
Submitted:
15 November 2024
Posted:
15 November 2024
You are already at the latest version
Abstract
The evaluation of the broad variety of allelochemicals present in allelopathic plants allows detecting safer bioherbicides with new mechanisms of action. This study tested two phenolic compounds of Cistus ladanifer essential oil (2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol), which are commercially available. At 0.01mM, these compounds, both separately and in combination, inhibited up to over 50% of germination, cotyledon emergence and seedling growth of Lactuca sativa for the tests conducted on paper. Against Allium cepa, cotyledon emergence and seedling growth was inhibited at 0.5mM. When the tests were carried out in soil, the mixture of the two study compounds significantly inhibited the germination of L. sativa and A. cepa when applied at 0.5 and 1mM, respectively, and seedling growth inhibition was greater for the latter in the paper tests. The greatest inhibitions were observed with the highest concentrations analysed. Although there was no statistically significant difference among treatments, 2-Methoxyphenol seemed to affect germination and cotyledon emergence to a greater extent, whereas 2,6-Di-tert-butyl-4-methylphenol had a greater impact on seedling size. The effect of the mixture was greater than that of both compounds separately.
Keywords:
phytotoxicity
; phenolic compounds
; allelopathy
; bioherbicides
; essential oil
1. Introduction
This is an age in the geological and biological history of Earth in which the signs of human action are evident all over the planet and, in this scenario, agriculture faces the challenge of feeding a population in constant growth. Traditional agricultural practices, such as the extensive use of herbicides, have been effective in terms of productivity; however, they have contributed significantly to the emergence of resistant weeds and environmental deterioration through the loss of biodiversity and soil contamination [1].
The proliferation of weeds that compete for nutrients reduces crop yield, affects agricultural infrastructure (e.g., water channeling systems), and hinders the harvest and commercialization processes, which requires an active search for more natural alternatives to the use of synthetic herbicides.
Since antiquity, different studies have shown the harmful effects of certain plants on the growth of other plants, as well as their interaction with microorganisms. This interaction among plants mediated by secondary metabolites is known as allelopathy [2]. Allelopathy has gained interest in agricultural systems, with research focusing on the interference between crop plants and weeds to regulate the latter. Different practices are being implemented, such as allelopathic topsoil, cultural management and natural herbicides, with the aim of improving weed control. However, despite the fact that ecological management and restoration methods are improving, as a result of exhaustive research on allelopathy, this phenomenon continues to be underused in modern agricultural practices [3].
A better understanding of the types of allelopathic substances produced by plants, as well as their purposes and mechanisms of action, offers promising solutions for weed management, crop protection and environmental conservation [4]. It would be ideal to obtain and employ natural, water-soluble compounds with potential herbicide properties at low concentration, low molecular weight and simple formulae, which would facilitate their extraction or synthesis and application; in addition, they should be species-specific and biodegradable and present low toxicity, thereby being safer for humans, animals and the environment. Most phenolic compounds have several of these properties, which make them promising candidates for the development of bioherbicides. Phenolic acids are common secondary metabolites that can act as allelochemicals and suppress the germination and growth of competing plants in the soil [5]. Despite their broad variety and the evidence on the molecular mechanisms of many of them [6-8], most studies are focused on allelopathic compounds of extracts, with a lack of information about the phytotoxicity of many species [9]. Some of these compounds are volatile and can be concentrated by essential oil extraction. Some essential oils, such as those of Cinnamomum zeylanicum, Thymus vulgaris, Eucalyptus globulus, Rosmarinus officinalis and Lavandula angustifolia, have been proved to significantly inhibit the germination and growth of several species of common and problematic weeds [10-12]. These results suggest that essential oils may be as effective as some synthetic herbicides, with the additional benefit of being biodegradable and less toxic for the environment. Evaluating the activity of all phenolic compounds would allow finding compounds with new mechanisms of action and uncharted places of action that could lead to solving the problem of weed resistance to conventional herbicides [13].
Cistus ladanifer Linneo (1753) is a typical allelopathic species of the Mediterranean ecosystem. When the labdanum tree is exploited, cutting it to extract its essential oil or labdanum, plant richness increases immediately in the area, although it decreases in time when the population of labdanum tree regenerates [14]. Several studies indicate that this reduction of biodiversity is caused by the allelopathic potential of this species [15-18], whose essential oil also presents phytotoxic activity [19,20]. The labdanum and essential oil of its leaves contain a high concentration of phenolic compounds, some of which have not been studied to date [21-28].
In this context, the current study aims to contribute to understanding the allelopathic potential of Cistus ladanifer and to identifying possible structures that may serve as sources of safer and less polluting natural herbicides. To this end, we evaluated the phytotoxic activity of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol, which are two phenolic compounds present in C. ladanifer essential oil that have not been studied to date [17,18,29,30]. 2-Methoxyphenol or Guaiacol (Figure 1A) exists naturally in guaiac wood resin and essential oil. It is used as a non-stick agent in surface coating, a flavouring agent, and an antioxidant (for fats, oils and vitamins), and with medicinal purposes as an expectorant, an antiseptic and a local anesthetic [31]. On the other hand, 2,6-Di-tert-butyl-4-methylphenol or Butylhydroxytoluene (Figure 1B) is found in its natural form in the lychee fruit. It is used as a food additive (E-321), and an antioxidant in cosmetics, pharmaceuticals, airplane fuels, rubber, petroleum derivatives, electrical transformer oil, and embalming fluid [32].
The phytotoxicity of phenolic compounds may vary considerably as a function of their chemical structure, the concentration applied, the presence of other compounds and the species and substrate used [6,18,21,33-37]. Therefore, this study proposes a bioassay that contemplates all these variables, using commercial seeds of Lactuca sativa (dicotyledonous) and Allium cepa (monocotyledonous), which are characterized by their uniform sensitivity and response capacity in phytotoxic tests [6, 38-40]. Whatman filter paper and commercial substrate were employed to verify whether the activity of the compounds was affected by the physical-chemical and biological conditions of the soil. The compounds were applied separately and as a mixture, in order to detect possible interactions, and at low concentrations (0.01 to 1mM) and in a sufficiently wide range to identify possible hormetic effects [41]. The effects of these compounds were analyzed in terms of germination, cotyledon emergence and seedling growth to determine their viability as natural herbicides.
2. Results
2.1. Effect of 2-Methoxyphenol and 2,6-Di-Tert-Butyl-4-Methylphenol on the Germination of Lactuca sativa L.
In the tests conducted on paper, 2-Methoxyphenol, 2,6-Di-tert-butyl-4-methylphenol and the mixture of both compounds inhibited the total germination (%Gt) and germination rate (%GR) of Lactuca sativa at the four concentrations analysed, with this inhibition being up to 75% for the highest concentration (Figure 2). However, in the tests performed in soil, although %GR was inhibited with all treatments, %Gt was only inhibited when the compounds were applied as a mixture at 0.5 and 1mM.
It is worth highlighting that the greatest effects were observed at the highest concentrations. The effect of the mixture of compounds on %Gt and %GR showed a significant positive linear correlation with concentration (R2=0.98 and R2=0.91, respectively), i.e., the higher the concentration, the greater the inhibition. The half-maximal inhibitory concentration (IC50) was 0.5mM and 0.01mM, respectively (Figure 2). Moreover, the inhibitory effect was greater when the compounds were applied as a mixture with respect to the effect of the separate compounds. This difference was statistically significant at 1mM for %Gt, %GR on paper and at 0.5 and 1mM for %Gt in soil.
It is also important to point out that, on paper, the effect of 2-Methoxyphenol at 1mM on %GR was similar to that of the mixture and significantly greater than that of 2,6-Di-tert-butyl-4-methylphenol.
2.2. Effect of 2-Methoxyphenol and 2,6-Di-Tert-Butyl-4-Methylphenol on the Cotyledon Emergence of Lactuca sativa L.
The analysed compounds presented the same behaviour on total cotyledon emergence (%Ct) and cotyledon emergence rate (%CR) as on %Gt and %GR, although inhibition was greater in all analysed concentrations both on paper and in soil. On paper, %Ct and %CR were strongly inhibited up to over 75% at the highest concentrations, whereas the effect was lower in soil; however, in the latter case, both the separate compounds and their mixture, in addition to inhibiting cotyledon emergence rate, also inhibited %Ct at all analysed concentrations. Furthermore, it was observed that, at any of the applied concentrations, 2-Methoxyphenol delayed cotyledon emergence to a significantly greater extent compared to 2,6-Di-tert-butyl-4-methylphenol (Figure 3).
Similarly, the greatest inhibitory effects were observed at the highest concentrations, and the effect of the mixture of compounds on %Ct and %CR showed a significant positive correlation with concentration (R2=0.97 and R2=0.99, respectively). IC50 was lower than 0.01mM in both cases, and, when the mixture was applied at 1mM in soil, the inhibitory effect on %Ct and %CR was significantly greater than that of the separate compounds (Figure 3).
2.3. Effect of 2-Methoxyphenol and 2,6-Di-Tert-Butyl-4-Methylphenol on The Seedling Growth of Lactuca sativa L.
2-Methoxyphenol, 2,6-Di-tert-butyl-4-methylphenol and the mixture of both phenols also significantly inhibited the seedling growth of Lactuca sativa at all analysed concentrations in the paper tests. 2,6-Di-tert-butyl-4-methylphenol inhibited radicle length (%Radicle length) and hypocotyl length (%Hypocotyl length) to a greater extent than 2-Methoxyphenol when applied at 0.5 and 1mM (Figure 4).
In soil, the effects of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol were low, with only 2-Methoxyphenol significantly inhibiting radicle and hypocotyl growth at 1mM. Moreover, 2,6-Di-tert-butyl-4-methylphenol significantly stimulated hypocotyl growth at 0.01 and 0.1mM. On its part, the mixture also stimulated hypocotyl growth at 0.1mM, although it inhibited %Radicle length and %Hypocotyl length at 0.5 and 1mM (Figure 4).
Both in soil and on paper, the greatest inhibitory effects on seedling growth were observed at the highest concentrations. On paper, the effect of the mixture of compounds on %Radicle length and %Hypocotyl length showed a significant logarithmic correlation with concentration (R2=0.97 and R2=0.91, respectively). IC50 was lower than 0.1mM in both cases. It is also important to highlight that, when the mixed compounds were applied at 0.5mM in soil, radicle and hypocotyl growth was inhibited, whereas the separate compounds at this concentration did not show a significant difference with the control (Figure 4).
2.4. Effect of 2-Methoxyphenol and 2,6-Di-Tert-Butyl-4-Methylphenol on the Germination of Allium cepa L.
The paper test with seeds of Allium cepa revealed that 2-Methoxyphenol and the mixture of the two compounds inhibited neither total germination (%Gt) nor germination rate (%GR) at any of the four analysed concentrations. Only 2,6-Di-tert-butyl-4-methylphenol at 1mM produced significantly lower values of %Gt and %GR with respect to the control (Figure 5). The soil test showed that 2,6-Di-tert-butyl-4-methylphenol inhibited %Gt and %GR at low concentrations. In turn, as was expected, 2-Methoxyphenol and the mixture of both compounds inhibited %Gt and %GR at high concentrations (0.5 and 1 mM). It is worth pointing out that the mixture applied at 1mM in soil inhibited %Gt, whereas the separate compounds at the same concentration did not have an inhibitory effect, and the mixture applied at 0.5mM inhibited %GR, while the separate compounds at the same concentration did not have an inhibitory effect either (Figure 5).
2.5. Effect of 2-Methoxyphenol and 2,6-Di-Tert-Butyl-4-Methylphenol on the Cotyledon Emergence of Allium cepa L.
On paper, 2-Methoxyphenol inhibited total cotyledon emergence (%Ct) from a concentration of 0.1mM and cotyledon emergence rate (%CR) at all four analysed concentrations. In soil, although %Ct was only inhibited at 1mM, the effect on %CR was 20% greater at all concentrations (Figure 6).
On paper, 2,6-Di-tert-butyl-4-methylphenol only inhibited %Ct at 1mM without significantly affecting %Ct; however, in soil, this compound inhibited %Ct at 0.01mM and %CR at all concentrations (Figure 6).
Similarly, the mixture of both phenolic compounds also presented a greater inhibitory effect in soil. On paper, the mixture inhibited %CR at 0.5 and 1mM, whereas, in soil, it inhibited %CR at all analysed concentrations (Figure 6).
For all paper treatments, the greatest inhibitory effects were observed at the highest concentrations, which was not observed in any case in the soil tests.
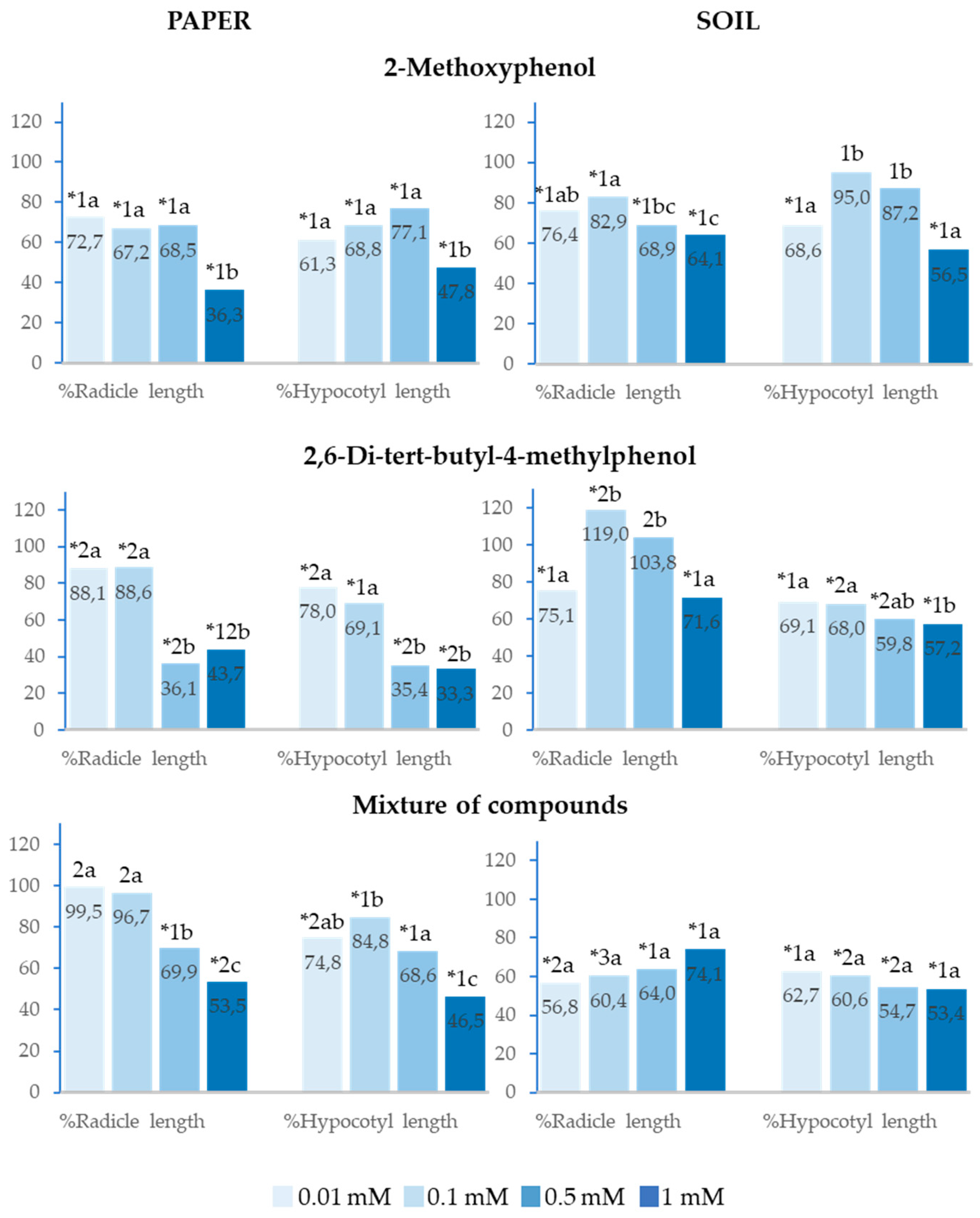
2.6. Effect of 2-Methoxyphenol and 2,6-Di-Tert-Butyl-4-Methylphenol on the Seedling Growth of Allium cepa L.
2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two phenols significantly inhibited the %Radicle length and %Hypocotyl length of the seedlings of Allium cepa at all analysed concentrations when applied both on paper and in soil, except for %Hypocotyl length at 0.1 and 0.5mM when 2-Methoxyphenol was applied, %Radicle length at 0.1 and 0.5mM when 2,6-Di-tert-butyl-4-methylphenol was applied in soil, and %Radicle length at 0.01 and 0.1mM when the mixture was applied on paper (Figure 7).
It is important to underline that, when the mixed compounds were applied in soil, radicle and cotyledon growth was inhibited to a greater extent than when the compounds were applied separately (Figure 7).
3. Discussion
The repeated and frequent use of non-selective herbicides, such as glyphosate, has led to environmental health problems and the appearance of resistant weeds. In the face of possible prohibitions by institutions that advocate for the change toward more sustainable practices and safer agriculture, it is necessary to search for more natural products with new mechanisms of action that decompose rapidly in soil and that can be used in ecological agriculture.
Essential oils of allelopathic plants may contain allelochemicals that inhibit germination [42], apparently by damaging meristematic cells [43]. Essential oils are classified as "generally regarded as safe" (GRAS) and can be used as a viable technology for the control of weeds in organic agricultural systems, although it is necessary to obtain basic information about their compounds and the phytotoxic potential of each of these compounds. An important group which the phytotoxic activity of essential oils has been attributed to is constituted by phenolic compounds [42,43].
After determining which phenolic compounds that are present in Cistus ladanifer essential oil are commercially available and have not been evaluated as possible strong herbicides, the objectives of this study were to: 1) analyse the phytotoxic activity of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol against monocotyledons and dicotyledons; 2) determine whether the activity of these compounds is maintained when they are applied in soil; and 3) identify interactions among these compounds that may increase or reduce their effects. To this end, we conducted standardised bioassays with low concentrations (0.01 to 1mM) of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol, separately and as a mixture, against seeds of Lactuca sativa and Allium cepa. The results showed that these phenolic compounds, as others that have been previously studied, presented phytotoxic activity to a greater or lesser extent depending on the substrate, the species and the concentration analysed [36,41,44].
For the tests conducted on paper with seeds of Lactuca sativa, 2-Methoxyphenol, 2,6-Di-tert-butyl-4-methylphenol and the mixture of both compounds not only delayed cotyledon germination and emergence significantly, but they also directly inhibited germination, cotyledon emergence and seedling growth at all concentrations analysed. The fact that these compounds showed up to over 50% inhibitory activity at very low concentrations (0.01mM) suggests that they could be considered as candidates for possible sources of bioherbicides. In general, high concentrations of phenols tend to inhibit germination and seedling growth, whereas low doses may stimulate these processes through the phenomenon known as hormesis [45-46]. This study was focused on assays with very low concentrations, although the greatest inhibitory effects were obtained with the highest concentration (1mM) and, when the mixture was applied, there was a dependence with the dose in all indexes measured, detecting neither hormetic nor synergistic effects. Pearson’s correlation analysis showed that R2>0.95, calculating an IC50 of 0.5mM for %Gt, 0.01mM for %GR, %Radicle length and %Hypocotyl length, and less than 0.01mM for %Ct and %CR. The fact that the effect depends on concentration is further evidence of the phytotoxic potential of phenolic compounds. This finding of allelochemicals with the characteristic of presenting phytotoxic activity at very low concentrations is significant for ensuring the effectiveness of the product and minimising the costs and environmental impact in a possible application.
Some species may be sensitive to certain compounds, whereas others may show greater resistance. When the assays were carried out on paper with seeds of Allium cepa, the effects of 2-Methoxyphenol, 2,6-Di-tert-butyl-4-methylphenol and the mixture of both were lower than those observed in Lactuca sativa. Against this monocotyledonous species, none of the treatments had a significant effect on germination or %GR. Cotyledon emergence and %CR were only inhibited at the highest concentrations, and, although the different solutions significantly inhibited seedling growth, the effects were lower than in the assays with Lactuca sativa. This delay in cotyledon emergence and the inhibition of radicle and hypocotyl growth may be as detrimental for the establishment of the seedling as the inhibition of germination [48]. Phenolic acids increase the activity of phenylalanine ammonia-lyase (PAL) and β-glucosidase, whereas they also reduce the activity of phenol-β-glucosyltransferase, inhibiting radicle growth. They can also affect DNA and RNA integrity and inhibit protein synthesis [49]. Some phenolic acids, such as benzoic acid and cinnamic acid, reduce the content of chlorophyll, thereby inhibiting photosynthesis [50]. Other phenolic acids reduce the growth of receptive plants through the alteration of hormones [51-53], membranes [52,54-56] and respiration [55,57]. Other studies show that phenolic compounds, such as chlorogenic acid, caffeic acid and catechol inhibit germination by altering the functionality of λ-phosphorylase, which is a key enzyme in the process [2,55]. This enzyme is present in both monocotyledons and dicotyledons, thus it is deduced that, in this case, the mechanism of action of these compounds could be different, making them more specific to dicotyledons. Understanding these mechanisms is essential to optimise the application of these compounds in crop management [58]. This specificity could be used to develop more specific and effective weed control strategies [59].
The assays conducted with different substrates showed that the efficacy of phenols can be affected by environmental factors. When the treatments were carried out in universal substrate, the inhibitory effects on %Gt were lower, and only the mixture of phenolic compounds significantly inhibited the germination of lettuce and onion seeds at 0.5 and 1mM, respectively. Microbial degradation affects the final destination and concentration of allelochemical substances in the surrounding soil and their activity [60-63]. Isolated microorganisms from a hydroponic lettuce crop supplied with ferulic acid degraded this allelochemical compound, reducing its phytotoxic effects [64]. This would explain the lower effects observed when the analysed solutions were applied in soil. The rest of the indexes measured in soil were also lower than on paper with Lactuca sativa seeds; however, a different behaviour was detected in Allium cepa, observing that, when any of the solutions were applied in this substrate, the inhibition of %CR in A. cepa was greater than on paper. The same occurred with %Radicle length and %Hypocotyl length; when the mixture was applied in soil, the inhibition of seedling growth in A. cepa was greater than on paper. Once released, allelochemicals accumulate in the soil [37] and endure the processes of the latter, such as retention, transport and transformation [65]. These processes may be different for each compound. Due to its physicochemical characteristics, 2-Methoxyphenol is expected to volatilise in humid soil and show very high mobility and low absorption; in the atmosphere, this phenol is expected to be susceptible to solar photolysis and to have a half-life of few hours [31]. In turn, 2,6-Di-tert-butyl-4-methylphenol is expected to be immobile in soil and to degrade in a few days, as well as to be unstable in water and to be susceptible to photolysis [32]. Moreover, transformation or biodegradation may lead to other compounds that are more or less active with a recipient plant, which could explain the difference found among species. 2-Methoxyphenol can lose its methoxy group (-OCH₃), thereby becoming a catechol (1,2-dihydroxybencene), which becomes 1,2-benzoquinone. In its decomposition, 2,6-Di-tert-butyl-4-methylphenol could lead to more simple phenolic compounds and benzoquinones. Catechol, as was previously mentioned, may alter the functioning of essential enzymes and proteins for photosynthesis and respiration, by reacting with their functional groups [2,66]. Some quinones, such as sorghum and juglone, also affect plant growth, as they prevent photosynthesis and cell respiration, by decoupling electron transport in mitochondria and chloroplasts [67]. Nowadays, there are herbicides based on these types of synthetic compounds. Thus, the differences between monocotyledonous and dicotyledonous in radicle structure, cell wall composition, metabolism and stress response lead to variations in the degree and type of damage of the assayed allelochemicals and their degradation products generated in the soil.
Comparing the activity between the analysed phenols, there was no clear difference in their behaviour, although we identified some significant differences between them. On paper, against both lettuce and onion, 2-Methoxyphenol had a significantly greater effect on %GR and %Ct, whereas 2,6-Di-tert-butyl-4-methylphenol inhibited radicle and hypocotyl length to a greater extent at 0.5mM. In soil, 2-Methoxyphenol also showed a greater effect than 2,6-Di-tert-butyl-4-methylphenol on %Ct, %CR and seedling growth of Lactuca sativa, whereas in Allium cepa, 2,6-Di-tert-butyl-4-methylphenol presented a greater effect on the measured indexes. This confirms not only that the effects depend on the compounds and the concentration applied, but also that their efficacy can be affected by different environmental factors that may influence chemical degradation or transformation, as well as the influence of microorganisms and the physicochemical properties of the substrate.
It is worth highlighting that, when the mixture of the two compounds was applied at 1mM in soil, the germination of Lactuca sativa and Allium cepa was inhibited, whereas their separate application at this concentration did not show significant differences with respect to the control. Furthermore, %Ct, %CR and hypocotyl length of lettuce was significantly lower with the mixture of phenols compared to the separate phenols. In addition, when the mixture was applied at 0.1mM on onion seeds, cotyledon emergence and radicle length were significantly inhibited, which was not observed with the separate phenols. This confirms the interaction of the compounds. The allelopathic capacity of a species often results from the collective or synergistic response of several allelochemicals rather than from a single compound [47].
To sum up, 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol presented phytotoxic properties that may interfere with the germination, cotyledon emergence and seedling growth of monocotyledonous and dicotyledonous plants (Allium cepa and Lactuca sativa). The tests conducted at different concentrations and in different substrates revealed that the efficacy of these phenols may be particularly affected by the conditions of the assay. The chemical formula of the compound could be key to its effectiveness against a target species, although this aspect may be less relevant among compounds of the same chemical group if soil degradation leads to the same products. Factors such as concentration can also be an essential aspect for the activity of the compound, which could range from inhibition to stimulation. Due to biodegradation processes, activity in soil is expected to be lower than on paper; however, the chemical transformation of some compounds may lead to other products that are more phytotoxic than the precursor. Another condition that must be considered is the combination of the different compounds that may be found in the substrate, which often results in greater phytotoxic activity compared to that of isolated compounds. Lastly, compounds can show different effects when they interact with different species. Making use of these characteristics, we can develop more selective and effective formulae at lower concentrations, thereby minimising the negative environmental impacts. The comparison of these mechanisms is crucial to optimise their possible application in crop management.
Therefore, it is concluded that, in the current situation, it is urgent to search for safe, natural compounds with herbicide properties that minimise the environmental impact when applied to crops in weed control. Essential oils of allelopathic plants show great potential as a source of natural herbicides [43]. Some of their components, such as phenolic compounds, present unique properties that make them especially appropriate candidates. This work not only contributes to understanding the herbicide activity of two phenolic compounds present in Cistus ladanifer essential oil, but it also offers guidelines on the methodology to follow in the study of these natural products.
4. Materials and Methods
4.1. Reagents, Seeds and Substrates
Reagents of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol with a purity of >98% were acquired from Aldrich-Chemical to prepare the solutions of 0.01mM, 0.1mM, 0.5mM and 0.1mM of each of the compound separately. For the mixture of the two phenols, four solutions of these four concentrations were prepared with equimolar concentrations of each of the compounds. The pH varied between 6.1 and 6.3 from one solution to another. There were no significant differences in pH among solutions.
We acquired commercial seeds of Lactuca sativa, romaine verte maraîchère variety (Vilmorin jardin—CS70110—38291 St Quentin Fallavier Cedex—France), as a representative of dicotyledons, and seeds of Allium cepa, Bianca di Maggio variety (Vilmorin Jardin—CS70110—38291 St Quentin Fallavier Cedex—France), as a representative of monocotyledons. These species are the ideal for phytotoxicity bioassays [68] and presented a total germination of over 98% after being watered with distilled water.
The substrates used were Whatman No. 118 paper and substrate of universal type, based on 95% peat, 5% green compost, and 1.3 Kg/m3 fertiliser: 12N + 12P + 17K (Geolia, Aki Brico-lage España S.L. B-839857—Spain). The commercial universal soil presented the following characteristics: organic matter per dry matter (60%), electrical conductivity (40 mS/m), apparent dry density (320 g/L), grain size (0–20 mm), and pH 5.5–6.5.
None of the materials (seeds and substrate) were sterilised before the experiment.
4.2. Phytotoxicity Bioassay
To evaluate the phytotoxic activity of the selected phenolic compounds, a standardised test was followed [39,69]. In Petri dishes, 50 seeds of Lactuca sativa or Allium cepa were placed on paper or on 25g of universal substrate, and they were watered with Milli-Q water for the controls and with different solutions for the treatments (5ml for paper and 16ml for soil). Four replicates for each treatment were sealed with Parafilm to prevent evaporation for 5 and 6 days for the seeds of L. sativa and A. cepa, respectively, in a germination chamber at 15h of light and 9h of darkness at a constant temperature of 22 ºC.
4.3. Measured Indexes to Quantify the Phytotoxic Effect
Total germination (%Gt) and total cotyledon emergence (%Ct): On the last day of the experiment, the number of germinated seeds and the number of germinated seedlings with cotyledons were counted. With these data, we calculated the mean values of the four replicates, which are expressed as percentage relative to the control [36,41,44].
Germination rate (%GR) and cotyledon emergence rate (%CR): In order to measure a possible delay in germination and cotyledon emergence, we counted the number of germinated seeds and cotyledons that appeared every day, and the indexes were calculated using the following equation (1):
Radicle growth (%Radicle length) and hypocotyl growth (%Hypocotyl length): To determine a possible alteration of growth, on the last day, 10 seedlings selected at random from each Petri dish were measured, and we calculated the mean values, which were expressed as percentage relative to the control [70].
4.4. Statistical Analysis
The significance level of the comparisons among treatments was estimated using the Mann-Whitney U test. The differences were considered significant when p<0.05. The interrelationships between germination and seed growth with the concentration of phenolic compounds were determined by Pearson′s determination coefficient. The effective concentrations required to induce half-maximal inhibition of growth (IC50) were calculated according to the linear or logarithmic relationship between concentration and percent inhibition of plant growth. All statistical analyses were conducted using the statistical software IBM SPSS Statistics version 26.
Author Contributions
Conceptualization, T.S.D.; methodology, D.O.D; validation, T.S.D., D.F.F. and A.M.S.; formal analysis, D.O.D; investigation, D.O.D and T.S.D.; resources, T.S.D. and D.F.F.; data curation, A.M.S.; writing—original draft preparation, T.S.D.; writing—review and editing, D.O.D; visualization, D.F.F. and A.M.S.; supervision, D.F.F and A.M.S.; project administration, T.S.D.; funding acquisition, T.S.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Regional Government of Extremadura and the European Regional Development Fund and the project PID2020-116144RB-I00/AEI/10.13039/501100011033.IB18105.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic Interactions and Allelochemicals: New Possibilities for Sustainable Weed Management. Crit. Rev. Plant Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy; Academic Press: Orlando, FL, USA, 1984; p. 422. [Google Scholar]
- Cheng, F.; Cheng, Z. Research Progress on the Use of Plant Allelopathy in Agriculture and the Physiological and Ecological 678 Mechanisms of Allelopathy. Front. Plant Sci. 2015, 6, 160714. [Google Scholar] [CrossRef]
- Blanco, Y. La utilización de la alelopatía y sus efectos en diferentes cultivos agrícolas. Cultiv. Trop. 2006, 27, 5–16. [Google Scholar]
- Kong, C.-H.; Li, Z.; Li, F.-L.; Xia, X.-X.; Wang, P. Chemically Mediated Plant–Plant Interactions: Allelopathy and Allelobiosis. Plants 2024, 13, 626. [Google Scholar] [CrossRef] [PubMed]
- Einhellig, F.A. Mode of Allelochemical Action of Phenolic Compounds. In Allelopathy: Chemistry and mode of action of allelochemicals; Macías, F.A., Galindo, J.C.G., Molinillo, J.M.G.V., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 217–238. [Google Scholar] [CrossRef]
- Inderjit. Plant Phenolics in Allelopathy. Bot. Rev. 1996, 62, 186–202. [Google Scholar] [CrossRef]
- Kuiters, A.T. Effects of phenolic acids on germination and early growth of herbaceous woodland plants. J. Chem. Ecol. 1989, 15, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Al Harun, M.A.; Johnson, J.; Uddin, MN.; Robinson, R.W. Identification and Phytotoxicity Assessment of Phenolic Compounds in Chrysanthemoides monilifera subsp. monilifera (Boneseed). PLoS One 2015, 10, e0139992. [Google Scholar] [CrossRef] [PubMed]
- Tworkoski, T. Herbicide effects of essential oils. Weed Sci. 2002, 50, 425–431. [Google Scholar] [CrossRef]
- Vokou, D.; Douvli, P.; Blionis, G.J.; Halley, J.M. Effects of monoterpenoids, acting alone or in pairs, on seed germination and subsequent seedling growth. J. Chem. Ecol. 2003, 29, 2281–2301. [Google Scholar] [CrossRef]
- Angelini, L.G.; Carpanese, G.; Cioni, P.L.; Morelli, I.; Macchia, M.; Flamini, G. Essential oils from Mediterranean Lamiaceae as weed germination inhibitors. J. Agric. Food Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Molinillo, J.M.; Varela, R.M.; Galindo, J.C. Allelopathy--a natural alternative for weed control. Pest Manag Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Márquez-García, F.; García-Alonso, D.; Vázquez, F.M. Notes of Changes in Biodiversity in the Exploited Populations of Cistus ladanifer, L., (Cistaceae) from SW Iberian Peninsula. J. Fungi. 2022, 2, 1–7. [Google Scholar]
- Agra-Coelho, C.; Rosa, M.L.; Moreira, I. Allelopathic effects of Cistus ladanifer L. III Natl. Symp. Herbol. 1980, 1, 165–178. [Google Scholar]
- Pereira, I.P. Allelopathic potential of Cistus ladanifer L. and Cistus salvifolius L. Master’s Thesis, Instituto Superior de Agronomia, Lisboa, Portugal, 1992. [Google Scholar]
- Dias, A.S.; Costa, C.T.; Dias, L.S. Allelopathic Plants. XVII. Cistus ladanifer L. Allelopathy. J. 2005, 16, 1–30. [Google Scholar]
- Chaves, N.; Alías, J.C.; Sosa, T. Phytotoxicity of Cistus ladanifer L.: Role of allelopathy. Allelopathy. J. 2016, 38, 113–132. [Google Scholar]
- Pérez-Izquierdo, C.; Serrano-Pérez, P.; Rodríguez-Molina, M.C. Chemical composition, antifungal and phytotoxic activities of Cistus ladanifer L. essential oil and hydrolate. Biocatal. Agric. Biotechnol. 2022, 45, 102527. [Google Scholar] [CrossRef]
- Verdeguer, M.; Blázquez, M.A.; Boira, H. Chemical composition and herbicidal activity of the essential oil from a Cistus ladanifer L. population from Spain. Nat. Prod. Res. Former. Nat. Prod. Lett. 2012, 26, 1602–1609. [Google Scholar] [CrossRef]
- Frazão, D.F.; Martins-Gomes, C.; Steck, J.L.; Keller, J.; Delgado, F.; Gonçalves, J.C.; Bunzel, M.; Pintado, C.M.B.S.; Díaz, T.S.; Silva, A.M. Labdanum Resin from Cistus ladanifer L.: A Natural and Sustainable Ingredient for Skin Care Cosmetics with Relevant Cosmeceutical Bioactivities. Plants 2022, 11, 1477. [Google Scholar] [CrossRef]
- Chaves, N.; Sosa, T.; Alías, J.C.; Escudero, J.C. Identification and effects of interaction phytotoxic compounds from exudate of Cistus ladanifer leaves. J. Chem. Ecol. 2001, 27, 611–621. [Google Scholar] [CrossRef]
- Chaves, N.; Sosa, T.; Escudero, J.C. Plant growth inhibiting flavonoids in exudate of Cistus ladanifer and in associated soils. J. Chem. Ecol. 2001, 27, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Chaves, N.; Sosa, T.; Alías, J.C.; Escudero, J.C. Germination inhibition of herbs in Cistus ladanifer L. soil: Possible involvement of allelochemicals. Allelopathy. J. 2003, 11, 31–42. [Google Scholar]
- Días, A.S.; Dias, L.S.; Pereira, I.P. Activity of water extracts of Cistus ladanifer and Lavandula stoechas in soil on germination and early growth of wheat and Phalaris minor. Allelopathy. J. 2004, 14, 59–64. [Google Scholar]
- Herranz, J.M.; Farrandis, P.; Copete, M.A.; Duro, E.M.; Zalacaín, A. Effect of allelopathic compounds produced by Cistus ladanifer on germination of 20 Mediterranean taxa. Plant Ecol. 2006, 184, 259–272. [Google Scholar] [CrossRef]
- Sosa, T.; Valares, C.; Alías, J.C.; Chaves, N. Persistence of flavonoids in Cistus ladanifer soils. Plant Soil. 2010, 377, 51–63. [Google Scholar] [CrossRef]
- Ramalho, P.S.; de Freitas, V.A.P.; Macedo, A.; Silva, G.; Silva, A.M.S. Volatile components of Cistus ladanifer leaves. Flavour Fragr. J. 1999, 14, 300–302. [Google Scholar] [CrossRef]
- García-Martín, D.; García-Vallejo, M.C. Contribution à la connaissance de l´huile essentielle de Cistus ladaniferus L. Var. Maculatus Dun Ciste commun (jara) d´Espagne. Parfumerie Cosmétique Savons 1969, 12, 283–290. [Google Scholar]
- Simon-Fuentes, A.; Sendra, J.M.; Cuñat, P. Neutral volátiles of Cistus ladaniferus L. essential oil. An. R. Soc. Esp. Quím. 1987, 83, 201–204. [Google Scholar]
- National Center for Biotechnology Information (2024). PubChem Compound Summary for CID 460, Guaiacol. Retrieved October 24, 2024 from https://pubchem.ncbi.nlm.nih.gov/compound/Guaiacol.
- National Center for Biotechnology Information (2024). PubChem Compound Summary for CID 31404, Butylated Hydroxytoluene. Retrieved October 24, 2024 from https://pubchem.ncbi.nlm.nih.gov/compound/2_6-ditert-butyl-4-methylphenol.
- Kato-Noguchi, H.; Seki, T.; Shigemori, H. Allelopathy and allelopathic substance in the moss Rhynchostegium pallidifolium. J. Plant Phys. 2010, 167, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Einhellig, F.A.; Schon, M.K.; Rasmussen, J.A. Synergistic effects of four cinnamic acid compounds on grain sorghom. Plant Growth Regul. 1983, 1, 251–258. [Google Scholar]
- Batish, D.R.; Singh, H.P.; Kaur, S.; Kohli, R.K.; Yadav, S.S. Caffeic acid affects early growth, and morphogenetic response of hypocotyl cuttings of mung bean (Phaseolus aureus). J. Plant Phys. 2008, 165, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Requesón, E.; Osuna, D.; del Rosario Santiago, A.; Sosa, T. Evaluation of the Activity of Estragole and 2-Isopropylphenol, Phenolic Compounds Present in Cistus ladanifer. Agronomy 2022, 12, 1139. [Google Scholar] [CrossRef]
- Cheng, H.H. Characterization of the mechanisms of allelopathy: Modeling and experimental approaches. In Allelopathy: Organisms, Processes, and Applications; Inderjit, Dakshini, K.M.M., Einhellig, F.A., Eds.; American Chemical Society: Washington, WA, USA, 1995; pp. 132–141. [Google Scholar]
- Einhellig, F.A.; Leather, G.R.; Hobbs, L.L. Use of Lemna minor L. as a bioassay in allelopathy. J. Chem. Ecol. 1985, 11, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Li, H.H.; Inoue, M.; Nishimura, H.; Mizutani, J.; Tsuzuki, E. Interactions of trans-cinnamic acid, its related phenolic allelochemicals, and abscisic acid in seedling growth and seed germination of lettuce. J. Chem. Ecol. 1993, 19, 1775–1787. [Google Scholar] [CrossRef]
- Waller, G.R. Allelochemicals: Role in Agriculture and Forestry. ACS Symposium Series 330; American Chemical Society: Washington, WA, USA, 1987. [Google Scholar] [CrossRef]
- Espinosa-Colín, M.; Hernandez-Caballero, I.; Infante, C.; Gago, I.; García-Muñoz, J.; Sosa, T. Evaluation of Propiophenone, 4-Methylacetophenone and 2',4'-Dimethylacetophenone as Phytotoxic Compounds of Labdanum Oil from Cistus ladanifer L. Plants 2023, 12, 1187. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dudai, N.; Poljakoff-Mayber, A.; Mayer, A.M.; Putievsky, E.; Lerner, H.R. Essential oils as allelochemicals and their potential use as bioherbicides. J. Chem. Ecol. 1999, 25, 1079–1089. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic effects and mechanism of action of essential oils and terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef]
- Tena, C.; del Rosario Santiago, A.; Osuna, D.; Sosa, T. Phytotoxic Activity of p-Cresol, 2-Phenylethanol and 3-Phenyl-1-Propanol, Phenolic Compounds Present in Cistus ladanifer L. Plants 2021, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Li Liu, D.; An, M.; Johnson, I.R.; Lovett, J.V. Mathematical modeling of allelopathy. III. A model for curve-fitting allelochemical dose responses. Nonlinearity Biol. Toxicol. Med. 2003, 1, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Paradigm lost, paradigm found: the re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ. Pollut. 2005, 138, 378–411. [Google Scholar] [CrossRef]
- Malato-Belíz, J.; Escudero, J.C.; Buyolo, T. Application of Traditional Indices and of Diversity to an Ecotonal Area of Different Biomes. In The State of the Art in Vegetation Science; International Association for Vegetation Science (IAVS): Toledo, Spain, 1992; Volume 75. [Google Scholar]
- Wacker, T.; Safir, G.R.; Stephens, C.T. Effects of ferulic acid on Glomus fasciculatum and associated effects on phosphorus uptake an growth of asparagus (Asparagus officinalis L.). J. Chem. Ecol. 1990, 16, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Baziramakenga, R.; Simard, R. R.; Leroux, G. D. Effects of Benzoic and Cinnamic Acid on Growth, Chlorophyll and Mineral Contents of Soybean. J. Chem. Ecol. 1994, 20, 2821–2833. [Google Scholar] [CrossRef]
- Muller, C.H. The role of chemical inhibition (allelopathy) in vegetation composition. Bull. Torrey Bot. Club 1966, 93, 332–351. [Google Scholar] [CrossRef]
- Ballester, A. Estudio de sustancias de crecimiento aislados de Ericacinerea L. Acta Científica Compostel 1971, 8, 79–84. [Google Scholar]
- Tukey, H.B. Leaching of metabolites from above grown plant parts and its implication. Bull Torrey. Bot. Club 1966, 93, 385–401. [Google Scholar] [CrossRef]
- Del Moral, R.; Muller, C.H. Effects of Eucalyptus camaldulensis. Am. Midl. Nat. 1975, 83, 254–282. [Google Scholar] [CrossRef]
- Einhelling, F.A. Allelopathy: Current Status and Future Goals. In Allelopathy: Organisms, Processes, and Applications; Inderjit, K.M., Dakshini, M., Einhelling, F.A., Eds.; American Chemical Society: Washington, DC, USA, 1995; pp. 1–24. [Google Scholar]
- Rabotnov, T.A. On the allelopathy in the phytocenosis. Izo Akad Nauk Ser Biol. 1974, 6, 811–820. [Google Scholar]
- An, M.; Pratley, J.; Haig, T. Allelopathy: From Concept to Reality. In Proceedings of the 9th Australian Agronomy Conference, Wagga Wagga, Australia, 20–23 July 1998; pp. 563–566. [Google Scholar]
- Lehman, M.E.; Blum, U. Influence of pretreatment stresses on inhibitory effects of ferulic acid, an allelopathic phenolic acid. J. Chem. Ecol. 1999, 25, 1517–1529. [Google Scholar] [CrossRef]
- Bhowmik, P.C. Inderjit, Challenges and opportunities in implementing allelopathy for natural weed management. Crop Prot. 2003, 22, 661–671. [Google Scholar] [CrossRef]
- Li, X.J.; Xia, Z.C.; Kong, C.H.; Xu, X.H. Mobility and microbial activity of allelochemicals in soil. J. Agric. Food Chem. 2013, 61, 5072–5079. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.Y.; Kong, C.H.; Wang, P.; Huang, Q.L. Temporal variation of soil friedelin and microbial community under different land uses in a long-term agroecosystem. Soil Biol. Biochem. 2014, 69, 275–281. [Google Scholar] [CrossRef]
- Shi, K.; Shao, H. Changes in the soil fungal community mediated by a Peganum harmala allelochemical. Front. Microbiol. 2022, 13, 911836. [Google Scholar] [CrossRef] [PubMed]
- Inderjit. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 2005, 274, 227–236. [Google Scholar] [CrossRef]
- Caspersen, S.; Alsanius, B.W.; Sundin, P.; Jensen, P. Bacterial amelioration of ferulic acid toxicity to hydroponically grown lettuce (Lactuca sativa L.). Soil Biol Biochem 2000, 32, 1063. [Google Scholar] [CrossRef]
- Kong, C.H.; Wang, P.; Gu, Y.; Xu, X.H.; Wang, M.L. The fate and impact on microorganisms of rice allelochemicals in paddy soil. J. Agric. Food Chem. 2008, 56, 5043–5049. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, A.; Gentili, A.; Isidori, M.; Monaco, P.; Nardelli, A.; Parrella, A.; Temussi, F. Environmental effects caused by olive mill wastewaters: toxicity comparison of low-molecular-weight phenol components. J. Agric. Food Chem. 2003, 51, 1005–1009. [Google Scholar] [CrossRef]
- Pan, Z.; Baerson, S.R.; Wang, M.; Bajsa-Hirschel, J.; Rimando, A.M.; Wang, X; Nanayakkara, N.P.D.; Noonan, B.P.; Fromm, M.E.; Dayan, F.E.; Khan, I.A.; Duke, S.O. A cytochrome P450 CYP71 enzyme expressed in Sorghum bicolor root hair cells participates in the biosynthesis of the benzoquinone allelochemical sorgoleone. New Phytol. 2018, 218, 616. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. Literature review on higher plants for toxicity testing. Water Air Soil Poll. 1991, 59, 381–400. [Google Scholar] [CrossRef]
- Chiapusio, G.; Sánchez, M.A.; Reigosa, J.M.; González, L.; Pellissier, F. Do germination indices adequately reflect allelochemical effects on the germination process? J. Chem. Ecol. 1997, 23, 11. [Google Scholar] [CrossRef]
- Jaderlund, A.; Zackrisson, O.; Nilsson, M.C. Effects of bilberry (Vaccinium myrtillus L.) litter on seed germination and early seedling growth of four boreal tree species. J. Chem. Ecol. 1996, 22, 973–986. [Google Scholar] [CrossRef]
Figure 1.
Chemical structure of 2-Methoxyphenol (A), 2,6-Di-tert-butyl-4-methylphenol (B). Source: https://echa.europa.eu/ (accessed on 28 October 2024).
Figure 1.
Chemical structure of 2-Methoxyphenol (A), 2,6-Di-tert-butyl-4-methylphenol (B). Source: https://echa.europa.eu/ (accessed on 28 October 2024).
Figure 2.
The effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the total germination (%Gt) and germination rate (%GR) of Lactuca sativa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 × 50 = 200 seeds in total for each solution). * Significantly different from the controls; 1, 2 and 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Figure 2.
The effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the total germination (%Gt) and germination rate (%GR) of Lactuca sativa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 × 50 = 200 seeds in total for each solution). * Significantly different from the controls; 1, 2 and 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
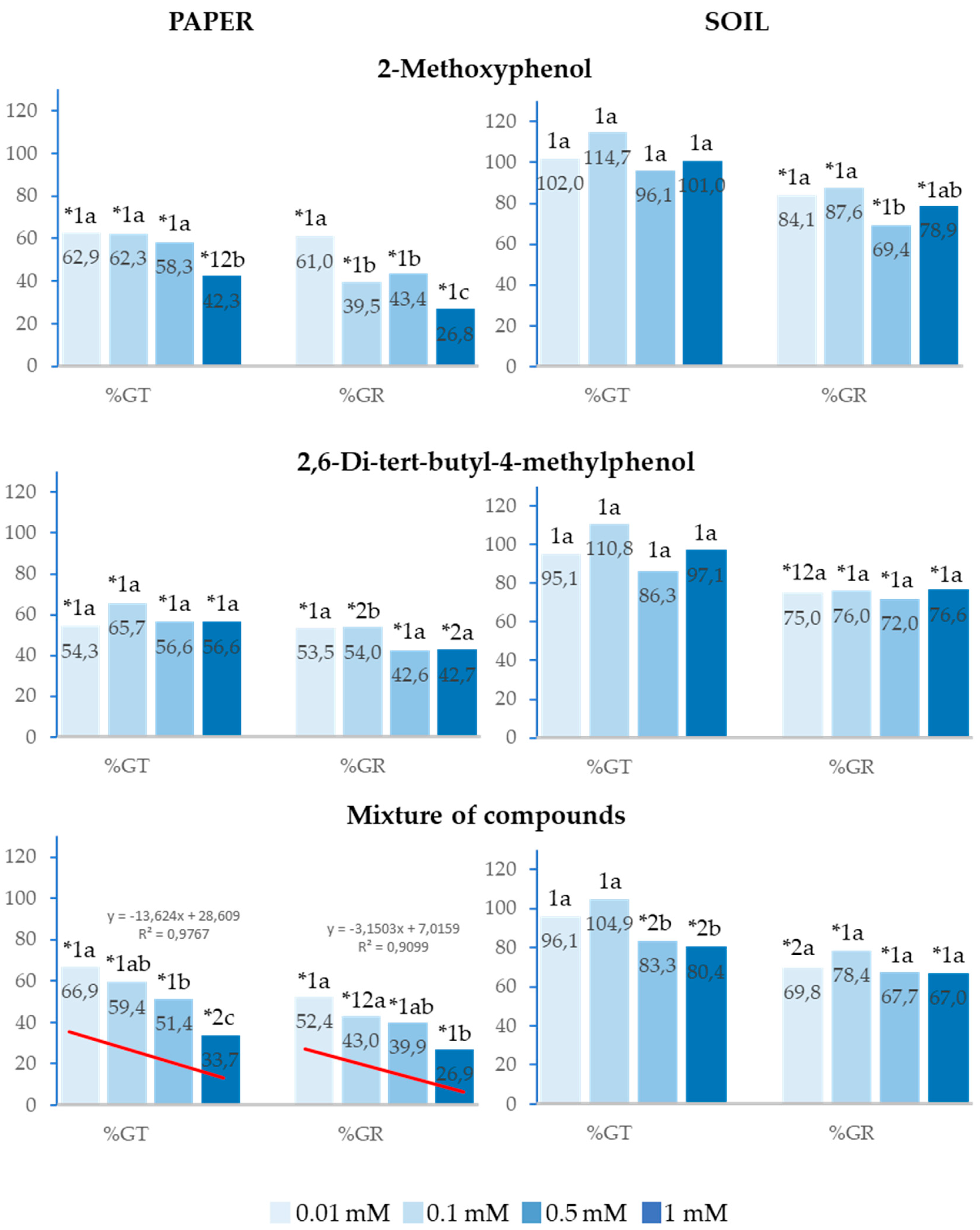
Figure 3.
The effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the total cotyledon emergence (%Ct) and cotyledon emergence rate (%CR) of Lactuca sativa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 × 50 = 200 seeds in total for each solution). * Significantly different from the controls; 1, 2 and 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Figure 3.
The effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the total cotyledon emergence (%Ct) and cotyledon emergence rate (%CR) of Lactuca sativa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 × 50 = 200 seeds in total for each solution). * Significantly different from the controls; 1, 2 and 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
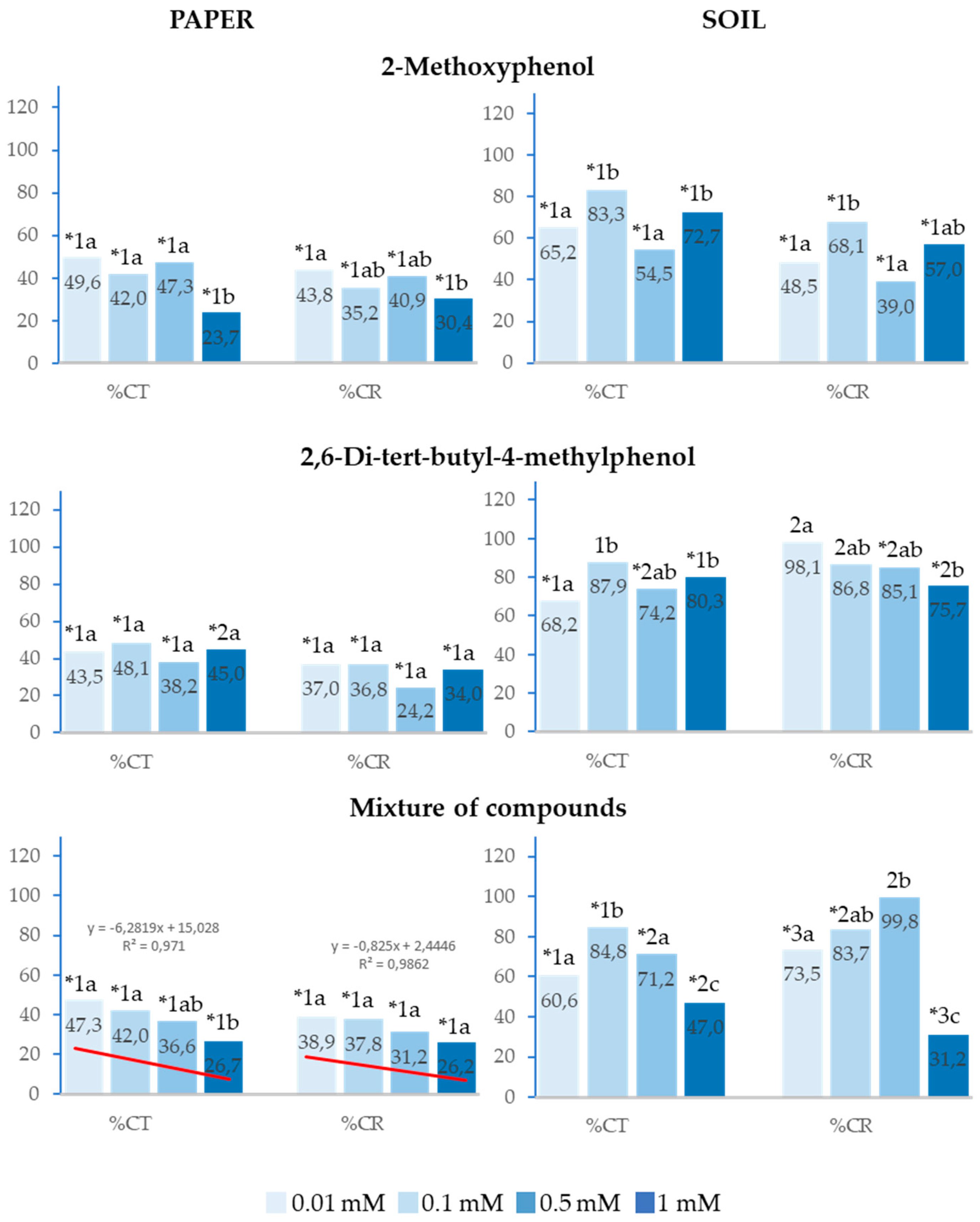
Figure 4.
Effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the radicle and hypocotyl length of Lactuca sativa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 x 50 = 200 seeds in total for each solution). * Significantly different from the controls. 1, 2, 3: different numbers indicate significant differences between treatment of the same index and for each concentration. a, b, c: different in letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Figure 4.
Effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the radicle and hypocotyl length of Lactuca sativa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 x 50 = 200 seeds in total for each solution). * Significantly different from the controls. 1, 2, 3: different numbers indicate significant differences between treatment of the same index and for each concentration. a, b, c: different in letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
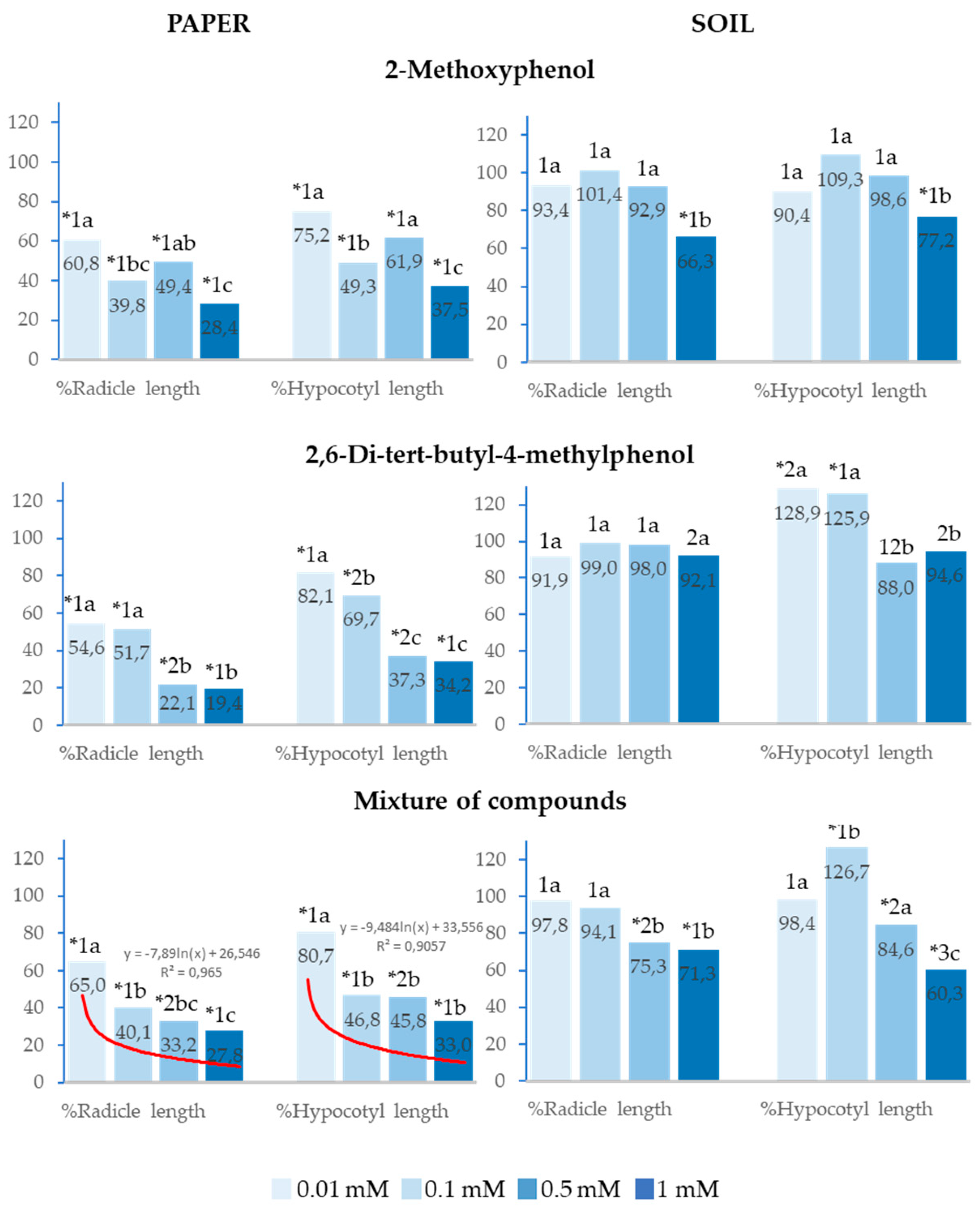
Figure 5.
The effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the total germination (%Gt) and germination rate (%GR) of Allium cepa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 × 50 = 200 seeds in total for each solution). * Significantly different from the controls; 1, 2 and 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Figure 5.
The effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the total germination (%Gt) and germination rate (%GR) of Allium cepa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 × 50 = 200 seeds in total for each solution). * Significantly different from the controls; 1, 2 and 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
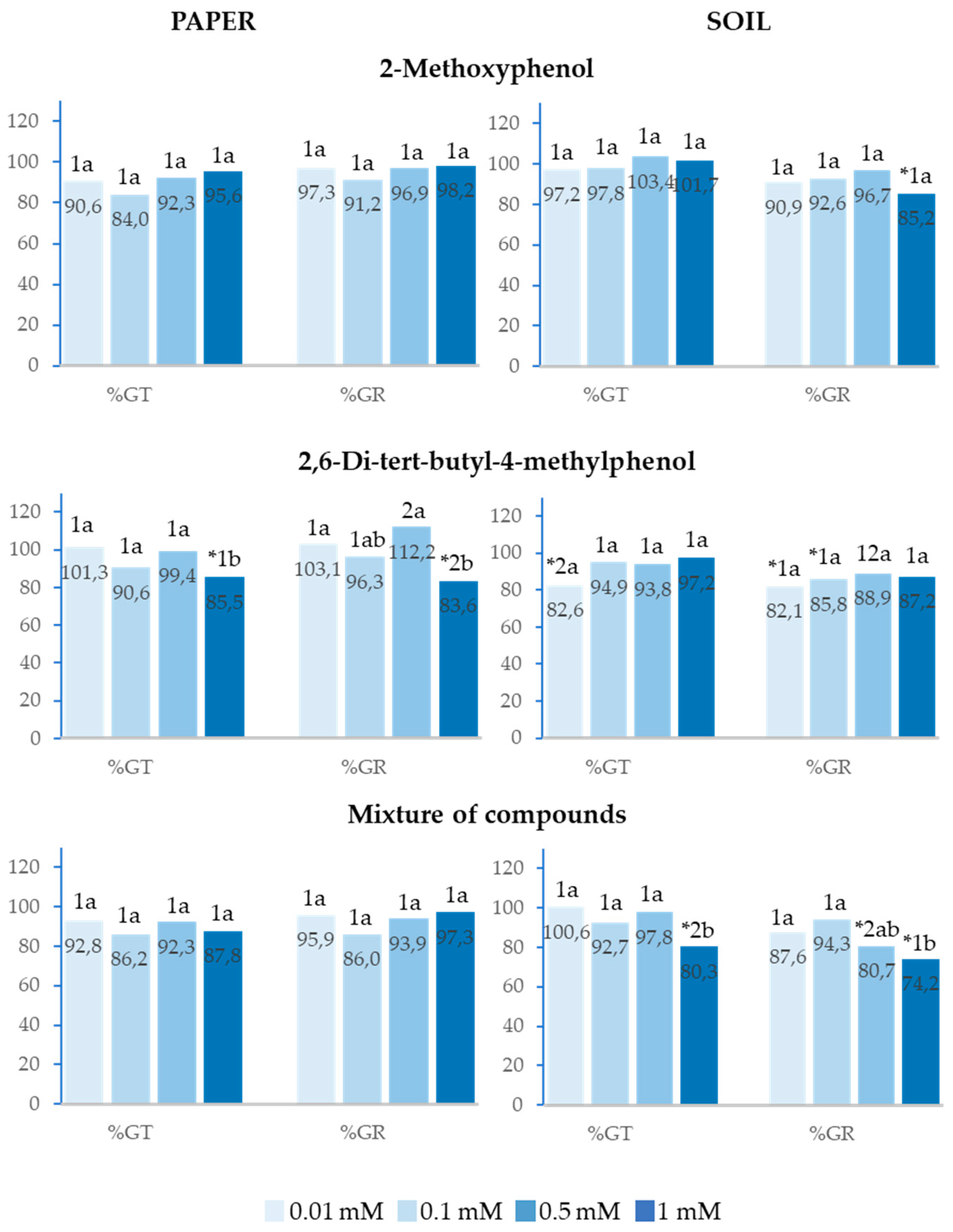
Figure 6.
Effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the radicle and hypocotyl length of Allium cepa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 x 50 = 200 seeds in total for each solution). * Significantly different from the controls. 1, 2, 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Figure 6.
Effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the radicle and hypocotyl length of Allium cepa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 x 50 = 200 seeds in total for each solution). * Significantly different from the controls. 1, 2, 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
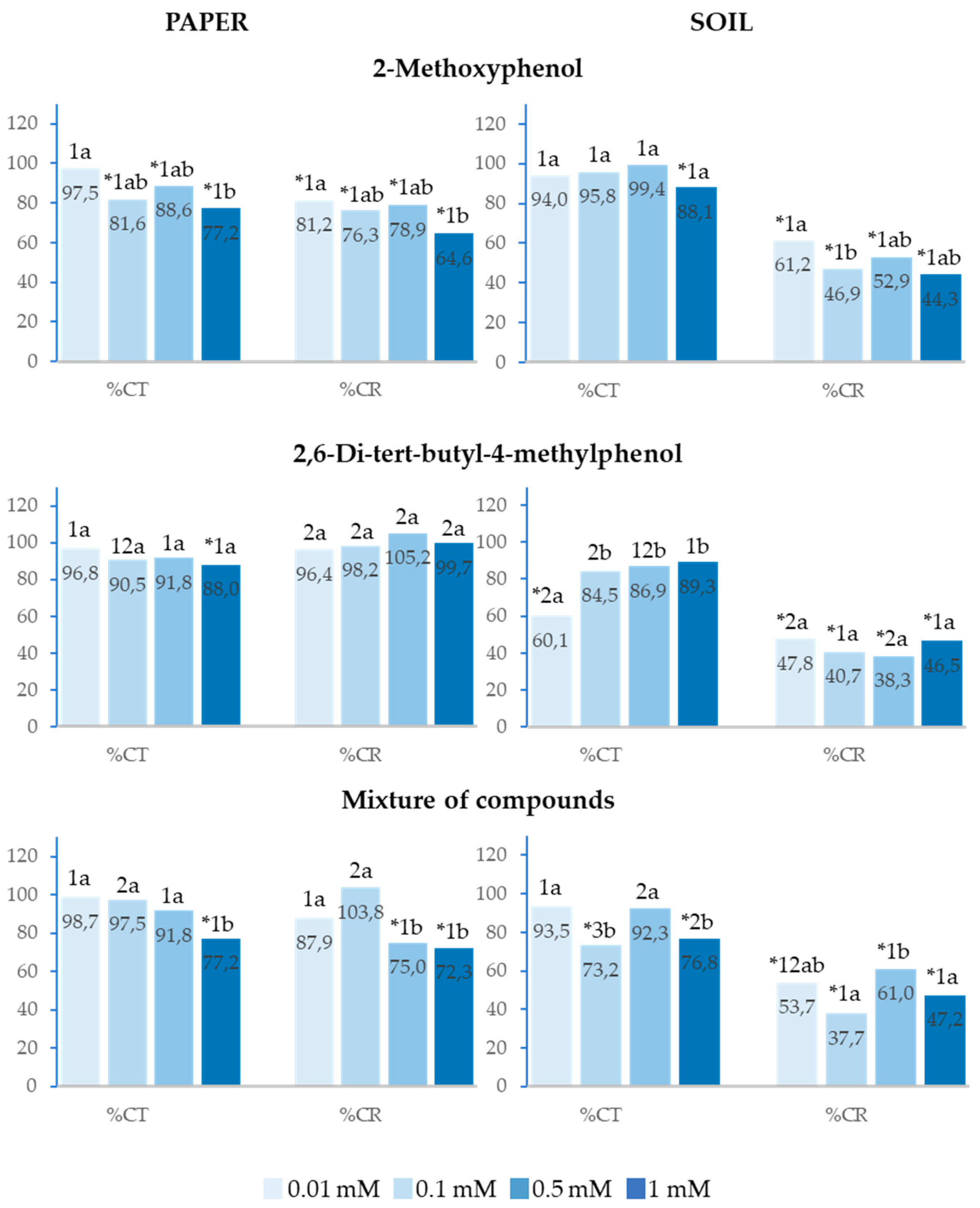
Figure 7.
Effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the radicle and hypocotyl length of Allium cepa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 x 50 = 200 seeds in total for each solution). * Significantly different from the controls. 1, 2, 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Figure 7.
Effect of different concentrations of 2-Methoxyphenol and 2,6-Di-tert-butyl-4-methylphenol and the mixture of the two compounds on the radicle and hypocotyl length of Allium cepa, expressed as percentage relative to the control. Four replicates of each treatment were performed (n = 4 x 50 = 200 seeds in total for each solution). * Significantly different from the controls. 1, 2, 3: different numbers indicate significant differences between treatments of the same index and for each concentration. a, b, c: different letters indicate significant differences between concentrations of the same index and for each treatment. P < 0.05 (Mann–Whitney U test).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



