
Submitted:
18 November 2024
Posted:
19 November 2024
You are already at the latest version
Abstract
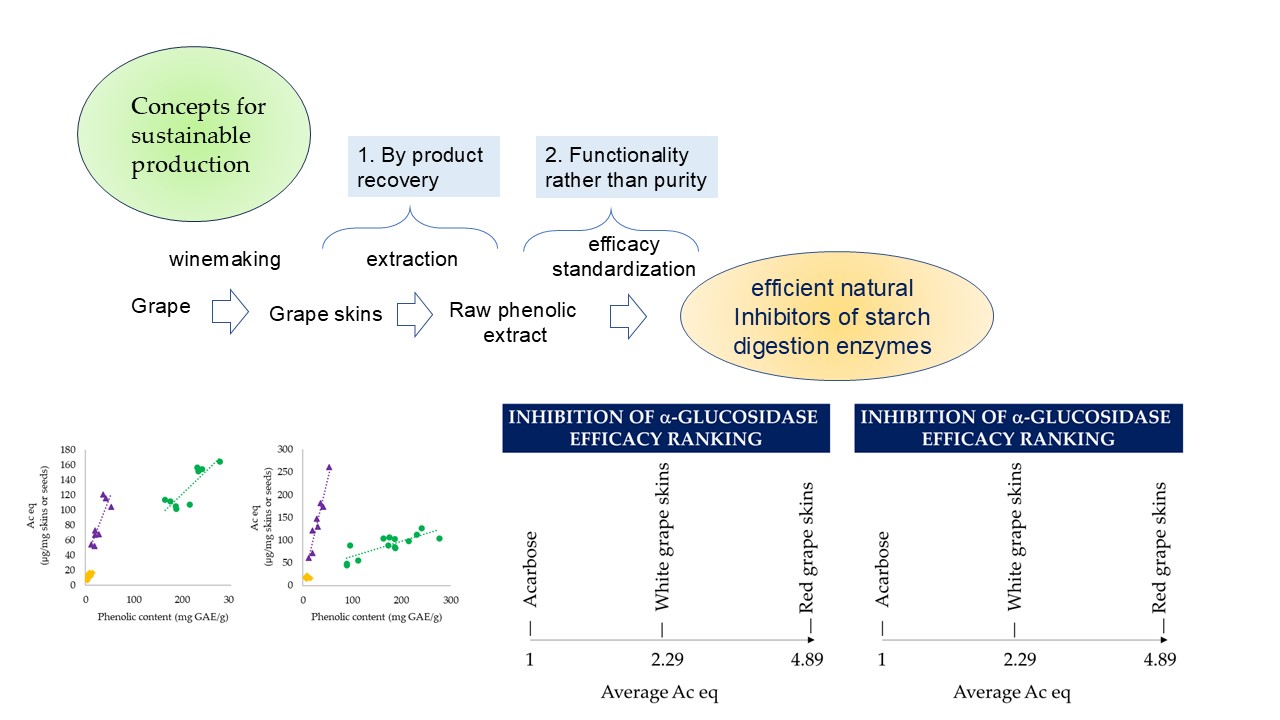
The increase in the incidence of hyperglycemia and diabetes poses the challenge to search cost-effective natural inhibitors of starch digestion enzymes. Among natural compounds, phenolics have been considered as promising candidates. The aims of this study were: a) to investigate if winemaking byproducts are effective as inhibitors of the intestinal brush border alpha-glucosidase and the pancreatic alpha-amylase in vitro; b) to calculate an efficacy index for their phenolic pool, despite their variability due to grape cultivar and winemaking process; c) to compare winemaking byproducts’ efficacy with those of other plants. Winemaking by-products were characterized for the contents of total phenolics, flavanols, flavonols and anthocyanins, as well as the inhibitory activity towards the intestinal brush border alpha-glucosidase and pancreatic alpha-amylase. The inhibition towards alpha-glucosidase resulted to be on average 2.29 and 4.89 µg acarbose equivalents/µg total phenolics (µg Ac eq/µg GAE) for white and red grape skins, respec-tively. The inhibition towards alpha-amylase was 0.5 and 1.6 µg Ac eq/µg GAE for white and red grape skins, respectively. The inhibitory activity of the phenolic pool of red grape skins was higher than those of isolated phenolic compounds, probably due to synergistic effects among compounds. Hence, bioactive phenolic fractions could be produced with the focus on functionality rather than purity, in line with the principles of sustainable processing. A correlation was observed between the total phenolic contents of red grape skins and their inhibitory effectiveness, which is useful to standardize the efficacy of phenolic extracts. Compared to other plants reported in the literature red grape skins resulted to be cost-effective sources of starch digestion enzyme inhibitors.
Keywords:
alpha-glucosidase
; alpha-amylase
; grape
; phenolic
; acarbose
1. Introduction
The ability of phenolics as inhibitors of starch digestion enzymes such as α-amylase and α-glucosidase, has been the focus of different investigations, which have also been carefully recompiled by review studies [1]. In fact, the pharmacological approach to treat type-2 diabetes consists of inhibiting starch digestion enzymes by synthetic drugs, especially acarbose, for which however, several side effects are described [2]. Hence, it would be advisable to find efficient natural inhibitors. Among the different potential sources of natural inhibitors, grape and grape-derived products have emerged as the most promising [3,4]. In particular, a large part of grape bioactive compounds, including anthocyanins, flavonols, monomeric, dimeric and oligomeric flavanols remains in grape pomace (skins and seeds) after the winemaking process, mainly due to their hydrophobicity and hydrogen bonds [3]. The first report on the potential anti-glycemic effect of grape pomace proved that red grape pomace (Cabernet Franc variety) extract suppresses postprandial hyperglycemia in diabetic mice following a potato starch challenge, and the effect was attributed to α-glucosidase inhibition [5]. In a further study, red grape skin extract (Norton variety) was found to significantly inhibit mammalian intestinal α-glucosidases in the streptozocin-treated male mice following starch challenge, but it was not effective on other digestive enzymes including pancreatic α-amylase [6]. A subsequent study evaluated in vivo inhibition of α-amylase and α-glucosidase by running starch and maltose tolerance tests in rats with or without administration of a red grape pomace (Merlot variety) extract [7]. The positive effect observed was attributed to the inhibition of both pancreatic and salivary α-amylases, while no inhibition on α-glucosidase was detected [7]. A pilot study performed on eight human subjects showed that postprandial plasma glucose concentrations after ingestion of high carbohydrate meal with grape seed extract were significantly lower than that of the control [8].
To get insights about the potential of grape phenolics as inhibitors of starch digestion enzymes, isolated compounds were assessed in vitro towards α-glucosidase [9,10,11,12,13] and α-amylase [11,15,16,17]. Moreover, the combination of procyanidins with anthocyanins improved starch digestion enzymes inhibition, likely due to synergistic effects [9]. Based on this latter finding, it is important to consider the raw phenolic extract as a pool, with potentially higher effectiveness than the isolated components. This latter approach is also in line with the principle of sustainable food production, since food products/ingredients can become more sustainable in case they are produced with the focus on functionality rather than purity [23]. Moreover, the possibilities to upgrade food by-products is another principle of sustainable food production [23].
Hence, the aim of this manuscript was to investigate the effectiveness of different winemaking byproducts, which are notably rich in phenolic compounds, as inhibitors of starch digestion enzymes.
2. Materials and Methods
2.1. Chemicals
Standard phenolics, namely, catechin, epicatechin, procyanidin B1, procyanidin B2, quercetin 3-O glucoside, quercetin, kaempferol, delphinidin 3-O-glucoside, cyanidin 3-O-glucoside, petunidin 3-O-glucoside; peonidin 3-O-glucoside; malvidin 3-O-glucoside, rat intestinal acetone powder (N1377-5G), p-nitrophenyl α-D-glucoside (p-NPG), acarbose, porcine pancreatic α-amylase, type VI-B (A3176), p-nitrophenyl α-D-maltopentaoside (p-NPGP) and all other chemicals were from Sigma Aldrich (Milan, Italy).
2.2. Grape Pomace
Pomace samples of the red grape varieties Barbera (BA), Croatina (CR), Freisa (FR), Dolcetto (DO 01 and DO 02, corresponding to the same variety processed by two different wineries), Grignolino (GR), Neretto (NR) and Nebbiolo (NE) and of the white grape varieties Arneis (AR), Moscato (MO), Muller Thurgau (MT), Erbaluce (ER), Chardonnay (CH), Nascetta (NA) e Riesling (RI), as well as a heterogeneous batch of red grape pomace, were kindly provided by wineries located in Northern Italy. At the winery, the pomace was sieved (with a 5 mm sieve) to separate the skins from the seeds and frozen to inhibit microbial growth. Grape skins and seeds were transported frozen to the lab and dried at 50 °C for about 8 h using a Ignis model AKS201/IX/01 ventilated oven (Whirpool, Milan, Italy). Dried grape skins were then milled and sieved by using the Octagon Digital sieve shaker (Endecotts Ltd., London, UK), with certified sieves to obtain the fraction with particle sizes in the range 250–500 µm.
2.3. Phenolic Extraction
For sample comparison, about 100 mg of grape skin powder was extracted with 8 mL methanol:water:HCl (80:20:0.1, v/v/v), for 2 h at room temperature with continuous stirring. The mixture was centrifuged at 10,000g for 10 min, the supernatant was recovered and the solid residue was re-extracted using 6 mL of the same solvent twice. The three supernatants were pooled, dried under vacuum at 35 °C and the residue was suspended in 10 mL of methanol:water:HCl (80:20:0.1, v/v/v) for further analysis. Red grape skins separated from the heterogeneous pomace batch were extracted according to a food-grade protocol, by 60% aqueous ethanol with continuous stirring for 2 h at 60 °C [24].
2.4. Total Phenolics
For the Folin–Ciocalteu assay the reaction mixture contained 6.0 mL of distilled water, 0.5 mL of the extracts, 0.5 mL of Folin–Ciocalteu reagent and 3 mL of 10% Na2CO3. The mixtures were incubated for 90 min at room temperature and then the absorbance was recorded at 760 nm against a blank with no extract addition. For each extract, 2–4 dilutions were assessed in duplicate. A calibration curve was built using gallic acid. Total phenolics were expressed as milligrams of gallic acid equivalents (GAE) per gram of dry product or milligrams of gallic acid equivalents (GAE) per liter of extract.
2.5. Oligomeric Procyanidins
Oligomeric procyanidin content was analysed as described previously [25]. Briefly, 1 mL of the extract was added to 6 mL of n-butanol/HCl (95:5, v/v) and 0.2 mL of 2% NH4Fe(SO4)2·12 H2O in 2 M HCl. Hydrolysis was carried out at 95 °C for 40 min. The reaction mixtures were cooled and the absorbance was measured at 550 nm against a blank made as for the sample but incubated at room temperature. For each extract, 2–4 dilutions were assessed in duplicate. Oligomeric procyanidin amount was expressed as milligrams per liter of extract, using 1.79 as a conversion factor [25].
2.6. Phenolic Characterization by HPLC
The phenolic contents were analyzed in duplicate as described previously [26], using a model Shimadzu LC-20 AD pump coupled to a model Shimadzu SPD-M20A photodiode array detector (DAD) and an RF-20 AXS operated by Labsolution Software 5.5 Shimadzu, Kyoto, Japan). A 2.6 µm Kinetex C18 column (150 × 4.6 mm; Phenomenex, Bologna, Italy) was used for the separation at a flow-rate of 1.5 mL/min. The column was maintained at 40 °C. The separation was performed by means of a linear gradient elution. Eluents were: (A) 0.1% H3PO4; (B) acetonitrile. The gradient was as follows: from 6% B to 20% B in 18 min; from 20% B to 60% B in 7 min; from 60% B to 90% B in 19 min; 90% B for 10 min and then 6% B for 5 min. DAD analysis was carried out in the range of 200–600 nm. Quantification of phenolics was performed by calibration curves built with cyanidin 3-O-glucoside with the DAD set at 520 nm for anthocyanins, quercetin 3-O-glucoside with the DAD set at 354 nm for flavonols and catechin with the fluorimetric detector set at λex 230 and λem 320. Results were expressed as milligrams of phenolic compound per liter of extract.
2.7. In Vitro α-Glucosidase and α-Amylase Inhibition Assay
A crude α-glucosidase solution was prepared from rat intestinal acetone powder. Briefly, 200 mg of rat intestinal acetone powder was dissolved in 4 mL of 50 mM ice cold phosphate buffer (pH 6.8) and sonicated for 15 min at 4 °C. The suspension was vortexed for 20 min and then centrifuged at 10,000 g at 4 °C for 30 min. The resulting supernatant was used for the assay. For the α-glucosidase activity assay, 650 µL of 50 mM phosphate buffer, pH 6.8, 100 µL of the enzyme solution and 50 µL of grape skin extract were added in Eppendorf tubes and pre-incubated for 5 min at 37 °C. Then, 200 µL of 1 mM pNPG was added as substrate and the mixture was further incubated at 37 °C for 25 min. For the pancreatic α-amylase assay, 550 µL of 50 mM phosphate buffer, pH 6.8, 200 µL of the enzyme solution (10 μM in the same buffer) and 50 µL of grape skin extract were added in Eppendorf tubes, pre-incubated for 5 min at 37 °C. Then, 200 µL of 1 mM pNPGP was added to the tubes as the substrate and the mixture was further incubated at 37 °C for 55 min. For both enzymatic reactions, the assay mixture was centrifuged at 10,000 g for 3 min and the absorbance of the clear supernatant was recorded at 405 nm. The control was run by adding the extraction solvent replacing the sample. A sample blank and a control blank were run without addition of substrate and without addition of both substrate and sample, respectively [27]. Acarbose was used as a reference inhibitor for both enzymatic reactions. Dose–response curves were made for samples and acarbose. The I50 values i.e. concentration that inhibited the reaction by 50% were expressed as milligram of skins per milliliter and microgram of acarbose equivalents (Ac eq) per milligram of skins. Moreover, the ratio between I50 of acarbose (µg/ml) and I50 of samples (µg GAE/ml) was calculated to assess the efficacy of the phenolic pool [27].
2.8. Statistical Analysis of Data
Experimental data were analyzed by one-way ANOVA using the least significant difference (LSD) as a multiple range test, and by non-linear regression analyses using Statgraphics 5.1 (STCC Inc.; Rockville, MD, USA). Results are reported as average ± standard deviation (SD).
3. Results
3.1. α-glucosidase and α-amylase inhibition
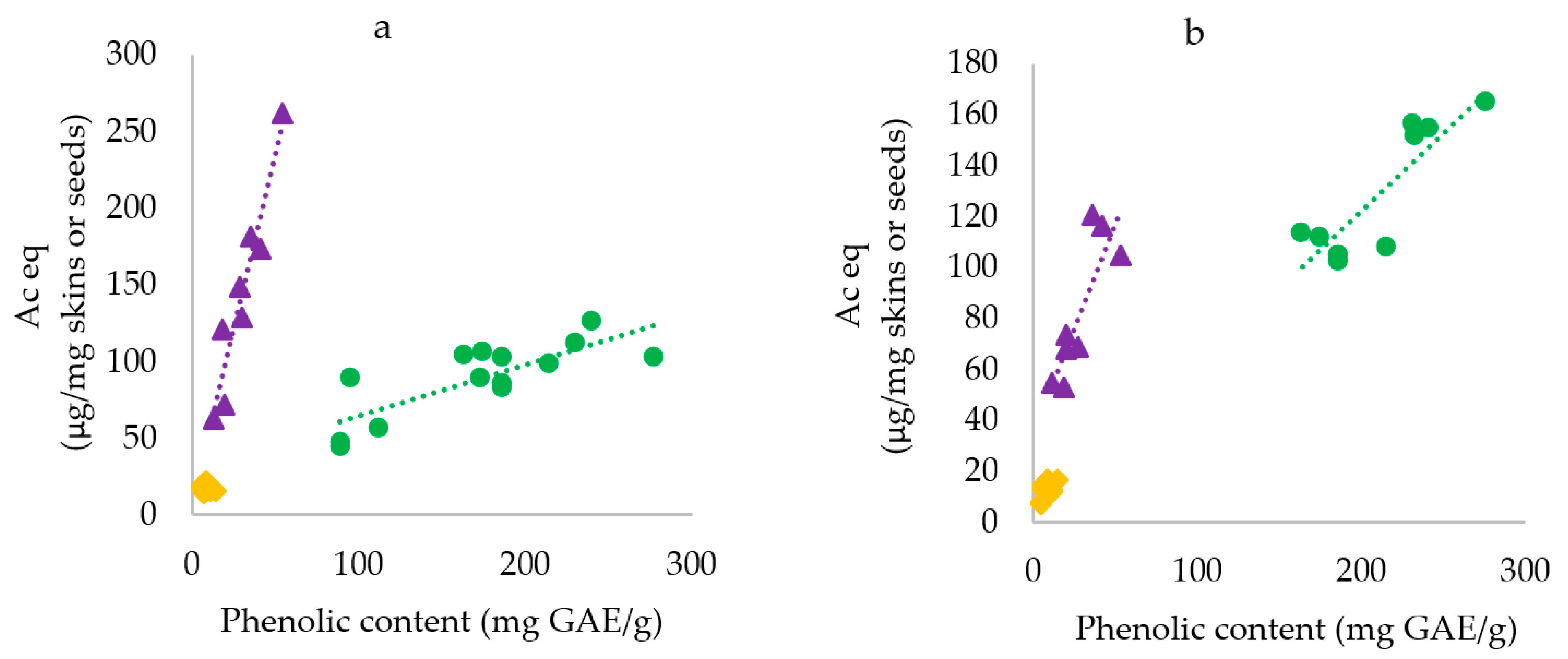
Grape skins phenolics were extracted from eight red grape skins and seven white grape skins batches recovered from winemaking by-products and the inhibitory activity of the extracts toward starch digestion enzymes was assessed (Table 1). The correlation between total phenolic content of the extracts and starch digestion enzymes inhibition was then investigated (Figure 1a,b).
3.1.1. α-Glucosidase Inhibition
The I50 values for α-glucosidase inhibition ranged between 0.38 and 1.66 mg skins/mL, corresponding to 261 – 62 µg Ac eq/mg skins for red grape skins and between 4.62 and 6.45 mg skins/mL corresponding to 21.6 and 15.5 µg Ac eq/mg skins (Table 1). A previous study has shown that fourteen batches of grape seeds had I50 values ranging from 0.80 to 2.37 mg seeds/mL, corresponding to 126 - 44 µg Ac eq/mg seeds [27].
Considering red grape skins, the inhibition effectiveness toward α-glucosidase was correlated to procyanidin content (R = 0.922, p < 0.05) and anthocyanin content (R = 0.893, p < 0.05), while no correlation was found with flavonol content (not shown). However, the best correlation was observed between α-glucosidase and total phenolic content (R = 0.966, p < 0.05, Figure 1a). Hence, on average, the inhibitory activity of red grape skins was 4.89 µg Ac eq/µg GAE.
Among white grape skins, the inhibitory activity toward α-glucosidase was similar and lower than for red grape skins. No significant correlation was observed between the inhibitory activity and the contents of procyanidins, flavonols and total phenolic. An average value of 2.29 µg Ac eq/µg GAE was observed. As shown previously, the inhibitory activity of grape seeds toward α-glucosidase was correlated to the total phenolic content (R = 0.8033, p < 0.05), with an average value of 0.33 µg Ac eq/µg GAE [27]. Hence, the phenolic pool of red grape skins was 10-fold more efficient than that of grape seeds.
3.1.2. α-Amylase Inhibition
Considering α-amylase inhibition, the I50 values of red grape skins ranged between 0.24 and 0.57 mg skins/mL, corresponding to 118 - 52 µg Ac eq/mg skins. For white grape skins, the I50 values ranged between 1.77 and 2.44 mg skins/mL, corresponding to 16.9 - 12.4 µg Ac eq/mg skins (Table 1). In a previous study, fourteen batches of grape seeds were found to have I50 values ranging from 0.19 to 0.31 mg seeds/mL, corresponding to 164 - 98 µg Ac eq/mg seeds [27].
For red grape skins a significant correlation was observed between the inhibition effectiveness toward α-amylase and phenolic content (R = 0.836, p < 0.05, Figure 1b), as well as anthocyanin content (R = 0.760, p < 0.05) and procyanidin content (R = 0.755, p < 0.05) but no correlation was observed with flavonol content. From the slope of the correlation curve between enzyme inhibition and total phenolic content, it can be derived that red grape skins can provide an average of 1.6 µg Ac eq/µg GAE.
White grape skins inhibitory activity was slightly correlated to the content of total phenolics (R = 0.57, p < 0.05) and procyanidins (R = 0.72, p < 0.01) but no correlation was observed with the contents of flavonols. On average, white grape skins phenolic provided 0.50 µg Ac eq/µg GAE.
The phenolic pool of red grape skins was generally a more efficient inhibitor that that of grape seeds, which had 0.61 µg Ac eq/µg GAE [27]. However, grape seeds generally have higher phenolic content than grape skins and hence they result in higher acarbose equivalents per mass unit.
Figure 1.
Correlation between enzyme inhibition efficacy and phenolic content for red grape skins (▲), white grape skins (♦) and grape seeds (●). a) α-glucosidase inhibition; b) α-amylase inhibition. Data for grape seeds were recalculated from Lavelli et al., 2015 [27].
Figure 1.
Correlation between enzyme inhibition efficacy and phenolic content for red grape skins (▲), white grape skins (♦) and grape seeds (●). a) α-glucosidase inhibition; b) α-amylase inhibition. Data for grape seeds were recalculated from Lavelli et al., 2015 [27].
3.2. Characterization of Food-Grade Extracts of Red Grape Skins
The phenolics fraction of a heterogeneous batch (mixture of varieties) of red grape skins was then recovered by a procedure that yields a food-grade extract with a good stability, showing its potential to be considered for food applications [24]. In fact, since the grape pomace is a byproduct, it is generally collected as a whole, without any separation according to grape varieties or winemaking process. As expected, the phenolic profile, included anthocyanins, flavonols, monomeric, dimeric and oligomeric procyanidins (Table 2). The inhibitory activity towards starch digestion enzymes was then determined and found to be 5.2 and 2.0 µg Ac eq/µg GAE for α-glucosidase and α-amylase, respectively (Table 3 and Table 4). These values are close to those expected from the correlations between total phenolic content and inhibitory efficacy for α-glucosidase (4.89 µg Ac eq/µg GAE) and α-amylase (1.6 Ac eq/µg GAE), suggesting that the correlation found could be used to predict the enzyme inhibition effectiveness based on the phenolic content.
4. Discussion
Previous studies have found that natural compounds had higher inhibitory activity towards yeast α-glucosidase than rat intestinal α-glucosidase [28,29]. Hence, the inhibitory activity of winemaking by products was compared to those of standard compounds and plant extracts assessed using rat intestinal α-glucosidase. Moreover, the comparison was made only when acarbose was used as a positive control and the activity could be measured as acarbose equivalents, in order to minimize possible differences due to the assay conditions (Table 3).

Table 3.
Effectiveness of standard phenolics (µg Ac eq/µg), red grape skin extract and different plant extracts (µg Ac eq/µg GAE) as rat intestinal α-glucosidase inhibitors.
Table 3.
Effectiveness of standard phenolics (µg Ac eq/µg), red grape skin extract and different plant extracts (µg Ac eq/µg GAE) as rat intestinal α-glucosidase inhibitors.
| Compound/plant | Solvent | µg Ac eq/µg | Ref. |
|---|---|---|---|
| catechin | < 0.047 | [13] | |
| epicatechin | < 0.047 | [13] | |
| procyanidin B1 | < 0.047 | [13] | |
| malvidin 3-O-glucoside | 3.99 | [10] | |
| cyanidin 3-O-glucoside | 0.26, 0.46, 0.52 | [9,10,11] | |
| delphinidin 3-O-glucoside | 3.46 | [10] | |
| petunidin 3-O-glucoside | 3.12 | [10] | |
| quercetin | 0.22, 0.09 | [12,13] | |
| quercetin 3-O-glucoside | 0.28 | [14] | |
| kaempferol | 0.093 | [13] | |
| kaempferol 3-O-glucoside | 0.12, 0.16 | [13,14] | |
| Blackcurrant (Ribes nigrum) | fruit, 0.2% formic acid in water | 2.0 | [9] |
| Rowanberry (Sorbus aucuparia) | fruit, 0.2% formic acid in water | 1.3 | [9] |
| Raspberry (Rubus idaeus) | fruit, 0.2% formic acid in water | < 0.2 | [9] |
| Cloudberry (Rubus chamaemorus) | fruit, 0.2% formic acid in water | < 0.2 | [9] |
| Blackberry (Rubus grandifolius) | fruit, 80% methanol, 7% acetic acid | 0.4 | [11] |
| Blackberry (Rubus grandifolius) | fruit, 80% methanol, 7% acetic acid | 0.4 | [11] |
| Blackberry (Rubus grandifolius) | leaves, 80% methanol, 7% acetic acid | 0.5 | [11] |
| Blackberry (Rubus grandifolius) | leaves, 80% methanol, 7% acetic acid | 0.6 | [11] |
| Red cabbage (Brassica oleracea) | edible part, 70% methanol | 5.8 | [30] |
| Red cabbage (Brassica oleracea) | edible part, 70% methanol | 8.3 | [30] |
| Red cabbage (Brassica oleracea) | edible part, 70% methanol | 5.4 | [30] |
| Red cabbage (Brassica oleracea) | edible part, 70% methanol | 6.6 | [30] |
| Red cabbage (Brassica oleracea) | edible part, 70% methanol | 4.3 | [30] |
| Coffee (Coffea arabica) | ground roasted beans, water | 1.7 | [31] |
| Mushroom (Grifola frondose) | fruiting body, 95% ethanol | 4.5 | [29] |
| Grape (Vitis vinifera) | red skins, 60% ethanol | 5.2 ± 0.1 | this study |
The main grape anthocyanin, i.e. malvidin 3-O-glucoside was found to be a more effective inhibitor than acarbose (3.99 µg Ac eq/µg) [10]; similarly, the other grape anthocyanins are more efficient than acarbose [10], except for cyanidin 3-O-glucoside [9,10,11]. Flavonols are generally 10-fold less efficient than acarbose [12,14], except for quercetin glucoside [14]. Catechin, epicatechin and procyanidin B1 were found to be 20 times less efficient than acarbose [13]. For cyanidin 3-O-glucoside, quercetin and kaempferol 3-O-glucoside, the observed effectiveness was not the same for the different studies. This result could be due to the different assay conditions used.
Molecular docking studies showed that anthocyanins can enter in depth into the active pocket of α-glucosidase and form hydrophobic interactions with Glu276, Asp214, Ala278, Tyr71, Asp349 residues; besides, their glycosyl groups can interact with Asp408 via hydrogen bonds [10]. Overall, the phenolic pool of red grape skins observed in the current study was found to be more efficient than the purified compounds, confirming the importance of the synergistic effects among antioxidants as previously observed [9]. The inhibitory activity towards rat intestinal α-glucosidase was also investigated for different berries (Table 3), including blackcurrant, rowanberry, raspberry, cloudberry, and blackberry [9,11]. In all these fruits the inhibitory activity was lower than that found in the red grape skins. Red cabbage was found to be an efficient source of rat intestinal α-glucosidase inhibitors, comparable to that of red grape skins, with a variety-dependent effect [30]. The high inhibitory activity of red cabbage was attributed to the content of anthocyanins, especially the diacylated forms, which could act synergistically with hydroxycinnamic acids [30]. Among potential rat intestinal α-glucosidase inhibitors, roasted ground coffee was one anthocyanin-free source, less efficient than red grape skins, whose activity was tentatively attributed not only to phenolics, but to the Amadori compounds and melanoidins [31]. Another anthocyanin-free source which is receiving attention is the fruiting body of mushrooms [29]. Indeed, an inhibitory activity comparable to that found in grape skins was observed in mushroom, which was attributed to pyrrole alkaloids [29].
Regarding α-amylase inhibition by standard compounds and plant extracts, only studies in which porcine α-amylase was used and acarbose was included as a positive control were considered (Table 4).
Table 4.
Effectiveness of standard phenolics (µg Ac eq/µg), red grape skin extract and different plant extracts (µg Ac eq/µg GAE) as porcine pancreatic α-amylase inhibitors1.
Table 4.
Effectiveness of standard phenolics (µg Ac eq/µg), red grape skin extract and different plant extracts (µg Ac eq/µg GAE) as porcine pancreatic α-amylase inhibitors1.
| Compound/plant | Solvent | µg Ac eq/µg | Ref. |
|---|---|---|---|
| catechin | 0.0021, 0.043 | [15,16] | |
| epicatechin | nd | [15,16], [17] | |
| cyanidin 3-O-glucoside | 0.020, 0.044 | [11,18] | |
| quercetin | < 0.027 | [19] | |
| kaempferol | < 0.028 | [19] | |
| PA, mDP not specified | 6.1, 1.2 | [20,21] | |
| PA mDP 1.35 %G 5.43) | 0.28 | [22] | |
| PA mDP 6.97 %G 13.27) | 6.77 | [22] | |
| Rowanberry (Sorbus aucuparia) | fruit, 0.2% formic acid in water | 0.2 | [9] |
| Hibiscus (Hibiscus deflersii) | aerial parts, water | 0.6 | [32] |
| Hibiscus (Hibiscus calyphyllus) | aerial parts, water | 0.3 | [32] |
| Hibiscus (Hibiscus micranthus) | aerial parts, water | 0.2 | [32] |
| Dogwoods (Cornus mas) | fruits, 60% ethanol extract | 0.8 | [33] |
| Dogwoods (Cornus alba) | fruits, 60% ethanol extract | 0.7 | [33] |
| Elderberry (Sambucus nigra) | edible part, 50% ethanol | 8.34 | [35] |
| Saffron (Crocus sativus) | stigma, water extract | 1.98 | [34] |
| Saffron (Crocus sativus) | tepal, water extract | 2.32 | [34] |
| Coffee (Coffea arabica) | ground roasted beans, water | 1.49 | [31] |
| Red pepper (Capsicum annuum) | edible part, 70% ethanol | 8.42 | [36] |
| Grape (Vitis vinifera) | red skins, 60% ethanol | 2.0 ± 0.1 | this study |
1 mDP, mean polymerization degree.
Very low inhibitory activity was observed for anthocyanins [11,17,18], flavonols [19] and monomeric flavanols [15,16,17]. Conversely, grape seed procyanidins resulted to be more effective than acarbose (6.1 µg Ac eq/µg) [20]. However, in a further study, lower effectiveness was observed for procyanidins with respect to acarbose (0.793 µg Ac eq/µg) [16]. In another approach, red grape phenolics were fractionated and the efficacy of procyanidin fraction resulted to be higher (1.19 µg Ac/µg) with respect to those of the anthocyanin fraction (0.74 µg Ac eq/µg) [21]. The difference in enzyme inhibition effectiveness of procyanidins could be due to different procedures for purification, resulting in different molecular weight of the fractionated compounds. Indeed, the procyanidin fraction with mean polymerization degree of 1.35 resulted to have 0.28 µg Ac eq/µg, while that with mean polymerization degree of 6.97 showed 6.77 µg Ac eq/µg [22]. Molecular docking studies demonstrated that grape procyanidins can interact with the active site of α-amylase through hydrogen interactions, hydrophobic and electrostatic interactions, which are stronger for procyanidin tetramer than for the trimer and the dimer [22]. In fact, in the catalytic core of α-amylase, seven hydrogen bonds can be formed between procyanidins tetramer and the enzyme; in the proposed binding mode procyanidins tetramer binds to the enzyme via π-π stacking contact with Trp59, His201 and Tyr151, respectively, Asp300 and Glu233 have hydrophobic effects on tetramer.
Overall, the phenolic pool of red grape skins was more efficient than acarbose as inhibitor of amylase. However, higher efficacy could be obtained upon purification of the procyanidin faction. Various plants considered as sources of α-amylase inhibitors, such as rowanberry, hibiscus and dogwoods, displayed lower effectiveness with respect to red grape skins, while coffee and saffron displayed a comparable effectiveness [9,31,32,33,34] (Table 4). On the other hand, elderberry was found to be more effective than red grape skins, most probably due procyanidins [35]. Moreover, red sweet pepper has been highlighted as a possible potent α-amylase inhibitor, more efficient than red grape skins, due to its content in carvacrol [36].
5. Conclusions
All winemaking byproducts, including red and white grape skins and grape seeds showed excellent ability to inhibit starch digestion enzymes, compared to the drug acarbose, with the highest effectiveness found for red grape skins phenolics. Comparing different grape varieties, a correlation was observed between total phenolic contents (GAE) and acarbose equivalents for both α-glucosidase and α-amylase inhibition. Total phenolic content in grape skins varies markedly, depending on the cultivar, climate, location of the vineyard and winemaking procedure. Therefore, the observed correlation provides a useful tool to design the inhibitory efficacy of red grape skins extracts based on the standardization of their total phenolic content, thus facilitating the scaling-up this application.
Regarding the inhibition of α-glucosidase, the efficacy of the phenolic pool of red grape skins was found to be on average 4.89 µg Ac eq/µg GAE, which was higher than that of isolated phenolic compounds, probably due to synergistic effects among compounds. On the other hand, the effectiveness of inhibition of α-amylase was 1.6 µg Ac eq/µg GAE while isolation of procyanidin fraction could achieve higher activity.
Compared to the phenolic pools of other plants, the inhibitory activity of red grape skins was generally higher, especially towards α-glucosidase. However, various plant extracts considered in the literature as sources of starch digestion enzymes’ inhibitors were not included in the comparison because of the lack of specification of the enzymes source used in the assay or the use of yeast α-glucosidase instead of rat intestinal α-glucosidase, or the absence of an internal control (acarbose). Moreover, different varieties of a potential plant sources should be analysed for the inhibitory efficacy, to consider the impact of biodiversity.
Despite these limitations, the role of red grape skins as a potential source of inhibitors of starch digestion enzymes resulted to be promising. Additionally, the use of unpurified fractions instead of isolated compounds and the opportunity to upgrade food by-products are in line with the principles of sustainable food processing.
Author Contributions
Conceptualization, P.S.C.S.H. and V.L.; methodology, P.S.C.S.H. and V.L.; formal analysis, P.S.C.S.H. and V.L; investigation, P.S.C.S.H. and V.L; data curation, P.S.C.S.H. and V.L; writing—original draft preparation, V.L.; writing—review and editing, P.S.C.S.H. and V.L.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: a review. Crit. Rev. Food Sci. Nutr. 2020, 60, 4, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Rosak, C.; Mertes, G. Critical evaluation of the role of acarbose in the treatment of diabetes: Patient considerations. Diabetes Metab. Syndr. Obes. 2012, 5, 357. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Yupanqui, M.; Lante, A.; Mihaylova, D.; Krastanov, A.I.; Rizzi, C. The α-amylase and α-glucosidase inhibition capacity of grape pomace: A review. Food Bioprocess Technol. 2022, 16, 691–703. [Google Scholar] [CrossRef]
- Taladrid, D.; Rebollo-Hernanz, M.; Martin-Cabrejas, M.A.; Moreno-Arribas, M.V.; Bartolomé, B. Grape pomace as a cardiometabolic health-promoting ingredient: activity in the intestinal environment. Antioxidants 2023, 12, 979. [Google Scholar] [CrossRef]
- Hogan, S.; Zhang, L.; Li, J.; Sun, S.; Canning, C.; Zhou, K. Antioxidant rich grape pomace extract suppresses postprandial hyperglycemia in diabetic mice by specifically inhibiting α-glucosidase. Nutr. Metab., 2010, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hogan, S.; Li, J.; Sund, S.; Canning, C.; Zheng, S.J.; Zhou, Z. Grape skin extract inhibits mammalian intestinal α-glucosidase activity and suppresses postprandial glycemic response in streptozocin-treated mice. Food Chem. 2011, 126, 466–471. [Google Scholar] [CrossRef]
- Kato-Schwartz, C.G.; Corrêa, R.C.G.; de Souza Lima, D.; de Sá-Nakanishi, A.B.; de Almeida Gonçalves, G.; Seixas, F.A.V.; Haminiuk, C.W.I.; Barros, L.; Ferreira, I.C.F.R.; Bracht, A.; et al. Potential anti-diabetic properties of Merlot grape pomace extract: An in vitro, in silico and in vivo study of-amylase and-glucosidase inhibition. Food Res. Int. 2020, 137, 109462. [Google Scholar] [CrossRef] [PubMed]
- Sapwarobol, S.; Adisakwattana, S.; Changpeng, S.; Ratanawachirin, W.; Tanruttanawong,K.; Boonyarit, W. Postprandial blood glucose response to grape seed extract in healthy participants: A pilot study. Pharmacogn. Mag. 2012, 8, 31, 192–196. [CrossRef]
- Boath, A. S.; Stewart, D.; McDougal, G. J. Berry components inhibit α-glucosidase in vitro: Synergies between acarbose and polyphenols from black currant and rowanberry. Food Chem. 2012, 135, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xi, L.; Liu, Y.; Chen, W. Pelargonidin-3-O-rutinoside as a novel α-glucosidase inhibitor for improving postprandial hyperglycemia. Chem. Commun. 2019, 55, 39. [Google Scholar] [CrossRef] [PubMed]
- Spinola, V.; Pinto, J.; Llorent-Martínez, E. J.; Tomás, H.; Castilho, P. C. Evaluation of Rubus grandifolius L. (wild blackberries) activities targeting management of type-2 diabetes and obesity. Food Chem. Toxicol, 2019; 123, 443–452. [Google Scholar] [CrossRef]
- Jo, S.-H.; Ha, K.-S.; Moon, K.-S.; Lee, O.-H.; Jang, H.-D.; Kwon, Y.-I. In vitro and in vivo anti-hyperglycemic effects of Omija (Schizandra chinensis) Fruit. Int. J. Mol. Sci. 2011, 12(2), 1359–1370. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Ramdath, D. D.; Tang, Y.; Chen, P. X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on a-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Sari, S.; Barut, B.; Özel, A. Discovery of potent α-glucosidase inhibitor flavonols: Insights into mechanism of action through inhibition kinetics and docking simulations. Bioorg.Chem. 2018, 79, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kong, H.; Chitrakar, B.; Ban, X.; Gu, Z.; Hong, Y.; Cheng, L.; Li, Z.; Li, C. The substitution sites of hydroxyl and galloyl groups determine the inhibitory activity of human pancreatic α-amylase in twelve tea polyphenol monomers. Int. J. Biol. Macromol. 2024, 259, 129189. [Google Scholar] [CrossRef] [PubMed]
- Yilmazer-Musa, M. A. M.; Griffith, A. J.; Schneider, M. E.; Frei, B. Grape seed and tea extracts and catechin 3-gallates are potent inhibitors of a-amylase and a-glucosidase activity. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef]
- Kaeswurm, J. A. H., B. Claasen, M. P. Fischer, and M. Buchweitz. 2019. Interaction of structurally diverse phenolic compounds with porcine pancreatic a-amylase. J. Agric. Food Chem. 2019, 67, 11108–11118. [Google Scholar] [CrossRef] [PubMed]
- Ho, G. T.T.; Kase, E. T.; Wangensteen, H.; Barsett, H. Phenolic elderberry extracts, anthocyanins, procyanidins, and metabolites influence glucose and fatty acid uptake in human skeletal muscle cells J. Agric. Food Chem. 2017, 65, 2677–2685. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.; Sun, W.; Xing, Y.; Xiu, Z.; Zhuang, C.; Dong, Y. Dietary flavonoids and acarbose synergistically inhibit α-glucosidase and lower postprandial blood glucose. J. Agric. Food Chem. 2017, 65, 8319–8330. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Song, L.; Wang, H.; Huang, D.A. High-through put assay for quantification of starch hydrolase inhibition based on turbidity measurement. J. Agric. Food Chem. 2011; 59, 9756–9762. [Google Scholar]
- Ostberg-Potthoff, J.J.; Berger, K.; Richling, E.; Winterhalter, P. Activity-guided fractionation of red fruit extracts for the identification of compounds influencing glucose metabolism. Nutrients, 2019, 11(5), 1166. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Zhang, L.; Li, W.; Zhang, S.; Luo, L.; Wang, J.; Sun, B. In vitro evaluation of the anti-digestion and antioxidant effects of grape seed procyanidins according to their degrees of polymerization. J. Func. Foods 2018, 89, 85–95. [Google Scholar] [CrossRef]
- van der Goot, A. J.; Pelgrom, P. J. M.; Berghout, J. A. M.; Geerts, M. E. J.; Jankowiak, L.; Hardt, N. A.; Keijer, J.; Schutyser, M. A. I.; Nikiforidis, C. D.; Boom, R. M. Concepts for further sustainable production of foods. J. Food Eng. 2016, 168, 42–51. [Google Scholar] [CrossRef]
- Amendola, D.; De Faveri, D. M.; Spigno, G. Grape marc phenolics: Extraction kinetics, quality and stability of extracts. J. Food Eng. 2010, 97, 384–392. [Google Scholar] [CrossRef]
- Pedrali, D.; Barbarito, S.; Lavelli, V. Encapsulation of grape seed phenolics from winemaking byproducts in hydrogel microbeads – Impact of food matrix and processing on the inhibitory activity towards α-glucosidase. LWT - Food Sci. Technol. 2020, 133, 109952. [Google Scholar] [CrossRef]
- Bonassi, G.; Lavelli, V. Hydration and fortification of common bean (Phaseolus vulgaris L.) with grape skin phenolics—Effects of ultrasound application and heating. Antioxidants, 2024, 13, 615. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Sri Harsha, P. S. C.; Fiori, L. Screening grape seeds recovered from winemaking byproducts as sources of reducing agents and mammalian α-glucosidase and α-amylase inhibitors. Int. J. Food Sci. Technol. 2015, 50, 1182–1189. [Google Scholar] [CrossRef]
- Kim, G.-N.; Shin, J.-G.; Jang, H.-D. Antioxidant and antidiabetic activity of Dangyuja (Citrus grandis Osbeck) extract treated with Aspergillus saitoi. Food Chem. 2009, 117(1), 35–41. [Google Scholar] [CrossRef]
- Chen, S.; Yong, T.; Xiao, C.; Su, J.; Zhang, Y.; Jiao, C.; Xie, Y. Pyrrole alkaloids and ergosterols from Grifola frondosa exert anti-α-glucosidase and anti-proliferative activities. J. Func. Foods, 2018, 43, 196–205. [Google Scholar] [CrossRef]
- Podsędek, A.; Majewska, I.; Kucharska, A. Z. Inhibitory potential of red cabbage against digestive enzymes linked to obesity and type 2 diabetes. J. Agric. Food Chem. 2017, 65, 7192–7199. [Google Scholar] [CrossRef]
- Mitiku, H.; Kim, T. Y.; Kang, H.; Apostolidis, E.; Lee, J. Y.; Kwon, Y.I. Selected coffee (Coffea arabica L. ) extracts inhibit intestinal α-glucosidases activities in-vitro and postprandial hyperglycemia in SD rats BMC Complement. Med. Ther. 2022, 22, 249. [Google Scholar] [CrossRef]
- Al Yousef, H.M.; Hassan, W.H.B.; Abdelaziz, S.; Amina, M.; Adel, R.; El Sayed, M.A. UPLC-ESI-MS/MS Profile and antioxidant, cytotoxic, antidiabetic, and antiobesity activities of the aqueous extracts of three different Hibiscus species. J. Chem. 2020, 6749176. [Google Scholar] [CrossRef]
- Swierczewska, A.; Buchholz, T.; Melzig, M. F., Czerwinska, M. E. In vitro a-amylase and pancreatic lipase inhibitory activity of Cornus mas L. and Cornus alba L. fruit extracts. J. Food Drug Anal. 2019, 27, 1, 249–258. [CrossRef] [PubMed]
- Bellachioma, L.; Morresi, C.; Albacete, A.; Martínez-Melgarejo, P.A.; Ferretti, G.; Giorgini, G.; Galeazzi, R.; Damiani, E.; Bacchetti, T. Insights on the hypoglycemic potential of Crocus sativus Tepal polyphenols: An in vitro and in silico study. Int. J. Mol. Sci. 2023, 24, 9213. [Google Scholar] [CrossRef] [PubMed]
- Terzi, M.; Cavi, D.; Majki, T., Zengin, G.; Radojkovi, M. Could elderberry fruits processed by modern and conventional drying and extraction technology be considered a valuable source of health-promoting compounds? Food Chem. 2023, 405, 134766. [CrossRef] [PubMed]
- Di Sotto, A.; Vecchiato, M., Abete, L.; Toniolo, C.; Giusti, A. M. L. Mannina, Locatelli, M.; Nicoletti, M., Di Giacomo, S. Capsicum annuum L. var. Cornetto di Pontecorvo PDO: Polyphenolic profile and in vitro biological activities. J. Func. Foods, 2018, 40, 679-691. [CrossRef]
Table 1.
Efficacy of inhibition of rat intestinal α-glucosidase and porcine pancreatic α-amylase by phenolic extracts of grape skins recovered from winemaking byproducts.
Table 1.
Efficacy of inhibition of rat intestinal α-glucosidase and porcine pancreatic α-amylase by phenolic extracts of grape skins recovered from winemaking byproducts.
| Sample | α-Glucosidase Inhibition | α-Amylase Inhibition | ||
|---|---|---|---|---|
| I50 mg skins/mL | µg Ac eq/mg skins | I50 mg skins/mL | µg Ac eq/mg skins | |
| Red skins | ||||
| DO 01 | 0.77ab ± 0.03 | 130cd ± 7 | 0.24a ± 0.02 | 118e ± 3.9 |
| BA | 0.68ab ± 0.04 | 148de ± 12 | 0.44ab ± 0.01 | 68.3c ± 5.3 |
| FR | 0.38a ± 0.02 | 261f ± 21 | 0.29ab ± 0.00 | 105d ± 0.9 |
| CR | 0.58ab ± 0.03 | 173e ± 13 | 0.26a ± 0.03 | 116e ± 3.2 |
| NE | 1.39b ± 0.29 | 72c ± 8 | 0.42ab ± 0.02 | 70c ± 3.5 |
| NR | 1.66c ± 0.26 | 62b ± 13 | 0.55b ± 0.02 | 55b ± 1.9 |
| GR | 0.83ab ± 0.05 | 121cd ± 10 | 0.57b ± 0.00 | 52b ± 1.5 |
| DO 02 | 0.55ab ± 0.03 | 181e ± 10 | 0.25a ± 0.01 | 118e ± 2.3 |
| White skins | ||||
| MO | 5.51ef ± 0.28 | 18.1a ± 0.1 | 2.23d ± 0.18 | 14.0a ± 1.1 |
| AR | 4.62d ± 0.08 | 21.6a ± 0.4 | 1.77c ± 0.10 | 16.9a ± 0.1 |
| ER | 5.36e ± 0.54 | 18.8a ± 0.1 | 1.85c ± 0.11 | 16.2a ± 0.9 |
| MT | 6.42g ± 0.10 | 15.6a ± 0.1 | 1.82c ± 0.01 | 16.5a ± 0.1 |
| CH | 5.88fg ± 0.44 | 16.7a ± 0.6 | 2.26d ± 0.24 | 13.1a ± 1.4 |
| NA | 6.45g ± 0.29 | 15.5a ± 0.7 | 2.44d ± 0.30 | 12.4a ± 1.5 |
| RI | 5.69efg ± 0.20 | 17.1a ± 0.6 | 2.38d ± 0.12 | 12.6a ± 0.6 |
1 Values are the means ± SD. Different apices in the same column indicate significant differences (LSD, p < 0.05).
Table 2.
Phenolic contents (mg/L) of the 60% ethanol extract of a heterogeneous batch of red grape skins.
Table 2.
Phenolic contents (mg/L) of the 60% ethanol extract of a heterogeneous batch of red grape skins.
| Parameter | |
|---|---|
| delphinidin-3-O-glucoside | 34.5 ± 1.4 |
| cyanidin-3-O-glucoside | 14.9 ± 0.6 |
| petunidin-3-O-glucoside | 44.4 ± 1.8 |
| peonidin-3-O-glucoside | 17.9 ± 0.7 |
| malvidin-3-O-glucoside | 93.4 ± 3.7 |
| malvidin-p-coumaroyl-glucoside | 17.9 ± 0.7 |
| quercetin 3-O-glucoside | 29.7 ± 1.2 |
| quercetin | 27.2 ± 1.1 |
| kaempferol | 5.2 ± 0.2 |
| procyanidin B1 | 6.6 ± 0.3 |
| catechin | 37.3 ± 1.5 |
| procyanidin B2 | 10.3 ± 0.4 |
| epicatechin | 17.5 ± 0.7 |
| oligomeric procyanidins | 3180 ± 130 |
| total phenolics (GAE) | 5490 ± 220 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



