of the fruit reaching maturation [3,4]. For this reason, many efforts had been devoted to determine the causes and significance of this massive flower and fruit abortion in cultivated fruit trees as well as in wild species. In this regard, different theories try to explain the benefits of the formation of these surplus flowers. Since the cost of over-initiating flowers and fruit is important, the benefits of their formation has to largely compensate the expenditure incurred in order to reasonably explain why this character is fixed and extended in the evolution of Higher Plants.
In olive, different research teams have explored the mechanisms operating in the massive flower and fruit abscission in order to suggest crop management techniques that might increase the low fruit set characteristic of this crop. An olive tree in its “on” year may produce as many as 500,000 flowers, but only 1-2% of them reach harvest as ripen fruit [5]. The remaining 98-99% of flowers abscise mainly within the first two weeks after bloom [6]. In this study, I check current hypotheses formulated elsewhere to explain the significance of the premature fruit abscission in olive. These theories are reviewed and confronted using the results of previous published and unpublished investigations as well as new knowledge about the reproductive biology of Olea europaea, with the aim to determine the ultimate reasons why olive produce many more flowers than fruit. Stephenson [1], in his prestigious review about this topic, distinguished proximate causes, as the mechanisms responsible for the failure of flowers to set fruit, from the ultimate reasons, that is, the functions surplus flowers may play in the reproduction of plants that persistently fail to reach maturation. This distinction is pertinent to the aim of this review.
2. The Plant
Domesticated olive tree (Olea europaea subsp. europaea L.) is a woody wind-pollinated, preferentially allogamous crop of immense importance in the Mediterranean area. Its longevity and history reach the category of Myth in several monotheist religions of the Mediterranean area, being frequently mentioned in the sacred texts of Christianism, Judaism, and Islamism. Olive is andromonoecious, forming hermaphrodite and staminate flowers in panicles of between 15 and 30 flowers in average, depending the number on the cultivar and on the growing conditions [7]. The panicles develop clustered in a high number in the axil of leaves of 1-year-old shoots (Figure 1).
Olive panicles are ramified and, in addition to the apical flower (“king flower”), have primary, secondary, and, eventually, tertiary branches [8]. Hermaphrodite flowers are mostly displayed in primary branches, while the staminate flowers are more frequent in secondary and higher order branches [9,10] (Figure 2).
Olive exhibits a strong alternative bearing habit, with “on” years of massive flowering followed by “off” years with no or few flowers. In its “on” year, an adult olive trees blooms profusely producing as many as 500,000 flowers [5]. However, not all these flowers reach harvest because up to 98-99% of them drop, as flowers or developing fruitlets, in the five-seven weeks following bloom [6]. Three main waves of abscission may be distinguished in olive. First wave of abscission affect to the staminate flowers that drop a few days after bloom, once they accomplished their mission of exporting their pollen grains [9]. The second wave of abscission affects to unfertilized, but fertile, hermaphrodite flowers that drop in the second week after bloom. Despite the magnitude of the drop of these two phases can be important, olive still initially set a much larger proportion of fruit that those reaching harvest. Thus, after fertilization of the flowers, many developing fruitlets start to growth triggering an intense fruitlet competition resolved with the abscission of many of them and the stabilization of the fruit population in the third wave of fruitlet abscission occurring five-seven weeks after flowering [11,12]. This last drop may partially overlap with the abscission of unfertilized flowers [13]. Whatever the reason for the failure of the flowers and fruit to reach maturity, sound reasons are needed for explaining the formation of these many surplus flowers and for over-initiating fruit that later irrevocably abscise. Below, I explore different hypotheses trying to explain the benefits of forming such an extraordinary number of surplus flowers in olive.
3. Hypotheses to Explain Massive Fruitlet Abortion
Six valid not mutually exclusive hypotheses try to explain the massive flower and fruitlet abscission in many hermaphroditic Angiosperm. The hypotheses explaining the production of surplus flowers are: (1) Pollinator attraction by which flowers serve to attract a higher number of biotic pollination vectors; (2) Abscission due to pollination deficits, either by the scarcity or the inadequacy of the pollen grains landing in the stigma, or because pollinators do not visit at all the flowers; (3) Excess flowers playing solely a male function, exporting their pollen and thus enhancing male and, consequently, the total fitness of the progenitor; (4) Flower drop due to resource, nutrient limitation, including here water deficits; (5) Bet-hedging theory that suggests that there is not surplus flowers, but that they are formed just in case unexpected good seasons might allow full fruit set; and (6) Sexual selection and selective abortion among developing fruitlet based on fruit and seed sink strength (genotype based).
The appropriateness of these hypotheses to explain massive flower and fruitlet abscission is examined based on my own and other colleagues’ previously published research on olive reproductive biology, and confronted to determine the ultimate reasons for the formation of surplus flowers in olive.
3.1. Pollinator Attraction
First theory postulates that an excess number of flowers are formed, especially when bloom occurs in inflorescences [14] to attract more intensively biotic pollinating agents (pollinators) seeking floral rewards at a lower cost. In this regard, the basic unit of attraction to pollinators is the inflorescence. However, olive is wind pollinated. Certainly, olive belong to the family Oleaceae, family where many species are insect-pollinated. In fact, olive flowers still possess a conspicuous white corolla and emit sweet aroma that attract to different insects, mainly honeybees [8]. This ambophily has been identified in the close relative Olea ferruginea [15]. Nonetheless, it is clear that olive has evolved toward anemophily and in its “on” year, adult plants might produce billions of pollen grains (around 50,000 million of pollen grains per tree extrapolating data from Cuevas and Polito [9] and Rojas-Gómez et al. [16]) (Figure 2); pollen that is readily transported by the wind as many allergic persons in the Mediterranean countries note and suffer (Figure 3).
Even although a relistic function of surplus flowers attracting insects cannot be left out of consideration, the true is that if surplus flowers played that role in olive, then an increasing number of flowers (as it is formed in “on” years) will produce a higher attraction to pollinators and then will result in higher reproductive success [17]. However, the truth is that the olive trees with more flowers set proportionally fewer fruit (Figure 4). On the contrary, trees in their “off” year set more fruit in average [18]. Consequently, surplus flowers do not function to attract pollinators in olive.
3.2. Pollination Deficits
A different theory explains that fruit set is limited because of pollination deficits. These deficits can be due to a shortage or inadequacy of the pollen grains or because pollinators absence [19,20]. A stated before, olive is wind-pollinated and a single adult tree might produce billions of pollen grains. Although wind is a random pollination vector, many field pollination experiments have demonstrated that most, if not all, hermaphrodite flowers receive enough pollen grains on the stigma to achieve fertilization. This has been proved in many different experiments in which pollen load on the stigmas has been counted in flowers left exposed under conditions of open free-pollination [21,22,23,24,25,26]. Lack of pollen in some stigmas has been rarely documented [12] and, when found, they might reflect lack of functionality of the stigma due to late abortion in a small portion of flowers rather than a problem of pollen transport.
A different approach here is to suggest that not the quantity, but the quality of the pollen might lead to the lack of fertilization, and therefore, to flower abscission in olive orchards. Self-incompatibility in olive has been recently confirmed to be of the sporophytic type and to severely affect to many olive cultivars [27,28]. But, while olive has been repeatedly confirmed as a self-incompatible species [21,27,28,29], the truth is that even under optimal cross-pollination treatments, massive flower and fruitlet abscission occurs. In fact, successful fertilization achieved by cross-pollen advances fruitlet abscission, because earlier and higher levels of flower fertilization enhance fruit growth and trigger the abscission of other less competitive fruitlets [12].
Fertilization levels of 40-50% of the flowers has been reported for different olive cultivars under cross-pollination conditions [21,22,30,31]. Furthermore, Rapoport and Rallo [6] found out that more than 50% of the abscised flowers of cultivar Manzanillo (syn. Manzanilla de Sevilla) were fertilized, ruling out pollination deficit as a cause of their drop. Unfortunately, high levels of fertilization in olive do not guarantee similar levels of final fruit set that, even under the best conditions, do not reach levels above 10% of the hermaphrodite flowers formed [12,21]. Certainly, the benefits of cross-pollination are undeniable, significantly increasing fruit set and yield in olive orchards [30,32,33,34], but pollination deficits, neither in quantity, nor in quality, are not enough to justify the cost of producing many more flowers than fruit in olive. Therefore, cross-pollination significantly increases fruit set in most cultivars, and although some level of pollination deficits might appear in large monovarietal orchards due to the strong self-incompatibility behavior of the olive, but even under the best pollination scenario, massive June drop occurs. Therefore, the pollination deficits theory does not operate in cultivated olive.
3.3. Male Function
The formulation of this hypothesis is that the surplus flowers that later drop are in fact accomplishing only a male function; that is, exporting pollen grains to achieve fertilization in flowers of other plants [35,36,37,38]. In other words, according to this theory, despite morphologically hermaphrodite, all or most abscised flowers are, in fact, functionally male, and this flower surplus formation is explained by enhancing the male competition component of sexual selection [39]. Extensive analyses of a high number of species demonstrate that in monoecious and/or dioecious species female flower abortion is lower than in hermaphrodite plants [2]. This pattern is explained because in monoecious and dioecious species, plants may dissociate the investments in male and female flower organs, adjusting therefore more tightly the resources invested in the formation of pistillate flowers to those potentially reaching maturation. If this were the case for olive, it is not easily understandable then, why olive has not transitioned toward monoecy, to form some of them just as staminate ones, saving resources in surplus pistil formation. In hermaphrodite plants, the resources invested in plant reproduction are divided between male (pollen) and female organs (fruit and seeds [1,40]; the latter much more expensive to produce. Evolution toward monoecy and/or dioecy would allow the plant to discern the investment in male and female organs according to the available resources and environmental constrains, but in olive, andromonoecy seems largely fixed [9] and the formation of hermaphrodite flowers in excess cannot be explained solely by the male function theory. On the other hand, the formation of an important number of fruitlet that later abscised during June drop period also denies that these flowers were only formed to accomplish the male function by exporting their pollen grains.
Staminate flowers are the product of pistil abortion (Figure 5B and D). Pistil abortion in olive may occur in different moments of flower development, but, in most instances, pistil growth is early arrested and flowers completely lack or present rudimentary pistils [41]. In some cases, however, pistil formation is almost complete, although the stigma papillae fail to develop fully so the flowers are not able to adhere pollen grains and, then, they are unable to set fruit (Figure 5D) [42]. The occurrence of pistil abortion in various stages of development suggests that olive tree is continuously balancing the amount of the available resources, water included, to invest in pistil development with poorer growing conditions leading to higher rates of pistil abortion. In this regard, the production of staminate flowers is extremely variable among cultivars, years, trees, branches and in even among panicles of the same shoot, but it is higher in dry seasons and under nutritional deficits [41].
Staminate flowers are also formed in less favorable sites within the panicles, suggesting again resource limitation as the cause of their formation. The selective placement of staminate flowers in worse nurtured sites within the panicle (secondary and tertiary branches) [9,10] (Figure 2) leads to some architectural negative effects on their fate that reinforce the idea that competition for resources plays a role in their formation (Seifi et al., 2008). Plant architecture (pedicel thickness) regulates and/or is associated with critical components of female reproductive success such as fruit and seed set, seed number and size [43,44]. In an apparent contradiction, the resources saved not developing a pistil in the staminate flowers of olive are not invested in the other organs of the same flower, as the smaller size (petal and sepals) of staminate flower compared to hermaphrodite ones indicate [9]. Nonetheless, the staminate flowers form an equal number of pollen grains per anther, pollen grains that are of the same size and viability, and with the same capacity to perform ovule fertilization in hermaphrodite flowers of a different cultivar [9]. The results then allow concluding that staminate flowers in olive serve to enhance male and total plant fitness at a lower cost, since the staminate flowers have the capacity to father the same number of embryos in other olive plants and are cheaper to form than producing complete flowers.
Therefore, certainly, an important proportion of olive flowers are staminate, and hence does not have the capacity to become fruit. Their effective achievement of the male function explains the formation of those staminate flowers, but cannot explain then why extra hermaphrodite flowers are still formed.
3.4. Resource Limitation
If the proportion of staminate flowers is, at least, partly explained by a shortage of resources, we may think that, perhaps, the reduced fruit set in hermaphrodite flowers is also due to the lack of resources, being this lack of resources circumstantial and not structural [45,46]. If this hypothesis is true and explains the formation of surplus flowers, then it implies that in some conditions, all or most hermaphrodite flowers might become fruit. Nonetheless, fruit set in olive is always very low, and there is not seasonal or spatial (orchard) variation in it that allows all flowers becoming mature fruit. Certainly good orchard management (especially irrigation and fertilization) increases fruit set in olive, but, unfortunately only a little, and never a in a percentage higher than 4-5% of the flowers formed.
It is clear, however, that competition for resources among developing fruitlets seems to be the proximate cause, the mechanism, triggering the intensive post-anthesis fruitlet abscission in olive, first by competition of the young fruitlet within the panicle, and later among fruit of nearby panicles. This conclusion is based on the results of different thinning experiments and by the observation that early fruit growth immediately precedes the abscission of fertilized and unfertilized flowers [12,13]. In this regard, thinning experiments have made clear that removing more than 50% of panicles increases fruit set of remaining flowers and does not reduce the final number of fruit at harvest (Lavee et al., 1996). This result confirms the surplus conditions of a high number of flowers (in some experiments up to 60%). The same conclusion is reached if the whole inflorescence is removed or if flower thinning is performed by eliminating some flowers within the panicle. The fact that the result is also the same regardless if flower removal is performed before or days after bloom [11,47,48] indirectly confirms again that extra flowers do not serve the purpose of attracting pollinating insects.
It is worth mentioning that small-fruited cultivars have a higher fruit set than large-fruited cultivars, although productivity per tree does not differ widely. In addition, small-fruited cultivars have smaller ovaries at bloom than large sized fruit cultivars [49,50], so they are less competitive, less selfish we may say. Similar results were observed when ‘Galego’ olive trees in pots were grown at low temperatures (20/14ºC). The lower growth rates of the fruit at this low temperature reduced fruitlet competition and allowed to double fruit set in comparison with potted trees growing at 25/20ºC, although the final size of the fruit at harvest was half at the lower temperature [51]. Cool springs have also a positive effects on olive fruit set.
Therefore, although good management may and should increase fruit set and yield in olive orchards, it would never do in such as extent to avoid June drop. On the other hand, flower thinning experiments confirm that competition for resources is the proximate cause of the abscission, but not the ultimate reason of why surplus flowers are formed in olive. A different explanation for the production of surplus hermaphrodite flowers is still needed.
3.5. Bet Hedging: Ovary Reserve Hypothesis
Bet hedging hypothesis has some point in common with resource limitation theory as the former justifies the formation of extra flowers based on higher fruit set in favorable seasonal conditions. In this regard, bet hedging theory states that the extra flowers are formed to take advantage of unexpected prodigious seasons in unpredictable environments [1,52]; that is, extraordinary seasons when plants plenty of resources (in nature, for Olea europaea, we will say rain) and high pollinator activity would allow full setting. However, in domesticated olive, there is not spatial nor temporal variation in fruit set that allow full fruit setting; neither, as stated before, under optimal orchard management, nor in exceptionally good years. The fact that many of the olive flowers drop after fertilization and at various stages of embryo development [13] suggests also that olive is incapable of maturing all hermaphrodite flowers formed and hoping that all of them will reach maturation is unrealistic. On the other hand, the activity of pollinating insect plays no role in olive reproduction, since, as stated before, olive is mostly wind-pollinated. Therefore, expecting optimal pollination in some extraordinary seasons when pollinators activity is great is not a suitable explanation for olive massive flower production. Consequently, bet hedging theory fails to justify the formation of surplus hermaphrodite flowers in cultivated olive trees.
A variant of bet-hedging theory is the ovary reserve hypothesis that explains that many flowers are formed to confront unpredictable external mortality non-maternally regulated [53], especially, but not only, herbivory [4]. In this theory, surplus flowers would act as an insurance against flower losses [54], or even as a bait for herbivores achieving seed predators satiation forming more flowers (and seeds) than needed [45,52]. If this were the case, we expect that this sacrifice of part of the pistils in benefit of the remainders would be extended through the whole reproductive season, offering siblings protection against seed predators from bloom to fruit maturation and seed dispersal. However, in olive, most juvenile fruit drop very early in the season leaving exposed sibling fruits to predators most of the reproductive period. It is still possible that the plant strategy is just to cover safely the pre-bloom period, with resource competition among fruitlets causing the abscission of weaker juvenile fruits. Ehrlen [53] proposed that the number of surplus flowers increases with increasing fruit-cost/flower-cost ratio and ovary mortality, and then, that fruit abortion would be higher in species forming costly fruit and small flowers as it happens in olive.
Guitián et al. [55] found that if flower mortality is high in Cornus sanguinea, then subsequent fruit abortion is lower, suggesting that in Cornus surplus flowers act, at least partially, as an insurance against predators. Similar conclusion is reached in Prunus mahaleb [56]. Certainly, in olive, larvae of some pests, like the second generation of Prays oleae, depredates on hermaphrodite flowers (eating the ovary), while the third generation of Prays damages small fruitlets, but not in such an extent to threaten olive production. On the other hand, if ovary reserve theory works out in olive, then we may expect ample seasonal variations in fruit set depending on the incidence of herbivory, and the truth is that fruit set in olive is always low and massive flower and fruitlet abscission also occurs in the absence of pest damages. We may argue that domestic olive cultivation protect the crop to avoid extensive damages in the orchards, although this is not always true. However, wild olives (Olea europaea subsp. sylvestris) and close relatives to olive as Olea europaea subsp. cuspidata do not set either many fruit per panicle and present similar levels of flower and fruit abscission [57,58].
Therefore, the conclusion is that surplus flowers are in fact formed in olive and that the elimination of many of them by herbivores or by thinning (or pruning) is compensated by enhancing fruit set in the survival flowers, but not up to a level to reach full setting. Therefore, circumstantial high levels of herbivory to the flowers do not seem reason enough for the expenditure achieved by olive forming twenty-fold more flowers than fruit at harvest.
3.6. Sexual Selection, Sibling Competition and Selective Abortion
Previous hypotheses fail to fully explain the ultimate reasons and significance of surplus flower production and massive fruit abortion in olive. The last hypothesis here discussed is that olive form more flowers and fruit than needed for selecting the best of them based on the quality of the next generation (“the survival of the fittest”; [59]) Selective abortion to enhance offspring quality has been reported in many species of multi-seeded fruit, where mother investment in fruit development is optimized by aborting fruit with less or weaker seeds [1,60,61]. Selective fruit abortion in a drupe is mostly based on the size and strength of the only one seed formed, since drupes rarely contains more than one seed. Certainly, some olive cultivars are prone to produced bi-seeded fruit, especially under cross-pollination. Being the endocarp the dispersal unit, the presence of two seeds within a stone is detrimental for the size of each individual seed [62], and likely for seedlings survival too by stressing sibling competition on the same ground site after bird dispersal of the endocarp.
I suggest that mother olive tree forms extra flowers and fruitlets for letting many of them drop. As stated before, this is not a behavior exclusive of olive trees, as many other woody plants, among them, many fruit crops, produce many more flowers than fruit. The right question itself extends to why many plants produce many more ovules than seeds, having in mind that seeds constitute the next generation of sporophytes, being the fruit only a vehicle for seed dispersal. Whichever the best question is, the right answer to me is that olive may form more flowers and ovules than fruit and seeds to allow the survival of the fittest seedlings, promoting the selection of the best embryos and the abortion of the weaker fruitlets. Lee [63] beautifully developed the gametophyte competition theory by which plants selectively mature fruit from ovaries in which pollen tube competition has been greater. He developed the hypothesis that embryos resulting from the fertilization of the ovules by faster pollen tubes will produce sporophytes with higher fitness. The limitations of resources and the differences in the quality of the pollen achieving fertilization will thus open the possibility for sexual selection of the developing fruits [60]. Reports of experimentation giving support to the theory of selective abortion, sexual selection and sibling competition and how it operates in olive follows.
Olive proceeds to select the best embryos by selecting the most rapid pollen tube (containing the male gametes), the most attractive ovule (enclosing the embryo sac with the egg cell and polar nuclei) for double fertilization, and the strongest siblings after fertilization. First step in the sexual selection in olive takes place in the transmitting tissue of the recipient pistil. One single olive pistil may receive in the stigma more than 1,000 pollen grains, many of which germinate and grow forming pollen tubes carrying the male gametes in their tips. The first selection of the male gamete is based on the adequacy of the genotype of the pollen grain, since most olive cultivars are preferentially allogamous and discriminate against self-pollen by means of self-incompatibility reactions. Nonetheless, under open- and cross-pollination conditions is common to see inside many flowers dozens of pollen tubes growing in the transmitting tissue of the pistil, although only one pollen tube, very rarely two, is able to reach the base of the style and enter the ovary (Figure 6). An intense pollen tube growth attrition is responsible for this only one pollen tube gaining access to the ovary, where now, the winner fastest pollen tube finds four viable ovules within the ovary.
Next, not only male gametes are a subject of intense competition with the transmitting tissue of the pistil, but also the female gametes contained within the ovules can also be subjected to selective pressures, because after analyzing possible reasons for surplus flowers, one may rightfully ask why olive flowers contains four ovules when only one becomes a seed? Ovules in olive are equally-sized at bloom, occupy symmetric positions within the ovary and do not seem to differ in fertility or longevity. Secondary ovules exist in the flowers of stone fruit crops, but they differ in development with the primary ovule in a more advanced stage being normally the only one fertilized. However, in olive, there are no obvious reasons why the selected pollen tube accessing to the ovary may choose one or another ovule. Nonetheless, this election cannot be done by chance. Cuevas et al. [64] have proposed that this scenario allows ovule competition for the only pollen tube accessing to the ovary, an unusual case of female competition and male choice.
As the pollen tube exits the transmitting tissue, it responds to attractant signals from the ovule, and extends its growth along the surface of the funiculus, and the proceeds towards the micropyle of the ovule to perform the double fertilization process exclusive of Angioserm. In this regard, we know that synergids cells are implicated in the attraction of the pollen tube to the filiform apparatus [65,66]. Ion Calcium is, on this point, a crucial signaling element for pollen tube guidance and fertilization. As the pollen tube approaches the filiform apparatus of the ovule, the attractants excreted by the synergid cells affect the Ca2+ concentration at the pollen tube tip, facilitating the interaction between the pollen tube and the synergid cell receiving the discharge of the male gametes [67]. The synergids also provide some resources in this last step of pollen tube growth before discharging the two male gametes into the embryo sac [66,68]. A differential capacity of the synergids cell of different ovules to attract the pollen tube might explain in olive the preference of the winner pollen tube to select that ovule based on its strength, and the mere formation of four ovules per flower instead only one becoming seed. This could represent the first reported case of ovule (female) selection and male choice in Flowering Plants [64], even though this circumstance may be present in many other multi-ovulated ovaries sharing the same characteristics than olive. It is worth noting that synergids genes are not transcending to the next generation, as the egg cells genes do, after double fertilization. Thus, the role of the synergids, sisters of the egg cell, may represent a singular case of altruism.
If the strong selection of male and female gametes would not be enough, olive promotes the competition of the siblings formed after fertilization, by displaying many flowers together in a single panicle (the unit of fructification). This display enhances competition by the limited available resources, first within the panicle, and then followed by competition among developing fruit of nearby panicles. Flower display in large inflorescence and simultaneous flower opening reinforce effective sexual selection and competition among siblings by aborting those weaker fruitlets that grow at a lower rate [35]. Similar observation have been carried out in Prunus mahaleb, where the inflorescence also functions as the fructification unit [56]. In this regard, fruitlet abscission has been demonstrated to mainly affect the smaller fruitlets, either because the fertilization was achieved later (by a slower pollen tube) or because the seed is weaker and able to grow only in a lower rate. Either of those reasons diminish fruitlets competitiveness for survival leading to their drop. In olive, it has not yet been elucidated which flowers will become fruit, if the first pistils being fertilized or the pistils growing faster [12]. On the contrary, Hiei and Ohara [69] have observed in Melampyrum roseum that the flower better located in the inflorescence and opening earlier show higher fruit set. Medrano et al. [70] suggest that the sequestration of resources by the earliest developing fruit in Pancratium maritimum is also the cause of the abscission of the smaller fruitlets.
Simultaneous and massive full bloom is a prerequisite to make sexual selection more effective, since scattered flowering reduces sibling competition and sexual selection. In olive, only slight differences in phenology occurs within the panicle and also with nearby panicles, with bloom mostly co-occurring. In some species, it has been demonstrated that the percentage of fruitlet product of self-fertilization diminish along the period of fruitlet abscission because mother plants select fruitlet produce of cross-fertilization [1]. In avocado, this selection leads to the production of larger fruit, thus benefiting farmers’ profits [71,72]. This demonstration presents some challenges in olive for the difficulty extracting DNA from abscised fruitlet and dead seeds. Sexual selection in domestic olive might indirectly benefit to the olive farmer, since we have probed that seed weight is linearly related to fruit weight, reflecting seed sink strength to attract photoassimilates for fruit growth [62]. Seed size, in turn, is related to seedling survival and might be critical in harsh Mediterranean environments. The final aim of such strong selection of the seedlings in wild olives lies in producing seeds of better quality able to survive after germination in a harsh Mediterranean environment where prolonged drought and poor soils are common.
4. Consequences on Orchard Management: Future Prospects
Consequences of the strong seedling selection extends, however, to different aspects of orchard management in domestic olive. In this regard, farmers have to understand that massive flower and fruitlet abscission are part of the reproductive system of olive, and cannot be eliminated by increasing fertilization and/or by over-irrigation in their orchards. Lack of resources is not the ultimate reasons why olive produce so many flowers and abort regularly many fruit. Certainly, flower thinning in olive increases fruit/flower ratio, but not the number of fruit per panicle. Therefore, the formation of the surplus flowers is not explained by resource limitation, although the competition for resources is the mechanism by which olive selectively abort less competitive fruitlets. Therefore, extra flowers are mostly formed in olive to achieve the selection of the best embryos by means of the selective abortion of the weaker fruitlets. Therefore, neither irrigation (water is the main limiting factor for olive production), nor heavy orchard fertilization would avoid fruitlet competition and drop, although these orchard management techniques increase fruit set and, hence the percentage of flowers reaching harvest, by enhancing available plant resources.
Flower thinning and pruning limit the population of flowers and then reduce fruitlet competition. However, since fruit competition starts within the panicle, and extends later between nearby panicles, any of these techniques would not be able to make disappear fruitlet abscission in olive. The fact that fruit load at harvest does not change after removing a large proportion of flowers at bloom means that fruit set is proportionally increased by the limitation of the population of flowers. Nonetheless, heavy flowering in an “on” year usually leads to high yields, what in turn, triggers the inhibition of flower induction for the next season [73], and therefore to a much reduced crop in the following “off” year, a behavior typical of alternate bearer cultivars. This suggests that the selection of cultivars with lower flowering levels will diminish the wastage of resources due to heavy flowering and reduce olive alternate bearing habit [5]. It is important to note, however, that this selection should be based on genotypes with less flower per panicle and panicles uniformly distributed around the canopy to minimize fruitlets competition. Then, chemical fruit thinning in olive oil cultivars should be explored.
I mentioned before that pruning and flower thinning reduce sibling competition by limiting the number of flowers, keeping in the last case (flower thinning) the same level of resources and not affecting the source of photoassimilates, the leaves. These techniques increase fruit size as it is well known in many fruit crops. However, diminishing competition with fewer fruitlet would not allow a selection as effective as when many siblings compete. On the other hand, cross-pollination, other factors no limiting, allows reaching full production potential in olive, increasing initial fruit set and seed size, but an increasingly larger fruit may cause heavier fruit drop, if the available resources for fruiting remain the same. In this regard, small-fruited olive cultivars set a higher proportion of flowers by reducing fruit abscission [50].
On the other hand, more genetically similar fruit lead to less sibling rivalry and likely less fruit abortion [46]. On the contrary, seeds that are more diverse enhance fruit competition and differences in seed and fruit size. It is well-known that seed size is a fundamental trait affecting seed germination and seedlings survival and growth [74,75,76]. In any case, cross-pollination benefits in self-incompatible olive cultivars are unquestionable. In order to maximize fruit set, cross-pollination must occur extensively in all panicles, so a proper pollination design and a suitable selection of pollinizers including their number and placement in the orchard must be achieved. The selection of the pollinizer must be based on their inter-compatibility relationships and contemporaneous blooming. Destination of the olive fruit (oil versus table), regular bearing and similar vigor are some other criteria for selecting pollinizers [26]. On the contrary, no evidence of xenia has been found in olive [33], although seeds (and fruit) are slightly heavier under compatible cross-pollination treatments (Cuevas et al., in prep.)
The final result is that from 500,000 flowers formed in “on” olive trees, only around 6,000 fruit will reach ripening, and this still constitutes a good yield [5]. Orchard management techniques must procure this level of fructification. This analysis can be applicable to many other fruit crops especially to those blooming in panicles like mango, avocado and loquat, although the suitability of the hypotheses must consider the specific reproductive strategies, pollination requirements and seeding patterns.
Author Contributions
J.C. performed the conceptualization and writing-original and edited- of this review. J.C. has read and agreed to the published version of this manuscript.
Funding
Julián Cuevas received partial support by the University of Almeria’s programme for re-search and knowledge transfer years 2023 and 2024.
Data Availability Statement
No new data were created in this study. Data sharing is not applicable to this article.
Acknowledgments
To all who inspire this review, in special to the colleagues of the Department of Agronomy at the University of Córdoba (Spain) who performed many of the experiments here cited.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stephenson, A.G. Flower and fruit abortion: Proximate causes and ultimate functions. Ann. Rev. Ecol. Syst. 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Sutherland, S. Patterns of fruit-set: what controls fruit-flower ratios in plants? Evolution 1986, 40, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.G. Sexual strategies in plants. I. An hypothesis of serial adjustment of maternal investment during one reproductive season. New Phytol 1980, 86, 69–80. Available online: https://www.jstor.org/stable/2434416. [CrossRef]
- Stephenson, A.G. Fruit set, herbivory, fruit reduction, and the fruiting strategy of Catalpa speciosa (Bignoniaceae). Ecology 1980, 61, 57–64. [Google Scholar] [CrossRef]
- Martin, G.C. Olive flower and fruit population dynamics. Acta Hortic. 1990, 286, 141–154. [Google Scholar] [CrossRef]
- Rapoport, H.F.; Rallo, L. Postanthesis flower and fruit abscission in ‘Manzanillo’ olive. J. Amer. Soc. Hort. Sci. 1991, 116, 720–723. [Google Scholar] [CrossRef]
- Seifi, E.; Guerin, J.; Kaiser, B.; Sedgley, M. Flowering and fruit set in olive: a review. Iran. J. Plant Physiol. 2015, 5, 1263–1272. [Google Scholar]
- Lavee, S. Olea europaea. In Handbook of Flowering; Halevy, A.H., Ed.; CRC Press: Boca Raton, Fl, USA, 1985; Volume 6, pp. 423–434. [Google Scholar]
- Cuevas, J.; Polito, V.S. The role of staminate flowers in the breeding system of Olea europaea (Oleaceae): an andromonoecious, wind-pollinated taxon. Ann. Bot. 2004, 93, 547–553. [Google Scholar] [CrossRef]
- Seifi, E.; Guerin, J.; Kaiser, B.; Sedgley, M. Inflorescence architecture of olive. Sci. Hortic. 2008, 116, 273–279. [Google Scholar] [CrossRef]
- Rallo, L.; Fernández-Escobar, R. Influence of cultivar and flower thinning within the inflorescence on competition among olive fruit. J. Amer. Soc. Hort. Sci. 1985, 110, 303–308. [Google Scholar] [CrossRef]
- Cuevas, J.; Rapoport, H.F.; Rallo, L. Relationship among reproductive processes and fruitlet abscission in 'Arbequina' olive. Adv. Hortic. Sci. 1995, 9, 92–96. [Google Scholar]
- Rapoport, H.F.; Rallo, L. Fruit set and enlargement in fertilized and unfertilized olive ovaries. HortSci. 1991, 29, 890–898. [Google Scholar] [CrossRef]
- Willson, M.F.; Price, P.W. The evolution of inflorescence size in Asclepias (Asclepiadaceae). Evolution, 1977; 31, 495–511. [Google Scholar] [CrossRef]
- Khan, S.; Kumari, P.; Verma, S. Ambophily in Olea ferruginea: a transitional state in the pollination syndrome. Plant Biosystems 2022, 157, 221–232. [Google Scholar] [CrossRef]
- Rojas-Gómez, M.; Moral, J.; López-Orozco, R.; Cabello, D.; Oteros, J.; Barranco, D.; Galán, C.; Díez, C.M. Pollen production in olive cultivars and its interannual variability. Ann. Bot, 2023; 132, 1145–1158. [Google Scholar] [CrossRef]
- Guitian, J. 1993. Why Prunus mahaleb (Rosaceae) produces more flowers than fruits. Am. J. Bot. 1993, 80, 1305–1309. [Google Scholar] [CrossRef]
- Cuevas, J.; Rallo, L.; Rapoport, H.F. Crop load effects on floral quality in olive. Sci. Hortic. 1994, 59, 123–130. [Google Scholar] [CrossRef]
- Willson, M.F.; Schemske, D.W. Pollinator limitation, fruit production, and floral display in pawpaw (Asimina triloba). Bull. Torrey Bot. Club 1980, 107, 401–408. [Google Scholar] [CrossRef]
- Bawa, K.S.; Webb, C.J. Flower, fruit and seed abortion in tropical forest trees: implications for the evolution of paternal and maternal reproductive patterns. Am. J. Bot. 1984, 71, 736–751. [Google Scholar] [CrossRef]
- Bradley, M.V.; Griggs, W.H. Morphological evidence of incompatibility in Olea europaea L. Phytomorphology 1963, 13, 141–156. [Google Scholar]
- Ateyyeh, A.F.; Stösser, R.; Qrunfleh, M. Reproductive biology of the olive (Olea europaea L.) cultivar 'Nabali Baladi'. J. App. Bot. 2000, 74, 255–270. [Google Scholar]
- Seifi, E.; Guerin, J.; Kaiser, B.; Sedgley, M. Sexual compatibility and floral biology of some olive cultivars. N. Z. J. Crop Hortic. Sci. 2011, 39, 141–151. [Google Scholar] [CrossRef]
- Sánchez-Estrada, A.; Cuevas, J. ‘Arbequina’ olive is self-incompatible. Sci. Hortic. 2018, 230, 50–55. [Google Scholar] [CrossRef]
- Sánchez-Estrada, A.; Cuevas, J. Pollen-pistil interaction in ‘Manzanillo’ olive (Olea europaea L.) under self-, free- and cross-pollination. Rev. Chapingo Ser. Hortic. 2019, 25, 141–150. [Google Scholar] [CrossRef]
- Cuevas, J.; Chiamolera, F.M.; Pinillos, V.; Rodríguez, F.; Salinas, I.; Cabello, D.; Arbeiter, A.B.; Bandelj, D.; Raboteg Božiković, M.; Vuletin Selak, G. Arbosana olive is self-incompatible, but inter-compatible with some other low-vigor olive cultivars. Horticulturae 2024, 10, 739. [Google Scholar] [CrossRef]
- Breton, C.M.; Bervillé, A. New hypothesis elucidates self-incompatibility in the olive tree regarding S-alleles dominance relationships as in the sporophytic model. C. R. Biol. 2012, 335, 563–572. [Google Scholar] [CrossRef]
- Saumitou-Laprade, P.; Vernet, P.; Vekemans, X.; Billiard, S.; Gallina, S.; Essalouh, L.; Mhaïs, A.; Moukhli, A.; El Bakkali, A.; Barcaccia, G.; Alagna, F.; Mariotti, R.; Cultrera, N.G.M.; Pandolfi, S.; Rossi, M.; Khadari, B.; Baldoni, L. Elucidation of the genetic architecture of self-incompatibility in olive: Evolutionary consequences and perspectives for orchard management. Evol. Appl. 2017, 10, 867–880. [Google Scholar] [CrossRef]
- Cuevas, J. Incompatibilidad polen-pistilo. In Variedades de olivo cultivadas en España, 1st ed.; Barranco, D., Caballero, J.M., Martín, A., Rallo, L., Del Río, C., Tous, J., Trujillo, I., Eds.; Junta de Andalucía, Mundi-Prensa and COI: Seville, Spain, 2004; pp. 303–308. [Google Scholar]
- Cuevas, J.; Polito, V. S. Compatibility Relationships in `Manzanillo' Olive. HortSci. 1997, 32, 1056–1058. [Google Scholar] [CrossRef]
- Vuletin Selak, G.; Cuevas, J.; Goreta Ban, S.; Perica, S. Pollen tube performance in assessment of compatibility in olive (Olea europaea L.) cultivars. Sci. Hortic. 2014, 165, 36–43. [Google Scholar] [CrossRef]
- Bradley, M.V.; Griggs, W.H.; Hartmann, H.T. Studies on self- and cross-pollination of olives under varying temperature conditions. Calif. Agric. 1961, 15, 4–5. [Google Scholar] [CrossRef]
- Griggs, W.H.; Hartmann, H.T.; Bradley, M.V.; Iwakiri, B.T.; Whisler, J.E. Olive pollination in California, Bulletin 869; University of California – Agricultural Experiment Station: Berkeley, USA, 1975; pp. 1–49. [Google Scholar]
- Lavee, S.; Datt, A.C. The necessity of cross-pollination for fruit set of Manzanillo olives. J. Horticult. Sci. 1978, 53, 261–266. [Google Scholar] [CrossRef]
- Willson, M.F. , Rathcke, B.J. Adaptative design of the floral display in Asclepias syriaca L. Am. Midland Naturalist 1974, 92, 47–57. [Google Scholar] [CrossRef]
- Sutherland, S.; Delph, L.F. On the importance of male fitness in plants: patterns of fruit-set. Ecology 1093, 65, 1093–1104. [Google Scholar] [CrossRef]
- Sutherland, S. Why hermaphroditic plants produce many more flowers than fruits: experimental tests with Agave mckelveyana. Evolution, 1987; 41, 750–759. [Google Scholar] [CrossRef]
- Queller, D. Pollen removal, paternity, and the male function of flowers. Am. Nat. 1997, 149, 585–594. [Google Scholar] [CrossRef]
- Queller, D. Sexual selection in a hermaphroditic plant. Nature 1983, 305, 706–707. [Google Scholar] [CrossRef]
- Lloyd, D.G. Parental strategies of angiosperms. N. Z. J. Bot. 1979, 17, 595–606. [Google Scholar] [CrossRef]
- Uriu, K. Periods of pistil abortion in the development of the olive flower. Proc. Amer. Soc. Hort. Sci. 1959, 73, 194–202. [Google Scholar]
- Cuevas, J.; Pinney, K.; Polito, V.S. Flower differentiation, pistil development and pistil abortion in olive. Acta Hortic. 1999, 474, 293–296. [Google Scholar] [CrossRef]
- Diggle, P.K. Architectural effects and the interpretation of patterns of fruit and seed development. Annu. Rev. Ecol. Evol. Syst. 1995, 26, 531–552. [Google Scholar] [CrossRef]
- Wolfe L., M.; Denton, W. Morphological constraints on fruit size in Linaria canadensis. Int. J. Plant. Soil Sci. 2002, 162, 1313–1316. [Google Scholar] [CrossRef]
- Kelly, D.; Sork, V.L. Mast seeding in perennial plants: why, how, where? Ann. Rev. Ecol. and System. 2002, 33, 427–447. [Google Scholar] [CrossRef]
- Pías, B.; Salvande, M.; Guitián, P. Variation in predispersal losses in reproductive potential in rowan (Sorbus aucuparia L. Rosaceae) in the NW Iberian Peninsula. Plant Ecol. 2007, 188, 191–203. [Google Scholar] [CrossRef]
- Lavee, S.; Rallo, L.; Rapoport, H.F.; Troncoso, A. The floral biology of the olive: effect of flower number, type and distribution on fruitset. Sci. Hortic. 1996, 66, 149–158. [Google Scholar] [CrossRef]
- Suarez, M.P.; Fernández-Escobar, R.; Rallo, L. Competition among fruits in olive II. Influence of inflorescence or fruit thinning and cross-pollination on fruit set components and crop efficiency. Acta Hortic 1984, 149, 131–143. [Google Scholar] [CrossRef]
- Rosati, A.; Zipanćič, M.; Caporali, S.; Padula, G. Fruit weight is related to ovary weight in olive (Olea europaea L.). Sci. Hortic. 2009, 122, 399–403. [Google Scholar] [CrossRef]
- Rosati, A.; Zipanćič, M.; Caporali, S.; Paoletti, A. Fruit set is inversely related to flower and fruit weight in olive (Olea europaea L.). Sci. Hortic. 2010, 126, 200–204. [Google Scholar] [CrossRef]
- Cuevas, J.; Rallo, L. Respuesta a la polinización cruzada en olivo bajo diferentes temperaturas. Actas de Horticultura 1988, 1, 203–208. [Google Scholar]
- Janzen, D.H. Escape of Cassia grandis L. beans from predators in time and space. Ecology 1971, 52, 964–979. [Google Scholar] [CrossRef]
- Ehrlen, J. Why do plants produce surplus flowers? A reserve-ovary model. Am. Nat. 1991, 138, 918–933. [Google Scholar] [CrossRef]
- Brown, A.O.; McNeil, J.N. Fruit production in cranberry (Vaccinium macrocarpon): a bet-hedging strategy to optimize reproductive effort. Am. J. Bot. 2006, 93, 910–916. [Google Scholar] [CrossRef]
- Guitián, J.; Guitián, P.; Navarro, L. Fruit set, fruit reduction, and fruiting strategy in Cornus sanguinea (Cornaceae). Am. J. Bot. 1996, 83, 744–748. [Google Scholar] [CrossRef]
- Guitian, J. Selective fruit abortion in Prunus mahaleb (Rosaceae). Am. J. Bot. 1994, 81, 1555–1558. [Google Scholar] [CrossRef]
- Mulas, M. Characterisation of olive wild ecotypes. Acta Hortic. 1999, 474, 121–124. [Google Scholar] [CrossRef]
- Khadivi, A.; Mirheidari, F.; Saeidifar, A.; Moradi, Y. Morphological characterizations of Olea europaea subsp. cuspidata. Genet. Resour. Crop Evol. 2024, 71, 1837–1853. [Google Scholar] [CrossRef]
- Darwin, C.R. 1869. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. 5th ed.; John Murray: London, Great Britain.
- Willson, M.F. Sexual selection in plants. Am. Nat. 1979, 113, 777–790. [Google Scholar] [CrossRef]
- Stephenson, A.G.; Winsor, J.A. Lotus corniculatus regulates offspring quality through selective fruit abortion. Evolution 1986, 40, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.; Oller, R. Olive seed set and its impact on seed and fruit weight. Acta Hortic. 2002, 586, 485–488. [Google Scholar] [CrossRef]
- Lee, T.D. Patterns of fruit maturation: a gametophyte competition hypothesis. Am. Nat. 1984, 123, 427–432. [Google Scholar] [CrossRef]
- Cuevas, J.; Rallo, L.; Rapoport, H.F. Pollen tube growth and ovule abortion in Olea europaea (Oleaceae): A case if ovule selection. In Pollination Mechanisms, Ecology and Agricultural Advances; Raskin, N.D., Vuturro, P.T., Eds.; Nova Science Publisher Inc.: New York, USA, 2011; pp. 57–72. [Google Scholar]
- Sauter, M. A. A guided tour: Pollen tube orientation in flowering plants. Chin. Sci. Bull. 2009, 54, 2376–2382. [Google Scholar] [CrossRef]
- Kanaoka, M.M. Cell–cell communications and molecular mechanisms in plant sexual reproduction. J. Plant Res. 2018, 131, 37–47. [Google Scholar] [CrossRef]
- Iwano, M.; Ngo, Q.A.; Entani, T.; Shiba, H.; Nagai, T.; Miyawaki, A.; Isogai, A.; Grossniklaus, U.; Takayama, S. Cytoplasmic Ca2+ changes dynamically during the interaction of the pollen tube with synergid cells. Development 2012, 139, 4202–4209. [Google Scholar] [CrossRef]
- Takeuchi, H.; Higashiyama, T. Attraction of tip-growing pollen tubes by the female gametophyte. Curr. Opin. Plant Biol. 2011, 14, 614–621. [Google Scholar] [CrossRef]
- Hiei, K.; Ohara, M. Variation in fruit- and seed set among and within inflorescences of Melampyrum roseum var. japonicum (Scrophulariaceae). Plant Species Biol. 2002, 17, 13–23. [Google Scholar] [CrossRef]
- Medrano, M.; Guitián, P.; Guitián, J. Patterns of fruit and seed set within inflorescences of Pancratium maritimum (Amaryllidaceae): nonuniform pollination, resource limitation, or architectural effects? Am. J. Bot. 2000, 87, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Hapuarachchi, N.S.; Kämper, W.; Hosseini Bai, S.; Ogbourne, S.M.; Nichols, J.; Wallace, H.M.; Trueman, S.J. Selective retention of cross-fertilised fruitlets during premature fruit drop of Hass avocado. Horticulturae 2024, 10, 591. [Google Scholar] [CrossRef]
- Alcaraz, M.L.; Hormaza, J.I. Fruit set in avocado: pollen limitation, pollen load size, and selective fruit abortion. Agronomy 2021, 11, 1603. [Google Scholar] [CrossRef]
- Stutte, G.W.; Martin, G.C. Effect of killing the seed on return bloom of olive. Sci. Hortic. 1986, 29, 107–113. [Google Scholar] [CrossRef]
- Stanton, M.L. Seed variation in wild radish: effect of seed size on components of seedling and adult fitness. Ecology 1984, 65, 1105–1112. [Google Scholar] [CrossRef]
- Nakamura, R. Seed abortion and seed variation within fruits of Phaseolus vulgaris: pollen donor and resource limitation effects. Am. J. Bot. 1988, 75, 1003–1010. [Google Scholar] [CrossRef]
- Zhang, J.; Maun, M.A. Seed size variation and its effects on seedling growth in Agropyron psammophilum. Bot. Gaz. 1990, 151, 106–113. [Google Scholar] [CrossRef]
Figure 1.
Massive bloom in an olive in its “on” year. Panicles develop clustered on 1-year-old shoots.
Figure 1.
Massive bloom in an olive in its “on” year. Panicles develop clustered on 1-year-old shoots.

Figure 2.
Olive panicles showing hermaphrodite and staminate flowers. Left: Intact panicle. Right: the petals have been removed to better expose the well-developed pistil (white arrow) or the rudimentary pistil (black arrow). Source: Esmaeil Seifi Ph.D Thesis; with permission.
Figure 2.
Olive panicles showing hermaphrodite and staminate flowers. Left: Intact panicle. Right: the petals have been removed to better expose the well-developed pistil (white arrow) or the rudimentary pistil (black arrow). Source: Esmaeil Seifi Ph.D Thesis; with permission.

Figure 3.
Olive pollen clouds common during blooming season in Andalusia (Spain).

Figure 4.
Fruitful shoots taken from an “on” and an “off” olive tree. Despite “off” trees set a proportionally higher number of flowers, the number of fruit is still much larger in “on” trees, because the number of inflorescences is much higher in the former. Note different shoot length. Fruit ripening occurs before in “off” trees.
Figure 4.
Fruitful shoots taken from an “on” and an “off” olive tree. Despite “off” trees set a proportionally higher number of flowers, the number of fruit is still much larger in “on” trees, because the number of inflorescences is much higher in the former. Note different shoot length. Fruit ripening occurs before in “off” trees.

Figure 5.
Scanning Electron Microscopy pictures of hermaphrodite (left) and staminate (right) olive flowers. A. Detail of well-developed stigma (SG) in front of one stamen (ST). B. Pistil aborted flower showing rudimentary pistil (PI). C. Pollen germination on well-developed stigma papillae. D. Functionally male, late pistil-aborted flower, showing stigma with stigma papillae.
Figure 5.
Scanning Electron Microscopy pictures of hermaphrodite (left) and staminate (right) olive flowers. A. Detail of well-developed stigma (SG) in front of one stamen (ST). B. Pistil aborted flower showing rudimentary pistil (PI). C. Pollen germination on well-developed stigma papillae. D. Functionally male, late pistil-aborted flower, showing stigma with stigma papillae.
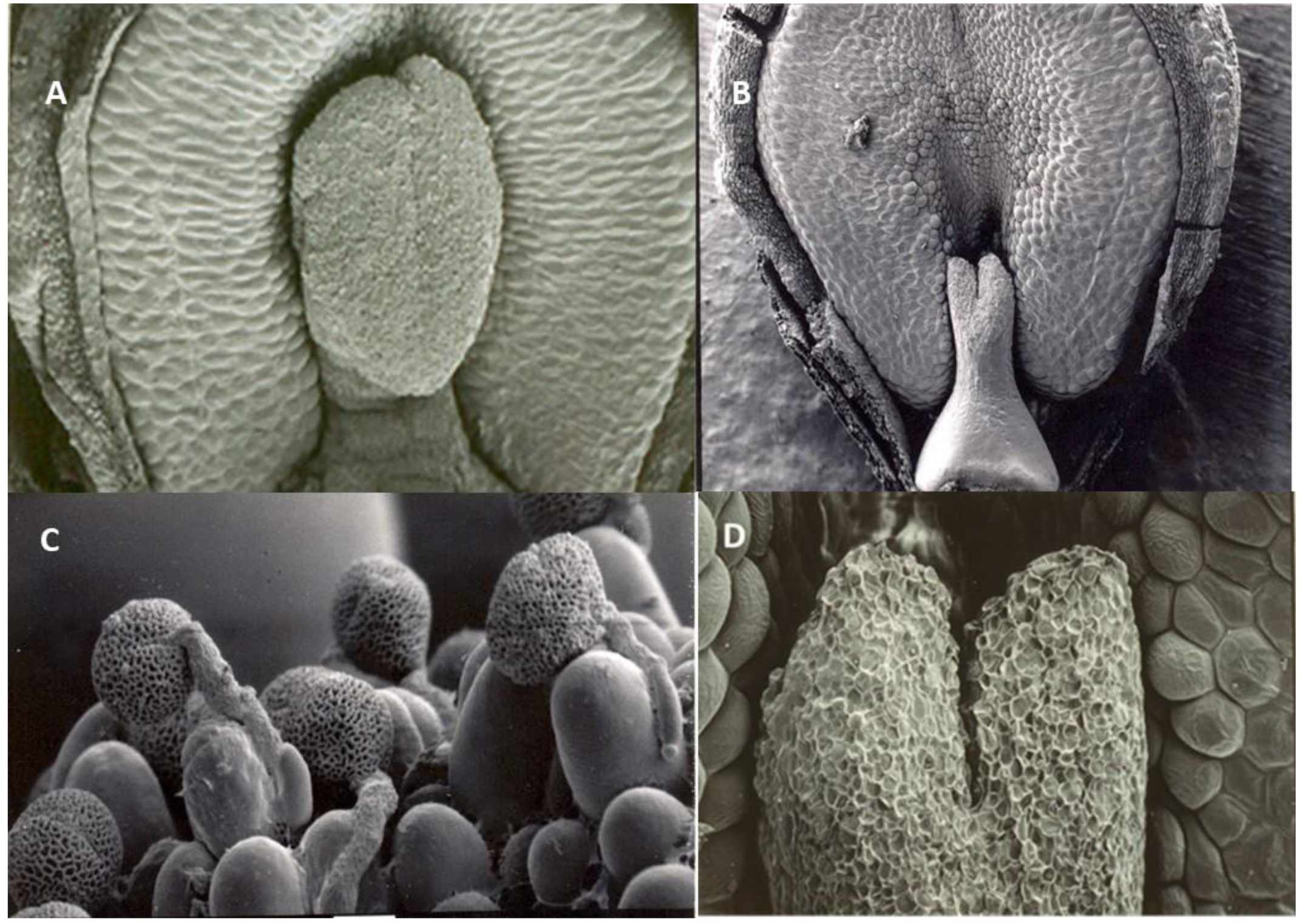
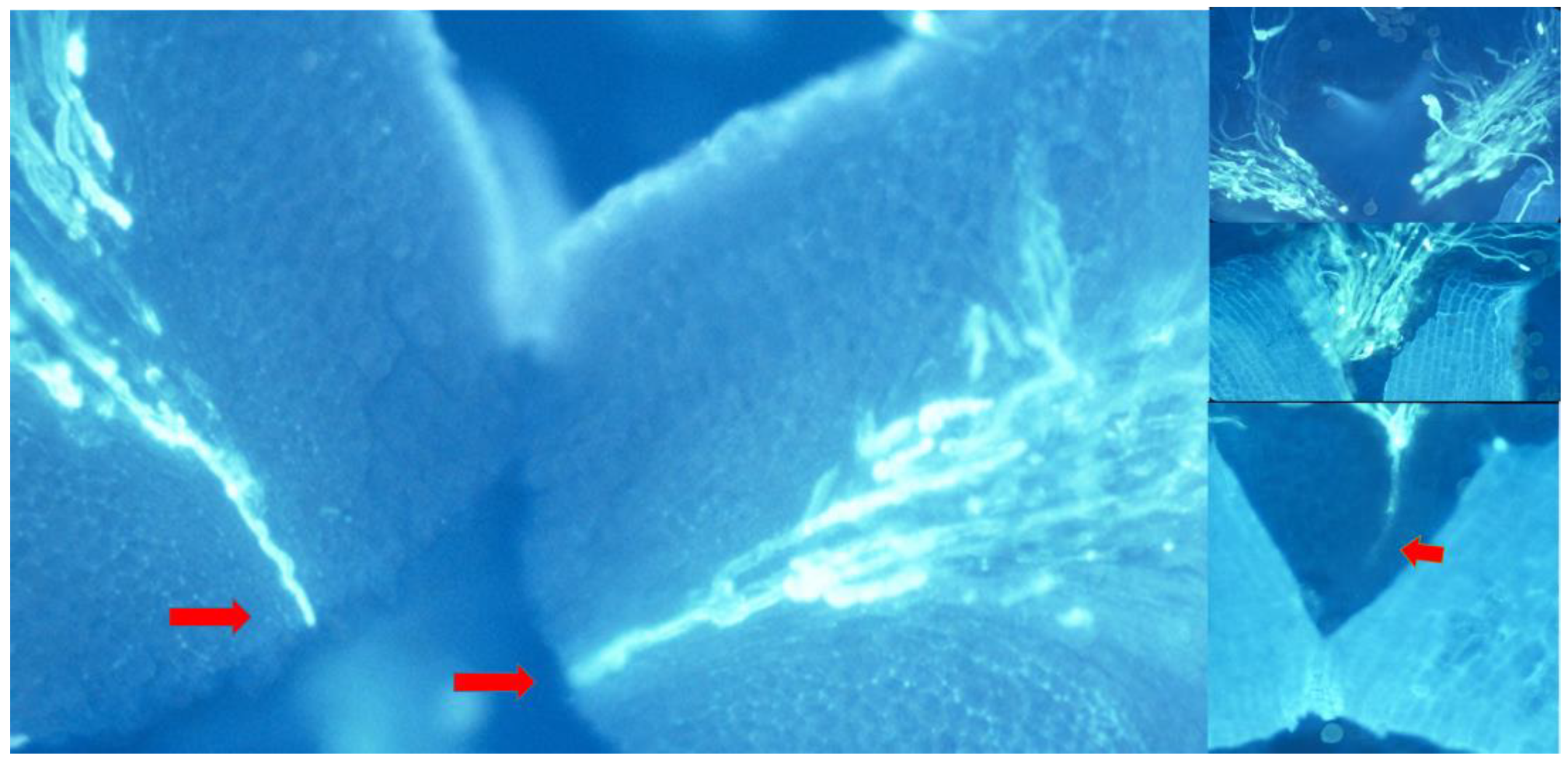
Figure 6.
Composition of pollen tube attrition in cross-pollinated olive flowers. Left: The flower has been dissected, softened with NaOH 1N, squashed and stained with aniline blue and then observed under fluorescence microscopy. Right: Different pistil sections showing progressive pollen tube attrition. Up: stigma; down: style. Red arrows showing winner pollen tubes.
Figure 6.
Composition of pollen tube attrition in cross-pollinated olive flowers. Left: The flower has been dissected, softened with NaOH 1N, squashed and stained with aniline blue and then observed under fluorescence microscopy. Right: Different pistil sections showing progressive pollen tube attrition. Up: stigma; down: style. Red arrows showing winner pollen tubes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



