
Submitted:
04 November 2024
Posted:
06 November 2024
You are already at the latest version
Abstract
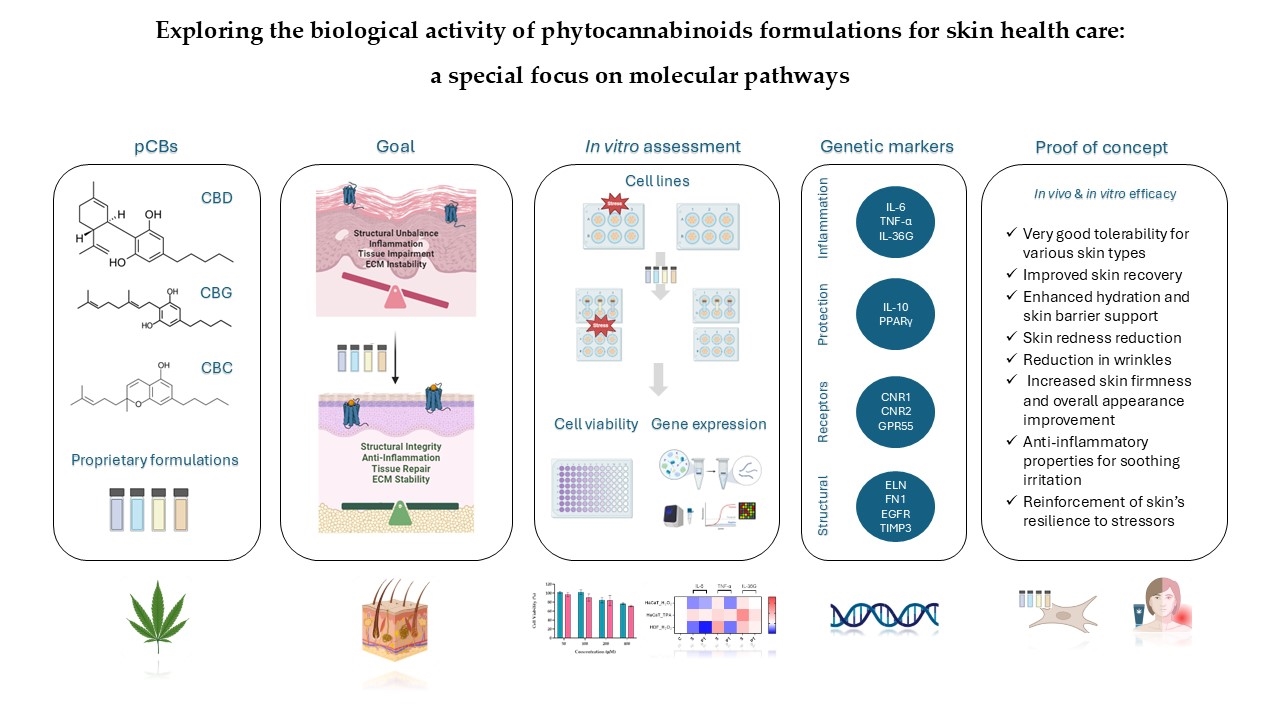
Background: Recent advancements have highlighted the potential of cannabis and its phy-tocannabinoids (pCBs) in skin health applications. These compounds, through their interac-tion with the endocannabinoid system (ECS), show promise for skin health products. Their ability to regulate inflammation, oxidative stress, and cellular proliferation makes them bene-ficial in addressing skin issues like inflammation, scarring, healing, acne, and aging, posi-tioning them as valuable tools for innovative skincare solutions.
Methods: We investigated the cellular and molecular effects of proprietary pCB-based for-mulations on ECS modulation, inflammation, and skin regeneration. Using human dermal fibroblasts (HDF) and keratinocytes (HaCat), we assessed the formulations in both pre-treatment and treatment scenarios following exposure to stress-inducing agents. Key molecular markers were analyzed to tackle their efficacy in mitigating inflammation and promoting structural integrity and regeneration.
Results: Our findings show that these formulations significantly reduce inflammation, pro-mote skin regeneration, and improve structural functions. In vivo studies confirmed that the formulations were well-tolerated and led to noticeable improvements in skin health, including enhanced barrier function.
Conclusions: This study demonstrates the safety and efficacy of pCB-based formulations for cosmeceutical applications. By combining molecular analysis with in vivo testing, our research provides new insights into the therapeutic potential of pCBs for managing various skin con-ditions.
Keywords:
Endocannabinoid system
; skin homeostasis
; skin inflammation
; skin regeneration
; phytocannabinoids
; cosmeceutical applications
; skin therapeutics
1. Introduction
Cannabis sativa is a complex plant with a long and outstanding history spanning over 4,000 years, particularly in Asia, with significant use in India for religious, recreational, and medicinal purposes [1]. In the last years, there has been an increased interest by the scientific community in cannabis and its constituents aiming to explore their therapeutic potential [2]. The diverse phytochemical profile of Cannabis sativa contributes to its wide-ranging effects and potential medical applications. The plant contains over 500 distinct compounds, including terpenes, flavonoids, alkaloids, and cannabinoids [3]. To date, approximately 125 cannabinoids have been identified and characterized [1], and these bioactive compounds are generally classified based on their origin [4,5]. The three primary classes of cannabinoids are:
- -
- Endocannabinoids, which are naturally synthesized in the human body. This class includes anandamide (EAE) and 2-arachidonoylglycerol (2-AG), which interact with the endocannabinoid system (ECS).
- -
- Phytocannabinoids (pCBs), which are produced by plants. Among the most studied pCBs are Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD), though many others have been identified and are currently under investigation for their potential therapeutic effects.
- -
Our research specifically focused on pCBs from Cannabis sativa. To date, more than 120 distinct pCBs have been identified, reflecting the plant’s remarkable chemical diversity [6]. These pCBs interact with the human ECS, influencing various physiological processes.
The unique properties of different pCBs have led to intense research in areas such as pain management and neurological disorders. In dermatology, the most studied pCBs include CBD, cannabigerol (CBG), cannabichromene (CBC), cannabinol (CBN), and THC [4,5]. These compounds have been explored for their potential roles in modulating skin homeostasis, including effects on inflammation, cell proliferation, sebum production, skin barrier function, and wound healing.
In vitro studies have provided substantial evidence for the pharmacological effects of these pCBs on various skin-related cell lines, including keratinocytes, sebocytes, and fibroblasts [4]. For instance, CBD has been shown to inhibit the proliferation of keratinocytes, suggesting its potential role in managing hyperproliferative skin disorders like psoriasis [5]. Similarly, CBG has been found to induce apoptosis in human skin fibroblasts, highlighting its potential utility in managing fibrotic skin diseases [6]. These findings underscore the therapeutic potential of pCBs in skin disorders, warranting further investigation into their mechanisms of action and their applications in both cosmeceuticals and clinical settings.
The ECS is a complex signaling system present in both the central and peripheral nervous system, as well as in many peripheral tissues[7,8]. It comprises cannabinoid receptors (CB1R and CB2R), endocannabinoids such as AEA and 2-AG, and the enzymes responsible for their synthesis and degradation [9,10]. Initially discovered in the brain, the ECS has since been found to extend its regulatory functions to various peripheral tissues, influencing numerous physiological pathways including pain perception, gastrointestinal activity, hormonal regulation, cardiovascular function, immune system modulation, and inflammatory responses [11,12].
The ECS plays a crucial role in maintaining skin homeostasis, regulating inflammation, and promoting skin regeneration by influencing various physiological processes, including cell proliferation, differentiation, and apoptosis. The interaction between endocannabinoids and their receptors in the skin suggests a significant role in maintaining skin health and responding to stressors. Recent scientific advancements have highlighted the potential therapeutic applications of cannabinoids in dermatological formulations, particularly in addressing skin disorders through the modulation of the ECS.
pCBs such as CBD, CBG, and CBC, have garnered attention for their potential benefits in cosmeceutical formulations. Importantly, pCBs have different affinities towards human receptors and the biological effects are often the result of the interaction with multiple molecular targets [4,13,14,15].
These compounds interact with ECS receptors in the skin, offering anti-inflammatory and antioxidant properties and hence the topical application of pCBs can potentially address various skin disorders, including eczema, psoriasis, and acne, by modulating inflammatory pathways and promoting skin regeneration [16,17,18].
CBD, one of the most studied non-psychoactive pCBs, has been shown to possess anti-inflammatory, antioxidant, and anti-proliferative properties. It modulates skin homeostasis by influencing key signaling pathways involved in skin cell proliferation and differentiation. Also, it has demonstrated efficacy in regulating the production of sebum in sebocytes and in reducing inflammation in keratinocytes, making it a promising candidate for the treatment of acne vulgaris and other inflammatory skin conditions [19,20]. Previous studies reported that in keratinocytes, the pCBs, CBD, CBG and CBC, were able to reduce the inflammation [21,22]. Sangiovanni and colleagues evaluated the effect of CBD on the wound healing and inflammation processes, using TNF-α as a proinflammatory stimulus. They found that CBD not only protected the cells but also induced the expression of genes related to antioxidant and detoxification systems, as well as those involved in keratinocyte differentiation and skin development. [23].In human fibroblasts, CBD was able to reduce inflammation and regulate the extracellular matrix (ECM). The authors observed a decrease on the cytokines’ expression, and an activation of proteinases that regulate the extracellular matrix components [21,24]. Atalay et al. (2020) investigated the antioxidant properties of CBD in keratinocytes exposed to UVB radiation and hydrogen peroxide. The study found that CBD upregulated the expression of several antioxidant genes, including superoxide dismutase and catalase. This upregulation provided significant protection against oxidative damage, suggesting that CBD could be used to prevent or mitigate oxidative stress-related skin aging and damage [25].
CBG, although less abundant in the cannabis plant, has also shown potential benefits for skin care. Research indicates that CBG may have antimicrobial, anti-inflammatory and analgesic properties, which could be beneficial in treating skin conditions characterized by inflammation and pain. CBG has been observed to influence the ECS in the skin, particularly through its interaction with cannabinoid receptors, potentially modulating inflammatory responses and skin cell proliferation [26,27]. Pérez et al. (2022) conducted in vitro and clinical evaluations of CBG using gene expression assays. The study demonstrated that CBG has a broad range of anti-inflammatory effects, modulating the expression of various genes involved in skin inflammation and immune responses. This work expands the scope of cannabinoids studied for dermatological applications beyond CBD, indicating that CBG could also play a significant role in skin health [18].
CBC has demonstrated potential as an anti-inflammatory and antimicrobial agent. Its ability to inhibit the lipoxygenase enzyme suggests that CBC may play a role in modulating inflammatory processes within the skin. Furthermore, CBC has shown potential in promoting skin homeostasis by influencing the activity of sebaceous glands, thereby offering a possible therapeutic avenue for conditions like acne [26].
A study by Palmieri et al. (2019) evaluated the effects of a CBD-enriched ointment on inflammatory skin diseases, demonstrating improvements in various clinical parameters and quality of life measures [28].
In the context of skin physiology, cannabinoid receptors play pivotal roles in maintaining skin homeostasis across various cell types like keratinocytes, fibroblasts, and immune cells. Tóth et al. (2019) underscored the critical influence of CB1 and CB2 receptors on processes such as cell proliferation, differentiation, and immune response regulation in the skin. In dermal fibroblasts, CBD's interaction with CB1 and CB2 receptors influences collagen production and skin elasticity, suggesting applications in anti-aging and wound healing. Moreover, the expression of structural and repair-associated genes provides insights into the regenerative effects of pCB-based formulations. Key genes like fibronectin (FN1), epidermal growth factor receptor (EGFR), and metallopeptidase inhibitor 3 (TIMP3) are crucial for skin differentiation, barrier function, and tissue repair [29]. Casares et al. (2019) examined CBD's impact on gene expression using a 3D model of human epidermis, highlighting CBD's ability to modulate FN1, EGFR, and TIMP3 expression. This modulation promotes skin health by enhancing barrier formation and tissue integrity, emphasizing CBD's potential therapeutic benefits in skincare formulations [23].
The antioxidant and anti-inflammatory properties of CBD, CBG and CBC make them highly promising candidates for enhancing overall skin health and promoting the regeneration of damaged skin tissues [18,25,30]. As an example, the anti-inflammatory effects of pCBS are crucial in managing skin disorders characterized by chronic inflammation, such as psoriasis, eczema, and acne. In these conditions, inflammation can cause redness, swelling, and discomfort, exacerbating the symptoms and delaying the healing process [31]. pCBs can help reduce this inflammation by modulating the skin's immune response, leading to calmer, less irritated skin. These compounds possess the ability to neutralize free radicals, which are reactive molecules that cause oxidative stress—a major factor in skin aging and the development of various skin conditions. By mitigating oxidative stress, pCBs help protect the skin from environmental damage, such as UV radiation and pollution, which can lead to premature aging, wrinkles, and loss of skin elasticity. By reducing both oxidative and chemical stress, pCBs support the skin's natural barrier function, helping to maintain hydration and prevent the entry of harmful substances. These properties suggest that pCBs could be valuable components in skincare formulations aimed at improving skin resilience, reducing the signs of aging, and treating chronic inflammatory skin diseases.
Considering the significant cosmetic and therapeutic potential of pCBs, the primary objective of the present study was to evaluate, in vitro, the effects of pCBs-based formulations on various human cell lines subjected to stress conditions. To achieve this, we used keratinocytes (HaCat) and human dermal fibroblasts (HDF). The expression levels of target genes associated with inflammation and regenerative processes were assessed and compared between cells treated with pCBs-based formulations and untreated controls.
To make it easier for readers, a list of abbreviations was provided.
2. Results & Discussion
2.1. Evaluation of Cellular Viability of Cell Lines Exposed to Stress Inducing Agents: Optimization of the Experimental Conditions
In previous published work, the safety of pure pCBs, CBD, CBC, CBG and pCBs-based formulations was evaluated following its incubation in HaCat and HDF cell lines. The cell viability obtained was above 90% for all the pure pCBs and pCBs formulations. These results demonstrated the safety of pCBs formulations to be used in skin disorders associated with inflammation and cellular homeostasis [32,33].
For each inducing agent different concentrations were tested and the respective cell viability after incubation in the cell lines under study was evaluated. These preliminary assays allowed to select the concentration of the inducing agent that would result in a loss of cell viability below 20%.
2.1.1. H2O2
H2O2 is an oxidizing agent that induces cellular damage at inappropriate concentrations [34,35]. It has been extensively used in research to induce cellular oxidative stress aiming to modulate redox-regulated cellular pathways [36]. Its role as both a signaling molecule and a source of oxidative stress makes it crucial in understanding various skin conditions, their progression, and potential treatments. Fibroblasts, the cells responsible for producing collagen and other ECM components, react to H₂O₂ during wound healing. In keratinocytes, H₂O₂ can induce oxidative damage that disrupts the skin barrier function.
In Figure 1 is presented the cell viability obtained for HaCat and HDF cell lines incubated for 24 h with H2O2. The concentrations of H2O2 ranged from 50 to 400 μM. The cell viability at a concentration of 50 μM was above 97%, at 100 μM higher than 90%, at 200 μM around 84% and at 400 μM higher than 71% for both skin cell lines tested.
Based on these results, 200 μM was the H2O2 concentration chosen for the evaluation of the molecular variations after stress induction, as a reduction on cell viability below 20%, for the cell lines under study, was achieved.
2.1.2. TPA
Phorbol 12-myrstate 13-acetate (TPA), plays a critical role in various cellular pathways, including cell proliferation, differentiation, apoptosis and has the capacity to generate reactive oxygen species [37]. TPA is a potent activator of protein kinase C (PKC) influencing multiple cellular processes, such as gene expression, modulation of the immune response and cell cycle regulation. TPA is also recognized as a cellular stress factor, inducing oxidative stress by generating reactive oxygen species (ROS) and activating stress-related signaling pathways [38,39].
TPA is used to induce inflammatory responses in cell cultures and animal models, enabling the study of inflammatory pathways to evaluate anti-inflammatory therapies.
Figure 2 depicts the results obtained for HaCat cell line incubated 24 h with TPA. The concentrations tested for TPA ranged from 5 to 20 μM. For a concentration of 5 μM cell viability above 93% was achieved, while for 10 μM the cell viability was superior to 84% for both cell lines. TPA at concentrations of 15 and 20 μM induced a HaCat cell reduction of 21 and 33%, respectively.
Based on these results, the concentration selected for the evaluation of the molecular variations after stress induction for HaCat was 10 μM, as a reduction on cell viability below 20% was achieved.
2.2. Evaluation of Gene Expression Markers in Cell Lines
The use of HaCaT and HDF cell lines in studies of cannabinoid formulations provides valuable insights into their effects on inflammation, regeneration, and wound healing. Keratinocytes are the predominant cell type in the epidermis and play a significant role in the skin's response to injury and inflammation. Cannabinoid research using HaCaT has shown that pCBs can exert anti-inflammatory effects by modulating cytokine production and promote wound healing by enhancing keratinocyte migration [1,20]. This migration is a vital step in the re-epithelialization process during wound repair, thus restoring the skin's barrier function. HDF cells are also relevant for studying wound healing and regeneration as pCBs can influence fibroblast activity, which is crucial for tissue repair and ECM remodeling which provide structural support to the skin, affecting processes such as cell proliferation, migration, and collagen production. For instance, pCBs have shown to promote fibroblast proliferation and migration, which are critical for wound closure and tissue repair modulate the balance between inflammation and regeneration, which are crucial for effective wound healing and minimizing scarring [40].
Using both HaCaT and HDF cell lines to model the complex interactions keratinocytes and fibroblasts allows us to better understand how pCBs-based formulations might modulate inflammation and regeneration highlighting their potential for cosmeceutical and/or therapeutic use in dermatology.
These cell lines express several key proteins and pathways involved in stress responses, inflammation, and tissue homeostasis. PKC is expressed in both HDF and HaCaT cells, playing a crucial role in signal transduction. In HaCaT cells, PKC activation, such as by phorbol esters like TPA, can stimulate the nuclear factor kappa B (NF-κB) pathway and activate Matrix Metalloproteinases (MMPs). The NF-κB pathway, active in both cell types, is a central mediator of inflammatory responses. Upon activation in HaCaT cells, NF-κB translocate to the nucleus, promoting the transcription of pro-inflammatory genes including Tumor necrosis factor alpha (TNF-α), interleukin 36G (IL-36G), and interleukin 6 (IL-6). Peroxisome Proliferator-Activated Receptor gamma (PPARγ) is expressed in both HDF and HaCaT cells. While its specific role in these cell lines requires further elucidation, PPARγ is known to be involved in regulating inflammation and maintaining skin homeostasis. MMPs are also expressed in both cell types, with their activation in HaCaT cells linked to PKC stimulation under conditions of chemical stress. pCBs, while not endogenously expressed, exert significant effects on both HDF and HaCaT cells. Recent studies have demonstrated that pCBs modulate inflammatory responses in HaCaT cells [22,41,42,43]. These effects are mediated through the ECS and signaling pathways. It is important to note that while both HDF and HaCaT cells express these proteins and pathways, their expression levels and responses to stimuli may differ due to their distinct origins and functions within the skin. This underscores the importance of considering cell type-specific responses when interpreting results from in vitro studies using these models in skin biology and inflammation research.
Figure 3 illustrates the complex interplay between oxidative stress, chemical stress, and the ECS in modulating gene expression pathways within keratinocytes and fibroblasts. Oxidative stress inducers, such as H₂O₂, lead to an overproduction of ROS that activate the NF-κB signaling pathway [44]. Once activated, NF-κB translocate to the cell nucleus, where it drives the transcription of pro-inflammatory genes, including TNF-α, IL-36G, and IL-6 [45]. Chemical stressors, such as TPA, activate PKC, which in turn stimulates the NF-κB pathway, resulting in the transcription of the same inflammatory genes as seen in the oxidative stress pathway [46]. Additionally, PKC activation triggers another signaling cascade that leads to the activation of MMPs. These zinc-dependent enzymes degrade components of the ECM, such as collagens and elastin (ELN), which can compromise ECM stability, tissue integrity, and cellular behavior [46]. Cannabinoid receptors, including CNR1, CNR2, and GPR55, play vital roles in regulating various skin physiological processes like inflammation, pain perception, cell proliferation, and differentiation [47]. CNR1 is linked with pain modulation, neuroprotection, and anti-inflammatory effects, CNR2 is primarily involved in anti-inflammatory and immunomodulatory functions and G protein-coupled receptor 55 (GPR55), sometimes referred to as the "third" cannabinoid receptor, influences inflammatory responses and cell proliferation. Although pCBs may not always directly activate these receptors, they can modulate receptor expression and thereby influence the ECS [30,47]. Activation of these receptors can lead to PPARγ pathway activation, that is crucial for anti-inflammatory responses, tissue repair, and ECM maintenance. One significant gene regulated by PPARγ is TIMP3, which inhibits MMPs, preventing excessive ECM degradation and promoting tissue homeostasis. The activation of cannabinoid receptors and the PPARγ pathway can inhibit NF-κB by various mechanisms, such as preventing NF-κB from translocating to the nucleus, interfering with its DNA binding, or modulating upstream signals that activate NF-κB [30,48].
Stressors & Stress-baseline response
H₂O₂ induces oxidative stress, which leads to cellular damage and triggers responses such as inflammation, apoptosis, or senescence [49,50,51]. TPA, on the other hand, activates signaling pathways, particularly through PKC [46], which plays a crucial role in processes like signal transduction, cell proliferation, differentiation, apoptosis, and immune responses, often resulting in inflammation and potential tumorigenesis [52,53,54].
Exposure to H₂O₂ and TPA typically stimulates an inflammatory response, increasing the production of pro-inflammatory cytokines such as IL-6, TNF-α, and IL-36G. IL-6 is essential during the acute phase of inflammation, while IL-36G is crucial in skin-related inflammatory responses and conditions like psoriasis. TNF-α promotes the recruitment of immune cells and enhances vascular permeability, further intensifying inflammation by stimulating other pro-inflammatory cytokines.
The regulation of anti-inflammatory markers like IL-10 and PPARγ is equally critical in managing damage and initiating repair. IL-10 reduces the expression of pro-inflammatory cytokines, limiting tissue damage and promoting healing. PPARγ plays a key role in controlling inflammation, regulating lipid metabolism, and promoting cellular differentiation and stress response. Both IL-10 and PPARγ are expected to be up-regulated by treatments aimed at reducing inflammation and aiding tissue repair.
2.2.1. Molecular Insights into the Pro-Inflammatory Modulation by F1CR1, F1CR2, F2CAA, and F3TAC Formulations in Pre-Treatment and Treatment Conditions
The effect of each one of the proprietary formulations on pro-inflammatory markers (IL-6, TNF-α and IL-36G), was assessed in HaCat and HDF cells following exposition to H2O2 and TPA, either in pre-treatment or treatment conditions (Figure 4). Detailed results can be found in supplementary material, “S1 – Effect of formulations on pro-inflammatory markers”.
F1CR1 Pre-Treatment: resulted in a significant downregulation of key pro-inflammatory cytokines (IL-6, TNF-α, and IL-36G) in both HaCaT and HDF cells (Figure 4 A). This effect was consistent across both oxidative (H₂O₂) and inflammatory (TPA) stress conditions, indicating the broad anti-inflammatory potential of F1CR1. These results suggest that F1CR1 effectively primes skin cells to withstand oxidative and inflammatory stress, making it highly protective against skin damage. By reducing the expression of these pro-inflammatory markers, F1CR1 may help prevent chronic inflammation and associated tissue damage.
F1CR1 Treatment: F1CR1 also demonstrated significant efficacy when applied post-stress (Figure 4 E). After oxidative or inflammatory stress, it reduced pro-inflammatory markers, suggesting its ability to control inflammation even after damage has occurred. This post-treatment effect indicates a potential therapeutic role for F1CR1 in managing ongoing skin inflammation, especially in conditions characterized by excessive oxidative stress or inflammation. Overall, F1CR1 modulates both inflammatory and regenerative processes in HaCat and HDF, offering protection and aiding recovery in various stress conditions.
F1CR2 Pre-Treatment: F1CR2 exhibited stress-dependent and cell-type-specific modulation of pro-inflammatory markers (Figure 4 B). In HaCaT cells IL-6 was upregulated, while TNF-α and IL-36G were downregulated, suggesting that F1CR2 helps modulate the inflammatory response and offers protection against oxidative stress. In HDF cells, a strong downregulation of TNF-α and an upregulation of IL-36G was observed, highlighting a tailored inflammatory response that supports tissue repair. Pre-treatment before TPA-induced inflammatory stress in HaCaT cells resulted in a strong downregulation of IL-6, TNF-α, and IL-36G, suggesting that F1CR2 helps control inflammation and prevents chronic skin damage under inflammatory conditions.
F1CR2 Treatment: the results varied depending on the type of stressor (Figure 4 F). After oxidative stress, both HaCaT and HDF cells showed an upregulation of IL-6 and downregulation of TNF-α, indicating that F1CR2 helps manage inflammation while promoting tissue regeneration. IL-36G was downregulated in HaCaT cells but upregulated in HDF cells, suggesting a role in supporting tissue repair, especially in HDF. However, when F1CR2 was applied post-TPA stress, there was an upregulation of IL-6, TNF-α, and IL-36G, indicating that F1CR2 may be less effective in controlling inflammation once the pro-inflammatory cascade is fully activated. This highlights the importance of timing when using F1CR2 for its anti-inflammatory and regenerative properties.
F2CAA Pre-Treatment: In HaCaT cells an upregulation of IL-6 and downregulation of TNF-α and IL-36G, suggesting a compensatory response to oxidative stress was achieved (Figure 4 C). In HDF cells, F2CAA led to a consistent downregulation of all three pro-inflammatory markers, particularly TNF-α, indicating strong anti-inflammatory potential in fibroblasts. In HaCaT, F2CAA caused significant downregulation of IL-6, TNF-α, and IL-36G, showing its ability to effectively prime the skin against inflammatory stressors.
F2CAA Treatment: After oxidative stress it was demonstrated an upregulation of IL-6 and a downregulation of TNF-α in both HaCaT and HDF cells, highlighting a selective modulation of inflammatory pathways, with stronger effects on HDF (Figure 4 G). After TPA-induced stress, F2CAA caused a slight upregulation of IL-6 and TNF-α but a strong downregulation of IL-36G, indicating its effectiveness in targeting pathways like IL-36G, which are linked to chronic skin inflammation.
Overall, F2CAA demonstrates strong anti-inflammatory potential in both pre-treatment and treatment scenarios, with its ability to modulate key cytokines making it a valuable agent in managing skin inflammation
F3TAC Pre-Treatment: In HaCaT cells, pre-treated before H₂O₂, a strong upregulation of IL-6 and downregulation of TNF-α and IL-36G, indicating complex modulation of the inflammatory response was observed (Figure 4 D). In HDF cells, pre-treated with F3TAC, all three pro-inflammatory cytokines (IL-6, TNF-α, IL-36G) were consistently downregulated, with TNF-α showing the most significant decrease. These results suggest that F3TAC has potent anti-inflammatory effects, particularly in HDF, where it significantly reduces inflammation under oxidative stress. Additionally, pre-treatment with F3TAC, before TPA-induced stress in HaCaT cells, resulted in a marked downregulation of IL-6, TNF-α, and IL-36G, highlighting F3TAC’s potential to prime keratinocytes against inflammatory damage.
F3TAC Treatment: F3TAC’s effects on pro-inflammatory markers were more limited (Figure 4 H). In HaCaT cells exposed to H₂O₂, a slight upregulation of IL-6 and downregulation of TNF-α and IL-36G, indicating that F3TAC can still modulate inflammation after oxidative damage, though less effectively than in pre-treatment scenarios was attained. In HDF cells, post-treatment with F3TAC led to an upregulation of IL-6 but strong downregulation of TNF-α, suggesting a continued anti-inflammatory effect in fibroblasts even after stress. After TPA-induced stress, F3TAC slightly upregulated IL-6 but strongly downregulated IL-36G, suggesting that it remains effective in modulating specific inflammatory pathways triggered by inflammatory stress.
The differential effects of F3TAC between pre-treatment and post-treatment scenarios highlight the importance of timing in modulating the inflammatory response in skin cells. Pre-treatment appears to prime cells to handle stress more effectively, while treatment shows F3TAC’s continued ability to manage inflammation and aid in tissue repair. These findings suggest that F3TAC has both protective and therapeutic properties depending on when it is administered.
2.2.2. Molecular Insights into the Anti-Inflammatory Effects of F1CR1, F1CR2, F2CAA, and F3TAC Formulations in Pre-Treatment and Treatment Conditions
The effect of each one of the proprietary formulations on anti-inflammatory markers (IL-10 and PPARγ), was assessed in HaCat and HDF cells following exposition to H2O2 and TPA, either in pre-treatment or treatment conditions (Figure 5). Detailed results can be found in supplementary material, “S2 – Effect of formulations on anti-inflammatory markers”.
F1CR1 Pre-Treatment: In HaCaT cells exposed to H₂O₂ stress, no significant changes were observed in IL-10 and PPARγ regulation, suggesting that F1CR1 does not significantly alter the anti-inflammatory response in keratinocytes under oxidative stress (Figure 5 A). However, in HDF cells, F1CR1 pre-treatment led to a notable downregulation of both IL-10 and PPARγ, indicating that F1CR1 may shift anti-inflammatory mechanisms in fibroblasts. When subjected to TPA stress, both HaCaT and HDF cells showed significant downregulation of IL-10 and PPARγ, suggesting that F1CR1’s effects vary based on cell type and stress conditions, potentially modulating inflammation through alternative pathways.
F1CR1 Treatment: In HaCaT cells after H₂O₂ stress, there was a downregulation of PPARγ, suggesting modulation of inflammatory responses and keratinocyte differentiation (Figure 5 E). In HDF cells, F1CR1 led to an upregulation of IL-10 and a strong downregulation of PPARγ, highlighting its anti-inflammatory potential and modulation of metabolic and inflammatory pathways in HDF. Following TPA stress, HaCaT cells showed a downregulation of IL-10 without significant changes in PPARγ, suggesting stress-specific responses and different compensatory mechanisms depending on the type of stress.
F1CR2 Pre-Treatment: H2O2 stress led to a strong downregulation of IL-10 and a significant upregulation of PPARγ in both HaCaT and HDF cells (Figure 5 B). In HaCaT cells, this suggests that F1CR2 prepares cells to handle oxidative stress by reducing the need for IL-10, potentially promoting stress tolerance and repair mechanisms. In HDF cells, F1CR2 reduced IL-10 dependency and enhanced PPARγ expression, indicating its role in regulating inflammation and promoting tissue repair.
F1CR2 Treatment: When F1CR2 was applied after H₂O₂ stress, no significant changes in IL-10 were observed in HaCaT cells, but PPARγ remained upregulated, indicating the continued activation of recovery pathways (Figure 5 F). In HDF cells, IL-10 was upregulated, suggesting an active role in managing inflammation and supporting tissue repair. The consistent upregulation of PPARγ in both cell lines highlights F1CR2’s role in promoting anti-inflammatory responses and aiding recovery from oxidative stress, particularly in fibroblasts.
F2CAA Pre-Treatment: In HaCaT cells exposed to H₂O₂ stress, IL-10 was slightly upregulated, suggesting enhanced anti-inflammatory responses, while PPARγ remained stable (Figure 5 C). In HDF cells, both IL-10 and PPARγ were downregulated, indicating a more targeted and cell-specific inflammation modulation. Under TPA stress, both IL-10 and PPARγ were downregulated in HaCaT cells, indicating stress-specific effects of F2CAA.
F2CAA Treatment: In HaCaT cells after H₂O₂ stress, there was a strong downregulation of both IL-10 and PPARγ, suggesting a more pronounced inflammatory modulation (Figure 5 G). In HDF cells, IL-10 was downregulated, but PPARγ was upregulated, indicating an anti-inflammatory response. After TPA stress, HaCaT cells exhibited an upregulation of both IL-10 and PPARγ, indicating a protective response against inflammation. These results show that F2CAA’s effects are cell-type and stress-specific, with differing impacts on anti-inflammatory mediators.
F3TAC Pre-Treatment: In HaCaT cells, regarding H₂O₂ stress, there was a slight upregulation of IL-10 with no significant changes in PPARγ, indicating a potential enhancement of anti-inflammatory responses in keratinocytes (Figure 5 D). However, in HDF cells, both IL-10 and PPARγ were downregulated, indicating cell-specific effects of F3TAC in modulating pro- and anti-inflammatory signals in fibroblasts. Pre-treatment with F3TAC before TPA stress led to significant downregulation of both IL-10 and PPARγ in HaCaT cells, suggesting F3TAC’s effects are stress-dependent and may involve other compensatory mechanisms.
F3TAC Treatment: Post-treatment with F3TAC in HaCaT cells after H₂O₂ stress resulted in slight but non-significant downregulation of IL-10 and PPARγ, suggesting limited effects on these markers after oxidative stress (Figure 5 H). In HDF cells, there was a consistent downregulation of IL-10 and PPARγ, indicating that F3TAC has a more pronounced effect on modulating anti-inflammatory pathways in fibroblasts. After TPA stress, HaCaT cells showed no significant changes in IL-10 or PPARγ, further indicating that F3TAC’s effects are context-dependent
2.2.3. Molecular Insights into the Structural Modulation by F1CR1, F1CR2, F2CAA, and F3TAC Formulations in Pre-Treatment and Treatment Conditions
The effect of each one of the proprietary formulations on structural markers (ELN, FN1, EGFR and TIMP3), was assessed in HaCat and HDF cells following exposition to H2O2 and TPA, either in pre-treatment or treatment conditions (Figure 6). Detailed results can be found in supplementary material, “S3 – Effect of formulations on structural markers”.
F1CR1 Pre-Treatment: In HaCaT cells, exposed to H₂O₂ stress, F1CR1 induced a strong downregulation of FN1 and an upregulation of both EGFR and TIMP3 (Figure 6 A). This suggests a role in modulating ECM production and promoting cell survival and proliferation under oxidative stress. In HDF cells, under the same conditions, F1CR1 led to the downregulation of ELN, FN1, and TIMP3, highlighting cell-specific responses, possibly aimed at preventing fibrosis. Under TPA-induced stress, HaCaT cells experienced downregulation of ELN, FN1, TIMP3, and EGFR, indicating a protective response against inflammation and excessive cell proliferation. F1CR1 may help prepare cells to better cope with stress by modulating genes involved in ECM production, cell proliferation, and tissue remodeling.
F1CR1 Treatment: In HaCaT cells, after H₂O₂ stress, the FN1 downregulation suggests modulation of ECM production, potentially preventing excessive fibrosis (Figure 6 E). The upregulation of EGFR points to the restoration of normal cell proliferation and differentiation, while the stronger upregulation of TIMP3 indicates enhanced ECM remodeling and protection against oxidative damage. In HDF cells, F1CR1 led to a slight upregulation of ELN, indicating a mild protective effect on skin elasticity, with a subtle downregulation of FN1, EGFR, and TIMP3, suggesting less pronounced remodeling activity. After TPA-induced stress, HaCaT cells showed no significant changes in ELN, TIMP3, and EGFR, but FN1 was upregulated, potentially reflecting an enhanced wound healing response to inflammation.
F1CR2 Pre-Treatment: In HaCaT cells exposed to H₂O₂ stress, there was a downregulation of ELN and FN1, suggesting a protective mechanism against oxidative damage and potential prevention of excessive fibrosis (Figure 6 B). The upregulation of EGFR and TIMP3 indicates enhanced cellular proliferation and ECM remodeling, aiding tissue repair. In HDF cells, F1CR2 pre-treatment led to upregulation of ELN and FN1, suggesting enhanced structural support and wound healing under stress.
F1CR2 Treatment: When F1CR2 was applied after H₂O₂ stress, HaCaT cells showed enhanced ECM production (upregulated FN1) and tissue regeneration (upregulated EGFR and TIMP3) (Figure 6 F). In HDF cells, there was a strong downregulation of ELN and FN1, possibly to prevent excessive scarring, with a slight upregulation of TIMP3 indicating controlled matrix remodeling. After TPA-induced stress, HaCaT cells demonstrated upregulation of ELN and TIMP3, with slight downregulation of EGFR, indicating a focus on structural support and matrix stabilization in response to inflammation.
F2CAA Pre-Treatment: In HaCaT cells, regarding H₂O₂ stress, a strong downregulation of FN1 and EGFR, along with a strong upregulation of TIMP3, suggesting a protective mechanism against oxidative stress by reducing ECM degradation and fibrosis was observed (Figure 6 C). In HDF cells, F2CAA pre-treatment led to a downregulation of ELN and FN1, with an upregulation of EGFR, indicating modulation of ECM production and cell proliferation in fibroblasts.
F2CAA Treatment: In post-treatment and after H₂O₂ stress, HaCaT cells showed downregulation of ELN, FN1, and EGFR, alongside strong upregulation of TIMP3, indicating protection against ECM degradation (Figure 6 G). In HDF cells, downregulation of FN1, ELN, and TIMP3 was observed, suggesting a complex modulation of ECM balance in fibroblasts under oxidative stress. After TPA stress, HaCaT cells exhibited slight upregulation of ELN, FN1, and TIMP3, with slight downregulation of EGFR, indicating tissue repair promotion and moderation of cell proliferation under inflammatory conditions.
F3TAC Pre-Treatment: In HaCaT cells, considering H₂O₂ stress, the upregulation of ELN suggests improved skin elasticity, while the strong downregulation of FN1 and EGFR indicates reduced cell proliferation and migration (Figure 6 D). The very strong upregulation of TIMP3 suggests enhanced protection against ECM degradation. In HDF cells pre-treated with F3TAC under H₂O₂ stress, downregulation of ELN, FN1, and TIMP3 indicates reduced ECM production and remodeling, with stable EGFR levels suggesting maintained cell proliferation.
F3TAC Treatment: When HaCaT cells were treated with F3TAC after H₂O₂ stress, FN1 remained stable, suggesting preserved ECM production (Figure 6 H). The strong downregulation of EGFR indicates reduced cell proliferation, while the strong upregulation of TIMP3 points to enhanced matrix protection. In HDF cells treated after H₂O₂ stress, downregulation of ELN, FN1, and TIMP3 suggests reduced matrix production and remodeling, with stable EGFR levels maintaining cell proliferation. After TPA stress, HaCaT cells treated with F3TAC showed upregulation of ELN, TIMP3, and slight upregulation of FN1, indicating enhanced matrix production and protection, while EGFR remained unchanged, reflecting stable cell proliferation capacity.
2.2.4. Molecular Evaluation of Cannabinoid Receptor Modulation by F1CR1, F1CR2, F2CAA, and F3TAC Formulations in Pre-Treatment and Treatment Scenarios
The effect of each one of the proprietary formulations on cannabinoid receptor markers (GPR55, CNR2 and CNR1), was assessed in HaCat and HDF cells following exposition to H2O2 and TPA, either in pre-treatment or treatment conditions (Figure 7). Detailed results can be found in supplementary material, “S4 – Effect of the formulations on cannabinoid receptor markers”.
F1CR1 Pre-Treatment: Exposed to H₂O₂ stress, GPR55 and CNR2 were downregulated in HaCat cells, suggesting potential anti-inflammatory actions, while CNR1 remained unchanged (Figure 7 A). In contrast, HDF cells showed an upregulation of GPR55 and downregulation of CNR2, highlighting cell-specific responses, with GPR55 potentially aiding wound healing in fibroblasts. Under TPA-induced stress, HaCaT cells experienced upregulation of GPR55 and downregulation of CNR2, reinforcing the stress-specific nature of F1CR1’s effects.
F1CR1 -Treatment: After H₂O₂ treatment, F1CR1 upregulated CNR2 and downregulated CNR1 in HaCaT cells, indicating modulation of HaCat inflammation, while HDF showed mild upregulation of GPR55 and CNR1 (Figure 7 E). Post-TPA treatment, all three receptors—GPR55, CNR2, and CNR1—were upregulated, suggesting F1CR1 activates the ECS to reduce inflammation and support regeneration under inflammatory stress. These results highlight the complex and context-dependent effects of F1CR1 on the ECS in skin cells.
F1CR2 Pre-Treatment: The application of F1CR2 prior to H₂O₂-induced oxidative stress in HaCaT cells resulted in the downregulation of GPR55 and CNR2, accompanied by an upregulation of CNR1 (Figure 7 B). The downregulation of GPR55 suggests that F1CR2 may be attenuating pro-inflammatory effects, while the upregulation of CNR1 points to enhanced protective mechanisms against oxidative stress. In HDF cells, F1CR2 led to a strong upregulation of GPR55 and CNR2, with downregulation of CNR1, indicating activation of pathways involved in wound healing and tissue remodeling.
F1CR2 -Treatment: In HaCaT cells, after H₂O₂-induced stress, CNR1 remained upregulated, but CNR2 and GPR55 were downregulated, indicating a reduced pro-inflammatory response (Figure 7 F). In HDF cells, GPR55 and CNR2 were upregulated, promoting regenerative and anti-inflammatory responses, while CNR1 was downregulated, potentially indicating a shift towards cellular repair. This highlights that the timing of F1CR2 application is crucial for maximizing its anti-inflammatory and regenerative benefits.
F2CAA Pre-Treatment: In HaCaT cells, and before H₂O₂ stress, GPR55 was downregulated while CNR1 and CNR2 were upregulated, indicating a protective, anti-inflammatory effect (Figure 7 C). In HDF cells, F2CAA pre-treatment also downregulated GPR55 and CNR2, with CNR1 upregulated, suggesting cell-specific protective responses. Under TPA stress, F2CAA caused GPR55 upregulation and CNR2 downregulation in HaCaT cells, pointing to a modulation of inflammatory pathways depending on the stress type.
F2CAA Treatment: Treatment with F2CAA, after H₂O₂ stress, led to the downregulation of all three receptors in HaCaT cells, suggesting a focus on reducing oxidative stress effects (Figure 7 G). In HDF cells, only GPR55 was slightly downregulated, indicating a milder modulation. After TPA stress, F2CAA caused upregulation of all three receptors, showing enhanced cannabinoid signaling and tissue repair post-inflammation. These results emphasize the importance of timing when administering F2CAA, as it appears to provide both protective and regenerative benefits depending on when it is applied.
F3TAC Pre-Treatment: In HaCaT cells, and before H₂O₂ stress, GPR55 and CNR2 were slightly upregulated, and CNR1 was strongly upregulated, suggesting protection against oxidative stress through anti-inflammatory and antioxidant effects (Figure 7 D). In HDF cells, F3TAC pre-treatment led to slight upregulation of GPR55 and CNR1, with downregulation of CNR2, indicating cell-specific protective mechanisms in the dermal layer. Under TPA stress, HaCaT cells pre-treated with F3TAC showed upregulation of GPR55, downregulation of CNR2, and unchanged CNR1, reflecting modulation of stress-specific inflammatory pathways.
F3TAC Treatment: In HaCaT cells, and after H₂O₂ stress, CNR2 was significantly downregulated, GPR55 was slightly upregulated, and CNR1 remained unchanged, indicating a shift in inflammatory modulation and regeneration post-stress (Figure 7 H). In HDF cells, GPR55 was downregulated, while CNR1 and CNR2 were strongly upregulated, enhancing cannabinoid signaling and promoting anti-inflammatory and regenerative responses. Post-TPA treatment in HaCaT cells led to significant upregulation of all three receptors, suggesting enhanced cannabinoid signaling for repair and recovery from inflammatory stress.
The most noticeable results are indicated in Table 1, highlighting the potential pathways involved in modulation of pro-inflammatory, anti-inflammatory and cannabinoid receptor markers and ECM regulation suggested from the in vitro assays.
The observed effects across different cell lines suggest that F1CR1, F1CR2, F2CAA and F3TAC formulations may influence cell-specific and stress-dependent responses, highlighting their capacity to modulate inflammatory and regenerative processes in a controlled environment. These findings point to possible mechanisms that could be further explored for their therapeutic relevance.
2.3. In-Vivo Efficacy Assays in Human Volunteers: A Proof of Concept
The regenerator cream, formulated with the F1CR1 combination, demonstrated significant efficacy in promoting skin recovery and regeneration across multiple dimensions. This aligns with the previously discussed properties of F1CR1, which include potent anti-inflammatory effects, enhancement of cellular repair mechanisms, and modulation of key structural and inflammatory pathways.
The application of the F1CR1-associated regenerator cream for 28 days significantly accelerated the reduction of skin redness, decreasing the recovery time by 2.7 days (12%, p<0.05) compared to the control. This outcome is consistent with F1CR1's ability to downregulate pro-inflammatory cytokines, such as TNF-α, which are responsible for prolonged inflammation and redness in the skin. The cream also effectively impacted microcirculation, reducing recovery time by 1.9 days (6.6%, p<0.05) compared to the control. This improvement could be due to F1CR2’s influence on the ECS, particularly through its modulation of cannabinoid receptors, which play a role in maintaining microcirculation and reducing vascular inflammation. The cream enhanced skin's barrier function, shortening the recovery time by 2.7 days (11%, p<0.05) versus the control. The cream exhibited excellent skin compatibility and acceptability, with no adverse reactions reported throughout the study, reflecting the formulation’s safety and tolerability.
Subjective efficacy results at the study's conclusion were also positive. 79% of participants reported softer, more hydrated skin with an overall improved appearance, likely due to F1CR1’s role in promoting skin hydration and repair. 64% of subjects agreed that their skin was effectively repaired, with 36% stating complete skin reparation, underscoring F1CR1’s ability to enhance the skin's natural healing processes. 93% observed a noticeable reduction in skin redness, consistent with the anti-inflammatory properties of F1CR1. In terms of cosmetic qualities, the cream was well-received by all participants. 100% of subjects appreciated the cream's texture, agreeing that it spread and absorbed easily, which likely contributed to the overall positive user experience and efficacy in daily use.
The application of F1CR2 for 28 days resulted in a reduction in erythema recovery time by 2.2 days (9.5%) compared to the control. The anti-inflammatory properties observed in pre-treated HaCaT and HDF support this result. Erythema, often accompanied by increased blood flow, was similarly affected, with F1CR2 reducing blood flow recovery time by 0.8 days (1.6%) compared to the control. The slight reduction in blood flow recovery time may be linked to F1CR2's ability to modulate the skin's ECS, particularly through the upregulation of CNR1 and CNR2 receptors. This modulation could help improve microcirculation and vascular function, leading to faster recovery from increased blood flow associated with erythema. The use of F1CR2 reduced barrier recovery time by 1.4 days (5.4%) when compared to the control. F1CR2's impact on ECM components and structural markers such as TIMP3 and FN1, as seen in vitro, likely contributes to its efficacy in enhancing skin barrier recovery.
No adverse reactions were reported during the study, indicating that F1CR2 exhibited excellent skin compatibility and acceptability. Regarding the subjective efficacy of F1CR2, 79% of participants agreed that the appearance of their skin had improved, 86% stated that their skin felt soft and hydrated, 86% agreed that their skin was repaired, with 50% noting full repair, 93% observed a reduction in skin redness. In terms of cosmetic qualities, 100% of participants liked the texture of F1CR2, and 92% agreed that it was easily absorbed and spread on the skin.
The application of the F2CAA cream resulted in a modest reduction across various skin spot metrics, including a 3.6% decrease in the count of visible spots, a 5.6% decrease in their area, a 4.3% reduction in UV spot area, and a 3.7% decrease in red spot count. The antioxidant capacity of the cream was evaluated by inducing an oxidation reaction with UVA radiation. Using β-carotene, a yellow chromophore that loses its color upon oxidation, it was observed that the untreated control area exhibited a 53.6% greater color variation compared to the area treated with the cream. This statistically significant result (p<0.05) indicates that the cream effectively mitigates oxidation under UVA exposure.
The cream demonstrated a statistically significant reduction (p<0.05) in the count of wrinkles and fine lines by 21.2%. Additionally, the application of the cream significantly improved (p<0.05) skin firmness by 11.9% and significantly decreased (p<0.05) skin roughness, while a non-significant increase of 1.2% in skin elasticity was observed. The efficacy of the cream in reducing wrinkles was further corroborated through clinical evaluation using the crow’s feet clinical score. After 28 days of application, a significant reduction (p<0.05) of 9.5% in the crow’s feet score was recorded, highlighting the cream’s effectiveness in diminishing the appearance of wrinkles.
Post-treatment analysis revealed a statistically significant decrease of 21.4% in the maximum microcirculation value and a significant 40.6% increase in the onset time to vasodilation. These findings suggest that the cream effectively reduces vasodilation and delays its onset when induced by histamine, thereby demonstrating its soothing properties. These outcomes suggest that the cream not only reduces the extent of vasodilation—a key marker of inflammation—but also delays its onset, indicating a soothing effect on the skin. The study also reported a statistically significant 19.1% reduction in Trans Epidermal Water Loss (TEWL), indicating an enhancement of the skin barrier function. In terms of hydration, the treatment led to a statistically significant 32.3% increase in skin hydration, further substantiating the cream’s efficacy in improving both hydration and skin barrier integrity. The reduction in TEWL and the improvement in skin barrier function further support the anti-inflammatory conclusion, as a well-functioning skin barrier is essential for protecting against environmental stressors and reducing inflammatory responses.
No adverse reactions were documented throughout the study, confirming the cream's excellent skin compatibility and acceptability.
Regarding the subjective efficacy of the cream, 100% of participants reported that their skin felt softer, 86% noted immediate hydration, 93% observed that their skin appeared hydrated, glowing, and radiant, 86% felt their skin was firmer, 64% noted a reduction in the appearance of lines and wrinkles, 86% reported that their skin felt restored and rejuvenated, 71% agreed that their skin looked and felt plumped and youthful, 86% felt their skin was nourished, 93% agreed that their skin felt energized, brighter, and that its texture was improved and more even, 50% observed a reduction in hyperpigmentation, 86% noticed a reduction in pore size, and 64% reported a reduction in skin redness.
Regarding the cosmetic formulation, 100% of participants expressed satisfaction with the texture and found the product easy to absorb and spread.
In conclusion, these findings suggest that the F2CAA cream is effective in improving the appearance of aging skin, provides antioxidant protection, and exhibits soothing effects after 28 days of consistent use.
For all the creams, no adverse reactions were reported during the study. This aligns with the in vitro findings for which, each formulation demonstrated a capacity to modulate inflammation without triggering excessive pro-inflammatory responses. The high percentage of participants reporting skin repair, hydration, and improved appearance can be attributed to the formulation role in upregulating key regenerative markers. These factors contribute to enhanced tissue repair, collagen synthesis, and overall skin health, as seen in both HaCat and HDF. The reduction in skin redness reported by participants directly correlates with anti-inflammatory properties demonstrated in vitro.
Regarding the application of F3TAC, there was an increase in the count and area of red spots, a slight decrease in the count of visible spots accompanied by an increase in their area, and an increase in the lipidic index, suggesting that the anti-acne tonic may not be performing as expected in reducing signs of acne. A possible explanation for these outcomes can be that sometimes, topical treatments, especially those targeting acne, can trigger an initial purging phase. This is a period where the skin begins to expel impurities from beneath the surface, leading to a temporary increase in spots and redness. The increase in red spots and their area could be indicative of this purging process, where the skin is reacting to the active ingredients in the tonic by bringing underlying issues to the surface.
Moreover, a positive outcome was that there was a 10.4% decrease in both porphyrins count and area. Porphyrins are produced by bacteria such as Cutibacterium acnes, and known as virulence factor involved in the pathogenesis of acne [55,56]. High levels are associated with increased acne severity, whereas a reduction in porphyrins correlates with acne improvement [57].
Regarding hydration and skin barrier function, the product demonstrated a 12.9% increase in skin hydration and a 20.7% increase in TEWL
Subjective assessments of the tonic’s efficacy revealed that 67% of participants agreed that their skin appeared brighter, felt protected, and that skin irritation was reduced with regulated sebum secretion, 83% agreed that oil production was controlled and skin texture improved, 75% reported a reduction in blemishes and breakouts,67% observed a reduction in the size and color of blemishes, with faster disappearance, 58% felt their skin was more balanced and noted improvement as early as the fifth day of use, 41% agreed that their skin felt more hydrated, 75% felt their skin was calm, soothed, and non-greasy after application, 41% noticed a reduction in hyperpigmentation, 58% saw a reduction in scarring associated with blemishes or acne, 50% agreed that pore size and skin redness were reduced. Regarding the cosmetic formulation, 92% of participants liked the texture and 83% agreed that the product absorbed easily.
No adverse reactions were reported during the study, confirming that the tonic exhibited good skin compatibility and acceptability.
In conclusion, while the product produced changes across all measured parameters, these differences were not statistically significant, the results indicate a trend towards a reduction in pore appearance and porphyrins, aligning with the intended purpose of an anti-acne tonic.
To enhance the efficacy of the tonic, several strategies could be considered, focusing on both formulation improvements and complementary approaches, by increasing the concentration of key active ingredients (within safe limits), can be combined with F1CR2 and / or tested during more time. The efficacy of acne treatments should typically be monitored over a period of 8 to 12 weeks as the skin cycle, or the time it takes for new skin cells to form and reach the surface, is about 4 to 6 weeks. Monitoring for at least two full skin cycles allows for a more accurate assessment of how well the treatment is working. Most topical acne treatments require several weeks to start showing noticeable effects. Initial improvements in acne lesions, reduction in inflammation, and overall skin texture often become evident after 4 to 8 weeks of consistent use.
This work confirmed the effectiveness and safety of the pCBs-based formulations for skin applications, highlighting their potential to improve its regeneration, and enhance structural processes. This research further underscores the significant promise of cannabis-based formulations in both the cosmetic and dermatological industries, offering substantial value to consumers seeking advanced skincare solutions. With increasing consumer demand for natural and effective products, cannabis-based products are poised to attract growing interest from both the public and industries, potentially reshaping the landscape of modern skincare and therapeutic applications.
4. Materials and Methods
4.1. Materials
Phosphate Phosphate buffered saline (PBS), 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and hydrogen peroxidase (H2O2) were obtained from Sigma-Aldrich (St. Louis, USA). TRK Lysis Buffer was acquired from VWR Life Sciences (PA, USA). The culture media, fetal bovine serum (FBS), antibiotics and phorbol 12-myrstate 13-acetate (TPA) were purchased from Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA). All other chemicals and reagents were of analytical grade.
4.2. Cell Lines Culture Conditions
Human keratinocytes (HaCaT) (CLS 300493) and human dermal fibroblasts (HDF) (ATCC® PCS-201-012™) were maintained in Dulbecco’s Modified Eagle’s medium (DMEM) with high-glucose (4,500 mg/L), supplemented with 10% FBS and 100 IU/mL of penicillin and 100 μg/mL streptomycin, henceforth designated as complete culture medium. All cells were maintained at 37°C in a 5% CO2 environment and medium were changed, and cells were tripsinized every 3-4 days when 80% confluence was reached.
4.3. Evaluation of Cellular Viability of Cell Lines Exposed to Stress Inducing Agents
MTT assay was used to evaluate the cell viability of HaCat and HDF in the presence of increasing concentration of two stress inducers, namely H2O2 and TPA. Briefly, cells were seeded at a concentration of 2x104 cells/mL (200 μL) in 96-well plates and allowed to adhere for 24 h. On the next day, the medium was removed, and cells were incubated with stress inducers for 24 h at 37°C in a humidified air incubator under a 5 % CO2 atmosphere. After that, culture medium was removed, and cells were washed twice with PBS. Then, 50 μL of a MTT solution at 0.5 mg/mL in incomplete medium was added to the cells and allowed to incubate for 2-4 h. Later, 200 μL of DMSO was added to each well in order to dissolve the formazan crystals produced by the cells upon MTT reduction. The absorbance was then read at 570 nm using a BioTek ELx800 Absorbance Microplate Reader (BioTek Instruments, Inc., Winooski, VT, USA) and the cells viability (%) calculated according to equation:
where ODt is the optical density of the cells from testing groups and ODc the optical density of control cells (cells in complete medium corresponding to 100 % cell viability).
Table 2 describes the range of concentrations for the stress inducers tested for each cell line.
4.4. Formulation Development and Analysis
pCBs isolated from Cannabis sativa provided the source for 4 proprietary formulations F1CR1, F1CR2, F2CAA, F3TAC. CBD was obtained from BSPG (https://www.bspglab.co.uk/), CBG, and CBC from ecsbrands (https://ecsbrands.com/isolates/). Also, purified CBG and CBC samples (>99.5%) were obtained by Centrifuge Partition Chromatography (CPC) technology using an rCPC device (RotaChrom, Purified Solutions, Budapest, Hungary). The method was internally developed through optimizing the best solvent combination alongside with the most appropriate solvent ratio (proprietary data). Briefly, CPC is a liquid–liquid preparative chromatographic technique that makes use of two immiscible liquid phases, the solvent system, representing the stationary and mobile phases of a typical chromatographic apparatus. The main goal of this technology is to isolate specific compounds with a high purity grade (>95%) and high recovery mass yield (>85%). Formulations were kept in dark amber glass flasks at room temperature. The cannabinoids and formulations were analyzed by high-performance liquid chromatography (HPLC, Cannabis Analyzer™ for Potency, Shimadzu).
The 4 proprietary formulations were designed with the potential for anti-inflammatory and regenerative actions (F1CR1, F1CR2, F2CAA) and additionally for anti-acne (F3TA). The formulations contain different ratios between each one of the cannabinoids.
The cosmetic matrices used in this study were expertly produced by a specialized cosmetic company (COSMETEK - Cosmetic Lab & Consulting, https://www.cosmetek.org/sobre-nos/) ensuring high-quality and standardized matrices.
4.5. Evaluation of pCBs Effect in Pre-Treatment and Treatment Conditions
The effect of pCBs-based formulations on gene expression in the studied cell lines, HaCat and HDF, was evaluated under both prophylactic (pre-treatment) and therapeutic (treatment). In these two approaches, stress was induced either after (pre-treatment) or before (treatment) incubation with the pCBs formulations. The molecular response under all experimental stress conditions was assessed by measuring the expression of relevant genes using qRT-PCR.
For the gene expression assay, cells were seeded at a concentration of 1.7x105 cells/mL (3 mL) in 6-well plates and allowed to adhere for 24 h.
- Pre-treatment effect
Cells were allowed to adhere for 24 h. After this period, the supernatant was removed and pCBs formulations solubilized in complete medium were added [32,33]. Complete medium was also added to negative and positive controls. After 24 h medium was discarded in all wells and stress inducer solution was added in tested wells and positive control and complete medium in negative control. In Figure 8 a scheme of the protocol. is represented.
- Treatment effect
Cells were allowed to adhere for 24 h. After this period, the medium was discarded, and cells were incubated with stress inducer at the pre-selected concentrations or complete medium (negative control). After a 24 h incubation period, medium was discarded and cells were incubated with pCBs formulations [32,33] or with complete medium either for negative (control cells) or positive controls (cells incubated with stress inducers).
After 24 h the supernatant was removed, and cells were washed with PBS twice to remove cellular debris. Cells were removed from each well, collected in pairs and centrifuged at 1000 g, for 20 min. at 4 °C. The pellet was then lysated with TRK lysin buffer and 2-Mercaptoethanol was added to inhibit RNase activity.
The concentrations of the stress inducing agents were chosen based on previous cell viability results performed in 96 well plates. Therefore, the concentration of H2O2 used in HaCat and HDF cells was 1700 μM. The stress inducer TPA was tested in HaCat cell line at a concentration of 85 μM.
4.6. Real-Time Polymerase Chain Reaction (RT-qPCR)
The gene expression level for each of the genes selected as a molecular marker was determined by real-time quantitative PCR using TaqMan probes (Thermo Scientific) (Figure 9). The quantitative real-time PCR reaction allowed the quantification of the gene expression of the genes of interest. This method combines PCR amplification technology with real-time product detection and quantification using fluorophores. The advantage of this technique is that it relies on the amount of product formed in the exponential phase, where the DNA amplification reaction conditions are optimal with a strong correlation between the threshold and the initial amount of DNA in the test sample.
Total RNA was extracted from different cell lines and exposed to several compounds and pCBs-based formulations, using a commercial kit according to manufacturer’s instruction (Thermo Scientific), and after extraction, the mRNA was quantified with a NanoDrop. cDNA synthesis was performed using a commercial kit (Thermo Scientific), the isolated mRNA was reverse transcribed in a final volume of 20 µL, following incubation in a thermocycler for 5 min at 22◦C, 30 min at 42 °C and 5 min at 85 °C. RT-qPCR was performed using the PerfeCTa qPCR FastMix II Low ROX in an QuantStudio™ 5 Real-Time PCR System. Each PCR reaction (20 µL final volume) was carried out with 5 µL of cDNA, 0.4 µL of primers Forward + Reverse (200 nM mix), 0.4 µL probe (200 nM). The qPCR reaction was performed with the following conditions: holding at 95 °C for 5 min, cycling at 95 °C for 30 s and 54 °C for 1 min (for 45 cycles). The relative expression of the different amplicons was calculated using the ∆∆Ct method and then converted into the relative expression ratio (2−∆∆Ct) and log2(2−∆∆Ct) for graphic elaboration. All data were normalized to the endogenous reference gene rRNA18S.
4.7. In Vivo Efficacy and Safety Studies
The efficacy of the formulations F1CR2, F1CR1, F2CAA and F3TAC was assessed through the development of two regenerators and one anti-aging creams and an anti-acne tonic. Each product was infused with pCBS as primary active ingredients. To enhance their effectiveness, additional active ingredients were selected based on their specific benefits. For acne treatment, salicylic acid was incorporated at low concentrations for its well-known acne-control properties. Essential oils, including lavender, juniper, and tea tree oil, were added selectively to the creams for their antimicrobial and soothing effects.
A single center, blinded, randomized, and controlled study was conducted to evaluate the efficacy, compatibility, and acceptability of the creams and tonic. This study, carried out by PhdTrials (https://phdtrials.com/) in accordance with Good Clinical Practices, also evaluated the cosmetic qualities and subjective efficacy of the three products.
To assess skin recovery and regeneration quantitatively, a panel of 14 subjects (comprising both genders, aged 18-35) participated in the study. Controlled skin irritation was induced using Sodium Lauryl Sulphate (SLS) exposure, followed by the daily application of the regenerator cream for 28 days. The study included a known skin regenerator as a positive control and no product application as a negative control for comparative purposes. Objective measurements of skin regeneration efficacy were conducted using instrumental methods to evaluate changes in skin color, barrier function, and microcirculation at various points before, during, and after the treatment period.
To evaluate anti-aging properties, the study involved the daily application of the cream for 28 days on a group of 14 women aged 50-65 years. The product’s revitalizing efficacy was objectively and quantitatively assessed using various instrumental measurements, including UV spots, visible spots, red spots, skin roughness, wrinkles, skin barrier function, firmness, elasticity, hydration, and microcirculation. Clinical scores were also evaluated by a dermatologist before, during, and after treatment.
To evaluate the efficacy for acne prone skins, the study consisted in applicating the tonic daily for 28 days, on a panel of 12 subjects of both genders with the following characteristics: i) 18-25 years old; ii) mixed and oily skin; iii) presence of comedones. The efficacy was evaluated objectively and quantitatively, by instrumental measurements of the skin lipidic index, hydration, skin barrier function, porphyrines, pores, visible spots and red spots.
Instrumental efficacy data were analyzed using the Student's t-test for continuous data, while subjective efficacy data were analyzed with the Binomial test. Statistical analyses were performed using SPSS 23 (IBM) with a 95% significance level. The acceptability and compatibility were controlled by visual examination of the experimental area by dermatologists and by the daily registry of any discomfort by the subjects. The subjective cosmetic qualities and efficacy were assessed at the end of the study using a target questionnaire. The subjective data of efficacy was submitted to a Binomial test for statistical treatment. Participants also provided feedback on the cosmetic qualities and perceived efficacy of the cream through a targeted questionnaire.
5. Conclusions
The study investigates the potential of pCBs for skin health applications, focusing on their ability to modulate skin ECS, inflammation and structural processes, demonstrating their potential for cosmeceutical applications and treatment of various skin diseases The pCBs CBD, CBG, and CBC are known to interact with the ECS, influencing various biological processes critical to skin health. However, the specific effects of these cannabinoids, both individually and in combination, on skin cells under stress conditions remain underexplored. This study aimed to compare the effects of four distinct cannabinoid-based formulations on key parameters involved in inflammation, regeneration, and homeostasis of skin. The tested formulations included F1CR2 (CBD + CBG), F1CR1 (CBD + CBC), F2CAA (CBD + CBG + CBC), and F3TAC (CBC + CBG) and were evaluated in both pre-treatment and treatment scenarios in human HaCaT and HDF cells, focusing on their modulation of pro-inflammatory and anti-inflammatory cytokines, cannabinoid receptor expression, and structural gene regulation.
Our study is highly innovative and represents the first of its kind to combine a deep molecular approach with in vivo efficacy assessments, focusing on the therapeutic potential of pCBs for skin health applications. By exploring the interaction of key cannabinoids—CBD, CBG, and CBC—with the skin's ECS, we provide new insights into how these compounds modulate inflammation, regeneration, and structural processes. The study offers a comprehensive comparison of four distinct cannabinoid-based formulations, evaluating their effects both in pre-treatment and treatment scenarios. This integrated approach not only enhances our understanding of pCBs in cosmeceutical applications but also opens new avenues for treating various skin diseases through targeted cannabinoid therapies.
Key Findings
F1CR1 holds significant potential as a treatment for skin inflammation by modulating key inflammatory mediators in skin cells exposed to oxidative stress. Its ability to reduce pro-inflammatory cytokine expression in both keratinocytes and fibroblasts indicates a broad-spectrum anti-inflammatory effect, making it useful for various dermatological applications. F1CR1 shows strong anti-inflammatory properties, particularly when used as a pre-treatment, and influences the expression of anti-inflammatory markers (IL-10, PPARγ) differently in keratinocytes and fibroblasts, suggesting cell type-specific effects. F1CR1 also affects cannabinoid receptors in a cell type-specific manner, potentially enhancing protective and regenerative responses. The downregulation of pro-inflammatory cytokines may primarily be driven by CBD’s anti-inflammatory properties, with CBC potentially enhancing these effects. Structural gene modulation, such as ELN and FN1, might also be influenced by both pCBs, and the differential responses observed in keratinocytes and fibroblasts could be related to varying cannabinoid receptor expression. Also, F1CR1 impacts the ECM by downregulating FN1 under oxidative stress, indicating its potential to prevent fibrosis, particularly in the treatment of hypertrophic scars. Additionally, its upregulation of EGFR and TIMP3 supports cell proliferation and matrix remodeling, which are vital for wound healing and skin regeneration. The combination of CBD and CBC in F1CR1 formulations provides a more comprehensive approach to addressing complex skin conditions involving both inflammatory and regenerative processes, making it more effective than single-compound treatments. This positions F1CR1 as a versatile option for managing both inflammation and structural damage in skin conditions.
F1CR2 plays a key role in modulating the inflammatory response in skin cells, striking a balance between immediate immune activation and long-term tissue repair. Its effects are tailored to specific skin cell types, making it a promising therapeutic agent for reducing inflammation and promoting regeneration after oxidative stress. F1CR2 is more effective in downregulating pro-inflammatory genes following H₂O₂-induced stress, which supports tissue repair and inflammation resolution. Conversely, under TPA-induced stress, F1CR2 faces a more persistent pro-inflammatory environment that is harder to reverse, though it still offers benefits in mitigating inflammation. F1CR2 modulates cannabinoid receptors differently in keratinocytes and fibroblasts, suggesting a coordinated cellular response designed to protect against oxidative damage and fine-tune inflammation. Its combination of CBD and CBG likely enhances therapeutic effects compared to single-compound formulations, with CBD acting through its anti-inflammatory effects on NF-κB pathways and CBG modulating additional inflammatory mediators. This synergy influences key structural genes like ELN and FN1, which are crucial for skin cell proliferation and differentiation. CBD’s interaction with CB2 receptors, along with CBG's partial agonist activity at both CB1 and CB2, suggests that F1CR2 engages the ECS comprehensively in skin cells. In keratinocytes, F1CR2 increases IL-6 levels while decreasing TNF-α and IL-36G, showing its ability to balance oxidative stress response and inflammation control. This regulation helps manage conditions where inflammation must be contained without compromising tissue repair. The downregulation of TNF-α in both keratinocytes and fibroblasts further underscores F1CR2’s capacity to reduce chronic inflammation, prevent tissue damage, and support healing. Its ability to upregulate PPARγ across both cell types enhances metabolic and regenerative functions, critical for treating chronic wounds. Moreover, F1CR2’s modulation of structural genes such as ELN, FN1, EGFR, and TIMP3 under oxidative stress points to its potential in preventing fibrosis, promoting ECM remodeling, and restoring skin elasticity. The formulation also differentially modulates cannabinoid receptors across cell types: in keratinocytes, it downregulates GPR55 and CNR2 while upregulating CNR1, providing protection against oxidative stress and reducing inflammation. In fibroblasts, however, F1CR2 upregulates GPR55 and CNR2 while downregulating CNR1, indicating a shift toward tissue remodeling and wound healing. In summary, F1CR2 demonstrates a versatile approach to managing skin inflammation and supporting regeneration. Its tailored effects across different cell types, modulation of key genes, and cannabinoid receptor activity make it a promising therapeutic option for various skin conditions characterized by inflammation, oxidative stress, and structural damage.
F2CAA exhibits a comprehensive and multifaceted modulation of inflammation, cannabinoid receptors, and structural genes in skin cells under various stress conditions. This formulation, which combines CBD, CBG, and CBC, harnesses the unique properties of each cannabinoid, contributing to its wide-ranging effects on skin health. F2CAA's anti-inflammatory effects are evident through its downregulation of pro-inflammatory cytokines such as TNF-α, a key driver of chronic inflammation. The formulation also upregulates IL-10 in keratinocytes, highlighting its dual role in suppressing harmful inflammation while promoting anti-inflammatory signaling. This positions F2CAA as an effective treatment for inflammatory skin conditions. Additionally, the upregulation of TIMP3, which helps maintain the ECM and prevent its degradation, underscores F2CAA's regenerative properties, especially beneficial for aging skin, chronic wounds, and preventing fibrosis. F2CAA also modulates crucial structural genes, including FN1 and ELN, which play vital roles in preserving skin elasticity, promoting wound healing, and preventing scar formation. Moreover, F2CAA’s influence on cannabinoid receptors, particularly its reduction of GPR55 and increase in CNR1 and CNR2 in keratinocytes, suggests a protective mechanism against oxidative stress. In fibroblasts, this receptor modulation strengthens the skin’s resilience, potentially aiding in wound healing and protecting against further damage. Overall, F2CAA emerges as a highly versatile and effective formulation with a broad-spectrum impact on inflammation, tissue repair, and structural integrity, making it suited for managing complex skin conditions involving both oxidative and inflammatory stress across different cell types.
F3TAC exhibits variable and context-dependent effects on inflammation and skin repair across different cell types and stress conditions. This differential response is likely due to the specific interactions of CBG and CBC with cannabinoid receptors like GPR55, CNR1, and CNR2. CBG, known as a partial agonist of both CB1 and CB2 receptors, and CBC, which interacts with CB2 receptors and TRP channels, each contribute distinct anti-inflammatory and skin-regeneration effects. Under oxidative and inflammatory stress conditions, F3TAC modulates key structural genes like ELN, FN1, EGFR, and TIMP3, which are crucial for ECM remodeling and wound healing. The formulation potentially offers skin benefits, particularly by reducing oxidative stress and inflammation through CBC’s role as a CB2 receptor activator and free radical scavenger. CBG complements this by providing anti-inflammatory and analgesic effects through its partial agonism at both CB1 and CB2 receptors. F3TAC’s modulation of structural components like FN1 and TIMP3 highlights its capacity to support ECM remodelling and wound healing. The more focused approach could be beneficial in specific inflammatory scenarios where broader effects are less desirable. This makes F3TAC especially useful for managing targeted inflammatory conditions and promoting healing in conditions like acne, where inflammation control and scar prevention are critical.
Based on discussions and conclusions about the formulations' efficacy, a strategic approach can be formulated to address a range of skin health concerns, including aging, skin regeneration, and acne control. Each formulation has unique strengths, and by utilizing them either individually or in combination, more targeted and effective solutions can be achieved.
Aging -For aging skin, the primary goals are to reduce wrinkles, prevent further skin damage, maintain elasticity, and promote collagen production. The F2CAA formulation shows significant potential for addressing aging skin by targeting key processes such as wrinkle reduction, collagen production, and skin elasticity maintenance. Its effectiveness lies in the modulation of important structural genes like TIMP3, FN1, and ELN, which prevent fibrosis and support skin elasticity. F2CAA also exhibits strong anti-inflammatory properties by downregulating pro-inflammatory cytokines like TNF-α and upregulating IL-10, a critical factor in tissue repair and immune balance. F2CAA’s anti-aging capabilities are further enhanced through cannabinoid receptor modulation, particularly the synergistic interaction between CBD, CBG, and CBC, which supports wound healing, antioxidant protection, and tissue repair. This mechanism protects against oxidative stress while promoting resilience across different skin cell types. The combined effects of CBD, CBG, and CBC support collagen production, ECM remodeling, and the prevention of scarring, making F2CAA an ideal formulation for reducing the signs of aging. Combining F2CAA with F1CR2 could amplify these effects. While F2CAA focuses on ECM remodeling and preventing skin degradation, F1CR2 enhances tissue repair and mitigates oxidative stress, leading to a comprehensive anti-aging strategy that targets both inflammation control and skin regeneration.
Skin regeneration - For skin regeneration, the primary goal is to enhance healing, boost collagen synthesis, and maintain or restore skin integrity. F1CR2 and F1CR1 formulations were evaluated each demonstrating unique strengths in promoting regeneration, managing inflammation, and supporting tissue repair. F1CR2 stands out for its ability to promote long-term skin regeneration. It achieves this by upregulating key structural genes such as PPARγ, EGFR, TIMP3, and FN1, which enhance collagen production, stabilize the ECM, and maintain skin elasticity. This makes F1CR2 particularly effective in supporting the deeper layers of the skin, especially the dermis. Its strong activation of fibroblasts, essential for skin structure and wound healing, further underscores its suitability for deeper tissue repair, particularly in cases of chronic oxidative stress or skin aging. The upregulation of PPARγ by F1CR2 plays a vital role in regulating inflammation and aiding skin cells in their metabolic recovery, making this formulation highly effective for long-term regeneration and recovery from oxidative damage. F1CR1 is primarily focused on immediate inflammation control. It effectively reduces pro-inflammatory cytokines such as TNF-α and IL-6, making it well-suited for managing acute skin inflammation. F1CR1 also improves microcirculation, which helps reduce redness and irritation, crucial for the early stages of skin recovery. This formulation has been shown to excel in vivo, where its ability to modulate the skin’s response to external stressors leads to quicker recovery from inflammation and improved barrier function. The combination of CBD and CBC in F1CR1 enhances its anti-inflammatory properties and allows for deep skin penetration, promoting faster recovery and protecting the skin from environmental stress. Both formulations modulate cannabinoid receptors (CNR1, CNR2, GPR55) to promote healing and inflammation control. F1CR2 is particularly effective in fibroblast activation under oxidative stress, whereas F1CR1 demonstrates broader anti-inflammatory effects across both keratinocytes and fibroblasts, addressing inflammation and healing at both the surface (epidermis) and deeper layers (dermis) of the skin. The molecular insights behind the active ingredients show that CBD, with its moderate skin permeability, interacts with CB2 receptors to promote wound healing, while CBG, known for its role in collagen synthesis and antioxidant protection, supports long-term dermal repair. CBC also plays a role in modulating CB2 receptors in immune cells and fibroblasts, reducing inflammation and supporting wound healing. In vitro studies showed that F1CR2 outperformed F1CR1 in promoting tissue repair and structural integrity under controlled conditions. However, in vivo studies revealed that F1CR1 was more effective at reducing redness, improving microcirculation, and accelerating skin recovery. This difference is attributed to F1CR1’s ability to interact with a complex in vivo environment that involves vascularization, hydration, and immune responses. Combining F1CR2 and F1CR1 provides a synergistic approach to skin care. F1CR2 supports long-term regeneration, collagen production, and ECM remodeling, while F1CR1 acts as a maintenance solution to control inflammation and protect the skin from daily stressors. Using F1CR1 during the day for inflammation control and F1CR2 at night for deeper repair ensures comprehensive skin care, addressing both immediate and long-term needs.
Acne control - For acne control, the focus is on reducing inflammation and preventing scarring. F3TAC is highly effective for targeting the initial stages of acne formation. The F3TAC formulation is designed for acne prone skin, leveraging its anti-inflammatory and antimicrobial properties. Key ingredients include CBC, which regulates inflammation, and CBG, which acts as an antimicrobial agent to reduce acne-causing bacteria. F3TAC was shown to lower pro-inflammatory cytokines like TNF-α and IL-6 in vitro, although in vivo results were less pronounced, due to the skin's complex environment, including factors like permeability and microbiome balance. Ongoing studies on the skin microbiome and sebocytes will provide further insights into how F3TAC affects acne by regulating sebum production. The combined use of F3TAC and F1CR2 offers a synergistic approach for acne-prone skin. F3TAC addresses immediate inflammation, while F1CR2 provides long-term anti-inflammatory benefits that help prevent acne lesions from worsening and support healing. Together, these formulations can reduce scarring and maintain clear skin.
The findings on the four formulations emphasize the importance of selecting the appropriate cannabinoid-based formulation according to the specific skin condition and therapeutic goals. The different effects of these formulations highlight the potential of cannabinoid combinations in enhancing skin health through targeted modulation of inflammatory, regenerative, and structural pathways. As research into cannabinoid-based therapies continues to evolve, these formulations represent promising candidates for advancing the management of a wide range of dermatological conditions. The formulations can be applied either as standalone treatments or in synergistic combinations, depending on the specific skin health challenge being addressed. This targeted approach maximizes the strengths of each formulation to achieve optimal outcomes in skin care.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, S1 – Effect of formulations on pro-inflammatory markers; S2 – Effect the formulations on anti-inflammatory markers; S3 – Effect of formulations on structural markers; S4 – Effect of formulations on cannabinoid receptor markers.
Author Contributions
Conceptualization and methodology M.M.G., C.P.R., S.S; formal analysis G.T., M.C., M.M., I.L., M.M.G., C.P.R., S.S.; investigation, G.T., M.C., C.F., M.M., I.L., M.M.G., C.P.R., S.S.; resources, M.M.G., C.P.R., S.S..; writing—original draft preparation, G.T., M.C.; writing—review and editing, M.M.G., C.P.R., S.S.; supervision, M.M.G., C.P.R, S.S.; project administration, M.M.G., C.P.R., S.S.; funding acquisition, M.M.G., C.P.R., S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Union through the European Regional Development Fund (FEDER), under the programs Compete2020, Portugal2020, Lisboa2020, and Norte2020, with project references LISBOA-01-0247-FEDER-047213 and NORTE-01-247-FEDER. The project, titled "SkanABility – Proof of Concept for the Biological Activity of Cannabinoids and Terpenes – Cosmeceutical Applications," involved the following beneficiaries: EXMceuticals Portugal Lda (Coordinator), COSMETEK Lda, VINIDEAS (Desenvolvimento Enológico), Faculdade de Farmácia da Universidade de Lisboa, Universidade do Minho, COFAC – Universidade Lusófona de Humanidades e Tecnologias, and LM – Laboratório Nacional do Medicamento (Estado Maior do Exército and Fundacão para a Ciência e Tecnologia (FCT) through projects UIDB/04138/2020 and UIDP/04138/2020.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors due commercial reasons. Restrictions apply to the availability of these data. Data were obtained from ExMceuticals and are available [from the authors/at URL] with the permission of EXMceuticals.
Conflicts of Interest
Authors SS, GT, IL, and MM was employed by company EXMceuticals Portugal Lda. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
2-AG – 2-arachidonoylglycerol; AEA – anandamide; CB1 – cannabinoid receptor type 1; CB2 – cannabinoid receptor type 2; CNR1 – gene cannabinoid receptor type 1; CNR2 CBC – cannabichromene; CBD – cannabidiol; CBG – cannabigerol; CBN – cannabinol; ECM – extracellular matrix; ECS – endocannabinoid system; EGFR – epidermal growth factor receptor; ELN – elastin; FN1 – fibronectin; GPR55 – G protein-coupled receptor 55; H2O2 – hydrogen peroxidase; HaCat – human keratinocytes; HDF – human dermal fibroblast; IL-10 – Interleukin 10; IL-36G – Interleukin-36 gamma; IL-6 – Interleukin 6; MMPs – matrix metalloproteinases; NF – кB – nuclear factor kappa B; pCBs – phytocannabinoids; PKC – protein kinase C; PPARγ – proliferator – activated receptor gamma; ROS – reactive oxygen species; TEWL – trans epidermal water loss; THC – Δ9-tetrahydrocannabinol; TIMP3 – metallopeptidase inhibitor 3; TNF-α – Tumor necrosis factor alfa; TPA – phorbol 12-myrstate 13-acetate
References
- ElSohly, M.A.; Slade, D. Chemical Constituents of Marijuana: The Complex Mixture of Natural Cannabinoids. Life Sci 2005, 78, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Legare, C.A.; Raup-Konsavage, W.M.; Vrana, K.E. Therapeutic Potential of Cannabis, Cannabidiol, and Cannabinoid-Based Pharmaceuticals. Pharmacology 2022, 107, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Radwan, M.M.; Chandra, S.; Gul, S.; ElSohly, M.A. Cannabinoids, Phenolics, Terpenes and Alkaloids of Cannabis. Molecules 2021, 26, 2774. [Google Scholar] [CrossRef]
- Martinelli, G.; Magnavacca, A.; Fumagalli, M.; DellʼAgli, M.; Piazza, S.; Sangiovanni, E. Cannabis Sativa and Skin Health: Dissecting the Role of Phytocannabinoids. Planta Med 2022, 88, 492–506. [Google Scholar] [CrossRef]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An Overview of the Cannabinoid Type 2 Receptor System and Its Therapeutic Potential. Curr Opin Anaesthesiol 2018, 31, 407–414. [Google Scholar] [CrossRef]
- Scheau, C.; Badarau, I.A.; Mihai, L.-G.; Scheau, A.-E.; Costache, D.O.; Constantin, C.; Calina, D.; Caruntu, C.; Costache, R.S.; Caruntu, A. Cannabinoids in the Pathophysiology of Skin Inflammation. Molecules 2020, 25, 652. [Google Scholar] [CrossRef]
- Katona, I.; Freund, T.F. Multiple Functions of Endocannabinoid Signaling in the Brain. Annu Rev Neurosci 2012, 35, 529–558. [Google Scholar] [CrossRef]
- Mackie, K. Distribution of Cannabinoid Receptors in the Central and Peripheral Nervous System. In Cannabinoids; Springer-Verlag: Berlin/Heidelberg, 2005; pp. 299–325. [Google Scholar]
- Piomelli, D. The Molecular Logic of Endocannabinoid Signalling. Nat Rev Neurosci 2003, 4, 873–884. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int J Mol Sci 2018, 19, 833. [Google Scholar] [CrossRef]
- Rodríguez de Fonseca, F.; Del Arco, I.; Bermudez-Silva, F.J.; Bilbao, A.; Cippitelli, A.; Navarro, M. The Endocannabinoid system: physiology and pharmacology. Alcohol and Alcoholism 2005, 40, 2–14. [Google Scholar] [CrossRef]
- Osafo, N.; Yeboah, O.K.; Antwi, A.O. Endocannabinoid System and Its Modulation of Brain, Gut, Joint and Skin Inflammation. Mol Biol Rep 2021, 48, 3665–3680. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.P.; Duarte, P.; Calado, M.; Almeida, A.J.; Reis, C.P.; Gaspar, M.M. The Current Role of Cannabis and Cannabinoids in Health: A Comprehensive Review of Their Therapeutic Potential. Life Sci 2023, 329, 121838. [Google Scholar] [CrossRef] [PubMed]
- Žugić, A.; Martinović, M.; Tadić, V.; Rajković, M.; Racić, G.; Nešić, I.; Koren, A. Comprehensive Insight into Cutaneous Application of Hemp. Pharmaceutics 2024, 16, 748. [Google Scholar] [CrossRef] [PubMed]
- GL*, S. Scientific Evidence for the Cosmetic Skin and Hair Benefits of Hemp-Derived Cannabinoids: A Review. Clinical Dermatology Open Access Journal 2024, 9. [Google Scholar] [CrossRef]
- Wilkinson, J.D.; Williamson, E.M. Cannabinoids Inhibit Human Keratinocyte Proliferation through a Non-CB1/CB2 Mechanism and Have a Potential Therapeutic Value in the Treatment of Psoriasis. J Dermatol Sci 2007, 45, 87–92. [Google Scholar] [CrossRef]
- Wang, L.-L.; Zhao, R.; Li, J.-Y.; Li, S.-S.; Liu, M.; Wang, M.; Zhang, M.-Z.; Dong, W.-W.; Jiang, S.-K.; Zhang, M.; et al. Pharmacological Activation of Cannabinoid 2 Receptor Attenuates Inflammation, Fibrogenesis, and Promotes Re-Epithelialization during Skin Wound Healing. Eur J Pharmacol 2016, 786, 128–136. [Google Scholar] [CrossRef]
- Perez, E.; Fernandez, J.R.; Fitzgerald, C.; Rouzard, K.; Tamura, M.; Savile, C. In Vitro and Clinical Evaluation of Cannabigerol (CBG) Produced via Yeast Biosynthesis: A Cannabinoid with a Broad Range of Anti-Inflammatory and Skin Health-Boosting Properties. Molecules 2022, 27, 491. [Google Scholar] [CrossRef]
- Turcotte, C.; Blanchet, M.-R.; Laviolette, M.; Flamand, N. The CB2 Receptor and Its Role as a Regulator of Inflammation. Cellular and Molecular Life Sciences 2016, 73, 4449–4470. [Google Scholar] [CrossRef]
- Oláh, A.; Tóth, B.I.; Borbíró, I.; Sugawara, K.; Szöllõsi, A.G.; Czifra, G.; Pál, B.; Ambrus, L.; Kloepper, J.; Camera, E.; et al. Cannabidiol Exerts Sebostatic and Antiinflammatory Effects on Human Sebocytes. Journal of Clinical Investigation 2014, 124, 3713–3724. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Fumagalli, M.; Pacchetti, B.; Piazza, S.; Magnavacca, A.; Khalilpour, S.; Melzi, G.; Martinelli, G.; Dell’Agli, M. Cannabis Sativa L. Extract and Cannabidiol Inhibit in Vitro Mediators of Skin Inflammation and Wound Injury. Phytotherapy Research 2019, 33, 2083–2093. [Google Scholar] [CrossRef]
- Petrosino, S.; Verde, R.; Vaia, M.; Allarà, M.; Iuvone, T.; Di Marzo, V. Anti-Inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis. Journal of Pharmacology and Experimental Therapeutics 2018, 365, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Casares, L.; García, V.; Garrido-Rodríguez, M.; Millán, E.; Collado, J.A.; García-Martín, A.; Peñarando, J.; Calzado, M.A.; de la Vega, L.; Muñoz, E. Cannabidiol Induces Antioxidant Pathways in Keratinocytes by Targeting BACH1. Redox Biol 2020, 28, 101321. [Google Scholar] [CrossRef] [PubMed]
- Styrczewska, M.; Kostyn, A.; Kulma, A.; Majkowska-Skrobek, G.; Augustyniak, D.; Prescha, A.; Czuj, T.; Szopa, J. Flax Fiber Hydrophobic Extract Inhibits Human Skin Cells Inflammation and Causes Remodeling of Extracellular Matrix and Wound Closure Activation. Biomed Res Int 2015, 2015, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants 2019, 9, 21. [Google Scholar] [CrossRef]
- Giacoppo, S.; Mandolino, G.; Galuppo, M.; Bramanti, P.; Mazzon, E. Cannabinoids: New Promising Agents in the Treatment of Neurological Diseases. Molecules 2014, 19, 18781–18816. [Google Scholar] [CrossRef]
- Donvito, G.; Nass, S.R.; Wilkerson, J.L.; Curry, Z.A.; Schurman, L.D.; Kinsey, S.G.; Lichtman, A.H. The Endogenous Cannabinoid System: A Budding Source of Targets for Treating Inflammatory and Neuropathic Pain. Neuropsychopharmacology 2018, 43, 52–79. [Google Scholar] [CrossRef]
- Palmieri, B.; Laurino, C.; Vadalà, M. A Therapeutic Effect of Cbd-Enriched Ointment in Inflammatory Skin Diseases and Cutaneous Scars. Clin Ter 2019, 170, e93–e99. [Google Scholar] [CrossRef]
- Tóth, K.F.; Ádám, D.; Bíró, T.; Oláh, A. Cannabinoid Signaling in the Skin: Therapeutic Potential of the “C(Ut)Annabinoid” System. Molecules 2019, 24, 918. [Google Scholar] [CrossRef]
- Baswan, S.M.; Klosner, A.E.; Glynn, K.; Rajgopal, A.; Malik, K.; Yim, S.; Stern, N. Therapeutic Potential of Cannabidiol (CBD) for Skin Health and Disorders. Clin Cosmet Investig Dermatol 2020, Volume 13, 927–942. [Google Scholar] [CrossRef]
- Martins, A.M.; Gomes, A.L.; Vilas Boas, I.; Marto, J.; Ribeiro, H.M. Cannabis-Based Products for the Treatment of Skin Inflammatory Diseases: A Timely Review. Pharmaceuticals 2022, 15, 210. [Google Scholar] [CrossRef]
- Coelho, P., M.; Duarte, P.A.; Calado, M.; Trigo, G.; Santos, S.; Reis, P., C.; Gaspar, M.M. Cannabinoid-Based Topical Formulations for Skin Disorders.; Lisbon, 2023.
- Coelho, P., M.; Duarte, P.A.; Calado, M.; Almeida, J., A.; Trigo, G.; Santos, S.; Reis, P., C.; Gaspar, M.M. Application of Cannabinoids in Cosmeceutical Formulations.; Varna, 2023.
- Heo, S.; Kim, S.; Kang, D. The Role of Hydrogen Peroxide and Peroxiredoxins throughout the Cell Cycle. Antioxidants 2020, 9, 280. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, H.; Xu, F.; Jiang, X.; Ma, H.; Seeram, N.P. Cannabidiol Protects Human Skin Keratinocytes from Hydrogen-Peroxide-Induced Oxidative Stress via Modulation of the Caspase-1–IL-1β Axis. J Nat Prod 2021, 84, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.B.; Smallwood, M.J.; Smerdon, G.R.; Winyard, P.G. Cellular Pre-Adaptation to the High O2 Concentration Used in Standard Cell Culture Confers Resistance to Subsequent H2O2-Induced Cell Death. Antioxidants 2024, 13, 269. [Google Scholar] [CrossRef] [PubMed]
- PubChem Phorbol 12-Myristate 13-Acetate. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Phorbol-12-myristate-13-acetate (accessed on 22 April 2024).
- Chang, S.N.; Dey, D.K.; Oh, S.T.; Kong, W.H.; Cho, K.H.; Al-Olayan, E.M.; Hwang, B.S.; Kang, S.C.; Park, J.G. Phorbol 12-Myristate 13-Acetate Induced Toxicity Study and the Role of Tangeretin in Abrogating HIF-1α-NF-ΚB Crosstalk In Vitro and In Vivo. Int J Mol Sci 2020, 21, 9261. [Google Scholar] [CrossRef]
- Traore, K.; Trush, M.A.; George, M.; Spannhake, E.Wm.; Anderson, W.; Asseffa, A. Signal Transduction of Phorbol 12-Myristate 13-Acetate (PMA)-Induced Growth Inhibition of Human Monocytic Leukemia THP-1 Cells Is Reactive Oxygen Dependent. Leuk Res 2005, 29, 863–879. [Google Scholar] [CrossRef]
- Cialdai, F.; Risaliti, C.; Monici, M. Role of Fibroblasts in Wound Healing and Tissue Remodeling on Earth and in Space. Front Bioeng Biotechnol 2022, 10, 958381. [Google Scholar] [CrossRef]
- Mazzantini, C.; El Bourji, Z.; Parisio, C.; Davolio, P.L.; Cocchi, A.; Pellegrini-Giampietro, D.E.; Landucci, E. Anti-Inflammatory Properties of Cannabidiol and Beta-Caryophyllene Alone or Combined in an In Vitro Inflammation Model. Pharmaceuticals 2024, 17, 467. [Google Scholar] [CrossRef]
- Di Meo, C.; Tortolani, D.; Standoli, S.; Angelucci, C.B.; Fanti, F.; Leuti, A.; Sergi, M.; Kadhim, S.; Hsu, E.; Rapino, C.; et al. Effects of Rare Phytocannabinoids on the Endocannabinoid System of Human Keratinocytes. Int J Mol Sci 2022, 23, 5430. [Google Scholar] [CrossRef]
- Tortolani, D.; Di Meo, C.; Standoli, S.; Ciaramellano, F.; Kadhim, S.; Hsu, E.; Rapino, C.; Maccarrone, M. Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway. Int J Mol Sci 2023, 24, 2721. [Google Scholar] [CrossRef]
- Liu, H.-M.; Cheng, M.-Y.; Xun, M.-H.; Zhao, Z.-W.; Zhang, Y.; Tang, W.; Cheng, J.; Ni, J.; Wang, W. Possible Mechanisms of Oxidative Stress-Induced Skin Cellular Senescence, Inflammation, and Cancer and the Therapeutic Potential of Plant Polyphenols. Int J Mol Sci 2023, 24, 3755. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-ΚB: A Key Role in Inflammatory Diseases. Journal of Clinical Investigation 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Radaszkiewicz, K.A.; Beckerová, D.; Woloszczuková, L.; Radaszkiewicz, T.W.; Lesáková, P.; Blanářová, O.V.; Kubala, L.; Humpolíček, P.; Pachernik, J. 12-O-Tetradecanoylphorbol-13-Acetate Increases Cardiomyogenesis through PKC/ERK Signaling. Sci Rep 2020, 10, 15922. [Google Scholar] [CrossRef] [PubMed]
- Milando, R.; Friedman, A. Cannabinoids: Potential Role in Inflammatory and Neoplastic Skin Diseases. Am J Clin Dermatol 2019, 20, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M. Regulation of Tissue Homeostasis by NF-ΚB Signalling: Implications for Inflammatory Diseases. Nat Rev Immunol 2009, 9, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Pantelis, P.; Theocharous, G.; Lagopati, N.; Veroutis, D.; Thanos, D.-F.; Lampoglou, G.-P.; Pippa, N.; Gatou, M.-A.; Tremi, I.; Papaspyropoulos, A.; et al. The Dual Role of Oxidative-Stress-Induced Autophagy in Cellular Senescence: Comprehension and Therapeutic Approaches. Antioxidants 2023, 12, 169. [Google Scholar] [CrossRef]
- Terao, R.; Ahmed, T.; Suzumura, A.; Terasaki, H. Oxidative Stress-Induced Cellular Senescence in Aging Retina and Age-Related Macular Degeneration. Antioxidants 2022, 11, 2189. [Google Scholar] [CrossRef]
- Saputra, F.; Kishida, M.; Hu, S.-Y. Oxidative Stress Induced by Hydrogen Peroxide Disrupts Zebrafish Visual Development by Altering Apoptosis, Antioxidant and Estrogen Related Genes. Sci Rep 2024, 14, 14454. [Google Scholar] [CrossRef]
- He, S.; Li, Q.; Huang, Q.; Cheng, J. Targeting Protein Kinase C for Cancer Therapy. Cancers (Basel) 2022, 14, 1104. [Google Scholar] [CrossRef]
- Neagu, M.; Constantin, C. Correction to: Signal Transduction in Immune Cells and Protein Kinases. In; 2021; pp. C1–C1.
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Abdul Hamid, Z.; Mohamad Anuar, N.N.; Jalil, J.; Mohd Nor, N.A.; Budin, S.B. The Role of PKC-MAPK Signalling Pathways in the Development of Hyperglycemia-Induced Cardiovascular Complications. Int J Mol Sci 2022, 23, 8582. [Google Scholar] [CrossRef]
- Lee, W.L.; Shalita, A.R.; Poh-Fitzpatrick, M.B. Comparative Studies of Porphyrin Production in Propionibacterium Acnes and Propionibacterium Granulosum. J Bacteriol 1978, 133, 811–815. [Google Scholar] [CrossRef]
- Kjeldstad, B. Photoinactivation of Propionibacterium Acnes by Near-Ultraviolet Light. Zeitschrift für Naturforschung C 1984, 39, 300–302. [Google Scholar] [CrossRef]
- Borelli, C.; Merk, K.; Schaller, M.; Jacob, K.; Vogeser, M.; Weindl, G.; Berger, U.; Plewig, G. In Vivo Porphyrin Production by P. Acnes in Untreated Acne Patients and Its Modulation by Acne Treatment. Acta Derm Venereol 2006, 86, 316–319. [Google Scholar] [CrossRef]
Figure 1.
Assessment of cell viability of HaCat and HDF exposed 24 h to the stress inducing agent, H2O2, tested over a concentration range between 50 and 400 µM. Results correspond to 6 replicates and 3 independent assays.
Figure 1.
Assessment of cell viability of HaCat and HDF exposed 24 h to the stress inducing agent, H2O2, tested over a concentration range between 50 and 400 µM. Results correspond to 6 replicates and 3 independent assays.
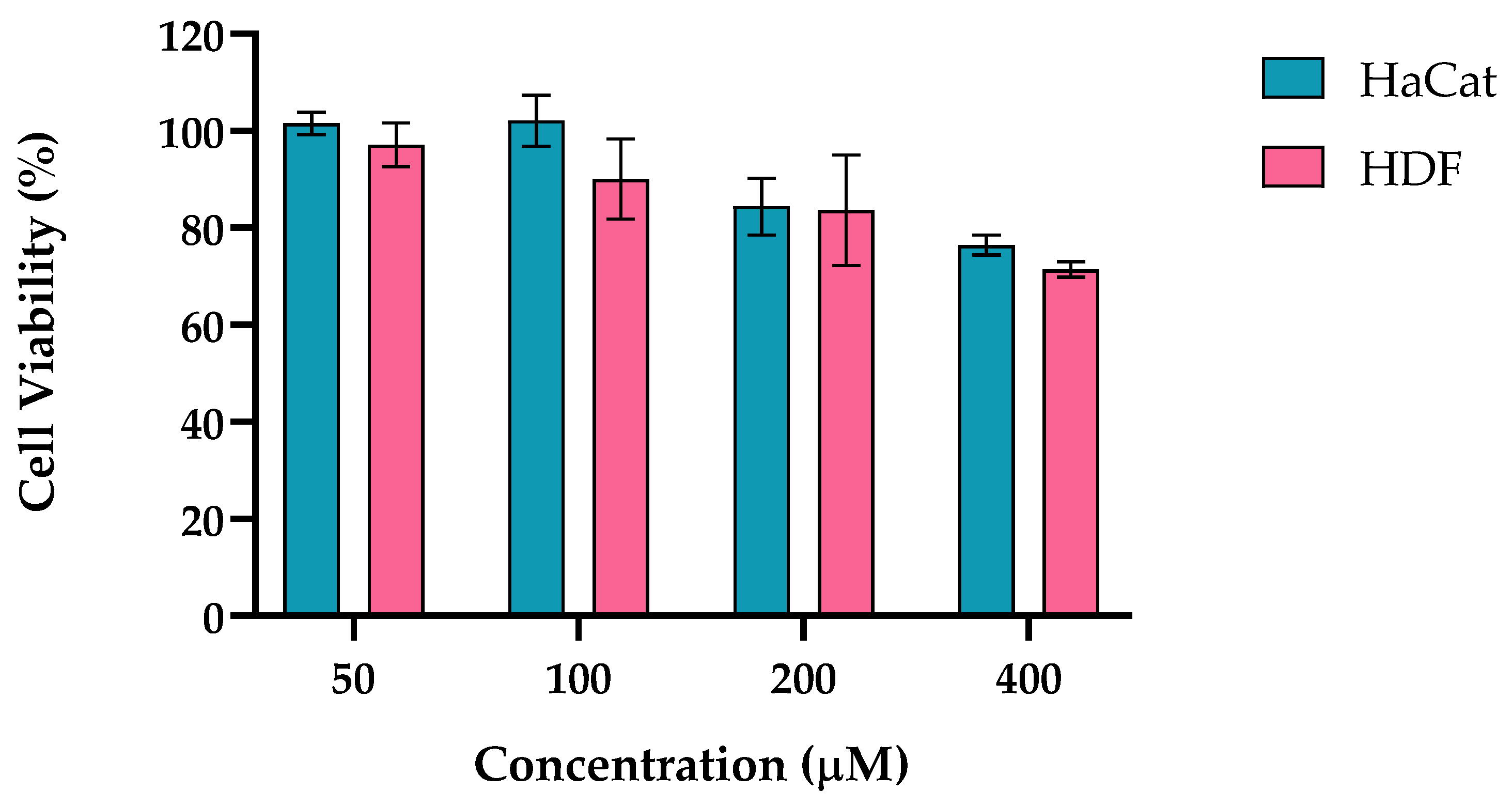
Figure 2.
Assessment of cell viability of HaCat exposed 24 h to the stress inducing agent, TPA, tested over a range of concentrations between 5 and 20 µM. Results correspond to 6 replicates and 3 independent assays.
Figure 2.
Assessment of cell viability of HaCat exposed 24 h to the stress inducing agent, TPA, tested over a range of concentrations between 5 and 20 µM. Results correspond to 6 replicates and 3 independent assays.
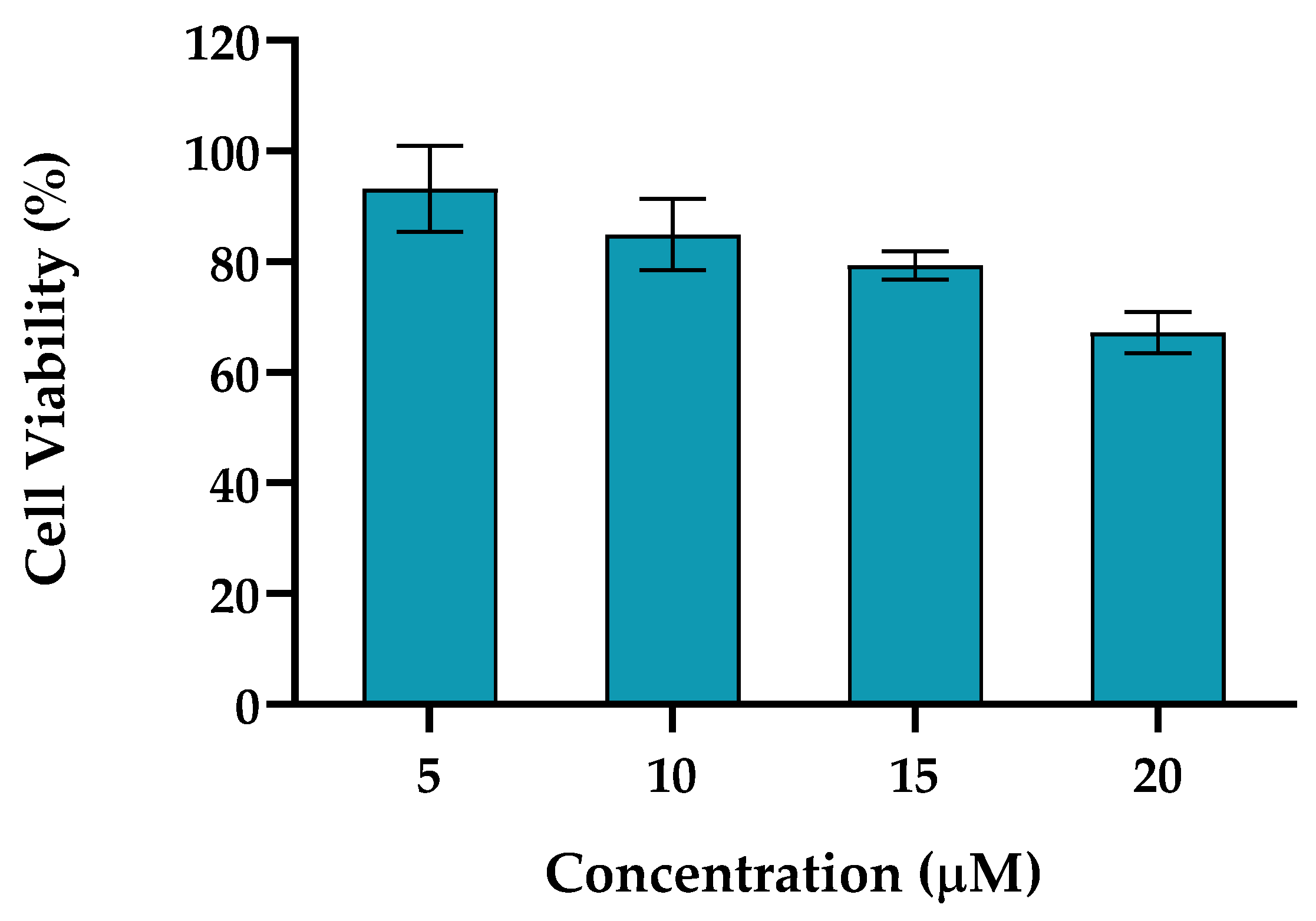
Figure 3.
Schematic representation of gene expression pathways modulated by stressor and pCBs highlighting their potential effects on keratinocytes and fibroblasts. (1) Oxidative Stress (OS) pathway. H₂O₂ generates an excess of ROS that triggers the activation of NF-κB signaling cascade. NF-κB translocate from the cytoplasm to the cell nucleus promoting the transcription of various pro-inflammatory genes, including TNF-α (a cytokine involved in systemic inflammation), IL-36G (a cytokine family involved in inflammatory responses) and IL-6 (pro- and anti-inflammatory properties) (2) Chemical stress (CS) pathway. TPA, activates PKC that a) Stimulates the NF-κB pathway, leading to the transcription of the same set of inflammatory genes as in the OS pathway; b) Triggers a separate signaling cascade that results in the activation of MMPs that can significantly affect ECM stability, tissue integrity, and cellular behavior. (3) Cannabinoid receptors, CB1R, CB2R and GPR55. While pCBs cannabinoids may not always directly activate these receptors, they can influence their expression and modulate the ECS. The activation of the receptors leads to a) Stimulation of the PPARγ pathway, that can inhibit the inflammatory process by directly suppressing the NF-κB pathway; b) Direct inhibition of the NF-κB pathway through multiple mechanisms, which may include preventing NF-κB translocation to the nucleus, interfering with NF-κB DNA binding. Solid Arrows: Direct activation or positive regulation, Bar Arrows: Degradation or inhibitory steps, Dashed Arrows: Secondary or indirect steps, Blue Boxes: Components of the inflammatory process, Brown Box: MMPs process, Blue Circles: Pro-inflammatory genes expressed, Green Boxes: Components of the PPARγ pathway, Yellow Circles: Anti-inflammatory and tissue repair genes expressed. Created using Biorender.com.
Figure 3.
Schematic representation of gene expression pathways modulated by stressor and pCBs highlighting their potential effects on keratinocytes and fibroblasts. (1) Oxidative Stress (OS) pathway. H₂O₂ generates an excess of ROS that triggers the activation of NF-κB signaling cascade. NF-κB translocate from the cytoplasm to the cell nucleus promoting the transcription of various pro-inflammatory genes, including TNF-α (a cytokine involved in systemic inflammation), IL-36G (a cytokine family involved in inflammatory responses) and IL-6 (pro- and anti-inflammatory properties) (2) Chemical stress (CS) pathway. TPA, activates PKC that a) Stimulates the NF-κB pathway, leading to the transcription of the same set of inflammatory genes as in the OS pathway; b) Triggers a separate signaling cascade that results in the activation of MMPs that can significantly affect ECM stability, tissue integrity, and cellular behavior. (3) Cannabinoid receptors, CB1R, CB2R and GPR55. While pCBs cannabinoids may not always directly activate these receptors, they can influence their expression and modulate the ECS. The activation of the receptors leads to a) Stimulation of the PPARγ pathway, that can inhibit the inflammatory process by directly suppressing the NF-κB pathway; b) Direct inhibition of the NF-κB pathway through multiple mechanisms, which may include preventing NF-κB translocation to the nucleus, interfering with NF-κB DNA binding. Solid Arrows: Direct activation or positive regulation, Bar Arrows: Degradation or inhibitory steps, Dashed Arrows: Secondary or indirect steps, Blue Boxes: Components of the inflammatory process, Brown Box: MMPs process, Blue Circles: Pro-inflammatory genes expressed, Green Boxes: Components of the PPARγ pathway, Yellow Circles: Anti-inflammatory and tissue repair genes expressed. Created using Biorender.com.
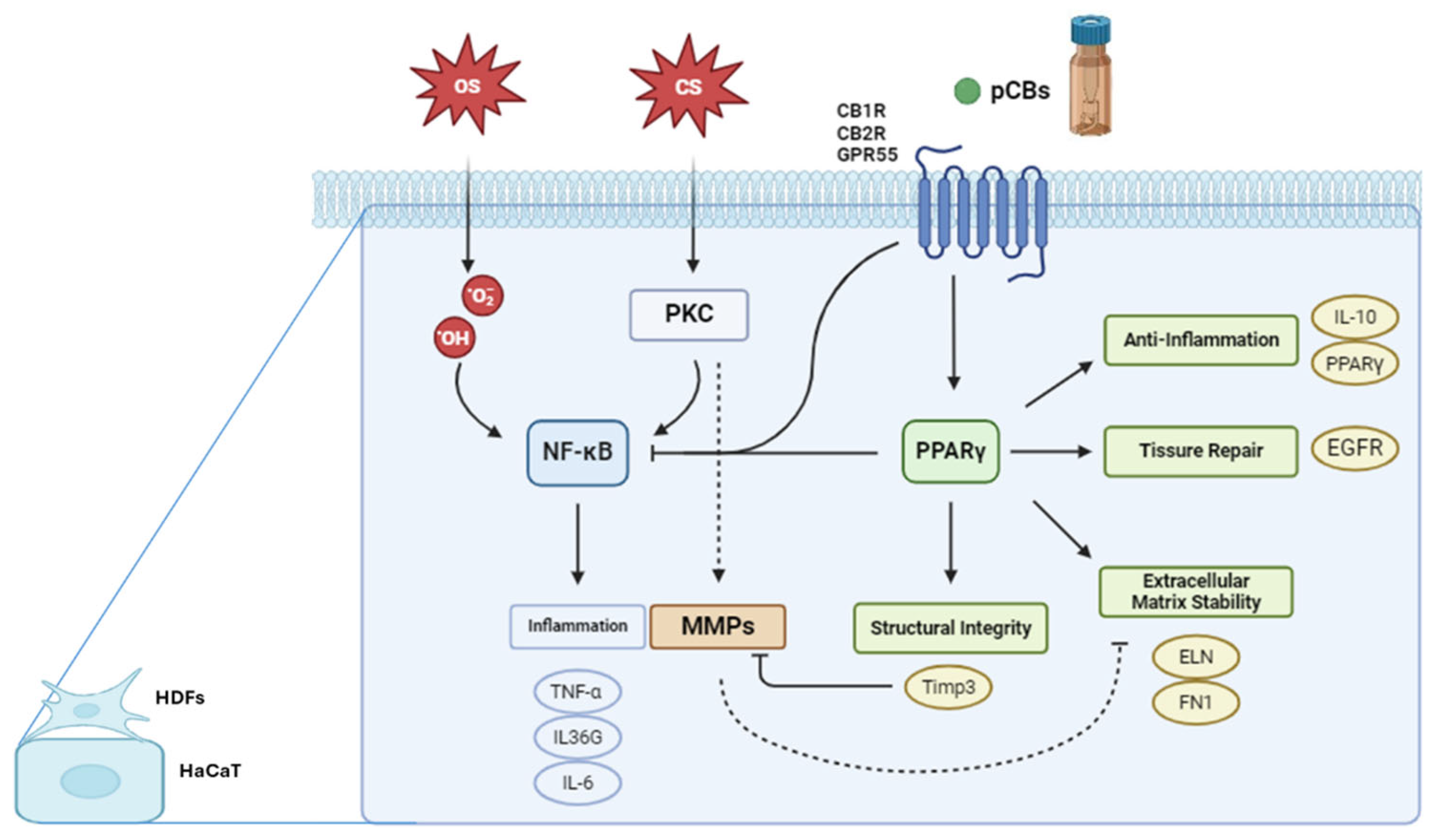
Figure 4.
Heat map of pro-inflammatory genetic markers (IL-6, TNF-α and IL-36G) in HaCaT and HDF cells induced with H2O2 and TPA and pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
Figure 4.
Heat map of pro-inflammatory genetic markers (IL-6, TNF-α and IL-36G) in HaCaT and HDF cells induced with H2O2 and TPA and pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
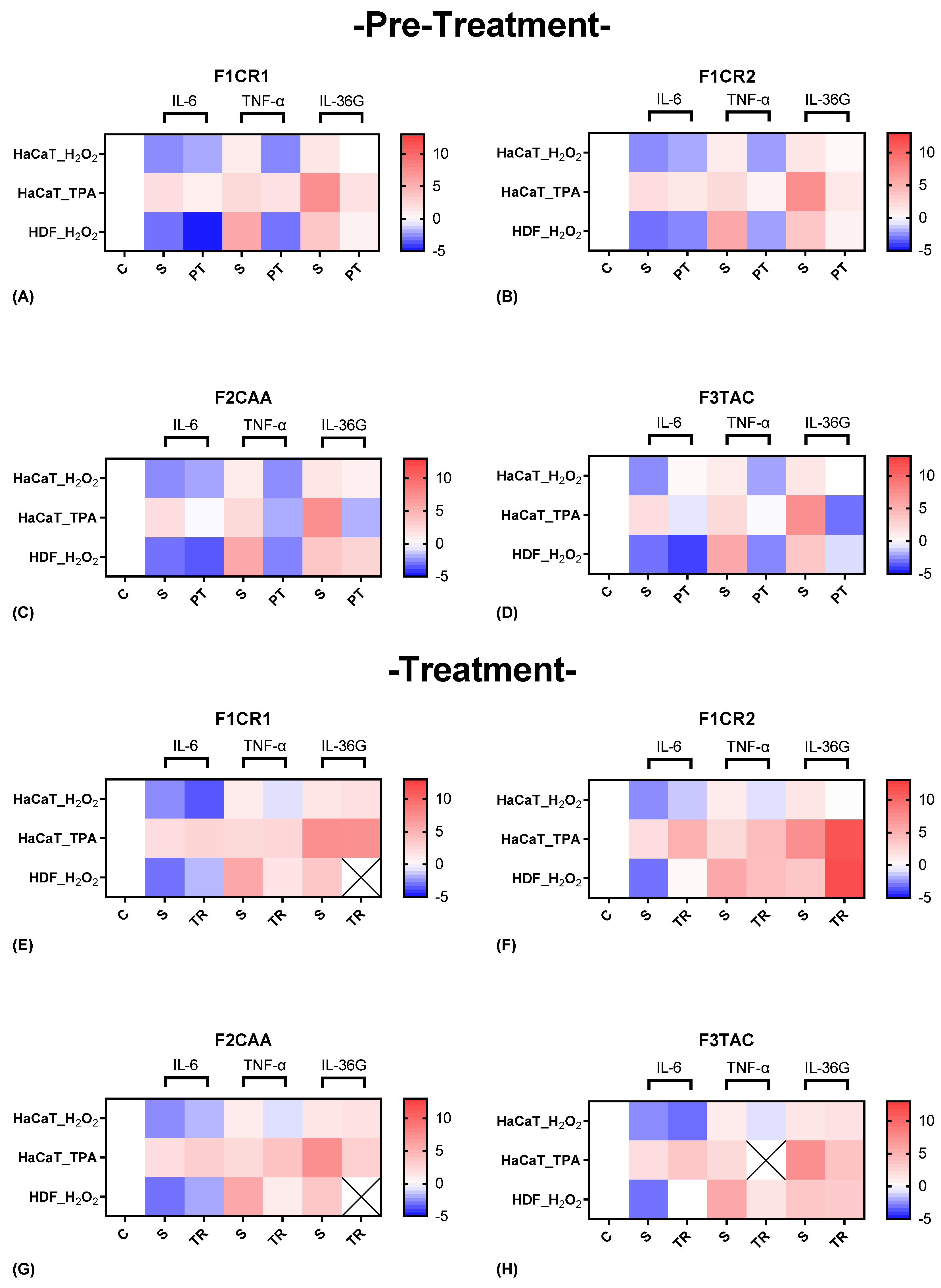
Figure 5.
Heat map of anti-inflammatory genetic markers (IL-10 and PPARγ) in HaCaT and HDF cells induced with H2O2 and TPA and pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
Figure 5.
Heat map of anti-inflammatory genetic markers (IL-10 and PPARγ) in HaCaT and HDF cells induced with H2O2 and TPA and pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
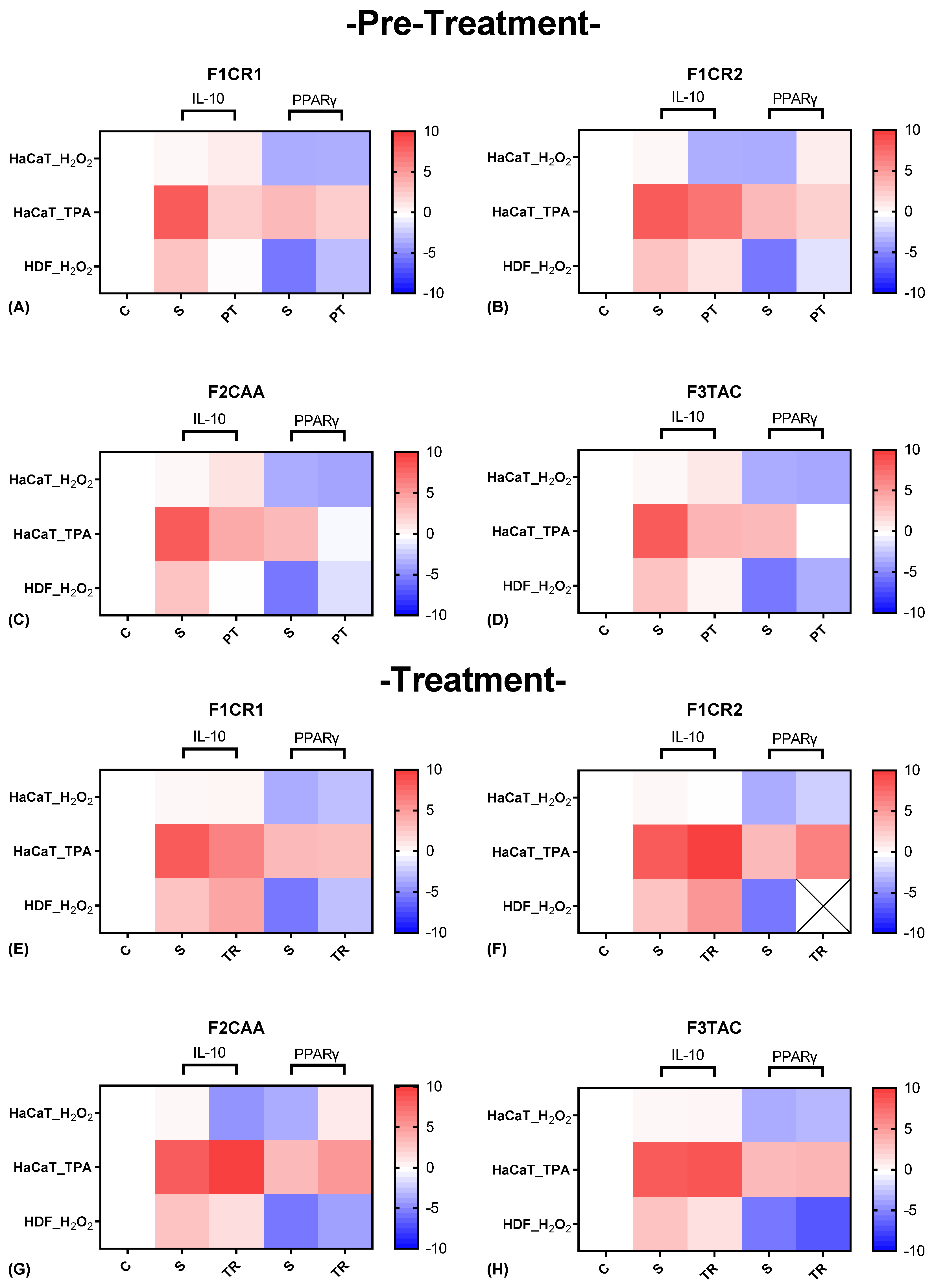
Figure 6.
Heat map of structural genetic markers (ELN, FN1, EGFR and TIMP3) in HaCaT and HDF cells induced with H2O2 and TPA pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
Figure 6.
Heat map of structural genetic markers (ELN, FN1, EGFR and TIMP3) in HaCaT and HDF cells induced with H2O2 and TPA pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
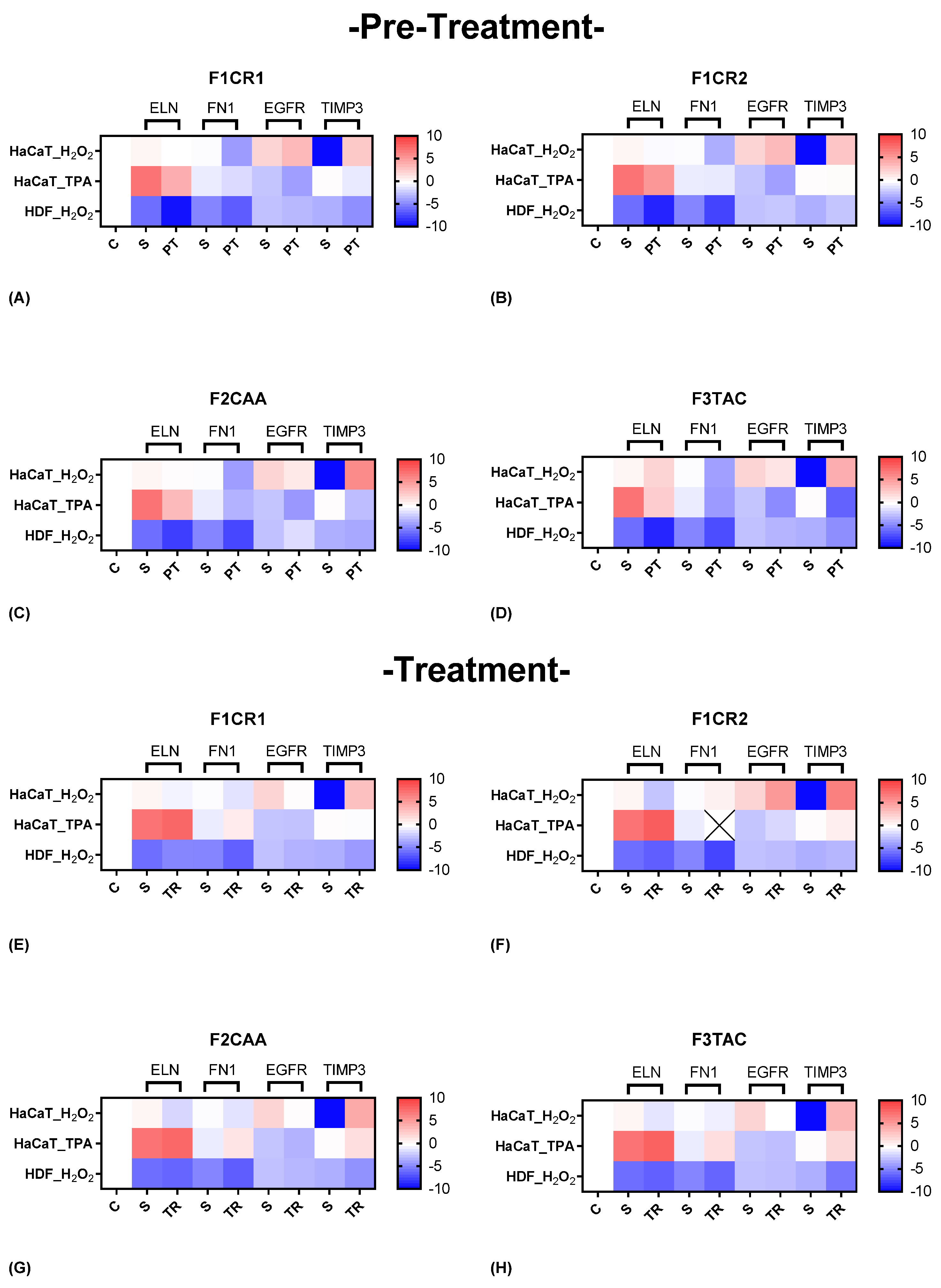
Figure 7.
Heat map of cannabinoid receptors genetic markers (GPR55, CNR2 and CNR1) in HaCaT and HDF cells induced with H2O2 and TPA and pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
Figure 7.
Heat map of cannabinoid receptors genetic markers (GPR55, CNR2 and CNR1) in HaCaT and HDF cells induced with H2O2 and TPA and pre-treated (A, B, C, D) and treated (E, F, G, H) with the formulations F1CR1, F1CR2, F2CAA and F3TAC. The blue color indicates downregulation of the gene, whereas the red represents upregulation. C= control, S= stress condition, PT= pre-treatment with Formulation, TR= treatment with Formulation, Cross= no detectable expression level. Data are expressed as mean (Log2 (Fold Change)) of sets of experiments for each condition (n ≥ 2).
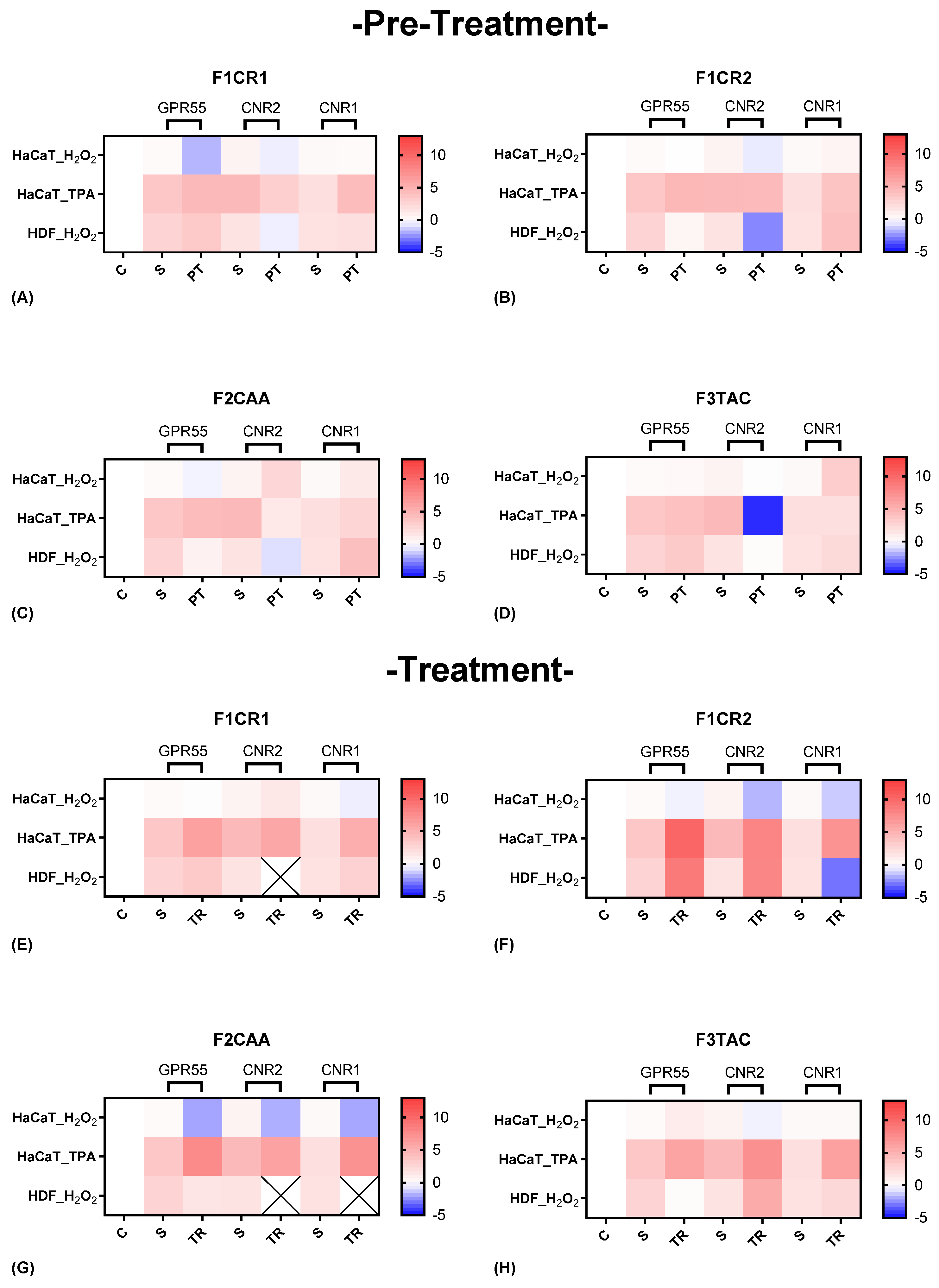
Figure 8.
Scheme of the two in vitro schedules (pre-treatment and treatment) used assessing the ability of pCBs formulations to modify gene expression to basal levels.
Figure 8.
Scheme of the two in vitro schedules (pre-treatment and treatment) used assessing the ability of pCBs formulations to modify gene expression to basal levels.
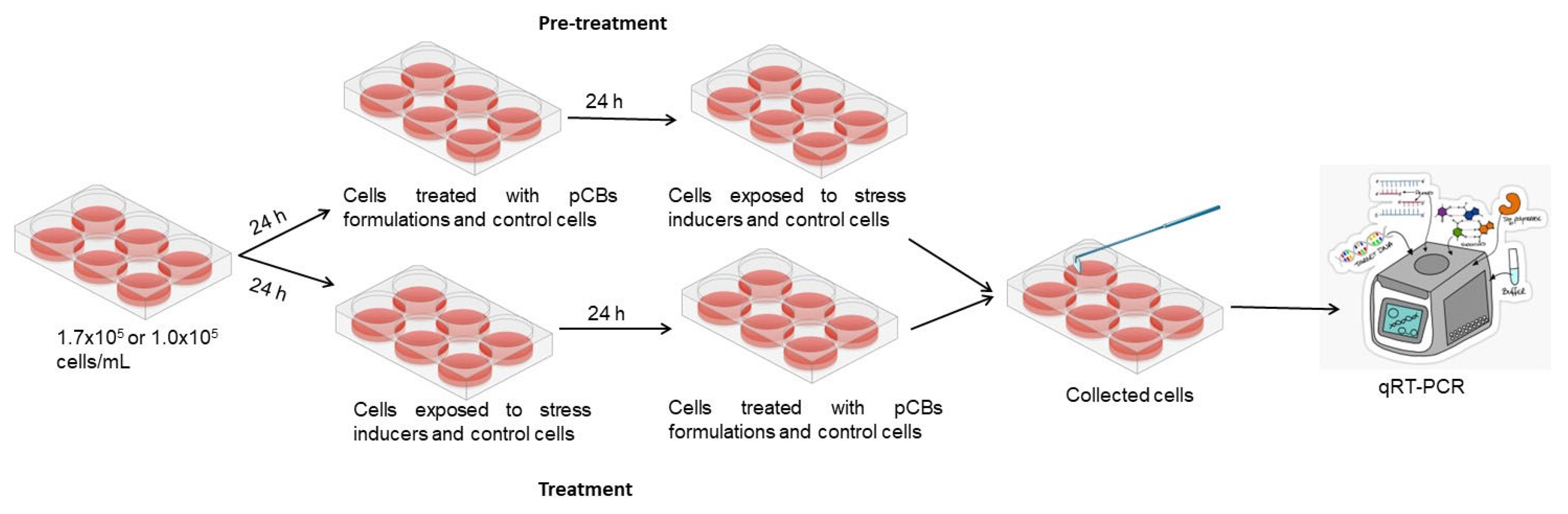
Figure 9.
Scheme illustrating the time points used in the in vitro assays for (A) pre-treatment and (B) treatment, designed to evaluate the efficacy of pCB formulations and their impact on gene expression modulation. Created using Biorender.com.
Figure 9.
Scheme illustrating the time points used in the in vitro assays for (A) pre-treatment and (B) treatment, designed to evaluate the efficacy of pCB formulations and their impact on gene expression modulation. Created using Biorender.com.
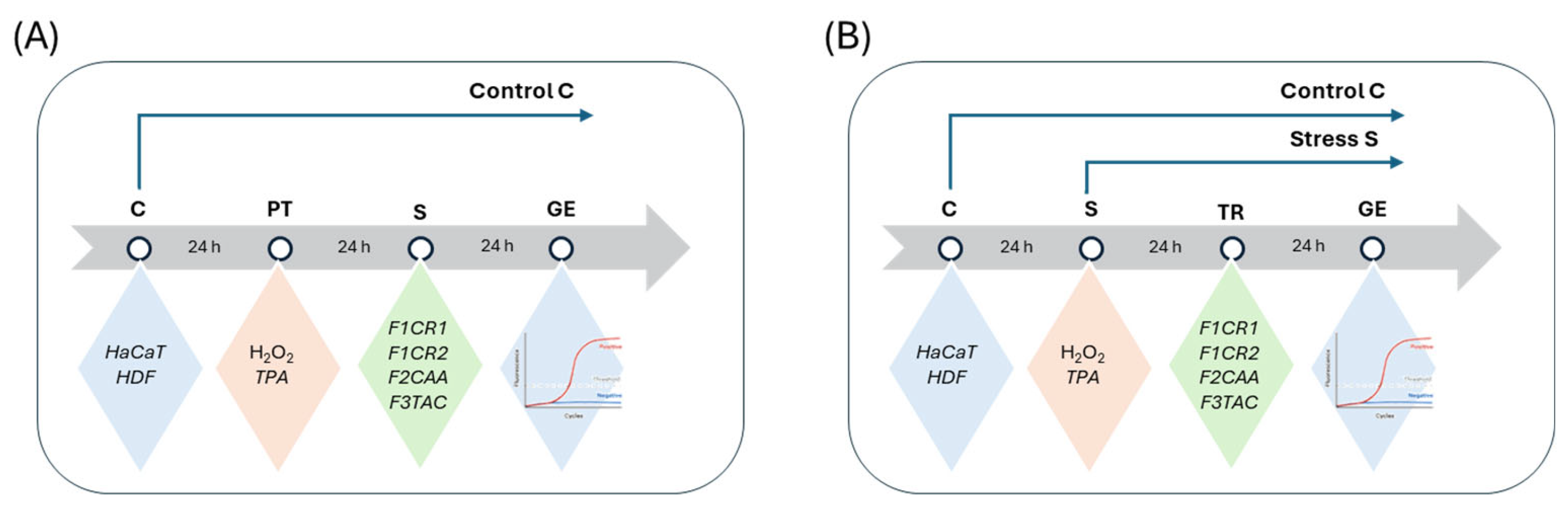

Table 1.
Effect of F1CR1, F1CR2, F2CAA, F3TAC formulations on the pathways suggested from the in vitro assays.
Table 1.
Effect of F1CR1, F1CR2, F2CAA, F3TAC formulations on the pathways suggested from the in vitro assays.
| pCBs | Pro-inflammation | Anti-Inflammation | Structural | Cann-receptor | |||||||
| CBD | CBG | CBC | PT | T | PT | T | PT | T | PT | T | |
| F1CR1 | ✓ | ✓ | HaCaT and HDF: anti-inflammatory against OS HaCaT: protective against inflammation from TPA stress |
HaCaT: complex regulatory action HDF: anti-inflammatory effect (dermis) HaCaT: no significant changes from TPA stress |
HaCaT: no significant changes from OS HDF: complex modulation of inflammatory effect |
HaCaT: modulation of inflammation and differentiation HDF: anti-inflammatory action |
HaCaT: modulation of ECM production and cell proliferation |
HaCaT: protection against tissue breakdown |
HaCaT: anti-inflammatory action HDF: enhanced wound healing |
HaCaT modulation of the inflammatory action HDF: medium protective action |
|
| F1CR2 | ✓ | ✓ | HaCaT:protective against OS, cellular repair HDF: anti-inflammatory effect in dermal layer HaCaT: protective in reducing inflammation (TPA stress) |
HaCaT and HDF: selective modulation of pro- inflammation and inflammation HaCaT: targeted effect from inflammatory stimuli (TPA stress) |
HaCaT: enhanced anti-inflammatory action HDF: complex modulation of anti-inflammatory signals in the dermis. |
HaCaT: inflammation modulation - shift HDF: anti-inflammatory response HaCaT: protective for inflammation (TPA-stress) |
HaCaT: protection against ECM breakdown, reduced fibrosis. HDF: modulation of matrix production and cell proliferation |
HaCaT and HDF: reduced fibrosis, protection against ECM degradation. HaCaT: tissue repair and protection against ECM degradation. |
HaCaT: enhanced anti-inflammatory and antioxidant effect HDF: enhanced cannabinoid signaling HaCaT: enhanced response to cannabinoid signaling and potential repair mechanisms. |
HDF: cellular proliferation and tissue remodeling | |
| F2CAA | ✓ | ✓ | ✓ | HaCaT: anti-inflammatory and regeneration action for OS and TPA stress HDF: anti-inflammatory effect |
HaCaT: complex modulation of inflammation HDF: anti-inflammatory effect HaCaT: selective modulation of inflammation |
HaCaT: anti-inflammatory action for OS HDF: selective modulation of inflammation |
HaCaT: modulation of inflammation and metabolic pathway HDF: protection against OS HaCaT: protection against inflammation (TPA stress) |
HaCaT: ECM protection and reduced fibrosis under OS HDF: ECM production. |
HaCaT: protection against tissue breakdown HDF: complex modulation of ECM production and protection HaCaT: tissue repair and protection (TPA stress) |
HaCaT: enhanced anti-inflammatory signaling HDF: cell-specific modulation |
HaCaT: modulate excessive signaling post-stress. HDF: protective effect |
| F3TAC | ✓ | ✓ | HaCaT: selective modulation of inflammatory pathways HDF: anti-inflammatory effect in dermal layer HaCaT anti-inflammatory action (TPA stress) |
HaCaT: limited efficacy HDF: selective modulation of inflammatory responses HaCaT: selective modulation of inflammatory pathway |
HaCaT: moderate anti-inflammatory support HDF: complex modulation of anti-inflammatory pathways |
HaCaT: limited post-stress effects HDF: stronger impact on anti-inflammatory pathways HaCaT: no significant changes, stress-specific modulation (TPA stress) |
HaCaT: enhanced ECM protection. HDF: reduced ECM production and remodeling |
HaCaT: enhanced protection against ECM degradation HDF: reduced ECM production HaCaT: enhanced ECM production and protection against degradation |
HaCaT: enhanced ECS activation HDF: protective mechanism |
HaCaT: selective modulation HDF: enhanced ECS activation |
|
Table 2.
Stress inducers tested for each human cell line.
| Human cell line | Stress inducers | Range of concentrations |
|---|---|---|
| HaCat | H2O2 | 50-400 μM |
| TPA | 5-20 μM | |
| HDF | H2O2 | 50-400 μM |
Table 3.
Cannabinoid composition per formulation F1CR1, F1CR2, F2CAA, F3TA.
| Formulations | CBD | CBG | CBC |
| F1CR1 | ● | ● | |
| F1CR2 | ● | ● | |
| F2CAA | ● | ● | ● |
| F3TAC | ● | ● |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



