1. Introduction
Living organisms are highly ordered, spatially distinct, semi-open systems that maintain a stable low entropy state while far from thermodynamic equilibrium [
1,
2]. Living organisms, unlike all other structures in nature, use information to maintain this non-equilibrium state for prolonged periods of time. Information components of living systems include heritable instructions or blueprints for macromolecules in the genome. In addition, living organisms must continuously access and process information in the environment to identify risks and opportunities, which may vary both over time and across space [
1].
Here we investigate the origin of information dynamics in living systems. Virtually all origin of life scenarios include the critical role of information, but most focus on pre-biotic synthesis of the molecular building blocks that encode information for replication. From the standpoint of evolutionary biology, this work, which has made impressive strides in recent decades, addresses the proximate causal basis [sensu Mayr [
2]] for the evolution of life by proposing potential mechanisms for how geochemistry evolved into life (biochemistry). In contrast, we focus on the evolutionary conditions in which information becomes critically linked to survival and proliferation and, therefore, both allows and is subject to Darwinian selection. From the standpoint of evolutionary biology, our proposal concentrates on ultimate causality [
2], i.e., why were certain protocells successful in the transition from geochemistry to life.
Evolutionary strategies are frequently framed through fundamental trade-offs, one of which is food and safety [
3]. That is, acquiring food [resources] necessarily entails greater exposure to risk. We argue that this tradeoff existed during the earliest development of protocells and imposed selection forces that promoted information dynamics observable in all past and current living systems.
We propose the initial source of information could not be environmental factors that are constant or vary stochastically. Rather, early pre-biotic systems could develop a “non-cognizant intelligence” to optimize their replicative ability only if the perturbation were predictably variable. Thus, we suggest biological information dynamics were evolutionarily selected to optimize fitness in the changing conditions caused by night/day transitions on primordial earth.
These regularly varying environmental characteristics imposed different opportunities and hazards. We propose initial biological information was specifically about time and allowed living systems to anticipate and adapt to the predictably varying environmental conditions during day/night cycles. Other, longer cyclical phenomena related to the orbit of the earth around the sun [and the moon around the earth] include the tides, climate, and ocean currents also represent informational cycles that drive additional adaptations. However, we hypothesize these dynamics could impose selection forces only after a genetic storage system was in place and remained stable over prolonged periods of time.
Here we focus on the role of diurnal environmental variations in evolution of primordial cellular structures and information dynamics. In addition to regular cyclical temperature changes, daytime hours, in the absence of an oxygen atmosphere in early earth, included intense solar UV radiation. While this promoted pre-biotic chemistry that synthesized necessary organic molecules [resources], it could also fatally damage critical components of the [proto]cell. This represents a classic [and perhaps first] food/safety trade-off in living systems. Daytime hours with increased temperature and UV irradiation accelerated chemical reactions to produce more resources and probably increased diffusion and convection currents in their environment allow greater delivery of substrate. Thus, to succeed, a protocell would necessarily need to capture scarce organic molecule resources while also avoiding the damaging effects of UV radiation. We thus hypothesize that a protocell would need to move in its environment in a way that is referred to as “viability-based behavior” [
4], defined as “a way that simple entities can adaptively regulate their environment in response to their health, and in so doing, increase the likelihood of their survival.”
We hypothesize these cyclical variations in environmental conditions selected for temporal information dynamics in protocells containing self-replicating polynucleotides that allowed them to anticipate critical environmental selection forces and adapt to them quickly or a priori to optimize fitness.
If the initial evolutionary selection for information processing capabilities in early living systems emerged in response to the day/night cycle, we anticipate these properties would be observed across extant life forms. Diurnal variations in the state of biological systems are, indeed, ubiquitous across the tree of life. In past work, we have noted that the 4 nucleotide components of DNA in all living systems critically share an asymmetry that allows rapid synthesis of a daughter strand of DNA or RNA from the template [
5,
6,
7]. Because increasing temperature during the day will cause separation of double stranded polynucleotides, synthesis must be completed during cooler nighttime temperatures applying strong selection for faster replication and, therefore, selective inclusion of asymmetric nucleotides.
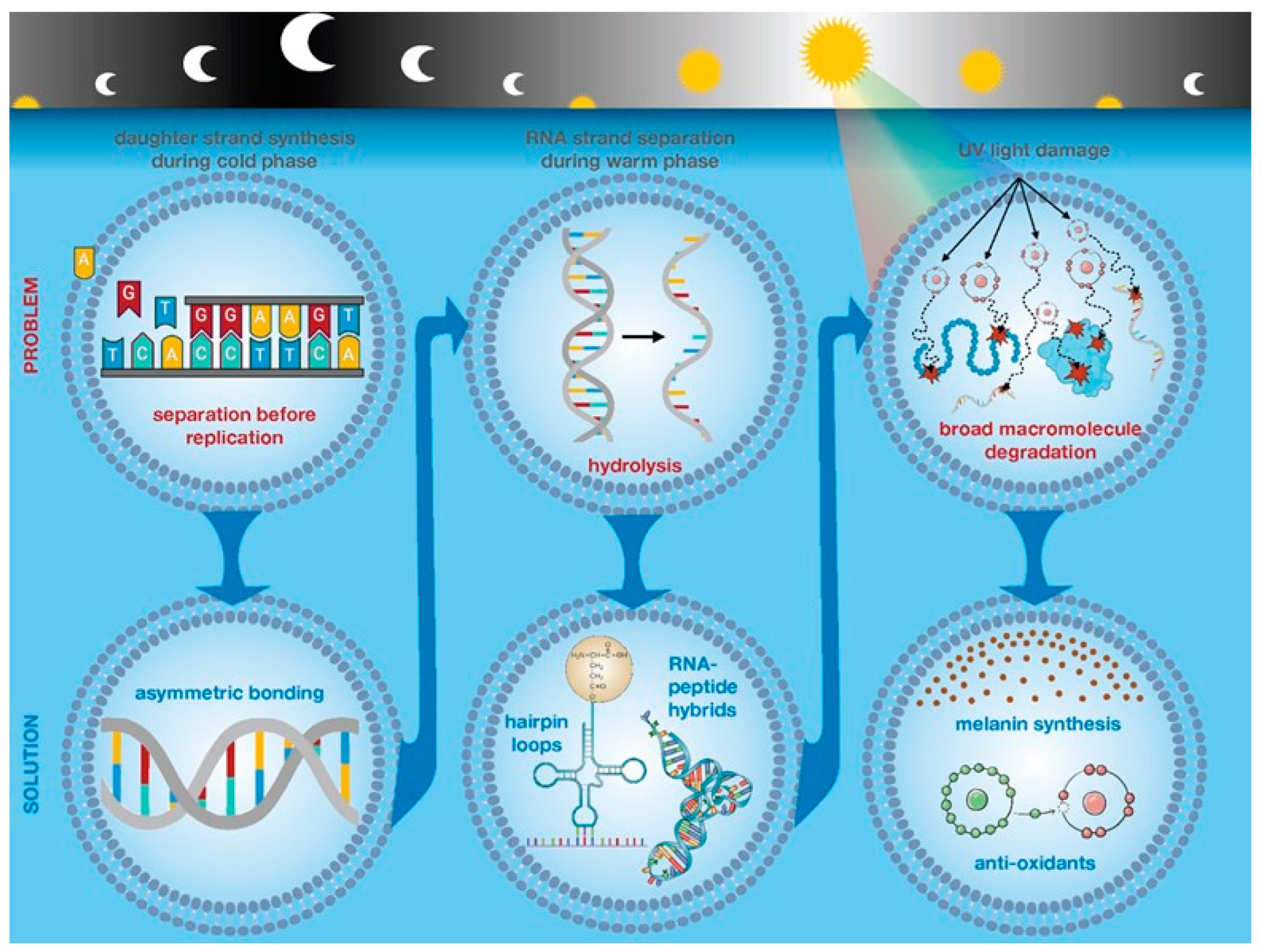
Here we focus on other selection forces during daytime hours as separation of polynucleotide strands, particularly RNA, are subject to hydrolysis as well as intense UV radiation that broadly damages all the macromolecular components of protocells. We propose these Darwinian selection forces, when applied to self-replicating RNA, favored nucleotide sequences that could reduce hydrolysis by self-bonding in hairpin loops or forming RNA-peptide hybrid molecules. In turn, these molecules, when trapped within a cell, could promote their replicative success by protecting the cell from UV damage through scavenging free radicals or by catalyzing synthesis of melanin from tyrosine.
We hypothesize these dynamics persist in modern living systems as Transfer RNA [tRNA], the RNA templated synthesis of proteins, ubiquitous diurnal rhythms, and melanosomes, which are evolutionarily ancient [
8,
9] membrane bound organelles in which melanin is synthesized. Additionally, we propose early living systems could limit UV damage by vertical migration facilitated by replacing intracellular Na+ by heavier K+ ions leading to the transmembrane ion gradients ubiquitously observed in modern life. This indeed may constitute the origins of the ubiquitous diel vertical migrations observed across numerous aquatic organisms today [
10].
2. Building a Model of Information in Early Life
We assume initial conditions of an aquatic environment during the Hadean Eon [4.3-4.0 billion years ago; Ga] or possibly the Eoarchean Era [4.0-3.6 Ga]. At this time, small continents [
11] and volcanic islands had formed [
12]. Putative chemical evidence of early life is preserved in sedimentary rocks from this age (3.95Ga) [
13]. The origin of life is often discussed to be associated to hydrothermal vent system, because it offers ample chemical energy, and the deep ocean niche was shielded from UV radiation and interruptive meteorite bombardment as late as ~3.5 Ga [
14]. In contrast, shallow shelf surfaces of early continents indeed would be exposed to not only bombardment of meteorites but also daytime UV radiation, which was much higher than at present, owing to a more active [although less luminous] sun [
15] and absence of an ozone layer [
16]. Despite the assumed destructive nature of meteorite impacts, the high temperatures generated may have resulted in reactions that produced necessary feedstocks from which RNA, proteins and lipids ultimately arose [
17].
Our assumed initial conditions include availability of lipids, polypeptides, and nucleotides that formed through condensation reactions [
18]. Amphiphilic compounds [those with polar and nonpolar components] can spontaneously self-assemble into bi-molecular layers forming closed membranous vesicles [
19]. Such vesicles in an aqueous environment rich in organic molecules could entrap those organic molecules, including self-replicating polynucleotides as well as mononucleotides and amino acids.
3. Fulfilling Darwin’s Postulates in Primordial Conditions
We propose a critical requirement for the origin of life is a non-dissipative environment, like a tidal pool or small lake, that physically constrains both self-replicating polynucleotides and their essential molecular building blocks [
20]. Here, nucleotides, amino acids, fatty acids, and other molecules are present in the environment from local pre-biotic synthesis with sustained or periodic influx of substrate [
21] but in limited quantities. Thus, self-replicated polynucleotides and protocells within a constrained space must compete for limited substrate and space.
We note these conditions fulfill Darwin’s three postulates [
22] necessary for evolution by natural selection. Self-replicating polynucleotides that can affect the phenotypic properties of the protocell provide the necessary heritable variations. The diurnal fluctuations of benefits and threats, in a resource-limited, non-diffusive environment (see below), apply selection pressures that determine proliferation [
23]. Those most adapted to the cycling opportunities and hazards will survive and replicate, at the expense of those less capable generating a “struggle for existence.”
4. Diurnal Variations in Evolutionary Selection Pressures
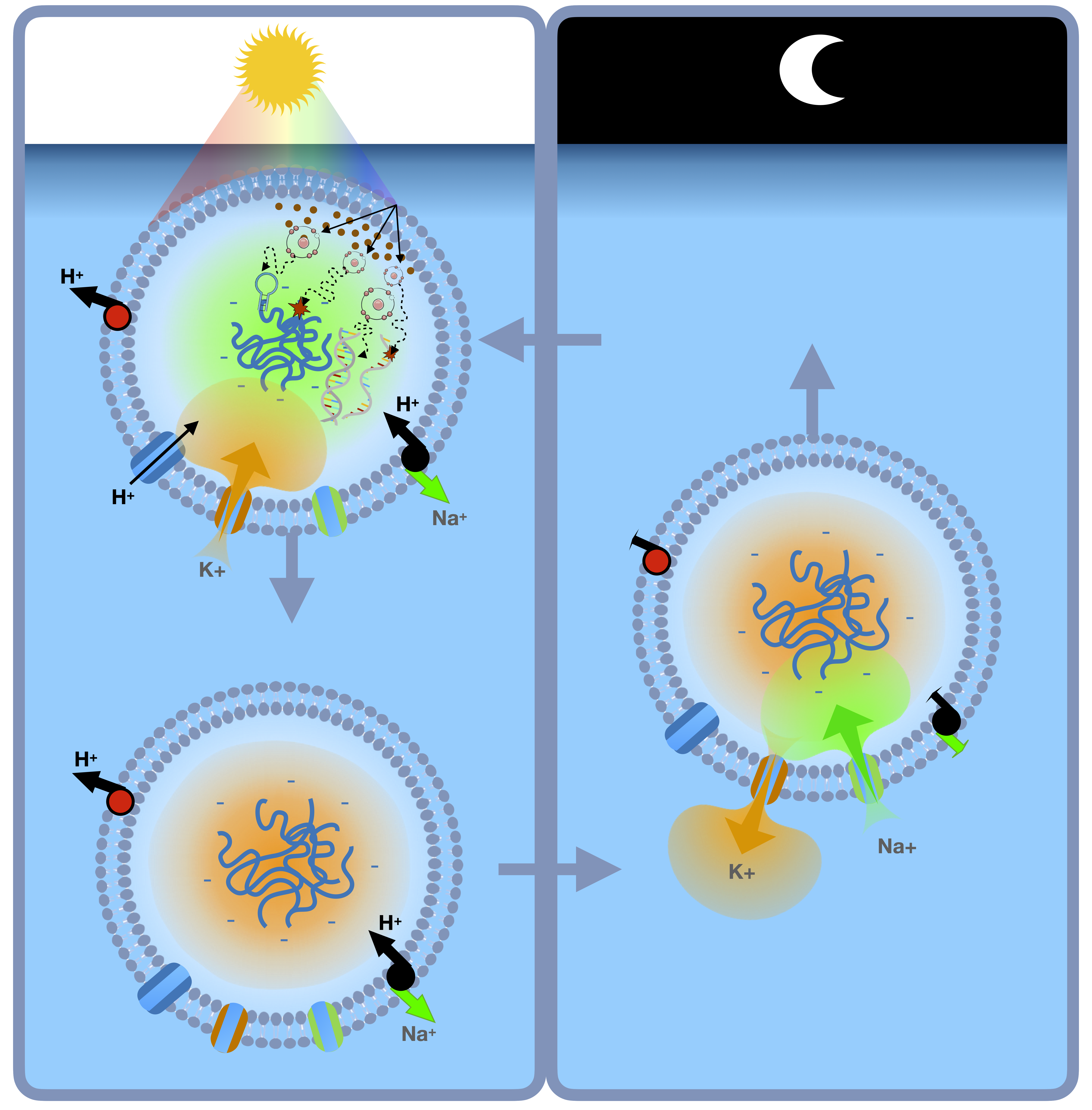
We next hypothesize that critical information available to early self-replicators in this environment arises through variation in selection forces during the regular day/night cycles (
Figure 1). Although tides in the deep ocean niches pace variations in temperature and energy near hydrothermal vents [
24] , the influence of time on vents is an open question [
25] . However, oscillation in shallow marine settings encompass sharper diurnal variations in UV light, oxidants, and temperature.
In daylight hours, intense UV exposure could drive pre-biotic chemistry, including formation of iron sulfur clusters [
26]. Noting that solar radiation is the paramount source of energy on Earth and can lead to synthesis of organic molecules from nonorganic building blocks, Green et al. [
27] identified photochemical pathways or schemes relevant to prebiotic molecules and reactions (such as condensation of biomolecules). A growing literature reports that UV irradiation can induce reactions that produce prebiotic precursors to the chemical building blocks of life [
17,
28] in plausible environmental conditions of early earth. As stated by Nunes Palmeira et al. [
29] “The idea that metabolism emerged from a geochemical protometabolism therefore looks increasingly persuasive”.
However, daytime production of the substrate from prebiotic chemistry is countered by hazards generating a food and safety tradeoff. UV light could also damage nascent living systems [
30]. Green et al. [
19] emphasize that, even as UV radiation provides energy for formation of chemical bonds and synthesis of biomolecules, it can also drive breakdown of organic molecules via photolysis. Mulkidjanian and Junge [
31,
32] proposed modern photosynthetic reaction centers evolved from earlier molecular “technologies” that evolved to protect “primordial” cells from UV damage. This destructive nature of UV radiation is the driving force of numerous protective strategies in modern organisms [
33].
In addition, diurnal variations produce daily temperature changes. These regular temperature cycles, similar to Polymerase Chain Reactions (PCR), could promote self-replicating polynucleotide strands [
7]. However, warmer daytime conditions may “melt” hydrogen bonds causing separation of double stranded polynucleotides. Subramanian et al [
6,
7] demonstrated this strongly selected monomers that permitted maximally fast synthesis of complementary strands for polynucleotides to complete strand duplication prior to onset of daytime warmer temperatures and strand separation. Molecular modeling of strand synthesis [
6,
7] demonstrated strong selection for asymmetric monomers that produce increased energy in covalent bonds in the existing strand, thus reducing the probability of breakage, while decreasing bonding energy at the developing edge of the growing strand to increase the probability of bonding with a monomer in solution.
While melting of double stranded polynucleotides provides theoretic opportunity for self-replication, single stranded polynucleotides, particularly RNA, will be subject to rapid hydrolysis [
33]. Evolutionarily this precludes self-replication as well as loss of any potential function of the RNA molecule.
An additional selection force in increased daytime temperatures is denaturation of folded polypeptides and increase the permeability of lipid bilayers. The former yields a prediction that proteins selected in early life would remain stably folded through the range of extant environmental temperatures. Consistent with this, we note the E. coli has a more thermostable proteome than human cells [
34] and critical proteins, such as those in the respiratory chains, are more thermally stable across the tree of life. Interestingly, Tyrosinase, critical for synthesis of melanin (see below) is stable between 20 and 65 C [
35]. However, increased cell membrane permeability may allow more building blocks into the protocell and thus contribute to the food part of the tradeoff.
5. Evolutionary Heritage of Prebiotic Information Acquisition
5.1. Evolution of Circadian Clocks
We propose that cycling selection forces in pre-biotic life specifically favored an ability to assess and respond to information about time. As noted by Green et al. [
27], early life would have evolved primitive mechanisms to take advantage of the benefits of UV radiation while simultaneously avoiding its corrosive effect of photolysis would have promoted some mechanism to time their activities. Similarly, Baluška and Reber [
36] noted “circadian clocks are inherently cognitive in nature”, and they must have “co-evolved with the first cells to safeguard their survival”.
5.2. Building a Diurnal Clock in Cells
We emphasize onset of daylight hours produces three significant changes that constitute strong selection forces on protocells: (1) UV radiation that can broadly damage the macromolecular structures of the cell, (2) increased temperature, that can cause separation of double strands of polynucleotides, and (3) single stranded polynucleotides, particularly RNA, are subject to rapid degradation by hydrolysis, thus effectively eliminating the possibility that long polynucleotide strands could consistently self-replicate.
We propose these (roughly) simultaneous selection forces were critical for development of capabilities to assess temporal information that anticipated the onset of UV and thus, prevented hydrolysis while simultaneously producing changes that protected the protocell from UV damage. That is, the melting of double stranded RNA due to increasing temperature during daytime hours and the subsequent adaptations to prevent hydrolysis also produced molecular machinery to shield cellular components (e.g. membrane) from damage from UV light. We hypothesize this is the origin of informational dynamics still observed in the circadian changes of extant living systems [
37].
Several scenarios may provide protective effects. Recent work has shown peptides can bind to single stranded RNA producing a hybrid molecule that is relatively stable [
38,
39]. This could ultimately lead to the triplet RNA code for amino acids. Furthermore, we note that RNA strands that form hairpin configurations, similar to the structure of modern tRNA, are relatively shielded from hydrolysis [
39].
We note several potential mechanisms by which strand melting and adaptation to hydrolysis could shield the protocell from UV radiation. We assume that early self-replicating polynucleotides would be relatively short and, thus, produce capable of producing small peptides which can have both antioxidant and UV shielding properties. Recent work also reports that a hybrid RNA/protein molecule can synthesize melanin in the absence of tyrosinase [
40]. Since tyrosine can be generated by pre-biotic synthesis [
41], it seems plausible that even very primitive early living systems could have synthesized melanin to protect from UV damage. We note such cells would likely have been highly successful competitors and these dynamics may be preserved in modern life as melanosomes, which are evolutionarily ancient [
8] membrane enclosed intracellular structures that are broadly involved in stress response and specialize in melanin production [
42].
The melanin family of proteins are diverse and can form heterogenous (often cross-linked) and assembled oligomers that interact with metals and can form amyloid fibrils [
43]. While they are most recognized for shielding cells from UV radiation, melanin proteins have diverse cellular functions including immune interactions, calcium homeostasis [
44], and endosomal sorting [
8,
42]. Melanosomes are found in all living organisms from bacteria to mammals and observed in early evolutionary life forms [
45,
46]. Melanosomes are ubiquitous across the tree of life [
8]. Moreover, melanosome formation is strongly linked to intraluminal pH which is maintained by extrusion of Na+ via TPC2 [
47,
48], which may influence buoyancy and further adaptations to avoid UV light (see below).
5.3. Evolutionary Selection for the Transmembrane Ion Gradient
One of the characteristics demanding explanation in origin of life research is the relative concentrations of potassium (K) and sodium (Na) in the cytoplasm compared to extracellular fluids and sea water, which is often hypothesized to be the environment in which early life originated. The cell cytoplasm of all living organisms contains a high concentration of K
+ and a low concentration of Na
+ relative to their environment. About 1/3 of a cell’s energy budget is used to pump ions against a concentration gradient [
49] but the corresponding benefit to justify this expenditure or resources remains unclear. As pointed out by Hansma [
50], either life originated in an environment with a high concentration of potassium, or, after life’s origin, early cells evolved a preference for potassium over the more ubiquitous sodium (which occurs in high concentration in sea water). Even if early living systems developed in a high K
+, low Na
+ environment, maintaining these intracellular concentrations in a seawater environment would require large investments of energy. We suggest that this strongly indicates that the high K
+ low Na
+ intracellular environment confers a pronounced evolutionary benefit.
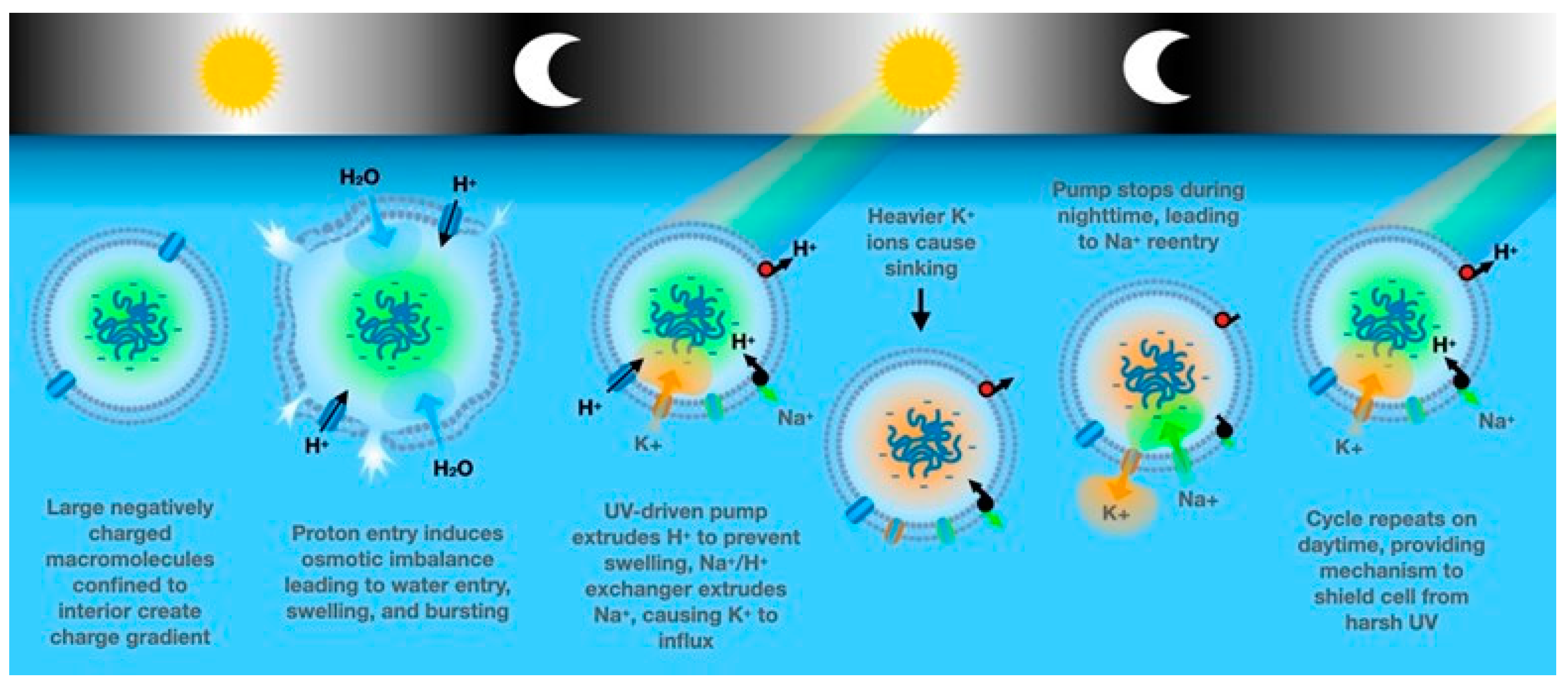
Within the context of our hypothesis, we propose any mechanism that may allow a protocell to control its depth in the thermal pond (or sea) with respect to the light-dark circadian cycle would be highly advantageous – allowing the cell to benefit from the UV -driven selective synthesis of biomolecules during daylight, while simultaneously avoiding UV-driven photolysis of those biomolecules. For instance, many aquatic prokaryote species possess a gas vacuole, which confers the cell with buoyancy, and their widespread occurrence among bacteria and archaea suggests that these organelles evolved very early [
51]. Before the evolution of the gas vacuole, a rather sophisticated cell organelle, we hypothesize that an earlier mechanism that could have provided the protocell with a capacity of buoyancy arose from ion channels, transporters, and pump-like membrane proteins that early cells or protocells must have developed to counter the Gibbs-Donnan effect [
52]. Sephus et al. [
53] suggest that a likely early proton pump providing the active transport needed to avoid cell rupture consisted of ancestral rhodopsins that evolved over “a span of time encompassing Earth’s early history of inhabitation". Morowitz ([
54] p. 166) offered a similar perspective: ”the conversion of photon energy to a chemically useful form may have been contemporaneous with life’s origin."
Assuming early or protocells contained impermeant molecules like nucleic acids and negatively charged metabolites, the Gibbs-Donnan effect will cause cellular swelling and rupture if not countered by the extrusion of sodium by a sodium pump. The “pump-leak” model [
55] proposes a combination of Cl
-, Na
+, K
+-channels, together with Na
+/H
+-exchangers and a bacteriorhodopsin-like H
+-pump must have allowed LUCA (Last Universal Common Ancestor) to resolve the Gibbs-Donnan effect. With these mechanisms in place, we suggest that selective intake of K
+ and extrusion of Na
+, because of the greater mass of K
+, would cause the protocell to sink a few cm in the water column, thus providing a small amount of shielding from UV radiation (
Figure 2). If this occurred at a time that benefited the protocell, this selectivity toward K
+ over Na
+ may eventually become an adaptive strategy to control buoyancy in a fitness-enhancing manner that is another step in what Trapp [
56] refers to as “a transition from chemical to biological systems”.
Thus, we hypothesize that, collectively, early evolution of protective effects of RNA-peptide hybrids in scavenging free radical and producing melanin and selective permeability of cations in early protocell membranes provide mechanisms by which early protocells could effectively shield themselves from UV radiation, as suggested by Green et al. [
27].
5.4. Evolution Homochirality
Like the cytoplasmic concentrations of K
+ and Na
+ discussed above, a unique property of life (but see Glavin et al [
57]) is the homochirality of amino acids and sugars, with the left-handedness (L) of amino acids, and the right-handedness (D) of sugars. Since the seminal publication of Frank [
58] numerous papers have examined the potential origins of biological homochirality. As discussed by Blackmond [
59,
60], these hypotheses can be classified as probabilistic or deterministic, abiotic or biotic, biological, physical, or chemical, among perhaps others. [
60].
Most of the proposed mechanisms exhibit no obvious relationship to diurnal fluctuations of environmental conditions. A possible exception is a recent publication by Bechtel et al. [
61], which identified a photoredox “pathway for symmetry breaking, chiral amplification, and enantioselective organocatalysis”. These authors conclude that their work opens “access to homochirality through efficient asymmetric catalysis and amplification,” which may then contribute to the resources necessary for growth and reproduction – i.e., the food part of food/safety tradeoff. Bechtel et al. [
61] propose that day and night cycles may provide the “necessary driving force” of such photoredox organocatalysis systems critical to the transition from prebiotic chemistry “to an evolving system with higher complexity”. Photoredox catalysis is a highly active area of chemical research (e.g., Nicewicz and MacMillan [
62]), and we believe that further insights into the origin of biological homochirality may be further elucidated by such studies. Regardless of its origin, biological homochirality, once it has evolved, provides distinct evolutionary advantages [
7,
63].
6. Discussion
Living systems, singularly in nature, use information to produce a highly ordered state that is far from thermodynamic equilibrium [
3,
4]. Here we present a hypothesis that diurnal variations in environmental conditions provided the selection forces by which information became critically linked to survival and proliferation and, therefore, subject to Darwinian optimization.
The origin of life on Earth is a topic of extensive prior research. Much of this work appropriately investigates the pre-biotic chemistry that produces the molecular building blocks of extant living systems. Here we consider the evolutionary selection forces that could plausibly select for the integration of information into the dynamics of systems that would eventually be defined as “living.”
We propose the initial source of information required a process that was neither stable nor stochastically changing. Rather information, specifically about time, can be obtained only from a process that is regularly and predictably variable. Thus, we propose that the information dynamics central to living systems originated in diurnal variations in environmental conditions. In particular, the daytime conditions simultaneously imposed two different opportunities and hazards: warming temperatures and high levels of UV irradiation.
These conditions would accelerate pre-biotic chemistry that produced the molecules necessary for life. However, the UV radiation could produce potentially fatal damage in self-reproducing systems and the increased temperature could cause separation of self-replicating polynucleotides. The latter exposes polynucleotides, particularly RNA, to degradation by hydrolysis and effectively limits the size of self-replicating RNA.
We note the temporal coincidence of these processes selects for simultaneous adaptive strategies that were critically interrelated. Thus, self-replicating polynucleotides, subject to selection pressure of strand separation during warmer daytime hours, must optimize the speed of self-replication. This will select for [
5,
6,
7] asymmetric nucleotides that accelerate daughter strand synthesis by increasing the energy of hydrogen bonds between strands proximal to each added nucleotide (reducing probability of separation) while decreasing the energy needed to form hydrogen bonds at the leading edge (maximizing the probability of binding by the next nucleotide in the growing daughter strand). The legacy of these dynamics persist as the nucleotides in extant living systems are all asymmetric [
6,
7].
In addition, self-replicating RNA can adapt to hydrolysis by producing strands that self-bond in hairpin configuration or by forming peptide-RNA hybrid molecules. These dynamics can, in turn, benefit survival of the whole protocell by scavenging free radicals to limit photolysis. Furthermore, a peptide-RNA hybrid has been shown to catalyze synthesis of melanin from tyrosine and may persist in modern organisms as melanosomes which are membrane bounded organelles that specialize in melanin production and are associated with stress response [
64].
In general, a broad prediction of any theoretical model for the earliest dynamics in life is that the resulting dynamics should be observable in modern organisms. Thus, we note: 1) self-replicating RNA strands that avoid hydrolysis via formation of hair-pin loops persist in the molecular configuration of tRNA. 2) Rapid synthesis of daughter strands of polynucleotides before melting during daytime warming would select for the asymmetric nucleotides observed in all living systems. 3) RNA-peptide hybrids anticipate mRNA-directed protein synthesis in extant life.
Our proposed role for a central role of melanin synthesis in protecting protocells can be tested experimentally. The predictions that adapted protocells may persist as melanosomes is difficult to prove beyond noting melanosome appear to be evolutionarily ancient structures [
8]. Finally, we note adaptations driven by either the self-replication polynucleotides or lipid bilayers could promote K+/Na+ exchange in the membrane to generate a higher intracellular K+ concentration allowing the protocell to sink below the surface thus reducing radiation exposure.
7. Conclusions
In conclusion, although the shallow marine environment of early Earth presented a limited and demanding habitat for life, it was not without opportunities. We hypothesize diurnal fluctuations of these limitations, demands, and opportunities are directly responsible for the information dynamics that distinguish life on earth from all other processes in nature.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, R.A.G. and C.J.W; writing—original draft preparation, R.A.G., C.J.W.; writing—review and editing, R.A.G, J.G., H.S., E.U.H, C.J.W.; visualization, J.G.; funding acquisition, R.A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Moffitt Physical Science Oncology Center NIH grant U54 CA143970.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schmidt KA MF, Szymkowiak J. Resurrecting Shannon’s surprise: landscape heterogeneity complements information use and population growth. Oikos. 2022. [CrossRef]
- Mayr E. Cause and effect in biology. Science. 1961;134(3489):1501-6. [CrossRef]
- Garland T, Jr., Downs CJ, Ives AR. Trade-Offs (and Constraints) in Organismal Biology. Physiol Biochem Zool. 2022;95(1):82-112. [CrossRef]
- Egbert M, Hanczyc MM, Harvey I, Virgo N, Parke EC, Froese T, et al. Behaviour and the Origin of Organisms. Origins Life Evol B. 2023;53(1-2):87-112. [CrossRef]
- Subramanian H, Gatenby RA. Evolutionary advantage of anti-parallel strand orientation of duplex DNA. Sci Rep. 2020;10(1):9883. [CrossRef]
- Subramanian H, Gatenby RA. Evolutionary advantage of directional symmetry breaking in self-replicating polymers. J Theor Biol. 2018;446:128-36. [CrossRef]
- Subramanian H, Brown J, Gatenby R. Prebiotic competition and evolution in self-replicating polynucleotides can explain the properties of DNA/RNA in modern living systems. BMC Evol Biol. 2020;20(1):75. [CrossRef]
- D’Alba L, Shawkey MD. Melanosomes: Biogenesis, Properties, and Evolution of an Ancient Organelle. Physiol Rev. 2019;99(1):1-19. [CrossRef]
- Edwards NP MP, Wogelius RA. Pigments through time. Pigment Cell and Melanoma Research. 2014;27(5):684-9. [CrossRef]
- Brierley AS. Diel vertical migration. Curr Biol. 2014;24(22):R1074-6. [CrossRef]
- Harrison, TM. Hadean Earth. Switzerland: Springer Nature; 2020.
- Deamer DW. Assembling life : how can life begin on earth and other habitable planets? New York, NY: Oxford University Press; 2019. xvii, 166 pages, 24 pages of plates p.
- Lepot, K. Signatures of early microbial life from the Archean (4 to 2.5 Ga) eon. Earth-Science Reviews. 2020;209:103296. [CrossRef]
- Bottke WF, Norman MD. The Late Heavy Bombardment. Annual Review of Earth and Planetary Sciences. 2017;45:619-47. [CrossRef]
- Obridko VN, Ragulskaya MV, Khramova EG. Young Sun, galactic processes, and origin of life. Journal of Atmospheric and Solar-Terrestrial Physics. 2020;208:105395. [CrossRef]
- Cockell CS, Horneck, G. The History of the UV Radiation Climate of the Earth—Theoretical and Space-based Observations. Photochemistry and Photobiology. 2007;73(4):447-51. [CrossRef]
- Patel BH, Percivalle C, Ritson DJ, Duffy CD, Sutherland JD. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat Chem. 2015;7(4):301-7. [CrossRef]
- Ross DS, Deamer D. Dry/Wet Cycling and the Thermodynamics and Kinetics of Prebiotic Polymer Synthesis. Life (Basel). 2016;6(3). [CrossRef]
- Deamer D, Dworkin JP, Sandford SA, Bernstein MP, Allamandola LJ. The first cell membranes. Astrobiology. 2002;2(4):371-81. [CrossRef]
- Toner JD CD. Alkaline lake settings for concentrated prebiotic cyanide and the origin of life. Geochimica et Cosmochimica Acta. 2019;260:124-32. [CrossRef]
- Sutherland JD. The origin of life—out of the blue. Angewandte Chemie International Edition. 2016;55(1):104-21. [CrossRef]
- Kingsolver J, Pfennig. Testing Darwin’s postulates. Trends in Ecology & Evolution. 2001;16(3). [CrossRef]
- Baum DA, Peng Z, Dolson E, Smith E, Plum AM, Gagrani P. The ecology-evolution continuum and the origin of life. J R Soc Interface. 2023;20(208):20230346. [CrossRef]
- Garrett C. Internal tides and ocean mixing. Science. 2003;301(5641):1858-9. [CrossRef]
- Mat AM, Sarrazin J, Markov GV, Apremont V, Dubreuil C, Eche C, et al. Biological rhythms in the deep-sea hydrothermal mussel Bathymodiolus azoricus. Nat Commun. 2020;11(1):3454. [CrossRef]
- Bonfio C, Valer L, Scintilla S, Shah S, Evans DJ, Jin L, et al. UV-light-driven prebiotic synthesis of iron-sulfur clusters. Nat Chem. 2017;9(12):1229-34. [CrossRef]
- Green NJ, Xu J, Sutherland JD. Illuminating Life’s Origins: UV Photochemistry in Abiotic Synthesis of Biomolecules. J Am Chem Soc. 2021;143(19):7219-36. [CrossRef]
- Whitaker DE CB, Powner MW. Nucleotide photochemistry on the early earth. In: Smoukov SK SJ, Gordon R, editor. Conflicting Models for the Origin of Life: Scrivener Publishing LLC; 2023. p. 369-405. [CrossRef]
- Nunes Palmeira R, Colnaghi M, Harrison SA, Pomiankowski A, Lane N. The limits of metabolic heredity in protocells. Proc Biol Sci. 2022;289(1986):20221469. [CrossRef]
- Ranjan S, Kufner CL, Lozano GG, Todd ZR, Haseki A, Sasselov DD. UV Transmission in Natural Waters on Prebiotic Earth. Astrobiology. 2022;22(3):242-62. [CrossRef]
- Mulkidjanian AY, Junge, W. Primordial UV-Protectors As Ancestors of the Photosynthetic Pigment-Proteins. . In: Peschek GA, Löffelhardt, W., Schmetterer, G. , editor. The Phototrophic Prokaryotes. Boston MA: Springer; 1999. [CrossRef]
- Mulkidjanian AY, Junge, W. On the origin of photosynthesis as inferred from sequence analysis. Photosynthesis Research. 1997;51:27-42. [CrossRef]
- Zhang K, Hodge J, Chatterjee A, Moon TS, Parker KM. Duplex Structure of Double-Stranded RNA Provides Stability against Hydrolysis Relative to Single-Stranded RNA. Environ Sci Technol. 2021;55(12):8045-53. [CrossRef]
- Mateus A, Bobonis J, Kurzawa N, Stein F, Helm D, Hevler J, et al. Thermal proteome profiling in bacteria: probing protein state in vivo. Mol Syst Biol. 2018;14(7):e8242. [CrossRef]
- Majidi D A, N. Stability of Tyrosinase Enzyme from Funalia Trogii American Journal of Microbiological Research. 2013;1(1):1-3. [CrossRef]
- Baluska F, Reber AS. CBC-Clock Theory of Life - Integration of cellular circadian clocks and cellular sentience is essential for cognitive basis of life. Bioessays. 2021;43(10):e2100121. [CrossRef]
- Kim YH, Lazar MA. Transcriptional Control of Circadian Rhythms and Metabolism: A Matter of Time and Space. Endocr Rev. 2020;41(5):707-32. [CrossRef]
- Muller F, Escobar L, Xu F, Wegrzyn E, Nainyte M, Amatov T, et al. A prebiotically plausible scenario of an RNA-peptide world. Nature. 2022;605(7909):279-84. [CrossRef]
- Wayment-Steele HK, Kim DS, Choe CA, Nicol JJ, Wellington-Oguri R, Watkins AM, et al. Theoretical basis for stabilizing messenger RNA through secondary structure design. Nucleic Acids Res. 2021;49(18):10604-17. [CrossRef]
- Netzer A, Katzir I, Baruch Leshem A, Weitman M, Lampel A. Emergent properties of melanin-inspired peptide/RNA condensates. Proc Natl Acad Sci U S A. 2023;120(44):e2310569120. [CrossRef]
- Miller SL. The atmosphere of the primitive Earth and the prebiotic synthesis of amino acids. Origins Life Evol Biosphere 1974;5(39):139-51. [CrossRef]
- Raposo G, Marks MS. Melanosomes--dark organelles enlighten endosomal membrane transport. Nat Rev Mol Cell Biol. 2007;8(10):786-97. [CrossRef]
- Hurbain I, Geerts WJ, Boudier T, Marco S, Verkleij AJ, Marks MS, et al. Electron tomography of early melanosomes: implications for melanogenesis and the generation of fibrillar amyloid sheets. Proc Natl Acad Sci U S A. 2008;105(50):19726-31. [CrossRef]
- Bush WD, Simon JD. Quantification of Ca(2+) binding to melanin supports the hypothesis that melanosomes serve a functional role in regulating calcium homeostasis. Pigment Cell Res. 2007;20(2):134-9. [CrossRef]
- Singh S, Malhotra AG, Pandey A, Pandey KM. Computational model for pathway reconstruction to unravel the evolutionary significance of melanin synthesis. Bioinformation. 2013;9(2):94-100. [CrossRef]
- Vinther J. A guide to the field of palaeo colour: Melanin and other pigments can fossilise: Reconstructing colour patterns from ancient organisms can give new insights to ecology and behaviour. Bioessays. 2015;37(6):643-56. [CrossRef]
- Wiriyasermkul P, Moriyama S, Nagamori S. Membrane transport proteins in melanosomes: Regulation of ions for pigmentation. Biochim Biophys Acta Biomembr. 2020;1862(12):183318. [CrossRef]
- Ambrosio AL, Boyle JA, Aradi AE, Christian KA, Di Pietro SM. TPC2 controls pigmentation by regulating melanosome pH and size. Proc Natl Acad Sci U S A. 2016;113(20):5622-7. [CrossRef]
- Guppy M, Kong SE, Niu X, Busfield S, Klinken SP. Method for measuring a comprehensive energy budget in a proliferating cell system over multiple cell cycles. J Cell Physiol. 1997;170(1):1-7. [CrossRef]
- Hansma HG. Potassium at the Origins of Life: Did Biology Emerge from Biotite in Micaceous Clay? Life (Basel). 2022;12(2). [CrossRef]
- Staley JT. The gas vacuole: An early organelle of prokaryote motility? Discover Life. 1980;10(111-116). [CrossRef]
- Naranjo D. A scenario for the origin of life: Volume regulation by bacteriorhodopsin required extremely voltage sensitive Na-channels and very selective K-channels. Bioessays. 2022;44(10):e2100210. [CrossRef]
- Sephus CD, Fer E, Garcia AK, Adam ZR, Schwieterman EW, Kacar B. Earliest Photic Zone Niches Probed by Ancestral Microbial Rhodopsins. Mol Biol Evol. 2022;39(5). [CrossRef]
- Morowitz HJ. Beginnings of cellular life : metabolism recapitulates biogenesis. New Haven: Yale University Press; 1992. x, 195 pp. ISBN 0-300-05483-1 cloth.
- Tosteson DC, Hoffman JF. Regulation of cell volume by active cation transport in high and low potassium sheep red cells. J Gen Physiol. 1960;44(1):169-94. [CrossRef]
- Trapp O. Origins of life research: A roadmap for the transition from chemistry to biology. Bioessays. 2022;44(10):e2200157. [CrossRef]
- Glavin DP, Burton AS, Elsila JE, Aponte JC, Dworkin JP. The Search for Chiral Asymmetry as a Potential Biosignature in our Solar System. Chem Rev. 2020;120(11):4660-89. [CrossRef]
- Frank FC. On spontaneous asymmetric synthesis. Biochim Biophys Acta. 1953;11(4):459-63. [CrossRef]
- Blackmond DG. The origin of biological homochirality. Cold Spring Harb Perspect Biol. 2010;2(5):a002147. [CrossRef]
- Sallembien Q, Bouteiller L, Crassous J, Raynal M. Possible chemical and physical scenarios towards biological homochirality. Chem Soc Rev. 2022;51(9):3436-76. [CrossRef]
- Bechtel M, Ebeling M, Huber L, Trapp O. (Photoredox) Organocatalysis in the Emergence of Life: Discovery, Applications, and Molecular Evolution. Acc Chem Res. 2023;56(20):2801-13. [CrossRef]
- Nicewicz DA, MacMillan DW. Merging photoredox catalysis with organocatalysis: the direct asymmetric alkylation of aldehydes. Science. 2008;322(5898):77-80. [CrossRef]
- Subramanian H, Gatenby RA. Chiral Monomers Ensure Orientational Specificity of Monomer Binding During Polymer Self-Replication. J Mol Evol. 2018;86(5):255-63.
- Burke JM, Kaczara P, Skumatz CM, Zareba M, Raciti MW, Sarna T. Dynamic analyses reveal cytoprotection by RPE melanosomes against non-photic stress. Mol Vis. 2011;17:2864-77. [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).