
Submitted:
17 November 2024
Posted:
19 November 2024
You are already at the latest version
Abstract
Human papillomavirus (HPV) is a prevalent sexually transmitted infection, implicated in various cancers, yet its influence in non-cancerous oesophageal tissue remains unclear. This study aims to investigate the gene expression changes associated with high-risk HPV (HR-HPV) in non-cancerous oesophageal tissue to elucidate potential early oncogenic mechanisms. Using RNA sequencing, we compared transcriptomic profiles of HPV-positive and HPV-negative non-cancerous oesophageal tissues. Differential gene expression analysis revealed significant upregulation of cell cycle and DNA replication pathways in HPV-positive samples, specifically involving key genes such as CCNA2, DSN1, and MCM10, which are known to regulate cellular proliferation and genomic stability. Additionally, kinase and transcription factor enrichment analyses highlighted HR-HPV-associated regulatory molecules, including E2F4 and CSNK2A1, suggesting HPV's role in modulating host cell cycle control. These findings support the hypothesis that HPV infection may initiate cellular alterations in oesophageal tissue, potentially predisposing it to malignancy. This study contributes to understanding HPV's impact in non-cancerous tissues and identifies possible biomarkers for early HPV-related cellular changes, offering insights into HPV-driven cancer development beyond traditionally associated sites.
Keywords:
1. Introduction
2. Results
2.1. Pathological Characteristics and Detection of HR-HPV
2.2. Transcriptomic Profiling of Non-Cancerous Oesophageal Tissue with and Without HR-HPV Infection
2.3. Analysis of Differentially Expressed Genes of Non-Cancerous Oesophageal Tissue with and Without HR-HPV Infection Using Pathway Enrichment Analysis
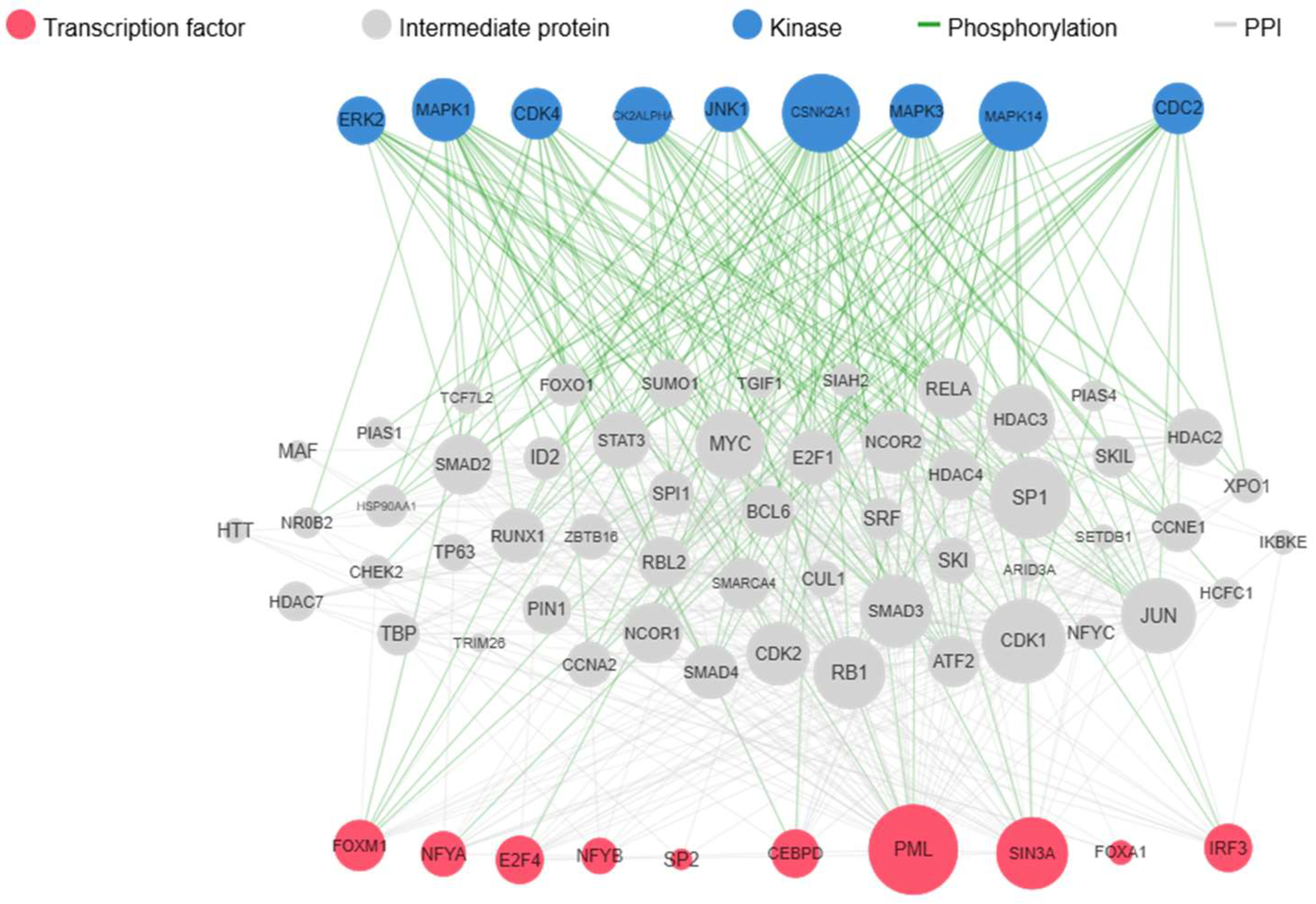
2.4. Investigation of Differentially Expressed Genes of Non-Cancerous Oesophageal Tissue with and Without HR-HPV Infection via Kinase Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Oesophageal Tissue Specimen Collection
4.2. Genomic Material Extraction and Purification
4.3. Detection and Genotyping of HPV DNA
4.4. RNA Sequencing
4.5. Bioinformatics Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graham, S. V. The Human Papillomavirus Replication Cycle, and Its Links to Cancer Progression: A Comprehensive Review. Clin Sci 2017, 131. [Google Scholar] [CrossRef] [PubMed]
- Kombe Kombe, A.J.; Li, B.; Zahid, A.; Mengist, H.M.; Bounda, G.-A.; Zhou, Y.; Jin, T. Epidemiology and Burden of Human Papillomavirus and Related Diseases, Molecular Pathogenesis, and Vaccine Evaluation. Front Public Health 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- de Sanjosé, S.; Brotons, M.; Pavón, M.A. The Natural History of Human Papillomavirus Infection. Best Pract Res Clin Obstet Gynaecol 2018, 47, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Tramontano, L.; Sciorio, R.; Bellaminutti, S.; Esteves, S.C.; Petignat, P. Exploring the Potential Impact of Human Papillomavirus on Infertility and Assisted Reproductive Technology Outcomes. Reprod Biol 2023, 23, 100753. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, G.H.; Haghshenas, M.R.; Marchetti, B.; O’Brien, P.M.; Campo, M.S. E5 Protein of Human Papillomavirus Type 16 Selectively Downregulates Surface HLA Class I. Int J Cancer 2005, 113, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, G.H.; Salman, N.A. Pathogenesis of Human Papillomavirus – Immunological Responses to HPV Infection. In Human Papillomavirus - Research in a Global Perspective; InTech, 2016.
- Bhattacharjee, R.; Das, S.S.; Biswal, S.S.; Nath, A.; Das, D.; Basu, A.; Malik, S.; Kumar, L.; Kar, S.; Singh, S.K.; et al. Mechanistic Role of HPV-Associated Early Proteins in Cervical Cancer: Molecular Pathways and Targeted Therapeutic Strategies. Crit Rev Oncol Hematol 2022, 174, 103675. [Google Scholar] [CrossRef]
- Jensen, J.E.; Becker, G.L.; Jackson, J.B.; Rysavy, M.B. Human Papillomavirus and Associated Cancers: A Review. Viruses 2024, 16, 680. [Google Scholar] [CrossRef]
- Pešut, E.; Đukić, A.; Lulić, L.; Skelin, J.; Šimić, I.; Milutin Gašperov, N.; Tomaić, V.; Sabol, I.; Grce, M. Human Papillomaviruses-Associated Cancers: An Update of Current Knowledge. Viruses 2021, 13, 2234. [Google Scholar] [CrossRef]
- Syrjänen, K.; Pyrhönen, S.; Aukee, S.; Koskela, E. Squamous Cell Papilloma of the Esophagus: A Tumour Probably Caused by Human Papilloma Virus (HPV). Diagn Histopathol 1982, 5, 291–296. [Google Scholar]
- Abbas, G.; Krasna, M. Overview of Esophageal Cancer. Ann Cardiothorac Surg 2017, 6, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Su, F.; Yang, H.; Xiao, Y.; Zhang, X.; Su, H.; Zhang, T.; Bai, Y.; Ling, X. E2F1 Transcriptionally Regulates CCNA2 Expression to Promote Triple Negative Breast Cancer Tumorigenicity. Cancer Biomarkers 2022, 33, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.; Kumar, S. NEDD1: Function in Microtubule Nucleation, Spindle Assembly and Beyond. Int J Biochem Cell Biol 2007, 39, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Wen, T.; Liu, D.; Wang, S.; Jiang, X.; Zhao, S.; Huang, G. DSN1 Is a Prognostic Biomarker and Correlated with Clinical Characterize in Breast Cancer. Int Immunopharmacol 2021, 101, 107605. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.-P.; Wang, J.-Y.; Jao, S.-W.; Wu, C.-C.; Chen, J.-H.; Hsiao, K.-H.; Lin, C.-Y.; Chen, S.-H.; Su, S.-Y.; Chen, Y.-J.; et al. Over-Expression of AURKA, SKA3 and DSN1 Contributes to Colorectal Adenoma to Carcinoma Progression. Oncotarget 2016, 7, 45803–45818. [Google Scholar] [CrossRef]
- Duensing, S.; Münger, K. Human Papillomaviruses and Centrosome Duplication Errors: Modeling the Origins of Genomic Instability. Oncogene 2002, 21, 6241–6248. [Google Scholar] [CrossRef]
- Scarth, J.A.; Patterson, M.R.; Morgan, E.L.; Macdonald, A. The Human Papillomavirus Oncoproteins: A Review of the Host Pathways Targeted on the Road to Transformation. Journal of General Virology 2021, 102. [Google Scholar] [CrossRef]
- Yeeles, J.T.P.; Deegan, T.D.; Janska, A.; Early, A.; Diffley, J.F.X. Regulated Eukaryotic DNA Replication Origin Firing with Purified Proteins. Nature 2015, 519, 431–435. [Google Scholar] [CrossRef]
- Cheng, J.; Lu, X.; Wang, J.; Zhang, H.; Duan, P.; Li, C. Interactome Analysis of Gene Expression Profiles of Cervical Cancer Reveals Dysregulated Mitotic Gene Clusters. Am J Transl Res 2017, 9, 3048–3059. [Google Scholar]
- Ghongane, P.; Kapanidou, M.; Asghar, A.; Elowe, S.; Bolanos-Garcia, V.M. The Dynamic Protein Knl1 – a Kinetochore Rendezvous. J Cell Sci 2014. [CrossRef]
- Cui, Y.; Zhang, C.; Ma, S.; Guo, W.; Cao, W.; Guan, F. <scp>CASC5</Scp> Is a Potential Tumour Driving Gene in Lung Adenocarcinoma. Cell Biochem Funct 2020, 38, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- BOYER, S.N.; WAZER, D.E.; BAND, V. E7 Protein of Human Papilloma Virus-16 Induces Degradation of Retinoblastoma Protein through the Ubiquitin-Proteasome Pathway. Cancer Res 1996, 56, 4620–4624. [Google Scholar] [PubMed]
- Bieging, K.T.; Mello, S.S.; Attardi, L.D. Unravelling Mechanisms of P53-Mediated Tumour Suppression. Nat Rev Cancer 2014, 14, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.T.; Schiffman, M.; Solomon, D. Prospective Follow-up Suggests Similar Risk of Subsequent Cervical Intraepithelial Neoplasia Grade 2 or 3 among Women with Cervical Intraepithelial Neoplasia Grade 1 or Negative Colposcopy and Directed Biopsy. Am J Obstet Gynecol 2003, 188, 1406–1412. [Google Scholar] [CrossRef]
- Kitchener, H.C.; Almonte, M.; Wheeler, P.; Desai, M.; Gilham, C.; Bailey, A.; Sargent, A.; Peto, J. HPV Testing in Routine Cervical Screening: Cross Sectional Data from the ARTISTIC Trial. Br J Cancer 2006, 95, 56–61. [Google Scholar] [CrossRef]
- Derchain, S.F.; Sarian, L.O.; Naud, P.; Roteli-Martins, C.; Longatto-Filho, A.; Tatti, S.; Branca, M.; Eržen, M.; Serpa-Hammes, L.; Matos, J.; et al. Safety of Screening with Human Papillomavirus Testing for Cervical Cancer at Three-Year Intervals in a High-Risk Population: Experience from the LAMS Study. J Med Screen 2008, 15, 97–104. [Google Scholar] [CrossRef]
- Gillison, M.L.; Broutian, T.; Pickard, R.K.L.; Tong, Z.; Xiao, W.; Kahle, L.; Graubard, B.I.; Chaturvedi, A.K. Prevalence of Oral HPV Infection in the United States, 2009-2010. JAMA 2012, 307, 693. [Google Scholar] [CrossRef]
- Giuliano, A.R.; Viscidi, R.; Torres, B.N.; Ingles, D.J.; Sudenga, S.L.; Villa, L.L.; Luiza Baggio, M.; Abrahamsen, M.; Quiterio, M.; Salmeron, J.; et al. Seroconversion Following Anal and Genital HPV Infection in Men: The HIM Study. Papillomavirus Research 2015, 1, 109–115. [Google Scholar] [CrossRef]
- Waghray, A.; Schober, M.; Feroze, F.; Yao, F.; Virgin, J.; Chen, Y.Q. Identification of Differentially Expressed Genes by Serial Analysis of Gene Expression in Human Prostate Cancer. Cancer Res 2001, 61, 4283–4286. [Google Scholar]
- Rakha, E.A.; Pinder, S.E.; Paish, E.C.; Robertson, J.F.; Ellis, I.O. Expression of E2F-4 in Invasive Breast Carcinomas Is Associated with Poor Prognosis. J Pathol 2004, 203, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Rakha, E.A.; Armour, J.A.L.; Pinder, S.E.; Paish, C.E.; Ellis, I.O. High-resolution Analysis of 16q22.1 in Breast Carcinoma Using DNA Amplifiable Probes (Multiplex Amplifiable Probe Hybridization Technique) and Immunohistochemistry. Int J Cancer 2005, 114, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.; Sage, J. Novel Functions for the Transcription Factor E2F4 in Development and Disease. Cell Cycle 2016, 15, 3183–3190. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Fu, Q.; Xu, J.; Gu, X.; Zhou, H.; Zhi, C. Transcription Factor E2F4 Is an Indicator of Poor Prognosis and Is Related to Immune Infiltration in Hepatocellular Carcinoma. J Cancer 2021, 12, 1792–1803. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Ren, Z.; Li, W. E2F4 Transcription Factor Is a Prognostic Biomarker Related to Immune Infiltration of Head and Neck Squamous Cell Carcinoma. Sci Rep 2022, 12, 12132. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.-B.; Li, X.-Z.; Zeng, S.; Liu, C.; Yang, S.-M.; Yang, L.; Hu, C.-J.; Bai, J.-Y. Regulation of the Master Regulator FOXM1 in Cancer. Cell Communication and Signaling 2018, 16, 57. [Google Scholar] [CrossRef]
- Rachmadi, L.; Billianti Susanto, Y.D.; Sitinjak, D.; Manatar, A.F.; Saraswati, M.; Adham, M. HPV Infection Is Associated with FoxM1 Overexpression in Dysplastic Changes of Sinonasal Inverted Papilloma. Asian Pac J Cancer Prev 2022, 23, 4293–4298. [Google Scholar] [CrossRef]
- Strum, S.W.; Gyenis, L.; Litchfield, D.W. CSNK2 in Cancer: Pathophysiology and Translational Applications. Br J Cancer 2022, 126, 994–1003. [Google Scholar] [CrossRef]
- Bae, J.S.; Park, S.-H.; Jamiyandorj, U.; Kim, K.M.; Noh, S.J.; Kim, J.R.; Park, H.J.; Kwon, K.S.; Jung, S.H.; Park, H.S.; et al. CK2α/CSNK2A1 Phosphorylates SIRT6 and Is Involved in the Progression of Breast Carcinoma and Predicts Shorter Survival of Diagnosed Patients. Am J Pathol 2016, 186, 3297–3315. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Ma, Z.; Zhang, G.; Yang, X.; Du, Q.; Wang, W. <p>CSNK2A1 Promotes Gastric Cancer Invasion Through the PI3K-Akt-MTOR Signaling Pathway</P>. Cancer Manag Res 2019, Volume 11, 10135–10143. [Google Scholar] [CrossRef]
- Baker, S.J.; Poulikakos, P.I.; Irie, H.Y.; Parekh, S.; Reddy, E.P. CDK4: A Master Regulator of the Cell Cycle and Its Role in Cancer. Genes Cancer 2022, 13, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.; Sicinski, P.; Hinds, P.W. Cyclins and Cdks in Development and Cancer: A Perspective. Oncogene 2005, 24, 2909–2915. [Google Scholar] [CrossRef] [PubMed]
- Jungert, K.; Buck, A.; Wichert, G. von; Adler, G.; König, A.; Buchholz, M.; Gress, T.M.; Ellenrieder, V. Sp1 Is Required for Transforming Growth Factor-β–Induced Mesenchymal Transition and Migration in Pancreatic Cancer Cells. Cancer Res 2007, 67, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Engeland, K. Cell Cycle Regulation: P53-P21-RB Signaling. Cell Death Differ 2022, 29, 946–960. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Guidance on Regulations for the Transport of Infectious Substances, 2023–2024; 2024. [Google Scholar]
- Jiang, H.; Lei, R.; Ding, S.-W.; Zhu, S. Skewer: A Fast and Accurate Adapter Trimmer for next-Generation Sequencing Paired-End Reads. BMC Bioinformatics 2014, 15, 182. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol 2014, 15, 550. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proceedings of the National Academy of Sciences 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database Hallmark Gene Set Collection. Cell Syst 2015, 1, 417–425. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M. V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr Protoc 2021, 1. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinformatics 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M. V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Xu, H.; Gordonov, S.; Lim, M.P.; Perkins, M.H.; Ma’ayan, A. Expression2Kinases: MRNA Profiling Linked to Multiple Upstream Regulatory Layers. Bioinformatics 2012, 28, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.J.B.; Kuleshov, M. V; Schilder, B.M.; Torre, D.; Duffy, M.E.; Keenan, A.B.; Lachmann, A.; Feldmann, A.S.; Gundersen, G.W.; Silverstein, M.C.; et al. EXpression2Kinases (X2K) Web: Linking Expression Signatures to Upstream Cell Signaling Networks. Nucleic Acids Res 2018, 46, W171–W179. [Google Scholar] [CrossRef] [PubMed]
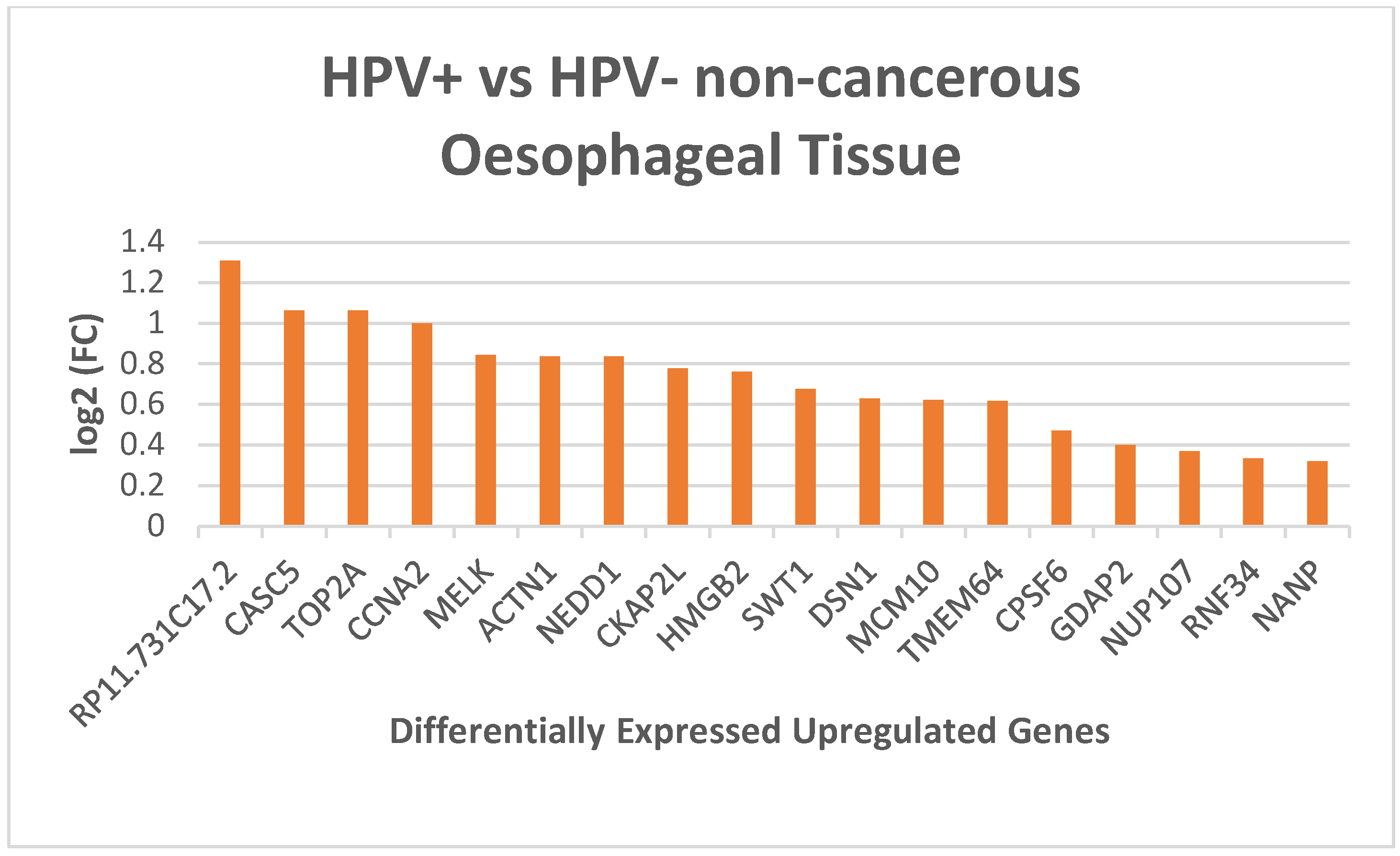

| Description | Number of overlap genes | p-Value | Overlap Genes |
|---|---|---|---|
| Cell Cycle | 7 | 7.49 x 10-8 | TOP2A; CCNA2; NEDD1; DSN1; NUP107; CASC5; MCM10 |
| DNA Replication | 5 | 8.69 x 10-5 | CCNA2; DSN1; NUP107; CASC5; MCM10 |
| A | |
| Transcription Factors | p-Value |
| E2F4 | 1.6 x 10-8 |
| FOXM1 | 1.5 x 10-4 |
| CEBPD | 9.1 x 10 -4 |
| B | |
| Regulatory Kinases | p-Value |
| CSNK2A1 | 2.2 x 10-15 |
| CDK4 | 7.0 x 10-15 |
| MAPK14 | 1.05 x 10-12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



