Submitted:
06 November 2024
Posted:
07 November 2024
You are already at the latest version
Abstract
Currently, therapeutic Fc-fusion protein is used for treating disease. ESAT6 (6-kDa early secretory antigenic target), secreted by Mycobacterium tuberculosis, is involved in immune regulation. Administered via intranasal immunization, ESAT6-Fc fusion protein significantly alleviated allergic airway inflammation and mucus hypersecretion, reduced the proportions of Th2 cells, Th17 cells and eosinophils with no observable histopathological injury to principal organs in ovalbumin (OVA)-induced allergic asthma (AA) model mice. A analysis of the transcriptome was conducted to explore its mechanisms of immune regulation, which confirmed that ESAT6 exerts an anti-AA effect by mainly suppressing some signaling pathways activation including Natural Killer Cell-Mediated Cytotoxicity, T Cell Receptor, Th1 and Th2 Cell Differentiation, Th17 Cell Differentiation, and Chemokine, which all belong to immune system in organismal systems. Meanwhile, The RT-qPCR validation results of Differently Expressed genes (DEGs) including Cd28, Icos, Cd48, Cd247, Cd40l, Itgal, Itgb2 and Ccl5 from the aforementioned pathways were consistent with the RNA-seq results. This study revealed the fundamental mechanisms of immune regulation underlying the effects of ESAT6 on OVA-induced allergic asthma. More importantly, this study provides valuable insights into the application of mucosal immunotherapy in treating AA, and ESAT6-Fc has potential as a secure mucosal immunotherapy agent for AA.
Keywords:
Allergic Asthma (AA)
; Mycobacterium tuberculosis (Mt.b)
; ESAT6
1. Introduction
Allergic asthma (AA) is characterized by high level of IgE, type 2 cytokines such as interleukin-4 (IL-4) and IL-13, Th17 cytokines such as interleukin (IL)-17A-F, IL-21 and IL-22, mucus hypersecretion, eosinophilia and airway hyperresponsiveness (AHR) [1,2].Dendritic cells (DCs),which represent the most potent antigen-presenting cells of the immune system, act as a bridge between innate and adaptive immunity [3].When stimulated by allergen, DCs induce the differentiation of naive T cells into Th2 and Th17cells [4].Th2 cytokines such as interleukin (IL)-4, and IL-13, facilitate recruitment and activation of eosinophils in the airway [5]. Th17 cytokines such as interleukin (IL)-17A-F, IL-21 and IL-22 induce mucous cell metaplasia and have pleotropic effects on airway smooth muscle resulting in airway narrowing [2,6]. Meanwhile, Th2 cells induce the B cells to product IgE, which binding to type I high-affinity IgE receptors (FcεRI) on mast cells and basophils, sensitizing them to get triggered when encountering with the same allergen [7], resulting in releasing of a large variety of inflammatory mediators, such as histamine and leukotrienes causing AHR [8,9].
In recent decades, the incidence of allergic diseases has been increasing in industrialized countries [10], and effective treatment may require regulation of the allergic inflammatory responses [11]. "Hygiene Hypothesis", which has evolved into the "Microbiota Hypothesis" demonstrated that microbial exposures could alter the risk of subsequent allergic disease and asthma [12]. Epidemiological studies have shown that the prevalence and severity of AA are negatively correlated with BCG vaccination, and the mechanism of Mycobacterium tuberculosis (Mt.b) components inhibiting allergic diseases remain unclear [13,14]. Mt.b ESAT6 (early secreted antigenic target of 6 kDa) secreted by Mt.b, is one of the major factors which regulating host immune responses [15,16]. The effect of ESAT6 on AA has been reported [17], however, the immune regulatory mechanisms underlying the ability of ESAT6 to alleviate the progression of AA remain unclear and need to clarify.
The role of IgGs is distinguished by their high circulating levels, long half-life and ability to transfer from mother to offspring attributing to interactions with neonatal Fc receptor (FcRn), which binding to the Fc portion of IgGs, allowing IgGs transport across polarized cells such as the endothelium and epithelium [18,19]. Based on the above principle, therapeutic Fc-fusion protein which composed of Fc region of IgG and a desired linked protein, was introduced for the treatment of diseases [20,21,22].
High-throughput transcriptome sequencing (RNA-Seq) is a novel technique that can discover novel RNAs, and identify differentially expressed genes (DEGs) under different conditions, which contribute to our understanding of disease-related changes [23]; and transcriptome analysis plays an essential role in identifying genetic networks underlying cellular, physiological and biochemical systems and establishing molecular biomarkers that respond to disease [24].
In this study, the eukaryotic expression plasmid pcDNA3.1 (+) Rv1875-Fc was constructed, and high-purity ESAT6-Fc fusion protein was obtained from Chinese hamster ovary cells (CHO) which is the predominant host cell line for the production of biopharmaceuticals [25,26].The aim of study is to investigate the effect of ESAT6-Fc fusion protein on OVA-induced AA by nasal mucosal immunization, and is followed by transcriptomics analysis to further explore the potential regulatory mechanism of ESAT6 on AA.
2. Results
2.1. Effects of ESAT6-Fc Fusion Protein on Allergic Asthma Model Mice
To assess whether ESAT6-Fc fusion protein alleviate AA. Firstly, the ESAT6-Fc fusion protein was expressed and purified (Supplementary Figure S1). After ESAT6-Fc fusion protein were administrated intranasally to M group mice, its ability to alleviate AA was evaluated. In the M group mice (1% OVA), the percentages of Th2, Th17, and eosinophils are significantly higher compared to the N group) (Figure 1A; P < 0.0001 for Th2, Th17 and eosinophils), indicating that 1% OVA activated immune signaling pathways to mimic AA. Conversely, the percentages of cells were lower in the T group mice (P < 0.05, P < 0.001, and P < 0.001, respectively), indicating that the ESAT6 protein likely suppresses activation of some immune signaling pathways in the lung. To assess whether ESAT6 protein modulate molecules expression involved in AA, levels of IL-4, IL-13, IL-17A and IgE in BALF were tested by ELISA. The results showed that levels of IL-4, IL-13, IL-17A, and IgE are significantly higher in BALF in M group mice compared with N group mice (P < 0.0001, P < 0.001, P < 0.0001 and P < 0.001, respectively). However, in T group, levels of IL-4, IL-13, IL-17A, and IgE are lower compared with M group(P < 0.001, P < 0.01, P < 0.01 and P < 0.05, respectively). In addition, histopathological analysis showed remarkable leukocyte infiltration in the peribronchial region of lung tissue of M group (P < 0.01). Meanwhile, the infiltration level of leukocytes was marked lower in T group mice compared to the M group (P < 0.01) (Figure 1C). The number of PAS-positive cells was substantially higher in the M group compared to the N group (P < 0.01), indicating higher mucus production. In the T group, the number of PAS-positive cells was significantly lower compared to the M group (P < 0.05), suggesting that ESAT6 suppressed mucus production (Figure 1D). HE staining had no observable histopathological changes in the heart, liver, spleen, and kidney samples of mice in the T group compared to the N group, indicating the safety profile of ESAT6-Fc (Figure 1E).
Figure 1.
Timelines for OVA-induced AA mice model and ESAT6-Fc treatment
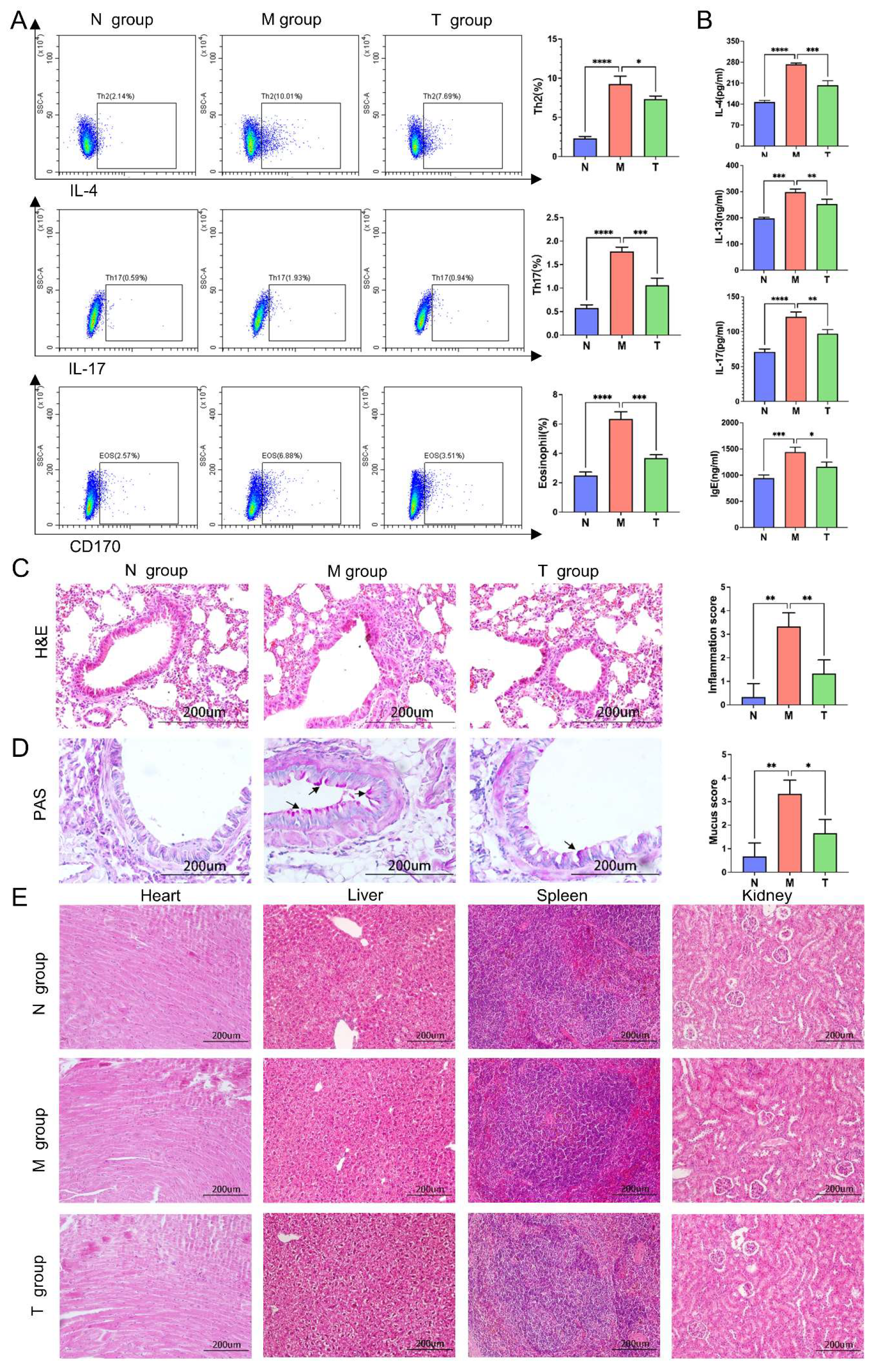
Figure 2.
Effects of ESAT6-Fc fusion protein on AA by intranasal immunization and security assessment. The percentage of Th2 cells, Th17 cells and eosinophils(A); Levels of IL-4 , IL-13, IL-17 and IgE in BALF (B); Effects of ESAT6 on airway inflammation in the peribronchiole region (H&E staining) (C); Goblet cell hyperplasia following ESAT6 treatment (PAS staining) (D). Data was expressed as mean ± standard deviation (SD) (n=4 per group). P < 0.05 means significant difference.
Figure 2.
Effects of ESAT6-Fc fusion protein on AA by intranasal immunization and security assessment. The percentage of Th2 cells, Th17 cells and eosinophils(A); Levels of IL-4 , IL-13, IL-17 and IgE in BALF (B); Effects of ESAT6 on airway inflammation in the peribronchiole region (H&E staining) (C); Goblet cell hyperplasia following ESAT6 treatment (PAS staining) (D). Data was expressed as mean ± standard deviation (SD) (n=4 per group). P < 0.05 means significant difference.
2.2. Transcriptomic Results
2.2.1. RNA-Seq Results and Quality Control
The statistical data for each sample RNA sequencing (RNA-seq) reads are shown in Table 2. The nine libraries yielded a total of 416.86M raw reads from Illumina sequencing, with an average of 46.32M reads per sample. Following quality-control filtering, the nine libraries yielded a total of 412.17M clean reads, with an average of 45.8M clean reads per sample. The average quality scores Q30 of the clean reads were 97.07%, indicating that the clean reads were high quality. Furthermore, the clean reads of the sample produced a high average content of 98.88 %.

Table 1.
Primers for the RT-qPCR.
| Gene | Reference sequence | Forward primer (F) | Reverse primer (R) |
|---|---|---|---|
| CD28 | NM_007642.4 | AACCAGAGAAGGCCAAGATTA | TATGTGTCAAGAGGCTGACT |
| Icos | NM_017480.3 | ATGGTGTTCTCTCTCTTCAGAT | CTCACACAGAAAGGACCG |
| CD48 | NM_007649.5 | TTCATCCCTAGCAGTGTTCC | GCAGACGTTCAGTAACACATT |
| CD247 | NM_031162.4 | TTCACCTGCTGATGTCACTT | CTCGTCATGAAATGGTGGC |
| Cd40lg | NM_011616.3 | AGGCACATAGAGCTGGAATA | GGGTTGCTGTTTCAGATTGTA |
| Itgal | NM_001253872.1 | GGAGAACTCCACTCTCTATATCA | TGTTGTGGTCATAGGCAGAT |
| Itgb2 | NM_008404.5 | TGGTAGGTGTCGTACTGATT | TCCTTCTCAAAGCGCCTGTA |
| CCL5 | NM_013653.3 | AACTATTTGGAGATGAGCTAGG | GGACTAGAGCAAGCAATGAC |
| β-Actin | NM_007393.5 | GGCTCCTAGCACCATGAAGA | AGCTCAGTAACAGTCCGCC |
Table 2.
Information of RawData (FASTQ) about RNA-Seq.
| Sample | Raw Reads (M) |
Raw Bases (G) |
Clean Reads (M) |
Clean Bases (G) |
Valid Bases (%) |
Q30 (%) |
|---|---|---|---|---|---|---|
| N1 | 41.49 | 6.14 | 40.87 | 6.05 | 98.5 | 97.15 |
| N2 | 44.00 | 6.54 | 43.54 | 6.47 | 98.94 | 97.2 |
| N3 | 48.09 | 7.13 | 47.46 | 7.03 | 98.68 | 97.1 |
| M1 | 47.62 | 7.06 | 47.03 | 6.97 | 98.76 | 97.17 |
| M2 | 47.10 | 7.00 | 46.61 | 6.92 | 98.95 | 97.04 |
| M3 | 46.40 | 6.88 | 45.85 | 6.8 | 98.83 | 97.05 |
| T1 | 47.45 | 7.06 | 47.03 | 7.00 | 99.12 | 97.12 |
| T2 | 47.66 | 7.09 | 47.23 | 7.03 | 99.11 | 96.95 |
| T3 | 47.04 | 6.99 | 46.58 | 6.92 | 99.02 | 96.85 |
1M=1million; 1G=109.
2.2.2. Principal Component Analysis (PCA) and Differentially Expressed Genes (DEGs)
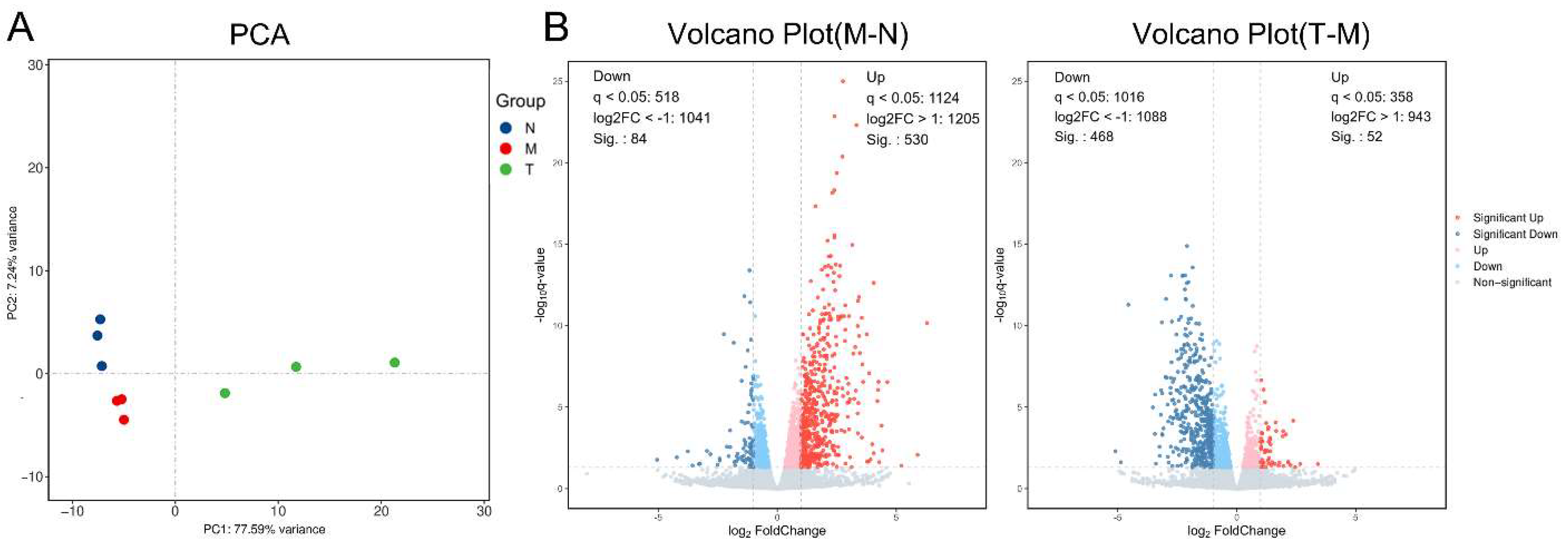
PCA was performed based on mRNA sequencing results, showing that the N, M and T groups could be distinguished. The N, M and T groups showed intra-group clustering and could be clearly separated during PCA, indicating mRNA of each group had good repeatability (Figure 2A); A total of 614 genes in the M group were differentially expressed genes (DEGs) with 530 upregulated and 84 downregulated genes comparing with the N group; and 520 genes in the T group were DEGs with 52 upregulated and 468 downregulated genes comparing with the M group (Figure 2B).
2.2.3. Pathway Enrichment Analysis of the DEGs Between the M and N Groups
To confirm the successful establishment of the AA model, the KEGG enrichment analysis of DEGs was performed. The KEGG pathway Classification was divided into five categories: organismal systems, metabolism, human disease, genetic information processing, environmental information processing and cellular processes. Most of the DEGs are involved in organismal systems, human disease and environmental information processing. In organismal systems, the immune system exhibits the highest number of DEGs (Figure 3A). Top20 KEGG enrichment analysis displayed on Figure 3B. DEGs Mainly enrich in the Cell cycle, Natural killer cell mediated cytotoxicity, T cell receptor signaling pathway, Th1 and Th2 cell differentiation, Th17 cell differentiation, Cell adhesion molecules, Cytokine-cytokine receptor interaction, and Chemokine signaling pathways. These signaling pathways are related to the initiation and progression of allergic inflammatory responses [31,32,33], indicating the successful generation of the AA model.
2.2.4. KEGG Enrichment Analysis of Pathways and Genes Regulated by ESAT6
To gain further insights into the biological functions of DEGs regulated by ESAT6, a KEGG enrichment analysis was conducted. A total of 231 different pathways were significantly enriched due to treatment with ESAT6-Fc. From them, top 20 pathways displayed on Figure 4, DEGs are mainly involved in human diseases, organismal systems, cellular processes, and environmental information processing. In organismal systems, DEGs are predominantly enriched in Natural Killer Cell-Mediated Cytotoxicity (mmu04650), which giving rise to a complex NK/dendritic cell (DC) cross-talk to help T(H)1 responses [34]; T Cell Receptor Signaling Pathway (mmu04660),critical for allergic airway disease, is triggered by signals of the T-cell receptor (TCR) and the cytokines generated during polarization [35]; Th1 and Th2 Cell Differentiation (mmu04658), which induce many of the hallmark features of asthma including airways hyperreactivity, eosinophilic by produce cytokines [36];Th17 Cell Differentiation (mmu4659), which plays an important role in the pathogenesis of AA and requires Notch signaling [37]. In Cellular Processes category, DEGs are primarily enriched in the Cell Cycle (mmu4110), which requires the cell adhesion to the extracellular matrix (ECM) via integrin receptors triggers signaling pathways [38]. In the Environmental Information Processing, DEGs are chiefly concentrated in Cell Adhesion Molecules (mmu04514), which is important for stabilizing the immunological synapse and inducing T-cell activation and proliferation [39]; and Cytokine-Receptor Interactions (mmu04060), in which cytokines regulate tissue regeneration and allow communication between various cell types by binding to cognate receptors [40].
To elucidate the immune regulation mechanism which the ESAT6 protein modulates AA in immune system of organismal systems, key regulatory factors involved in such signaling pathways as Natural Killer Cell-Mediated Cytotoxicity, T Cell Receptor Signaling Pathway, Th1 and Th2 Cell Differentiation, Th17 Cell Differentiation and Chemokine were identified and listed in Table 3.
2.4.5. Validation of RNA-Seq Results by RT-qPCR
Eight ESAT6 down-regulated DEGs including Cd28, Icos, Cd48, Cd247, Cd40l, Itgal, Itgb2 and Ccl5 from immune system were selected to validate the results of the RNA-seq. The results of RT-qPCR were in line with the results of transcriptome analysis (Figure 5).
3. Discussion
Asthma affects 3% of the global population, leading to over 0.25 million deaths annually [41]. As the main phenotype of asthma, Allergic asthma (AA) impose heavy economic and productivity burdens [42]. The conventional treatment modalities in AA are associated with side effects [43,44], emphasizing the need for alternative therapeutic agents. ESAT6 identified as a potent T-cell antigen, is secreted through the ESX-1 secretion system (Type VII) of M. tuberculosis [45]. The respiratory mucosal system plays an important role in the pathogenesis of AA, and the respiratory epithelium and mucosa provide a physical, functional, and immunologic barrier to protect the host from the inhaled environmental particles [46,47]. In recent years, respiratory mucosal immunotherapy exerts an important role in the treatment of asthma and other diseases [48,49].
Herein, the ESAT6-Fc fusion protein was intranasally administered and effectively inhibited the aggregation of Th2 cells, Th17 cells, and eosinophils in the lung. Besides, the ESAT6-Fc fusion protein inhibited the proliferation of bronchial goblet cells and concomitantly down-regulated the levels of IL-4, IL-13, IL-17, and IgE in the BALF of OVA-induced AA model mice. To examine the underlying mechanisms of immune regulation of the ESAT6 protein against AA, the signaling pathways of DEGs were analyzed using KEGG. Compared with N group, the up-regulated DEGs in M group are predominantly enriched in signaling pathways of Natural Killer Cell-Mediated Cytotoxicity, T Cell Receptor Signaling Pathway, Th1 and Th2 Cell Differentiation, Th17 Cell Differentiation, Chemokine, and Hematopoietic cell lineage in organismal systems, indicating that these signaling pathways have been activated by OVA.
T cells are crucial for immune functions to maintain health and prevent disease [50]. Natural Killer T (NKT) cells possess remarkable ability to modulate the immune response through the rapid secretion of various cytokines, and represent a promising target against AA [51]; CD4 T cells play critical roles in mediating adaptive immunity involved asthma and allergic responses. During TCR activation, naive CD4 T cells differentiate into one of several lineages of T helper (Th) cells, including Th1, Th2, and Th17 [52]. Immune cell trafficking is orchestrated by chemokine/chemokine receptor system in the allergic diseases [53].
After treated with ESAT6, in the organismal systems, the down-regulated DEGs are mainly enriched in Natural Killer Cell-Mediated Cytotoxicity, T Cell Receptor, Th1 and Th2 Cell Differentiation,Th17 Cell Differentiation, Chemokine, and Hematopoietic cell lineage signaling pathways compared with M group. These findings suggest that ESAT6 exerts an anti-AA effect by suppressing the activation of these signaling pathways. From the above signaling pathways, eight ESAT6 down-regulated DEGs including Cd28, Icos, Cd48, Cd247, Cd40l, Itgal, Itgb2 and Ccl5 were randomly selected for validation.
As well known that following T-cell receptor (TCR) activation, many co-receptors can enhance or suppress the TCR signal, and therapeutically targeting the co-receptors has been proven effective, and the B7-CD28 family is comprised of such immune-regulatory receptors [54,55];Inducible T-cell costimulator (ICOS) belongs to the CD28 family of cosignaling molecules, which binds to ICOS ligand (ICOSL) [56], and the ICOS signaling pathway plays main roles in coordinating the response of allergic reactions [57]; CD48, a costimulatory receptor and an effector molecule in asthma, has array of biological functions, such as adhesion, cellular activation and cytokine regulation, with potential as a target for therapy of allergic disease [58,59].CD247,also known as TCRZ, is expressed primarily in natural killer (NK) and T cells [60], and is relevant to allergic disease [61]. CD40 and CD40L are widely expressed in various types of cells, among which B cells and myeloid cells express high levels of CD40, and T cells and platelets express high levels of CD40L which acting as direct participants in asthma-related signaling pathways [62,63]. ITGAL (CD11a, gene name Itgal) and ITGB2(CD18, gene name Itgb2), the two subunits of LFA-1(integrin αLβ2, CD11a/CD18), is a member of the β2 integrin family localized on the surface of leukocytes [64]. T cells only express CD11a/CD18 (LFA-1) Of all β2-integrins [65], which playing key roles in the pathological progression of AA [66]. Chemokines play important roles in inflammation and in immune responses [67], and CCL5 is known to play an important role in the pathogenesis of AA [68]. Herein, these eight genes encode proteins are central to allergic and asthmatic responses.
When the ESAT6-Fc fusion protein infiltrates the respiratory epithelium of AA mice trafficked by FcRn, it is absorbed as a foreign macromolecule via pinocytosis or receptor-mediated endocytosis by phagocytes, especially dendritic cells (DC), neutrophils, and macrophages located in the respiratory submucosa. Simultaneously, antigens (ESAT6) are processed and presented by DC to initiate a Th1-driven adaptive immune response [69]. Therefore, the proportion of Th2 cells, Th17 cells and eosinophils in the lung, as well as the levels of IL-4, IL-13, IL-17A and IgE in the BALF are significantly decreased.
Overall, ESAT6 exerts an anti-AA effect by suppressing the OVA-induced signaling pathways activation of Natural Killer Cell-Mediated Cytotoxicity, T Cell Receptor, Th1 and Th2 Cell Differentiation, Th17 Cell Differentiation, and chemokine in immune system of organismal systems, which were confirmed by RT-qPCR.
4. Materials and Methods
4.1. Construction of Eukaryotic Plasmid and ESAT6-Fc Fusion Protein
To obtain ESAT6-Fc fusion protein, the pcDNA3.1(+)-Rv3875-Fc was constructed. Briefly 1116 bp nucleotide sequences encoding HSV2⁃gD signal peptide, M.tb ESAT6 and mouse IgG2a Fc were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China), and subsequently inserted between the polyclonal sites of HindIII and XhoI of pcDNA3.1 (+). Following this, pcDNA3.1-Rv1875-Fc was confirmed by HindⅢ/ XhoI (NEB, UK) enzyme digestion and sequencing (Beijing Ruibo Xingke Biotechnology Co., Ltd). Lastly, ESAT6-Fc fusion protein was produced and purified by Nanjing Mingyan Biotechnology Co., Ltd (Nanjin, China).
4.2. Animals and Grouping
A total of 18 female 7-week-old C57BL/6J (weight,18±1.0 g) mice were purchased from the Experimental Animal Center of Ningxia Medical University (certificate no. SCXK, 2020-0001).The mice were randomised into three groups (n=6/group): (1) normal group (N group); (2) AA modle group (M group), sensitised and challenged with 1% OVA; (3) ESAT6-Fc treatment group (T group), sensitised and challenged with 1% OVA, treated with ESAT6-Fc by intranasal immunization (Figure 1).
4.3. Allergic Asthma Mice Model
To establish the AA mouse model, Briefly, mice were intraperitoneally administered 1% ovalbumin (OVA, Solarbio, Beijing, China) three times over 14 days [27]. They were allowed ad libitum access to food for 14 days. Next, mice were nebulized with 1% OVA once daily for 20 minutes [27]. The timeline for AA model displayed in Figure 1. All procedures were approved by the Ethics Committee of Ningxia Medical University (approval no. 2024-G221; approval no.2024-N0023).
4.4. Ag85B-Fc Fusion Protein Treatment
To assess the effect of ESAT6-Fc on the symptoms of AA, the ESAT6-Fc fusion protein and a CpG adjuvant were intranasally administered to the AA model mice. Briefly, 14 days after intraperitoneal injection with 1% OVA, mice were intranasally administered 20 µL of a mixture composed of 10 µg Ag85B-Fc and 10 µg CpG adjuvant (ODN1826, invivogen, France). After 14 days, revaccination was performed to boost the immune reaction [28]. Thereafter, the mice were nebulized daily with 1% OVA for 5 days, with each session lasting 20 minutes [27]. The timeline for ESAT6-Fc treatment displayed in Figure 1. Finally, the mice were euthanized to collect bronchoalveolar lavage fluid (BALF), serum, as well as lung, heart, liver, spleen, and kidney samples for further experiments.
4.5. Flow Cytometry
Right lung lobes were cut into fragments, and digested in RPMI-1640 medium containing 1 mg/mL collagenase I (D8140, Solarbio, China) and 10 μg/mL deoxy ribonucnase I (D8071, Solarbio, China) for 60 min. The single-cell suspension was centrifuged before supernatant removing, and the cells were resuspended in staining buffer (E-CK-A107, Elabsence, Wuhan, China) and labeled with FITC-CD4 and Percp-efluor710-IL-4 (Thermo Fisher, America) for Th2; Efluor450-CD45、APC-CD11b and PE-CD170 (Thermo Fisher, America) for eosinophils;PE-IL-17A (Thermo Fisher, America) for Th17. The percentage of Th2, Th17 and eosinophils was analyzed by BD FACSCelesta Flow Cytometer (Becton, Dickinson and Company, Americ).
4.6. Enzyme Linked Immunosorbent Assay (ELISA)
levels of IL-4, IL-13, IL-17A and IgE in BALF were measured using ELISA kits (Jiangsu Jingmei Biotechnology Co., Ltd) according to the manufacturer's instructions.
4.7. Lung Histological Analysis
Left lungs were fixed with 4% paraformaldehyde (PFA) for 24 h at room temperature, and cut into 5 μm sections after embedded in paraffin. The deparaffinized and rehydrated tissue sections were stained with hematoxylin and eosin (H&E) solution or with periodic acid Schiff (PAS) stain. The severity of inflammation on H&E-stained lung sections was evaluated as the following grade: “0”, normal, “1”: few cells, “2”: a ring of inflammatory cells, 1 cell layer deep, “3”: a ring of inflammatory cells 2–4 cells deep, and “4”: a ring of inflammatory cells of > 4 cells deep [29]. Goblet-cell hyperplasia was assessed from PAS-stained lung sections according to the percentage of goblet cells in the epithelial cells: “0”, no goblet cells; “1”, < 25%; “2”, 25–50% ; “3”, 51–80% ; and “4” > 80% [30].
4.8. cDNA Library Construction and RNA-seq Analysis
Total RNA from lung tissue of each mice was extracted with the TRIzol Reagent (Invitrogen, CA, USA). The purity and quantification of RNA determined using the NanoDrop 2000 spectrophotometer (Thermo Scientific, USA), and RNA integrity was assessed using Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Then the libraries were constructed using the VAHTS Universal V6 RNA-seq Library Prep kit. RNA-Seq analysis were performed by Shanghai OE Biotechnology Co., Ltd. (Shanghai, China). Gene expression estimates and differentially expressed genes (DEGs) were identified based on fold change (FC) >2 or < 0.5 and P < 0.05. Based on DEG annotations, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were conducted. Raw Illumina sequences were deposited in the Gene Expression Omnibus (GEO) (accession number: GSE274427).
4.9. KEGG Pathways Enrichment Analysis
Pathway-based analysis helps to understand each gene’s biological functions. For this experiment, the enrichment level of DEGs in the KEGG pathways (http://www.genome.jp/kegg/) was obtained. Furthermore, pathways with p < 0.05 were considered significant enrichment.
4.10. Validation of RNA-seq Results
A total of eight DEGs were selected for the quantitative real-time PCR (RT-qPCR) experiment to validate the RNA-seq results and the relative gene expression was calculated with the 2−ΔΔCt. β-actin was used as a reference gene during the experiment, and primers for RT- qPCR are shown in Table 1. The RNA was the same as that used for cDNA library construction (Section 2.8). ABI 9700 system was used for the RT- qPCR. The experiment was conducted three times, with three replications for each treatment.
4.11. Statistical Analysis
Data were expressed as the mean ± standard deviation (SD) and analyzed using GraphPad Prism 6 software (GraphPad Software, Inc., San Diego, CA, USA). Bioinformatic analysis was performed using the OECloud tools at https://cloud.oebiotech.com. Student’s T-test was used to compare the difference between two groups. P-value < 0.05 was statistically significant.
5. Conclusions
In this study, pcDNA3.1(+)-Rv3875-Fc was successfully constructed, and high-purity ESAT6-Fc fusion protein was obtained. The ESAT6-Fc fusion protein immunized via nasal mucosa significantly inhibited the aggregation of Th2 cells, Th17 cells, and eosinophils in OVA-induced lungs, as well as inhibited the proliferation of bronchial goblet cells and down-regulated the levels of IL-4, IL-13, IL-17 and IgE in BALF. In addition, no histopathological changes were observed in the heart, liver, spleen, or kidney tissues of mice in the ESAT6-Fc fusion protein treated group compared to the control group.
According to KEGG pathway enrichment analysis, DEGs were significantly enriched in signaling pathways including Natural Killer Cell-Mediated Cytotoxicity, T Cell Receptor, Th1 and Th2 Cell Differentiation, Th17 Cell Differentiation, and Chemokine in immune system of organismal systems before and after ESAT6 treatment.
Additionally, validation results for Cd28, Icos, Cd48, Cd247, Cd40l, Itgal, Itgb2 and Ccl5 were in line with the results of transcriptome analysis. This study revealed the fundamental immune regulatory mechanism underlying the effects of ESAT6 on OVA-induced AA. More importantly, this study provides valuable insights into the application of mucosal immunotherapy in treating AA and ESAT6-Fc might act as a secure mucosal immunotherapy agent for AA.
Author Contributions
Data curation, formal analysis, J.W.(Jing Wang); resources, writing-original draft, M.Y (Maosheng yang); formal analysis, methodology, Y.Y. (Yuxin Yang); software, visualization, T.W. (Tiansong Wang); validation, conceptualization, M.H (Maiyan Hai); funding acquisition, writing-review and editing, W.Z.(Wei Zhang); supervision, project administration, Y.Q. (Yanyan Qin); funding acquisition, investigation, Y.Y.(Yuan Yang); data curation, resources, Z.H. (Zihan Dong); visualization, validation, Y.Y. (Yanyu Han); conceptualization, methodology, R.M.(Rui Ma); funding acquisition, project administration, Q.W. (Qiaofeng Wan).
Funding
This research was sponsored by the National Natural Science Foundation of China (No. 82360802), the Ningxia Natural Science Foundation (No. 2024AAC03283), and Innovation and Entrepreneurship Training Program for College Students (S202410752038).
Acknowledgments
The authors are grateful to Shanghai OE Biotechnology Co., Ltd. for bioinformatics analysis.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Conde, E.; Bertrand, R.; Balbino, B.; Bonnefoy, J.; Stackowicz, J.; Caillot, N.; Colaone, F.; Hamdi, S.; Houmadi, R.; Loste, A.; et al. Dual vaccination against IL-4 and IL-13 protects against chronic allergic asthma in mice. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Moreno-Córdova, V.; Berra-Romani, R.; Flores Mendoza, L.K.; Reyes-Leyva, J. Th17 Lymphocytes in Children with Asthma: Do They Influence Control? Pediatr Allergy Immunol Pulmonol. 2021, 34, 147–15. [Google Scholar] [CrossRef] [PubMed]
- Morianos, I.; Semitekolou, M. Dendritic Cells: Critical Regulators of Allergic Asthma. Int. J. Mol. Sci. 2020, 21, 7930. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef]
- Maspero, J.; Adir, Y.; Al-Ahmad, M.; Celis-Preciado, C.A.; Colodenco, F.D.; Giavina-Bianchi, P.; Lababidi, H.; Ledanois, O.; Mahoub, B.; Perng, D.-W.; et al. Type 2 inflammation in asthma and other airway diseases. ERJ Open Res. 2022, 8. [Google Scholar] [CrossRef]
- Luo, W.; Hu, J.; Xu, W.; Dong, J. ; Distinct spatial and temporal roles for Th1, Th2, and Th17 cells in asthma. Front Immunol. 2022, 12, 974066. [Google Scholar] [CrossRef]
- Akdis, C.A.; Arkwright, P.D.; Brüggen, M.-C.; Busse, W.; Gadina, M.; Guttman-Yassky, E.; Kabashima, K.; Mitamura, Y.; Vian, L.; Wu, J.; et al. Type 2 immunity in the skin and lungs. Allergy 2020, 75, 1582–1605. [Google Scholar] [CrossRef]
- Yamauchi, K.; Ogasawara, M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. Int. J. Mol. Sci. 2019, 20, 1733. [Google Scholar] [CrossRef]
- Yu, L.; Liu, Q.; Canning, B.J. Evidence for autocrine and paracrine regulation of allergen-induced mast cell mediator release in the guinea pig airways. Eur. J. Pharmacol. 2017, 822, 108–118. [Google Scholar] [CrossRef]
- Akdis, C.A. Does the epithelial barrier hypothesis explain the increase in allergy, autoimmunity and other chronic conditions? Nat. Rev. Immunol. 2021, 21, 739–751. [Google Scholar] [CrossRef]
- Sun, S.; Gu, Y.; Wang, J.; Chen, C.; Han, S.; Che, H. Effects of Fatty Acid Oxidation and Its Regulation on Dendritic Cell-Mediated Immune Responses in Allergies: An Immunometabolism Perspective. J. Immunol. Res. 2021, 2021, 1–10. [Google Scholar] [CrossRef]
- Johnson, C.C.; Ownby, D.R. The infant gut bacterial microbiota and risk of pediatric asthma and allergic diseases. Transl. Res. 2017, 179, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Kowalewicz-Kulbat, M.; Locht, C. BCG for the prevention and treatment of allergic asthma. Vaccine 2021, 39, 7341–7352. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Tang, A.-Z.; Xu, M.-L.; Chen, H.-L.; Wang, F.; Li, C.-Q. Mycobacterium vaccae attenuates airway inflammation by inhibiting autophagy and activating PI3K/Akt signaling pathway in OVA-induced allergic airway inflammation mouse model. Mol. Immunol. 2024, 173, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Anes, E.; Pires, D.; Mandal, M.; Azevedo-Pereira, J.M. ESAT-6 a Major Virulence Factor of Mycobacterium tuberculosis. Biomolecules 2023, 13, 968. [Google Scholar] [CrossRef] [PubMed]
- Zonghai, C.; Tao, L.; Pengjiao, M.; Liang, G.; Rongchuan, Z.; Xinyan, W.; Wenyi, N.; Wei, L.; Yi, W.; Lang, B. Mycobacterium tuberculosis ESAT6 modulates host innate immunity by downregulating miR-222-3p target PTEN. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2022, 1868, 166292. [Google Scholar] [CrossRef]
- Mustafa, A.S. Chemical and Biological Characterization of Mycobacterium tuberculosis-Specific ESAT6-Like Proteins and Their Potentials in the Prevention of Tuberculosis and Asthma. Med Princ. Pract. 2023, 32, 217–224. [Google Scholar] [CrossRef]
- Pyzik, M.; Kozicky, L.K.; Gandhi, A.K.; Blumberg, R.S. The therapeutic age of the neonatal Fc receptor. Nat. Rev. Immunol. 2023, 23, 415–432. [Google Scholar] [CrossRef]
- Qian, S.; Zhang, D.; Yang, Z.; Li, R.; Zhang, X.; Gao, F.; Yu, L. The role of immunoglobulin transport receptor, neonatal Fc receptor in mucosal infection and immunity and therapeutic intervention. Int. Immunopharmacol. 2024, 138, 112583. [Google Scholar] [CrossRef]
- Jafari, R.; Zolbanin, N.M.; Rafatpanah, H.; Majidi, J.; Kazemi, T. Fc-fusion Proteins in Therapy: An Updated View. Curr. Med. Chem. 2017, 24, 1228–1237. [Google Scholar] [CrossRef]
- Fan, Y.; Gao, D.; Zhang, Z. Telitacicept, a novel humanized, recombinant TACI-Fc fusion protein, for the treatment of systemic lupus erythematosus. Drugs Today 2022, 58, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, L.; Feng, X.; Gao, C.; Gao, L.; Zhan, Y.; Wang, Z.; Zhao, M.; Yin, H.; Lu, Q. TIGIT-Fc fusion protein alleviates murine lupus nephritis through the regulation of SPI-B-PAX5-XBP1 axis-mediated B-cell differentiation. J. Autoimmun. 2023, 139, 103087. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-X.; Li, H.; Bai, L.; Yao, S.; Zhang, W.; Wang, T.-S.; Wan, Q.-F. Bioinformatics analysis of ceRNA regulatory network of baicalin in alleviating pathological joint alterations in CIA rats. Eur. J. Pharmacol. 2023, 951, 175757. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhou, X.; Li, R.; Michal, J.J.; Zhang, S.; Dodson, M.V.; Zhang, Z.; Harland, R.M. Whole transcriptome analysis with sequencing: methods, challenges and potential solutions. Cell. Mol. Life Sci. 2015, 72, 3425–3439. [Google Scholar] [CrossRef]
- Rish, A.J.; Drennen, J.K.; Anderson, C.A. Metabolic trends of Chinese hamster ovary cells in biopharmaceutical production under batch and fed-batch conditions. Biotechnol. Prog. 2021, 38, e3220. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Dickson, A.J. Reprogramming of Chinese hamster ovary cells towards enhanced protein secretion. Metab. Eng. 2021, 69, 249–261. [Google Scholar] [CrossRef]
- Böll, S.; Ziemann, S.; Ohl, K.; Klemm, P.; Rieg, A.D.; Gulbins, E.; Becker, K.A.; Kamler, M.; Wagner, N.; Uhlig, S.; et al. Acid sphingomyelinase regulates TH2 cytokine release and bronchial asthma. Allergy 2019, 75, 603–615. [Google Scholar] [CrossRef]
- Li, W.; Wang, T.; Rajendrakumar, A.M.; Acharya, G.; Miao, Z.; Varghese, B.P.; Yu, H.; Dhakal, B.; LeRoith, T.; Karunakaran, A. An FcRn-targeted mucosal vaccine against SARS-CoV-2 infection and transmission. Nat Commun. 2023, 14, 711. [Google Scholar] [CrossRef]
- Myou, S.; Leff, A.R.; Myo, S.; Boetticher, E.; Tong, J.; Meliton, A.Y.; Liu, J.; Munoz, N.M.; Zhu, X. Blockade of Inflammation and Airway Hyperresponsiveness in Immune-sensitized Mice by Dominant-Negative Phosphoinositide 3-Kinase–TAT. J. Exp. Med. 2003, 198, 1573–1582. [Google Scholar] [CrossRef]
- Tanaka, H.; Masuda, T.; Tokuoka, S.; Komai, M.; Nagao, K.; Takahashi, Y.; Nagai, H. The effect of allergen-induced airway inflammation on airway remodeling in a murine model of allergic asthma. Inflamm. Res. 2001, 50, 616–624. [Google Scholar] [CrossRef]
- Chemmannur, S.V.; Badhwar, A.J.; Mirlekar, B.; Malonia, S.K.; Gupta, M.; Wadhwa, N.; Bopanna, R.; Mabalirajan, U.; Majumdar, S.; Ghosh, B.; et al. Nuclear matrix binding protein SMAR1 regulates T-cell differentiation and allergic airway disease. Mucosal Immunol. 2015, 8, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Li, K.; Song, Z.; Wang, Q.; Wang, J.; Li, X.; Li, Y.; Zhang, Q.; Zhu, Y.; Chen, H. Nitric oxide hinders club cell proliferation through Gdpd2 during allergic airway inflammation. FEBS Open Bio 2023, 13, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Netto, K.G.; Sokulsky, L.A.; Zhou, L.; Xu, H.; Liu, C.; Wang, M.; Wang, H.; Li, H.; Zhang, G.; et al. Single-cell RNA transcriptomic analysis identifies Creb5 and CD11b-DCs as regulator of asthma exacerbations. Mucosal Immunol. 2022, 15, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Scordamaglia, F.; Balsamo, M.; Scordamaglia, A.; Moretta, A.; Mingari, M.C.; Canonica, G.W.; Moretta, L.; Vitale, M. Perturbations of natural killer cell regulatory functions in respiratory allergic diseases. J. Allergy Clin. Immunol. 2008, 121, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Diaz-Meco, M.T.; Moscat, J. The signaling adapter p62 is an important mediator of T helper 2 cell function and allergic airway inflammation. EMBO J. 2006, 25, 3524–3533. [Google Scholar] [CrossRef]
- Tumes, D.J.; Papadopoulos, M.; Endo, Y.; Onodera, A.; Hirahara, K.; Nakayama, T. Epigenetic regulation of T-helper cell differentiation, memory, and plasticity in allergic asthma. Immunol Rev. 2017, 278, 8–19. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X.; Sheng, A.; Weng, C.; Zhu, T.; Zhao, W.; Li, C. γ-Secretase Inhibitor Alleviates Acute Airway Inflammation of Allergic Asthma in Mice by Downregulating Th17 Cell Differentiation. Mediat. Inflamm. 2015, 2015, 258168. [Google Scholar] [CrossRef]
- Kamranvar, S.A.; Rani, B.; Johansson, S. Cell Cycle Regulation by Integrin-Mediated Adhesion. Cells 2022, 11, 2521. [Google Scholar] [CrossRef]
- Na Kim, M.; Hong, J.Y.; Shim, D.H.; Sol, I.S.; Kim, Y.S.; Lee, J.H.; Kim, K.W.; Lee, J.M.; Sohn, M.H. Activated Leukocyte Cell Adhesion Molecule Stimulates the T-Cell Response in Allergic Asthma. Am. J. Respir. Crit. Care Med. 2018, 197, 994–1008. [Google Scholar] [CrossRef]
- Nikovics, K.; Favier, A.-L.; Rocher, M.; Mayinga, C.; Gomez, J.; Dufour-Gaume, F.; Riccobono, D. In Situ Identification of Both IL-4 and IL-10 Cytokine–Receptor Interactions during Tissue Regeneration. Cells 2023, 12, 1522. [Google Scholar] [CrossRef]
- Guo, H.-H.; Han, Y.-X.; Rong, X.-J.; Shen, Z.; Shen, H.-R.; Kong, L.-F.; Guo, Y.D.; Li, J.-Z.; Xu, B.; Gao, T.-L.; et al. Alleviation of allergic asthma by rosmarinic acid via gut-lung axis. Phytomedicine 2024, 126, 155470. [Google Scholar] [CrossRef]
- Papadopoulos, N.G.; Miligkos, M.; Xepapadaki, P. A Current Perspective of Allergic Asthma: From Mechanisms to Management. Handb Exp Pharmacol. 2022, 268, 69–93. [Google Scholar]
- Heffler, E.; Madeira, L.N.G.; Ferrando, M.; Puggioni, F.; Racca, F.; Malvezzi, L.; Passalacqua, G.; Canonica, G.W. Inhaled Corticosteroids Safety and Adverse Effects in Patients with Asthma. J. Allergy Clin. Immunol. Pract. 2018, 6, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Zuberi, F.F.; Haroon, M.A.; Haseeb, A.; Khuhawar, S.M. Role of Montelukast in Asthma and Allergic rhinitis patients. Pak. J. Med Sci. 2020, 36, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.K. The Role of ESX-1 in Mycobacterium tuberculosis Pathogenesis. Microbiol Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Chawes, B.L.; Wolsk, H.M.; Carlsson, C.J.; Rasmussen, M.A.; Følsgaard, N.; Stokholm, J.; Bønnelykke, K.; Brix, S.; Schoos, A.M.; Bisgaard, H. Neonatal airway immune profiles and asthma and allergy endpoints in childhood. Allergy 2021, 76, 3713–3722. [Google Scholar] [CrossRef] [PubMed]
- Hellings, P.W.; Steelant, B. Epithelial barriers in allergy and asthma. J. Allergy Clin. Immunol. 2020, 145, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.; Behrendt, A.-K.; Henken, S.; Wölbeling, F.; Maus, U.A.; Hansen, G. Pneumococcal pneumonia suppresses allergy development but preserves respiratory tolerance in mice. Immunol. Lett. 2015, 164, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Nieto, A.; Mazón, A.; Nieto, M.; Calderón, R.; Calaforra, S.; Selva, B.; Uixera, S.; Palao, M.J.; Brandi, P.; Conejero, L.; et al. Bacterial Mucosal Immunotherapy with MV130 Prevents Recurrent Wheezing in Children: A Randomized, Double-Blind, Placebo-controlled Clinical Trial. Am. J. Respir. Crit. Care Med. 2021, 204, 462–472. [Google Scholar] [CrossRef]
- Sun, L.; Su, Y.; Jiao, A.; Wang, X.; Zhang, B. T cells in health and disease. Signal Transduct. Target. Ther. 2023, 8, 235. [Google Scholar] [CrossRef]
- Gutiérrez-Vera, C.; García-Betancourt, R.; Palacios, P.A.; Müller, M.; Montero, D.A.; Verdugo, C.; Ortiz, F.; Simon, F.; Kalergis, A.M.; González, P.A.; et al. Natural killer T cells in allergic asthma: implications for the development of novel immunotherapeutical strategies. Front. Immunol. 2024, 15, 1364774. [Google Scholar] [CrossRef] [PubMed]
- Minnicozzi, M.; Sawyer, R.T.; Fenton, M.J. Innate immunity in allergic disease. Immunol. Rev. 2011, 242, 106–127. [Google Scholar] [CrossRef] [PubMed]
- Castan, L.; Magnan, A.; Bouchaud, G. Chemokine receptors in allergic diseases. Allergy 2016, 72, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Nagai, S.; Azuma, M. The CD28-B7 Family of Co-signaling Molecules. Adv Exp Med Biol. 2019, 1189, 25–51. [Google Scholar]
- West, S.M.; A Deng, X. Considering B7-CD28 as a family through sequence and structure. Exp. Biol. Med. 2019, 244, 1577–1583. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, X.; Tian, T.; Pang, W. The role of ICOS in allergic disease: Positive or Negative? Int Immunopharmacol. 2022, 103, 108394. [Google Scholar] [CrossRef]
- Dong, M.; Chang, J.; Lebel, M.; Gervais, N.; Fournier, M.; Gauthier, M.; Suh, W.; Melichar, H.J. The ICOS–ICOSL pathway tunes thymic selection. Immunol. Cell Biol. 2022, 100, 205–217. [Google Scholar] [CrossRef]
- Munitz, A.; Bachelet, I.; Levi-Schaffer, F. CD48 as a Novel Target in Asthma Therapy. Recent Patents Inflamm. Allergy Drug Discov. 2007, 1, 9–12. [Google Scholar] [CrossRef]
- Breuer, O.; Gangwar, R.S.; Seaf, M.; Barhoum, A.; Kerem, E.; Levi-Schaffer, F. Evaluation of Soluble CD48 Levels in Patients with Allergic and Nonallergic Asthma in Relation to Markers of Type 2 and Non-Type 2 Immunity: An Observational Study. J. Immunol. Res. 2018, 2018, 1–7. [Google Scholar] [CrossRef]
- Dexiu, C.; Xianying, L.; Yingchun, H.; Jiafu, L. Advances in CD247. Scand J Immunol. 2022, 96, e13170. [Google Scholar] [CrossRef]
- Vicente, C.T.; A Revez, J.; Ferreira, M.A.R. Lessons from ten years of genome-wide association studies of asthma. Clin. Transl. Immunol. 2017, 6, e165–e165. [Google Scholar] [CrossRef]
- Tang, T.; Cheng, X.; Truong, B.; Sun, L.; Yang, X.; Wang, H. Molecular basis and therapeutic implications of CD40/CD40L immune checkpoint. Pharmacol Ther. 2021, 219, 107709. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhu, H.; Huang, S.; Zhang, Y.; Shen, Y. Establishment of a diagnostic model based on immune-related genes in children with asthma. Heliyon 2024, 10, e25735. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, M.; Bolduan, V.; Haist, M.; Stege, H.; Hieber, C.; Johann, L.; Schelmbauer, C.; Blanfeld, M.; Karram, K.; Schunke, J. β2 Integrins on Dendritic Cells Modulate Cytokine Signaling and Inflammation-Associated Gene Expression, and Are Required for Induction of Autoimmune Encephalomyelitis. Cells 2022, 11, 2188. [Google Scholar] [CrossRef] [PubMed]
- Klaus, T.; Wilson, A.; Fichter, M.; Bros, M.; Bopp, T.; Grabbe, S. The Role of LFA-1 for the Differentiation and Function of Regulatory T Cells—Lessons Learned from Different Transgenic Mouse Models. Int. J. Mol. Sci. 2023, 24, 6331. [Google Scholar] [CrossRef] [PubMed]
- Lantero, S.; Alessandri, G.; Spallarossa, D.; Scarso, L.; Rossi, G. LFA-1 expression by blood eosinophils is increased in atopic asthmatic children and is involved in eosinophil locomotion. Eur. Respir. J. 1998, 12, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.E.; Guabiraba, R.; Russo, R.C.; Teixeira, M.M. Targeting CCL5 in inflammation. Expert Opin. Ther. Targets 2013, 17, 1439–1460. [Google Scholar] [CrossRef]
- Sandford, A.J.; Zhu, S.; Bai, T.R.; Fitzgerald, J.M.; Paré, P.D. The role of the C-C chemokine receptor-5 Delta32 polymorphism in asthma and in the production of regulated on activation, normal T cells expressed and secreted. J Allergy Clin Immunol. 2001, 108, 69–73. [Google Scholar] [CrossRef]
- Sereinig, S.; Stukova, M.; Zabolotnyh, N.; Ferko, B.; Kittel, C.; Romanova, J.; Vinogradova, T.; Katinger, H.; Kiselev, O.; Egorov, A. Influenza Virus NS Vectors Expressing theMycobacterium tuberculosisESAT-6 Protein Induce CD4+Th1 Immune Response and Protect Animals against Tuberculosis Challenge. Clin. Vaccine Immunol. 2006, 13, 898–904. [Google Scholar] [CrossRef]
Figure 3.
Principal component analysis (PCA) (A) and Statistics of Differently Expressed genes (DEGs) (B). N: normal group; M:1% OVA induced group; T: ESAT6-Fc treatment group.
Figure 3.
Principal component analysis (PCA) (A) and Statistics of Differently Expressed genes (DEGs) (B). N: normal group; M:1% OVA induced group; T: ESAT6-Fc treatment group.
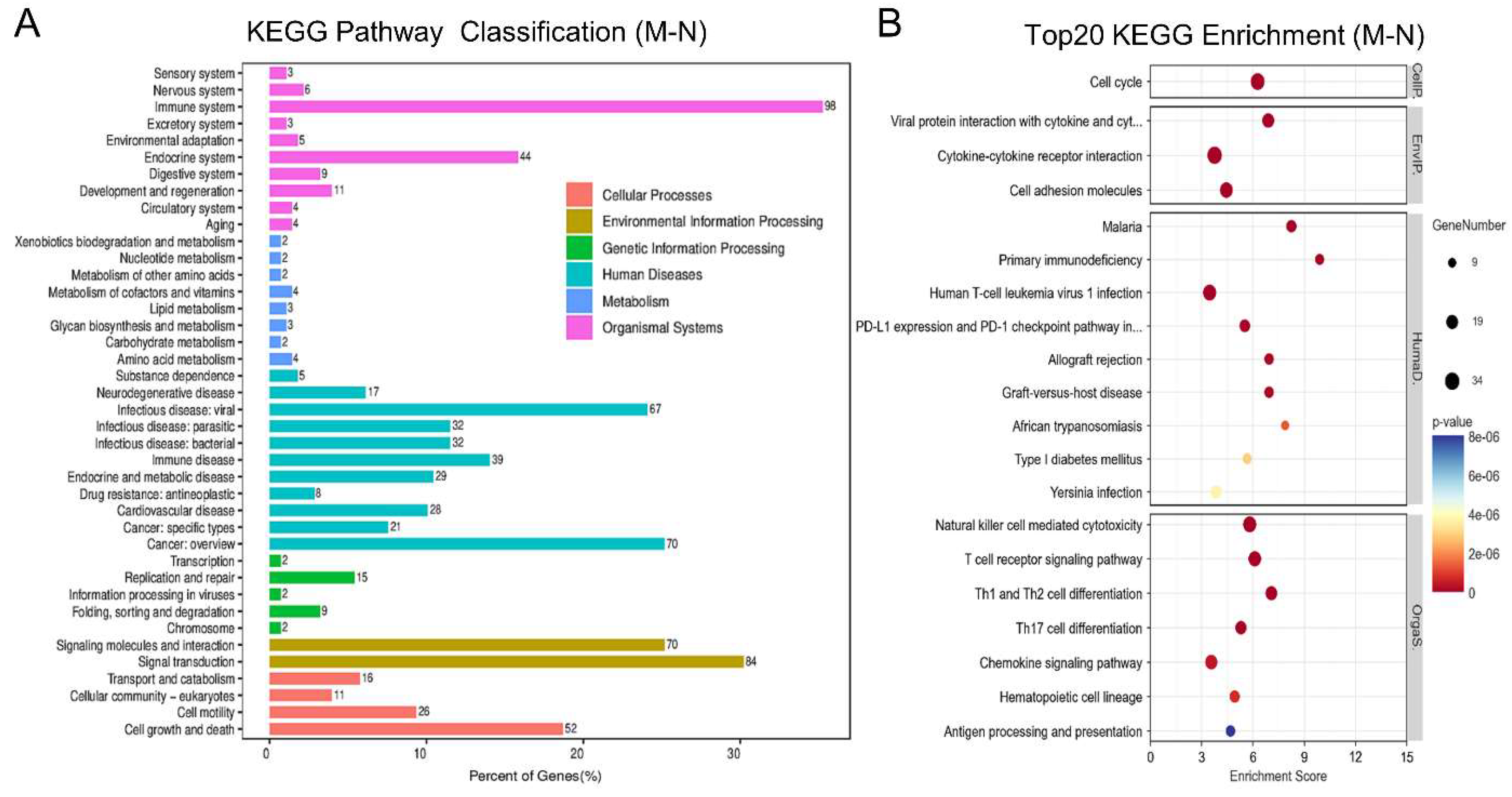
Figure 4.
Analysis of differentially expressed genes (DEGs) between the M and N groups. The gene KEGG pathway Classification (A); Top 20 KEGG enrichment pathways based on rich factor and P-value (B). N: normal group; M:1% OVA induced group.
Figure 4.
Analysis of differentially expressed genes (DEGs) between the M and N groups. The gene KEGG pathway Classification (A); Top 20 KEGG enrichment pathways based on rich factor and P-value (B). N: normal group; M:1% OVA induced group.
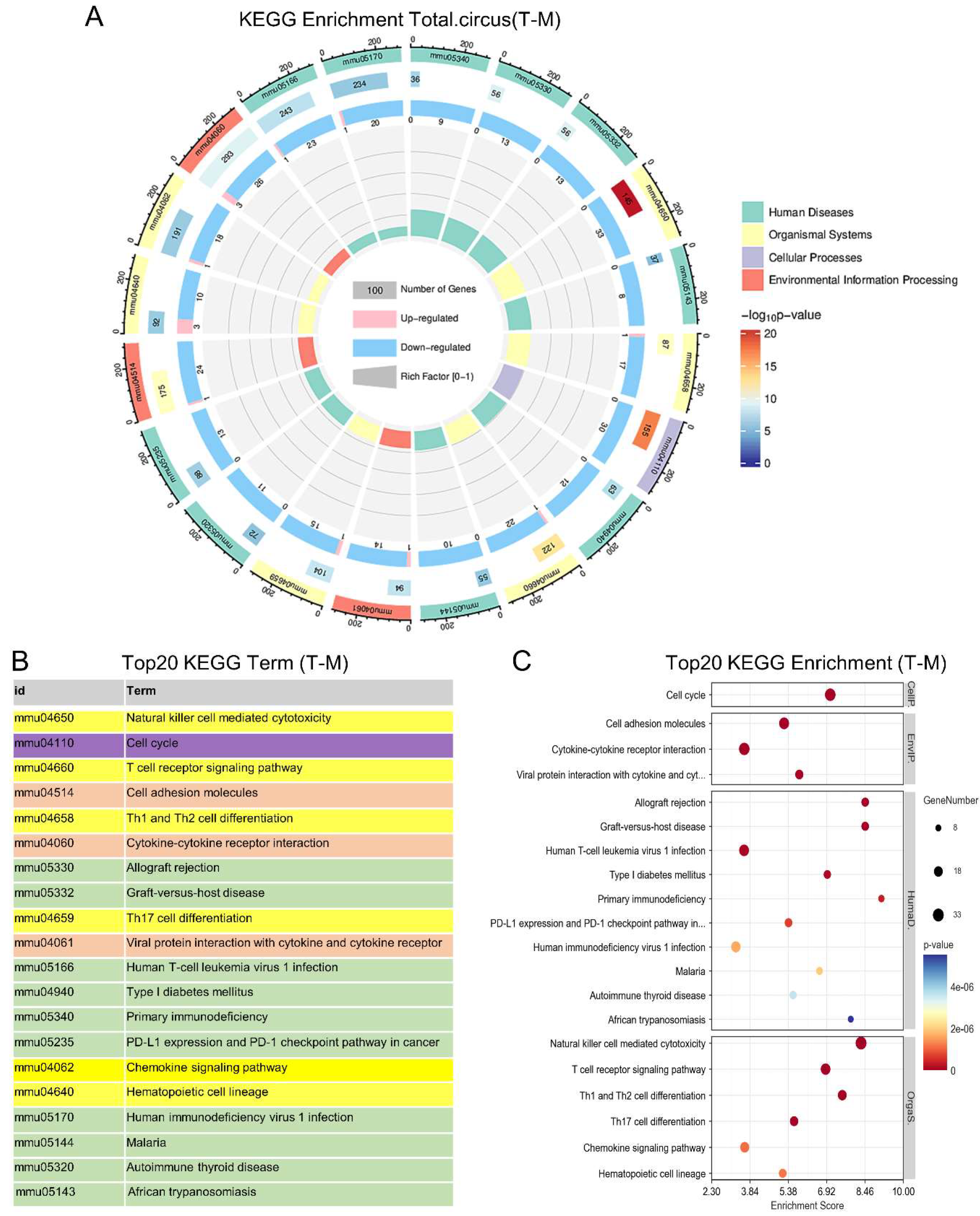
Figure 5.
KEGG enrichment analysis of pathways. KEGG enrichment circle chart (A): The first circle represents the pathway of the first 20 enrichments with different colors represent various classes; The second circle represents the number of genes involved in this pathway as well as the p-value; The third circle contains the number of differential genes that have been adjusted up and down, with pink color up, and blue color down; Top 20 KEGG enrichment term (B); Top 20 KEGG enrichment pathways (C). M:1% OVA induced group; T: ESAT6-Fc treatment group.
Figure 5.
KEGG enrichment analysis of pathways. KEGG enrichment circle chart (A): The first circle represents the pathway of the first 20 enrichments with different colors represent various classes; The second circle represents the number of genes involved in this pathway as well as the p-value; The third circle contains the number of differential genes that have been adjusted up and down, with pink color up, and blue color down; Top 20 KEGG enrichment term (B); Top 20 KEGG enrichment pathways (C). M:1% OVA induced group; T: ESAT6-Fc treatment group.
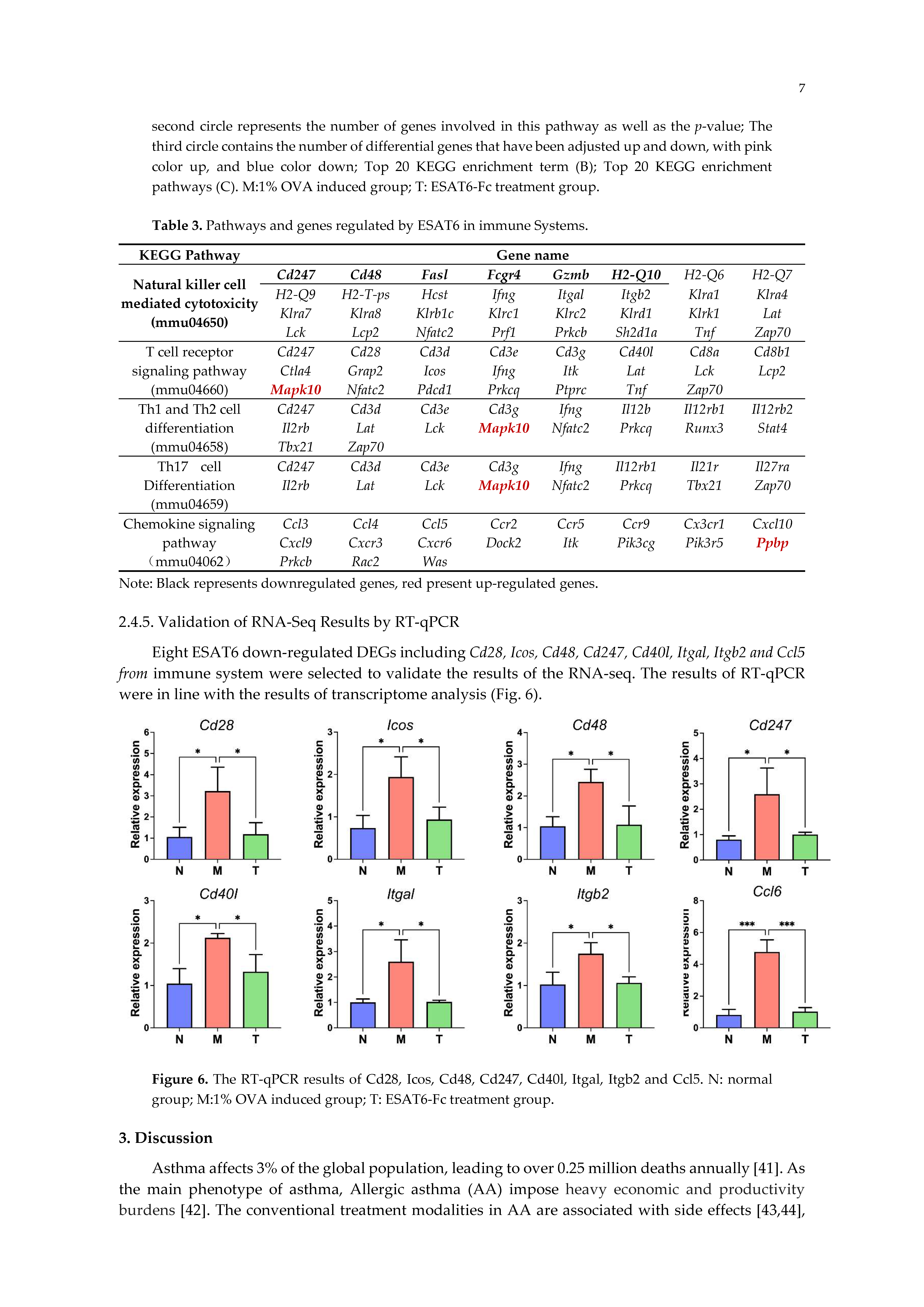
Figure 6.
The RT-qPCR results of Cd28, Icos, Cd48, Cd247, Cd40l, Itgal, Itgb2 and Ccl5. N: normal group; M:1% OVA induced group; T: ESAT6-Fc treatment group.
Figure 6.
The RT-qPCR results of Cd28, Icos, Cd48, Cd247, Cd40l, Itgal, Itgb2 and Ccl5. N: normal group; M:1% OVA induced group; T: ESAT6-Fc treatment group.
Table 3.
Pathways and genes regulated by ESAT6 in immune Systems.
| KEGG Pathway | Gene name | |||||||
|---|---|---|---|---|---|---|---|---|
|
Natural killer cell mediated cytotoxicity (mmu04650) |
Cd247 | Cd48 | Fasl | Fcgr4 | Gzmb | H2-Q10 | H2-Q6 | H2-Q7 |
| H2-Q9 | H2-T-ps | Hcst | Ifng | Itgal | Itgb2 | Klra1 | Klra4 | |
| Klra7 | Klra8 | Klrb1c | Klrc1 | Klrc2 | Klrd1 | Klrk1 | Lat | |
| Lck | Lcp2 | Nfatc2 | Prf1 | Prkcb | Sh2d1a | Tnf | Zap70 | |
| T cell receptor signaling pathway (mmu04660) |
Cd247 | Cd28 | Cd3d | Cd3e | Cd3g | Cd40l | Cd8a | Cd8b1 |
| Ctla4 | Grap2 | Icos | Ifng | Itk | Lat | Lck | Lcp2 | |
| Mapk10 | Nfatc2 | Pdcd1 | Prkcq | Ptprc | Tnf | Zap70 | ||
| Th1 and Th2 cell differentiation (mmu04658) |
Cd247 | Cd3d | Cd3e | Cd3g | Ifng | Il12b | Il12rb1 | Il12rb2 |
| Il2rb | Lat | Lck | Mapk10 | Nfatc2 | Prkcq | Runx3 | Stat4 | |
| Tbx21 | Zap70 | |||||||
| Th17 cell Differentiation (mmu04659) |
Cd247 | Cd3d | Cd3e | Cd3g | Ifng | Il12rb1 | Il21r | Il27ra |
| Il2rb | Lat | Lck | Mapk10 | Nfatc2 | Prkcq | Tbx21 | Zap70 | |
| Chemokine signaling pathway (mmu04062) |
Ccl3 | Ccl4 | Ccl5 | Ccr2 | Ccr5 | Ccr9 | Cx3cr1 | Cxcl10 |
| Cxcl9 | Cxcr3 | Cxcr6 | Dock2 | Itk | Pik3cg | Pik3r5 | Ppbp | |
| Prkcb | Rac2 | Was | ||||||
Note: Black represents downregulated genes, red present up-regulated genes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



