
Submitted:
14 November 2024
Posted:
14 November 2024
You are already at the latest version
Abstract
In spite of significant advancements in diagnosis and treatment, cancer remains one of the major threats to human health due to its ability to cause disease with high morbidity and mortality. A multifactorial and multitargeted approach is required towards intervention of the multitude of signaling pathways associated with carcinogenesis inclusive of angiogenesis and metastasis. In this context, plants provide an immense source of phytotherapeutics that show great promise as anticancer drugs. There is increasing epidemiological data that diets rich in vegetables and fruits could decrease the risks of certain cancers. Several studies have proved that natural plant polyphenols such as flavonoids, lignans, phenolic acids, alkaloids, phenylpropanoids, isoprenoids, terpenes and stilbenes could be used in anticancer prophylaxis and therapeutics by recruitment of mechanisms inclusive of antioxidant and anti-inflammatory activities and modulation of several molecular events associated with carcinogenesis. The current review discusses the anticancer activities of principal phytochemicals with focus on signaling circuits towards targeted cancer prophylaxis and therapy. Also addressed are plant-derived anti-cancer vaccines, nanoparticles, monoclonal antibodies and immunotherapies. This review article brings to light the importance of plants and plant-based platforms as invaluable, low-cost sources of anti-cancer molecules of particular applicability in resource-poor developing countries.
Keywords:
Cancer
; Phytochemicals
; PVNPs
; VLPs
; Mabs
; Phototherapy
; Plant viruses
1. Introduction
Cancer is one of the most lethal diseases affecting humans due to its capability to metastasize, failure to treat and manage the disease appropriately and lack of comprehensive knowledge of development mechanisms associated with cancer. As of recent times, the basic approaches in the treatment of cancer include chemotherapy, radiotherapy, and surgery. Such strategies do provide efficacy in treatment of patients having early-stage cancers and nonmetastatic cancers. However, they are by and large ineffective in achieving long-lasting beneficial effects in patients suffering from late-stage cancers and are often burdened with major impediments such as the elicitation of resistance rendering chemotherapy ineffective in the long run, high toxicity associated with prolonged use of chemotherapy [1] and the occurrence of severe adverse side-effects caused by overall radiation doses as well as the destructive impacts of cytotoxic drugs on healthy human cells and physiological functions [2]. In such a context of increased prevalence of cancer and augmented burden from a socioeconomic viewpoint, the need for providing alternate cancer prevention and therapeutic approaches is compellingly urgent.
In spite of environmental exposure of the human population to various chemical carcinogens, diet rich in vegetables and fruits forms a favored source of phytochemicals that preclude cancer. Phytochemicals confer properties such as detoxification, free radical scavenging and antioxidant activities that provide potent anti-cancer effects. Phytochemicals regulate cancer proliferative and cancer cell apoptotic pathways besides functioning as modulators of epigenetic mechanisms leading to chemoprevention of cancer [3,4,5]. Medicinal plants are promising as adjuvants to augment the efficiency of anti-cancer drugs mediated through targeting of multiple signaling pathways while minimizing detrimental side effects [6,7]. Therefore, there is a compelling need to investigate cellular pathways and explore their cross-link with phytochemicals in cancer therapy. Phytochemicals control multiple signaling networks by upregulation of tumor suppressor genes, inhibition of oncogenes and modulating the expression of upstream and downstream mediators to preclude cancer progression.
The burgeoning immunotherapy technology has shown tremendous potential in averting or suppressing the progression of cancer owing to its direct impact on malignant cancer cells with high efficacy to target and destroy cancer cells [8]. Recent investigations on antibody-enabled killing effects on tumor cells and the production of numerous antibodies against cancer cells has resulted in the generation of monoclonal antibodies (Mabs) that targetedly recognize specific antigens on the cancer cell surface. However, despite their benefits and treatment potential, their manufacture is not cost-efficient considering the requirements of high purity and quality of these antibodies in addition to the circumscribed scalability of the mammalian expression systems which impairs their widespread use as anticancer therapeutics. Further, Mabs produced in mammalian cells contain several mouse proteins and contaminants such as human pathogens. Plants provide a novel approach for generation of monoclonal antibodies against cancer. Plants inherently have high expression capability, inexpensive cultivation processes on large scales, lowered downstream processing steps subject to containment conditions while being bereft of human pathogens and ethical concerns associated with systems such as transgenic animals [9,10]. Hence, plants have tremendous potential as practically unlimited sources of Mabs, referred to as “plantibodies”.
Oncolytic viruses are of high clinical value for their efficacy against cancer. However, the medical use of these viruses is challenged by the likelihood of reversion mutations of attenuated oncolytic viruses into their virulent forms as well as the possibility of integration of genomic sequences of the virus into the host genome [11]. On the other hand, plant viruses are not capable of infecting mammalian cells and therefore lack the above drawbacks related to infection and afford a valuable tool for manipulation of tumors and induction of anti-tumor immunity [11,12]. Plant virus nanoparticles do not replicate in and destroy cancer cells in a direct manner but constitute a novel class of immunostimulatory agents [12]. There are two types of plant viruses namely, whole viruses referred to as viral nanoparticles (VNPs) and virus-like particles (VLPs). VLPs are genome-free equivalents of VNPs, lack the ability to replicate in plants and resemble the native structures of plant viruses. Both VLPs and VNPs can function as immune adjuvants and delivery systems for tumor-specific antigens which can be recognized by the human immune system. VNPs and VLPs are efficaciously taken up by antigen-presenting cells and can elicit strong immune responses. They have been used in cancer immunotherapy by direct injection into the tumors in order to induce anti-tumor immunity through the disruption of local immunosuppression, which renders support to local followed by systemic immunity against the tumor, a process referred to as “in situ vaccination”.
A recent review on plant-based platforms inclusive of plant-derived monoclonal antibodies, VNPs and phytochemicals to treat cancer is lacking. This comprehensive review aims to provide insights into the molecular mechanisms underlying the anticancer activities of plant-derived bioactive compounds / phytochemicals and their integration into phototherapeutic strategies. Further, it discusses the use of plant-based monoclonal antibodies and plant viral nanoparticles in combating cancer.
2. Phytochemicals in Cancer Prevention and Therapeutics: Recent Developments
Cancer continues to impose a formidable global health challenge, demanding a perpetual quest for innovative and effective therapeutic approaches. Anticancer plants, a captivating realm within the broader spectrum of medicinal flora, represent a rich source of bioactive compounds that have demonstrated remarkable potential in the fight against cancer. These plants, often celebrated in traditional medicine practices, harbour an array of phytochemicals such as alkaloids, flavonoids, polyphenols, and terpenoids, which manifest diverse biological activities with pronounced anticancer properties. Exploring these natural compounds has opened avenues for innovative and targeted therapeutic approaches in cancer treatment.
3. Major Benefits of Phytochemicals in Comparison with Synthetic Anti-Cancer Drugs
Chemotherapy is a therapeutic regime involving administration of combinations of synthetic drugs to the body. Significantly, this is the only one among a few therapeutic choices available to treat advanced stage or metastasized cancer. Nevertheless, an adverse drawback of chemotherapy is its lack of target selectivity [13]. Since cancer cells emerge from functional normal cells exhibiting unbridled growth, anticancer drugs show indiscriminate targeting of the growth and development of non-proliferative normal cells while inhibiting the growth of cancer cells. Such poor selectivity of most chemotherapeutic drugs leads to serious side effects impacting normal tissues including hair follicles, gastrointestinal tract and bone marrow [13]. Further, chemotherapeutic drugs compromise fertility and induce long-term damage to organs and even cause cancer. Additionally, these drugs pose toxicity issues to normal cells leading to several side effects of which some could be life-threatening. Many synthetic anticancer drugs have been shown to be associated with notable undesirable and adverse side effects such as ototoxicity due to the long-term administration of cisplatin [14], cardiotoxicity due to doxorubicin [15] and cognitive impairment due to 5-fluorouracil [16]. These side effects include fatigue, pain, problems of the mouth, skin, nail and hair, anemia, nausea dietary issues and weight change. Also, there occur other side effects which may not be as potent but severely circumscribe the patients’ quality of life leading to premature cessation of chemotherapy.
Synthetic anticancer drugs have poor solubility, diminished absorptions and decreased oral availability [17]. Also, other complications such as elicitation of multidrug resistance (MDR) emerge as these drugs are targeted to the DNA of a given cell wherein mutations occur and hence the cell develops resistance. It is because of the emergence of MDR, cancer cells continue to grow despite the administration of chemotherapeutic drugs. Manifestations of drug tolerance in cancer cells include changes in the potential drug target or enhancement of mechanisms for cell survival including alterations in apoptotic cycles caused by ceramide level changes, occurrence of DNA repair, inefficient p53 tumor suppressor protein or impact on cytochrome oxidases critical for cellular respiration. MDR also causes hyper-expression of efflux transporters based on the ATP binding cassette that in turn decrease the drug levels within the intracellular space to levels that are suboptimal [18]. Therefore, the adverse side effects of chemotherapy in addition to MDR development and the circumscribed therapeutic index of chemotherapeutic drugs, severely impact the therapeutic efficiency of chemotherapy. Such side effect severity requires reduction in dosage of anticancer drugs eventually leading to ineffectual treatment outcomes and potentiating metastasis. The effectiveness of synthetic anticancer drugs is circumscribed by complications such as frequent tumor relapse and initiation of metastasis [19]. Also, conventional chemotherapeutics drugs are prohibitively expensive. The delay in cancer diagnosis combined with non-responsive therapy cause high rates of mortality among several cancer patients. Such a situation has instigated the compelling necessity to look for alternative anticancer drugs sourced from plant systems.
Phytochemicals have been found to be less toxic, specific and selective to cancer cells due to which they show great promise as anticancer drugs. Phytochemicals preclude DNA damage, promote DNA repair, slow down cancer cell growth, regulate hormones and prevent the reproduction of damaged cells. They possess antioxidant properties, while neutralizing free radicals that damage the DNA. Phytochemicals have been shown to suppress pathways involved in promoting cancer and cancer progression, eventually inhibiting unbridled cell growth and causing cell death through apoptosis. They alter proteins associated with various signal transduction pathways and exert definitive chemotherapeutic and chemopreventive roles by integrating with specific molecular signals. They execute multiple biological functions such as antimutagenic, antiproliferative, antimetastatic, anti-angiogenesis, anti-inflammatory, antioxidant and immunomodulatory properties due to which they control cancer progression and intervene in different stages of cancer cell development. Besides, they are involved in the regulation of cell cycle as well as microRNA and lead to cancer cell death by promoting apoptosis and autophagy through ROS signaling. Chemopreventive phytochemicals substances vary in their efficacies based on the genotypes of individuals in the population. Hence, the combined administration of many phytochemicals contained in plant foods would enable suppression of carcinogenesis providing synergistic or additive effects against cancer when compared to treatment with single phytochemicals [20].
4. Molecular Insights into the Roles of Phytochemicals Against Cancer
Phytochemicals used to preclude cancer are categorized as tumor blockers or tumor suppressors wherein blockers target events involved in cancer initiation while suppressors preclude tumor promotion [21,22,23]. Several natural compounds such as polyphenols including curcumin, flavonoids, lignans, stilbenes, tannins and coumarins as well as glucosinolates and isothiocyanates block tumorigenesis by inducing the biosynthesis of detoxification enzymes [22,24], preventing DNA damage, genetic instability and mutation that instigate neoplastic transformation of cells. Such phase II detoxifying enzyme induction has been shown to be generated by Nrf2-ARE pathway activation [21,22,24].
On the other hand, suppressor agents inhibit tumor growth, development and tumor progression in cells that have already been transformed and promote removal of cancer cells within the tumor mass. Such suppressing phytochemical agents include isothiocyanates found in cabbage, cauliflower, broccoli and watercress, flavonoids occurring in citrus fruit, grapefruits, blueberries and parsley as well as coumarins sourced from cinnamon, tonka beans and sweet clover [21,24]. These suppressors hinder tumor promotion via several mechanisms including cell cycle arrest, angiogenesis inhibition, repression of pathways supporting cancer cells including NFkB [21,24].
Phytochemicals induce apoptosis by repressing release of Bcl-2, Bcl-xL and other anti-apoptotic proteins, stimulating release of Bak, Bax and other pro-apoptotic proteins that lead to cytochrome C release from the inner membrane of the mitochondria and apoptosome formation which results in activation and release of caspases 3, 6 and 7 effector proteins. This degrades intracellular proteins, leads to nuclear fragmentation, blebbing and cell shrinkage. Likewise, suppressors block the synthesis of hormones and hormone receptors within breast cancer cells that rely on hormones for tumor growth and development [21,22,24,25]. Phytochemicals are efficacious, widely available, non-toxic and possess many biological activities such as anticancer, pro-apoptotic, antioxidant, anti-proliferative, anti-angiogenic and anti-inflammatory properties [26,27,28].
5. Terpenes and Anticancer Effects
The Sweet wormwood, Artemisia annua contains artemisinin, a powerful sesquiterpene lactone that has anticancer effects. PH-sensitive nanoparticles (D/D NPs) containing dihydroartemisinin (DHA) and docetaxel (DTX) demonstrated anticancer activity in vitro in breast cancer cells. D/D NPs augmented ROS, decreased mitochondrial membrane potential, enhanced p53 expression and elicited release of cytochrome C into the cytoplasm that in turn activated caspase-3. This D/D NP combination therapy is a propitious strategy for treating metastatic breast cancers through ROS-generated mitochondrial apoptosis pathway [29].
The triterpene 3,7-dihydroxy-25-methoxycucurbita-5,23-diene-19-al of the cucurbitane-type (DMC) present in Momordica charantia L., the wild bitter melon shows anticancer effects in breast cancer cells wherein it inhibits mTOR-p70S6K signaling through activation of AMPK and downregulation of Akt, resulting in cytoprotective autophagy [30].
The sesquiterpene, furanodiene sourced from Rhizoma curcumae suppresses 95-D lung cancer cell line proliferation. Furanodiene blocked cell proliferation and inhibited the progression of the cell cycle in the G1 phase through downregulation of CDK6 and cyclin D1 protein levels and upregulation of the levels of p27 and p21 in 95-D cells [31]. It also downregulated the levels of Bcl-2, survivin, pro-caspase-7 and full PARP, and upregulated the level of cleaved PARP. It augmented the light chain 3-II (LC3-II) protein levels, implicating the involvement of autophagy in this process.
6. Alkaloids and Their Antitumor Effects
The benzylisoquinoline alkaloid, berberine is present in Berberis vulgaris as well as other Berberis plants. Berberine downregulates the growth of human gastric cancer cells in vivo and in vitro by eliciting cytostatic autophagy through inhibition of Akt and MAPK/mTOR/p70S6K pathways and affords a molecular basis for therapy of gastric cancer [32]. Treatment with berberine causes notable downregulation of HIF-1 and vascular endothelial growth factor (VEGF) which reversed resistance to radiotherapy [33].
In human lung adenocarcinoma spheroid models, caffeine shows anticancer toxicity via the decrease in the expression of Nrf2 and Claudin-2 leading to impairment of mitochondrial respiration and generation of ROS [34]. Exaggeration of cisplatin and doxorubicin toxicity was observed by caffeine therapy in these spheroids [34].
Deguelin found in peas and beans of the Leguminosae family induces apoptosis by augmenting the release of cytochrome C and protein levels of the apoptosis induction factors. Deguelin caused cell death in lung cancer cells through eliciting PUMA expression [35]. This induction was independent of p53 and was mediated through inhibition of the P13K/AKT pathway, thus triggering the binding of Foxo3a with the PUMA promoter to induce its transcription. Following activation, PUMA stimulated both Bax and the intrinsic mitochondrial cellular death pathway. Further, deguelin augmented doxorubicin chemotherapeutic sensitivity in vivo and in vitro associated with the potentiated induction of PUMA.
Piperlongumine, a phytoalkaloid is a bioactive therapeutic compound. It activates multiple cellular and molecular pathways via depolarization of cells, modulation of cell cycle growth factors, removal of oxidative stress as well as abnormally generated molecular factors and induction of apoptosis [36,37,38,39,40,41,42,43,44,45]. Piperlongumine regulates STAT3 (signal transducer and activator of transcription 3), nuclear factor kappa B, phosphatidylino-sitol 3-kinase/protein kinase B, cyclooxygenase-2, cyclin D1 and the glutathione pathway that are involved in cancer initiation, cellular proliferation and tumor progression. Piperlongumine acts as a multi-functional anticancer candidate that has been proved to have antioxidant and immune-promoting effects, thereby making it safe for healthy, normal cells.
Vincristine and vinblastine (Figure 1) sourced from Catharantus roseus, are naturally occurring alkaloids having antineoplastic properties. They are widely used for treatment of ovarian cancer, breast cancer, osteosarcoma, lung carcinoma, lymphoma, leukemia and gastric cancer. The phytochemicals including vinblastine, docetaxel and plinabulin are microtubule targeting agents (MTAs) [46,47]. When used in combination or alone, they disrupted the polymerization of microtubules by changing their conformation as well as the binding free energy of adjacent tubulin subunits. The combination of vinblastine and docetaxel synergistically augmented bending and depolymerization of microtubules
Vinblastine (Vin), when loaded on to graphene quantum dots (GQD) enhanced its cytotoxicty in cancer cells while at the same time exhibiting lowered toxicity towards normal cells [49]. The GQDs-Vin (1:5) composite showed more robust tumor inhibition compared to that of Vin alone wherein both GQDs and Vin exhibited synergistic effects. Upon evaluation of their efficiency in A549, CCF-STTG1, HGC-27, Hela and MCF-7 cancer cell lines, this composite demonstrated higher cytotoxicity compared to GQDs or Vin alone and lesser cytotoxicity towards normal cells which corroborated with similar in vivo effects in murine models.
The daratumumab-polymersome-vincristine (DP-VCR) combination was used as a nanotherapy directed towards CD38 in acute lymphoblastic leukemia (ALL) [50]. The DP-VCR demonstrated selective uptake of this composite in CD38+ Nalm-6-Luc and 697 ALL cells as well as robust anti-ALL activity wherein the IC50 was as low as 0.06 nM vincristine. This potency was 13.7-fold higher than free vincristine. Contrastingly, DP-VCR as high as 108.3 nM showed no toxicity towards human peripheral blood mononuclear cells. By means of apoptotic assays, DP-VCR was shown to be highly selective for CD38+ ALL cells. In vivo murine models treated with DP-VCR showed significant decrease of leukemia burden in the liver, spleen, blood and bone marrow and improved survival along with fewer side effects. This proved DP-VCR to be a potent and safe nanotherapy for CD38+ ALL.
Piperin, a compound in black pepper, suppresses glucuronidation of many chemopreventive substances, resulting in enhancement of their bioavailability, making piperin a robust inhibitor of drug metabolism [51]. Piperin at 75–150 µM dosage blocks the growth of many colon cancer cell lines wherein it inhibits the cell cycle and promotes apoptosis [52].
7. Organosulphur Compounds Against Cancer
Extracts from mature black garlic has been shown to negatively regulate the proliferation, invasion, migration and metastasis of the MDA-MB-361 and MCF-7 cell line ER+ breast cancer cells [53,54]. It induces apoptosis of these cells through blockage of BCL-2 and MCL-1 anti-apoptotic protein expression while eliciting the expression of BAK And BIM pro-apoptotic proteins [53]. The decrease in MCL-1 expression is mediated by the activation of JNK caused by an enhancement of ROS production in cancer cells [53]. Ripening black garlic hexane extract stimulates apoptosis in U937 human leukemia cells wherein caspase-determined apoptosis is initiated by both extrinsic and intrinsic pathways [55]. In HT29 colon cancer cells, ABGE blocks proliferation and stimulates apoptosis, possibly by modulating the PI3K/Akt signaling pathway, promoting PTEN expression and decreasing Akt and p-Akt expression [56].
In addition to S-Allyl-Cysteine (SAC), black garlic contains S-Allyl-Mercapto-Cysteine (SAMC). SAMC has anticancer effects in SW620 human colon cancer cell line caused by induction of apoptosis via the p38 and JNK pathways which in turn activate the p53 and Bax [57]. 80-90% inhibition is observed in mouse macrophage line (TIB-71), MCF-7 breast cancer, prostate cancer (PC-3), and Hep-G2 cells after 72 hours of administration of the black garlic extract, caused by inhibition of the cell cycle and cell proliferation, finally leading to apoptosis [58,59]. In HL-60 leukemia cells, black garlic had dose-determined cytotoxic effects [60]. In human gastric cancer cells SGC-7901 and in murine models, black garlic extracts exhibited immunomodulatory and anticancer effects wherein ABGE augmented GSH-Px and SOD activity and led to apoptosis and inhibition of cancer cell growth [61].
A natural phytochemical, sulforaphane obtained from cruciferous vegetables including broccoli and brussels sprouts is known for its anti-inflammatory and antioxidant potentials along with its abilities to repress the growth of cancer and associated cell proliferative capabilities of BCSCs and TNBC. Particularly, Notch and wnt/β-catenin BCSC-associated pathways are abrogated by sulforaphane [62].
Phytochemicals such as isothiocyanates are found in cruciferous vegetables and are known for their ability to preclude and treat cancer [63]. Isothiocyanate compounds have been explored for their anticancer activities against colon, liver, breast, prostate, bladder, pancreatic, lung, endometrial and glioblastoma cancer [64,65,66,67,68,69,70,71,72,73].
8. Polyphenols Against Cancer
Curcumin, a polyphenol found in Curcuma longa plants is known for its anti-cancer, antioxidant, anti-inflammatory and cytotoxic activities. In myeloid leukemia cell line, curcumin elicits apoptosis and autophagy by negative regulation of the Bcl-2 protein [74]. In human cancer cell lines, curcumin induces ER stress and malfunction of mitochondria to trigger apoptosis in HT-29 and AGS cell lines [75]. In glioma cells, curcumin causes increased expression of ING4 and p21, following which it upregulates BAX and downregulates the NF-B and Bcl-2 signaling pathways resulting in apoptosis [76,77]. Curcumin terminates the recognized wnt/β-catenin pathway, thus precluding β-catenin nuclear translocation and Slug transcription factor activation. This leads to restoration of the expression of E-cadherin and blockage of BCSC and EMT migration [78]. Moreover, curcumin terminates pathways maintaining breast cancer stemness, causes apoptosis and blocks the BCSC proliferative potential. Key signaling pathways such as Wnt/β-catenin and sonic hedgehog pathways in cancer stem cells are responsible for the drug resistance, aggression, remission and heterogeneity in breast cancer. Curcumin was shown to shut down these pathways in MCF-7 as well as in SUM159 sphere-forming cells [67] leading to the death of BCSCs. Curcumin has been shown to inhibit the PI3K/Akt/mTOR pathway which promotes cell survival and proliferation [79]. Curcumin has been shown to inhibit key signalling pathways involved in cancer progression, including nuclear factor-kappa B (NF-κB) and p38 mitogen-activated protein kinase (MAPK) pathways [80].
Epigallocatechin-3-gallate (EGCG), a polyphenol, occurs in Camellia sinensis (green tea) leaves and has anticancer and antioxidant properties. EGCG occurring in green tea catechin fractions, blocks proliferation of tumor cells, triggers apoptosis, precludes angiogenesis and cytokine synthesis [81]. It blocks the proliferative and angiogenic capability of breast cancer cells wherein it inhibits the expression of hypoxiainducible factor 1 subunit alpha (HIF-1α), activates NF-kB and expresses vascular endothelial growth factor (VEGF) in mouse models [82,83]. EGCG has been shown to downregulate matrix metalloproteinase-9 (MMP9) induced by EGF in ER+ breast cancer cells, resulting in cessation of metastasis and cellular invasion [84]. EGCG interferes with the PI3K/Akt pathway, impeding cancer cell survival and proliferation [85,86].
The phenolic gallic acid is found in onions, red fruits and tea. Gallic acid and cisplatin blocked colony formation and formation of tumor spheroids [87]. Also, they elicited apoptosis and inhibited the P13K/Akt pathway which upregulated the tumor suppressor protein, p53 that in turn, controlled proteins related to the cell cycle such as E1, Cyclin D1, p21 and p27 as well as intrinsic apoptotic proteins including cleaved caspase-3, Bax and Bcl-2. The anti-cancer properties of gallic acid were also confirmed in a murine in vivo model. Thus, gallic acid blocked progression of lung cancer by arresting the cell cycle and inducing apoptosis, thereby making it a promising therapeutic candidate to confront non-small cell lung cancer. Gallic acid can function as an adjuvant to promote the cytotoxicity of cisplatin towards lung cancer cells.
Magnolia officinalis bark contains a biphenolic neolignan, honokiol which has several therapeutic properties. Honokiol blocked glioblastoma cell proliferation by inciting slight arrest of the G0/G1 phase cell cycle and causing apoptosis through both caspase-dependent and caspase-independent pathways [88]. The apoptotic effect of honokiol involves blockage of STAT3 signaling and ERK1/2 in addition to the activation of p38 MAPK signaling pathway suggesting that treatment with honokiol could be a propitious strategy for human glioblastoma therapy.
A phenolic compound, oleocanthal occurring in virgin olive oils, suppresses proliferation, invasion and migration of prostate and breast cancer cells by inhibiting c-Met phosphorylation. Olive oil blocks progression of the cell cycle as well as cell proliferation, elicits oxidative stress and induces apoptosis while stimulating the immune system, thereby precluding carcinogenesis [89].
Phytochemicals including cinnamaldehyde and chlorogenic acid have proved to be efficacious in combating breast cancer. They block the initiation of tumor formation by detoxifying carcinogens, preventing the formation of DNA adducts, scouring electrophilic species, preventing peroxidation of lipids and protecting from mutagenesis. Additionally, they function as agents for tumor suppressing by inhibiting the growth of preneoplastic tissues, promoting autophagy and apoptosis, repressing tumor cell invasion and migration, disrupting the energy metabolism of cancerous tissues and blocking estrogen receptors [90,91,92,93]. Both natural compounds can arrest the cell cycle, promote apoptosis, inhibit cell proliferation, vascularization, capability of stemness, tumor metastasis and invasion, suppress proteins involved in drug efflux, restore normal cell metabolism and epigenetic markers and regulate the endocrine system in experimental models of breast cancer and in breast cancer cell lines.
A polyphenol phytoalexin, resveratrol is found in several kinds of grapes, blueberries and cranberries. In ovarian cancer cell lines, it initiates autophagy wherein it lowers the amount of mTOR and phosphorylated Akt [94]. It also induces the downregulation of the Wnt/β-Catenin signaling pathway and causes autophagy in BCSCs [95]. Resveratrol functions as a chemopreventive agent by intervening in the four principal phases of carcinogenesis, namely, the initiation, cancer cell promotion, cancer progression as well as metastasis [96] Resveratrol has proven anticancer efficacy in both in vivo and in vitro systems [97]. It has antitumor, antioxidant and anti-inflammatory properties which warrant its use as a complementary candidate for the traditional chemotherapy [98]. As demonstrated by several studies, it has been found to be efficacious against colorectal cancer [99], lung cancer [100], skin cancer [101] as well as prostate, pancreatic, hepatic and postmenopausal breast cancers in addition to haematological malignancies [102]. Resveratrol suppresses β-catenin expression and inhibits β-catenin nuclear translocation by perturbing MALAT1, the long noncoding RNA [103]. It downregulates the transcription factor Snail and the TGF-β/Smad-induced epithelial-mesenchymal transition (EMT) factor. Additionally, it reduces the IKK-induced TNF-β expression leading to blockage of proliferation of cancer cells via NF-ḳB deactivation. Further, it blocks nuclear accretion of FOXO3a mediated by p-PI3K/p-AKT. It downregulates Src-STAT3 phosphorylation and triggers cancer cell apoptosis. It inhibits activation of HIF-1α induced by AKT/MAPK and accelerates HIF-1α protein degradation driven by ubiquitination.
9. Phenolic Lipids Against Cancer
2-hydroxy-6-pentadecylbenzoic acid commonly called anacardic acid is found in cashew nut shells and has several anticancer properties. In A549 human lung cancer cells, it induces ER stress which promotes CHOP expression as well as cleavage of caspase-12 in addition to the disruption of Ca2+ homeostasis, resulting in apoptosis [104].
10. Flavonoids and Antitumor Effects
A flavonoid, ampelopsin sourced from Ampelopsis grossedentata (vine tea), has a diverse range of biological activites. Ampelopsin has protective antitumor effects in breast cancer cells wherein it causes apoptosis. In MCF-7 and MDA-MB-231 breast cancer cells, it induces intracellular ROS production and apoptosis associated with malfunction of mitochondria in breast cancer cells including loss of mitochondrial membrane potential, build-up of high levels of ROS and augmented expression of Bcl-2/Bax expression [105].
Apigenin, an anticancer flavonoid, is found in abundance in bell pepper, garlic, cabbage and celery. It is known to preclude cancer by inhibiting cell growth and promoting apoptosis specifically in lung cancer cells likely though enhancement of ROS generation while having no effect on normal cells. Following this, caspases 3 and 9 are induced leading to the death of A549 cells through apoptosis [106].
Another flavonoid, artocarpin sourced from the Artocarpus species promotes anticancer activity in lung cancer cells through induction of apoptosis. Artocarpin shows cytotoxic activities on non-small cell lung carcinoma (NSCLC, A549) cell lines by phosphorylating and activating cellular protein kinases inclusive of AktS473, p38 and Erk1/2 followed by apoptosis mediated by the elicitation of ROS [107]. Artocarpin activated p53-dependent apoptotic proteins inclusive of Apaf-1, caspase-3, cytochrome c and PUMA. In NSCLC cells, artocarpin-elicited apoptosis was mediated by the augmentation of both independent AktS473/NF-κB/c-Myc/Noxa and ERK/ p38/ p53-dependent cascades by ROS.
A flavonoid, butein found in Butea monosperma, has been shown to block growth of human ovarian cancer cells by inhibition of the interaction between IL-6/IL-6Rα and by regulation of the IL-6/STAT3/FoxO3a pathway attributed to butein’s high binding affinity to IL-6 [108]. This results in a reduction in cell proliferation, invasion and migration in addition to an enhancement of apoptosis and cell cycle arrest. Further, butein induced decrease in the ovarian cancer cell growth in mouse xenografts [108].
Chrysin is a natural flavone occurring in blue passionflower, propolis and honey which are known for their medicinal significance and economic value. Chrysin has anticancer, antioxidant and anti-inflammatory properties [109] as proven by various studies involving in vivo tumor models and cancer cell lines. It inhibits tumor growth by inducing apoptosis, altering the cell cycle, inhibiting invasion, angiogenesis and metastasis while being non-toxic to normal, healthy cells. Chrysin selectively modulates several cellular signaling pathways associated with tumor inflammation, angiogenesis, growth, invasion, survival and cancer cell metastasis. The broad-spectrum anticancer activity combined with low toxicity of chrysin highlights its translational value in cancer therapy. Chrysin augments the ratio of Bax/Bcl2, induces caspases 3 and 9 and stimulates lung cancer cell apoptosis [110].
The flavonoid delphinidin, elicits apoptosis in HCT116 human colon cancer cells through the generation of ROS. Activation of cytochrome C, caspase 3, 8 and 9, pro-apoptotic Bax as well as inhibition of the expression of anti-apoptotic proteins including ERK1/2, p38 and STAT-3 are all associated with apoptosis initiation [111].
Genistein, an isoflavone present in soybeans. Genistein has been shown to induce autophagy in ovarian cancer cells [112,113]. It induces cell death in ovarian cancer cells via caspase-independent pathway by inhibiting glucose uptake and leading to autophagy and apoptosis. Additionally, treatment with genistein results in diminished levels of phosphorylated Akt, that could contribute towards limiting utilization of glucose. Genistein has been used in the management of prostate and breast cancer. It inhibits inflammation-augmenting marker expression and ROS [114,115] that are associated with tumor proliferation and oxidative damage [116]. It causes induction of apoptosis and controls the spread and migration of neoplasms and has been found to be a widely used chemopreventive agent [117]. Genistein induces it apoptotic effects by modulating the Fas-FasL pathway, TRAIL-DR pathway, TNF-α-TNFR1 pathway, Bcl2-Bax pathway and targeting the PI3K-Akt-mTOR pathway and the JAK-STAT3 signal pathway. It exerts notable antiproliferative activities against ER+ human breast cancer cells by inducing G2-M arrest, p21 expression followed by apoptosis [118].
Kuwanon M (KWM) from the mulberry root bark decreases cell migration and proliferation, while augmenting apoptosis via the mitochondrial pathway, paraptosis by incrementing cytoplasmic vacuolation and by inducing ER stress in NCI-H292 and A549 lung cancer cells [119]. Mulberry Diels-Alder-type adducts (MDAAs) have anticancer properties associated with induction of apoptosis whereas KWM directly affects mitochondria, which underscores the significance of this organelle in phytochemical therapy of cancer cells [120].
The flavonoid, quercetin is found in many vegetables, onion, apples, green tea, berries and red wine. It is endowed with antioxidant and anti-inflammatory activities. Quercetin augments the accretion of hypoxia-induced factor 1 (HIF-1) in gastric cancer cells, in turn inhibiting mTOR signaling and stimulating the biosynthesis of BNIP3/BNIP3L. This process disrupts the Beclin 1/ Bcl-2 (Bcl-xL) complex, leading to the activation of autophagy [121]. Quercitin sourced from kale, capers and onion, initiates apoptotic cell death in CAL51 and MCF7 cell lines showing its capability to target triple-negative and ER+ breast cancer cell lines [122]. The novel complex, quercetin-zinc(II), induced apoptosis in colorectal adenocarcinoma and hepatocellular cell lines [123] wherein augmented absorption of this complex and activation of anticancer effects accompanied by increase in apoptosis levels were observed. These findings align well with other studies showing penetration of flavonoid metal complexes into lipid bilayers via hydrophobic protein pores, enhancing their intracellular intake [124]. The high expression levels of aldehyde dehydrogenase 1A1 (ALDH1A1) have been correlated with augmented tumorigenicity, invasiveness, clonogenicity and stemness characteristics of breast cancer. Quercitin downregulates ALDH1A1 activity [125]. The transmembrane glycoprotein Mucin 1 (MUC1) is highly expressed in breast cancer cells due to gene amplification and depletion of gene transcription as well as post-transcriptional regulatory networks. MUC1 protein is involved in interaction with EGFRs, activating signaling cascades related to cell proliferation, and leading to cancer cell invasion and tumor metastasis. Quercitin suppresses Mucin 1 (MUC1) expression by inhibiting cell proliferation and cancer metastasis in breast cancer cells. Quercitin downregulates the expression of epithelial cell adhesion molecule (EpCAM) implicated to be actively involved in inducing cancer stemness, cellular proliferation, angiogenesis, metabolism, drug resistance and epithelial to mesenchymal transition (EMT) in breast cancer cells as well as other carcinomas. EpCAM has been associated in crosstalk with p53, PI3K/AKT/mTOR, TGF-β/SMAD, and Wnt/β-catenin pathways in order maintain stemness of carcinomas and overall epithelial tumor survival [126]. Thus, the polyphenol, quercitin found in vegetables and fruits shuts down the stemness of breast tumor progenitor cells [125].
Silibinin, a polyphenolic flavonoid sourced from the seeds of Silybum marianum (or milk thistle) and Cynara scolymus (or artichoke) is the major constituent in silymarin plays several roles to affect various molecular targets present in breast cancer cells. Silibinin interacts with both α and β estrogen receptors (ERs) to mediate anticancer effects. Upon interaction with Erα it influences RAS/ERK and P13K/AKT/mTOR pathways of signal transduction, thereby including autophagy. Its interaction with Erβ enhances apoptosis. Likewise, silibinin blocks metastasis through EMT suppression by inhibiting the expression of TGF-β2. The anti-metastatic effects of silibinin is also associated with the Jak2/STAT3 pathway [127]. It has been reported that silibinin plays a therapeutic role by disrupting homeostasis of TNBC metabolism via modulation of the EGFR-MYC-TXNIP axis [128].
Different epidemiological investigations have shown inverse association between flavonoid consumption and breast cancer. Flavonoids induce tumor-suppressor gene expression resulting in a decrease in the progression of breast cancer and metastasis [129]. Flavonoids influence the pathway of NF-kB signaling. Flavonoids of different types show synergistic effects along with anthracyclines including daunorubicin, idarubicin, epirubicin and doxorubicin [130]. Several cancer types such as sarcoma, lymphoma and breast cancer are treated using anthracyclines which disrupt DNA integrity, bind to the cell membrane and increase oxidative stress mediated by enhancement in the production of free radicals [131]. The interaction between flavonoids and anthracyclines plays a vital role in this process [132].
11. Naphthoquinones and Their Anticancer Activities
The Carya catharsis green husk contains juglone which is known for its anticancer properties. Juglone causes ROS production in human endometrial cancer cells which in turn upregulates the expression of p21 mRNA and protein, concomitant with diminished levels of cyclin A, CHK1, cdc25A and CDK2. Further, it leads to downregulation of Bcl-xL and Bcl-2 and upregulation of cytochrome C, Bax and Bad suggesting its association with the mitochondrial pathway during apoptosis induced by juglone [133].
The natural naphthoquinone, MAM (2-Methoxy6acetyl7methyljuglone) sourced from Polygonum cuspidatum is widely used in Chinese medicine due to its anticancer, antioxidant and anti-inflammatory properties. Therapy with MAM results in necroptosis and production of nitric oxide in A549 lung cancer cells through activation of JNK. This augments peroxidation of lipids leading to the generation of peroxynitrite (ONOO–) which triggers apoptosis [134].
12. Saponins and Their Anticancer Effects
Saponins possess anti-proliferation, anti-angiogenesis, anti-metastasis properties in addition to their capability to reverse multidrug resistance (MDR). Saponins act by inducing apoptosis, promoting cell differentiation, modulating immune function, binding bile acids and ameliorating cell proliferation induced by carcinogens.
A plant saponin called dioscin possesses antitumor, antioxidative, anti-inflammatory and immunostimulatory activities. It reduces breast cancer stemness by arresting the cell cycle through regulation of AKT/mTOR and p38 MAPK signaling pathways [135]. Dioscin elicits the expression of p53 and p21 and blocks the expression of many cyclin-dependent kinases and cyclins.
Ginsenosides (triterpene saponins) obtained from Panax ginseng show remarkable anticancer properties. Ginsenosides can preclude and treat cancer through targeting many molecules and pathways by inducing autophagy, causing death of cancer cells by exercising control over the p53 pathway, neutralizing ROS, modulating miRNAs through decrease of Smad2 expression, regulation of Bcl-2 expression through NF-kB pathway normalization, blockage of inflammatory pathways through reduction of cytokine production, inciting cell cycle arrest by restriction of CDC2 and cyclin E1 and inducing apoptosis of cancerous cells [136].
13. Other Phytochemicals and Their Antitumor Activities
A polyisoprenylated benzophenone, garcinol sourced from Garcinia indica initiates apoptosis in SKBR3A, MDAMB231, and MCF7 breast cancer cell lines through the down regulation of the expression of anti-apoptotic proteins like Bax and Bcl-XL. Garcinol has also been implicated in eliciting cell cycle arrest followed by apoptosis in breast cancer cells overexpressing Her-2 [137] Additionally, it has been shown to cause loss of fragmentation of mitochondria and mitochondrial transmembrane potential leading to apoptosis in MCF-7 cells [137].
Propolis is a compound obtained from plants by honeybees and is widely used in conventional medicine. This natural substance causes ER stress in MCF-7 human breast cancer cells. The CCAAT/enhancerbinding homologous protein (CHOP) in turn elicits apoptosis in response to the ER stress [138].
The phytochemical, thymoquinone, is found in Nigella sativa plants. It has been shown to kill cancer cells and is known to preclude the spread of cancer. Thymoquinone causes autophagic cell death in head and neck squamous cell cancer cells. In human squamous carcinoma cells, thymoquinone elicits cell death through autophagy dependent on LC3-II activation and apoptosis dependent on caspase activation [139]. Thymoquinone caused strong cytotoxicity on the highly malignant head and neck squamous cell carcinoma cell line, SASVO3. It incited apoptotic cell death in these cells as shown by increased caspase-9 activation and Bax expression. In oral cancer cells, it caused cell death by means of anti-neoplastic effects that can elicit autophagy and apoptosis. Thymoquinone blocks bone metastasis associated with breast cancer cells by mediating disruption of NF-kB snd CXCR4 signaling axis [140]. Hence, thymoquinone is a propitious candidate in pathway-targeted, mechanistic and phytochemical-induced cancer prevention approaches.
Zingiber officinale Rosc (6-Shogaol Ginger) contains 6-Shogaol which triggers apoptosis in SMMC-7721 cells (human hepatocellular carcinoma cell line) while at the same time does not affect normal HL-7702 liver cells. Cells exposed to 6-Shogaol for long periods have the potential to undergo ER stress. In these cells, PERK/eIF2α dephosphorylation and induction of the expression of the downstream CHOP generated a caspase cascade effect that resulted in apoptosis [141].
The vitamin E, γ–Tocotrienol is found in annatto seeds, palm oil and rice bran oil. Beta-tocotrienol plays a role in prostate cancer prevention and has been shown to elicit autophagy, apoptosis and necrosis in LNCaP and PC-3 human prostate cancer cells. These activities were also accompanied by enhancement of intracellular dihydrosphingosine and dihydroceramide levels that indicate modulation of the sphingolipid biosynthetic pathway [142].
The Oryza officinalis species contains ω -Hydroxyundec-9- enoic acid (ω-HUA), a hydroxyl unsaturated fatty acid which is known for its anticancer properties towards human non-small cell lung cancer (NSCLC). In cells treated with xHUA, ω-HUA induces ROS. The biosynthesis of CHOP and phosphorylated p-eIF2α was suppressed by ROS along with NAC, revealing that ROS is vital for x-HUA-stimulated ER stress and caspase-enabled apoptosis in NSCLC cells [143].
Extracts from Potentilla species containing tannin, flavonoid and phenolics, exhibit anticancer effects in colon cancer cells by damaging the cell membranes [132,144,145]. Phytosterols regulate serum cholesterol and possess anticancer properties. Carotenoid consumption is inversely related to the recurrence of tumors and death because of breast cancer [146]. Polyphenols induce apoptosis and block hepatocarcinoma cell proliferation [147]. Cyclocarya paliurus contains a dammarane triterpenoid which exhibits anti-inflammatory action by decreasing the expression of IL-6, PGE2 and TNF-α [148].
Carotenoids constitute a diverse category of colorful, yellow, orange and red terpenoid pigments identified in vegetables, fruits, milk, meats, eggs, some crustacean seafoods and fish [149,150]. Lycopene occurs in brilliantly colored vegetables and fruits such as red grapefruits, beets, ripe tomatoes and watermelon [151]. β-Carotene is present in yellow and deep red fruits and vegetables wherein the orange pigmentation of carrots is attributed to β-Carotene. Green vegetables are composed of high quantities of xanthophylls and carotenes; the brilliant red color of peppers is caused by the presence of capsanthine while the red/pink pigmentation of crustaceans is caused by astaxanthin. In addition to their aesthetic characteristics, carotenoids possess antioxidant properties and are vital for the biosynthesis of nutrients like vitamin A [152]. The principal mechanisms by which carotenoids are shown to be implicated in cancer are associated with molecular pathways responsible for the development and death cells [153]. There are over 600 carotenoids including structural variants occurring naturally [154,155] of which the major carotenoids are lutein, lycopene, zeaxanthin, β-Carotene and astaxanthin [156]. Recent studies show that the carotenoid extract containing α-carotene, β-carotene, zeaxanthin and lutein sourced from Dunaliella salina suppressed the biosynthesis of pro-inflammatory cytokine molecules as well as enzymes including COX-2 and NO in LPS-elicited cells. Further, this extract showed anti-inflammatory potential by inhibiting JNK phosphorylation and NF-κB activation [157]. Saffron extract contains an abundance of carotenoids and has been found to diminish the viability of cells responsible for liver cancer in a time- and dose-dependent manner while decreasing cell proliferation, oxidative stress and inflammation. Furthermore, it elicited apoptosis in addition to the downregulation of inflammatory markers like NF-κB-p65, iNOS and COX-2 in vivo [158].
Emodin sourced from herbs has the potential for treatment of triple-negative breast cancer (TNBC) via targeting of transcriptional regulators SerRS and NCOR2 to inhibit the transcription of anti-vascular endothelial growth factor A (VEGFA) as well as tumor angiogenesis in murine models [159]. In investigation by Fu et al., 2020 demonstrated that daunorubicin liposomes modified with arginine8-glycine-aspartic acid (R8GD) along with emodin liposomes also modified with R8GD were strongly cytotoxic to MDA-MB-435S cells in vitro and in mouse models and efficiently suppressed the generation of VM (vasculogenic mimicry) channels and tumor cell metastasis occurring in invasive breast cancer [160]. Additionally, they induced the downregulation of some metastasis-associated proteins such as HIF-1α, TGF-β1, VE-cad and MMP-2. These combined liposomes enabled the chemotherapeutic drug to specifically accrue at the site of tumors, thereby showing a distinct anticancer effect. In yet another study by Shen et al., 2024, emodin was shown to inhibit colorectal cancer cell proliferation and elicit apoptosis [161]. Emodin reduced GSH content, expression of GPX4 and xCT while augmenting the generation of ROS, lipid peroxidation and MDA. Emodin inactivated the NF-κb pathway in these cells and in murine models wherein it inhibited tumor growth and elicited in vivo ferroptosis through ferritinophagy through the inactivation of the NF-κb pathway. Thus, emodin has been proposed as a favorable candidate for anticancer therapy. Table 1 enlists some of the phytochemicals currently under clinical trials.
Taken together, several phytochemicals use various mechanisms to enable suppression of the survival and growth of cancer cells and target pathways such as PI3K/AktmTOR, JAK/STAT pathways, Hedgehog, Notch, Wnt/β-catenin Hippo signaling pathways leading to shut down of cancer cells followed by suppression of the heterogeneity, aggression and remission of tumor cells [23].
14. Phytochemical-Based Nanoparticles in Cancer Prophylaxis and Therapy
The confluence of traditional healing wisdom and cutting-edge technology has given rise to phototherapy as a promising solution, offering targeted and minimally invasive strategies for the intricate battle against cancer [170]. Simultaneously, the rich reservoir of bioactive compounds inherent in various plant species has seized considerable attention due to its potential to mitigate the complex dynamics of cancer progression. Photo-mediated therapies such as photothermal therapy (PTT) and photodynamic therapy (PDT) have been found to be effectual in cancer therapy and operate by distinct damage mechanisms involving the production of heat and ROS respectively [171,172], which results in cellular death. Therefore, both PDT and PTT can be applied to treat many types of cancer [173,174].
PDT involves the use of photosensitizer drugs that can be stimulated by radiation. PDT is scarcely or non-invasive and very successful anticancer treatment strategy for several types of cancer [175]. PDT requires a light source, a photosensitizer (PS) drug and availability of oxygen whose interaction generates ROS [176]. Upon exposure to a specific wavelength of light, the PS absorb photons which results in conversion of the PS to its excited state as indicated in Figure 2. Subsequently, it crosses to a metastable triplet state that leads to type I PDT wherein the PS in its activated state can elicit a series of reactions involving biomolecules that produce radicals capable of reacting with molecular oxygen thereby generating ROS. In type II PDT, the PS directly transfers energy to oxygen which leads to elicitation of ROS [177,178]. These ROS molecules have powerful oxidizing and cytotoxic effects.
Curcumin is a polyphenol known for its antitumor effects and photosensitizing characteristics [179]. Curcumin has been encapsulated within solid lipid nanoparticles towards use in phototherapy. These nanoparticles demonstrated augmented uptake of the drug into cancerous lung cells resulting in elicitation of ROS under exposure to light and this proved to be propitious towards phototherapy [180]. Nano-emulsions harboring curcumin as the photosensitizer drug were demonstrated to be greatly phototoxic towards breast cancer cells in addition to eliciting ROS at high levels [181]. Preparations of nano-emulsions containing acai oil were combined along with irradiation of light which caused death of cancerous melanoma cells up to 85% [182]. This was further substantiated when the said nano-emulsion was shown to result in the reduction of tumor volume in murine models. A conjugate containing cyclodextrin and chlorophyll α when applied to colorectal human adenocarcinoma cells proved to be toxic to these cells under photo-induction thus enabling PDT applications [183].
Another therapeutic strategy, PTT uses near-infrared laser / light (NIR) to elevate the temperature within the tumor site and elicits death of cancer cells [184]. Other sources of radiation to cause hyperthermia involve microwaves, visible light, ultrasound and radiofrequency waves [185]. PTT exhibits high specificity while being barely invasive [186]. The PTT strategy operates through two mechanisms: one includes the exposure of the tumor site to elevated temperatures (over 45C) for a few minutes resulting in tumor cell death via thermal ablation, stasis within tumor vessels as well as hemorrhage that preclude its administration with other treatment strategies; the other includes the induction of mild hyperthermia wherein temperatures of 42-43C are set up that lead to tumor cell damage and augmented tumor vessel permeability that can be applied to promote the uptake of nanoparticles by tumors [172,187,188]. Tumor tissues show greater acidity and hypoxicity when compared to normal tissues [189] which make them more vulnerable to high temperatures thereby enabling PTT to destroy tumor cells selectively while protecting healthy cells around the tumor area [190]. This facilitates PTT administration along with other synergistic therapeutic strategies.
Single-walled carbon nanotubes containing polyvinylpyrrolidone and phosphatidylcholine were functionalized for delivery of curcumin which showed augmented curcumin delivery into cancerous cells within 4 hours [191]. This formulation showed enhanced uptake of curcumin up to 6-fold greater than native curcumin and increased the blood concentration of curcumin by as high as 18-fold. This photothermal ablation effect was further demonstrated in in vivo models wherein it led to reduction in tumor volume and weight.
Phytochemical compounds have been used along with magnetic nanoparticles towards phototherapy as well as drug delivery [192]. Iron oxide nanoparticles capped with eugenate (4-allyl-2-methoxyphenolate) were generated via green synthesis using Pimenta dioica, a medicinal plant [193]. This formulation demonstrated favorable biocompatibility in human embryonic kidney 293 cell line (HEK293) and the human cervical cancer (HeLa) cell line in addition to showing robust efficacy of hyperthermia production upon irradiation with laser at near infra-red (NIR) wavelength. Curcumin was loaded within Fe3O4 magnetic nanoparticles coated with silica towards generating singlet oxygen and hyperthermia [194]. Combined treatment with this formulation and PDT resulted in reduction of tumor volume by 58% while a combination of these nanoparticles with PDT and PTT showed 80% reduction in tumor volume which was attributed to the synergistic effects obtained by ROS production and hyperthermia at the tumor site [194]. EGCG when combined with PDT augments anticancer effects in vitro and in vivo [195].
Selective targeting of cancer cells occurs through various mechanisms. One approach involves the presence of specific markers on the surface of cancer cells that facilitate the preferential uptake of photosensitizers (PS) by cancer cells compared to healthy cells. Additionally, the tumor microenvironment, characterized by factors such as hypoxia and acidity, can enhance the sensitivity of cancer cells to ROS-induced damage, leading to selective destruction of cancer cells while sparing healthy cells. These combined factors contribute to the effectiveness of phototherapy in targeting and eliminating cancer cells while minimizing harm to surrounding healthy tissues [172].
15. Negative Effects of Phytochemicals
Certain phytochemicals have been found to be toxic to humans and these phytotoxins have been shown to act as anticholinergic, adverse gastrointestinal irritants, cyanogens, cardiac glycosides, stimulants of the central nervous system and hallucinogens [196]. Some polyphenols have been associated with genotoxic / carcinogenic effects and were shown interfere with the biosynthesis of thyroid hormone [197].
Neuropathy, specifically peripheral and autonomic sensory-motor neuropathy is a dose-circumscribing and dose-dependent negative effect often demonstrated in cancer patients undergoing vincristine treatment [198]. Neuropathy could be classified as chronic or acute and typically emerges within two weeks of initiation of treatment. The major common type, Vincristine-induced peripheral neuropathy (VIPN) frequently manifests as muscle weakness, paresthesia, areflexia, neurotic pain, wrist and foot drop [199].
Although several flavonoids can affect the normal function of the thyroid gland, phytoestrogens are the major substances of concern interfering with thyroid metabolism and function. Several studies have shown that quercetin and phytoestrogens could induce thyroid disruption [200]. Their toxic effects include mutation and carcinogenicity, kidney and liver toxicity, negative effects on thyroid and reproductive functioning and in addition to elicitation of disorders in intestinal flora. The toxicity mechanism is complex, and currently available evidence shows that naturally occurring flavonoid glycosides act on various targets at different doses in vitro and in vivo. Although most categories of flavonoids have been deemed safe, flavonoids recommended as food supplements must be assessed for tolerable maximal intake level due to reports of flavonoid toxicity.
Capsaicin functions as a co-carcinogen in 12-O-tetradecanoylphorbol-13-acetate (TPA)-promoted carcinogenesis of the skin in vivo and therefore caution must be exercised when administering capsaicin for topical application on a prolonged basis, particularly in the presence of cancer promoters including solar UV radiation exposure [201].
Cycasin as well as its metabolite namely, methylazoxymethanol (MAM), are usually extracted from roots and seeds of cycad plants [202]. These plants are potently poisonous and the toxicity due to ingestion of seeds is primarily caused by its misuse as a food source, as an agent to augment health, for precluding cancer for cosmetic purposes and for the treatment of gastrointestinal disorders. MAM is considered a genotoxic metabolite and has been shown to be involved in targeting of cellular processes associated with cancer development and neurodegeneration [203].
Genistein is an isoflavone phytoestrogen occurring in soybeans, fava beans and red clover. Another principal category of phytoestrogens are lignans of which matairesinol is found in several foods such as fruits, vegetables, whole grains and oil seeds [204]. Dietary phytoestrogens have been shown to contribute to the development of colorectal cancer in women and prostate cancer in men [205]. Localized production of estrogen is catalyzed by the enzyme aromatase which is differentially regulated in healthy and cancerous breast tissue. Soy supplements used to ameliorate menopausal symptoms have been shown to elicit the growth of MCF-7 breast cancer cells by increasing aromatase biosynthesis and activity associated with breast cancer [206]. Particularly, genistein has been shown to obstruct the inhibitory activities of aromatase inhibitors including letrozole [207] and fadrozole [206] against growth of MCF-7 breast cancer cells in a xenograft model and in vitro respectively. Hence, women under treatment with aromatase inhibitor must be cautioned against the consumption of soy products.
Aristolochic acids constitute a category of compounds used in traditional herbal remedies since ancient times. They are known for their anticancer effects [208] However, many studies have shown that aristolochic acid exposure leads to high occurrence of cancer involving the urinary tract and kidney [209].
Tetracyclic diterpenoids belong to the phorbol ester category of compounds known for their robust tumor promoting effects. The seed-derived oil of the Croton plant has been used in several herbal medicines for years and contain Phorbol 12-myristate 13-acetate which reportedly increases neutrophil and white blood cell counts in patients harboring solid tumors [210]. It also interferes with the migration and proliferation of thyroid cancer cells [211] and inhibits the growth of prostate cancer cells when used in combination with the anticancer drug, paclitaxel [212]. Nevertheless, it is also known to potently promote skin cancer [213]. Therefore, if this compound is used appropriately, it can treat lymphomas and leukemia despite their potential to induce skin cancer.
Pyrrolizidine alkaloids (PAs) riddelliine or comfrey occur in teas and are likely the most used among herbs in the present times. PAs have been reported to elicit liver cancer [214] in animal models. Dehydro-PAs interact with DNA and cellular proteins causing cancer and genotoxicity. Particularly, they cause skin cancer by generating ROS leading to lipid peroxidation [215].
Taken together, consumption of some phytochemicals can induce carcinogenesis, and the internet affords a huge marketplace for such kinds of products. Clinically notable adverse reactions to several unconventional remedies obtained via the internet have been observed [216,217]. Therefore, consumers need to be conscious that dietary supplements consisting of phytochemicals and related compounds are practically unregulated due to which manufacturers of such products are not required to demonstrate the health benefits and safety of these products before their release into the market.
16. Expression of Monoclonal Antibodies (Mabs) in Plants
There are other ways that plants can be a preferable solution to the world’s most pressing problems. Low cost, high scalability, and low risk of human pathogen contamination are the hallmarks of plant-based systems for producing Mabs [218]. Glycoengineering of plant hosts offers another advantage over mammalian cell-based systems to produce Mabs. Owing to their pluripotency, plants have the capability for regeneration from somatic cells [219]. Plants have been used for both transient and stable expression [218,220].
17. Stable Expression of Recombinant Mabs in Transgenic Plants
For stable transformation, particle bombardment and Agrobacterium-mediated transformation have been employed to enable the direct penetration of the coding sequence of antibody light and heavy chains into plant cells facilitating stable insertion of the cDNA into plant genomes [220]. Transgenic plants biosynthesizing either of these light or heavy chains can be chosen and crossed to develop transgenic lines that express both antibody chains. Recent, more successful and quicker strategies use binary vectors having both heavy and light chain encoding sequences within the same T-DNA. Hence, complete IgGs can be expressed through a single round of transformation in transgenic plants [221].
Transgenic plants are the most appropriate plant-based system for Mab production on a large scale. Vertical farming schemes or greenhouses are promising approaches for mass generation of Mabs in transgenic plant systems considering the fact that open field cultivation may not be suitable due to the lack of containment and inability to ensure adequate biosafety for Mab production [222].
Antibodies genes can be introduced into the genome of chloroplasts to produce chloroplast transgenic plants that express antibodies having proper folding in addition to disulfide bonds [223]. Such chloroplast transformation dispenses with pollen transmission of the transgenes in addition to displaying high level expression due to the inherently high polyploidy of the chloroplast genomes. Numerous plastids harboring multiple copies of the transgene can exist within a single plant cell leading to robust chloroplast expression as high as up to 25% of soluble plant proteins. Furthermore, chloroplast transformation dispenses with gene silencing as well as position effects. Therefore, if appropriate glycosylation is developed in the chloroplast, their transformation could emerge as a reliable expression system for stable production of anticancer antibodies [223].
18. Transient Expression of Plant Based Mabs
Mabs have also been generated by transient expression using recombinant plant viruses and agroinfiltration. Plant virus vectors can be employed for transient Mab expression more speedily than that of transgenic plants. For this purpose, viral vectors can be easily inoculated to rapidly express single-chain antibodies (scFv) that are customized for individual cancer patients distinguished by unique epitopes [224]. Further, TMV vectors harboring light and heavy chains have been used to express full-sized Mabs in N. benthamiana [224,225]. The expression platform commercially known as ‘MagnICON’ proved to be very effective for the high-yield production of Mabs in N. benthamiana. A successful example of the exploitation of this technology is the Phase 1 clinical study conducted on chimeric antibodies for the treatment of B cell follicular lymphoma essentially by using these molecules as individualized idiotype vaccines created from patient’s own cancer cells [226]. The rapid and high-level expression offered by plants permitted to proceed from biopsy to the individualized vaccine in less than 12 weeks [227].
Nevertheless, the plant viral scheme necessitates inoculation of the virus into the stem or the leaf every time owing to its transient expression and additionally this could often lead to gene mutation occurring in the process of virus replication in contrast to the stable transgenic expression system [228]. This calls for careful evaluation of the choice of the technique used for antibody gene expression [229].
Transient expression through epichromosomal transformation is not heritable and is therefore a batch-wise process. This decreases the hazard of environmental biosafety concerns associated with transgene propagation via pollen or seeds. Typically, high quantities of protein are expressed using this strategy within a brief period of time, commonly a few weeks or days which cannot be achieved using stable transformation [230]. Since each plant within a given batch has to be infiltrated with Agrobacteria, the major drawback of this quick expression methodology for large scale production is that this process of infiltration simply transfers the costs of bacterial fermentation [231].
19. Plant Cell Cultures for Mab Production
Additionally, other plant-based systems such as hairy roots cultures, plant tissue cultures, plant cell suspension cultures, and aquatic plants can be used similar to mammalian cell cultures. Such systems afford an efficient strategy for synthesis of heterologous proteins under sterile conditions accompanied by low risks of contamination by mammalian factors and pathogens. Further, both plant organs and transformed plant cells can be grown indefinitely while necessitating only simple nutrients. The candidate Mab proteins could be secreted into the prevailing culture medium which enables facile harvest and purification. However, poor protein yields and complexities in establishing large-scale recombinant Mab expression in bioreactors are the primary challenges for the use of these plant-based expression platforms in the future [222,230].
20. Recent Developments Involving the Expression of Plant-Based Monoclonal Antibodies Against Cancer
Bulaon et al., 2024, designed and produced a plant-based bispecific monoclonal antibody (bsAb) capable of recognizing both cytotoxic T-lymphocyte-associated protein 4 and programmed cell death ligand 1 within a single molecule called dual variable domain immunoglobulin atezolizumab × 2C8 [232]. This bsAb was expressed transiently in N. benthamiana wherein it demonstrated capability to bind cytotoxic T-lymphocyte-associated protein 4 and programmed cell death ligand 1 proteins in vitro. This antibody significantly blocked tumor growth in murine models harboring CT26 colorectal tumor while being tolerable and safe. Table 2 shows some examples of the recently generated Mabs in plant systems.
For many cancer malignancies, Durvalumab (called Imfinzi) that targets PD-L1 is currently being employed for immunotherapy. This IgG1 antibody Fc region has been genetically engineered to decrease FccR interactions towards enhancing the inhibition of interactions between PD-1 and PD-L1 without depleting immune cells expressing PD-L1 [240]. N. bethanmiana was engineered to express four Durvalumab variants namely, the wild-type IgG1 and LALAPG, its ‘Fc-effector-silent’ variant harboring modifications to enhance antibody half-life, as well as IgG4S228P and PVA, its variant having Fc mutations to reduce FccRI binding. Additionally, Durvalumab variants were generated in their afucosylated form and their decorated form with 1,6-core fucose [240]. Plant-based durvalumab variants interact with recombinant PD-L1 as well as gastrointestinal cancer cell PD-L1 to effectively inhibit their binding with PD-1 on T cells, resulting in enhancement of their activation. Moreover, those plant-derived antibody variants harboring core fucosylation and Fc amino acid mutations show positive impacts on their therapeutic potential. In comparison with Imfinzi, DL-IgG4 (PVA) S228P and DL-IgG1 (LALAPG) exhibit lesser affinity for the CD32B inhibitory receptor that can be therapeutically advantageous. Significantly, DL-IgG1 (LALAPG) demonstrated augmented FcRn binding, a vital determinant of IgG serum half-life [240].
Conventional Mabs like Trastuzumab face shortcomings during treatment of Human Epidermal Growth Factor Receptor 2 (HER2)-positive breast cancer, specifically in patients who develop drug resistance. Park et al., 2024, report a study wherein they express a plant-produced anti-HER2 variable fragments of camelid heavy chain domain (VHH) fragment crystallizable region (Fc) KEDL(K) antibody showing capability to function as a potent alternate treatment to overcome such limitations [244]. This plant-derived antibody proved to have specifically high affinity for breast cancer cells that are HER2-positive inclusive of those that are resistant to Trastuzumab. Further, in mice that are immune-deficient, this plant-based anti-HER2 VHH-FcK antibody shows superior anticancer activity, particularly against Trastuzumab-resistant tumors, underscoring its potential as effective immunotherapy for HER2-positive breast tumors that are Trastuzumab-resistant.
Jin et al., 2023, expressed anti-human epidermal growth factor receptor 2 (HER2) VHH-FcK mAbs and anti-colorectal cancer large single chain (LSC) CO17-1AK through cross-pollination of plants expressing anti-HER2 VHH-FcK and LSC CO17-1AK respectively, both of which targeted proteins in SKBR-3 human breast and SW620 human colorectal cancer cell lines correspondingly and inhibiting cell migration to levels equivalent to that of their respective parental antibodies [245].
Bulaon et al., 2023, report the production of anti-CTLA-4 antibody, 2C8 by rapid transient expression in N. benthamiana plants [237]. This anti-CTLA-4 2C8 mAb interacts with murine and human CTLA-4 proteins with comparable efficiency to that of one of the Fcg receptors. Additionally, it demonstrated equivalent antitumor efficacy to that of the commercially available anti-CTLA-4 mAb (Ipilimumab Yervoy®) in a humanized murine tumor model, implying that the plant-derived anti-CTLA-4 mAb has similar therapeutic potential to that of the clinically efficient Ipilimumab. Considering that Ipilimumab is expensive and unaffordable in developing world, plant-based production of anti-CTLA-4 2C8 mAb is more appealing for rapid expression, facile scale-up and economical manufacture of such recombinant therapeutics.
Immune checkpoint inhibitors (ICIs) are a category of immunotherapeutic agents with capability to alleviate the immunosuppressive environment exerted by neoplastic cells. Tumorigenic cells use one of the most universal checkpoints, the programmed cell death protein 1 (PD-1)/programmed death-ligand 1 (PD-L1) for evading the immune system by eliciting apoptosis and blocking cytokine production and proliferation of T lymphocytes. Presently, the most commonly used ICIs that target the PD-1/PD-L1 checkpoint are Mabs nivolumab and pembrolizumab that interact with PD-1 occurring on T-lymphocytes and block the binding with PD-L1 expressed on tumorigenic cells.
Nivolumab and pembrolizumab are the most commonly used ICIs for treatment of several cancers such as Hodgkin lymphoma, melanoma, lung, breast and colorectal cancers [246,247,248]. These two antibodies block the PD-1/PD-L1 immune checkpoint resulting in CTL activation and apoptosis induction in tumorigenic cells via T-cell-enabled cytotoxicity [159,249]. Both of these ICIs significantly augment the rates of survival of patients having a diverse range of cancer types. Nevertheless, the cost of these therapies currently in the market is exorbitantly high and therefore inaccessible to patients in developing countries [250], primarily owing to the expensive mammalian cell platform used for their expression. Hence, plant-based molecular farming of these antibodies is appealing due to its potential to greatly decrease the capital required for the manufacture of these ICIs [238,247]. Nivolumab and pembrolizumab have been transiently expressed in N. benthamiana leaves at yields as high as 140 mg/kg FLW and 340 mg/kg FLW respectively, which amounted to USD 4200 and 18,000 worth of these 2 antibodies correspondingly in 1 kg of leaves [242].
The first antibody that targeted the immune checkpoint PD-L1 was Atezolizumab (Tecentriq) which is currently one of the most widely used drugs in anticancer therapy. Nevertheless, this anti-PD-L1 antibody is expressed in mammalian cells that incurs high costs for manufacture, circumscribing the access of antibody treatment for cancer patients. Plant-based Atezolizumab upon transient expression in N. benthamiana showed high levels of expression within 4-6 days following infiltration. Purified plant-expressed Atezolizumab had no glycosylation and was able to bind to PD-L1 with equivalent affinity to that of Tecentriq [238]. Additionally, this plant-produced antibody inhibited tumor growth with an efficacy comparable to that of Tecentriq in murine models. This substantiates the capability of plants to serve as efficacious platforms for production of immunotherapeutic antibodies and therefore plants can be used to mitigate the cost of currently used anticancer drugs [238].
Varlilumab, a CD27-targeting monoclonal antibody, in its recombinant form was expressed in leaves of N. benthamiana within as little as 8 days of infiltration and was shown to assemble successfully [243]. The RNA silencing suppressor, p19 was co-expressed along with Varlilumab which resulted in the accumulation of the Mab averaging to about 174 µg/g of fresh leaf weight. This was greater when compared to the yield of nivolumab (140 µg/g of fresh leaf weight) produced in transient geminiviral expression system in N. benthamiana [248]. Purified plant-produced Varlilumab assembled properly into a tetramer and showed in vitro efficacy comparable to that of the commercial Varlilumab expressed in mammalian cells.
21. Plant-Based VNPs and VLPs Against Cancer
Plant viruses are a group of pathogens that infect plants. They contain a protein cage (capsid), and nucleic acid (RNA or DNA) [251,252]. The capsid naturally enclose the nucleic acid via a straightforward supramolecular self-assembly mechanism from many copies of one or a few different types of coat protein (CP) subunits [252]. CP subunits without the nucleic acid can assemble as virus-like particles (VLPs) under in vivo or in vitro conditions [253]. Thus, two key nanostructures based on plant viruses, as plant virus nanoparticle (PVNP) are available: the complete plant virus virion or virus nanoparticles (VNPs) and VLPs. PVNPs can be naturally isolated from plants, or produced by ‘molecular farming [254]. PVNPs have various morphologies, including icosahedral, road like, bacillus, and filamentous. PVNP’s noninfectious nature in animals, biocompatibility, biodegradability, and non-teratogenic properties significantly reduce their in vivo toxicity [255]. PVNPs can modify via genetically and physicochemical strategies to produce modified PVNPs [252]. Genetic modification involves incorporating non-native biological elements (specific amino acids, peptides, tags, and proteins) into the CP of PVNPs, or removal of residues from CP [256]. Genetic modification can be performed for displaying specific ligands or further modifications [256]. Chemical modification techniques depend on the utilization of activated conjugates to react with functional groups of natural or unnatural amino acids that are found on the capsid, within specific solution conditions of pH and salt concentration. Another method for manipulating is open pore structures on PVNPs. Pores are responsive to environmental stimuli, and undergoing conformational changes, allowing controlled molecule entrapment [256]. For instance, Red clover necrotic mosaic virus (RCNMV) encapsulates cargo through infusion triggered by stimulus-induced capsid pores opening and closing [257]. Self-assembly is an effective method for engineering of PVNPs, where purified CPs assemble around a specific cargo [251]. The extent of PVNP modification depends on charge, shape, size, shielding, and targeting, which are all influenced by various factors such as receptors or environmental factors [256]. The potential of PVNP-based modifications exhibit diverse yet distinct dimensions and configurations, making them highly customizable in terms of size and shape, and capable of loading various natural and synthetic payloads [252,256,258]. In context of cancer, modification of PVNPs can be applied to create and improve a variety of therapies including chemotherapy, gene therapy, immunotherapy and vaccines.
22. PVNPs as Delivery Nanosystem in Cancer
Delivery nanosystems are widely used to improve the safety and efficacy of encapsulated therapeutic/imaging agents [259,260]. Recently, PVNPs have been utilized as an innovative platform for delivery applications [256]. They can address therapeutic/imaging agent’s limitations by encapsulating them on their interior/exterior surfaces, and enhancing their effectiveness and safety [252,256]. Structurally, untargeted PVNP formulations have the potential to exhibit enhancement in their concentration within tumors via passive-targeting delivery, utilizing the "enhanced permeability and retention (EPR) effect [252]. However, the most important challenge of non-targeted PVNPs is clearance by phagocytes, even when modified with Polyethylene glycol (PEG), due to the existence of anti-PEG antibodies. To address this concern, a potential strategy involves the conjugation of serum albumin (SA) to PVNP -based nanocarriers. An investigation carried out on Balb/C mice revealed that SA-conjugated TMV, which was "camouflaged" using SA, exhibited decreased antibody recognition and enhanced pharmacokinetics. Furthermore, the attachment of specific ligands to the PVNP surface allows for the targeted delivery of PVNP formulations to target sites. Such ligands include GE11 (short peptide comprising 12 amino acids) [261], folate [262], Herclon (a monoclonal antibody (mAb)), peptide F3, epidermal growth factor-like domain 7 (EGFL7) [263] and CooP (a 9 aa peptide, sequence: CGLSGLGVA) [264].
Drug delivery: Anticancer therapeutic agents face numerous challenges including high resistance, frequent recurrence, rapid drug elimination, and non-targeted distribution, leading to toxicity and limited clinical effectiveness. The use of targeted delivery systems and dose reduction have the potential to enhance therapeutic outcomes. Recently, it was shown that PVNPs can be ideal for loading and delivery of anticancer agents (e.g. nucleic acids, peptides, proteins, small molecules and nanoparticles). They protect encapsulated payloads from degradation, facilitate their targeted delivery, selective cytotoxicity and sustained efficacy even at very low doses. Previous work has shown that PVNPs can encapsulate small molecule therapeutics (DOX, MTO, phenanthriplatin, gemcitabine, and cisPt), enhancing their accumulation in tumor tissues and causing cytotoxic effects (Figure 3A) [265,266,267,268]. For example, the loading of DOX utilizing CPMV, RCNMV, Johnson grass chlorotic stripe mosaic virus (JgCSMV), or the prodrug DOX via Physalis mottle virus (PhMV)-based VLPs have demonstrated significantly improved antitumor activity both in vitro and in vivo [269,270,271]. Notably, the DOX-loaded PVNPs elicited HCC cell death at IC50 values of 10–15 nM, which is 20 times more effective than free DOX. Studies find that the TMV can transport cisplatin and mitoxantrone, demonstrating exceptional cytotoxic effects, making it a promising candidate for reducing anticancer medication dosages (Figure 3A) [266,272].
Additionally, CPMV was utilized for the targeted delivery of mitoxantrone to treat glioblastomas, resulting in increased cytotoxicity against glioma cells [276]. TMV has been used to attach valine-citrulline monomethyl auristatin E, an antimitotic drug, to target non-Hodgkin's lymphoma. The TMV formulations showed potent cytotoxic effects against non-Hodgkin's lymphoma cell lines in vitro, with an IC50 value of around 250 nM. This targeted delivery capability was used to enhance the potency and effectiveness of phenanthriplatin, a cationic mono-functional DNA-binding Pt(II) anticancer drug, resulting in a significant reduction in tumor size [267,277,278]. Filamentous plant virus nanoparticles have shown potential for displaying protein drug delivery in cancer therapy. For example, PVX nanocarriers -based TRAIL/Herceptin delivery has shown enhanced therapeutic efficacy in cancer treatment through targeting specific receptors. The use of PVX nanocarriers for Herceptin delivery in breast cancer treatment has shown enhanced therapeutic efficacy, as it effectively induces apoptosis in HER2 positive cell lines, demonstrating the potential of this innovative approach [279]. Potato virus X has been used as a nanocarrier to deliver TRAIL, a protein drug that induces apoptosis in cancerous cells. This method reduced tumor growth in mouse models of human triple-negative breast cancer (Figure 3B) [273]. Le et al. developed a nanocarrier using potato virus X for targeted doxorubicin delivery, showing efficacy against ovarian, breast, and cervical cancer cell lines, inhibiting tumor growth in mouse models [265,280]. Recently, TBSV-based PVNPs have shown potential in drug delivery to Sonic hedgehog medulloblastoma. Targeted TBSV with CooP (a 9 aa peptide, sequence: CGLSGLGVA) (TBSV-CooP), selectively transports DOX to medulloblastoma, enhancing cell proliferation and apoptosis [264].
The bioconjugation process, involving the linking of epidermal growth factor-like domain 7 (EGFL7) (a protein predominantly expressed in endothelial cells) to CPMV, yielded a modified protein capable of targeting tumor-associated neovasculature with a remarkable level of precision [263]. Recently, a peptide-guided tomato bushy stunt virus (TBSV)-based nanocarrier system loaded with DOX has been utilized for cell-specific delivery. Marchetti et al. (2023) harnessed TBSV-based VNPs with CooP peptide, a homing peptide designed for medulloblastoma tumors. Encapsulating DOX within TBSV-CooP enhanced cell death and proliferation, underscoring their effectiveness in targeting brain tumors. The internalization of the TBSV-based nanocarrier platform, equipped with the C-terminal C-end rule (CendR) peptide RPARPAR (RPAR) (TBSV-RPAR), loaded with DOX, displayed selective cytotoxicity towards cells expressing the receptor neuropilin-1 (NRP-1) [281]. Another research effort showcases the utilization of iRGD peptides to target tumor neovasculature on PhMV-like nanoparticles, resulting in rapid uptake and heightened tumor homing. This strategy presents a promising avenue for delivering targeted molecular cargo to tumors [282].
PVNP can act as PTT and PDT agents in the treatment of tumors. PVNP-based PTT/ PDT agents, which absorb photons, have the capability to produce heat or ROS to eliminate cancer cells [283,284]. For example, the conjugation of a photothermal biopolymer polydopamine (PDA) on TMV followed by exposure to near-infrared laser in combination with immunotherapy and multimodal magnetic resonance/photoacoustic imaging presents a promising strategy and theranostic approach for cancer models in vivo [285]. Incorporating the porphyrin-based photosensitizer drug, Zn-Por, into TMV and tobacco mild green mosaic virus (TMGMV), has been proven to lead to a notable enhancement in the effectiveness of cell destruction. Notably, a five-fold increase in efficacy was observed in comparison to the free drug [284]. Researchers have developed a drug delivery approach using TMV protein nanotubes as a carrier for ovarian cancer treatment. The nanochannel was loaded with cisplatin, achieving a loading efficiency of 2700 cisPt2+ per TMV. TMV-cisPt showed superior efficacy against ovarian tumor cells, reducing tumor burden and increasing survival in mouse models [286].
Gene delivery: Nucleic acid therapeutics are widely used for treating diseases, but the main challenge is the safe and effective delivery of nucleic acids. Initially, mammalian viruses were preferred, but concerns about immunogenicity and integration led to alternative delivery methods like PVNPs, which combine viral and nonviral characteristics [287]. PVNPs have the capability to enhance suboptimal cellular uptake, instability caused by nucleases, and inefficiencies in the delivery of nucleic acids by encapsulating various types of RNA, such as heterologous RNA, siRNAs, mRNA, and CpG-ODNs [288]. Lam et al.'s research marked a milestone in PVNP-mediated genetic therapy, showing that an icosahedral PVNP can target and deliver a siRNAs targeting green fluorescent protein (GFP) or Forkhead box protein A1 (FOXA1). The study shows that siRNA molecules can be loaded into cowpea chlorotic mottle virus (CCMV) nanoparticles, and only CCMV with appended cell penetrating peptide (CPPs), such as M-lycotoxin peptide L17E, was effective in silencing the FOXA1 gene [289]. It has been demonstrated that VLPs with CCMV capsid along with mRNA-EGFP as cargo and reporter gene, have ability to directly transfect eukaryotic cell lines without adjuvants and deliver nucleic acid for translation [290]. CP-miR-26a (CP26a) VLPs were created by self-assembling purified CP from CCMV, retaining its structure and protecting miR-26a from digestion. CP26a showed similar cellular uptake efficiency, osteogenesis promotion ability, and better biocompatibility compared to Lipofectamine2000-miR-26a [291]. An illustration of this is seen with brome mosaic virus (BMV) and CCMV, which can be utilized to carry the siRNA Akt1 (siAkt1) for uptake by tumor cells [292]. Furthermore, CCMV, in combination with siRNAs designed to target FOXA1, a crucial transcription factor in the forkhead box (FOX) protein family, has been shown to induce gene silencing in the MCF-7 breast cancer cell line [289]. CCMV can be used for gene delivery, with a virus-like particle (VLP) encapsulating anti-miR-181a oligonucleotides to knock down ovarian cancer cells (Figure 3C). This study showed higher knockdown efficacy and reduced cancer cell invasiveness, highlighting the potential of plant-derived VLPs as nucleic acid carriers [274].
23. PVNPs as Imaging Agents
Nanoengineering of PVNPs presents a myriad of possibilities for the loading and modification of contrast agents [293]. Guanidinium agents, referred to as PVNP-based dyes, are commonly employed in preclinical diagnostic imaging. These contrast agents, based on PVNPs, hold promise for the development of features such as prolonged circulation, specific targeting capabilities, and efficient delivery to tumors in vivo. An example of this is the utilization of PhMV-like nanoparticles loaded with the fluorescent dye Cy5.5 and paramagnetic Gd(III) complexes, with PEGylated particles being linked to targeting peptides to monitor a human prostate tumor model using near-infrared fluorescence and magnetic resonance imaging (Figure 3D) [275]. PVNPs can also be adorned with bombesin peptides, PEG, and near-infrared fluorescent dyes [294]. Through the loading of Dy3+ and Cy7.5 into TMV nanoparticles and their conjugation with a Dy3+ dye and near-infrared fluorescence (NIRF) dye, significant transverse relaxation of targeted PC-3 prostate cancer cells and tumors was successfully achieved in vitro and in vivo under ultra-high-strength magnetic fields [295]
CPMV loaded with near-infrared dye (Alexa Fluor 647) and PEG, in addition to conjugation with the pan-bombesin analog, [β-Ala11, Phe13, Nle14] bombesin-(7–14), has the ability to specifically target the gastrin-releasing peptide receptor which is known to be highly expressed in human prostate cancers. The process of tumor homing was observed by utilizing human prostate tumor xenografts on the chicken chorioallantoic membrane model, employing intravital imaging techniques [296]. The elongated PVX can be genetically modified to exhibit either green fluorescent protein (GFP) or mCherry as markers for optical imaging in human cancer cells and in a preclinical mouse model [297]. Plant viruses, particularly TMV, can serve as a foundation for contrast agents in magnetic resonance imaging (MRI); TMV particles can be filled with Gd(DOTA) within the internal channel of TMV and the surface covered with silica, thus enhancing T1 relaxivities in comparison to uncoated Gd-loaded TMV [298]. The display of GE11 on PVX and the attachment of PVX-GE11 filaments with fluorescent markers can be precisely aimed at the epidermal growth factor receptor (EGFR). The detection and visualization of cells were illustrated using cell lines of colorectal adenocarcinoma, human skin epidermoid carcinoma, and triple-negative breast cancer (A-431, HT-29, MDA-MB-231), all of which displayed varying levels of EGFR upregulation [261].
24. PVNPs as Theranostic Agents
The distinctive structural and chemical characteristics of PVNPs render them highly appropriate for the integration of therapeutic and diagnostic agents' capabilities for in vivo applications [299]. Metal-phenolic networks (MPNs) derived from plant viruses like TMV, PVX, and CPMV have demonstrated advantageous optical properties, cytocompatibility, and remarkable cell-destructive performance during photothermal therapy, particularly when loaded with complexes of tannic acid (TA), metal ions (e.g., Fe3+, Zr4+, or Gd3+), or fluorescent dyes (e.g., rhodamine 6G and thiazole orange) and exposed to 808 nm irradiation [300]. Gd-loaded TMV particles coated with polydopamine (PDA) inspired by mussels represent biocompatible nanotheranostic agents that facilitate multimodal imaging and photothermal therapy (PTT) in PC-3 prostate cancer cells [301]. The utilization of SA-coated TMV laden with chelated gadolinium (DOTA) for detection via magnetic resonance imaging and the loading of DOX could enable the monitoring of disease progression, thus offering insights into the effectiveness of the drug delivery approach [301]. Engineered TMV-MOF (metal-organic framework) hybrid nanoparticles enhanced the retention of these VNPs in murine models [302]. By coating the TMV encapsulated with Cy5 with zeolitic imidazolate framework-8, particles of Cy5-TMV@ZIF were created, resulting in a 2.5 times higher fluorescence retention time compared to Cy5-TMV alone. These Cy5-TMV@ZIF particles exhibited resistance to harsh conditions, were non-toxic, and displayed high stability [302]. Tobacco mosaic virus (TMV) particles impregnated with a metal-free paramagnetic nitroxide organic radical contrast agent (ORCA) were developed as probes for electron paramagnetic resonance and magnetic resonance imaging to detect superoxide. These probes demonstrated enhanced in vitro r1 and r2 relaxivities, acting as both T1 and T2 contrast agents, highlighting their potential for preclinical and clinical MRI scanning [303]. In a separate study, TMV VNPs were engineered to target VCAM-1, the vascular cell adhesion molecule, and loaded with Gd-dodecane tetraacetic acid (GdDOTA). This led to highly sensitive identification and visualization of atherosclerotic plaques in ApoE-/- mice using minimal contrast agent doses, resulting in improved relaxivity and moderate tumbling of the Gd-DOTA-TMV carrier with enhanced signal-to-noise ratio. Furthermore, these conjugates exhibited heightened imaging sensitivity, leading to a 40-fold reduction in Gd dosage compared to standard clinical doses [304].
25. PVNPs as Vaccine and Immunotherapy Agents
Recent studies suggest that intratumoral immunotherapy (IT-IT) of certain PVNPs can induce anti-tumor immune responses within the tumor microenvironment [305,306]. PVNPs' immune-stimulation against tumors are due to non-self [288] recognition. They can be identified by innate immune cells' pattern recognition receptors (PRRs), specifically toll-like receptors (TLRs) [288]. PVNPs, upon interaction with surface or endosomal TLRs on antigen-presenting cells (APCs), trigger the secretion of cytokines, chemokines, and interferons, recruiting and activating anti-tumor immune cells [288]. Studies have shown that PVNP's immunostimulatory effects can be mediated by nucleic acid content and multimeric coat protein assemblies [288]. One example is CPMV, wherein TLR2 and TLR4 identify capsid or empty CPMV; TLR-7/8 are responsible for recognizing positive-strand RNA genomes (Figure 4A) [288,307,308,309].
Recent studies show that CPMV with two different RNA genomes packaged separately into identical protein capsids can activate innate immune cells, inducing pro-inflammatory cytokines and suppressing immunosuppressive cytokines, with comparable efficacy [307]. The use of intratumoral immunotherapy (IT-IT) of eCPMV in canine inflammatory mammary cancer (IMC) demonstrate notable rise in neutrophil populations, T and B lymphocytes within the tumor microenvironment, and induced the anti-tumor response [311{Valdivia, 2023 #60]. Based on several findings, it is evident that IT-IT of PVNPs such as PVX [312], papaya mosaic virus (PapMV) [313], TMV [314] and CCMV, Sesbania mosaic virus (SeMV) [315] and alfalfa mosaic virus (AMV) [316] are increasingly recognized as highly effective in situ vaccination agents for various types of cancers. Recently, Zhao et. al, have demonstrated that conjugation of targeted peptides of mannose receptor, namely CSPGAK (CD206s, 561.7 Da) and CSPGAKVRC (CD206, 920.1 Da) (CD206 and CD206s) to CPMV functions as a promising avenue for cancer immunotherapy directed at M2 macrophages [317].
While these PVNPs exhibit variability in their capacity to act as immune stimulants, they differ in additional characteristics, such as their potential to be modified for delivering cancer antigens [306]. Utilizing PVNP-based cancer vaccines enables the initiation of immune responses specific to tumor-associated antigens. Recent research has detailed the utilization of various PVNPs, including icosahedral CPMV, CCMV, SeMV, PhMV, and filamentous PVX in HER2-specific cancer vaccines (Figure 4B) [310,312,318]. CPMV has also been employed for the delivery of the immunogenic cancer-linked testis antigen NY-ESO-1 [319].
PVNPs can undergo modifications to load immunoadjuvants and improve antitumor efficacy. An illustration of this concept is the utilization of CCMV loaded with CpG oligonucleotides to stimulate the activation of tumor-associated macrophages (TAMs) both in vitro and in vivo (Figure 4C) [275,320,321]. The attachment of TLR3 or TLR7 agonists to various PVNPs such as CPMV and CCMV VNPs has shown to boost immune cell activation and the generation of the pro-inflammatory cytokine interleukin 6. Specifically, the linking of the TLR7 agonist 2-methoxyethoxy-8-oxo-9-(4-carboxybenzyl) adenine (1V209) to CPMV and CCMV resulted in diminished tumor growth and enhanced survival rates in mice [322]. Likewise, the administration of a nucleic acid based TLR3 agonist, polyinosinic acid with polycytidylic acid (poly(I:C)), through CPMV led to an increase in survival in mice. These results emphasize the significance of combining and co-delivering TLR agonists to enhance their antitumor efficacy, with the multivalent presentation, prolonged presence in tumors, and precise targeting of innate immune cells by the PVNP carriers being pivotal factors in improving effectiveness. The results indicate that active microneedle, Pluronic F127, implantable polymeric hydrogels formulations effectively maintain the structure and function of CPMV for enabling slow-release immunotherapy for cancer [322,323,324].
26. PVNPs -Based Combination Therapies
Combining intratumoral PVNPs with other tumor therapies is being explored to improve treatment outcomes as demonstrated by the potent efficacy of chemotherapy, immune checkpoint therapy (ICT), radiation therapy (RT), along with PVNPs in mouse tumor models. Results suggest that utilizing CPMV particles in combination with RT can turn an immunologically “cold” tumor (with low number of tumor infiltrating lymphocytes (TILs)) into an immunologically “hot” tumor (with increase in TILs) [325]. ICT in cancer treatment has shown promising results but is limited to a minority of patients. Heightened efficacy of combined CPMV IIT and anti- Lymphocyte-activation gene-3 (LAG-3) treatment in a mouse model of melanoma has been observed wherein LAG-3 functions as a next-generation inhibitory immune checkpoint with broad expression across multiple immune cell subsets. Its expression increases on activated T cells and contributes to T cell exhaustion. LAG-3 is a novel inhibitory immune checkpoint that increases activated T cells and promotes T cell exhaustion [326] Combination of CPMV and anti-4-1BB monoclonal antibody agonist is an effective dual therapy approach. Using murine models of metastatic colon carcinomatosis and intradermal melanoma, intratumorally administered CPMV + anti-4-1BB (CD137) dual provided a robust antitumor response, improved elimination of primary tumors, and reduced mortality compared to CPMV and anti-4-1BB monotherapies [327]. CPMV combination with anti-PD-1 peptides (SNTSESF) resulted in increased efficacy; however, increased potency against metastatic ovarian cancer was only observed when SNTSESF was conjugated to CPMV, and not added as a free peptide. This can be explained by the differences in the in vivo fates of the nanoparticle formulation vs. the free peptide; the larger nanoparticles are expected to exhibit prolonged tumor residence and favorable intratumoral distribution [328].
Furthermore, the coadministration of DOX via PVX+DOX enhanced the response of the PVX monotherapy through increased survival, which was also represented in the enhanced antitumor cytokine/chemokine profile stimulated by PVX+DOX when compared to PVX or DOX alone [329]. Combination therapy using CPMV and low doses of cyclophosphamide (CPA) has shown remarkable synergistic efficacy against 4T1 mouse tumors in vivo. The combination of CPMV and CPA increases the secretion of several cytokines, activates antigen-presenting cells, increases the abundance of tumor infiltrating T cells, and systematically reverses the immunosuppression. These results show that the combination of CPMV in situ vaccination with chemotherapy may become a potent new strategy for the treatment of tumors [330].
Photothermal therapy (PTT) is a promising treatment for cancer that targets tumors locally and enhances immune responses. Nevertheless, the efficacy of PTT when used alone is often limited systemically, prompting the exploration of combined treatment strategies. In pursuit of this objective, the TMV was utilized to deliver a small molecule immunomodulator, toll-like receptor 7 agonist (1V209), while its surface was modified with the photothermal biopolymer polydopamine (PDA). The resulting complex of 1V209-loaded TMV coated with PDA was employed in the treatment of B16F10 dermal melanoma in C57BL/6 mice. This formulation, known as 1V209-TMV-PDA, was administered by intratumoral injection followed by irradiation with an 808-nm near infrared laser. Notably, 60% of the mice treated with intratumoral 1V209-TMV-PDA and laser irradiation survived until the study endpoint, demonstrating a significant improvement compared to the 20% survival rate observed in the control group (Figure 4D) [285].The study aims to develop a vaccine for ovarian cancer using irradiated cancer cells (ICCs) as an antigen and cowpea mosaic virus (CPMV) adjuvants. Results show co-formulated CPMV–ICCs successfully withstood initial tumor challenges in mice, emphasizing the importance of simultaneous delivery [331].
27. Recent Developments of PVNPs Against Cancer
Nanoparticle delivery systems have the potential to improve pharmacokinetics, tissue targeting, and stability of encapsulated therapeutic and imaging agents [332]. Numerous scientific advancements have been achieved using bioinspired nanocarriers such as proteins, nucleic acids, and viruses. One of the natural nanocarriers are self-assembling viruses to effectively deliver cargos [235,333]. PVNPs are new candidates that can be used as promising nanoplatforms [332,334]. Currently, PhMV-based nanoparticles have shown potential as a nanocarrier platform for loading cargo into living organisms. However, their internal loading capacity is limited due to low reactivity. A structure-based approach has created mutants with enhanced reactivity towards thiol-reactive small molecules (e.g. doxorubicin and vcMMAE) and imaging agents (DOTA(Gd)), resulting in ten-fold increased reactivity towards chemotherapeutic and MRI imaging agents [332]. A nanomedicine platform using CCMV-based nanoparticles modified with elastin-like peptides (ELPs) has been developed to address chemotherapy difficulties. The nanoparticles deliver DOX into tumor cells, stimulate immune responses, and hinder tumor growth in cancer models. [335].
Protein-based vaccines from mammalian, bacteriophages, and plant viruses offer advantages in cancer immunotherapy due to their ability to modulate the immune system [336]. PVNPs, although non-infectious to mammals, are recognized by the immune system as strong adjuvants. Systemic administration of CPMV activates the innate immune system, aiding in the identification and destruction of cancer cells, leading to a lasting and adaptable immune response against metastatic cancers like colon, ovarian, melanoma, and breast cancer. [337]. A study on 11 companion dogs with canine mammary cancer found that neoadjuvant intratumoral immunotherapy with empty eCPMV led to tumor reduction in both treated and untreated tumors. The reduction was observed across different stages, sizes, grades, and molecular subtypes. The injected tumors showed decreased DNA replication and increased dendritic cell activity, with increased levels of neutrophils, T lymphocytes, and plasma cells. The eCPMV immunotherapy was effective and had no negative side effects [338]. A cancer vaccine targeting S100A9, a major inflammation regulator, has been developed using plant virus and bacteriophage nanotechnologies. The vaccine significantly reduced S100A9 levels in tumor-bearing mice, protecting against lung metastasis. The vaccine also increased immunostimulatory cytokines and decreased immunosuppressive cytokines, potentially having wide-ranging implications in preventing metastasis due to its prevalence in multiple cancer types [339]. The study found that CPMV, a cytokine, effectively treated a metastatic ovarian tumor model without causing organ toxicity. The viral RNA persisted and could be detected two weeks after final administration. The study also showed that systemic administration of CPMV is safe and widely available. Recently, polymeric hydrogels with CPMV were surgically implanted into the peritoneal cavity for cancer immunotherapy. PVNPs derived from TuMV have been found to attract immunoglobulins G (IgG) due to gene fusion. A fluorescent nanoplatform was created to target cancer cells overexpressing EGFR, presenting a promising avenue for cancer theranostics. [340].Small TLR agonists of poly(I:C) are being developed as intratumoral immunotherapies, with rapid washout and poor immune cell uptake. To address these issues poly(I:C) is enclosed in nanoparticles made from CCMV. These particles enhance the function of macrophages and demonstrate effectiveness in reducing tumor growth and extending survival in mouse models of colon cancer and melanoma. When combined with oxaliplatin, CCMV-poly(I:C) shows even greater efficacy, significantly inhibiting tumor growth and improving survival rates [341]. PVNPs, when combined with immunotherapy, offer promising cancer treatment. In the future, PVNPs-based cancer vaccines could be integrated with traditional methods like radiotherapy and chemotherapy. In murine cancer models, combining CPMV's immune cell activation with oxaliplatin's immunogenic cell death led to increased median survival rates. This combination therapy effectively altered the tumor microenvironment, leading to enhanced tumor cell death. The research underscores the potential for combining chemotherapy with PVNP intratumoral immunotherapy in clinical settings, highlighting the potential of PVNP-based cancer vaccines in the future [342]. Cryoablation and intratumoral CPMV, whether used separately or together, showed strong effectiveness against treated HCC tumors. However, only the combination of cryoablation and CPMV was able to inhibit the growth of untreated tumors, indicating an abscopal effect [343].Generally, Advancements in nanobiotechnology are expected to improve PVNPs, leading to advancements in tumor diagnosis, treatment, and prevention. Recent tumor screening and antigen discovery have contributed to personalized immunotherapy. By using optimized nanoadjuvants based on PVNPs, along with immune-related molecules like cytokines or immunopotentiators, nanocancer vaccines with targeted, safe, and controlled release can be developed.
28. Advantages of the Use of Plants for Production of Anti-Cancer Mabs, Plant Viral Nanoparticles and Phytochemicals Against Cancer
The major advantages of the use of phytochemicals are their capabilities to induce multiple signaling networks, that enable cells to have greater decoding and signal processing properties. They preclude the advancement of cancer by decreasing survival and growth signals of tumor cells. Further, diverse phytochemicals can control crosslinked biosynthetic pathways such as the NF-κB axis, glycolytic enzymes, DLC1 pathway, MAPKs and ROS driving multiple processes against the tumor. Phytochemicals by virtue of their effectiveness on complex signaling cascades, inhibit cancer cell evasion caused by single pathways [344].
Plant-produced antibodies could be full-sized, single-chain antibody fragments, membrane-anchored scFv, bispecific scFv fragments, Fab fragments and chimeric antibodies. Unlike mammalian cells, recombinant plants can efficiently express secretory IgA. Plants are available on a widespread basis, grow rapidly and commonly mature following one growth season. Also, it is facile to bring the plant-based Mabs to the market within brief timeframes and at large scales which reduces the production cost. Further, plants decrease costs for screening for viruses, prions and bacterial toxins as they do not introduce human pathogens in contrast to production platforms such as transgenic animals or mammalian cells [345]. Furthermore, plants possess an endomembrane system as well as secretory pathway comparable with human cells, that are distinct from prokaryotic systems and bacteria. Whereas animal-based Abs elicit immune responses involving foreign or non-self-agents, plant-based Abs discount with such reactions. Similar to animal cells, they possess posttranslational modification mechanisms that enable them to be considered as biofactories for production of therapeutic Abs [346]. Plants that are glycoengineered have substantially greater degree of glycan homogeneity. Consequently, plants provide a powerful expression system for anticancer Mabs production.
Plant viruses are appropriate for vaccine production due to their capability to be recognized as foreign molecules by the innate immune system via pathogen-associated molecular pattern (PAMP) receptors [347].They are not pathogenic to humans and elicit humoral antibody response as well as cell-mediated immune response [348,349] upon delivery via parenteral [350] and mucosal [351] routes. The genomes of plant viruses have been genetically engineered to enable the expression of foreign open reading frames.
VNPs can be used as novel nanomaterials having several favorable characteristics [352]. They self-assemble, are monodisperse, polyvalent, highly potent, dynamic and can be produced within brief time periods. They are superior when compared with synthetic nanomaterials as they are biodegradable and biocompatible. Ligands including proteins, peptides, small chemical modifiers and even other nanoparticles can be encapsulated within the VNPs by self-assembly using a diverse range of bioconjugation chemistries such as genetic engineering, chemical bioconjugation, mineralization and encapsulation [353,354]. VLPs serve as robust vaccine candidates as they mimic native virus conformations and induce intrinsic immunogenicity without dispensing with their inherent safety, which they accomplish by virtue of being bereft of the viral genome and hence being incapable of replication [299]. Consequently, VLPs are increasingly favored as subunit vaccines as they easily undergo internalization by the antigen presenting cells leading to efficacious immune reactions. They are inimitable platforms for processing of antigens and presentation of epitopes to the immune system. Further, VLPs are popular for cancer immunotherapy due to their natural ability to elicit immune responses which prime the microenvironment of tumors and launch antitumor immunity. VLPs present themselves as multivalent, repetitive molecular scaffolds as they are composed of viral capsid proteins in multitudinous copies that enable multivalent antigen presentation [355]. Vaccines based on VLPs provide superior immunogenicity in comparison to antigens occurring in their soluble forms. Also, they are characterized by their native adjuvant properties which dispense with the employment of further adjuvants to elicit potent immune responses.
Through genetic engineering, epitopes can be expressed on the plant virus capsids resulting in a homogenous formulation while mitigating the heterogeneity of chemical conjugation methodologies [356]. Such vaccines could be successfully developed and purified from plant hosts via molecular farming, hence decreasing downstream processing and production costs [357,358]. Additionally, these plant virus-derived vaccines can be incorporated within polymeric implants or devices that can promote its shelf life while enabling extended antigen release [359].
Plant virus diversity affords an array of biological materials having different structures such as icosahedrons, filamentous structures and nanotubes [360]. Their structural characteristics make them ideal templates for synthetic and chemical engineering including infusion, encapsulation, bioconjugation, shape-tuning or genetic loading with cargos that range from tiny molecules to polypeptides and synthetic nanoparticles [355,361,362,363]. VLPs and VNPs by virtue of being monodisperse, provide high structural uniformity, quality assurance and quality control [364].
29. Disadvantages of Plant-Based Platforms Against Cancer
Despite their potent anticancer properties, some of the phytochemicals have been shown to be toxic to humans. Some others have been shown to possess antinutrient characteristics that hinder nutrient absorption. Certain others like flavonoids and polyphenols may function as pro-oxidants when ingested in high amounts. Also, autophagy plays a role based on context in cancer patients, due to which targeting this critical cellular pathway would not always be favorable. Additionally, many phytochemicals are associated with targeting of multiple signaling pathways shared between multiple cellular systems, which poses a challenge to the use of anticancer drugs based on phytochemicals.
Further, natural phytochemicals are ridden with drawbacks such as low potency, lowered solubility and stability as well as poor pharmacokinetics. Therefore, further studies are required to promote the phytochemicals’ bioavailability with novel formulations or by developing analogues with more potency including nano-based drug delivery schemes [365], chitosan-pectin-core-shell nanoparticles loaded with phytochemicals [366], glycosylation agents [367] and encapsulated phytochemicals [368], to augment the polarity and reduce the poor pharmacokinetic profiles of natural phytochemicals, thereby enhancing their efficacy against cancer and driving their conversion from laboratory level to bedside. More studies are called for towards design and fabrication of novel nanocomplexes co-encapsulating two or more phytochemicals, that facilitate anti-cancer potency in a synergistic manner. In vivo and in vitro studies on modulation of autophagy and apoptosis mediated by phytochemicals is required. Further, an integrated systemic computational and pharmacological approach could be used to better comprehend the anticancer properties of phytochemicals.
Although production of plant-based monoclonal antibodies is advantageous in the treatment of cancer as stated above, there are significant caveats in their successful production. Protein G or protein A-based affinity chromatography has been used to purify antibodies that have been expressed in plant systems [369]. However, to achieve purification, the plant tissues have to be homogenized first in order to break open the cell walls to release cell debris, contaminants and noxious substances following which these have to be removed by means of various purification steps [220]. However, there are challenges to protein purification because of problems of clogging in the chromatography columns caused by the cell wall debris that is left over in the process of homogenization of the plant biomass and removal of impurities [369]. Additionally, the application of protein A columns is circumscribed by its prohibitively high costs.
As regards plant viral VLPs and VNPs, although they are highly efficacious as anti-cancer vehicles, there exist several drawbacks that need to be addressed. In some instances, VNP / VLP capsid formation would likely be impaired due to the fusion of antigenic peptides with the respective viral capsid proteins. The necessity of efficacious infection and generation of VNPs in plants circumscribes the expression of epitopes as the latter could result in the disruption of viral assembly and infection [10]. Additionally, procurement of reagents acceptable to the regulatory bodies poses several challenges. For manufacturing purposes, further research must evaluate optimization of codons between the chosen peptides and codon usage in plants to preclude the accumulation of unstable proteins [370,371]. Moreover, promoter usage, selection of untranslated regions and thylakoid localization have to be factored in [372]. VNPs possess fundamental characteristics present in most classes of nanoparticles that can impact their viability in vivo [373,374]. Biological in vivo barriers including interactions with antibodies, immune cells and serum could negatively impact the use of native or functionalized VNPs under clinical settings [375]. Also, in the human circulatory system, the surface of VNPs could be covered with proteins from the serum resulting in protein corona that augment their ingestion by phagocytic cells [376,377]. The formation of protein corona can be an impediment in the development of VNPs for in vivo uses. Antibodies against the VNPs could alter interactions of VNPs with the immune cells resulting in their elimination before they reach their target sites [378]. However, challenges to VNP delivery can usually be avoided or decreased by in situ vaccination. If the use of VNPs involves multiple VNP applications over many weeks, then it is likely that anti-VNP antibodies could be generated in the immune system that could restrict their half-life in the circulatory system and augment their elimination from the body [379,380].
30. Regulatory Aspects of Plant-Made Biopharmaceuticals
In general, it is well-established that drugs such as anticancer compounds are required to go through phase III clinical trials to obtain market permissions. The guidelines provided by the European Medicines Agency (EMA) and the Food and Drug Administration (FDA) specify that at least a single controlled Phase III trial having statistically notable results is necessary for allowing the license to market the respective plant-based biopharmaceutical product [381,382]. Excepting extraordinary circumstances, all these drugs must go through all clinical trial phases as per the EMA and FDA regulatory guidelines. Nevertheless, it has been noted that several pharmaceutical organizations deviate from established protocols and commence testing of the new drugs on human subjects prior to the defined timeline. The reason for these practices is to expedite the approval of the respective compounds due to pressure from investors [Apolone et al., 2005]. Therefore, such anticancer drugs are presented for approval despite having insufficient information on their safety, efficacy and quality. Although plant-derived biopharmaceuticals have been shown to have lesser toxicity when compared to traditional synthetic compounds, evidence has emerged regarding the undesirable side effects of unbridled and unregulated use of these compounds against cancer and other diseases. For example, Fagonia indica has shown robust anticancer activity to breast cancer upon testing in the MDA-MB-231 cell line [383]. Besides, F. indica has been used in traditional medicine to treat several disorders and some people have even used it as herbal tea to treat breast cancer. Nevertheless, there have been only a few reports on its anticancer activity.
Worldwide, the process of oncological drug development and sale is regulated by the recruitment of experts as well as advisory procedures mediated by the regulatory authorities [384]. There are many models of regulatory frameworks obtainable for prescribing these drugs, but harmony is required amongst regulatory bodies and enhancement in the regulatory process. The FDA has adopted guidelines for the regulation of the International Council for Harmonization regarding nonclinical assessment of drugs intended for cancer treatment. Regulatory authorities must be harmonious with other agencies to regulate plant-based anticancer compounds and should enhance the focus on integrating information obtained from traditional knowledge with scientific investigations regarding these drugs [385].
Further, plants belonging to the same species when cultivated in different areas show variation in their content of medicinal compounds [386]. This calls for the necessity to focus on the generation of plants having enhanced qualities with uniform profiles of metabolites that once investigated are deemed safe or not conclusively. This can be achieved by the in vitro growth and continuing genetic and biotechnological studies on such anticancer plants [387,388].
31. Conclusions and Future Perspectives
Chemoprevention by means of dietary phytochemicals is a viable clinical approach in the management of carcinogenesis due to its simplicity and low-cost. The multimodal clinical usage of phytochemicals in the form of multifunctional compounds is highly propitious due to the capability of these compounds to reverse or stop premalignant cells’ neoplastic transformation on a genomic level while preserving healthy cells and precluding the appearance of tumor phenotypes [389,390]. Phytochemicals primarily as natural mixtures forming part of whole plant foods provide effective antioxidant activities and function as principal chemopreventive agents during the initiation phase of neoplastic transformation [391]. The oncostatic roles of phytochemicals in the scavenging of free radicals, augmented endogenous biosynthesis of antioxidant enzymes, enhanced DNA repair, metabolic inactivation of carcinogens, inhibition of pro-oxidant enzymes and detoxification have been well documented [392,393,394,395]. As phytochemicals are able to interfere with molecular mechanisms associated with the growth of the tumors and their metastatic spread, chemoprevention strategies must be designed to preclude the initiation of cancer while suppressing angiogenesis, cancer cell proliferation and formation of malignant stem cells in addition to promoting apoptosis, regulation of epigenetic mechanisms and immunity [396,397,398,399,400,401,402,403].
The mechanistic data on the benefits of phytochemicals at the preclinical level could be combined with clinical studies to provide clinical recommendations on the use of these substances in the chemoprevention of malignant cancer at the initial, secondary and tertiary levels [404]. The pivotal aim of chemoprevention is the suppression of cancer incidence and progression. Evidently, the proclivity for plant-derived functional foods compared to single phytochemicals could provide the logical and efficacious approach to manage malignant diseases [391,405]. Nevertheless, there is an obvious paucity in terms of results substantiating these discoveries in clinical settings.
Navigating the challenges and opportunities presented by integrating phototherapy with plant-derived bioactive compounds opens the door to exciting future directions in cancer research and treatment. Further, elucidating the molecular mechanisms, optimizing treatment protocols, and conducting large scale and rigorous clinical trials are imperative steps in realizing the full potential of this advanced approach. The dynamic synergy between light-based therapies and the rich pharmacopeia of plant-derived compounds holds promise for revolutionizing cancer treatment. As we venture into uncharted territories, integrating plant-derived compounds into phototherapy offers enhanced efficacy and the potential for minimizing adverse effects, paving the way for a new era in personalized and effective cancer therapeutics.
Plant generated immunotherapy based on Mabs enables the immune system to recognize and target tumor cells and it is projected to provide a viable solution for cancer treatment. Design of Mab therapeutics has advanced and developed towards displaying enhanced capabilities and decreased systemic toxicities with respect to healthy cells. Using new molecular tools, scientists introduce the preferred genes into target plants to generate transgenic plants that are regarded as favorable systems for expression of human therapeutic antibodies having ideal traits. Careful evaluation of selected plant species is obligately required as each plant species is characterized by its own unique physiochemical and physiological properties influencing the biosynthesis and glycosylation of the respective antibody glycoproteins.
Plants function as propitious biofactories for antibody production on a large scale owing to their low cost, increased production capacity, improved scalability and facile growth procedures subject to containment conditions while dispensing with the need to screen for pathogens. Several plant-based Mabs have been expressed on large scales in accordance to cGMP regulations and have proven to meet up to the US FDA standards of purity, potency and identity in addition to demonstrating appropriate assembly, efficacious neutralization in vitro and potent efficacy in vivo in animal model systems [226]. The FDA has provided approval for over a dozen Mabs meant for therapeutics of several malignancies and many companies have been involved in the production of these molecules. Recent advancements in glycoengineering, posttranslational modifications and genetic engineering have afforded further advantages beyond the conventional benefits of economic feasibility, augmented safety and improved scalability and it is estimated that plants can function as excellent systems for Mab production in the future.
Nanoparticles are increasingly used in the treatment of cancer with specific focus on therapeutic agent delivery. Nevertheless, administration of nanoparticles systemically has revealed that only about 1% of these particles can accumulate within the tumor microenvironment [288]. Therefore, to make a significant impact, novel delivery systems are warranted. VNPs serve as building blocks having capability for therapeutic agent delivery or for in situ vaccinations as well as in their combined forms. Multifunctional VNPs capable of loading, protecting and regulating targeted cargo release in addition to being intrinsically immunomodulatory, can enable immunogenic death of tumor cells, thereby modifying the tumor microenvironment. VNPs can be used for in situ vaccinations along with other multiple therapeutic strategies within a single platform. Such combination therapies could pave the way for efficacious inhibition of tumors.
VNPs and VLPs are subject to rapid immunogenic clearance which decreases their therapeutic efficacies following repeated administrations [406]. Many surface passivation techniques have been employed to protect the surface of the VNPs and VLPs from non-specific serum protein adsorption or to preclude antibody recognition using coating of viral surfaces with polyethylene glycol or other polymers [407] or albumin camouflaging [408]. Multiple treatments of viral nanocarriers can be precluded using implants, patches and scaffolds that enable slow release of these particles. Biodegradable polymers enable release of the VNPs and VLPs over time while not impacting their structural integrity and elicited anti-VLP immune reactions equivalent to that accomplished by the repeated administrations of the soluble particles [359]. The advancement of VLP-derived oral vaccines is another promising area warranting future exploration.
Combination of VNPs and VLPs with radiotherapy and chemotherapy can potentiate anticancer responses by augmenting immunogenic cell death and increasing the pool of tumor-targeting T cells to promote their efficacy. With growing interest in this field, the count of immunotherapies based on plant viruses in pre-clinical trials is poised to expand and lead to clinical studies and regulatory approval of these plant virus-derived therapies to enhance cancer immunotherapy [288]. A major number of scientific investigations have been conducted on the efficacy of plant viral nanoparticles in animal models or in vitro cell culture systems. Therefore, the next principal challenge would be the design and execution of clinical trials in humans. Additionally, mechanisms of delivery of VNPs into patients have to be investigated in further detail and finally, regulatory policies have to be formulated to ensure the safe transition of VNP applicability from the bench level into the clinical phase [409].
Innovations in the manufacturing framework, financial backing and maturation of regulatory structure regarding plant-based pharmaceuticals are vital to their commercialization in developing nations. As a majority of plant-derived biotechnology research and development has been sourced from academia, transition from this phase to the corporate scenario will take tremendous effort at the very least [410]. Nevertheless, favorable features including minimal production costs, lesser purification steps along with increased levels of efficacy and safety make plants an ideal platform for generation of monoclonal antibodies, viral nanoparticles, phytochemicals and other related biopharmaceuticals. Ultimately, advanced medical strategies founded on personalized, pre-emptive and predictive medicine is deemed the future of cancer management.
Author Contributions
SV conceptualized, designed, wrote and revised the manuscript; GH, MS, AY and KH wrote and revised the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li S, Chen J, Wang Y, Zhou X, Zhu W: Moxibustion for the side effects of surgical therapy and chemotherapy in patients with gastric cancer: A protocol for systematic review and meta-analysis. Medicine 2020, 99, e21087. [CrossRef] [PubMed]
- Kaufman HL, Atkins MB, Subedi P, Wu J, Chambers J, Joseph Mattingly T, Campbell JD, Allen J, Ferris AE, Schilsky RL: The promise of Immuno-oncology: implications for defining the value of cancer treatment. Journal for immunotherapy of cancer, 2019; 7, 1–11.
- Tiffon C: The impact of nutrition and environmental epigenetics on human health and disease. International journal of molecular sciences 2018, 19, 3425. [CrossRef] [PubMed]
- Guo Y, Su Z-Y, Kong A-NT: Current perspectives on epigenetic modifications by dietary chemopreventive and herbal phytochemicals. Current pharmacology reports 2015, 1:245-257.
- Samec M, Liskova A, Kubatka P, Uramova S, Zubor P, Samuel SM, Zulli A, Pec M, Bielik T, Biringer K: The role of dietary phytochemicals in the carcinogenesis via the modulation of miRNA expression. Journal of cancer research and clinical oncology, 2019; 145, 1665–1679.
- Lin SR, Chang CH, Hsu CF, Tsai MJ, Cheng H, Leong MK, Sung PJ, Chen JC, Weng CF: Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. British journal of pharmacology 2020, 177, 1409–1423. [CrossRef]
- Mokhtari RB, Homayouni TS, Baluch N, Morgatskaya E, Kumar S, Das B, Yeger H: Combination therapy in combating cancer. Oncotarget 2017, 8, 38022. [CrossRef]
- Kamta J, Chaar M, Ande A, Altomare DA, Ait-Oudhia S: Advancing cancer therapy with present and emerging immuno-oncology approaches. Frontiers in oncology, 2017; 7, 64.
- Besufekad Y, Malaiyarsa P: Production of monoclonal antibodies in transgenic plants. J Adv Biol Biotechnol, 2017; 12, 1–8.
- Hefferon K: Reconceptualizing cancer immunotherapy based on plant production systems. Future science OA 2017, 3, FSO217. [CrossRef]
- Keshavarz M, Ebrahimzadeh MS, Miri SM, Dianat-Moghadam H, Ghorbanhosseini SS, Mohebbi SR, Keyvani H, Ghaemi A: Oncolytic Newcastle disease virus delivered by Mesenchymal stem cells-engineered system enhances the therapeutic effects altering tumor microenvironment. Virology journal, 2020; 17, 1–13.
- Shukla S, Hu H, Cai H, Chan S-K, Boone CE, Beiss V, Chariou PL, Steinmetz NF: Plant viruses and bacteriophage-based reagents for diagnosis and therapy. Annual review of virology 2020, 7, 559–587. [CrossRef] [PubMed]
- Gyanani V, Haley JC, Goswami R: Challenges of current anticancer treatment approaches with focus on liposomal drug delivery systems. Pharmaceuticals 2021, 14, 835. [CrossRef]
- Nonnekens J, Hoeijmakers JH: After surviving cancer, what about late life effects of the cure? EMBO molecular medicine 2017, 9, 4–6. [CrossRef]
- Carvalho FS, Burgeiro A, Garcia R, Moreno AJ, Carvalho RA, Oliveira PJ: Doxorubicin-induced cardiotoxicity: from bioenergetic failure and cell death to cardiomyopathy. Medicinal research reviews 2014, 34, 106–135. [CrossRef]
- Wigmore PM, Mustafa S, El-Beltagy M, Lyons L, Umka J, Bennett G: Effects of 5-FU. In: Chemo Fog: Cancer Chemotherapy-Related Cognitive Impairment. Springer; 2010: 157-164.
- Ioele G, Chieffallo M, Occhiuzzi MA, De Luca M, Garofalo A, Ragno G, Grande F: Anticancer drugs: recent strategies to improve stability profile, pharmacokinetic and pharmacodynamic properties. Molecules 2022, 27, 5436. [CrossRef]
- Feyzizadeh M, Barfar A, Nouri Z, Sarfraz M, Zakeri-Milani P, Valizadeh H: Overcoming multidrug resistance through targeting ABC transporters: Lessons for drug discovery. Expert Opinion on Drug Discovery 2022, 17, 1013–1027. [CrossRef]
- Naeem A, Hu P, Yang M, Zhang J, Liu Y, Zhu W, Zheng Q: Natural products as anticancer agents: current status and future perspectives. Molecules 2022, 27, 8367. [CrossRef]
- Talib WH, Awajan D, Hamed RA, Azzam AO, Mahmod AI, Al-Yasari IH: Combination anticancer therapies using selected phytochemicals. Molecules 2022, 27, 5452. [CrossRef]
- Steward W, Brown K: Cancer chemoprevention: a rapidly evolving field. British journal of cancer 2013, 109, 1–7. [CrossRef]
- Swetha M, Keerthana C, Rayginia TP, Anto RJ: Cancer chemoprevention: A strategic approach using phytochemicals. Frontiers in Pharmacology, 2022; 12, 809308.
- Olayiwola Y, Gollahon L: Natural Compounds and Breast Cancer: Chemo-Preventive and Therapeutic Capabilities of Chlorogenic Acid and Cinnamaldehyde. Pharmaceuticals 2024, 17, 361. [CrossRef]
- Chen C, Kong A-NT: Dietary cancer-chemopreventive compounds: from signaling and gene expression to pharmacological effects. Trends in pharmacological sciences 2005, 26, 318–326. [CrossRef]
- Mocanu M-M, Nagy P, Szöllősi J: Chemoprevention of breast cancer by dietary polyphenols. Molecules 2015, 20, 22578–22620. [CrossRef]
- Hussain SS, Kumar AP, Ghosh R: Food-based natural products for cancer management: Is the whole greater than the sum of the parts? In: Seminars in cancer biology: 2016. Elsevier: 233-246.
- Orlikova B, Dicato M, Diederich M: 1,000 Ways to die: natural compounds modulate non-canonical cell death pathways in cancer cells. Phytochemistry reviews, 2014; 13, 277–293.
- Israel BeB, Tilghman SL, Parker-Lemieux K, Payton-Stewart F: Phytochemicals: Current strategies for treating breast cancer. Oncology letters 2018, 15, 7471–7478.
- Tao J, Diao L, Chen F, Shen A, Wang S, Jin H, Cai D, Hu Y: pH-sensitive nanoparticles codelivering docetaxel and dihydroartemisinin effectively treat breast cancer by enhancing reactive oxidative species-mediated mitochondrial apoptosis. Molecular Pharmaceutics 2020, 18, 74–86.
- Weng J-R, Bai L-Y, Chiu C-F, Hu J-L, Chiu S-J, Wu C-Y: Cucurbitane Triterpenoid from Momordica charantia Induces Apoptosis and Autophagy in Breast Cancer Cells, in Part, through Peroxisome Proliferator-Activated Receptor γ Activation. Evidence-Based Complementary and Alternative Medicine 2013, 2013, 935675.
- Xu W-S, Li T, Wu G-S, Dang Y-Y, Hao W-H, Chen X-P, Lu J-J, Wang Y-T: Effects of furanodiene on 95-D lung cancer cells: apoptosis, autophagy and G1 phase cell cycle arrest. The American Journal of Chinese Medicine 2014, 42, 243–255. [CrossRef]
- Zhang Q, Wang X, Cao S, Sun Y, He X, Jiang B, Yu Y, Duan J, Qiu F, Kang N: Berberine represses human gastric cancer cell growth in vitro and in vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling pathways. Biomedicine & Pharmacotherapy, 2020; 128, 110245.
- Davoodvandi A, Sadeghi S, Alavi SMA, Alavi SS, Jafari A, Khan H, Aschner M, Mirzaei H, Sharifi M, Asemi Z: The therapeutic effects of berberine for gastrointestinal cancers. Asia-Pacific Journal of Clinical Oncology 2024, 20, 152–167. [CrossRef]
- Eguchi H, Kimura R, Onuma S, Ito A, Yu Y, Yoshino Y, Matsunaga T, Endo S, Ikari A: Elevation of anticancer drug toxicity by caffeine in spheroid model of human lung adenocarcinoma A549 cells mediated by reduction in claudin-2 and Nrf2 expression. International Journal of Molecular Sciences 2022, 23, 15447. [CrossRef]
- Wang A, Wang W, Chen Y, Ma F, Wei X, Bi Y: Deguelin induces PUMA-mediated apoptosis and promotes sensitivity of lung cancer cells (LCCs) to doxorubicin (Dox). Molecular and Cellular Biochemistry 2018, 442, 177–186. [CrossRef]
- Swain SS, Sahoo SK: Piperlongumine and its derivatives against cancer: A recent update and future prospective. Archiv der Pharmazie, 2024; e2300768.
- Singh P, Sahoo SK: Piperlongumine loaded PLGA nanoparticles inhibit cancer stem-like cells through modulation of STAT3 in mammosphere model of triple negative breast cancer. International journal of pharmaceutics, 2022; 616, 121526.
- Singh D, Mohapatra P, Kumar S, Behera S, Dixit A, Sahoo SK: Nimbolide-encapsulated PLGA nanoparticles induces mesenchymal-to-epithelial transition by dual inhibition of AKT and mTOR in pancreatic cancer stem cells. Toxicology in Vitro, 2022; 79, 105293.
- Wang F, Mao Y, You Q, Hua D, Cai D: Piperlongumine induces apoptosis and autophagy in human lung cancer cells through inhibition of PI3K/Akt/mTOR pathway. International journal of immunopathology and pharmacology 2015, 28, 362–373. [CrossRef]
- Thongsom S, Suginta W, Lee KJ, Choe H, Talabnin C: Piperlongumine induces G2/M phase arrest and apoptosis in cholangiocarcinoma cells through the ROS-JNK-ERK signaling pathway. Apoptosis, 2017; 22, 1473–1484.
- Rawat L, Hegde H, Hoti SL, Nayak V: Piperlongumine induces ROS mediated cell death and synergizes paclitaxel in human intestinal cancer cells. Biomedicine & Pharmacotherapy, 2020; 128, 110243.
- Tripathi SK, Biswal BK: Piperlongumine, a potent anticancer phytotherapeutic: Perspectives on contemporary status and future possibilities as an anticancer agent. Pharmacological Research, 2020; 156, 104772.
- Kung F-P, Lim Y-P, Chao W-Y, Zhang Y-S, Yu H-I, Tai T-S, Lu C-H, Chen S-H, Li Y-Z, Zhao P-W: Piperlongumine, a potent anticancer phytotherapeutic, induces cell cycle arrest and apoptosis in vitro and in vivo through the ROS/Akt pathway in human thyroid cancer cells. Cancers 2021, 13, 4266.
- Pan X, Chen G, Hu W: Piperlongumine increases the sensitivity of bladder cancer to cisplatin by mitochondrial ROS. Journal of Clinical Laboratory Analysis 2022, 36, e24452. [CrossRef]
- Yang Y, Sun S, Xu W, Zhang Y, Yang R, Ma K, Zhang J, Xu J: Piperlongumine inhibits thioredoxin reductase 1 by targeting selenocysteine residues and sensitizes cancer cells to erastin. Antioxidants 2022, 11, 710. [CrossRef]
- Chu Y, Tian Z, Yang M, Li W: Conformation and energy investigation of microtubule longitudinal dynamic instability induced by natural products. Chemical Biology & Drug Design 2023, 102, 444–456.
- Risinger AL, Du L: Targeting and extending the eukaryotic druggable genome with natural products: Cytoskeletal targets of natural products. Natural product reports 2020, 37, 634–652. [CrossRef]
- Dhyani P, Quispe C, Sharma E, Bahukhandi A, Sati P, Attri DC, Szopa A, Sharifi-Rad J, Docea AO, Mardare I: Anticancer potential of alkaloids: a key emphasis to colchicine, vinblastine, vincristine, vindesine, vinorelbine and vincamine. Cancer cell international 2022, 22, 206. [CrossRef]
- Au TH, Nguyen BN, Nguyen PH, Pethe S, Vo-Thanh G, Vu Thi TH: Vinblastine loaded on graphene quantum dots and its anticancer applications. Journal of microencapsulation 2022, 39, 239–251. [CrossRef]
- Zhang Y, An J, Shao Y, Yu N, Yue S, Sun H, Zhang J, Gu W, Xia Y, Zhang J et al: CD38-Directed Vincristine Nanotherapy for Acute Lymphoblastic Leukemia. Biomacromolecules 2022, 23, 377–387. [CrossRef]
- Atal C, Dubey RK, Singh J: Biochemical basis of enhanced drug bioavailability by piperine: evidence that piperine is a potent inhibitor of drug metabolism. Journal of Pharmacology and Experimental Therapeutics 1985, 232, 258–262.
- Yaffe PB, Power Coombs MR, Doucette CD, Walsh M, Hoskin DW: Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1 arrest and apoptosis triggered by endoplasmic reticulum stress. Molecular carcinogenesis 2015, 54, 1070–1085. [CrossRef]
- Yang Q, Li F, Jia G, Liu R: Aged black garlic extract inhibits the growth of estrogen receptor-positive breast cancer cells by downregulating MCL-1 expression through the ROS-JNK pathway. Plos one 2023, 18, e0286454.
- Stępień AE, Trojniak J, Tabarkiewicz J: Anti-Cancer and Anti-Inflammatory Properties of Black Garlic. International Journal of Molecular Sciences 2024, 25, 1801. [CrossRef]
- Park C, Park S, Chung YH, Kim G-Y, Choi YW, Kim BW, Choi YH: Induction of apoptosis by a hexane extract of aged black garlic in the human leukemic U937 cells. Nutrition Research and Practice 2014, 8, 132. [CrossRef]
- Dong M, Yang G, Liu H, Liu X, Lin S, Sun D, Wang Y: Aged black garlic extract inhibits HT29 colon cancer cell growth via the PI3K/Akt signaling pathway. Biomedical Reports 2014, 2, 250–254. [CrossRef]
- Zhang Y, Li HY, Zhang ZH, Bian HL, Lin G: Garlic-derived compound S-allylmercaptocysteine inhibits cell growth and induces apoptosis via the JNK and p38 pathways in human colorectal carcinoma cells. Oncology letters 2014, 8, 2591–2596. [CrossRef] [PubMed]
- Shang A, Cao S-Y, Xu X-Y, Gan R-Y, Tang G-Y, Corke H, Mavumengwana V, Li H-B: Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246.
- Bagul M, Kakumanu S, Wilson TA: Crude garlic extract inhibits cell proliferation and induces cell cycle arrest and apoptosis of cancer cells in vitro. Journal of medicinal food 2015, 18, 731–737. [CrossRef] [PubMed]
- Toledano Medina MÁ, Merinas-Amo T, Fernández-Bedmar Z, Font R, del Río-Celestino M, Pérez-Aparicio J, Moreno-Ortega A, Alonso-Moraga Á, Moreno-Rojas R: Physicochemical characterization and biological activities of black and white garlic: In vivo and in vitro assays. Foods 2019, 8, 220.
- Wang X, Jiao F, Wang Q-W, Wang J, Yang K, Hu R-R, Liu H-C, Wang H-Y, Wang Y-S: Aged black garlic extract induces inhibition of gastric cancer cell growth in vitro and in vivo. Molecular Medicine Reports 2012, 5, 66–72.
- Castro NP, Rangel MC, Merchant AS, MacKinnon G, Cuttitta F, Salomon DS, Kim YS: Sulforaphane suppresses the growth of triple-negative breast cancer stem-like cells in vitro and in vivo. Cancer Prevention Research 2019, 12, 147–158. [CrossRef]
- Na G, He C, Zhang S, Tian S, Bao Y, Shan Y: Dietary isothiocyanates: Novel insights into the potential for cancer prevention and therapy. International Journal of Molecular Sciences 2023, 24, 1962. [CrossRef]
- Li Y, Zhang T, Korkaya H, Liu S, Lee H-F, Newman B, Yu Y, Clouthier SG, Schwartz SJ, Wicha MS: Sulforaphane, a dietary component of broccoli/broccoli sprouts, inhibits breast cancer stem cells. Clinical Cancer Research 2010, 16, 2580–2590. [CrossRef]
- Jeon YK, Yoo DR, Jang YH, Jang SY, Nam MJ: Sulforaphane induces apoptosis in human hepatic cancer cells through inhibition of 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase4, mediated by hypoxia inducible factor-1-dependent pathway. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 2011, 1814, 1340–1348. [CrossRef] [PubMed]
- Keenan JI, Salm N, Wallace AJ, Hampton MB: Using food to reduce H. pylori-associated inflammation. Phytotherapy Research 2012, 26, 1620–1625. [Google Scholar] [CrossRef]
- He C, Huang L, Lei P, Liu X, Li B, Shan Y: Sulforaphane normalizes intestinal flora and enhances gut barrier in mice with BBN-induced bladder cancer. Molecular nutrition & food research 2018, 62, 1800427.
- Singh KB, Hahm E-R, Alumkal JJ, Foley LM, Hitchens TK, Shiva SS, Parikh RA, Jacobs BL, Singh SV: Reversal of the Warburg phenomenon in chemoprevention of prostate cancer by sulforaphane. Carcinogenesis 2019, 40, 1545–1556. [CrossRef] [PubMed]
- Yan Y, Zhou Y, Li J, Zheng Z, Hu Y, Li L, Wu W: Sulforaphane downregulated fatty acid synthase and inhibited microtubule-mediated mitophagy leading to apoptosis. Cell Death & Disease 2021, 12, 917.
- Li S-H, Fu J, Watkins DN, Srivastava RK, Shankar S: Sulforaphane regulates self-renewal of pancreatic cancer stem cells through the modulation of Sonic hedgehog–GLI pathway. Molecular and cellular biochemistry 2013, 373:217-227.
- Kumar R, de Mooij T, Peterson T, Johnson A, Daniels DJ, Parney IF: Modulating glioma-mediated myeloid-derived suppressor cell development with sulforaphane. In: NEURO-ONCOLOGY: 2016. OXFORD UNIV PRESS INC JOURNALS DEPT, 2001 EVANS RD, CARY, NC 27513 USA: 99-100.
- Rai R, Gong Essel K, Mangiaracina Benbrook D, Garland J, Daniel Zhao Y, Chandra V: Preclinical efficacy and involvement of AKT, mTOR, and ERK kinases in the mechanism of sulforaphane against endometrial cancer. Cancers 2020, 12, 1273. [CrossRef] [PubMed]
- Bao Y, Wang W, Zhou Z, Sun C: Benefits and risks of the hormetic effects of dietary isothiocyanates on cancer prevention. PLoS One 2014, 9, e114764.
- Rao J, Xu D-R, Zheng F-M, Long Z-J, Huang S-S, Wu X, Zhou W-H, Huang R-W, Liu Q: Curcumin reduces expression of Bcl-2, leading to apoptosis in daunorubicin-insensitive CD34+ acute myeloid leukemia cell lines and primary sorted CD34+ acute myeloid leukemia cells. Journal of translational medicine, 2011; 9, 1–15.
- Cao A, Li Q, Yin P, Dong Y, Shi H, Wang L, Ji G, Xie J, Wu D: Curcumin induces apoptosis in human gastric carcinoma AGS cells and colon carcinoma HT-29 cells through mitochondrial dysfunction and endoplasmic reticulum stress. Apoptosis, 2013; 18, 1391–1402.
- Liu E, Wu J, Cao W, Zhang J, Liu W, Jiang X, Zhang X: Curcumin induces G2/M cell cycle arrest in a p53-dependent manner and upregulates ING4 expression in human glioma. Journal of neuro-oncology, 2007; 85, 263–270.
- Mousavi SM, Hosseindoost S, Mahdian SMA, Vousooghi N, Rajabi A, Jafari A, Ostadian A, Hamblin MR, Hadjighassem M, Mirzaei H: Exosomes released from U87 glioma cells treated with curcumin and/or temozolomide produce apoptosis in naive U87 cells. Pathology-Research and Practice, 2023; 245, 154427.
- Mukherjee S, Mazumdar M, Chakraborty S, Manna A, Saha S, Khan P, Bhattacharjee P, Guha D, Adhikary A, Mukhjerjee S: Curcumin inhibits breast cancer stem cell migration by amplifying the E-cadherin/β-catenin negative feedback loop. Stem cell research & therapy, 2014; 5, 1–19.
- Borges GA, Elias ST, Amorim B, de Lima CL, Coletta RD, Castilho RM, Squarize CH, Guerra ENS: Curcumin downregulates the PI3K–AKT–mTOR pathway and inhibits growth and progression in head and neck cancer cells. Phytotherapy research 2020, 34, 3311–3324. [CrossRef]
- Shamsnia HS, Roustaei M, Ahmadvand D, Butler AE, Amirlou D, Soltani S, Momtaz S, Jamialahmadi T, Abdolghaffari AH, Sahebkar A: Impact of curcumin on p38 MAPK: Therapeutic implications. Inflammopharmacology 2023, 31, 2201–2212. [CrossRef] [PubMed]
- Almatroodi SA, Almatroudi A, Khan AA, Alhumaydhi FA, Alsahli MA, Rahmani AH: Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 2020, 25, 3146. [CrossRef]
- Hayakawa S, Ohishi T, Miyoshi N, Oishi Y, Nakamura Y, Isemura M: Anti-cancer effects of green tea epigallocatchin-3-gallate and coffee chlorogenic acid. Molecules 2020, 25, 4553. [CrossRef]
- Gu J-W, Makey KL, Tucker KB, Chinchar E, Mao X, Pei I, Thomas EY, Miele L: EGCG, a major green tea catechin suppresses breast tumor angiogenesis and growth via inhibiting the activation of HIF-1α and NFκB, and VEGF expression. Vascular cell, 2013; 5, 1–10.
- Sen T, Chatterjee A: Epigallocatechin-3-gallate (EGCG) downregulates EGF-induced MMP-9 in breast cancer cells: involvement of integrin receptor α5β1 in the process. European journal of nutrition, 2011; 50, 465–478.
- Wei R, Cortez Penso NE, Hackman RM, Wang Y, Mackenzie GG: Epigallocatechin-3-gallate (EGCG) suppresses pancreatic cancer cell growth, invasion, and migration partly through the inhibition of Akt pathway and epithelial–mesenchymal transition: Enhanced efficacy when combined with gemcitabine. Nutrients 2019, 11, 1856.
- Van Aller GS, Carson JD, Tang W, Peng H, Zhao L, Copeland RA, Tummino PJ, Luo L: Epigallocatechin gallate (EGCG), a major component of green tea, is a dual phosphoinositide-3-kinase/mTOR inhibitor. Biochemical and biophysical research communications 2011, 406, 194–199. [CrossRef]
- Ko E-B, Jang Y-G, Kim C-W, Go R-E, Lee HK, Choi K-C: Gallic acid hindered lung cancer progression by inducing cell cycle arrest and apoptosis in a549 lung cancer cells via PI3K/Akt pathway. Biomolecules & Therapeutics 2022, 30, 151.
- Zhang Y, Ren X, Shi M, Jiang Z, Wang H, Su Q, Liu Q, Li G, Jiang G: Downregulation of STAT3 and activation of MAPK are involved in the induction of apoptosis by HNK in glioblastoma cell line U87. Oncology reports 2014, 32, 2038–2046. [CrossRef] [PubMed]
- Moral R, Escrich E: Influence of olive oil and its components on breast cancer: Molecular mechanisms. Molecules 2022, 27, 477. [CrossRef] [PubMed]
- Li Y, Pu R, Zhou L, Wang D, Li X: Effects of a Chlorogenic Acid-Containing Herbal Medicine (LASNB) on Colon Cancer. Evidence-Based Complementary and Alternative Medicine 2021, 2021, 9923467.
- Lu H, Tian Z, Cui Y, Liu Z, Ma X: Chlorogenic acid: A comprehensive review of the dietary sources, processing effects, bioavailability, beneficial properties, mechanisms of action, and future directions. Comprehensive reviews in food science and food safety 2020, 19, 3130–3158. [CrossRef]
- Kim TW: Cinnamaldehyde induces autophagy-mediated cell death through ER stress and epigenetic modification in gastric cancer cells. Acta Pharmacologica Sinica 2022, 43, 712–723. [CrossRef]
- Mei J, Ma J, Xu Y, Wang Y, Hu M, Ma F, Qin Z, Xue R, Tao N: Cinnamaldehyde treatment of prostate cancer-associated fibroblasts prevents their inhibitory effect on T cells through toll-like receptor 4. Drug design, development and therapy, 2020; 3363–3372.
- Kueck A, Opipari Jr AW, Griffith KA, Tan L, Choi M, Huang J, Wahl H, Liu JR: Resveratrol inhibits glucose metabolism in human ovarian cancer cells. Gynecologic oncology 2007, 107, 450–457. [CrossRef]
- Fu Y, Chang H, Peng X, Bai Q, Yi L, Zhou Y, Zhu J, Mi M: Resveratrol inhibits breast cancer stem-like cells and induces autophagy via suppressing Wnt/β-catenin signaling pathway. PloS one 2014, 9, e102535.
- Jang M, Cai L, Udeani GO, Slowing KV, Thomas CF, Beecher CW, Fong HH, Farnsworth NR, Kinghorn AD, Mehta RG: Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. science 1997, 275, 218–220. [CrossRef]
- Baek SH, Ko J-H, Lee H, Jung J, Kong M, Lee J-w, Lee J, Chinnathambi A, Zayed M, Alharbi SA: Resveratrol inhibits STAT3 signaling pathway through the induction of SOCS-1: Role in apoptosis induction and radiosensitization in head and neck tumor cells. Phytomedicine 2016, 23, 566–577. [CrossRef]
- Harikumar KB, Kunnumakkara AB, Sethi G, Diagaradjane P, Anand P, Pandey MK, Gelovani J, Krishnan S, Guha S, Aggarwal BB: Resveratrol, a multitargeted agent, can enhance antitumor activity of gemcitabine in vitro and in orthotopic mouse model of human pancreatic cancer. International journal of cancer 2010, 127, 257–268.
- Carter LG, D'Orazio JA, Pearson KJ: Resveratrol and cancer: focus on in vivo evidence. Endocrine-related cancer 2014, 21, R209–R225. [CrossRef]
- Yousef M, Vlachogiannis IA, Tsiani E: Effects of resveratrol against lung cancer: In vitro and in vivo studies. Nutrients 2017, 9, 1231. [CrossRef] [PubMed]
- Adhami VM, Afaq F, Ahmad N: Suppression of ultraviolet B exposure-mediated activation of NF-κB in normal human keratinocytes by resveratrol. Neoplasia 2003, 5, 74–82. [CrossRef] [PubMed]
- Espinoza JL, Kurokawa Y, Takami A: Rationale for assessing the therapeutic potential of resveratrol in hematological malignancies. Blood Reviews, 2019; 33, 43–52.
- Ren B, Kwah MX-Y, Liu C, Ma Z, Shanmugam MK, Ding L, Xiang X, Ho PC-L, Wang L, Ong PS: Resveratrol for cancer therapy: Challenges and future perspectives. Cancer letters, 2021; 515, 63–72.
- Seong Y-A, Shin P-G, Yoon J-S, Yadunandam AK, Kim G-D: Induction of the endoplasmic reticulum stress and autophagy in human lung carcinoma A549 cells by anacardic acid. Cell biochemistry and biophysics, 2014; 68, 369–377.
- Li Y, Zhou Y, Wang M, Lin X, Zhang Y, Laurent I, Zhong Y, Li J: Ampelopsin inhibits breast cancer cell growth through mitochondrial apoptosis pathway. Biological and Pharmaceutical Bulletin 2021, 44, 1738–1745. [CrossRef]
- Lu H-F, Chie Y-J, Yang M-S, Lee C-S, Fu J-J, Yang J-S, Tan T-W, Wu S-H, Ma Y-S, Ip S-W: Apigenin induces caspase-dependent apoptosis in human lung cancer A549 cells through Bax-and Bcl-2-triggered mitochondrial pathway. International journal of oncology 2010, 36, 1477–1484.
- Tsai M-H, Liu J-F, Chiang Y-C, Hu SC-S, Hsu L-F, Lin Y-C, Lin Z-C, Lee H-C, Chen M-C, Huang C-L: Correction: Artocarpin, an isoprenyl flavonoid, induces p53-dependent or independent apoptosis via ROS-mediated MAPKs and Akt activation in non-small cell lung cancer cells. Oncotarget 2019, 10, 3430.
- Park S-A, Seo YJ, Kim LK, Kim HJ, Yoon KD, Heo T-H: Butein Inhibits Cell Growth by Blocking the IL-6/IL-6Rα Interaction in Human Ovarian Cancer and by Regulation of the IL-6/STAT3/FoxO3a Pathway. International Journal of Molecular Sciences 2023, 24, 6038. [CrossRef]
- Khoo BY, Chua SL, Balaram P: Apoptotic effects of chrysin in human cancer cell lines. International journal of molecular sciences 2010, 11, 2188–2199. [CrossRef]
- Samarghandian S, Azimi Nezhad M, Mohammadi G: Role of caspases, Bax and Bcl-2 in chrysin-induced apoptosis in the A549 human lung adenocarcinoma epithelial cells. Anti-Cancer Agents in Medicinal Chemistry (Formerly Current Medicinal Chemistry-Anti-Cancer Agents) 2014, 14, 901–909.
- Zhang Z, Pan Y, Zhao Y, Ren M, Li Y, Lu G, Wu K, He S: Delphinidin modulates JAK/STAT3 and MAPKinase signaling to induce apoptosis in HCT116 cells. Environmental Toxicology 2021, 36, 1557–1566. [CrossRef] [PubMed]
- Gossner G, Choi M, Tan L, Fogoros S, Griffith KA, Kuenker M, Liu JR: Genistein-induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecologic oncology 2007, 105, 23–30. [CrossRef] [PubMed]
- Nicosia SV, Bai W, Cheng JQ, Coppola D, Kruk PA: Oncogenic pathways implicated in ovarian epithelial cancer. Hematology/Oncology Clinics 2003, 17, 927–943. [CrossRef] [PubMed]
- Joshi H, Gupta DS, Abjani NK, Kaur G, Mohan CD, Kaur J, Aggarwal D, Rani I, Ramniwas S, Abdulabbas HS: Genistein: a promising modulator of apoptosis and survival signaling in cancer. Naunyn-Schmiedeberg's Archives of Pharmacology 2023, 396, 2893–2910. [CrossRef]
- Obinu A, Burrai GP, Cavalli R, Galleri G, Migheli R, Antuofermo E, Rassu G, Gavini E, Giunchedi P: Transmucosal solid lipid nanoparticles to improve genistein absorption via intestinal lymphatic transport. Pharmaceutics 2021, 13, 267. [CrossRef]
- Křížová L, Dadáková K, Kašparovská J, Kašparovský T: Isoflavones. Molecules 2019, 24, 1076. [CrossRef]
- Kim I-S: Current perspectives on the beneficial effects of soybean isoflavones and their metabolites for humans. Antioxidants 2021, 10, 1064. [CrossRef] [PubMed]
- Liu R, Yu X, Chen X, Zhong H, Liang C, Xu X, Xu W, Cheng Y, Wang W, Yu L: Individual factors define the overall effects of dietary genistein exposure on breast cancer patients. Nutrition research, 2019; 67, 1–16.
- Ma M, Luan X, Zheng H, Wang X, Wang S, Shen T, Ren D: A mulberry diels-alder-type adduct, Kuwanon M, triggers apoptosis and paraptosis of lung cancer cells through inducing endoplasmic reticulum stress. International Journal of Molecular Sciences 2023, 24, 1015. [CrossRef]
- Shu Y-h, Yuan H-h, Xu M-t, Hong Y-t, Gao C-c, Wu Z-p, Han H-t, Sun X, Gao R-l, Yang S-f: A novel Diels–Alder adduct of mulberry leaves exerts anticancer effect through autophagy-mediated cell death. Acta Pharmacologica Sinica 2021, 42, 780–790. [CrossRef]
- Wang K, Liu R, Li J, Mao J, Lei Y, Wu J, Zeng J, Zhang T, Wu H, Chen L: Quercetin induces protective autophagy in gastric cancer cells: involvement of Akt-mTOR-and hypoxia-induced factor 1α-mediated signaling. Autophagy 2011, 7, 966–978. [CrossRef]
- Mohammed HA, Sulaiman GM, Anwar SS, Tawfeeq AT, Khan RA, Mohammed SA, Al-Omar MS, Alsharidah M, Al Rugaie O, Al-Amiery AA: Quercetin against MCF7 and CAL51 breast cancer cell lines: apoptosis, gene expression and cytotoxicity of nano-quercetin. Nanomedicine 2021.
- Nakamura M, Urakawa D, He Z, Akagi I, Hou D-X, Sakao K: Apoptosis Induction in HepG2 and HCT116 Cells by a Novel Quercetin-Zinc (II) Complex: Enhanced Absorption of Quercetin and Zinc (II). International Journal of Molecular Sciences 2023, 24, 17457. [CrossRef] [PubMed]
- Tarahovsky YS, Kim YA, Yagolnik EA, Muzafarov EN: Flavonoid–membrane interactions: Involvement of flavonoid–metal complexes in raft signaling. Biochimica et Biophysica Acta (BBA)-Biomembranes 2014, 1838, 1235–1246. [CrossRef] [PubMed]
- Wang R, Yang L, Li S, Ye D, Yang L, Liu Q, Zhao Z, Cai Q, Tan J, Li X: Quercetin inhibits breast cancer stem cells via downregulation of aldehyde dehydrogenase 1A1 (ALDH1A1), chemokine receptor type 4 (CXCR4), mucin 1 (MUC1), and epithelial cell adhesion molecule (EpCAM). Medical Science Monitor: international medical journal of experimental and clinical research, 2018; 24, 412.
- Liu Y, Wang Y, Sun S, Chen Z, Xiang S, Ding Z, Huang Z, Zhang B: Understanding the versatile roles and applications of EpCAM in cancers: from bench to bedside. Experimental hematology & oncology 2022, 11, 97.
- Binienda A, Ziolkowska S, Pluciennik E: The anticancer properties of silibinin: its molecular mechanism and therapeutic effect in breast cancer. Anti-Cancer Agents in Medicinal Chemistry (Formerly Current Medicinal Chemistry-Anti-Cancer Agents) 2020, 20, 1787–1796.
- Iqbal MA, Chattopadhyay S, Siddiqui FA, Ur Rehman A, Siddiqui S, Prakasam G, Khan A, Sultana S, Bamezai RN: Silibinin induces metabolic crisis in triple-negative breast cancer cells by modulating EGFR-MYC-TXNIP axis: potential therapeutic implications. The FEBS journal 2021, 288, 471–485. [CrossRef]
- Feng X-L, Ho SC, Mo X-F, Lin F-Y, Zhang N-Q, Luo H, Zhang X, Zhang C-X: Association between flavonoids, flavonoid subclasses intake and breast cancer risk: a case-control study in China. European Journal of Cancer Prevention 2020, 29, 493–500. [CrossRef]
- Golonko A, Olichwier AJ, Swislocka R, Szczerbinski L, Lewandowski W: Why Do Dietary Flavonoids Have a Promising Effect as Enhancers of Anthracyclines? Hydroxyl Substituents, Bioavailability and Biological Activity. International Journal of Molecular Sciences 2022, 24, 391. [CrossRef]
- Dhingra R, Margulets V, Kirshenbaum L: Chapter 2—Molecular Mechanisms Underlying Anthracycline Cardiotoxicity: Challenges in Cardio-Oncology. Cardio-Oncology; Gottlieb, RA, Mehta, PK, Eds; Academic Press: Boston, MA, USA, 2017:25-34.
- Pons DG: Roles of Phytochemicals in Cancer Prevention and Therapeutics. In., vol. 25: MDPI; 2024: 5450.
- Zhang YY, Zhang F, Zhang YS, Thakur K, Zhang JG, Liu Y, Kan H, Wei ZJ: Mechanism of Juglone-Induced Cell Cycle Arrest and Apoptosis in Ishikawa Human Endometrial Cancer Cells. Journal of agricultural and food chemistry 2019, 67, 7378–7389. [CrossRef]
- Sun W, Bao J, Lin W, Gao H, Zhao W, Zhang Q, Leung C-H, Ma D-L, Lu J, Chen X: 2-Methoxy-6-acetyl-7-methyljuglone (MAM), a natural naphthoquinone, induces NO-dependent apoptosis and necroptosis by H2O2-dependent JNK activation in cancer cells. Free Radical Biology and Medicine, 2016; 92, 61–77.
- Ock CW, Kim GD: Dioscin decreases breast cancer stem-like cell proliferation via cell cycle arrest by modulating p38 mitogen-activated protein kinase and AKT/mTOR signaling pathways. Journal of cancer prevention 2021, 26, 183. [CrossRef]
- Shah MA, Abuzar SM, Ilyas K, Qadees I, Bilal M, Yousaf R, Kassim RMT, Rasul A, Saleem U, Alves MS: Ginsenosides in cancer: Targeting cell cycle arrest and apoptosis. Chemico-biological interactions, 2023; 110634.
- Aggarwal V, Tuli HS, Kaur J, Aggarwal D, Parashar G, Chaturvedi Parashar N, Kulkarni S, Kaur G, Sak K, Kumar M: Garcinol exhibits anti-neoplastic effects by targeting diverse oncogenic factors in tumor cells. Biomedicines 2020, 8, 103.
- Kamiya T, Nishihara H, Hara H, Adachi T: Ethanol extract of Brazilian red propolis induces apoptosis in human breast cancer MCF-7 cells through endoplasmic reticulum stress. Journal of agricultural and food chemistry 2012, 60, 11065–11070. [CrossRef] [PubMed]
- Chu S-C, Hsieh Y-S, Yu C-C, Lai Y-Y, Chen P-N: Thymoquinone induces cell death in human squamous carcinoma cells via caspase activation-dependent apoptosis and LC3-II activation-dependent autophagy. PloS one 2014, 9, e101579.
- Shanmugam MK, Ahn KS, Hsu A, Woo CC, Yuan Y, Tan KHB, Chinnathambi A, Alahmadi TA, Alharbi SA, Koh APF: Thymoquinone inhibits bone metastasis of breast cancer cells through abrogation of the CXCR4 signaling axis. Frontiers in pharmacology, 2018; 1294.
- Hu R, Zhou P, Peng Y-B, Xu X, Ma J, Liu Q, Zhang L, Wen X-D, Qi L-W, Gao N: 6-Shogaol induces apoptosis in human hepatocellular carcinoma cells and exhibits anti-tumor activity in vivo through endoplasmic reticulum stress. PloS one 2012, 7, e39664.
- Jiang Q, Rao X, Kim CY, Freiser H, Zhang Q, Jiang Z, Li G: Gamma-tocotrienol induces apoptosis and autophagy in prostate cancer cells by increasing intracellular dihydrosphingosine and dihydroceramide. International journal of cancer 2012, 130, 685–693. [CrossRef]
- Yang KM, Kim BM, Park J-B: ω-Hydroxyundec-9-enoic acid induces apoptosis through ROS-mediated endoplasmic reticulum stress in non-small cell lung cancer cells. Biochemical and Biophysical Research Communications 2014, 448, 267–273. [CrossRef] [PubMed]
- Augustynowicz D, Lemieszek MK, Strawa JW, Wiater A, Tomczyk M: Phytochemical profiling of extracts from rare Potentilla species and evaluation of their anticancer potential. International Journal of Molecular Sciences 2023, 24, 4836. [CrossRef]
- Augustynowicz D, Lemieszek MK, Strawa JW, Wiater A, Tomczyk M: Anticancer potential of acetone extracts from selected Potentilla species against human colorectal cancer cells. Frontiers in Pharmacology, 2022; 13, 1027315.
- Leitzmann C: Characteristics and health benefits of phytochemicals. Forschende Komplementärmedizin/Research in Complementary Medicine 2016, 23, 69–74.
- Li S, Tan HY, Wang N, Cheung F, Hong M, Feng Y: The potential and action mechanism of polyphenols in the treatment of liver diseases. Oxidative medicine and cellular longevity 2018, 2018, 8394818. [CrossRef]
- Elekofehinti OO, Iwaloye O, Olawale F, Ariyo EO: Saponins in cancer treatment: Current progress and future prospects. Pathophysiology 2021, 28, 250–272. [CrossRef]
- Moran NE, Mohn ES, Hason N, Erdman Jr JW, Johnson EJ: Intrinsic and extrinsic factors impacting absorption, metabolism, and health effects of dietary carotenoids. Advances in Nutrition 2018, 9, 465–492. [CrossRef] [PubMed]
- Rudzińska A, Juchaniuk P, Oberda J, Wiśniewska J, Wojdan W, Szklener K, Mańdziuk S: Phytochemicals in cancer treatment and cancer prevention—review on epidemiological data and clinical trials. Nutrients 2023, 15, 1896. [CrossRef]
- Khan UM, Sevindik M, Zarrabi A, Nami M, Ozdemir B, Kaplan DN, Selamoglu Z, Hasan M, Kumar M, Alshehri MM: Lycopene: Food sources, biological activities, and human health benefits. Oxidative medicine and cellular longevity 2021, 2021, 2713511. [CrossRef] [PubMed]
- Krinsky NI, Johnson EJ: Carotenoid actions and their relation to health and disease. Molecular aspects of medicine 2005, 26, 459–516. [CrossRef]
- Niranjana R, Gayathri R, Mol SN, Sugawara T, Hirata T, Miyashita K, Ganesan P: Carotenoids modulate the hallmarks of cancer cells. Journal of functional foods, 2015; 18, 968–985.
- Jomova K, Valko M: Health protective effects of carotenoids and their interactions with other biological antioxidants. European journal of medicinal chemistry, 2013; 70, 102–110.
- Rutz JK, Borges CD, Zambiazi RC, da Rosa CG, da Silva MM: Elaboration of microparticles of carotenoids from natural and synthetic sources for applications in food. Food chemistry, 2016; 202, 324–333.
- Tapiero H, Townsend DM, Tew KD: The role of carotenoids in the prevention of human pathologies. Biomedicine & Pharmacotherapy 2004, 58, 100–110.
- Yang D-J, Lin J-T, Chen Y-C, Liu S-C, Lu F-J, Chang T-J, Wang M, Lin H-W, Chang Y-Y: Suppressive effect of carotenoid extract of Dunaliella salina alga on production of LPS-stimulated pro-inflammatory mediators in RAW264. 7 cells via NF-κB and JNK inactivation. Journal of Functional Foods 2013, 5, 607–615. [Google Scholar]
- Amin A, Hamza AA, Bajbouj K, Ashraf SS, Daoud S: Saffron: a potential candidate for a novel anticancer drug against hepatocellular carcinoma. Hepatology 2011, 54, 857–867. [CrossRef]
- Zou G, Zhang X, Wang L, Li X, Xie T, Zhao J, Yan J, Wang L, Ye H, Jiao S: Herb-sourced emodin inhibits angiogenesis of breast cancer by targeting VEGFA transcription. Theranostics 2020, 10, 6839. [CrossRef]
- Fu M, Tang W, Liu J-J, Gong X-Q, Kong L, Yao X-M, Jing M, Cai F-Y, Li X-T, Ju R-J: Combination of targeted daunorubicin liposomes and targeted emodin liposomes for treatment of invasive breast cancer. Journal of drug targeting 2020, 28, 245–258. [CrossRef]
- Shen Z, Zhao L, Yoo S-a, Lin Z, Zhang Y, Yang W, Piao J: Emodin induces ferroptosis in colorectal cancer through NCOA4-mediated ferritinophagy and NF-κb pathway inactivation. Apoptosis, 2024; 1–14.
- Shi M, Chen Z, Gong H, Peng Z, Sun Q, Luo K, Wu B, Wen C, Lin W: Luteolin, a flavone ingredient: Anticancer mechanisms, combined medication strategy, pharmacokinetics, clinical trials, and pharmaceutical researches. Phytotherapy Research 2024, 38, 880–911. [CrossRef]
- Gu X, Peng Y, Zhao Y, Liang X, Tang Y, Liu J: A novel derivative of artemisinin inhibits cell proliferation and metastasis via down-regulation of cathepsin K in breast cancer. European Journal of Pharmacology, 2019; 858, 172382.
- Tong X, Chen L, He S-j, Zuo J-p: Artemisinin derivative SM934 in the treatment of autoimmune and inflammatory diseases: therapeutic effects and molecular mechanisms. Acta Pharmacologica Sinica 2022, 43, 3055–3061. [CrossRef] [PubMed]
- Holdhoff M, Nicholas MK, Peterson RA, Maraka S, Liu LC, Fischer JH, Wefel JS, Fan TM, Vannorsdall T, Russell M et al: Phase I dose-escalation study of procaspase-activating compound-1 in combination with temozolomide in patients with recurrent high-grade astrocytomas. Neuro-oncology advances 2023, 5, vdad087.
- Sminia P, van den Berg J, van Kootwijk A, Hageman E, Slotman BJ, Verbakel W: Experimental and clinical studies on radiation and curcumin in human glioma. J Cancer Res Clin Oncol 2021, 147, 403–409. [CrossRef]
- Medical University of South C: Phase I Assay-guided Trial of Anti-inflammatory Phytochemicals in Patients With Advanced Cancer. clinicaltrialsgov 2013.
- Paur I, Lilleby W, Bøhn SK, Hulander E, Klein W, Vlatkovic L, Axcrona K, Bolstad N, Bjøro T, Laake P: Tomato-based randomized controlled trial in prostate cancer patients: Effect on PSA. Clinical Nutrition 2017, 36, 672–679. [CrossRef]
- https://clinicaltrials.gov, 2024.
- Hamblin MR: Shining light on the head: photobiomodulation for brain disorders. BBA clinical, 2016; 6, 113–124.
- Zhen X, Cheng P, Pu K: Recent advances in cell membrane–camouflaged nanoparticles for cancer phototherapy. Small 2019, 15, 1804105. [CrossRef]
- Pivetta TP, Botteon CE, Ribeiro PA, Marcato PD, Raposo M: Nanoparticle systems for cancer phototherapy: An overview. Nanomaterials 2021, 11, 3132. [CrossRef] [PubMed]
- He Z, Zhao L, Zhang Q, Chang M, Li C, Zhang H, Lu Y, Chen Y: An acceptor–donor–acceptor structured small molecule for effective NIR triggered dual phototherapy of cancer. Advanced Functional Materials 2020, 30, 1910301. [CrossRef]
- Liu P, Yang W, Shi L, Zhang H, Xu Y, Wang P, Zhang G, Chen WR, Zhang B, Wang X: Concurrent photothermal therapy and photodynamic therapy for cutaneous squamous cell carcinoma by gold nanoclusters under a single NIR laser irradiation. Journal of Materials Chemistry B 2019, 7, 6924–6933. [CrossRef]
- Agostinis P, Berg K, Cengel KA, Foster TH, Girotti AW, Gollnick SO, Hahn SM, Hamblin MR, Juzeniene A, Kessel D: Photodynamic therapy of cancer: an update. CA: a cancer journal for clinicians 2011, 61, 250–281.
- Oniszczuk A, Wojtunik-Kulesza KA, Oniszczuk T, Kasprzak K: The potential of photodynamic therapy (PDT)—Experimental investigations and clinical use. Biomedicine & Pharmacotherapy.
- Kim M, Jung HY, Park HJ: Topical PDT in the treatment of benign skin diseases: principles and new applications. International journal of molecular sciences 2015, 16, 23259–23278. [CrossRef] [PubMed]
- Wen X, Li Y, Hamblin MR: Photodynamic therapy in dermatology beyond non-melanoma cancer: An update. Photodiagnosis and photodynamic therapy, 2017; 19, 140–152.
- Ailioaie LM, Litscher G: Curcumin and photobiomodulation in chronic viral hepatitis and hepatocellular carcinoma. International Journal of Molecular Sciences 2020, 21, 7150. [CrossRef] [PubMed]
- Jiang S, Zhu R, He X, Wang J, Wang M, Qian Y, Wang S: Enhanced photocytotoxicity of curcumin delivered by solid lipid nanoparticles. International Journal of Nanomedicine, 2017; 167–178.
- Machado FC, de Matos RPA, Primo FL, Tedesco AC, Rahal P, Calmon MF: Effect of curcumin-nanoemulsion associated with photodynamic therapy in breast adenocarcinoma cell line. Bioorganic & Medicinal Chemistry 2019, 27, 1882–1890.
- Monge-Fuentes V, Muehlmann LA, Longo JPF, Silva JR, Fascineli ML, de Souza P, Faria F, Degterev IA, Rodriguez A, Carneiro FP: Photodynamic therapy mediated by acai oil (Euterpe oleracea Martius) in nanoemulsion: A potential treatment for melanoma. Journal of Photochemistry and Photobiology B: Biology, 2017; 166, 301–310.
- Semeraro P, Chimienti G, Altamura E, Fini P, Rizzi V, Cosma P: Chlorophyll a in cyclodextrin supramolecular complexes as a natural photosensitizer for photodynamic therapy (PDT) applications. Materials Science and Engineering: C, 2018; 85, 47–56.
- Wang K, Xiang Y, Pan W, Wang H, Li N, Tang B: Dual-targeted photothermal agents for enhanced cancer therapy. Chemical science 2020, 11, 8055–8072. [CrossRef]
- Abadeer NS, Murphy CJ: Recent progress in cancer thermal therapy using gold nanoparticles. Nanomaterials and Neoplasms, 2021; 143–217.
- Zou L, Wang H, He B, Zeng L, Tan T, Cao H, He X, Zhang Z, Guo S, Li Y: Current approaches of photothermal therapy in treating cancer metastasis with nanotherapeutics. Theranostics 2016, 6, 762. [CrossRef]
- Wang J, Wu X, Shen P, Wang J, Shen Y, Shen Y, Webster TJ, Deng J: Applications of inorganic nanomaterials in photothermal therapy based on combinational cancer treatment. International Journal of Nanomedicine, 2020; 1903–1914.
- Fernandes N, Rodrigues CF, Moreira AF, Correia IJ: Overview of the application of inorganic nanomaterials in cancer photothermal therapy. Biomaterials science 2020, 8, 2990–3020. [CrossRef]
- Tafech A, Stéphanou A: On the importance of acidity in cancer cells and therapy. Biology 2024, 13, 225.
- Mendes R, Pedrosa P, Lima JC, Fernandes AR, Baptista PV: Photothermal enhancement of chemotherapy in breast cancer by visible irradiation of Gold Nanoparticles. Scientific Reports 2017, 7, 10872.
- Li H, Zhang N, Hao Y, Wang Y, Jia S, Zhang H: Enhancement of curcumin antitumor efficacy and further photothermal ablation of tumor growth by single-walled carbon nanotubes delivery system in vivo. Drug delivery 2019, 26, 1017–1026. [CrossRef]
- Bano S, Nazir S, Nazir A, Munir S, Mahmood T, Afzal M, Ansari FL, Mazhar K: Microwave-assisted green synthesis of superparamagnetic nanoparticles using fruit peel extracts: surface engineering, T 2 relaxometry, and photodynamic treatment potential. International Journal of Nanomedicine, 2016; 3833–3848.
- Kharey P, Dutta SB, Manikandan M, Palani I, Majumder S, Gupta S: Green synthesis of near-infrared absorbing eugenate capped iron oxide nanoparticles for photothermal application. Nanotechnology 2019, 31, 095705.
- Ashkbar A, Rezaei F, Attari F, Ashkevarian S: Treatment of breast cancer in vivo by dual photodynamic and photothermal approaches with the aid of curcumin photosensitizer and magnetic nanoparticles. Scientific reports 2020, 10, 21206.
- Mun ST, Bae DH, Ahn WS: Epigallocatechin gallate with photodynamic therapy enhances anti-tumor effects in vivo and in vitro. Photodiagnosis and photodynamic therapy 2014, 11, 141–147. [CrossRef] [PubMed]
- Zhao Y, Zhang L, Shen J, Ma L, Wang L: Effects of Nutrients/Nutrition on Toxicants/Toxicity. In: Nutritional Toxicology. Springer; 2022: 1-28.
- Duda-Chodak A, Tarko T: Possible side effects of polyphenols and their interactions with medicines. Molecules 2023, 28, 2536. [CrossRef]
- Alwhaibi AM, Alshamrani AA, Alenazi MA, Altwalah SF, Alameel NN, Aljabali NN, Alghamdi SB, Bineid AI, Alwhaibi M, Al Arifi MN: Vincristine-induced neuropathy in patients diagnosed with solid and hematological malignancies: the role of dose rounding. Journal of Clinical Medicine 2023, 12, 5662. [CrossRef] [PubMed]
- Madsen ML, Due H, Ejskjær N, Jensen P, Madsen J, Dybkær K: Aspects of vincristine-induced neuropathy in hematologic malignancies: a systematic review. Cancer chemotherapy and pharmacology, 2019; 84, 471–485.
- Tang Z, Zhang Q: The potential toxic side effects of flavonoids. Biocell 2022, 46, 357.
- Bode AM, Dong Z: Toxic phytochemicals and their potential risks for human cancer. Cancer prevention research 2015, 8, 1–8. [CrossRef]
- Laqueur G, Spatz M: Toxicology of cycasin. Cancer Research 1968, 28, 2262–2267.
- Kisby GE, Fry RC, Lasarev MR, Bammler TK, Beyer RP, Churchwell M, Doerge DR, Meira LB, Palmer VS, Ramos-Crawford A-L: The cycad genotoxin MAM modulates brain cellular pathways involved in neurodegenerative disease and cancer in a DNA damage-linked manner. PLoS One 2011, 6, e20911.
- Niemeyer HB, Honig DM, Kulling SE, Metzler M: Studies on the metabolism of the plant lignans secoisolariciresinol and matairesinol. Journal of agricultural and food chemistry 2003, 51, 6317–6325. [CrossRef]
- Ward HA, Kuhnle GG, Mulligan AA, Lentjes MA, Luben RN, Khaw K-T: Breast, colorectal, and prostate cancer risk in the European Prospective Investigation into Cancer and Nutrition–Norfolk in relation to phytoestrogen intake derived from an improved database. The American journal of clinical nutrition 2010, 91, 440–448. [CrossRef]
- van Duursen MB, Nijmeijer S, De Morree E, de Jong PC, van den Berg M: Genistein induces breast cancer-associated aromatase and stimulates estrogen-dependent tumor cell growth in in vitro breast cancer model. Toxicology 2011, 289(2-3):67-73.
- Ju YH, Doerge DR, Woodling KA, Hartman JA, Kwak J, Helferich WG: Dietary genistein negates the inhibitory effect of letrozole on the growth of aromatase-expressing estrogen-dependent human breast cancer cells (MCF-7Ca) in vivo. Carcinogenesis 2008, 29, 2162–2168. [CrossRef] [PubMed]
- Zhang H, Cifone M, Murli H, Erexson G, Mecchi M, Lawlor T: Application of simplified in vitro screening tests to detect genotoxicity of aristolochic acid. Food and chemical toxicology 2004, 42, 2021–2028. [CrossRef] [PubMed]
- Arlt VM, Stiborova M, Schmeiser HH: Aristolochic acid as a probable human cancer hazard in herbal remedies: a review. Mutagenesis 2002, 17, 265–277. [CrossRef] [PubMed]
- Han ZT, Tong YK, He LM, Zhang Y, Sun JZ, Wang TY, Zhang H, Cui YL, Newmark HL, Conney AH: 12-O-Tetradecanoylphorbol-13-acetate (TPA)-induced increase in depressed white blood cell counts in patients treated with cytotoxic cancer chemotherapeutic drugs. Proceedings of the National Academy of Sciences 1998, 95, 5362–5365. [CrossRef]
- Afrasiabi E, Ahlgren J, Bergelin N, Törnquist K: Phorbol 12-myristate 13-acetate inhibits FRO anaplastic human thyroid cancer cell proliferation by inducing cell cycle arrest in G1/S phase: Evidence for an effect mediated by PKCδ. Molecular and cellular endocrinology 2008, 292(1-2):26-35.
- Zheng X, Chang RL, Cui X-X, Avila GE, Hebbar V, Garzotto M, Shih WJ, Lin Y, Lu S-E, Rabson AB: Effects of 12-O-tetradecanoylphorbol-13-acetate (TPA) in combination with paclitaxel (Taxol) on prostate Cancer LNCaP cells cultured in vitro or grown as xenograft tumors in immunodeficient mice. Clinical cancer research 2006, 12, 3444–3451.
- Fürstenberger G, Berry D, Sorg B, Marks F: Skin tumor promotion by phorbol esters is a two-stage process. Proceedings of the National Academy of Sciences 1981, 78, 7722–7726. [CrossRef]
- Schoental R: Toxicology and carcinogenic action of pyrrolizidine alkaloids. Cancer Research 1968, 28, 2237–2246.
- Zhao Y, Xia Q, Yin JJ, Lin G, Fu PP: Photoirradiation of dehydropyrrolizidine alkaloids—formation of reactive oxygen species and induction of lipid peroxidation. Toxicology letters 2011, 205, 302–309. [CrossRef] [PubMed]
- Weisbord SD, Soule JB, Kimmel PL: Poison on line—acute renal failure caused by oil of wormwood purchased through the Internet. New England journal of medicine 1997, 337, 825–827. [CrossRef]
- Winickoff JP, Houck CS, Rothman EL, Bauchner H: Verve and Jolt: deadly new Internet drugs. Pediatrics 2000, 106, 829–830. [CrossRef]
- Komarova TV, Baschieri S, Donini M, Marusic C, Benvenuto E, Dorokhov YL: Transient expression systems for plant-derived biopharmaceuticals. Expert review of vaccines 2010, 9, 859–876. [CrossRef] [PubMed]
- Verdeil J-L, Alemanno L, Niemenak N, Tranbarger TJ: Pluripotent versus totipotent plant stem cells: dependence versus autonomy? Trends in plant science 2007, 12, 245–252. [CrossRef] [PubMed]
- Ko K, Steplewski Z, Glogowska M, Koprowski H: Inhibition of tumor growth by plant-derived mAb. Proceedings of the National Academy of Sciences 2005, 102, 7026–7030. [CrossRef] [PubMed]
- Kopertekh L, Schiemann J: Transient production of recombinant pharmaceutical proteins in plants: evolution and perspectives. Current Medicinal Chemistry 2019, 26, 365–380. [CrossRef] [PubMed]
- Buyel JF, Twyman RM, Fischer R: Very-large-scale production of antibodies in plants: The biologization of manufacturing. Biotechnology Advances 2017, 35, 458–465. [CrossRef]
- Daniell H: Medical molecular pharming: expression of antibodies, biopharmaceuticals and edible vaccines via the chloroplast genome. In: Plant Biotechnology 2002 and Beyond: Proceedings of the 10 th IAPTC&B Congress –28, 2002 Orlando, Florida, USA: 2003. Springer: 371-376. 23 June.
- McCormick AA, Kumagai MH, Hanley K, Turpen TH, Hakim I, Grill LK, Tusé D, Levy S, Levy R: Rapid production of specific vaccines for lymphoma by expression of the tumor-derived single-chain Fv epitopes in tobacco plants. Proceedings of the National Academy of Sciences 1999, 96, 703–708. [CrossRef]
- Verch T, Yusibov V, Koprowski H: Expression and assembly of a full-length monoclonal antibody in plants using a plant virus vector. Journal of immunological methods 1998, 220(1-2):69-75.
- Nessa MU, Rahman MA, Kabir Y: Plant-produced monoclonal antibody as immunotherapy for cancer. BioMed Research International 2020, 2020.
- Tusé D, Ku N, Bendandi M, Becerra C, Collins R, Langford N, Sancho SI, López-Díaz de Cerio A, Pastor F, Kandzia R: Clinical safety and immunogenicity of tumor-targeted, plant-made Id-KLH conjugate vaccines for follicular lymphoma. BioMed Research International 2015, 2015.
- Gronenborn B, Matzeit V: Plant gene vectors and genetic transformation: plant viruses as vectors. In: Molecular Biology of Plant Nuclear Genes. Elsevier; 1989: 69-100.
- Xu J, Dolan MC, Medrano G, Cramer CL, Weathers PJ: Green factory: plants as bioproduction platforms for recombinant proteins. Biotechnology advances 2012, 30, 1171–1184. [CrossRef]
- Donini M, Marusic C: Current state-of-the-art in plant-based antibody production systems. Biotechnology letters 2019, 41:335-346.
- Houdelet M, Galinski A, Holland T, Wenzel K, Schillberg S, Buyel JF: Animal component-free Agrobacterium tumefaciens cultivation media for better GMP-compliance increases biomass yield and pharmaceutical protein expression in Nicotiana benthamiana. Biotechnology Journal 2017, 12, 1600721.
- Bulaon CJI, Khorattanakulchai N, Rattanapisit K, Sun H, Pisuttinusart N, Phoolcharoen W: Development of Plant-Derived Bispecific Monoclonal Antibody Targeting PD-L1 and CTLA-4 against Mouse Colorectal Cancer. Planta Medica 2024.
- Lee JH, Park SR, Phoolcharoen W, Ko K: Expression, function, and glycosylation of anti-colorectal cancer large single-chain antibody (LSC) in plant. Plant Biotechnology Reports 2020, 14:363-371.
- Park SR, Lee J-H, Kim K, Kim TM, Lee SH, Choo Y-K, Kim KS, Ko K: Expression and in vitro function of anti-breast cancer llama-based single domain antibody VHH expressed in tobacco plants. International journal of molecular sciences 2020, 21, 1354. [CrossRef] [PubMed]
- González-Gamboa I, Caparco AA, McCaskill J, Fuenlabrada-Velázquez P, Hays SS, Jin Z, Jokerst JV, Pokorski JK, Steinmetz NF: Inter-coat protein loading of active ingredients into Tobacco mild green mosaic virus through partial dissociation and reassembly of the virion. Scientific Reports 2024, 14, 7168.
- Bulaon CJI, Khorattanakulchai N, Rattanapisit K, Sun H, Pisuttinusart N, Phoolcharoen W: Development of Plant-Derived Bispecific Monoclonal Antibody Targeting PD-L1 and CTLA-4 against Mouse Colorectal Cancer. Planta Medica 2024, 90, 305–315. [CrossRef]
- Bulaon CJI, Khorattanakulchai N, Rattanapisit K, Sun H, Pisuttinusart N, Strasser R, Tanaka S, Soon-Shiong P, Phoolcharoen W: Antitumor effect of plant-produced anti-CTLA-4 monoclonal antibody in a murine model of colon cancer. Frontiers in Plant Science, 2023; 14, 1149455.
- Rattanapisit K, Bulaon CJI, Strasser R, Sun H, Phoolcharoen W: In vitro and in vivo studies of plant-produced Atezolizumab as a potential immunotherapeutic antibody. Scientific Reports 2023, 13, 14146.
- Bulaon CJI, Sun H, Malla A, Phoolcharoen W: Therapeutic efficacy of plant-produced Nivolumab in transgenic C57BL/6-hPD-1 mouse implanted with MC38 colon cancer. Biotechnology Reports, 2023; 38, e00794.
- Izadi S, Gumpelmair S, Coelho P, Duarte HO, Gomes J, Leitner J, Kunnummel V, Mach L, Reis CA, Steinberger P: Plant-derived Durvalumab variants show efficient PD-1/PD-L1 blockade and therapeutically favourable FcR binding. Plant Biotechnology Journal 2024, 22, 1224–1237. [CrossRef]
- Shin JH, Oh S, Jang MH, Lee SY, Min C, Eu YJ, Begum H, Kim JC, Lee GR, Oh HB: Enhanced efficacy of glycoengineered rice cell-produced trastuzumab. Plant Biotechnology Journal 2024.
- Stark MC, Joubert AM, Visagie MH: Molecular farming of pembrolizumab and nivolumab. International Journal of Molecular Sciences 2023, 24, 10045. [CrossRef]
- Nguyen KD, Kajiura H, Kamiya R, Yoshida T, Misaki R, Fujiyama K: Production and N-glycan engineering of Varlilumab in Nicotiana benthamiana. Frontiers in Plant Science, 2023; 14, 1215580.
- Park C, Kim K, Kim Y, Zhu R, Hain L, Seferovic H, Kim M-H, Woo HJ, Hwang H, Lee SH: Plant-Derived Anti-Human Epidermal Growth Factor Receptor 2 Antibody Suppresses Trastuzumab-Resistant Breast Cancer with Enhanced Nanoscale Binding. ACS nano 2024.
- Jin C, Kang YJ, Park SR, Oh YJ, Ko K: Production, expression, and function of dual-specific monoclonal antibodies in a single plant. Planta 2024, 259, 15. [CrossRef]
- Lu R-M, Hwang Y-C, Liu I-J, Lee C-C, Tsai H-Z, Li H-J, Wu H-C: Development of therapeutic antibodies for the treatment of diseases. Journal of biomedical science, 2020; 27, 1–30.
- Phakham T, Bulaon CJI, Khorattanakulchai N, Shanmugaraj B, Buranapraditkun S, Boonkrai C, Sooksai S, Hirankarn N, Abe Y, Strasser R: Functional characterization of pembrolizumab produced in Nicotiana benthamiana using a rapid transient expression system. Frontiers in Plant Science, 2021; 12, 736299.
- Rattanapisit K, Phakham T, Buranapraditkun S, Siriwattananon K, Boonkrai C, Pisitkun T, Hirankarn N, Strasser R, Abe Y, Phoolcharoen W: Structural and in vitro functional analyses of novel plant-produced anti-human PD1 antibody. Scientific reports 2019, 9, 15205.
- Alsaab HO, Sau S, Alzhrani R, Tatiparti K, Bhise K, Kashaw SK, Iyer AK: PD-1 and PD-L1 checkpoint signaling inhibition for cancer immunotherapy: mechanism, combinations, and clinical outcome. Frontiers in pharmacology, 2017; 8, 561.
- Mattila PO, Babar Z-U-D, Suleman F: Assessing the prices and affordability of oncology medicines for three common cancers within the private sector of South Africa. BMC Health Services Research, 2021; 21, 1–10.
- Steele JF, Peyret H, Saunders K, Castells-Graells R, Marsian J, Meshcheriakova Y, Lomonossoff GP: Synthetic plant virology for nanobiotechnology and nanomedicine. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2017, 9, e1447.
- Shahgolzari M, Pazhouhandeh M, Milani M, Yari Khosroushahi A, Fiering S: Plant viral nanoparticles for packaging and in vivo delivery of bioactive cargos. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2020, 12, e1629.
- Steinmetz NF, Masarapu H, He H: Tymovirus virus and virus-like particles as nanocarriers for imaging and therapeutic agents. In.: Google Patents; 2023.
- Ortega-Rivera OA, Beiss V, Osota EO, Chan SK, Karan S, Steinmetz NF: Production of cytoplasmic type citrus leprosis virus-like particles by plant molecular farming. Virology.
- Nikitin N, Trifonova E, Karpova O, Atabekov J: Biosafety of plant viruses for human and animals. Moscow University biological sciences bulletin, 2016; 71, 128–134.
- Wen AM, Steinmetz NF: Design of virus-based nanomaterials for medicine, biotechnology, and energy. Chemical Society Reviews 2016, 45, 4074–4126. [CrossRef]
- Sherman MB, Guenther RH, Tama F, Sit TL, Brooks CL, Mikhailov AM, Orlova EV, Baker TS, Lommel SA: Removal of divalent cations induces structural transitions in red clover necrotic mosaic virus, revealing a potential mechanism for RNA release. Journal of virology 2006, 80, 10395–10406. [CrossRef]
- Czapar AE, Steinmetz NF: Plant viruses and bacteriophages for drug delivery in medicine and biotechnology. Current opinion in chemical biology, 2017; 38, 108–116.
- Kim SM, Faix PH, Schnitzer JE: Overcoming key biological barriers to cancer drug delivery and efficacy. Journal of Controlled Release, 2017; 267, 15–30.
- Cheng X, Xie Q, Sun Y: Advances in nanomaterial-based targeted drug delivery systems. Frontiers in bioengineering and biotechnology, 2023; 11, 1177151.
- Chariou PL, Lee KL, Wen AM, Gulati NM, Stewart PL, Steinmetz NF: Detection and imaging of aggressive cancer cells using an epidermal growth factor receptor (EGFR)-targeted filamentous plant virus-based nanoparticle. Bioconjugate chemistry 2015, 26, 262–269. [CrossRef]
- Destito G, Yeh R, Rae CS, Finn M, Manchester M: Folic acid-mediated targeting of cowpea mosaic virus particles to tumor cells. Chemistry & biology 2007, 14, 1152–1162.
- Cho C-F, Yu L, Nsiama TK, Kadam AN, Raturi A, Shukla S, Amadei GA, Steinmetz NF, Luyt LG, Lewis JD: Viral nanoparticles decorated with novel EGFL7 ligands enable intravital imaging of tumor neovasculature. Nanoscale 2017, 9, 12096–12109. [CrossRef]
- Marchetti L, Novelli F, Tanno B, Leonardi S, Hizam VM, Arcangeli C, Santi L, Baschieri S, Lico C, Mancuso M: Peptide-Functionalized and Drug-Loaded Tomato Bushy Stunt Virus Nanoparticles Counteract Tumor Growth in a Mouse Model of Shh-Dependent Medulloblastoma. International Journal of Molecular Sciences 2023, 24, 8911.
- Le DH, Lee KL, Shukla S, Commandeur U, Steinmetz NF: Potato virus X, a filamentous plant viral nanoparticle for doxorubicin delivery in cancer therapy. Nanoscale 2017, 9, 2348–2357. [CrossRef]
- Lin RD, Steinmetz NF: Tobacco mosaic virus delivery of mitoxantrone for cancer therapy. Nanoscale 2018, 10, 16307–16313. [CrossRef] [PubMed]
- Czapar AE, Zheng Y-R, Riddell IA, Shukla S, Awuah SG, Lippard SJ, Steinmetz NF: Tobacco mosaic virus delivery of phenanthriplatin for cancer therapy. ACS nano 2016, 10, 4119–4126. [CrossRef] [PubMed]
- Parhizkar E, Rafieipour P, Sepasian A, Alemzadeh E, Dehshahri A, Ahmadi F: Synthesis and cytotoxicity evaluation of gemcitabine-tobacco mosaic virus conjugates. Journal of Drug Delivery Science and Technology, 2021; 62, 102388.
- Alemzadeh E, Dehshahri A, Dehghanian AR, Afsharifar A, Behjatnia AA, Izadpanah K, Ahmadi F: Enhanced anti-tumor efficacy and reduced cardiotoxicity of doxorubicin delivered in a novel plant virus nanoparticle. Colloids and Surfaces B: Biointerfaces, 2019; 174, 80–86.
- Cao J, Guenther RH, Sit TL, Opperman CH, Lommel SA, Willoughby JA: Loading and release mechanism of red clover necrotic mosaic virus derived plant viral nanoparticles for drug delivery of doxorubicin. Small 2014, 10, 5126–5136. [CrossRef]
- Aljabali AA, Shukla S, Lomonossoff GP, Steinmetz NF, Evans DJ: Cpmv-dox delivers. Molecular pharmaceutics 2013, 10, 3–10. [CrossRef]
- Franke CE, Czapar AE, Patel RB, Steinmetz NF: Tobacco mosaic virus-delivered cisplatin restores efficacy in platinum-resistant ovarian cancer cells. Molecular pharmaceutics 2017, 15, 2922–2931.
- Le DH, Commandeur U, Steinmetz NF: Presentation and delivery of tumor necrosis factor-related apoptosis-inducing ligand via elongated plant viral nanoparticle enhances antitumor efficacy. ACS nano 2019, 13, 2501–2510.
- Chan SK, Steinmetz NF: microRNA-181a silencing by antisense oligonucleotides delivered by virus-like particles. Journal of Materials Chemistry B 2023, 11, 816–825. [CrossRef]
- Kim KR, Lee AS, Kim SM, Heo HR, Kim CS: Virus-like nanoparticles as a theranostic platform for cancer. Frontiers in Bioengineering and Biotechnology, 2023; 10, 1106767.
- Lam P, Lin RD, Steinmetz NF: Delivery of mitoxantrone using a plant virus-based nanoparticle for the treatment of glioblastomas. Journal of Materials Chemistry B 2018, 6, 5888–5895. [CrossRef]
- Kernan DL, Wen AM, Pitek AS, Steinmetz NF: Featured Article: Delivery of chemotherapeutic vcMMAE using tobacco mosaic virus nanoparticles. Experimental Biology and Medicine 2017, 242, 1405–1411. [CrossRef]
- Esfandiari N, Arzanani MK, Soleimani M, Kohi-Habibi M, Svendsen WE: A new application of plant virus nanoparticles as drug delivery in breast cancer. Tumor Biology, 2016; 37, 1229–1236.
- Esfandiari N: Targeting breast cancer with bio-inspired virus nanoparticles. Archives of Breast Cancer, 2018; 90–95.
- Shukla S, Roe AJ, Liu R, Veliz FA, Commandeur U, Wald DN, Steinmetz NF: Affinity of plant viral nanoparticle potato virus X (PVX) towards malignant B cells enables cancer drug delivery. Biomaterials science 2020, 8, 3935–3943. [CrossRef]
- Marchetti L, Simon-Gracia L, Lico C, Mancuso M, Baschieri S, Santi L, Teesalu T: Targeting of Tomato Bushy Stunt Virus with a Genetically Fused C-End Rule Peptide. Nanomaterials 2023, 13, 1428. [CrossRef] [PubMed]
- Barkovich KJ, Zhao Z, Steinmetz NF: iRGD-Targeted Physalis Mottle Virus Like Nanoparticles for Targeted Cancer Delivery. Small Science 2023, 3, 2300067. [CrossRef] [PubMed]
- Shahgolzari M, Venkataraman S, Osano A, Akpa PA, Hefferon K: Plant Virus Nanoparticles Combat Cancer. Vaccines 2023, 11, 1278. [CrossRef]
- Chariou PL, Wang L, Desai C, Park J, Robbins LK, von Recum HA, Ghiladi RA, Steinmetz NF: Let there be light: Targeted photodynamic therapy using high aspect ratio plant viral nanoparticles. Macromolecular Bioscience 2019, 19, 1800407.
- Nkanga CI, Ortega-Rivera OA, Steinmetz NF: Photothermal immunotherapy of melanoma using TLR-7 agonist laden tobacco mosaic virus with polydopamine coat. Nanomedicine: Nanotechnology, Biology and Medicine, 2022; 44, 102573.
- Zhao Z, Simms A, Steinmetz NF: Cisplatin-loaded tobacco mosaic virus for ovarian cancer treatment. Biomacromolecules 2022, 23, 4379–4387. [CrossRef] [PubMed]
- Lam P, Steinmetz NF: Plant viral and bacteriophage delivery of nucleic acid therapeutics. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2018, 10, e1487.
- Shahgolzari M, Dianat-Moghadam H, Yavari A, Fiering SN, Hefferon K: Multifunctional plant virus nanoparticles for targeting breast cancer tumors. Vaccines 2022, 10, 1431. [CrossRef]
- Lam P, Steinmetz NF: Delivery of siRNA therapeutics using cowpea chlorotic mottle virus-like particles. Biomaterials science 2019, 7, 3138–3142. [CrossRef]
- Villagrana-Escareño MV, Reynaga-Hernández E, Galicia-Cruz OG, Durán-Meza AL, la Cruz-González D, Hernández-Carballo CY, Ruíz-García J: VLPs derived from the CCMV plant virus can directly transfect and deliver heterologous genes for translation into mammalian cells. BioMed Research International 2019, 2019.
- Xue F, Cornelissen JJ, Yuan Q, Cao S: Delivery of MicroRNAs by plant virus-based nanoparticles to functionally alter the osteogenic differentiation of human mesenchymal stem cells. Chinese Chemical Letters 2023, 34, 107448. [CrossRef]
- Nuñez-Rivera A, Fournier PG, Arellano DL, Rodriguez-Hernandez AG, Vazquez-Duhalt R, Cadena-Nava RD: Brome mosaic virus-like particles as siRNA nanocarriers for biomedical purposes. Beilstein Journal of Nanotechnology 2020, 11, 372–382. [CrossRef] [PubMed]
- Masarapu H, Patel BK, Chariou PL, Hu H, Gulati NM, Carpenter BL, Ghiladi RA, Shukla S, Steinmetz NF: Physalis mottle virus-like particles as nanocarriers for imaging reagents and drugs. Biomacromolecules 2017, 18, 4141–4153. [CrossRef] [PubMed]
- Yildiz I, Shukla S, Steinmetz NF: Applications of viral nanoparticles in medicine. Current opinion in biotechnology 2011, 22, 901–908. [CrossRef]
- Hu H, Zhang Y, Shukla S, Gu Y, Yu X, Steinmetz NF: Dysprosium-modified tobacco mosaic virus nanoparticles for ultra-high-field magnetic resonance and near-infrared fluorescence imaging of prostate cancer. ACS nano 2017, 11, 9249–9258. [CrossRef]
- Steinmetz NF, Ablack AL, Hickey JL, Ablack J, Manocha B, Mymryk JS, Luyt LG, Lewis JD: Intravital imaging of human prostate cancer using viral nanoparticles targeted to gastrin-releasing peptide receptors. Small 2011, 7, 1664–1672. [CrossRef] [PubMed]
- Shukla S, Dickmeis C, Nagarajan A, Fischer R, Commandeur U, Steinmetz N: Molecular farming of fluorescent virus-based nanoparticles for optical imaging in plants, human cells and mouse models. Biomaterials science 2014, 2, 784–797. [CrossRef] [PubMed]
- Bruckman MA, Randolph LN, Gulati NM, Stewart PL, Steinmetz NF: Silica-coated Gd (DOTA)-loaded protein nanoparticles enable magnetic resonance imaging of macrophages. Journal of materials chemistry B 2015, 3, 7503–7510. [CrossRef]
- Chung YH, Cai H, Steinmetz NF: Viral nanoparticles for drug delivery, imaging, immunotherapy, and theranostic applications. Advanced Drug Delivery Reviews, 2020; 56, 214–235.
- Wu Z, Zhou J, Nkanga CI, Jin Z, He T, Borum RM, Yim W, Zhou J, Cheng Y, Xu M: One-step supramolecular multifunctional coating on plant virus nanoparticles for bioimaging and therapeutic applications. ACS applied materials & interfaces 2022, 14, 13692–13702.
- Pitek A, Hu H, Shukla S, Steinmetz N: Cancer theranostic applications of albumin-coated tobacco mosaic virus nanoparticles. ACS applied materials & interfaces 2018, 10, 39468–39477.
- Luzuriaga MA, Welch RP, Dharmarwardana M, Benjamin CE, Li S, Shahrivarkevishahi A, Popal S, Tuong LH, Creswell CT, Gassensmith JJ: Enhanced stability and controlled delivery of MOF-encapsulated vaccines and their immunogenic response in vivo. ACS applied materials & interfaces 2019, 11, 9740–9746.
- Dharmarwardana M, Martins AF, Chen Z, Palacios PM, Nowak CM, Welch RP, Li S, Luzuriaga MA, Bleris L, Pierce BS: Nitroxyl modified tobacco mosaic virus as a metal-free high-relaxivity MRI and EPR active superoxide sensor. Molecular pharmaceutics 2018, 15, 2973–2983. [CrossRef] [PubMed]
- Bruckman MA, Jiang K, Simpson EJ, Randolph LN, Luyt LG, Yu X, Steinmetz NF: Dual-modal magnetic resonance and fluorescence imaging of atherosclerotic plaques in vivo using VCAM-1 targeted tobacco mosaic virus. Nano letters 2014, 14, 1551–1558. [CrossRef]
- Valdivia G, Pérez-Alenza D, Barreno L, Alonso-Diez Á, de Oliveira JFA, Suárez-Redondo M, Fiering SF, Steinmetz NF, Peña L: Innovative CPMV immunotherapy: A canine model for poor-prognosis breast cancer treatment. Cancer Research, 2024; 84, (6_Supplement):6663-6663.
- Shahgolzari M, Fiering S: Emerging potential of plant virus nanoparticles (PVNPs) in anticancer immunotherapies. Journal of cancer immunology 2022, 4, 22.
- Jung E, Chung YH, Mao C, Fiering SN, Steinmetz NF: The Potency of Cowpea Mosaic Virus Particles for Cancer In Situ Vaccination Is Unaffected by the Specific Encapsidated Viral RNA. Molecular Pharmaceutics 2023, 20, 3589–3597. [CrossRef]
- Mao C, Beiss V, Fields J, Steinmetz NF, Fiering S: Cowpea mosaic virus stimulates antitumor immunity through recognition by multiple MYD88-dependent toll-like receptors. Biomaterials 2021, 275:120914.
- Shukla S, Wang C, Beiss V, Cai H, Washington T, Murray AA, Gong X, Zhao Z, Masarapu H, Zlotnick A: The unique potency of Cowpea mosaic virus (CPMV) in situ cancer vaccine. Biomaterials science 2020, 8, 5489–5503. [CrossRef] [PubMed]
- Hu H, Steinmetz NF: Development of a virus-like particle-based anti-HER2 breast cancer vaccine. Cancers 2021, 13, 2909. [CrossRef] [PubMed]
- Barreno L, Sevane N, Valdivia G, Alonso-Miguel D, Suarez-Redondo M, Alonso-Diez A, Fiering S, Beiss V, Steinmetz NF, Perez-Alenza MD: Transcriptomics of canine inflammatory mammary cancer treated with empty cowpea mosaic virus implicates neutrophils in anti-tumor immunity. International Journal of Molecular Sciences 2023, 24, 14034.
- Shukla S, Myers JT, Woods SE, Gong X, Czapar AE, Commandeur U, Huang AY, Levine AD, Steinmetz NF: Plant viral nanoparticles-based HER2 vaccine: Immune response influenced by differential transport, localization and cellular interactions of particulate carriers. Biomaterials 2017, 121:15-27.
- Lebel M-È, Chartrand K, Tarrab E, Savard P, Leclerc D, Lamarre A: Potentiating cancer immunotherapy using papaya mosaic virus-derived nanoparticles. Nano letters 2016, 16, 1826–1832. [CrossRef] [PubMed]
- Murray AA, Wang C, Fiering S, Steinmetz NF: In situ vaccination with cowpea vs tobacco mosaic virus against melanoma. Molecular pharmaceutics 2018, 15, 3700–3716. [CrossRef]
- Cai H, Shukla S, Wang C, Masarapu H, Steinmetz NF: Heterologous prime-boost enhances the antitumor immune response elicited by plant-virus-based cancer vaccine. Journal of the American Chemical Society 2019, 141, 6509–6518. [CrossRef]
- Shahgolzari M, Pazhouhandeh M, Milani M, Fiering S, Khosroushahi AY: Alfalfa mosaic virus nanoparticles-based in situ vaccination induces antitumor immune responses in breast cancer model. Nanomedicine 2020, 16, 97–107.
- Zhao Z, Chung YH, Steinmetz NF: Melanoma immunotherapy enabled by M2 macrophage targeted immunomodulatory cowpea mosaic virus. Materials Advances 2024, 5, 1473–1479. [CrossRef] [PubMed]
- Shukla S, Jandzinski M, Wang C, Gong X, Bonk KW, Keri RA, Steinmetz NF: A viral nanoparticle cancer vaccine delays tumor progression and prolongs survival in a HER2+ tumor mouse model. Advanced therapeutics 2019, 2, 1800139. [CrossRef]
- Patel BK, Wang C, Lorens B, Levine AD, Steinmetz NF, Shukla S: Cowpea mosaic virus (CPMV)-based cancer testis antigen NY-ESO-1 vaccine elicits an antigen-specific cytotoxic T cell response. ACS applied bio materials 2020, 3, 4179–4187. [CrossRef]
- Cai H, Shukla S, Steinmetz NF: The antitumor efficacy of CpG oligonucleotides is improved by encapsulation in plant virus-like particles. Advanced functional materials 2020, 30, 1908743. [CrossRef]
- Iravani S, Varma RS: Vault, viral, and virus-like nanoparticles for targeted cancer therapy. Materials Advances 2023, 4, 2909–2917. [CrossRef]
- Jung E, Chung YH, Steinmetz NF: TLR Agonists Delivered by Plant Virus and Bacteriophage Nanoparticles for Cancer Immunotherapy. Bioconjugate Chemistry 2023, 34, 1596–1605. [CrossRef]
- Shin MD, Jung E, Moreno-Gonzalez MA, Ortega-Rivera OA, Steinmetz NF: Pluronic F127 “nanoarmor” for stabilization of Cowpea mosaic virus immunotherapy. Bioengineering & Translational Medicine 2024, 9, e10574.
- Boone CE, Wang C, Lopez-Ramirez MA, Beiss V, Shukla S, Chariou PL, Kupor D, Rueda R, Wang J, Steinmetz NF: Active microneedle administration of plant virus nanoparticles for cancer in situ vaccination improves immunotherapeutic efficacy. ACS applied nano materials 2020, 3, 8037–8051. [CrossRef]
- Patel R, Czapar AE, Fiering S, Oleinick NL, Steinmetz NF: Radiation therapy combined with cowpea mosaic virus nanoparticle in situ vaccination initiates immune-mediated tumor regression. ACS omega 2018, 3, 3702–3707. [CrossRef]
- Karan S, Jung E, Boone C, Steinmetz NF: Synergistic combination therapy using cowpea mosaic virus intratumoral immunotherapy and Lag-3 checkpoint blockade. Cancer Immunology, Immunotherapy 2024, 73, 51. [CrossRef] [PubMed]
- Koellhoffer EC, Steinmetz NF: Cowpea mosaic virus and natural killer cell agonism for in situ cancer vaccination. Nano letters 2022, 22, 5348–5356. [CrossRef] [PubMed]
- Gautam A, Beiss V, Wang C, Wang L, Steinmetz NF: Plant viral nanoparticle conjugated with anti-PD-1 peptide for ovarian cancer immunotherapy. International Journal of Molecular Sciences 2021, 22, 9733. [CrossRef] [PubMed]
- Lee KL, Murray AA, Le DHT, Sheen MR, Shukla S, Commandeur U, Fiering S, Steinmetz NF: Combination of Plant Virus Nanoparticle-Based in Situ Vaccination with Chemotherapy Potentiates Antitumor Response. Nano Letters 2017, 17, 4019–4028. [CrossRef] [PubMed]
- Cai H, Wang C, Shukla S, Steinmetz NF: Cowpea mosaic virus immunotherapy combined with cyclophosphamide reduces breast cancer tumor burden and inhibits lung metastasis. Advanced science 2019, 6, 1802281. [CrossRef]
- Zhao Z, Ortega-Rivera OA, Chung YH, Simms A, Steinmetz NF: A co-formulated vaccine of irradiated cancer cells and cowpea mosaic virus improves ovarian cancer rejection. Journal of Materials Chemistry B 2023, 11, 5429–5441. [CrossRef]
- Barkovich KJ, Wu Z, Zhao Z, Simms A, Chang EY, Steinmetz NF: Physalis Mottle Virus-Like Nanocarriers with Expanded Internal Loading Capacity. Bioconjugate Chemistry 2023, 34, 1585–1595. [CrossRef]
- Almalki WH: An Up-to-date Review on Protein-based Nanocarriers in the Management of Cancer. Current Drug Delivery 2024, 21, 509–524. [CrossRef]
- Arul SS, Balakrishnan B, Handanahal SS, Venkataraman S: Viral nanoparticles: Current advances in design and development. Biochimie 2023.
- Shen L, Zhou P, Wang YM, Zhu Z, Yuan Q, Cao S, Li J: Supramolecular nanoparticles based on elastin-like peptides modified capsid protein as drug delivery platform with enhanced cancer chemotherapy efficacy. International Journal of Biological Macromolecules 2024, 256:128107.
- Shah S, Famta P, Tiwari V, Kotha AK, Kashikar R, Chougule MB, Chung YH, Steinmetz NF, Uddin M, Singh SB: Instigation of the epoch of nanovaccines in cancer immunotherapy. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2023, 15, e1870.
- Chung YH, Zhao Z, Jung E, Omole AO, Wang H, Sutorus L, Steinmetz NF: Systemic Administration of Cowpea Mosaic Virus Demonstrates Broad Protection Against Metastatic Cancers. Advanced Science 2024:2308237.
- Valdivia G, Alonso-Miguel D, Perez-Alenza MD, Zimmermann ABE, Schaafsma E, Kolling IV FW, Barreno L, Alonso-Diez A, Beiss V, Affonso de Oliveira JF: Neoadjuvant intratumoral immunotherapy with cowpea mosaic virus induces local and systemic antitumor efficacy in canine mammary cancer patients. Cells 2023, 12, 2241.
- Chung YH, Ortega-Rivera OA, Volckaert BA, Jung E, Zhao Z, Steinmetz NF: Viral nanoparticle vaccines against S100A9 reduce lung tumor seeding and metastasis. Proceedings of the National Academy of Sciences 2023, 120, e2221859120. [CrossRef]
- Truchado Martín DA, Juárez-Molina M, Rincón S, Zurita L, Tomé-Amat J, Lorz López MC, Ponz F: A multifunctionalized potyvirus-derived nanoparticle that targets and internalizes into cancer cells. 2024.
- Jung E, Foroughishafiei A, Chung YH, Steinmetz NF: Enhanced Efficacy of a TLR3 Agonist Delivered by Cowpea Chlorotic Mottle Virus Nanoparticles. Small Science 2024:2300314.
- Moreno-Gonzalez MA, Zhao Z, Caparco AA, Steinmetz NF: Combination of cowpea mosaic virus (CPMV) intratumoral therapy and oxaliplatin chemotherapy. Materials Advances 2024.
- Ghani MA, Bangar A, Yang Y, Jung E, Sauceda C, Mandt T, Shukla S, Webster NJ, Steinmetz NF, Newton IG: Treatment of Hepatocellular Carcinoma by Multimodal In Situ Vaccination Using Cryoablation and a Plant Virus Immunostimulant. Journal of Vascular and Interventional Radiology 2023, 34, 1247–1257. [CrossRef] [PubMed]
- Zheng Z, Zhang L, Hou X: Potential roles and molecular mechanisms of phytochemicals against cancer. Food & Function 2022, 13, 9208–9225.
- Oluwayelu DO, Adebiyi AI: Plantibodies in human and animal health: a review. African health sciences 2016, 16, 640–645. [CrossRef] [PubMed]
- Sheshukova E, Komarova T, Dorokhov Y: Plant factories for the production of monoclonal antibodies. Biochemistry (Moscow) 2016, 81:1118-113.
- Acosta-Ramírez E, Pérez-Flores R, Majeau N, Pastelin-Palacios R, Gil-Cruz C, Ramírez-Saldaña M, Manjarrez-Orduño N, Cervantes-Barragán L, Santos-Argumedo L, Flores-Romo L: Translating innate response into long-lasting antibody response by the intrinsic antigen-adjuvant properties of papaya mosaic virus. Immunology 2008, 124, 186–197.
- Yusibov V, Mett V, Mett V, Davidson C, Musiychuk K, Gilliam S, Farese A, MacVittie T, Mann D: Peptide-based candidate vaccine against respiratory syncytial virus. Vaccine, 2261.
- Kemnade JO, Seethammagari M, Collinson-Pautz M, Kaur H, Spencer DM, McCormick AA: Tobacco mosaic virus efficiently targets DC uptake, activation and antigen-specific T cell responses in vivo. Vaccine 2014, 32, 4228–4233. [CrossRef]
- Brennan F, Jones T, Longstaff M, Chapman S, Bellaby T, Smith H, Xu F, Hamilton W, Flock J-I: Immunogenicity of peptides derived from a fibronectin-binding protein of S. aureus expressed on two different plant viruses. Vaccine 1999, 17(15-16):1846-1857.
- Brennan FR, Bellaby T, Helliwell SM, Jones TD, Kamstrup S, Dalsgaard K, Flock J-I, Hamilton WD: Chimeric plant virus particles administered nasally or orally induce systemic and mucosal immune responses in mice. Journal of virology 1999, 73, 930–938. [CrossRef]
- Steinmetz NF: Viral nanoparticles as platforms for next-generation therapeutics and imaging devices. Nanomedicine: Nanotechnology, Biology and Medicine 2010, 6, 634–641. [CrossRef]
- Young M, Debbie W, Uchida M, Douglas T: Plant viruses as biotemplates for materials and their use in nanotechnology. Annu Rev Phytopathol 2008, 46:361-384.
- Steinmetz NF, Evans DJ: Utilisation of plant viruses in bionanotechnology. Organic & biomolecular chemistry 2007, 5, 2891–2902.
- Venkataraman S, Hefferon K: Application of plant viruses in biotechnology, medicine, and human health. Viruses 2021, 13, 1697. [CrossRef]
- Kumar S, Ochoa W, Singh P, Hsu C, Schneemann A, Manchester M, Olson M, Reddy V: Tomato bushy stunt virus (TBSV), a versatile platform for polyvalent display of antigenic epitopes and vaccine design. Virology 2009, 388, 185–190. [CrossRef] [PubMed]
- Lebel M-È, Chartrand K, Leclerc D, Lamarre A: Plant viruses as nanoparticle-based vaccines and adjuvants. Vaccines 2015, 3, 620–637. [CrossRef]
- Balke I, Zeltins A: Recent advances in the use of plant virus-like particles as vaccines. Viruses 2020, 12, 270. [CrossRef]
- Lee PW, Shukla S, Wallat JD, Danda C, Steinmetz NF, Maia J, Pokorski JK: Biodegradable viral nanoparticle/polymer implants prepared via melt-processing. ACS nano 2017, 11, 8777–8789. [CrossRef] [PubMed]
- Affonso de Oliveira JF, Chan SK, Omole AO, Agrawal V, Steinmetz NF: In Vivo Fate of Cowpea Mosaic Virus In Situ Vaccine: Biodistribution and Clearance. ACS nano 2022, 16, 18315–18328. [CrossRef] [PubMed]
- Rohovie MJ, Nagasawa M, Swartz JR: Virus-like particles: Next-generation nanoparticles for targeted therapeutic delivery. Bioengineering & translational medicine 2017, 2, 43–57.
- Nooraei S, Bahrulolum H, Hoseini ZS, Katalani C, Hajizade A, Easton AJ, Ahmadian G: Virus-like particles: preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. Journal of nanobiotechnology 2021, 19:1-27.
- Li L, Xu C, Zhang W, Secundo F, Li C, Zhang Z-P, Zhang X-E, Li F: Cargo-compatible encapsulation in virus-based nanoparticles. Nano Letters 2019, 19, 2700–2706. [CrossRef]
- Rizzo LY, Theek B, Storm G, Kiessling F, Lammers T: Recent progress in nanomedicine: therapeutic, diagnostic and theranostic applications. Current opinion in biotechnology 2013, 24, 1159–1166. [CrossRef]
- Lo Y-L, Wang C-S, Chen Y-C, Wang T-Y, Chang Y-H, Chen C-J, Yang C-P: Mitochondrion-directed nanoparticles loaded with a natural compound and a microRNA for promoting cancer cell death via the modulation of tumor metabolism and mitochondrial dynamics. Pharmaceutics 2020, 12, 756. [CrossRef] [PubMed]
- Ahmadi F, Ghasemi-Kasman M, Ghasemi S, Gholamitabar Tabari M, Pourbagher R, Kazemi S, Alinejad-Mir A: Induction of apoptosis in hela cancer cells by an ultrasonic-mediated synthesis of curcumin-loaded chitosan–alginate–sTPP nanoparticles. International journal of nanomedicine 2017:8545-8556.
- Ren J, Barton CD, Sorenson KE, Zhan J: Identification of a novel glucuronyltransferase from Streptomyces chromofuscus ATCC 49982 for natural product glucuronidation. Applied Microbiology and Biotechnology 2022, 106, 1165–1183. [CrossRef] [PubMed]
- Ahmad M, Gani A: Development of novel functional snacks containing nano-encapsulated resveratrol with anti-diabetic, anti-obesity and antioxidant properties. Food Chemistry, 2021, 352:129323.
- Valdés R, Gómez L, Padilla S, Brito J, Reyes B, Alvarez T, Mendoza O, Herrera O, Ferro W, Pujol M: Large-scale purification of an antibody directed against hepatitis B surface antigen from transgenic tobacco plants. Biochemical and Biophysical Research Communications 2003, 308, 94–100. [CrossRef] [PubMed]
- Peyret H, Lomonossoff GP: When plant virology met Agrobacterium: the rise of the deconstructed clones. Plant biotechnology journal 2015, 13, 1121–1135. [CrossRef]
- Ibrahim A, Odon V, Kormelink R: Plant viruses in plant molecular pharming: toward the use of enveloped viruses. Frontiers in Plant Science 2019, 10:457269.
- Morgenfeld M, Lentz E, Segretin ME, Alfano EF, Bravo-Almonacid F: Translational fusion and redirection to thylakoid lumen as strategies to enhance accumulation of human papillomavirus E7 antigen in tobacco chloroplasts. Molecular biotechnology 2014, 56, 1021–1031. [CrossRef]
- Lico C, Giardullo P, Mancuso M, Benvenuto E, Santi L, Baschieri S: A biodistribution study of two differently shaped plant virus nanoparticles reveals new peculiar traits. Colloids and Surfaces B: Biointerfaces, 2016, 148:431-439.
- Bruckman MA, Randolph LN, VanMeter A, Hern S, Shoffstall AJ, Taurog RE, Steinmetz NF: Biodistribution, pharmacokinetics, and blood compatibility of native and PEGylated tobacco mosaic virus nano-rods and-spheres in mice. Virology 2014, 449:163-173.
- Balke I, Zeltins A: Use of plant viruses and virus-like particles for the creation of novel vaccines. Advanced drug delivery reviews.
- Gulati N, Pitek A, Czapar A, Stewart P, Steinmetz N: The in vivo fates of plant viral nanoparticles camouflaged using self-proteins: overcoming immune recognition. Journal of Materials Chemistry B 2018, 6, 2204–2216. [CrossRef]
- Pitek AS, Wen AM, Shukla S, Steinmetz NF: The protein corona of plant virus nanoparticles influences their dispersion properties, cellular interactions, and in vivo fates. Small 2016, 12, 1758–1769. [CrossRef]
- Gonzalez MJ, Plummer EM, Rae CS, Manchester M: Interaction of Cowpea mosaic virus (CPMV) nanoparticles with antigen presenting cells in vitro and in vivo. PloS one 2009, 4, e7981.
- Fausther-Bovendo H, Kobinger GP: Pre-existing immunity against Ad vectors: humoral, cellular, and innate response, what's important? Human vaccines & immunotherapeutics 2014, 10, 2875–2884.
- Hwang C, Sanda M: Prospects and limitations of recombinant poxviruses for prostate cancer immunotherapy. Current opinion in molecular therapeutics 1999, 1, 471–479.
- Khan T, Ali M, Khan A, Nisar P, Jan SA, Afridi S, Shinwari ZK: Anticancer plants: A review of the active phytochemicals, applications in animal models, and regulatory aspects. Biomolecules 2019, 10, 47.
- Apolone G, Joppi R, Bertele V, Garattini S: Ten years of marketing approvals of anticancer drugs in Europe: regulatory policy and guidance documents need to find a balance between different pressures. British Journal of Cancer 2005, 93, 504–509. [CrossRef] [PubMed]
- Lam M, Carmichael AR, Griffiths HR: An aqueous extract of Fagonia cretica induces DNA damage, cell cycle arrest and apoptosis in breast cancer cells via FOXO3a and p53 expression. PloS one 2012, 7, e40152.
- Farrell A, Papadouli I, Hori A, Harczy M, Harrison B, Asakura W, Marty M, Dagher R, Pazdur R: The advisory process for anticancer drug regulation: a global perspective. Annals of oncology 2006, 17, 889–896. [CrossRef] [PubMed]
- Calixto J: Efficacy, safety, quality control, marketing and regulatory guidelines for herbal medicines (phytotherapeutic agents). Brazilian Journal of medical and Biological research 2000, 33:179-189.
- Silva TCd, Silva JMd, Ramos MA: What factors guide the selection of medicinal plants in a local pharmacopoeia? A case study in a rural community from a historically transformed atlantic forest landscape. Evidence-Based Complementary and Alternative Medicine 2018, 2018, 2519212. [CrossRef]
- Khan T, Abbasi BH, Khan MA, Shinwari ZK: Differential effects of thidiazuron on production of anticancer phenolic compounds in callus cultures of Fagonia indica. Applied biochemistry and biotechnology 2016, 179:46-58.
- Khan T, Abbasi BH, Khan MA, Azeem M: Production of biomass and useful compounds through elicitation in adventitious root cultures of Fagonia indica. Industrial crops and products 2017, 108:451-457.
- Liskova A, Stefanicka P, Samec M, Smejkal K, Zubor P, Bielik T, Biskupska-Bodova K, Kwon TK, Danko J, Büsselberg D: Dietary phytochemicals as the potential protectors against carcinogenesis and their role in cancer chemoprevention. Clinical and experimental medicine 2020, 20:173-190.
- Cojocneanu Petric R, Braicu C, Raduly L, Zanoaga O, Dragos N, Monroig P, Dumitrascu D, Berindan-Neagoe I: Phytochemicals modulate carcinogenic signaling pathways in breast and hormone-related cancers. OncoTargets and therapy 2015:2053-2066.
- Kapinova A, Stefanicka P, Kubatka P, Zubor P, Uramova S, Kello M, Mojzis J, Blahutova D, Qaradakhi T, Zulli A: Are plant-based functional foods better choice against cancer than single phytochemicals? A critical review of current breast cancer research. Biomedicine & Pharmacotherapy 2017, 96:1465-1477.
- Bag A, Chattopadhyay RR: Evaluation of synergistic antibacterial and antioxidant efficacy of essential oils of spices and herbs in combination. PloS one 2015, 10, e0131321.
- Rescigno T, Tecce MF, Capasso A: Protective and restorative effects of nutrients and phytochemicals. The Open Biochemistry Journal 2018, 12:46.
- Al-Gubory KH, Blachier F, Faure P, Garrel C: Pomegranate peel extract decreases small intestine lipid peroxidation by enhancing activities of major antioxidant enzymes. Journal of the Science of Food and Agriculture 2016, 96, 3462–3468. [CrossRef]
- Chandel P, Rawal RK, Kaur R: Natural products and their derivatives as cyclooxygenase-2 inhibitors. Future Medicinal Chemistry 2018, 10, 2471–2492. [CrossRef]
- Kapinova A, Kubatka P, Liskova A, Baranenko D, Kruzliak P, Matta M, Büsselberg D, Malicherova B, Zulli A, Kwon TK: Controlling metastatic cancer: the role of phytochemicals in cell signaling. Journal of Cancer Research and Clinical Oncology, 2019; 145, 1087–1109.
- Liskova A, Kubatka P, Samec M, Zubor P, Mlyncek M, Bielik T, Samuel SM, Zulli A, Kwon TK, Büsselberg D: Dietary phytochemicals targeting cancer stem cells. Molecules 2019, 24, 899. [CrossRef]
- Kubatka P, Uramova S, Kello M, Kajo K, Samec M, Jasek K, Vybohova D, Liskova A, Mojzis J, Adamkov M: Anticancer activities of Thymus vulgaris L. in experimental breast carcinoma in vivo and in vitro. International journal of molecular sciences 2019, 20, 1749. [Google Scholar] [CrossRef]
- Uramova S, Kubatka P, Dankova Z, Kapinova A, Zolakova B, Samec M, Zubor P, Zulli A, Valentova V, Kwon TK: Plant natural modulators in breast cancer prevention: status quo and future perspectives reinforced by predictive, preventive, and personalized medical approach. EPMA Journal 2018, 9:403-419.
- Jasek K, Kubatka P, Samec M, Liskova A, Smejkal K, Vybohova D, Bugos O, Biskupska-Bodova K, Bielik T, Zubor P: DNA methylation status in cancer disease: modulations by plant-derived natural compounds and dietary interventions. Biomolecules 2019, 9, 289.
- Kubatka P, Uramova S, Kello M, Kajo K, Kruzliak P, Mojzis J, Vybohova D, Adamkov M, Jasek K, Lasabova Z: Antineoplastic effects of clove buds (Syzygium aromaticum L. ) in the model of breast carcinoma. Journal of Cellular and Molecular Medicine 2017, 21, 2837–2851. [Google Scholar]
- Kubatka P, Kello M, Kajo K, Kruzliak P, Výbohová D, Mojžiš J, Adamkov M, Fialová S, Veizerová L, Zulli A: Oregano demonstrates distinct tumour-suppressive effects in the breast carcinoma model. European journal of nutrition, 2017; 56, 1303–1316.
- Abotaleb M, Samuel SM, Varghese E, Varghese S, Kubatka P, Liskova A, Büsselberg D: Flavonoids in cancer and apoptosis. Cancers 2018, 11, 28. [CrossRef] [PubMed]
- Kaur V, Kumar M, Kumar A, Kaur K, Dhillon VS, Kaur S: Pharmacotherapeutic potential of phytochemicals: Implications in cancer chemoprevention and future perspectives. Biomedicine & Pharmacotherapy 2018, 97:564-586.
- Wang C-Y, Bai X-Y, Wang C-H: Traditional Chinese medicine: a treasured natural resource of anticancer drug research and development. The American journal of Chinese medicine 2014, 42, 543–559. [CrossRef]
- Shukla S, Hu H, Cai H, Chan S-K, Boone CE, Beiss V, Chariou PL, Steinmetz NF: Plant viruses and bacteriophage-based reagents for diagnosis and therapy. Annual review of virology 2020, 7:559-587.
- Gulati NM, Stewart PL, Steinmetz NF: Bioinspired shielding strategies for nanoparticle drug delivery applications. Molecular pharmaceutics 2018, 15, 2900–2909. [CrossRef]
- Shukla S: Serum albumin ‘camouflage’of plant virus based nanoparticles prevents their antibody recognition and enhances pharmacokinetics. Biomaterials 2016, 89:89-97.
- Shoeb E, Badar U, Venkataraman S, Hefferon K: Frontiers in bioengineering and biotechnology: plant nanoparticles for anti-cancer therapy. Vaccines 2021, 9, 830. [CrossRef]
- Hefferon K: Plant virus expression vectors: a powerhouse for global health. Biomedicines 2017, 5, 44.
Figure 1.
The chemical structures of some prominent natural alkaloids and their semisynthetic derivatives serve as effective agents in combating cancer. Adapted from an open-access source Dhyani et al., 2022 [48].
Figure 1.
The chemical structures of some prominent natural alkaloids and their semisynthetic derivatives serve as effective agents in combating cancer. Adapted from an open-access source Dhyani et al., 2022 [48].
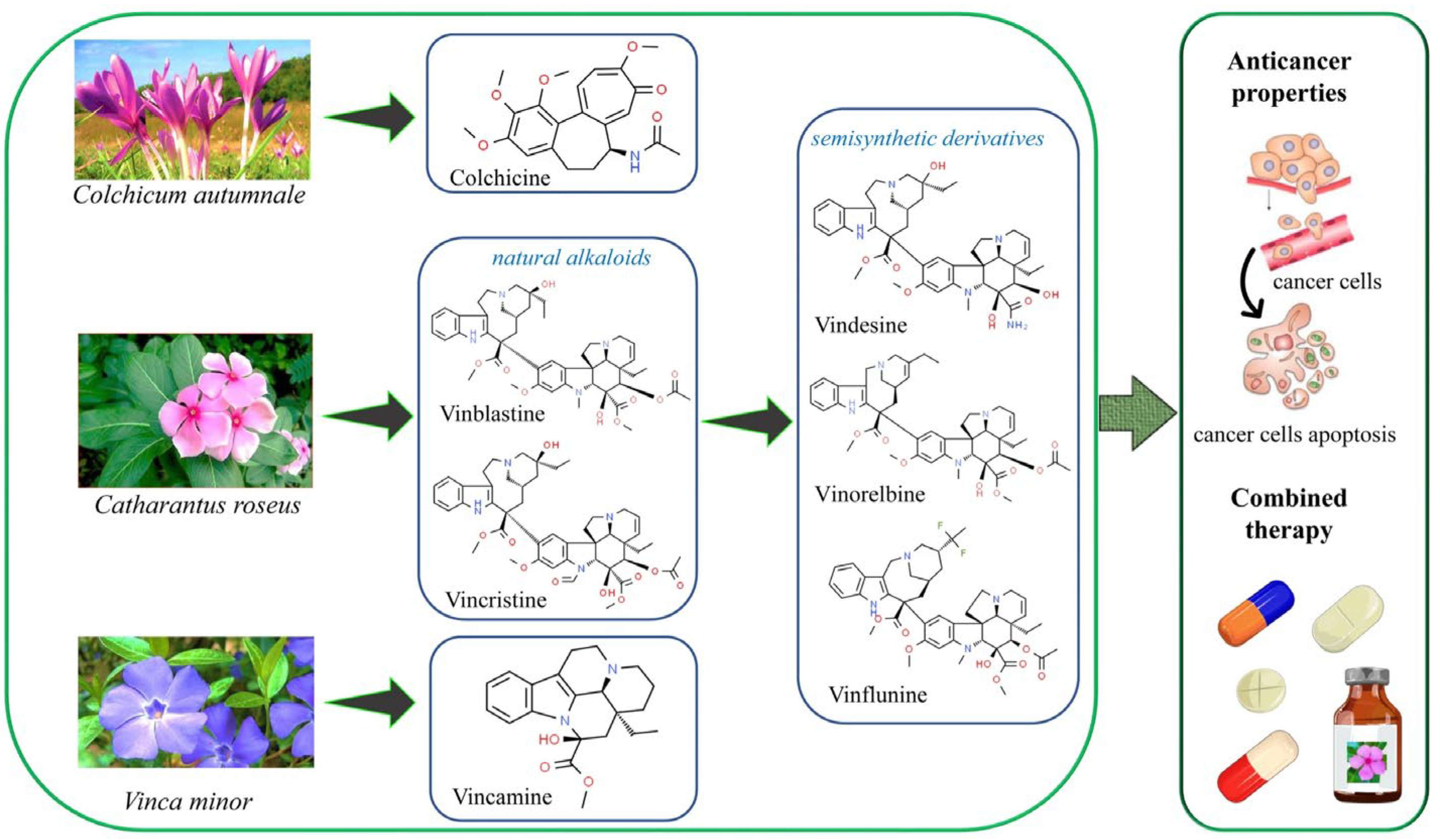
Figure 2.
The phototherapy mechanism of action. In photodynamic therapy (PDT), photosensitizers (PS) absorb light, transitioning to an excited state. This leads to two pathways: PDT Type I, where the PS reacts with biomolecules to create reactive oxygen species (ROS), and PDT Type II, where the PS transfers energy directly to oxygen, producing ROS. ROS exhibit high oxidizing power, causing cytotoxic effects, primarily near their generation due to their short lifespan. Adapted from an open access source Pivetta et al., 2021. [172].
Figure 2.
The phototherapy mechanism of action. In photodynamic therapy (PDT), photosensitizers (PS) absorb light, transitioning to an excited state. This leads to two pathways: PDT Type I, where the PS reacts with biomolecules to create reactive oxygen species (ROS), and PDT Type II, where the PS transfers energy directly to oxygen, producing ROS. ROS exhibit high oxidizing power, causing cytotoxic effects, primarily near their generation due to their short lifespan. Adapted from an open access source Pivetta et al., 2021. [172].
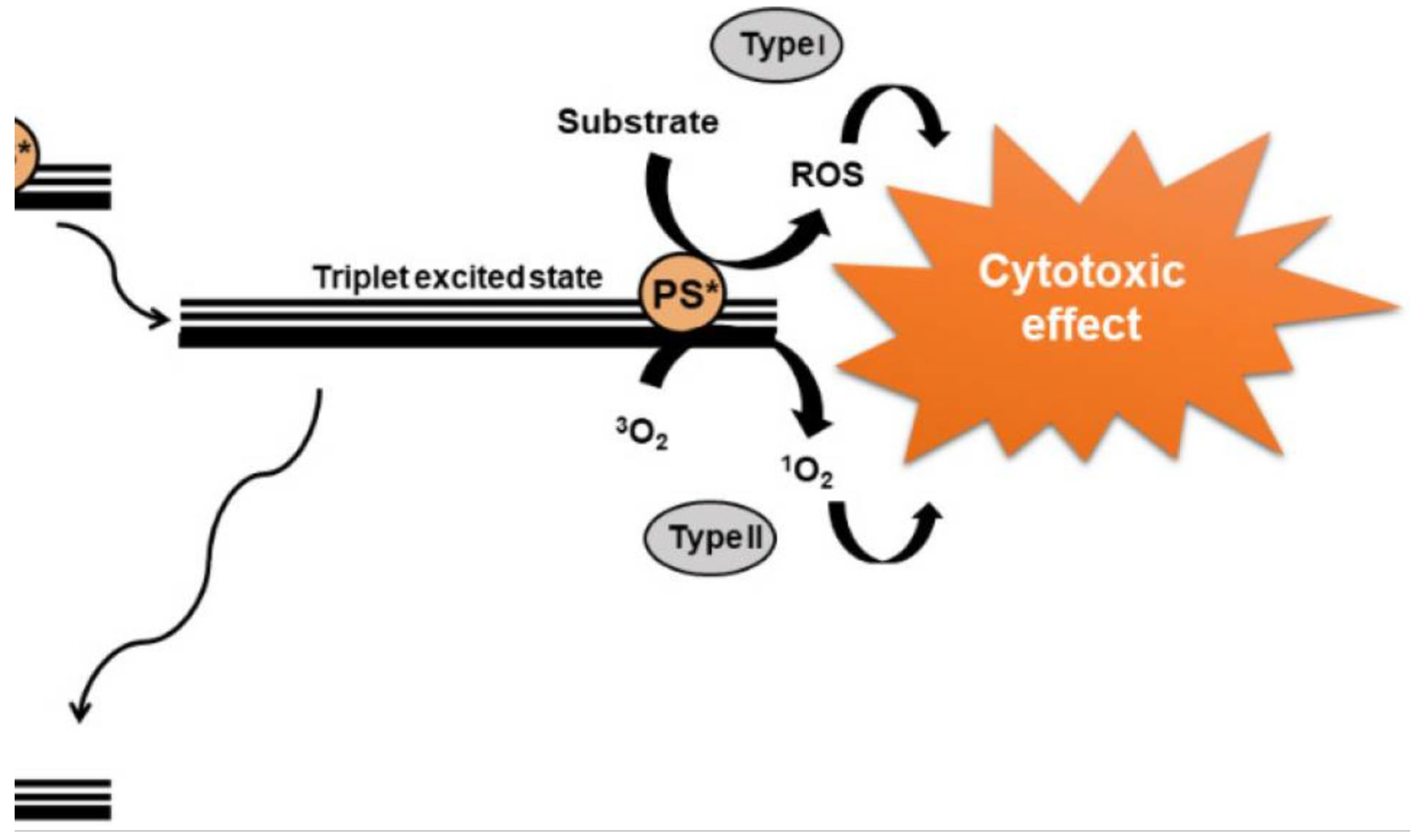
Figure 3.
PVNPs as delivery therapeutic and imaging agents in cancer. A) (A) Tobacco mosaic virus (TMV) for the targeted delivery of cisplatin in Pt-resistant ovarian cancer cells [272] (Permission obtained). B) The preparative process for potato virus X (PVX)-HisTRAIL by coordinating the bond between a Ni-nitrilotriacetic (NTA) group on the virus; the His-tag at the N-terminus of HisTRAIL is shown with a purple triangle. Multivalent display of HisTRAIL on the elongated PVX particle permits proper binding on death receptors DR4/5 (the trimers with blue color) for activating the caspase-dependent apoptosis in cancerous cells [273]. C) miR-181a is an important target for ovarian cancer therapy. qPCR data and cancer cell migration assays demonstrated higher knockdown efficacy when anti-miR-181a oligonucleotides were encapsulated and delivered using the VLPs resulting in reduced cancer cell invasiveness [274] [Citation credited and no permission needed].D) Schematic illustration of Gd-Cy5.5-PhMV-mPEG NPs for cancer imaging. In vivo NIR fluorescence images of PC-3 prostate tumors in athymic nude mice after the intravenous injection of Gd-Cy5.5-PhMV-DGEA [275] [Citation credited and no permission needed].
Figure 3.
PVNPs as delivery therapeutic and imaging agents in cancer. A) (A) Tobacco mosaic virus (TMV) for the targeted delivery of cisplatin in Pt-resistant ovarian cancer cells [272] (Permission obtained). B) The preparative process for potato virus X (PVX)-HisTRAIL by coordinating the bond between a Ni-nitrilotriacetic (NTA) group on the virus; the His-tag at the N-terminus of HisTRAIL is shown with a purple triangle. Multivalent display of HisTRAIL on the elongated PVX particle permits proper binding on death receptors DR4/5 (the trimers with blue color) for activating the caspase-dependent apoptosis in cancerous cells [273]. C) miR-181a is an important target for ovarian cancer therapy. qPCR data and cancer cell migration assays demonstrated higher knockdown efficacy when anti-miR-181a oligonucleotides were encapsulated and delivered using the VLPs resulting in reduced cancer cell invasiveness [274] [Citation credited and no permission needed].D) Schematic illustration of Gd-Cy5.5-PhMV-mPEG NPs for cancer imaging. In vivo NIR fluorescence images of PC-3 prostate tumors in athymic nude mice after the intravenous injection of Gd-Cy5.5-PhMV-DGEA [275] [Citation credited and no permission needed].
Figure 4.
PVNPs in cancer immune and combinational therapy A) Intratumoral administration of plant-derived Cowpea mosaic virus (CPMV) nanoparticles as an in situ vaccine overcomes the local immunosuppression and stimulates a potent anti-tumor response in several mouse cancer models and canine patients [309].B) The PhMV-based anti-HER2 vaccine PhMV-CH401, demonstrated efficacy as an anti-HER2 cancer vaccine. Our studies highlight that VLPs derived from PhMV are a promising platform to develop cancer vaccines [310].C) Schematic diagram of preparing CCMV VLPs containing ODN 1826 (CCMV-ODN1826) for cancer therapy [275]. D) Photothermal immunotherapy of melanoma using TLR-7 agonist laden TMV with polydopamine coat [285]. (citations credited and no permission needed).
Figure 4.
PVNPs in cancer immune and combinational therapy A) Intratumoral administration of plant-derived Cowpea mosaic virus (CPMV) nanoparticles as an in situ vaccine overcomes the local immunosuppression and stimulates a potent anti-tumor response in several mouse cancer models and canine patients [309].B) The PhMV-based anti-HER2 vaccine PhMV-CH401, demonstrated efficacy as an anti-HER2 cancer vaccine. Our studies highlight that VLPs derived from PhMV are a promising platform to develop cancer vaccines [310].C) Schematic diagram of preparing CCMV VLPs containing ODN 1826 (CCMV-ODN1826) for cancer therapy [275]. D) Photothermal immunotherapy of melanoma using TLR-7 agonist laden TMV with polydopamine coat [285]. (citations credited and no permission needed).

Table 1.
Clinical trials of phytochemicals against cancer.
| Study | Drugs involved | Conditions / Effects | Status | Identifier | References |
|---|---|---|---|---|---|
| Therapeutic effect of luteolin natural extract versus its nanoparticles on tongue squamous cell carcinoma cell line | Luteolin Nano-luteolin |
Tongue neoplasms Carcinoma |
Unknown | NCT03288298 | [162] |
| Artemisinin derivative SM934 inhibits expression of cathepsin K after forming a complex with testosterone | SM934 (a novel water-soluble artemisinin analog) | Inhibits proliferation and metastasis in breast cancer |
Phase II | NA |
[163,164] |
| Study of Liposomal Curcumin in combination with RT and TMZ in patients with newly diagnosed high-grade gliomas |
Curcumin combined with radiotherapy (RT) and Temozolomide (TMZ) | Glioblastoma | Phase I / Phase II | NCT05768919 | [165] |
| Curcumin Bioavailability in Glioblastoma Patients | Curcumin | Glioblastoma | Unknown | NCT01712542 | [166] |
| Phase I Assay-guided Trial of Anti-inflammatory Phytochemicals in Patients With Advanced Cancer |
Grape seed extract and Vitamin D | Solid cancers (gastrointestinal, lung, breast, prostate, lymphoma or cancer of the lymph nodes) | Phase 1 Completed |
NCT01820299 | [167] |
| Dietary Intervention With Phytochemicals and Polyunsaturated Fatty Acids in Prostate Cancer Patients | Tomato or a multi-diet consisting of grape juice, pomegranate juice, tomato, green tea, black tea, soy, selenium and PUFAs |
Prostate cancer | Phase 1 and Phase 2 Completed | NCT00433797 | [168] |
| Clinical Trial of Lung Cancer Chemoprevention With Sulforaphane in Former Smokers | Sulforaphane | Lung cancer | Phase 2 Completed | NCT03232138 | [169] |
| Black Raspberry Confection in Preventing Oral Cancer in Healthy Volunteers | Black raspberry confection | Oral cancer | Phase 1, Active, not recruiting | NCT01961869 | [169] |
| Docetaxel With a Phytochemical in Treating Patients With Hormone Independent Metastatic Prostate Cancer (PROTAXY) | Phytochemical dietary supplement with docetaxel | Prostate cancer | Phase 2, Completed | NCT01012141 | [169] |
| Tangerine or Red Tomato Juice in Treating Patients With Prostate Cancer Undergoing Surgery | Tangerine tomato juice or red tomato juice rich in lycopene | Prostate cancer | Not applicable | NCT02144649 | [169] |
Table 2.
Recent reports of anticancer mAb expression in plants.
| Type of cancer | Plant system used | Effects | Reference |
|---|---|---|---|
| Colorectal cancer | Transgenic tobacco expressing large single chain (LSC) antibody CO17-1A (LSC CO) and LSC CO tagged with the endoplasmic reticulum (ER) retention signal KDEL (LSC COK) | In vitro binding activity towards human colon cancer cell lines | [233] |
| Breast cancer | Transgenic tobacco expressing anti-HER2 VHH-FcK mAb | Bound to cancer cells in vitro and inhibited cell migration | [234] |
| Colorectal cancer and breast cancer | Transgenic tobacco expressing both mAbs LSC CO17-1AK and anti-HER2 VHH-FcK in the same plant | Demonstrated binding to human SW620 and SKBR-3 cancer cells and inhibition of cell migration in vitro |
[235] |
| Mouse colorectal cancer | Transient expression of recombinant bispecific monoclonal antibody for dual inhibition of programmed cell death protein 1/ programmed cell death ligand 1 and cytotoxic T-lymphocyteassociated protein 4 axes in Nicotiana benthamiana | Significant inhibition of tumor growth in vivo and reduction of tumor weight and volume | [236] |
| Murine colon cancer | Transient expression of anti-CTLA-4 2C8 mAb in N. benthamiana by agroinfiltration | Recognition and binding to both human and murine CTLA-4 in vitro as well as inhibition of in vivo tumor growth | [237] |
| Mouse colorectal tumor | Atezolizumab anti-PD-L1 antibody transiently produced in N. benthamiana | Mouse tumor growth inhibition and in vitro binding to PD-L1 | [238] |
| Mouse MC38 colon cancer | Recombinant anti-PD-1 Nivolumab was produced in Nicotiana benthamiana by transgenic technology | Reduced in vivo mouse tumor growth | [239] |
| Gastric and colorectal cancer | Transient expression of Durvalumab variants in Nicotiana benthamiana | Recognition and binding to recombinant PD-L1 and to PD-L1 expressed in gastrointestinal cancer cells, precluding its interaction with PD-1 on T cells thereby augmenting T-cell immunity | [240] |
| Breast cancer | Trastuzumab transgenically expressed in glycoengineered rice | Inhibition of BT-474 cancer cell line proliferation, increased ADCC efficacy against Jurkat cells, efficacious tumour uptake with lower liver uptake compared to TMab in a xenograft assay using the BT-474 murine model. | [241] |
| Hodgkin lymphoma, melanoma, lung colorectal and breast and cancer | Transient expression of pembrolizumab and nivolumab in Nicotiana benthamiana | PD-1/PD-L1 inhibitory activity in vitro; both immune checkpoint inhibitors (ICIs) inhibit the PD-1/PD-L1 immune checkpoint leading to CTL activation and the elicitation of apoptosis in tumorigenic cells via T-cell-mediated cytotoxicity | [242] |
| CD27- expressing lymphoma and leukemia, recurrent glioblastoma, advanced solid tumors | Transient generation of Varlilumab (anti-human CD27) in N. benthamiana | Co-expression with chimeric beta 1,4-GALT (beta 1,4- galactosyltransferase) successfully achieved biantennary b1,4- galactosylated Varlilumab | [243] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



