
Submitted:
31 October 2024
Posted:
01 November 2024
You are already at the latest version
Abstract
As one of the first live vaccines, Bacillus Calmette-Guérin (BCG) is experiencing a experiencing its second rise. The recent COVID-19 pandemic has spurred interest in BCG due to its ability to provide protection not only against mycobacteria, but also against unrelated pathogens. This ability is based on BCG's ability to non-specifically activate cells of the innate immune system, bringing them into a prepared state where they can be activated more quickly. The process known as trained immunity. This ability of BCG to elicit an immune response gives it the opportunity to become not only a basis for universal or pan-pandemic vaccines, but also a cancer vaccine. We have reviewed the main mechanisms underlying the immunoactivating effect of BCG and proposed ways to enhance this effect. Since the pandemic has greatly altered the research landscape in vaccinology and, to some extent, cancer immunotherapy, we have focused more on studies published during and after the pandemic, minimizing information from the pre-pandemic period where possible. We discuss the recent trend in the use of BCG-based therapies, with particular emphasis on ongoing clinical trials. Based on the data presented, we suggested the most optimal options for further research to develop effective BCG-based vaccines.
Keywords:
Bacillus Calmette-Guérin
; BCG
; vaccine
; adjuvant
; cancer
; pandemic
; trained immunity
; clinical trials
1. Introduction
The immune system (IS) is a barrier that protects the organism not only from external factors, but also from internal foes. External influences are mainly pathogens, while internal ones are the result of errors in cell proliferation and differentiation. IS cells undergo a complex selection process learning to distinguish self from non-self and to respond appropriately to danger. The most ancient innate immune system is activated first, and is able to identify invaders non-specifically, destroy them, and pass the baton to the cells of the adaptive immune system, which can recognize invaders specifically and remember how to act against them. Such a synchronous duet allows the organism to be better prepared for the impact in the future. To be trained.
This property of the immune system has been the basis for the development of vaccines - drugs that mimic the pathogen and can train the immune system to fight a real pathogen. Similar properties are used in the antitumor immunotherapy approach, where IS cells are trained to recognize tumor neoantigens. To recognize, to be activated, to respond, to remember - this is the common basis of IS action. These pathways are similar, which allows us to try finding some basic universal activators for IS. One such activator can be Bacillus Calmette-Guérin (BCG) - a conditional pathogen that can teach IS how to act in the face of real danger. Here we will discuss the properties of BCG and the signaling pathways it triggers, which may allow it to become the basis for creating a pool of vaccines. We will consider the concept of pan-pandemic or universal vaccines that could be used in future pandemics of a pathogen of unknown etiology, and we will look at BCG-based cancer vaccines - highlighting the common features of BCG action and the possibilities for improving such drugs. In addition, we will consider how BCG is being used in current clinical trials - in which combinations and for which conditions. Based on the analysis of the data presented, we will outline what we expect to be the most promising research directions for the development of effective BCG-based drugs.
2. Towards Concept of Pan-Pandemic Vaccines
2.1. The Beginning
Tuberculosis is the ninth leading cause of death worldwide, according to experts. Tuberculosis (TB) remained the second leading cause of death from a single infectious agent worldwide in 2022, causing an estimated 1.30 million deaths and 7.5 million new cases of TB [1]. The spread of tuberculosis is widespread despite preventive measures. TB is an airborne infection caused by Mycobacterium tuberculosis (M. tuberculosis, Mtb), a member of the Mycobacterium group. These bacteria most commonly affect the respiratory tract and lungs due to their high tropism for the epithelium of the respiratory tract and alveoli [2]. Once in the human body, Mtb are phagocytosed mainly by macrophages (MΦ) and dendritic cells (DC), which then aggregate into structures called granulomas. The ability of bacteria to disrupt autophagy ensures their long-term intracellular persistence. The immunocompetence of patients is crucial for the development of TB, as groups with weakened immunity experience more severe manifestations of the disease [3]. Infants, persons with diabetes mellitus, HIV-infected patients, and patients with immunosuppression are considered the most susceptible groups of the population at risk of developing active tuberculosis [4]. Without treatment, the probability of experiencing TB during a lifetime is 5-10% [4].
The situation is complicated by the emergence of drug-resistant M. tuberculosis and HIV-TB co-infection, which has significantly worsened the prognosis and treatment of TB. In 2016, 490,000 cases of multidrug-resistant tuberculosis and 110,000 cases of rifampicin-resistant and isoniazid-sensitive TB were registered [5].
2.2. The Role of BCG in Modern Vaccination Against Tuberculosis and the Need of an Improved Vaccine
The first bacterial vaccine against human tuberculosis was developed more than 100 years ago. Bacille Calmette-Guérin (BCG), a live attenuated vaccine derived from Mycobacterium bovis, was first used in medical practice in 1921. Despite years of intensive research, BCG remains the only licensed, highly effective anti-tuberculosis vaccine [6]. It is still used clinically to protect against tuberculosis, especially in children. Universal recommendations for the BCG vaccination are widespread in most countries [7]. The World Health Organization (WHO) recommends that newborns in areas with a high incidence of tuberculosis and/or leprosy be vaccinated with a single dose of BCG vaccine. Some studies have shown that vaccination at birth is characterized by greater induction of CD4+ and CD8+ T cells expressing interferon γ (IFNγ) and proliferating cells capable of producing IFN-γ, tumor necrosis factor α (TNFα) and interleukin 2 together, compared to delayed vaccination [8]. Thus, different patterns of T-cell induction may occur depending on when the BCG vaccine was administered. However, results may depend on geographic and population heterogeneity.
This is consistent with other findings suggesting that BCG vaccination at birth is effective in preventing tuberculosis in infants but ineffective in adolescents and adults. Immunoprotection therefore needs to be boosted in older populations [9]. The effectiveness of the vaccine varies from 0 to 80% [10]. The main adverse reactions with conventional intradermal administration of the vaccine include itching, redness, swelling and other signs of inflammatory reaction [11]. To avoid complications, efforts have been made to convert the existing intradermal vaccine into an oral formulation [12]. Oral administration is cheaper, does not require specific medical skills, pharmacokinetics are easier to predict and provides greater patient compliance [13]. However, the direct effect of the vaccine on the mucosa is considered to be highly immunogenic, therefore the development of an oral vaccine capable of interfering with the production of gastric juice, pH-dependent degradation of antigens in the stomach and intestines, the low permeability of the mucus barrier, limits its development [14,15]. Nevertheless, the development of various methods of administration of BCG vaccine continues in many research groups [16].
As a result of numerous studies in the field of tuberculosis vaccinology, many new vaccine candidates have emerged and are in clinical and preclinical trials, with the aim of either replacing the old vaccine or increasing the effectiveness of BCG. However, there are no clear data on the efficacy of any of the new TB vaccines, and the possibility of their use is currently under investigation [17].
2.3. Lessons from COVID-19 and Heterologous Protection Against Other Pathogens
The COVID-19 pandemic struck the modern world by sudden - the development of vaccines against a new, previously unknown pathogen required time that the healthcare system did not have, as it may not cope with the number of cases. The period from the emergence of a new pathogen capable of causing a pandemic to the appearance of specific vaccines was a time of searching and observing for drugs that could somehow help in the control of the disease.
2.3.1. Heterologous Protection Against Pathogens Induced by BCG
Previously, a number of researchers have demonstrated the great potential provided by the use of BCG against heterologous infections. It was found that vaccinating infants against tuberculosis with BCG led to a decrease in the overall level of infant mortality not caused by tuberculosis [9]. In other words, the BCG vaccine had some protective effect that created a non-specific defense barrier against other non-tuberculous infections [18]. One of the reasons for the development of such an effect, which is not directly related to the vaccination pathogen, is the activation of the first line of defense of the innate immune mechanisms. This phenomenon, known as “training” of innate immunity, can lead to an increased readiness of immune cells to respond to various pathogens, not necessarily associated with a specific vaccine antigen. Thus, trained innate immunity (TRIM) is characterized by its ability to confer cross-protection against various infectious diseases following stimulation by bacteria, fungi, viruses, and other pathogens. This protective effect safeguards against a number of unrelated pathogens [19,20].
The use of BCG has demonstrated the important role of innate immune cells in protection against acute respiratory tract infections in people of all ages, including infants and the elderly. For example, in Guinea-Bissau, measles-naïve infants with acute lower respiratory tract infections were more likely to be BCG-negative and have no BCG scar [21]. In elderly people (60-75 years), a significant reduction in the incidence of acute upper respiratory tract infections and an increase in IFN-γ and IL-10 levels were shown in the BCG group relative to placebo [22]. Subcutaneous BCG vaccination enhances innate protection against pneumococcal (Streptococcus pneumoniae) infection by increasing pulmonary neutrophils, independent of circulating monocytes. These results may help in the development of new effective vaccination strategies against unrelated bacterial respiratory pathogens [23].
In addition to its effect on protection against respiratory infections, the protection provided by trained innate immunity against other viruses and bacteria is being studied extensively. Some strains of BCG not only protect against TB disseminated disease, but also reduce overall mortality by providing protection against infections caused by a variety of pathogens. Experimental studies in mice have confirmed the protective effects of BCG against several viruses, including herpes and influenza [24].
Several studies have shown that BCG immunotherapy has protective efficacy in the immunotherapy of warts [25]. In a study of patients with viral warts, patients with negative tuberculin test results received intradermal BCG vaccination, while those with positive results received standard treatment with salicylic acid. A complete response to treatment was observed in 9.8% and 5.3%, respectively. The use of BCG demonstrated higher cure rates for genital (22.2% vs. 7.7%) and common warts (8.5% vs. 0%) [26].
The immune response and protective efficacy of BCG vaccine against controlled malaria infection were evaluated. After malaria infection, BCG-vaccinated individuals showed earlier and more severe clinical manifestations, as well as significantly earlier appearance of NK cell activation markers and a tendency for earlier monocyte activation. BCG vaccination was shown to alter the clinical and immune response to malaria, which opens possibilities for the development of malaria vaccines [27].
BCG protects against experimental infection with an attenuated strain of yellow fever virus. The reduction in viremia correlated with an increase in IL-1β, which is associated with trained immunity. Thus, BCG induces epigenetic reprocessing of monocytes that provides protection against unrelated viral infections, with IL-1β playing a key role in this process [28].
These data were crucial for the design of vaccination programs and future vaccination strategies. In this regard, it is worth noting the potential significance of the spread of BCG vaccination, since the basis of the non-specific protective effect of BCG is the phenomenon of trained immunity, which transfers the immune system to a more prepared phase due to the reprogramming of some cells of the immune system. Such an effect of BCG vaccination can allow it to perform the function of temporary control of infection until the emergence of specific vaccines. This approach was tested during the COVID-19 pandemic.
2.3.2. Ambiguity of COVID-19 protection data
Early evidence suggested that BCG vaccination induced a trained immunity that provided significant protection against a variety of viral infections. It was also hypothesized that BCG might reduce the incidence and severity of COVID-19, but randomized clinical trials were needed to confirm this hypothesis. Data analysis showed that countries with mandatory BCG vaccination had lower COVID-19 mortality, but causality was not established and needed to be confirmed [29].
At the beginning of the pandemic in the 2020s, quite a number of publications appeared giving some hope for the positive effect of BCG vaccination in preventing severe COVID-19 infection. More than a dozen clinical trials in different populations were conducted to study the effect of BCG vaccination on COVID-19 cases [19]. An early study in 2020 observed a correlation between the degree of country-wide BCG vaccination and COVID-19 mortality in several socially similar European countries. The authors concluded that a 10% increase in BCG vaccination was associated with a 10.4% decrease in COVID-19 mortality [30]. Recent BCG vaccination has been shown to be safe and does not lead to an increase in respiratory symptoms during the COVID-19 outbreak [31].
In a statistical analysis of the first 30 days of COVID-19 outbreaks by country, the incidence of countries that had mandatory BCG vaccination until at least 2000 was compared with countries that did not [32]. The models used in the study showed a significant effect of mandatory BCG policy on the growth rate of COVID-19 incidence. The authors believe that BCG vaccination can only be effective in the presence of herd immunity, which does not allow the virus to spread easily among the population. Thus, the work suggests a positive role for mandatory BCG vaccination, although it should be taken into account that the BCG vaccination studied in the work was performed long before the pandemic [32].
The incidence of SARS-CoV-2 infection was studied in a large cohort of healthcare workers in Los Angeles. Nearly one-third of the study volunteers had been previously vaccinated with BCG. These workers had a significantly lower number (30-40%) of confirmed COVID-19 diagnoses, defined by positive PCR tests and seropositive tests for anti-SARS-CoV-2. BCG vaccination was associated with a lower incidence of COVID-19, despite the age of the patients and the presence of a large number of comorbidities. COVID-19-related symptoms were also significantly less severe in the BCG-vaccinated group than in the unvaccinated group. Such beneficial effects have not been observed after administration of other vaccines, such as influenza, meningococcal or pneumococcal vaccines; the effect described above appears to be unique to BCG immunization [33].
Although epidemiologic studies have shown that countries with mandatory BCG vaccination have lower COVID-19 mortality, there is still insufficient evidence to confirm or refute this hypothesis due to some systemic biases [19,34]. Thus, some recent studies indicate that there is insufficient evidence to confirm the efficacy of BCG vaccination in preventing COVID-19 infection and severe forms of COVID-19, to avoid overestimating the role of BCG vaccination, leading to its misuse [35]. To evaluate the impact of BCG on COVID-19 morbidity and mortality among healthcare workers in the Western Cape, South Africa, a study was conducted with 1000 participants, median age 39 years. Hospitalization for COVID-19 occurred in 15 participants, the majority in the BCG group. However, no statistically significant protection was observed. Thus, BCG vaccination did not protect healthcare workers from SARS-CoV-2 infection and severe forms of COVID-19 [36].
Clinical trials (NCT04327206, NCT04328441) were conducted to evaluate the efficacy of BCG in reducing the incidence and severity of COVID-19 in healthcare workers in the Netherlands and Australia. These studies NCT04328441 [37] found no significant differences between participants receiving BCG and placebo in rates of COVID-19 incidence, hospitalization, or disease severity [38].
At the same time, other authors suggest that trained immunity obtained as a result of BCG vaccination has probably reduced the incidence of COVID-19 in some populations [20,39]. The ACTIVATE-2 study was initiated to evaluate the efficacy of BCG vaccination against coronavirus infection COVID-19. In a multicenter, double-blind trial, 301 volunteers over 50 years of age were evenly distributed to receive BCG vaccine or placebo. The primary endpoints of the study included the incidence of COVID-19 and the presence of antibodies to SARS-CoV-2, which were monitored for 6 months after the intervention. The results showed a relative reduction in the risk of developing COVID-19 in the BCG-vaccinated group compared to placebo. Of the 8 patients hospitalized with COVID-19, 6 were in the placebo group and 2 were in the BCG group. Antibodies to SARS-CoV-2 were detected in 1.3% of subjects in the placebo group and 4.7% in the BCG group at 3 months post-intervention. Although the primary endpoint of reducing the risk of COVID-19 at 3 months was not met, the secondary endpoint of reducing the risk at 6 months was met. The data suggest that BCG vaccination may be a promising approach in the control of the COVID-19 pandemic [40].
A Phase III clinical trial conducted late in the pandemic evaluated whether a multidose BCG vaccination provided protection against COVID-19 and other infectious diseases in American adults with type 1 diabetes. From April 2021 to November 2022, the Tokyo strain BCG vaccine provided significant protection against COVID-19 and strong protection against all infectious diseases. The efficacy of BCG vaccination was not affected by concomitant COVID-19 vaccination [41].
2.3.3. Possible Reasons for Ambiguous Data
There may be several reasons why the data on BCG vaccination may differ in inducing a non-specific immune response: BCG is not a perfect vaccine and has two major limitations: its efficacy against tuberculosis varies considerably between pediatric and adult populations, and it can cause disseminated BCG disease in immunocompromised individuals. There are several substrains of BCG that are genetically distinct from each other, but the impact of these differences on vaccine efficacy remains largely unclear.
A comparative analysis of the virulence and efficacy of 13 BCG strains representing different genetic lines was performed in SCID and BALB/c mice. The results showed that the DU2 IV group strains (BCG-Phipps, BCG-Frappier, BCG-Pasteur, BCG-Tice) have the highest virulence, while the DU2 II group strains (BCG-Sweden, BCG-Birkhaug) have the lowest virulence. These differences may be due to specific duplications and deletions of genomic DNA. More virulent BCG strains usually show greater protection against Mtb infection [42]. In addition, a recent study showed that the season of vaccination also matters. BCG vaccination in winter resulted in a greater increase in proinflammatory cytokine production by peripheral blood mononuclear cells (PBMC) upon stimulation with various bacterial and fungal antigens compared to spring. In contrast, winter BCG vaccination was associated with lower levels of IFNγ release in PBMC. NK cells from winter-vaccinated individuals showed a greater capacity to produce proinflammatory cytokines and IFNγ upon heterologous stimulation. Vaccination had little effect on the monocyte transcriptome at 3 months. In contrast, seasonal epigenetic changes were detected in monocytes and NK cells, which partly explain the increased reactivity of immune cells in the winter-vaccinated group. These results suggest that BCG vaccination in winter promotes a more robust trained immune response through activation and reprogramming of immune cells, especially NK cells (Dutch clinical trial registry no. NL58219.091.16) [43].
Thus, the analysis of clinical trials conducted in a number of countries brings additional clarity to the question of how BCG can be used in a pandemic and what role it may play in future pandemics, especially in low- and middle-income countries where infrastructure and access to vaccines are underdeveloped. Economically and technically, the potential use of BCG is an excellent solution in terms of epidemiological safety [44]. In this vein, we would like to point out the potential significance of the spread of BCG vaccination, since the non-specific protective effect of BCG is based on the effect of trained immunity, which moves the immune system into a more prepared phase by reprogramming some cells of the immune system. This effect of the BCG vaccination can perform the function of temporary containment of the infection until the advent of specific vaccines.
2.4. Trained Immunity and Unspecific Activation of the Immune System after BCG Administration
Until recently, it was considered that only the adaptive arm of the immune system was capable of developing an immunologic memory that would allow for more effective protection against secondary infections. It turns out that this is not entirely true. After primary stimulation, innate immune cells are able to change their state by introducing epigenetic marks into the promoters of genes necessary to initiate a non-specific immune response. When a new immune-activating stimulus arises, the cells are able to respond much more rapidly and with greater amplitude (Figure 1B). This phenomenon has been termed trained immunity. Recently, a large number of studies, including ours, have been written to investigate its mechanism in detail (see reviews in [19,20,45]).
In brief, the main pathway of BCG activation is based on the interaction of pattern recognition receptors (PRRs) of immune cells with their ligands, which primarily include surface TLR2, TLR4, the non-TLR receptor Dectin-1, endosomal TLR9 and the cytosolic sensor NOD2 (Figure 1A). Through their cascades, they trigger signaling pathways associated with 1) Translocation of transcription factors (TFs) into the nucleus to activate synthesis of inflammatory cytokines IL6/IL1b/TNFa/IL12 (NF-κB cascade); and synthesis of interferons (IFNs); 2) Metabolic rewiring - metabolic cascades primarily associated with NOD2 and Dectin-1 [46,47,48], acting through the AKT/mTOR pathway, activate glycolysis (recently reviewed in [49]). Initiation of the TCA cycle (Krebs/tricarboxylic acid cycle) and increasing levels of acetyl-coenzyme A (Ac-CoA) promote the acetylation of H3K27ac, resulting in epigenetic rewriting [19]. Metabolic intermediates such as acetyl-CoA, fumarate, succinate, nicotinamide adenine dinucleotide (NAD+), and mevalonate, which are formed as a result of this activation-induced metabolic reprogramming, regulate the epigenetic landscape [50]. The result of the above processes is 3) Epigenetic reprogramming of immune cells as a result of activation of histone modification processes, in particular acquisition of activating marks H3K27ac and H3K4me3, reduction of histone methylation levels (namely repressive mark H3K9me3 at promoters of genes of proinflammatory cytokines such as IL6, TNFα and IL-1β). Histone acetylation transfers chromatin regions necessary for triggering the immune response to a more open state [20]. The main effects induced by the interaction of the immune system with BCG are summarized in Table 1.
If the process of transcription factor translocation and consequent inflammatory activation is a response to infection (which BCG generally is), then the processes leading to epigenetic reprogramming are associated with the emergence of trained immunity (TRIM). The degree and strength of such processes affect how much faster and stronger the immune response can develop during a secondary infection. Thus, factors that inhibit the development of a response to pathogens may result in a failure to control microbial infection, thereby initiating systemic responses and aberrant inflammation [51], and most likely lead to impairment of TRIM development. Another significant factor influencing the optimal functioning of the immune system is the maintenance of an equilibrium between pro- and anti-inflammatory cells and signals, as well as the activation and resolution of the immune response [52]. This also has a critical impact on the development of TRIM [53].
It should be noted that the factors contributing to the development of TRIM are usually associated with the development of an inflammatory state of the immune system. In general, the emergence of trained immunity can be characterized as an inflammatory process that occurs in response to a primary pathogen stimulus, associated with inflammation and epigenetic remodeling of innate immune cells. Once the pathogen is eliminated, the inflammatory response must be rapidly resolved to allow tissue repair and regeneration. Ineffective resolution of the inflammation can lead to the development of chronic disease and tissue destruction [54,55]. Signaling through NF-kB plays an important role in this process [54]. At the same time, the promoters of inflammatory genes involved in this process enter a more activated state due to epigenetic remodeling and euchromatinization [50]. As a result, the IS is in a more prepared state and responds more quickly and robustly to secondary infection.
2.4.1. Impairing the Innate Immunity Training
As mentioned above, many factors can influence the development of TRIM, which subsequently leads to the development of a non-specific primary response during secondary stimulation by pathogens. Obviously, the simplest factor in the dysfunction of the immune system is the absence or improper functioning of the PRRs. Thus, factors that inhibit the development of a response to infection, such as NOD2 deficiency [51], can lead to impairment of TRIM emergence. Thus, NOD2-deficient patients showed impaired induction of trained immunity [122].
Certain cellular chemicals and metabolites may also have an inhibitory effect on TRIM formation (summarized in Table 2). Inhibition of the mTOR or glycolysis pathway with rapamycin, 2-deoxyglucose (2-DG), or metformin [136] during TRIM formation disrupts the production of pro-inflammatory cytokines and leads to a loss of protection against infection.
It is worth noting that factors associated with the formation of TRIM are in many cases factors that contribute to the development of tumors (Table 1, column “In cancer”), which, if we recall one of the old characteristics of a tumor as a “non-healing wound”, clearly links the pro-inflammatory phenotype with the pro-tumor phenotype. In the case of TRIM inhibitors, they possibly may act as anti-tumor agents. Thus, research in the field of TRIM can be used in the search for new anti-tumor targets.
2.4.2. Persistence of the Innate Immunity
It has long been unclear how some innate immune cells, which have a relatively short persistence in the blood, can provide TRIM. Recent studies have shown that trained innate immunity can arise in hematopoietic stem and progenitor cells (HSPCs) in addition to mature myeloid cells in the bloodstream. For example, administration of TRIM inducers to mice induced reprogramming of myeloid precursors in the bone marrow [140].
Functional reprogramming induced by BCG vaccination occurs at the level of myeloid progenitors in the bone marrow of mice, where BCG vaccination biases HSCs toward myelopoiesis using IFNγ signaling, resulting in the formation of trained differentiated cells [141]. An increase in the representation of open chromatin after BCG stimulation in mice is shown at the level of HSCs in the bone marrow [141].
Human studies have shown that the functional reprogramming induced by BCG vaccination also occurs at the level of myeloid progenitors in the bone marrow. Changes in the transcriptional programs of HSPCs in BCG-vaccinated individuals were observed 3 months after vaccination, associated with a bias toward myelopoiesis and granulopoiesis [142].
Therefore, some studies on the mechanisms of long-term induction of innate immune memory have focused on the memory response in bone marrow-derived immune progenitor cells, where the trained phenotype is transferred to circulating innate immune cells, the so-called central trained immunity [140,141,142].
Other research has shown that tissue-resident macrophages can also develop immune memory, referred to as peripheral trained immunity [143]. Peripheral trained immunity occurs after exposure of bone marrow progenitors to a specific stimulus for trained immunity, resulting in their reprogramming and differentiation into mature peripheral innate immune cells with an enhanced inflammatory response upon repeated stimulation. For example, increased secretion of IL-1β by peripheral mononuclear immune cells 3 months after BCG vaccination following secondary exposure to C. albicans [142].
2.4.3. Cells of the Immune System in the Formation of TRIM
Immunological memory in vertebrates is often entirely associated with the function of T and B lymphocytes. Recently, it has been hypothesized that enhanced and prolonged non-specific immune responses after an initial infection may also provide protection against reinfection. Several research groups have focused on identifying the cells that mediate the effects of trained immunity. Researchers have chosen experimental mouse models with a deficiency in adaptive immunity to test the protective effect of innate immunity.
NK cells isolated from naïve volunteers after BCG vaccination have been shown to increase IFNγ production 4-7-fold and two-fold increase the release of monocytic cytokines such as TNFα and IL-1β in response to unrelated bacterial and fungal pathogens. The enhanced functionality of circulating monocytes persisted for at least 3 months after vaccination and was accompanied by an increase in activation markers (CD11b, TLR4). These effects are mediated by the NOD2 receptor through an increase in H3K4 histone trimethylation. BCG vaccination results in increased survival of SCID mice, which is partially dependent on natural killer (NK) cells. The results suggest that NK cells may contribute to the non-specific (heterologous) beneficial effects of BCG vaccination [144].
Similar data have been obtained with other TRIM activators. For example, in a proof-of-concept study, mice lacking functional T and B lymphocytes were shown to be protected from reinfection with Candida albicans by monocytes. C. albicans and fungal wall β-glucans induced functional reprogramming of monocytes, resulting in enhanced cytokine production both in vivo and in vitro. Training required the β-glucan receptor Dectin-1 and the non-classical Raf-1 signaling pathway. Training of monocytes by β-glucans was associated with robust changes in histone trimethylation at H3K4, suggesting that epigenetic mechanisms are involved in this phenomenon. Functional reprogramming of monocytes, reminiscent of similar properties of NK cells, supports the TRIM concept and may be used to develop improved vaccination strategies [145].
Overall, it should be noted that one of the critical cellular components of TRIM are natural killer (NK) cells, which play a key effector function in innate immunity. However, some recent studies have revealed the involvement of a rather rare and unusual fraction of immune system cells in the formation of TRIM, namely γδ T cells. These cells are considered to be part of the innate immunity in which the specific T cell receptor (TCR) can function as a pattern recognition receptor [146]. The MIC molecule also serves as a ligand for an activating receptor belonging to the NKG2 family of C-type lectin-like receptors (NKG2D), which is also expressed on γδ T cells [147]. It has been shown previously that human γδ T cells expand in response to tuberculosis infection [146]. γδ T cells have been called the “bridge between innate and adaptive responses” [148] because most of them recognize ligands that are significantly different from the short peptides recognized by α/β T cells in the context of MHC class I or class II molecules. Thus, human Vδ2+ T cells (the predominant subset of γδ T cells) recognize small bacterial phosphoantigens, alkylimines, and synthetic amino-bisphosphonates, whereas Vδ1 T cells recognize stress-inducible MHC-related molecules such as MICA/B, as well as several other ligands. Functionally, γδ T cells rapidly produce a variety of cytokines and typically exhibit potent cytotoxic activity, including against many tumor cells. The proposed role of γδ T cells as a bridge between the innate and adaptive immune systems is based on the assumption that γδ T cells use their TCR as a pattern recognition receptor.
The in vivo induction of innate memory in γδ T cells following BCG vaccination in healthy volunteers was investigated using a combination of single-cell RNA sequencing technologies and functional immune assays. BCG did not alter total γδ T cell numbers and membrane activation markers. However, vaccination altered γδ T cell transcriptional programs and enhanced their responsiveness to heterologous bacterial and fungal stimuli, including lipopolysaccharide and Candida albicans, as evidenced by increased production of tumor necrosis factor and IFNγ weeks after vaccination. Adult human γδ T cells demonstrate the potential to develop a TRIM phenotype following BCG vaccination [149].
2.4.4. Interaction of Adaptive and Innate Immune Systems in the Formation of TRIM
Thus, BCG vaccination promotes the transition of IS to a prepared state. Recent research in the field of TRIM indicates that for maximum effectiveness, the interaction of both arms of immunity - innate and adaptive - is necessary, as seen in the example of γδ T cells. The interaction of the adaptive and innate immune systems with non-hematopoietic cells in tissues is key to the formation of strong TRIM, or antigen-agnostic protective immunity, as some researchers call it.
Key points in the interaction of innate and adaptive immunity include the following mechanisms:
- Primary immune recognition by innate IS cells via PRRs and other “danger signal” receptors triggers activation of antigen-presenting cells (APCs), which interact with naïve CD4+ and CD8+ T cells to bridge the innate and adaptive immune systems [150].
- APCs initiate four critical signals for the induction of the adaptive immune response: 1) engagement of the T/B cell receptor (TCR/BCR) by antigenic peptides on the major histocompatibility complex (MHC), 2) ligation of immune checkpoint molecular pairs (co-stimulation and co-inhibition, e.g., CD80, CD86 and CD40), 3) cytokine stimulation, 4) recognition of “danger signals” by metabolic sensors [150,151,152].
- Activated immune cells undergo metabolic reprogramming characterized by increased glycolysis and decreased oxidative phosphorylation, leading to increased ATP production and pro-inflammatory functions. This metabolic shift is associated with epigenetic changes such as increased acetylation and decreased methylation, which contribute to the pro-inflammatory state [153].
- Metabolic reprogramming results in increased production of reactive oxygen species (ROS) [150] and activation of the NF-κB pathway, which enhances the innate immune response and promotes the differentiation of DCs into APCs. In this context, ROS are required for T and B cell activation, differentiation and survival, modulation of T cell subset differentiation, Treg functionality and maintenance of T cell homeostasis. ROS affect the expression of MHC and immune checkpoint molecules on APCs, thereby enhancing T cell activation [150].
- The interplay between APC activation and immune checkpoint responses influences the differentiation of different immune cell subsets [150]: monocytes can differentiate into anti-inflammatory (CD14+CD16- monocytes in humans, CD11b+Ly6C- monocytes in mice, CD14+CD40- monocytes and M2/Mhem macrophages), secreting anti-inflammatory cytokines (e.g., IL-10), leading to tissue repair, and pro-inflammatory (CD14+CD16+ monocytes in humans, CD1b+Ly6C+ monocytes in mice, CD14+CD40+ monocytes, M1 and M4 macrophages), which promote tissue inflammation by producing TNFα, IL-1β, ROS and other inflammatory mediators [154]
- Naïve CD8+ T cells can be activated into cytotoxic T lymphocytes to kill infected and tumor cells. In contrast, naive CD4+ T cells can differentiate into a variety of effector T cells, including proinflammatory Th1/2/9/17/22 subsets, anti-inflammatory regulatory T cells [155].
Prof. Pulendran’s research group has proposed that interactions between the adaptive and innate immune systems and non-hematopoietic cells in tissues are key to so-called antigen-agnostic protective immunity [156,157].
The researchers assumed that adjuvants had great potential to increase the effectiveness of vaccines. Adjuvants are components of subunit vaccines that enhance the magnitude, breadth and duration of the immune response. They have great potential to increase the effectiveness of vaccines, their addition allows to reduce the antigen amount without compromising immunogenicity, while reducing the adverse events associated with the antigen [158,159]. Thus, it was shown that TLR7/8 agonist - 3M-052, induces long-term humoral immunity in non-human primates, currently the obtained effect is being evaluated in clinical trials and in humans. It was found that a compound agonist formulated in alum to produce 3M-052-Alum for ovalbumin immunization (OVA) induced a potent program of antiviral and interferon responses. Activation of myeloid cells in draining lymph nodes persists until day 28, and single-cell level analysis reveals putative gene regulatory programs based on transcription factors in different myeloid cells and monocyte heterogeneity[160].
Subsequent work showed that mice vaccinated with intravenous BCG experienced less weight loss and/or better viral clearance when challenged with coronavirus (SARS-CoV-2 B.1.351) or influenza virus PR8. Protection began between 14 and 21 days after vaccination and lasted for approximately 3 months. Notably, BCG induced a biphasic innate response and potent antigen-specific Th1 responses in the lung. MyD88 signaling was required for innate and Th1 cellular responses and protection against SARS-CoV-2. Deletion of CD4(+) T cells or IFNγ activity prior to infection completely abolished innate activation and protection. Single-cell and spatial transcriptomics revealed CD4-dependent expression of IFN-stimulated genes in myeloid and lung epithelial cells. Notably, BCG also provided protection against weight loss following infection with mouse-adapted coronaviruses such as SARS-CoV-2 BA.5, SARS-CoV, and SHC014. Thus, BCG induces an integrated organ immune response in which CD4(+) T cells target myeloid and epithelial tissue cells to generate long-lasting and broad innate antiviral resistance [157].
Based on previous findings that the BCG vaccine induces heterologous immunity via CD4 T cell-derived IFNγ, which imprints an antiviral state on the innate immune system and epithelial cells, a vaccine containing ovalbumin antigen and GLA (TLR-4 agonist)/3M-052 (TLR7/8 agonist) adjuvants, which induce IFNγ responses, was developed. In vaccinated mice, antigen-stimulated T cells imprinted long-lasting and broad innate antiviral resistance in myeloid and epithelial cells, protecting the mice against SARS-CoV-2 and influenza viruses unrelated to the challenged antigen.
Thus, the authors introduce the concept of “integrated organ immunity,” which describes the interaction between the innate and adaptive immune systems, as well as non-hematopoietic cells in tissues, that allows the formation of robust and long-lasting protective immunity against a variety of pathogens, regardless of their specific antigens. By viewing immune responses through this integrated lens, the researchers propose the development of a new class of vaccines, known as “universal vaccines”, that would not be restricted to specific pathogens [156]
2.5. The Pan-Pandemic Vaccine Concept
Climate change is believed to be contributing to an increase in the number of potential pandemics, along with continued population growth, with potentially catastrophic economic and social consequences. At the same time, the nature of the pathogens causing pandemics may vary, making it impossible to rapidly develop specific vaccines. For this reason, it is necessary to create a buffer period for the development of specific vaccination of the population. During this period, the simplest methods of immunity activation can be used - for example, the use of so-called PAMP adjuvants, such as outer membrane vesicles of gram-negative bacteria, lipopolysaccharide (LPS) and other PRR agonists that activate the innate immune response [39].
PAMPs (pathogen-associated molecular patterns) are sets of molecules associated with exogenous pathogens, such as bacterial LPS, endotoxins, peptidoglycans, RNA species, DNA, and flagellin [161,162]. The interaction of these molecules with PRRs triggers a response to “danger signals” within the cells and in their microenvironment. According to the danger theory, the main driving force of the immune system is the need to detect and defend against danger [163]. Recent results on the activation of the innate immune system by PRRs support the idea that tissue injury, various forms of cell death, as well as metabolic and nutritional sensors can modulate the innate immune system to activate adaptive immunity [164]. Adjuvants have been used previously for similar purposes. It is expected that the use of new adjuvants (PRR activators) in light of new concepts and expanding knowledge of innate immunity and systemic vaccinology will advance the research and development of new adjuvants for use in vaccines against COVID-19 and future pandemics, which is especially relevant in conditions where rapid and effective immune solutions are needed to protect the population [164]. However, the disadvantage of the above mentioned PAMPs adjuvants may be less effective TRIM activation, which could result in a transient response.
Another approach could be the development of so-called “universal vaccines”, which are able to achieve a stronger activation of TRIM and thus put the cells of the immune system on high alert for unknown pathogens. However, instead of the term “universal” vaccines, we would prefer to use the term “pan-pandemic” vaccines when talking about vaccines used during pandemics of pathogens of unknown etiology and aimed at enhancing non-specific immunity in the population based on TRIM. This term is more accurate because it does not mislead regarding the use of such vaccines as anticancer. Thus, pan-pandemic vaccines are vaccines for emergency use against an unknown pathogen, based on the activation of TRIM, designed to protect the population until a specific vaccine is developed.
The BCG vaccine is a very good candidate for the role of such a pan-pandemic vaccine since it is one of the most potent inducers of TRIM. Above, we considered examples of the use of BCG and its potential, noting that, for example, in the case of the spread of COVID-19, its usefulness caused serious debate and controversial opinions. We believe that the insufficiency of BCG vaccination for non-specific activity could be compensated by the synthesis of special factors in the composition of recombinant BCG (rBCG). The reliable safety and immunogenicity profile of BCG is the rationale for using it as a vector for the development of vaccines against other infectious diseases. By introducing genes into BCG whose products contribute to better activation and/or proliferation of innate immune cells, it will be possible to obtain rBCG that are more effective as a means of emergency prophylaxis against viral diseases. Such rBCG can be used to reduce the incidence of new infections and/or alleviate their consequences in situations where no other specific prophylactic means are available.
2.6. Reinforcement of Non-Specific BCG Protection
In general, BCG, as a typical mycobacterium, affects PRRs. In some cases, this effect is mimicked by PAMP adjuvants. However, BCG is more effective because it triggers a stronger response, which is reflected in the formation of TRIM. However, this may not be enough, and enhancing the action of BCG in various ways may have a greater chance of creating a truly pan-pandemic vaccine. Below, we consider several strategies for such enhancement of BCG action.
2.6.1. Booster Strategy as an Example
As noted above, the easiest way to enhance the immune response without immunotoxicity is to use adjuvants. Several studies have shown the effectiveness of combining adjuvants with attenuated vaccines, including BCG (see, for example, [39]). The use of adjuvants to increase the amplitude of non-specific immune response and more stable formation of TRIM (including in a larger number of cells initially). The simplest possibility is the use of alum. Thus, the combination of alum and BCG is partially used in the development of vaccines against leishmaniasis, mainly in African countries. This approach aims to create relatively inexpensive drugs in low-income countries that suffer most from this disease.
Thus, pilot studies conducted in Sudan showed that first-generation vaccines for the treatment of visceral leishmaniasis (VL) based on a combination of autoclaved Leishmania major (ALM) and BCG are safe, but do not exceed BCG alone in protective properties. Attempts were made to create an enhanced version of the vaccine by adding the adjuvant alum. 24 healthy volunteers received different doses of leishmanial protein in the ALM + BCG vaccine with aluminum. Side effects were minimal, and a reaction to the skin test for leishmania was observed in all group participants. The vaccine mixture was safe, induced a strong delayed type hypersensitivity (DTH) reaction in the study volunteers [165]. Similar trials were conducted in leishmanin-unresponsive children in the field conditions. The study included 544 healthy children who received a single injection of Alum/ALM + BCG or placebo. The observation period was 2 years with an interval of 6 months. Leishmanin skin test conversion (>or=5 mm) was shown in a significant proportion of participants after vaccination at different stages of observation. In the placebo group, these indicators were significantly lower. No significant increase in antibodies was found. Parasitological VL was confirmed in four patients in the placebo group. Presumably, multiple vaccinations may be required to achieve results [166].
In a study to identify a suitable adjuvant for a leishmaniasis vaccine, low Th2 cytokine and high IgG2 antibody responses were used. Groups of vervet monkeys were immunized with Leishmania donovani antigen (Ag) either alone or in combination with alum-BCG (AlBCG), monophosphoryl lipid A (MPL), or montanide ISA 720 (MISA) as adjuvants. Immunization was performed subcutaneously three times at 4- and 6-week intervals. The results showed relatively higher levels of IL-4 and IL-10 after immunization with MPL + Ag compared to AlBCG + Ag or MISA + Ag. Significantly higher IgG2 antibody responses were associated with AlBCG + Ag or MISA + Ag compared to MPL + Ag. It was shown that the use of AlBCG and MISA may be effective in the development of leishmaniasis vaccines [167].
However, there is no licensed vaccine for leishmaniasis available for human use yet [168,169]. It can be noted that the addition of alum slightly increased the protective effect of vaccination. Thus, some further studies in animals led to the assumption that an additional boost is required for an effective immune response [170].
2.6.2. Recombinant BCG
Initially, it was thought that BCG would be used as a carrier for the secretion of various immune-activating factors. Historically, recombinant BCGs were primarily aimed at increasing the efficacy against TB. Thus, the 2018 article provides an overview of such studies and examines in detail the approaches used to create anti-tuberculosis rBCGs [171].
In terms of the possibility for developing pan-pandemic vaccines, rBCG may have the advantage of utilizing cytokines to enhance certain aspects of the immune response necessary for providing protection. This can be achieved by several approaches, the simplest of which is the use of cytokines that can activate a specific set of immune cells. We will discuss such approaches below, primarily using examples from the development of a more effective TB vaccine. The principles from these studies can be translated to improve the response of the immune system in general. The rational development of new vaccines that target key components of the host immune response is critical to the control of infectious diseases.
The development and analysis of modified BCG strains using the pMV261 vector and the M.bovis Ag85B secretory peptide has been intensively investigated in the group of Professor Triccas. The chemokine MCP-3, which binds to the CCR1, CCR2 and CCR3 receptors, was used [172]. These receptors are expressed on mononuclear phagocytes, T cells, B cells, NK cells and especially on immature dendritic cells. Mice lacking CCR2 are highly susceptible to Mtb infection and show a significant reduction in early macrophage recruitment and a later defect in DC and T cell migration [173]. Thus, chemokines may play a key role in the response to infection by being required for the migration of cells involved in generating a protective immune response. Mice vaccinated with BCG/MCP-3 showed increased lymphocyte migration and enhanced antigen-specific T cell responses compared to mice vaccinated with BCG alone [172].
In a similar study, the use of a modified BCG/mGM-CSF strain was shown to confer increased protection against Mtb. Vaccination with BCG/mGM-CSF resulted in a 10-fold increase in protection (protective effect) against disseminated Mtb infection compared to the BCG control. [174]. GM-CSF produced by subcutaneous administration of BCG/mGM-CSF increases the number of APCs in the draining lymph nodes, with the most significant increase occurring on day 7 after infection. After vaccination, a change in the number of cells with a MΦ phenotype (CD11c-CD11b-F480+) was observed, while no change in the number of other cells of the innate immune response was observed; there was no significant difference in the number of NK1.1+. It was shown that differentiated APCs under the influence of BCG’s GM-CSF became potent producers of IL-12p40, which contributed to the activation of naïve CD4+ T cells, confirming the above mentioned necessity of interaction between innate and adoptive immune cells. GM-CSF secreted by BCG increases the number of APCs in the areas draining the immunization site, but not in distant lymph nodes, i.e., the activation of APCs occurs only locally. It has also been shown that BCG/mGM-CSF has a long-term effect on the activation and maintenance of Ag-specific T cells generated as a result of vaccination.
BCG is able to stimulate functional CD8 T cells, suggesting that efficient delivery of Ag to sites of T cell activation by vaccines may be a key requirement for an optimal CD8 T cell response to control mycobacterial infection [175].
A study of targeted delivery of the rBCG-expressed cytokine mGM-CSF to the lungs showed an increase in the number of pulmonary DCs and secretion of the immunoregulatory cytokine IL-12 compared to BCG immunization. This effect of BCG/mGM-CSF on the number of APCs led to accelerated priming of antigen-specific CD4+ T cells in mediastinal lymph nodes and increased migration of activated CD4+ T cells to the lungs. Intranasal administration of BCG/mGM-CSF significantly increased the protection of mice against Mtb infection compared to mice vaccinated with BCG alone. BCG/mGM-CSF vaccination demonstrated an improved safety profile and significantly increased the lifespan of the mice. Data show that manipulation of immune cells in the lung by administration of BCG-based mGM-CSF can promote protective mucosal immunity against pulmonary bacterial infection [176].
Thus, in animal studies, recombinant BCG vaccines expressing various viral proteins are likely to induce TRIM and enhance protection [177]. A vaccine expressing the S1 subunit of pertussis toxin provided 100% protection against intracerebral challenge with Bordetella pertussis in newborn mice [178]. rBCG-S1PT was also shown to protect mice against challenge with B. pertussis and C. albicans, and rBCG-S1PT reduced the allergic response to ovalbumin in asthmatic mice. rBCG-S1PT induces an enhanced inflammatory response in human mononuclear cells [179]. rBCG with HIV antigens induced a T-cell response, although it did not induce HIV-specific antibodies [180]. Combination of rBCG with viral vector boosters enhanced T cell responses against HIV [181]. A vaccine expressing the hMPV phosphoprotein reduced viral load and tissue damage [182,183,184]. rBCG containing the hRSV nucleoprotein provided protection against viral infection by inducing TH1/TH17 memory and neutralizing antibodies [185].
A recent study showed that mucosal vaccination with the recombinant rBCGPPE27 strain, which produces the mycobacterial survival-promoting antigen PPE27 [186], significantly enhanced the TRIM in mice, which contributed to a more effective protective response against Mtb and heterologous bacterial reinfection in mice compared to BCG. Mucosal immunization with rBCGPPE27 increased the production of innate cytokines by alveolar macrophages, which is associated with activated glycolytic metabolism typical of trained immunity. Interestingly, rBCGPPE27, rBCGESAT6 and rBCGRv3763 significantly increased the secretion of TNFα, IL-6 and IL-1β in alveolar macrophages, BMDMs or human monocytes compared to BCG [187].
It is described that mTORC2 and hexokinase 1 deficiency rescued immunometabolic and epigenetic rearrangements in mouse alveolar MΦ following mucosal vaccination with rBCGPPE27. Importantly, exploiting the pronounced TRIM effects of rBCGPPE27, a single mucosal immunization with rBCGPPE27-adjuvanted coronavirus disease (CoV-2) vaccine induced the rapid development of virus-specific IgG antibodies, increased the level of neutralizing antibodies against pseudoviruses, and enhanced the release of Th1-biased cytokines by vaccine-specific T cells compared to the BCG/CoV-2 vaccine. Mucosally administered rBCG was shown to induce pulmonary memory MΦs and enhance TRIM through retraining of mTORC2/HK1-mediated aerobic glycolysis, providing new vaccination strategies to improve vaccination against TB or coronavirus variants and to target innate immunity through mucosal surfaces [187].
A combination of two strategies, rBCG and an adjuvant, can also be observed. In a study conducted on K18-hACE2 transgenic mice expressing the human ACE2 receptor used by severe acute respiratory syndrome coronavirus (SARS-CoV) to gain cellular entry, rBCG carrying spike-derived immunogenic epitopes from SARS-CoV-2 rBCG-ChD6 was used in combination with recombinant nucleocapsid and spike chimera (rChimera) [188]. They represent epitopes of the Nucleocapsid (serine-glycine-arginine-rich domain (SGRD) and serine-arginine-rich domain (SRD)) and Spike (receptor binding domain (RBD)) structural proteins of SARS-CoV-2. Primary immunization of animals with rBCG-ChD6 followed by boosting with rChimera-Alum demonstrated the highest titers of IgG and IgG2c antibodies to rChimera with neutralizing activity against SARS-CoV-2 Wuhan strain compared to control groups [189]. IFNγ and IL-6 production in spleen cells and a decrease in lung viral load were also demonstrated. No viable virus was detected in these mice, which was associated with a decrease in lung pathology compared to the BCG WT-rChimera/alum or rChimera/alum control groups. Thus, we see the potential for combination vaccination strategies to be effective.
3. Cancer Vaccines
Tumor diseases can be characterized as the result of immune malfunction, occurring when the IS fails to recognize a newly formed neoplastic cell, allowing it to survive and, through a complex path of clonal selection that promotes immune evasion, develop into a tumor. The innate immune system is the first line of defense against malignant cells, with its cells designed to recognize and destroy any aberrant cells that differ from those that should be “here and now”. Cytotoxic NK cells, which are capable of eliminating tumor cells, and MΦs, which can perform phagocytosis, play the main role in the destruction of such cells. Destruction of cancer cells by cells of the innate immune system triggers adaptive immunity [190]. The inability of the immune system to ensure the elimination of neoplastic cells leads to tumor diseases and is associated with the inability of the IS to recognize a tumor.
3.1. Highjacking the Innate Immunity
One of the goals of anti-tumor therapy is to make the tumor visible to the immune system. Thus, the success of anti-tumor immunotherapy is based on the initiation or stimulation of a self-sustaining anti-tumor immune response. A number of sequential events leading to this have been summarized in the concept of the cancer-immunity cycle [191]. Initially, this cycle was considered in the context of the adaptive system, where tumor neoantigens are presented by antigen-presenting cells, leading to the priming and activation of a pool of cytotoxic CD8 T cells. However, further works discussed that the interaction of adaptive and innate immune cells may enhance antitumor efficacy. In our previous work, we considered such a possible enhancement in the context of the augmented cancer-immunity cycle [192]. This possibility was based on the properties of the innate immune system, which is activated when pathogens are encountered. As discussed above, from the immune system’s point of view, pathogens are PRR ligands that trigger a signaling cascade leading to the activation of innate immune cells. Since BCG acts as a pathogen (danger signal) that activates several types of PRRs, but is not harmful to an immunocompetent person, it appears to be an optimal candidate for the development of anti-tumor immunotherapies.
Notably, that in 1990, BCG became the first immunotherapy of any type to be approved by the FDA and is still used for the treatment of early-stage bladder cancer. It was in 1959 when Lloyd J. Old and colleagues reported the use of the BCG vaccine as an immunotherapeutic agent that could inhibit tumor growth in mice [193]. Studies have shown that although BCG therapy for bladder cancer has a negative impact on the quality of sleep of patients with non-invasive bladder cancer, this therapy still demonstrates positive efficacy in the treatment of this disease [194]. Today, the BCG vaccine is an FDA-approved treatment for bladder cancer, latest Guideline updates for Non-Muscle Invasive Bladder Cancer were amended at 2024 [195].
Other malignancies such as lymphoma and melanoma also respond to the BCG vaccine [196]. BCG has been shown to reduce the risk of melanoma [197] and can also be used to treat patients with stage III melanoma [198,199]. Another study showed that BCG is effective against kidney cancer and prostate cancer [200]. In addition, studies have shown that early BCG vaccination of infantes can help reduce the risk of cancer and mortality from tumor diseases such as leukemia and lung cancer [201,202]. The latter fact suggests that BCG vaccination may have a protective antitumor effect, potentially related to processes similar to those involved in vaccination.
3.2. TRIM in Cancer
Many studies have linked the antitumor effect of BCG to the induction of TRIM. It has been shown that immunoinducing TRIM agents, such as BCG and fungal β-glucan, are able to prevent the immunosuppressive tumor microenvironment (TME), enhance T-cell responses, and eventually lead to tumor regression [203]. Molecules produced in TME cells that act as PAMPs and DAMPs to train myeloid cells mediate the antitumor effect of BCG. DAMPs (damage-associated molecular patterns) are a type of “danger signals” more associated with cellular damage, representing extracellular proteins such as decorin and tenascin C, as well as intracellular proteins such as high-mobility group box 1 (HMGB1), histones, S100 proteins, heat shock proteins (HSPs) [204]. Transcriptional reprogramming of cells is a key feature of TRIM, which enhances the pro-inflammatory phenotype. Repeated pathogen administration induces the infiltration of MΦs, DCs and NK cells, as well as the production of cytokines, including NF-κB cascade cytokines and interferons. This disrupts tumor homeostasis and enhances antitumor immunity, leading to the activation of tumor-infiltrating T cells[19].
Thus, it has recently been shown that intravesical administration of BCG induces TRIM, which is reflected in increased production of TNF and IL-1β after heterologous stimulation of blood PBMC ex vivo after 6-12 weeks [205]. Only BCG-naive patients were included in this study. It is believed that TRIM is formed and is responsible for antitumor effects in a model of colon cancer [206], gastric cancer [207], squamous cell carcinoma of the head and neck [208], as well as in hematologic malignancies (cited by [19]).
3.3. Autophagy Perplexity
Many studies conducted in model cell lines have shown the induction of autophagy and apoptosis after exposure to BCG. For example, a study conducted on the gastric cancer cell line MGC-803 showed that BCG induced apoptosis and autophagy in a dose-dependent manner [207]. Autophagy plays an essential role in monocyte-macrophage differentiation [209]. M1 macrophages are known for their antitumor effect based on producing high levels of ROS in the activated state [210], they can also distinguish between tumor cells and normal cells [211]. Autophagy is thought to play a dual role in tumor suppression and promotion across various types of cancer. It is assumed that in the early stages of tumor formation, autophagy may be one of the ways to counteract tumor progression. However, in the advanced tumor process, autophagy helps to protect tumor cells by adapting them to stress conditions, lack of oxygen and nutritional factors [212].
Some researchers suggest that the use of autophagy regulators may play a therapeutic role, for which rapamycin and chloroquine can be used [213]. The anti-cancer role of rapamycin is well discussed [137,138]. Thus, the main target of rapamycin is mTORC1, it blocks the early stages of autophagy by phosphorylation-dependent inhibition of Atg13 and ULK1 [214,215]. When mTORC1 is inhibited, autophagy is enhanced. Inhibition of mTORC1 has been shown to increase autophagic degradation, reducing the demand for energy and nutrients. Therefore, rapamycin turns off mTORC1, which leads to the activation of autophagy. Remember that rapamycin is also a TRIM inhibitor (see section 2.4.1). It is clear that the complex interplay between autophagy, TRIM and antitumor activity is far from being understood.
There is also a point to be made here - there is evidence that BCG blocks autophagy in MΦs and may promote intracellular survival of mycobacteria. At the same time, autophagy is a critically important protective mechanism to clear MΦs of mycobacterial infection [216]. Simultaneously, preliminary induction of autophagy by various means might overcome the imposed block [216]. Thus, it has been shown that infection with alive or thermally inactivated BCG results in a strong inhibition of autophagy. However, when autophagy is induced in M. smegmatis in advance, this effect is not observed [217]. In co-infection with BCG and heat-killed M. smegmatis, an enhanced antigen-specific response of CD4+ T cells is observed, along with reduced mycobacterial survival. BCG can block phagosome maturation and reduce the level of MHCII expression of APCs, which affects T-cell activation. This effect can be circumvented by using a rBCG strain expressing the TLR2 activator, the C5 Mtb peptide, in combination with Ag85B. Such additional stimulation of PRRs results in the induction of autophagy in MΦs, which enhances MHCII-dependent antigen presentation [218]. Typically, such effects were not observed in TLR2 knockout mice. Thus, we see that enhancing the effect on one of the BCG activation pathways through various PRRs can enhance the effect of targeting the IS by enhancing the immune response and TRIM. Another approach to bypass the blockade of autophagy may be the development of recombinant strains in which fragments of the genome that can block the activation of autophagy are deleted [219]. In this case, genetic modification of BCG allows bypassing the factors limiting its use.
3.4. Merging the Antitumor Features of BCG
The potential mechanisms of action of BCG-based cancer vaccines could therefore be outlined as follows:
- Breaking cancer tolerance by inducing an innate immune response leading to theexpansion of the augmented cancer-immunity cycle, which may be at least partially based on:
- TRIM-inducing effect of BCG
- Enhancement of autophagy by specially designed rBCG
Thus, using BCG as a basis for cancer vaccines, we can potentially improve the immune response and extend its efficacy (e.g., antimetastatic). To achieve this, several strategies can be explored ranging from additional targeting of the pathways involved in the activation of innate IS to bypassing the inherent limitations of BCG.
4. A Brief Overview of the Current Clinical Trials That Use BCG
Scientific discoveries do not always reflect the contribution of a particular technology to clinical trials (CT). To determine which BCG technologies are currently most in demand in the clinic, we conducted a brief analysis of clinical trials using BCG. For this purpose, the parameters “Other terms” were entered into the СT database https://clinicaltrials.gov/: Bacillus Calmette-Guerin OR BCG/Vaccine OR Vaccination, yielding a total of 287 studies. Of these, 15 observational studies were removed. We excluded all trials that were Terminated (17) and Withdrawn (10). A total of 245 trials remained. Of these, 154 were “Completed studies” and were excluded from the analysis. Eight studies not directly related to BCG were also excluded, including those on other types of vaccines (such as oral polio vaccine, Prevenar, etc.) and social studies related to different types of patient reminders.
The remaining 83 studies were analyzed in detail to ensure that they specifically used certain types of BCG or utilized BCG as an adjuvant. From these studies, we excluded trials that included BCG non-responders (resistant or relapsing) or non-tolerant patients. We also excluded studies of the MTBVAC vaccine, a live attenuated strain of M. tuberculosis, and other subunit vaccines using BCG fragments. For the COVID-19 condition, we also excluded trials as irrelevant if more than 2 years had elapsed from the specified primary completion date (estimated). Thus, from the previously obtained list of 287 studies, we left only 62 CTs that interested us as a representative cross-section of modern ongoing CT on BCG. These studies were classified according to eight key conditions: Cancer, COVID-19, Diabetes, HIV, Immunity, Mortality, Neurodegenerative and Tuberculosis (See Figure 2).
Interestingly, the number of studies on cancer was dominant, with more than twice as many as studies on tuberculosis (28 versus 13). All other studies made up about 1/3 of the two most represented studies and were relatively evenly distributed. Below are brief comments on the CTs included in the analysis and their composition.
4.1. Cancer
The largest group combining the use of BCG in cancer research is predominantly Non-Muscle Invasive Bladder Cancer (NMIBC) and other forms of bladder cancer (about two dozen), which is not surprising given that BCG is the first immunotherapeutic drug for this type of cancer (see Chapter 2). There are about an order of magnitude less studies for various types of melanoma, prostate, colon, and breast cancer.
Thus we observe a fairly predictable distribution by nosology. Of greater interest in this group are the various ways of using BCG, ranging from its use as an adjuvant for other vaccines to its combination with different agents. It is worth noting that in this group exhibits the greatest variety of BCG use all the clinical trials included in the analysis. The most abundant combination of BCG is with PD1/PD-L1 immune checkpoint inhibitors (ICH). Thus, ICH therapy is one of the most promising and advanced immunotherapies, but it has the limitation only a subset of patients respond to this therapy [59,220,221]. The ICH/BCG combination can increase the proportion of such responders. Thus, BCG immunotherapy can be combined with other treatments, such as checkpoint inhibitors, to enhance the immune response against various cancers [222]. The efficacy of BCG can be enhanced by attaching tumor-specific peptides to its outer membrane, thereby inducing stronger T-cell-specific immune responses [223]. This novel approach, when combined with immune checkpoint inhibitors such as anti-PD-1, increases the proportion of successful responses to cancer therapy [223].
The next most common CTs are the use of BCG alone or in combination with any type of chemotherapy. The efficacy of using a combination of BCG with gemcitabine alone or gemcitabine + docetaxel (GEMDOCE), as well as in combination with mitomycin, an antibiotic with antitumor activity, has been shown [224,225]. BCG is also used as an adjuvant, in combination with cellular derivatives, and for antigen enhancement - in general, all these actions can be combined as designed to enhance the immune response against tumor neoantigens. Probably by TRIM induction as discussed above. Some studies include combinations with an oncolytic adenovirus, the beta-blocker propranolol, and even studies with recombinant BCG - VPM1002BC (See Table 3). VPM1002BC is based on the rBCG strain BC1002 originally developed for the treatment of tuberculosis and formulated for intravesical immunotherapy of non-muscle invasive bladder cancer. BC1002 is a recombinant live BCG strain in which the urease C gene is deleted by insertion of the gene encoding the hemolysin listeriolysin (LLO). The rBCGΔureC::hly construct was designed to facilitate the targeting of phagosomal antigens to the major histocompatibility complex I pathway, thereby enhancing the induction of CD4+ and CD8+ T cell-mediated immune responses. To achieve this, the listeriolysin (hly) gene from Listeria monocytogenes was integrated into the urease C gene (ureC). This insertion disrupts urease C activity, preventing the enzyme from maintaining pH stability within the host cell phagosome. As a result, the pH decreases, activating listeriolysin (LLO), which has peak activity at pH 5.5. LLO is a hemolysin that can perforate membranes, and its function within the phagosome enables the translocation of proteins into the cytosol of infected host or cancer cells [226].
4.2. Tuberculosis
This category covers studies conducted mainly in countries most affected by Mtb. Various types of studies on safety, revaccination and other ways to activate immunity against Mtb, mainly in patients previously vaccinated with BCG or compared to patients without such vaccination. The vast majority used different native BCG strains and two studies using recombinant VPM1002 (rBCG) (refer to Table 3).
4.3. COVID-19
An interesting situation has occurred with COVID-19: initially there were about 20 studies related to BCG in the search, but after analysis, only 3 studies remained relevant - one with rBCG and two others that have not yet changed their status to from “Unknown” to “Complete”, despite a considerable amount of time has passed. In Chapter 2.3 we discussed the situation with COVID-19 and BCG, and it is unlikely that these studies will be able to add anything new. In addition, the pandemic is over and the relevance of such data is no longer as critical.
4.4. Diabetes
The search included several diabetes-related clinical trials. For example, a 2018 survey showed that the BCG vaccine can reduce blood glucose levels in patients with type 1 diabetes mellitus (T1DM) to near-normal values, and this effect persists for a long time [229]. BCG restores gene expression in Tregs in type 1 diabetics to a pattern consistent with non-type 1 control subjects. In addition, the authors demonstrated in diabetic mouse models that BCG vaccination improves glucose metabolism by systemically switching from oxidative phosphorylation to aerobic glycolysis (a state with high glucose consumption).
A strategy of multiple BCG administration to patients with type 1 diabetes showed specific killing of some disease-causing white blood cells, as well as a short-term effect of restoring pancreatic insulin secretion. BCG also had a positive metabolic effect, increasing glucose uptake and steadily reducing the level of glycated hemoglobin in the subjects. For type 2 diabetes, a strategy of BCG administration to patients with pre-diabetes may be possible, with the aim of reducing chronic inflammation in hyperinsulinemia [230].
However, these studies are beyond the scope of this review (You may refer to Faustman Lab’s publications [231]). In addition, Chapter 2.4 outlines the mechanisms of glycolysis activation upon BCG administration.
4.5. HIV (Human Immunodeficiency Virus)
The studies are primarily concerned with the use of BCG vaccination in HIV-infected patients in terms of safety and efficacy. Exposure to various infectious diseases may influence early progression to AIDS. BCG vaccine is usually given at birth to most children, including those born to HIV-positive mothers. However, HIV-infected infants are at risk of developing disseminated Mycobacterium bovis infection, which may be caused by the vaccine. In addition, the BCG vaccine may activate immune responses that promote the spread of the HIV virus and accelerate the progression of the disease to AIDS. These clinical trials are aimed at investigating the possibility of such a phenomenon and its impact on the outcome of clinical conditions in this group of patients.
4.6. Immunity
Presented by diverse studies on the contribution of BCG to TRIM and heterologous protection. From the possibility of inheriting methylation profiles of activated immunity to protection against various infections, including when used as an adjuvant in the development of other vaccines.
4.7. Mortality
Studies of infant vaccination outcomes in relation to mortality, morbidity and mortality in hospitalization and other factors are presented. There is an association between BCG vaccination and a reduction in infant morbidity and early mortality during the first year of life for both normal weight and low birth weight infants. Vaccination of women of reproductive age with BCG vaccine before pregnancy aims to study the additional effects of early administration of the vaccine to their children on the development of specific and non-specific immunity and on the general health of mother and child.
Thus, recent studies have examined the non-specific effects of BCG vaccination on infant morbidity and mortality. While studies in Greenland [232] and Denmark [233] found no significant reduction in hospitalization rates due to infectious diseases among BCG-vaccinated children, other research has suggested potential benefits. A study in Spain reported lower hospitalization rates for respiratory infections and sepsis in BCG-vaccinated infants, with protective fractions ranging from 32.4% to 69.6% across different age groups [234]. Similarly, a study in Guinea-Bissau found that early BCG vaccination reduced in-hospital mortality rates, particularly for sepsis cases, although it did not affect overall hospitalization rates [235]. BCG vaccination was associated with lower mortality among children aged 1-5 years in Uganda [236].
4.8. Neurodegenerative
This small group includes conditions associated with neurodegenerative processes and the potential use of BCG to treat such conditions. These mainly include studies in Alzheimer’s disease (AD) and radiologically isolated syndrome, a subtype of early stage of multiple sclerosis (MS) in which there are no symptoms or outward signs of the disease, but MRI results of the brain and spinal cord show foci of MS. The studies are based primarily on pilot data on the potential of BCG to affect such conditions.
Thus, recent studies suggest that BCG immunotherapy for bladder cancer may reduce the risk of AD. Multiple retrospective analyses of bladder cancer patients found that those treated with intravesical BCG had a significantly lower incidence of subsequent AD diagnosis compared to untreated patients [237,238,239]. The mechanism may involve BCG-induced modulation of the immune system, particularly through elevated IL-2 levels and Treg cell expansion, which could inhibit AD-associated inflammation [237]. While these findings are promising, they are based on retrospective data, and prospective studies are needed to confirm the potential of BCG as a preventive measure against AD in the general population [237,238].
As can be seen from the above, the most relevant studies for clinical trials are those for antitumor therapy, closely followed by clinical trials for the treatment and prevention of tuberculosis. These are the most obvious and accessible applications of BCG based on its properties as a conditional pathogen and TRIM activator. However, researchers are trying to exploit all possible aspects of BCG. For example, BCG’s ability to activate glycolysis can be used to combat or prevent diabetes. Such clinical trials using BCG to combat diabetes and neurodegenerative diseases highlight the potential broader impact of BCG vaccination beyond tuberculosis prevention and cancer immunotherapy. At the same time, studies related to the possibility of creating so-called pan-pandemic vaccines are very modestly presented, focusing mainly on preventing heterologous infections or reducing mortality from non-tuberculous infections in infants. In general, this is in line with the trend of pharmaceutical companies to look for a solution to existing threats. Research into the possibility of developing pan-pandemic vaccines is more in the realm of scientific research, but may be needed at any time in the event of a pandemic. Separately, it should be noted that recombinant vaccines are still poorly represented (only 4 studies, see Table 3), which may be due to some bias in the translation of research into clinical work.
5. General Considerations
Overall, despite some confusion in evaluating the efficacy of BCG vaccination as a non-specific vaccine against COVID-19, its role in activating various components of the immune system is beyond doubt. It can serve as a good basis for both pan-pandemic and anti-tumor vaccines. It has been found that the degree of TRIM and antitumor effect can be improved by modifying BCG to express higher levels of key PAMP molecules [240]. At the same time, researchers expect that TRIM can help achieve long-term therapeutic benefits against cancer [203]. Pharmacological compounds capable of activating or inhibiting TRIM regulatory pathways offer new opportunities to modulate specific and non-specific vaccine effects. For example, metformin inhibits TRIM induction by BCG, while experimental cancer vaccination studies have demonstrated its ability to enhance immune responses. We have noticed that factors that promote TRIM are often pro-tumorigenic (see Table 2, TRIM inhibitors). From this point of view, TRIM inhibitors can be considered as potential anti-tumor agents. At the same time, the opposite situation is possible if genes with tumor suppressor potential are involved in TRIM inhibition. For example, the transcription factor RORα, which is associated with circadian rhythms, inflammation and lipid metabolism, is suppressed in breast cancer [241]. It is thought that under certain conditions it may act as a tumor suppressor. It has been shown that RORα is able to block the epithelial-mesenchymal transition that promotes tumor progression [242,243,244], directly repressing Snail transcription [245]. Studies conducted this year have shown that RORα agonists are able to suppress TRIM that occurs during BCG induction, suggesting that RORα may be a TRIM inhibitor [246]
In general, we may summarize the BCG features as a base for pan-pandemic and cancer vaccines by action mechanisms as similar – impact primary on innate immunity, with certain bias towards TRIM activation and somehow controversial features as factors enhancing TRIM are in general pro-inflammatory and pro-tumorigenic. This principle can differentiate approaches to the development of BCG-based drugs. Despite the basic molecular mechanism of action being centered on the effect on PRRs, the subsequent effect of the drug on the signaling pathways that require enhancement for optimal efficacy may vary. Hence, despite the co-directionality and similarity of the immune system pathways, one should be careful when creating vaccines, especially cancer vaccines. Thus, some non-obvious things may occur, for example, from recent work that examine in detail the possibility of vaccinating different cohorts of patients. In a study by Moorlag et al., it was shown that individuals with condensed chromatin in genes involved in innate immunity prior to vaccination exhibited strong trained immunity induced by BCG and acquired open chromatin in these genes. At the same time, “non-responders” had initially open chromatin in similar regions prior to BCG vaccination, resulting in a lack of further enhancement of immune responses after vaccination [53]. Thus, in cancer, persistent inflammation may be detrimental to the formation of a long-term immune response because cells cannot switch from one activation state to another. This may represent another and less obvious problem of persistent inflammation during tumor progression as one of the types of immune response evasion. We may speculate that in cancer therapy, the BCG-induced response should preferably occur against the background of the introduction of secretory or additional anti-inflammatory cytokines, or precede such introduction. Indeed, persistent activation of some pathways in cancer leads to anergy and dysfunction of immune cells. TRIM is not formed, leading to cancer relapse.
Based on the molecular mechanisms of BCG action on the host immune system, we can suggest the following possible research strategies for the development of effective drugs:
- 1)
-
Driven Improvement (Specific activation of different signaling pathways)rBCG improved with various cytokines may be used to improve immune activation. A particular case of such amplification could be the use of additional effectors in cells with the NOD2 pathway turned off to compensate for it, or similar solutions. However, it is possible that the use of rBCG alone may be insufficient - this conclusion is based on the observation that BCG/rBCG has been shown to be more effective in studies using it in combination with other compounds. However, as a base, such formulations may be more effective than native BCG.
- 2)
-
Adjuvant Empowerment (Unspecific amplification of TRIM)Use of adjuvants to increase the magnitude of non-specific immune response and more stable formation of TRIM (including in a larger number of cells initially). The most straightforward option is the use of alum, which has been successfully used not only in subunit vaccines but also in combination with an inactivated pathogen (see 2.6.1). With particular emphasis on cancer vaccines, a logical evolution of this approach is to replace alum with metallic nanoparticles that can themselves specifically activate TRIM [247,248] and/or have more focused immunomodulatory properties [192].
- 3)
-
Combination:The combined use of rBGC and adjuvants or nanoparticles with specific properties could make a more significant contribution with relatively simple technological solutions. Thus, depending on the development of technologies, in the easiest way adjuvants can be used not only for adsorption of BCG molecules, as in the studies of chapter 2.6.1, but also for covalent binding with biotechnologically synthesized proteins - immune activators, as in the works of the Proff. Wittrup’s group [249,250]. An optional solution could be the use of constitutively modified rBGC, which are limited by a low level of expression of the introduced genes [251], with adjuvants or nanoparticles to enhance the effect.
6. Conclusion
Thus, having considered the various pathways activated by BCG that lead to an immune response, we can assume that in vaccinology, studying the detailed mechanisms of action of different vaccine components can enhance understanding of how to efficiently assemble a pan-pandemic vaccine and then subsequently develop a specific vaccine. In cancer research, the situation is more complicated - an increase in data often leads to new contradictions and doubts. As an example of the ambiguity of mechanisms and possible actions, the example of rapamycin / mTORC1 / autophagy / TRIM or the dual role of some cytokines in cancer shows that the abundance of contradictory details can lead to a possible dead end. After all Nixon’s ‘war on cancer’ is still not won [252,253]. Perhaps the clue to the solution comes from observations of equilibrium chemical reactions - the equilibrium can be shifted by a catalyst, and under certain conditions the reaction can be made unidirectional. Thus, when acting on a tumor that contains many fluctuating factors such as “double-edged sword,” “dual role,” “controversial,” etc., it is necessary to provide a catalytic impulse to shift the equilibrium in a particular direction. Potentially, with the right choice of components, BCG can play an important role here, due to its features and the possibility of modification.
Author Contributions
Conceptualization, VVP; software, VVP and LGK; validation, MVZ and LGK; formal analysis, VVP; investigation, VVP; resources, IVA; data curation, IVA; writing—original draft preparation, VVP; writing—review and editing, VVP, LGK, MVZ, IVA; visualization, LGK; supervision, VVP; project administration, VVP; funding acquisition, IVA. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Russian Science Foundation (project No. 22-14-00308), “https://rscf.ru/en/project/22-14-00308/ (accessed on 27 October 2024)”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- WHO. Global tuberculosis report 2023. 2023.
- World Health, O. BCG vaccine: WHO position paper, February 2018 - Recommendations. Vaccine 2018, 36, 3408–3410. [Google Scholar] [CrossRef] [PubMed]
- Lyon, S.M.; Rossman, M.D. Pulmonary Tuberculosis. Microbiology spectrum 2017, 5. [Google Scholar] [CrossRef]
- Trauer, J.M.; Moyo, N.; Tay, E.L.; Dale, K.; Ragonnet, R.; McBryde, E.S.; Denholm, J.T. Risk of Active Tuberculosis in the Five Years Following Infection... 15%? Chest 2016, 149, 516–525. [Google Scholar] [CrossRef]
- Bouzeyen, R.; Javid, B. Therapeutic Vaccines for Tuberculosis: An Overview. Frontiers in immunology 2022, 13, 878471. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki-Nakashimada, M.A.; Unzueta, A.; Berenise Gamez-Gonzalez, L.; Gonzalez-Saldana, N.; Sorensen, R.U. BCG: a vaccine with multiple faces. Human vaccines & immunotherapeutics 2020, 16, 1841–1850. [Google Scholar] [CrossRef]
- Ozeki, Y.; Yokoyama, A.; Nishiyama, A.; Yoshida, Y.; Ohara, Y.; Mashima, T.; Tomiyama, C.; Shaban, A.K.; Takeishi, A.; Osada-Oka, M.; et al. Recombinant mycobacterial DNA-binding protein 1 with post-translational modifications boosts IFN-gamma production from BCG-vaccinated individuals’ blood cells in combination with CpG-DNA. Scientific reports 2024, 14, 9141. [Google Scholar] [CrossRef]
- Lutwama, F.; Kagina, B.M.; Wajja, A.; Waiswa, F.; Mansoor, N.; Kirimunda, S.; Hughes, E.J.; Kiwanuka, N.; Joloba, M.L.; Musoke, P.; et al. Distinct T-cell responses when BCG vaccination is delayed from birth to 6 weeks of age in Ugandan infants. The Journal of infectious diseases 2014, 209, 887–897. [Google Scholar] [CrossRef]
- Martinez, L.; Cords, O.; Liu, Q.; Acuna-Villaorduna, C.; Bonnet, M.; Fox, G.J.; Carvalho, A.C.C.; Chan, P.C.; Croda, J.; Hill, P.C.; et al. Infant BCG vaccination and risk of pulmonary and extrapulmonary tuberculosis throughout the life course: a systematic review and individual participant data meta-analysis. The Lancet. Global health 2022, 10, e1307–e1316. [Google Scholar] [CrossRef]
- Roy, A.; Eisenhut, M.; Harris, R.J.; Rodrigues, L.C.; Sridhar, S.; Habermann, S.; Snell, L.; Mangtani, P.; Adetifa, I.; Lalvani, A.; et al. Effect of BCG vaccination against Mycobacterium tuberculosis infection in children: systematic review and meta-analysis. Bmj 2014, 349, g4643. [Google Scholar] [CrossRef]
- Villanueva, P.; Crawford, N.W.; Garcia Croda, M.; Collopy, S.; Araujo Jardim, B.; de Almeida Pinto Jardim, T.; Marshall, H.; Prat-Aymerich, C.; Sawka, A.; Sharma, K.; et al. Safety of BCG vaccination and revaccination in healthcare workers. Human vaccines & immunotherapeutics 2023, 19, 2239088. [Google Scholar] [CrossRef]
- Foster, M.; Hill, P.C.; Setiabudiawan, T.P.; Koeken, V.; Alisjahbana, B.; van Crevel, R. BCG-induced protection against Mycobacterium tuberculosis infection: Evidence, mechanisms, and implications for next-generation vaccines. Immunological reviews 2021, 301, 122–144. [Google Scholar] [CrossRef]
- Cho, T.; Khatchadourian, C.; Nguyen, H.; Dara, Y.; Jung, S.; Venketaraman, V. A review of the BCG vaccine and other approaches toward tuberculosis eradication. Human vaccines & immunotherapeutics 2021, 17, 2454–2470. [Google Scholar] [CrossRef]
- Ferluga, J.; Yasmin, H.; Al-Ahdal, M.N.; Bhakta, S.; Kishore, U. Natural and trained innate immunity against Mycobacterium tuberculosis. Immunobiology 2020, 225, 151951. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Du, G.; Chen, X.; Sun, X. Advancedoral vaccine delivery strategies for improving the immunity. Advanced drug delivery reviews 2021, 177, 113928. [Google Scholar] [CrossRef]
- Luca, S.; Mihaescu, T. History of BCG Vaccine. Maedica 2013, 8, 53–58. [Google Scholar]
- Fatima, S.; Kumari, A.; Das, G.; Dwivedi, V.P. Tuberculosis vaccine: A journey from BCG to present. Life sciences 2020, 252, 117594. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.; Cords, O.; Horsburgh, C.R.; Andrews, J.R.; Pediatric, T.B.C.S.C. The risk of tuberculosis in children after close exposure: a systematic review and individual-participant meta-analysis. Lancet 2020, 395, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xiang, D.; Zhang, X.; Zhang, L.; Wang, S.; Jin, K.; You, L.; Huang, J. The mechanisms and cross-protection of trained innate immunity. Virology journal 2022, 19, 210. [Google Scholar] [CrossRef]
- Alekseenko, I.V.; Vasilov, R.G.; Kondratyeva, L.G.; Kostrov, S.V.; Chernov, I.P.; Sverdlov, E.D. The cellular and epigenetic aspects of trained immunity and prospects for creation of universal vaccines on the eve of more frequent pandemics. Russ J Genet 2023, 59, 851–868. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Nante, E.; Jensen, I.P.; Kofoed, P.E.; Poulsen, A.; Jensen, H.; Newport, M.; Marchant, A.; Aaby, P. Acute lower respiratory tract infections and respiratory syncytial virus in infants in Guinea-Bissau: a beneficial effect of BCG vaccination for girls community based case-control study. Vaccine 2005, 23, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Wardhana; Datau, E. A.; Sultana, A.; Mandang, V.V.; Jim, E. The efficacy of Bacillus Calmette-Guerin vaccinations for the prevention of acute upper respiratory tract infection in the elderly. Acta medica Indonesiana 2011, 43, 185–190. [Google Scholar]
- Kang, A.; Ye, G.; Singh, R.; Afkhami, S.; Bavananthasivam, J.; Luo, X.; Vaseghi-Shanjani, M.; Aleithan, F.; Zganiacz, A.; Jeyanathan, M.; et al. Subcutaneous BCG vaccination protects against streptococcal pneumonia via regulating innate immune responses in the lung. EMBO molecular medicine 2023, 15, e17084. [Google Scholar] [CrossRef]
- Moorlag, S.J.C.F.M.; Arts, R.J.W.; van Crevel, R.; Netea, M.G. Non-specific effects of BCG vaccine on viral infections. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases 2019, 25, 1473–1478. [Google Scholar] [CrossRef]
- Podder, I.; Bhattacharya, S.; Mishra, V.; Sarkar, T.K.; Chandra, S.; Sil, A.; Pal, S.; Kumar, D.; Saha, A.; Shome, K.; et al. Immunotherapy in viral warts with intradermal Bacillus Calmette-Guerin vaccine versus intradermal tuberculin purified protein derivative: A double-blind, randomized controlled trial comparing effectiveness and safety in a tertiary care center in Eastern India. Indian journal of dermatology, venereology and leprology 2017, 83, 411. [Google Scholar] [CrossRef] [PubMed]
- Al-Yassen, A.Q.; Al-Maliki, S.K.; Al-Asadi, J.N. The Bacillus Calmette-Guerin (BCG) Vaccine: Is it a better choice for the treatment of viral warts? Sultan Qaboos University medical journal 2020, 20, e330–e336. [Google Scholar] [CrossRef]
- Walk, J.; de Bree, L.C.J.; Graumans, W.; Stoter, R.; van Gemert, G.J.; van de Vegte-Bolmer, M.; Teelen, K.; Hermsen, C.C.; Arts, R.J.W.; Behet, M.C.; et al. Outcomes of controlled human malaria infection after BCG vaccination. Nature communications 2019, 10, 874. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Moorlag, S.; Novakovic, B.; Li, Y.; Wang, S.Y.; Oosting, M.; Kumar, V.; Xavier, R.J.; Wijmenga, C.; Joosten, L.A.B.; et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell host & microbe 2018, 23, 89–100 e105. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: can it offer protection against COVID-19? Nature reviews. Immunology 2020, 20, 335–337. [Google Scholar] [CrossRef]
- Escobar, L.E.; Molina-Cruz, A.; Barillas-Mury, C. BCG vaccine protection from severe coronavirus disease 2019 (COVID-19). Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 17720–17726. [Google Scholar] [CrossRef]
- Moorlag, S.J.C.F.M.; van Deuren, R.C.; van Werkhoven, C.H.; Jaeger, M.; Debisarun, P.; Taks, E.; Mourits, V.P.; Koeken, V.; de Bree, L.C.J.; Ten Doesschate, T.; et al. Safety and COVID-19 Symptoms in Individuals Recently Vaccinated with BCG: a Retrospective Cohort Study. Cell reports. Medicine 2020, 1, 100073. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.K.; Yu, Q.; Salvador, C.E.; Melani, I.; Kitayama, S. Mandated Bacillus Calmette-Guerin (BCG) vaccination predicts flattened curves for the spread of COVID-19. Science advances 2020, 6, eabc1463. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.N.; Ebinger, J.E.; Wu, M.; Sun, N.; Braun, J.; Sobhani, K.; Van Eyk, J.E.; Cheng, S.; Arditi, M. BCG vaccination history associates with decreased SARS-CoV-2 seroprevalence across a diverse cohort of health care workers. The Journal of clinical investigation 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Aaby, P.; Benn, C.S.; Flanagan, K.L.; Klein, S.L.; Kollmann, T.R.; Lynn, D.J.; Shann, F. The non-specific and sex-differential effects of vaccines. Nature reviews. Immunology 2020, 20, 464–470. [Google Scholar] [CrossRef]
- Wen, J.; Liu, Q.; Tang, D.; He, J.Q. Efficacy of BCG Vaccination against COVID-19: Systematic Review and Meta-Analysis of Randomized Controlled Trials. Journal of clinical medicine 2023, 12. [Google Scholar] [CrossRef]
- Upton, C.M.; van Wijk, R.C.; Mockeliunas, L.; Simonsson, U.S.H.; McHarry, K.; van den Hoogen, G.; Muller, C.; von Delft, A.; van der Westhuizen, H.M.; van Crevel, R.; et al. Safety and efficacy of BCG re-vaccination in relation to COVID-19 morbidity in healthcare workers: A double-blind, randomised, controlled, phase 3 trial. EClinicalMedicine 2022, 48, 101414. [Google Scholar] [CrossRef]
- Claus, J.; Ten Doesschate, T.; Gumbs, C.; van Werkhoven, C.H.; van der Vaart, T.W.; Janssen, A.B.; Smits, G.; van Binnendijk, R.; van der Klis, F.; van Baarle, D.; et al. BCG Vaccination of Health Care Workers Does Not Reduce SARS-CoV-2 Infections nor Infection Severity or Duration: a Randomized Placebo-Controlled Trial. mBio 2023, 14, e0035623. [Google Scholar] [CrossRef]
- Gong, W.; Du, J. Excluding Participants With Mycobacteria Infections From Clinical Trials: A Critical Consideration in Evaluating the Efficacy of BCG Against COVID-19. Journal of Korean medical science 2023, 38, e343. [Google Scholar] [CrossRef]
- Portilho, A.I.; De Gaspari, E. Trained-immunity and cross-reactivity for protection: insights from the coronavirus disease 2019 and monkeypox emergencies for vaccine development. Explor Immunol 2023, 3, 276–285. [Google Scholar] [CrossRef]
- Tsilika, M.; Taks, E.; Dolianitis, K.; Kotsaki, A.; Leventogiannis, K.; Damoulari, C.; Kostoula, M.; Paneta, M.; Adamis, G.; Papanikolaou, I.; et al. ACTIVATE-2: A Double-Blind Randomized Trial of BCG Vaccination Against COVID-19 in Individuals at Risk. Frontiers in immunology 2022, 13, 873067. [Google Scholar] [CrossRef]
- Kuhtreiber, W.M.; Hostetter, E.R.; Wolfe, G.E.; Vaishnaw, M.S.; Goldstein, R.; Bulczynski, E.R.; Hullavarad, N.S.; Braley, J.E.; Zheng, H.; Faustman, D.L. Late in the US pandemic, multi-dose BCG vaccines protect against COVID-19 and infectious diseases. iScience 2024, 27, 109881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ru, H.W.; Chen, F.Z.; Jin, C.Y.; Sun, R.F.; Fan, X.Y.; Guo, M.; Mai, J.T.; Xu, W.X.; Lin, Q.X.; et al. Variable Virulence and Efficacy of BCG Vaccine Strains in Mice and Correlation With Genome Polymorphisms. Molecular therapy : the journal of the American Society of Gene Therapy 2016, 24, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Kilic, G.; Debisarun, P.A.; Alaswad, A.; Baltissen, M.P.; Lamers, L.A.; de Bree, L.C.J.; Benn, C.S.; Aaby, P.; Dijkstra, H.; Lemmers, H.; et al. Seasonal variation in BCG-induced trained immunity. Vaccine 2024, 42, 126109. [Google Scholar] [CrossRef] [PubMed]
- Amirlak, L.; Haddad, R.; Hardy, J.D.; Khaled, N.S.; Chung, M.H.; Amirlak, B. Effectiveness of booster BCG vaccination in preventing Covid-19 infection. Human vaccines & immunotherapeutics 2021, 17, 3913–3915. [Google Scholar] [CrossRef]
- Vuscan, P.; Kischkel, B.; Joosten, L.A.B.; Netea, M.G. Trained immunity: General and emerging concepts. Immunological reviews 2024, 323, 164–185. [Google Scholar] [CrossRef]
- Rothfuchs, A.G.; Bafica, A.; Feng, C.G.; Egen, J.G.; Williams, D.L.; Brown, G.D.; Sher, A. Dectin-1 interaction with Mycobacterium tuberculosis leads to enhanced IL-12p40 production by splenic dendritic cells. Journal of immunology 2007, 179, 3463–3471. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; van Loenhout, J.; de Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proceedings of the National Academy of Sciences of the United States of America 2012, 109, 17537–17542. [Google Scholar] [CrossRef]
- Bisiaux, A.; Boussier, J.; Duffy, D.; Quintana-Murci, L.; Fontes, M.; Albert, M.L.; Milieu Interieur, C. Deconvolution of the Response to Bacillus Calmette-Guerin Reveals NF-kappaB-Induced Cytokines As Autocrine Mediators of Innate Immunity. Frontiers in immunology 2017, 8, 796. [Google Scholar] [CrossRef]
- Baydemir, I.; Dulfer, E.A.; Netea, M.G.; Dominguez-Andres, J. Trained immunity-inducing vaccines: Harnessing innate memory for vaccine design and delivery. Clinical immunology 2024, 261, 109930. [Google Scholar] [CrossRef]
- Ochando, J.; Mulder, W.J.M.; Madsen, J.C.; Netea, M.G.; Duivenvoorden, R. Trained immunity - basic concepts and contributions to immunopathology. Nature reviews. Nephrology 2023, 19, 23–37. [Google Scholar] [CrossRef]
- Al Nabhani, Z.; Dietrich, G.; Hugot, J.P.; Barreau, F. Nod2: The intestinal gate keeper. PLoS pathogens 2017, 13, e1006177. [Google Scholar] [CrossRef] [PubMed]
- Cicchese, J.M.; Evans, S.; Hult, C.; Joslyn, L.R.; Wessler, T.; Millar, J.A.; Marino, S.; Cilfone, N.A.; Mattila, J.T.; Linderman, J.J.; et al. Dynamic balance of pro- and anti-inflammatory signals controls disease and limits pathology. Immunological reviews 2018, 285, 147–167. [Google Scholar] [CrossRef] [PubMed]
- Moorlag, S.J.C.F.M.; Folkman, L.; Ter Horst, R.; Krausgruber, T.; Barreca, D.; Schuster, L.C.; Fife, V.; Matzaraki, V.; Li, W.; Reichl, S.; et al. Multi-omics analysis of innate and adaptive responses to BCG vaccination reveals epigenetic cell states that predict trained immunity. Immunity 2024, 57, 171–187 e114. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhong, Z.; Karin, M. NF-kappaB: A Double-Edged Sword Controlling Inflammation. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of Inflammation: What Controls Its Onset? Frontiers in immunology 2016, 7, 160. [Google Scholar] [CrossRef]
- Lotfi, N.; Zhang, G.X.; Esmaeil, N.; Rostami, A. Evaluation of the effect of GM-CSF blocking on the phenotype and function of human monocytes. Scientific reports 2020, 10, 1567. [Google Scholar] [CrossRef]
- Bender, A.T.; Ostenson, C.L.; Giordano, D.; Beavo, J.A. Differentiation of human monocytes in vitro with granulocyte-macrophage colony-stimulating factor and macrophage colony-stimulating factor produces distinct changes in cGMP phosphodiesterase expression. Cellular signalling 2004, 16, 365–374. [Google Scholar] [CrossRef]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. Journal of leukocyte biology 2016, 100, 481–489. [Google Scholar] [CrossRef]
- Alekseenko, I.; Kuzmich, A.; Kondratyeva, L.; Kondratieva, S.; Pleshkan, V.; Sverdlov, E. Step-by-Step Immune Activation for Suicide Gene Therapy Reinforcement. International journal of molecular sciences 2021, 22. [Google Scholar] [CrossRef]
- Kumar, A.; Taghi Khani, A.; Sanchez Ortiz, A.; Swaminathan, S. GM-CSF: A Double-Edged Sword in Cancer Immunotherapy. Frontiers in immunology 2022, 13, 901277. [Google Scholar] [CrossRef]
- Waller, E.K. The role of sargramostim (rhGM-CSF) as immunotherapy. The oncologist 2007, 12 Suppl 2, 22–26. [Google Scholar] [CrossRef]
- Kim, I.K.; Koh, C.H.; Jeon, I.; Shin, K.S.; Kang, T.S.; Bae, E.A.; Seo, H.; Ko, H.J.; Kim, B.S.; Chung, Y.; et al. GM-CSF Promotes Antitumor Immunity by Inducing Th9 Cell Responses. Cancer immunology research 2019, 7, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, M.M.; Abuharfeil, N.M.; Darmani, H. The role of IL-1beta during human immunodeficiency virus type 1 infection. Reviews in medical virology 2023, 33, e2400. [Google Scholar] [CrossRef] [PubMed]
- Broderick, L.; Hoffman, H.M. IL-1 and autoinflammatory disease: biology, pathogenesis and therapeutic targeting. Nature reviews. Rheumatology 2022, 18, 448–463. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) pathway. Science signaling 2010, 3, cm1. [Google Scholar] [CrossRef]
- Bent, R.; Moll, L.; Grabbe, S.; Bros, M. Interleukin-1 Beta-A Friend or Foe in Malignancies? International journal of molecular sciences 2018, 19. [Google Scholar] [CrossRef]
- Aarreberg, L.D.; Esser-Nobis, K.; Driscoll, C.; Shuvarikov, A.; Roby, J.A.; Gale, M., Jr. Interleukin-1beta Induces mtDNA Release to Activate Innate Immune Signaling via cGAS-STING. Molecular cell 2019, 74, 801–815 e806. [Google Scholar] [CrossRef]
- Mantovani, A. Cancer: an infernal triangle. Nature 2007, 448, 547–548. [Google Scholar] [CrossRef]
- Rebe, C.; Ghiringhelli, F. Interleukin-1beta and Cancer. Cancers 2020, 12. [Google Scholar] [CrossRef]
- Malkova, A.M.; Gubal, A.R.; Petrova, A.L.; Voronov, E.; Apte, R.N.; Semenov, K.N.; Sharoyko, V.V. Pathogenetic role and clinical significance of interleukin-1beta in cancer. Immunology 2023, 168, 203–216. [Google Scholar] [CrossRef]
- Zhou, J.; Ottewell, P.D. The role of IL-1B in breast cancer bone metastasis. Journal of bone oncology 2024, 46, 100608. [Google Scholar] [CrossRef] [PubMed]
- Kaplanov, I.; Carmi, Y.; Kornetsky, R.; Shemesh, A.; Shurin, G.V.; Shurin, M.R.; Dinarello, C.A.; Voronov, E.; Apte, R.N. Blocking IL-1beta reverses the immunosuppression in mouse breast cancer and synergizes with anti-PD-1 for tumor abrogation. Proceedings of the National Academy of Sciences of the United States of America 2019, 116, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nature reviews. Immunology 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Bonjardim, C.A. Interferons (IFNs) are key cytokines in both innate and adaptive antiviral immune responses--and viruses counteract IFN action. Microbes and infection 2005, 7, 569–578. [Google Scholar] [CrossRef]
- Mertowska, P.; Smolak, K.; Mertowski, S.; Grywalska, E. Immunomodulatory Role of Interferons in Viral and Bacterial Infections. International journal of molecular sciences 2023, 24. [Google Scholar] [CrossRef]
- Zannikou, M.; Fish, E.N.; Platanias, L.C. Signaling by Type I Interferons in Immune Cells: Disease Consequences. Cancers 2024, 16. [Google Scholar] [CrossRef]
- Goldszmid, R.S.; Caspar, P.; Rivollier, A.; White, S.; Dzutsev, A.; Hieny, S.; Kelsall, B.; Trinchieri, G.; Sher, A. NK cell-derived interferon-gamma orchestrates cellular dynamics and the differentiation of monocytes into dendritic cells at the site of infection. Immunity 2012, 36, 1047–1059. [Google Scholar] [CrossRef]
- Tough, D.F. Modulation of T-cell function by type I interferon. Immunology and cell biology 2012, 90, 492–497. [Google Scholar] [CrossRef]
- Huber, J.P.; Farrar, J.D. Regulation of effector and memory T-cell functions by type I interferon. Immunology 2011, 132, 466–474. [Google Scholar] [CrossRef]
- Kuka, M.; De Giovanni, M.; Iannacone, M. The role of type I interferons in CD4(+) T cell differentiation. Immunology letters 2019, 215, 19–23. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, Y.; Wang, Y.; Zhang, M.; Sun, W.; Dai, T.; Wang, A.; Wu, X.; Zhang, S.; Wang, S.; et al. A Dual Role of Type I Interferons in Antitumor Immunity. Advanced biosystems 2020, 4, e1900237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, S.; Zhu, Y.; Zhang, M.; Zhao, Y.; Yan, Z.; Wang, Q.; Li, X. Double-edged effects of interferons on the regulation of cancer-immunity cycle. Oncoimmunology 2021, 10, 1929005. [Google Scholar] [CrossRef] [PubMed]
- Abdolvahab, M.H.; Darvishi, B.; Zarei, M.; Majidzadeh, A.K.; Farahmand, L. Interferons: role in cancer therapy. Immunotherapy 2020, 12, 833–855. [Google Scholar] [CrossRef] [PubMed]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-gamma in tumor progression and regression: a review. Biomarker research 2020, 8, 49. [Google Scholar] [CrossRef]
- Song, M.; Ping, Y.; Zhang, K.; Yang, L.; Li, F.; Zhang, C.; Cheng, S.; Yue, D.; Maimela, N.R.; Qu, J.; et al. Low-Dose IFNgamma Induces Tumor Cell Stemness in Tumor Microenvironment of Non-Small Cell Lung Cancer. Cancer research 2019, 79, 3737–3748. [Google Scholar] [CrossRef]
- Beziaud, L.; Young, C.M.; Alonso, A.M.; Norkin, M.; Minafra, A.R.; Huelsken, J. IFNgamma-induced stem-like state of cancer cells as a driver of metastatic progression following immunotherapy. Cell stem cell 2023, 30, 818–831 e816. [Google Scholar] [CrossRef] [PubMed]
- Lo, U.G.; Pong, R.C.; Yang, D.; Gandee, L.; Hernandez, E.; Dang, A.; Lin, C.J.; Santoyo, J.; Ma, S.; Sonavane, R.; et al. IFNgamma-Induced IFIT5 Promotes Epithelial-to-Mesenchymal Transition in Prostate Cancer via miRNA Processing. Cancer research 2019, 79, 1098–1112. [Google Scholar] [CrossRef]
- Schalper, K.A.; Carvajal-Hausdorf, D.; McLaughlin, J.; Altan, M.; Velcheti, V.; Gaule, P.; Sanmamed, M.F.; Chen, L.; Herbst, R.S.; Rimm, D.L. Differential Expression and Significance of PD-L1, IDO-1, and B7-H4 in Human Lung Cancer. Clinical cancer research : an official journal of the American Association for Cancer Research 2017, 23, 370–378. [Google Scholar] [CrossRef]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The Role of Tumor Necrosis Factor Alpha (TNF-alpha) in Autoimmune Disease and Current TNF-alpha Inhibitors in Therapeutics. International journal of molecular sciences 2021, 22. [Google Scholar] [CrossRef]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: a road to inflammation. Nature reviews. Immunology 2023, 23, 289–303. [Google Scholar] [CrossRef]
- Laha, D.; Grant, R.; Mishra, P.; Nilubol, N. The Role of Tumor Necrosis Factor in Manipulating the Immunological Response of Tumor Microenvironment. Frontiers in immunology 2021, 12, 656908. [Google Scholar] [CrossRef] [PubMed]
- van Deventer, S.J. Transmembrane TNF-alpha, induction of apoptosis, and the efficacy of TNF-targeting therapies in Crohn’s disease. Gastroenterology 2001, 121, 1242–1246. [Google Scholar] [CrossRef] [PubMed]
- Faraj, S.S.; Jalal, P.J. IL1beta, IL-6, and TNF-alpha cytokines cooperate to modulate a complicated medical condition among COVID-19 patients: case-control study. Annals of medicine and surgery 2023, 85, 2291–2297. [Google Scholar] [CrossRef]
- Ham, B.; Fernandez, M.C.; D’Costa, Z.; Brodt, P. The diverse roles of the TNF axis in cancer progression and metastasis. Trends in cancer research 2016, 11, 1–27. [Google Scholar]
- Alim, L.F.; Keane, C.; Souza-Fonseca-Guimaraes, F. Molecular mechanisms of tumour necrosis factor signalling via TNF receptor 1 and TNF receptor 2 in the tumour microenvironment. Current opinion in immunology 2024, 86, 102409. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, B.P. TNF-alpha/NF-kappaB/Snail pathway in cancer cell migration and invasion. British journal of cancer 2010, 102, 639–644. [Google Scholar] [CrossRef]
- Tang, D.; Tao, D.; Fang, Y.; Deng, C.; Xu, Q.; Zhou, J. TNF-Alpha Promotes Invasion and Metastasis via NF-Kappa B Pathway in Oral Squamous Cell Carcinoma. Medical science monitor basic research 2017, 23, 141–149. [Google Scholar] [CrossRef]
- Shin, M.R.; Kang, S.K.; Kim, Y.S.; Lee, S.Y.; Hong, S.C.; Kim, E.C. TNF-alpha and LPS activate angiogenesis via VEGF and SIRT1 signalling in human dental pulp cells. International endodontic journal 2015, 48, 705–716. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harbor perspectives in biology 2014, 6, a016295. [Google Scholar] [CrossRef]
- Dienz, O.; Eaton, S.M.; Bond, J.P.; Neveu, W.; Moquin, D.; Noubade, R.; Briso, E.M.; Charland, C.; Leonard, W.J.; Ciliberto, G.; et al. The induction of antibody production by IL-6 is indirectly mediated by IL-21 produced by CD4+ T cells. The Journal of experimental medicine 2009, 206, 69–78. [Google Scholar] [CrossRef]
- Li, B.; Jones, L.L.; Geiger, T.L. IL-6 Promotes T Cell Proliferation and Expansion under Inflammatory Conditions in Association with Low-Level RORgammat Expression. Journal of immunology 2018, 201, 2934–2946. [Google Scholar] [CrossRef]
- Ngwa, D.N.; Pathak, A.; Agrawal, A. IL-6 regulates induction of C-reactive protein gene expression by activating STAT3 isoforms. Molecular immunology 2022, 146, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Tie, R.; Li, H.; Cai, S.; Liang, Z.; Shan, W.; Wang, B.; Tan, Y.; Zheng, W.; Huang, H. Interleukin-6 signaling regulates hematopoietic stem cell emergence. Experimental & molecular medicine 2019, 51, 1–12. [Google Scholar] [CrossRef]
- Li, Y.S.; Ren, H.C.; Cao, J.H. Roles of Interleukin-6-mediated immunometabolic reprogramming in COVID-19 and other viral infection-associated diseases. International immunopharmacology 2022, 110, 109005. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.T.; Appenheimer, M.M.; Evans, S.S. The two faces of IL-6 in the tumor microenvironment. Seminars in immunology 2014, 26, 38–47. [Google Scholar] [CrossRef]
- Raskova, M.; Lacina, L.; Kejik, Z.; Venhauerova, A.; Skalickova, M.; Kolar, M.; Jakubek, M.; Rosel, D.; Smetana, K., Jr.; Brabek, J. The Role of IL-6 in Cancer Cell Invasiveness and Metastasis-Overview and Therapeutic Opportunities. Cells 2022, 11. [Google Scholar] [CrossRef]
- Zhao, G.; Zhu, G.; Huang, Y.; Zheng, W.; Hua, J.; Yang, S.; Zhuang, J.; Ye, J. IL-6 mediates the signal pathway of JAK-STAT3-VEGF-C promoting growth, invasion and lymphangiogenesis in gastric cancer. Oncology reports 2016, 35, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Bent, E.H.; Millan-Barea, L.R.; Zhuang, I.; Goulet, D.R.; Frose, J.; Hemann, M.T. Microenvironmental IL-6 inhibits anti-cancer immune responses generated by cytotoxic chemotherapy. Nature communications 2021, 12, 6218. [Google Scholar] [CrossRef]
- Zhang, K.W.; Wang, D.; Cai, H.; Cao, M.Q.; Zhang, Y.Y.; Zhuang, P.Y.; Shen, J. IL-6 plays a crucial role in epithelial-mesenchymal transition and pro-metastasis induced by sorafenib in liver cancer. Oncology reports 2021, 45, 1105–1117. [Google Scholar] [CrossRef]
- Ekkens, M.J.; Shedlock, D.J.; Jung, E.; Troy, A.; Pearce, E.L.; Shen, H.; Pearce, E.J. Th1 and Th2 cells help CD8 T-cell responses. Infection and immunity 2007, 75, 2291–2296. [Google Scholar] [CrossRef]
- Muraille, E.; Leo, O.; Moser, M. TH1/TH2 paradigm extended: macrophage polarization as an unappreciated pathogen-driven escape mechanism? Frontiers in immunology 2014, 5, 603. [Google Scholar] [CrossRef] [PubMed]
- Howard, F.H.N.; Kwan, A.; Winder, N.; Mughal, A.; Collado-Rojas, C.; Muthana, M. Understanding Immune Responses to Viruses-Do Underlying Th1/Th2 Cell Biases Predict Outcome? Viruses 2022, 14. [Google Scholar] [CrossRef]
- Swanson, M.A.; Lee, W.T.; Sanders, V.M. IFN-gamma production by Th1 cells generated from naive CD4+ T cells exposed to norepinephrine. Journal of immunology 2001, 166, 232–240. [Google Scholar] [CrossRef]
- Nonaka, K.; Saio, M.; Umemura, N.; Kikuchi, A.; Takahashi, T.; Osada, S.; Yoshida, K. Th1 polarization in the tumor microenvironment upregulates the myeloid-derived suppressor-like function of macrophages. Cellular immunology 2021, 369, 104437. [Google Scholar] [CrossRef]
- Lee, J.; Lozano-Ruiz, B.; Yang, F.M.; Fan, D.D.; Shen, L.; Gonzalez-Navajas, J.M. The Multifaceted Role of Th1, Th9, and Th17 Cells in Immune Checkpoint Inhibition Therapy. Frontiers in immunology 2021, 12, 625667. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Jiang, L.; Zhang, F.; Li, M.; Yang, B.; Zhang, F.; Guo, X. Acetate promotes SNAI1 expression by ACSS2-mediated histone acetylation under glucose limitation in renal cell carcinoma cell. Bioscience reports 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Villa, M.; Sanin, D.E.; Buck, M.D.; O’Sullivan, D.; Ching, R.; Matsushita, M.; Grzes, K.M.; Winkler, F.; Chang, C.H.; et al. Acetate Promotes T Cell Effector Function during Glucose Restriction. Cell reports 2019, 27, 2063–2074 e2065. [Google Scholar] [CrossRef]
- Zhang, S.; Lv, K.; Liu, Z.; Zhao, R.; Li, F. Fatty acid metabolism of immune cells: a new target of tumour immunotherapy. Cell death discovery 2024, 10, 39. [Google Scholar] [CrossRef]
- Xu, M.; Jiang, Z.; Wang, C.; Li, N.; Bo, L.; Zha, Y.; Bian, J.; Zhang, Y.; Deng, X. Acetate attenuates inflammasome activation through GPR43-mediated Ca(2+)-dependent NLRP3 ubiquitination. Experimental & molecular medicine 2019, 51, 1–13. [Google Scholar] [CrossRef]
- Miller, K.D.; O’Connor, S.; Pniewski, K.A.; Kannan, T.; Acosta, R.; Mirji, G.; Papp, S.; Hulse, M.; Mukha, D.; Hlavaty, S.I.; et al. Acetate acts as a metabolic immunomodulator by bolstering T-cell effector function and potentiating antitumor immunity in breast cancer. Nature cancer 2023, 4, 1491–1507. [Google Scholar] [CrossRef]
- Guertin, D.A.; Wellen, K.E. Acetyl-CoA metabolism in cancer. Nature reviews. Cancer 2023, 23, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Godkowicz, M.; Druszczynska, M. NOD1, NOD2, and NLRC5 Receptors in Antiviral and Antimycobacterial Immunity. Vaccines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Miao, Z.; Qin, X.; Li, B.; Han, Y. NOD2 deficiency confers a pro-tumorigenic macrophage phenotype to promote lung adenocarcinoma progression. Journal of cellular and molecular medicine 2021, 25, 7545–7558. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Advances in nutrition 2018, 9, 21–29. [Google Scholar] [CrossRef]
- Sun, J.; Chen, S.; Zang, D.; Sun, H.; Sun, Y.; Chen, J. Butyrate as a promising therapeutic target in cancer: From pathogenesis to clinic (Review). International journal of oncology 2024, 64. [Google Scholar] [CrossRef] [PubMed]
- Valcarcel-Jimenez, L.; Frezza, C. Fumarate hydratase (FH) and cancer: a paradigm of oncometabolism. British journal of cancer 2023, 129, 1546–1557. [Google Scholar] [CrossRef]
- Fitzsimmons, C.M.; Mandler, M.D.; Lunger, J.C.; Chan, D.; Maligireddy, S.S.; Schmiechen, A.C.; Gamage, S.T.; Link, C.; Jenkins, L.M.; Chan, K.; et al. Rewiring of RNA methylation by the oncometabolite fumarate in renal cell carcinoma. NAR cancer 2024, 6, zcae004. [Google Scholar] [CrossRef]
- Yang, M.; Soga, T.; Pollard, P.J.; Adam, J. The emerging role of fumarate as an oncometabolite. Frontiers in oncology 2012, 2, 85. [Google Scholar] [CrossRef]
- Gegotek, A.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid. Antioxidants 2022, 11. [Google Scholar] [CrossRef]
- Cheng, S.C.; Quintin, J.; Cramer, R.A.; Shepardson, K.M.; Saeed, S.; Kumar, V.; Giamarellos-Bourboulis, E.J.; Martens, J.H.; Rao, N.A.; Aghajanirefah, A.; et al. mTOR- and HIF-1alpha-mediated aerobic glycolysis as metabolic basis for trained immunity. Science 2014, 345, 1250684. [Google Scholar] [CrossRef]
- Intravenous Vitamin C (PDQ(R)): Health Professional Version. In PDQ Cancer Information Summaries; Bethesda (MD), 2002.
- Gonzalez-Montero, J.; Chichiarelli, S.; Eufemi, M.; Altieri, F.; Saso, L.; Rodrigo, R. Ascorbate as a Bioactive Compound in Cancer Therapy: The Old Classic Strikes Back. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Liao, Y.; Na, J.; Wu, L.; Yin, Y.; Mi, Z.; Fang, S.; Liu, X.; Huang, Y. The Involvement of Ascorbic Acid in Cancer Treatment. Molecules 2024, 29. [Google Scholar] [CrossRef]
- Najeeb, H.A.; Sanusi, T.; Saldanha, G.; Brown, K.; Cooke, M.S.; Jones, G.D. Redox modulation of oxidatively-induced DNA damage by ascorbate enhances both in vitro and ex-vivo DNA damage formation and cell death in melanoma cells. Free radical biology & medicine 2024, 213, 309–321. [Google Scholar] [CrossRef]
- Maekawa, T.; Miyake, T.; Tani, M.; Uemoto, S. Diverse antitumor effects of ascorbic acid on cancer cells and the tumor microenvironment. Frontiers in oncology 2022, 12, 981547. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Frias, M.A.; Chatterjee, A.; Yellen, P.; Foster, D.A. The Enigma of Rapamycin Dosage. Molecular cancer therapeutics 2016, 15, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Cancer prevention with rapamycin. Oncotarget 2023, 14, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Repas, J.; Zupin, M.; Vodlan, M.; Veranic, P.; Gole, B.; Potocnik, U.; Pavlin, M. Dual Effect of Combined Metformin and 2-Deoxy-D-Glucose Treatment on Mitochondrial Biogenesis and PD-L1 Expression in Triple-Negative Breast Cancer Cells. Cancers 2022, 14. [Google Scholar] [CrossRef]
- Mitroulis, I.; Ruppova, K.; Wang, B.; Chen, L.S.; Grzybek, M.; Grinenko, T.; Eugster, A.; Troullinaki, M.; Palladini, A.; Kourtzelis, I.; et al. Modulation of Myelopoiesis Progenitors Is an Integral Component of Trained Immunity. Cell 2018, 172, 147–161 e112. [Google Scholar] [CrossRef]
- Kaufmann, E.; Sanz, J.; Dunn, J.L.; Khan, N.; Mendonca, L.E.; Pacis, A.; Tzelepis, F.; Pernet, E.; Dumaine, A.; Grenier, J.C.; et al. BCG Educates Hematopoietic Stem Cells to Generate Protective Innate Immunity against Tuberculosis. Cell 2018, 172, 176–190 e119. [Google Scholar] [CrossRef]
- Cirovic, B.; de Bree, L.C.J.; Groh, L.; Blok, B.A.; Chan, J.; van der Velden, W.; Bremmers, M.E.J.; van Crevel, R.; Handler, K.; Picelli, S.; et al. BCG Vaccination in Humans Elicits Trained Immunity via the Hematopoietic Progenitor Compartment. Cell host & microbe 2020, 28, 322–334 e325. [Google Scholar] [CrossRef]
- Yao, Y.; Jeyanathan, M.; Haddadi, S.; Barra, N.G.; Vaseghi-Shanjani, M.; Damjanovic, D.; Lai, R.; Afkhami, S.; Chen, Y.; Dvorkin-Gheva, A.; et al. Induction of Autonomous Memory Alveolar Macrophages Requires T Cell Help and Is Critical to Trained Immunity. Cell 2018, 175, 1634–1650 e1617. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Jacobs, C.; Xavier, R.J.; van der Meer, J.W.; van Crevel, R.; Netea, M.G. BCG-induced trained immunity in NK cells: Role for non-specific protection to infection. Clinical immunology 2014, 155, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Quintin, J.; Saeed, S.; Martens, J.H.A.; Giamarellos-Bourboulis, E.J.; Ifrim, D.C.; Logie, C.; Jacobs, L.; Jansen, T.; Kullberg, B.J.; Wijmenga, C.; et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell host & microbe 2012, 12, 223–232. [Google Scholar] [CrossRef]
- Morita, C.T.; Mariuzza, R.A.; Brenner, M.B. Antigen recognition by human gamma delta T cells: pattern recognition by the adaptive immune system. Springer seminars in immunopathology 2000, 22, 191–217. [Google Scholar] [CrossRef]
- Xu, B.; Pizarro, J.C.; Holmes, M.A.; McBeth, C.; Groh, V.; Spies, T.; Strong, R.K. Crystal structure of a gammadelta T-cell receptor specific for the human MHC class I homolog MICA. Proceedings of the National Academy of Sciences of the United States of America 2011, 108, 2414–2419. [Google Scholar] [CrossRef]
- Holtmeier, W.; Kabelitz, D. gammadelta T cells link innate and adaptive immune responses. Chemical immunology and allergy 2005, 86, 151–183. [Google Scholar] [CrossRef]
- Suen, T.K.; Moorlag, S.; Li, W.; de Bree, L.C.J.; Koeken, V.; Mourits, V.P.; Dijkstra, H.; Lemmers, H.; Bhat, J.; Xu, C.J.; et al. BCG vaccination induces innate immune memory in gammadelta T cells in humans. Journal of leukocyte biology 2024, 115, 149–163. [Google Scholar] [CrossRef]
- Sun, L.; Wang, X.; Saredy, J.; Yuan, Z.; Yang, X.; Wang, H. Innate-adaptive immunity interplay and redox regulation in immune response. Redox biology 2020, 37, 101759. [Google Scholar] [CrossRef]
- Jain, A.; Pasare, C. Innate Control of Adaptive Immunity: Beyond the Three-Signal Paradigm. Journal of immunology 2017, 198, 3791–3800. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nature immunology 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yang, X.; Yuan, Z.; Wang, H. Metabolic Reprogramming in Immune Response and Tissue Inflammation. Arteriosclerosis, thrombosis, and vascular biology 2020, 40, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.F.; Wang, H. Monocyte and macrophage differentiation: circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomarker research 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Li, X.; Dai, J.; Cole, L.; Camacho, J.A.; Zhang, Y.; Ji, Y.; Wang, J.; Yang, X.F.; Wang, H. Immune cell subset differentiation and tissue inflammation. Journal of hematology & oncology 2018, 11, 97. [Google Scholar] [CrossRef]
- Pulendran, B. Integrated organ immunity: a path to a universal vaccine. Nature reviews. Immunology 2024, 24, 81–82. [Google Scholar] [CrossRef]
- Lee, A.; Floyd, K.; Wu, S.; Fang, Z.; Tan, T.K.; Froggatt, H.M.; Powers, J.M.; Leist, S.R.; Gully, K.L.; Hubbard, M.L.; et al. BCG vaccination stimulates integrated organ immunity by feedback of the adaptive immune response to imprint prolonged innate antiviral resistance. Nature immunology 2024, 25, 41–53. [Google Scholar] [CrossRef]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Human vaccines & immunotherapeutics 2015, 11, 477–488. [Google Scholar] [CrossRef]
- Vermaelen, K. Vaccine Strategies to Improve Anti-cancer Cellular Immune Responses. Frontiers in immunology 2019, 10, 8. [Google Scholar] [CrossRef]
- Lee, A.; Scott, M.K.D.; Wimmers, F.; Arunachalam, P.S.; Luo, W.; Fox, C.B.; Tomai, M.; Khatri, P.; Pulendran, B. A molecular atlas of innate immunity to adjuvanted and live attenuated vaccines, in mice. Nature communications 2022, 13, 549. [Google Scholar] [CrossRef]
- Gallucci, S.; Matzinger, P. Danger signals: SOS to the immune system. Current opinion in immunology 2001, 13, 114–119. [Google Scholar] [CrossRef]
- Chaudhuri, N.; Sabroe, I. Basic science of the innate immune system and the lung. Paediatric respiratory reviews 2008, 9, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Matzinger, P. Tolerance, danger, and the extended family. Annual review of immunology 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Pulendran, B.; P, S.A.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nature reviews. Drug discovery 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Kamil, A.A.; Khalil, E.A.; Musa, A.M.; Modabber, F.; Mukhtar, M.M.; Ibrahim, M.E.; Zijlstra, E.E.; Sacks, D.; Smith, P.G.; Zicker, F.; et al. Alum-precipitated autoclaved Leishmania major plus bacille Calmette-Guerrin, a candidate vaccine for visceral leishmaniasis: safety, skin-delayed type hypersensitivity response and dose finding in healthy volunteers. Transactions of the Royal Society of Tropical Medicine and Hygiene 2003, 97, 365–368. [Google Scholar] [CrossRef]
- Khalil, E.A.; Musa, A.M.; Modabber, F.; El-Hassan, A.M. Safety and immunogenicity of a candidate vaccine for visceral leishmaniasis (Alum-precipitated autoclaved Leishmania major + BCG) in children: an extended phase II study. Annals of tropical paediatrics 2006, 26, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Mutiso, J.M.; Macharia, J.C.; Gicheru, M.M. Immunization with Leishmania vaccine-alum-BCG and montanide ISA 720 adjuvants induces low-grade type 2 cytokines and high levels of IgG2 subclass antibodies in the vervet monkey (Chlorocebus aethiops) model. Scandinavian journal of immunology 2012, 76, 471–477. [Google Scholar] [CrossRef]
- Selvapandiyan, A.; Puri, N.; Reyaz, E.; Beg, M.A.; Salotra, P.; Nakhasi, H.L.; Ganguly, N.K. Worldwide Efforts for the Prevention of Visceral Leishmaniasis Using Vaccinations. In Challenges and Solutions Against Visceral Leishmaniasis, Selvapandiyan, A., Singh, R., Puri, N., Ganguly, N.K., Ed.; Springer, Singapore: 2024.
- Ayala, A.; Llanes, A.; Lleonart, R.; Restrepo, C.M. Advances in Leishmania Vaccines: Current Development and Future Prospects. Pathogens 2024, 13. [Google Scholar] [CrossRef]
- Barati, M.; Mohebali, M.; Khamesipour, A.; Bahrami, F.; Darabi, H.; Khaze, V.; Riazi-Rad, F.; Habibi, G.; Ajdary, S.; Alimohammadian, M.H. Evaluation of Cellular Immune Responses in Dogs Immunized with Alum-Precipitated Autoclaved Leishmania major along with BCG and Imiquimod. Iranian journal of parasitology 2021, 16, 348–356. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Kaufmann, S.H.E. Next-Generation Vaccines Based on Bacille Calmette-Guerin. Frontiers in immunology 2018, 9, 121. [Google Scholar] [CrossRef]
- Ryan, A.A.; Spratt, J.M.; Britton, W.J.; Triccas, J.A. Secretion of functional monocyte chemotactic protein 3 by recombinant Mycobacterium bovis BCG attenuates vaccine virulence and maintains protective efficacy against M. tuberculosis infection. Infection and immunity 2007, 75, 523–526. [Google Scholar] [CrossRef]
- Peters, W.; Scott, H.M.; Chambers, H.F.; Flynn, J.L.; Charo, I.F.; Ernst, J.D. Chemokine receptor 2 serves an early and essential role in resistance to Mycobacterium tuberculosis. Proceedings of the National Academy of Sciences of the United States of America 2001, 98, 7958–7963. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.A.; Wozniak, T.M.; Shklovskaya, E.; O’Donnell, M.A.; Fazekas de St Groth, B.; Britton, W.J.; Triccas, J.A. Improved protection against disseminated tuberculosis by Mycobacterium bovis bacillus Calmette-Guerin secreting murine GM-CSF is associated with expansion and activation of APCs. Journal of immunology 2007, 179, 8418–8424. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.A.; Nambiar, J.K.; Wozniak, T.M.; Roediger, B.; Shklovskaya, E.; Britton, W.J.; Fazekas de St Groth, B.; Triccas, J.A. Antigen load governs the differential priming of CD8 T cells in response to the bacille Calmette Guerin vaccine or Mycobacterium tuberculosis infection. Journal of immunology 2009, 182, 7172–7177. [Google Scholar] [CrossRef]
- Nambiar, J.K.; Ryan, A.A.; Kong, C.U.; Britton, W.J.; Triccas, J.A. Modulation of pulmonary DC function by vaccine-encoded GM-CSF enhances protective immunity against Mycobacterium tuberculosis infection. European journal of immunology 2010, 40, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Covian, C.; Fernandez-Fierro, A.; Retamal-Diaz, A.; Diaz, F.E.; Vasquez, A.E.; Lay, M.K.; Riedel, C.A.; Gonzalez, P.A.; Bueno, S.M.; Kalergis, A.M. BCG-Induced Cross-Protection and Development of Trained Immunity: Implication for Vaccine Design. Frontiers in immunology 2019, 10, 2806. [Google Scholar] [CrossRef]
- Nascimento, I.P.; Dias, W.O.; Quintilio, W.; Christ, A.P.; Moraes, J.F.; Vancetto, M.D.; Ribeiro-Dos-Santos, G.; Raw, I.; Leite, L.C. Neonatal immunization with a single dose of recombinant BCG expressing subunit S1 from pertussis toxin induces complete protection against Bordetella pertussis intracerebral challenge. Microbes and infection 2008, 10, 198–202. [Google Scholar] [CrossRef]
- Kanno, A.I.; Boraschi, D.; Leite, L.C.C.; Rodriguez, D. Recombinant BCG Expressing the Subunit 1 of Pertussis Toxin Induces Innate Immune Memory and Confers Protection against Non-Related Pathogens. Vaccines 2022, 10. [Google Scholar] [CrossRef]
- Yu, J.S.; Peacock, J.W.; Jacobs, W.R., Jr.; Frothingham, R.; Letvin, N.L.; Liao, H.X.; Haynes, B.F. Recombinant Mycobacterium bovis bacillus Calmette-Guerin elicits human immunodeficiency virus type 1 envelope-specific T lymphocytes at mucosal sites. Clinical and vaccine immunology : CVI 2007, 14, 886–893. [Google Scholar] [CrossRef]
- Chapman, R.; Stutz, H.; Jacobs, W., Jr.; Shephard, E.; Williamson, A.L. Priming with recombinant auxotrophic BCG expressing HIV-1 Gag, RT and Gp120 and boosting with recombinant MVA induces a robust T cell response in mice. PloS one 2013, 8, e71601. [Google Scholar] [CrossRef]
- Lay, M.K.; Cespedes, P.F.; Palavecino, C.E.; Leon, M.A.; Diaz, R.A.; Salazar, F.J.; Mendez, G.P.; Bueno, S.M.; Kalergis, A.M. Human metapneumovirus infection activates the TSLP pathway that drives excessive pulmonary inflammation and viral replication in mice. European journal of immunology 2015, 45, 1680–1695. [Google Scholar] [CrossRef]
- Palavecino, C.E.; Cespedes, P.F.; Gomez, R.S.; Kalergis, A.M.; Bueno, S.M. Immunization with a recombinant bacillus Calmette-Guerin strain confers protective Th1 immunity against the human metapneumovirus. Journal of immunology 2014, 192, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Soto, J.A.; Galvez, N.M.S.; Pacheco, G.A.; Canedo-Marroquin, G.; Bueno, S.M.; Kalergis, A.M. Induction of Protective Immunity by a Single Low Dose of a Master Cell Bank cGMP-rBCG-P Vaccine Against the Human Metapneumovirus in Mice. Frontiers in cellular and infection microbiology 2021, 11, 662714. [Google Scholar] [CrossRef]
- Cespedes, P.F.; Rey-Jurado, E.; Espinoza, J.A.; Rivera, C.A.; Canedo-Marroquin, G.; Bueno, S.M.; Kalergis, A.M. A single, low dose of a cGMP recombinant BCG vaccine elicits protective T cell immunity against the human respiratory syncytial virus infection and prevents lung pathology in mice. Vaccine 2017, 35, 757–766. [Google Scholar] [CrossRef]
- Yang, G.; Luo, T.; Sun, C.; Yuan, J.; Peng, X.; Zhang, C.; Zhai, X.; Bao, L. PPE27 in Mycobacterium smegmatis Enhances Mycobacterial Survival and Manipulates Cytokine Secretion in Mouse Macrophages. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research 2017, 37, 421–431. [Google Scholar] [CrossRef]
- Peng, X.; Zhou, Y.; Zhang, B.; Liang, X.; Feng, J.; Huang, Y.; Weng, S.; Xu, Y.; Su, H. Mucosal recombinant BCG vaccine induces lung-resident memory macrophages and enhances trained immunity via mTORC2/HK1-mediated metabolic rewiring. The Journal of biological chemistry 2024, 300, 105518. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.C.; de Magalhaes, M.T.Q.; Homan, E.J. Immunoinformatic Analysis of SARS-CoV-2 Nucleocapsid Protein and Identification of COVID-19 Vaccine Targets. Frontiers in immunology 2020, 11, 587615. [Google Scholar] [CrossRef] [PubMed]
- Mambelli, F.; Marinho, F.V.; Andrade, J.M.; de Araujo, A.; Abuna, R.P.F.; Fabri, V.M.R.; Santos, B.P.O.; da Silva, J.S.; de Magalhaes, M.T.Q.; Homan, E.J.; et al. Recombinant Bacillus Calmette-Guerin Expressing SARS-CoV-2 Chimeric Protein Protects K18-hACE2 Mice against Viral Challenge. Journal of immunology 2023, 210, 1925–1937. [Google Scholar] [CrossRef]
- Yi, M.; Li, T.; Niu, M.; Mei, Q.; Zhao, B.; Chu, Q.; Dai, Z.; Wu, K. Exploiting innate immunity for cancer immunotherapy. Molecular cancer 2023, 22, 187. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Oncology meets immunology: the cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef]
- Naletova, I.; Tomasello, B.; Attanasio, F.; Pleshkan, V.V. Prospects for the Use of Metal-Based Nanoparticles as Adjuvants for Local Cancer Immunotherapy. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Old, L.J.; Clarke, D.A.; Benacerraf, B. Effect of Bacillus Calmette-Guerin infection on transplanted tumours in the mouse. Nature 1959, 184 (Suppl 5), 291–292. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Nishimura, N.; Oda, Y.; Owari, T.; Hori, S.; Morizawa, Y.; Gotoh, D.; Nakai, Y.; Anai, S.; Torimoto, K.; et al. Intravesical Bacillus Calmette-Guerin treatment-induced sleep quality deterioration in patients with non-muscle invasive bladder cancer: functional outcome assessment based on a questionnaire survey and actigraphy. Supportive care in cancer : official journal of the Multinational Association of Supportive Care in Cancer 2022, 30, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Holzbeierlein, J.M.; Bixler, B.R.; Buckley, D.I.; Chang, S.S.; Holmes, R.; James, A.C.; Kirkby, E.; McKiernan, J.M.; Schuckman, A.K. Diagnosis and Treatment of Non-Muscle Invasive Bladder Cancer: AUA/SUO Guideline: 2024 Amendment. The Journal of urology 2024, 211, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Mulder, W.J.M.; Ochando, J.; Joosten, L.A.B.; Fayad, Z.A.; Netea, M.G. Therapeutic targeting of trained immunity. Nature reviews. Drug discovery 2019, 18, 553–566. [Google Scholar] [CrossRef]
- Pfahlberg, A.; Kolmel, K.F.; Grange, J.M.; Mastrangelo, G.; Krone, B.; Botev, I.N.; Niin, M.; Seebacher, C.; Lambert, D.; Shafir, R.; et al. Inverse association between melanoma and previous vaccinations against tuberculosis and smallpox: results of the FEBIM study. The Journal of investigative dermatology 2002, 119, 570–575. [Google Scholar] [CrossRef]
- Sfakianos, J.P.; Salome, B.; Daza, J.; Farkas, A.; Bhardwaj, N.; Horowitz, A. Bacillus Calmette-Guerin (BCG): Its fight against pathogens and cancer. Urologic oncology 2021, 39, 121–129. [Google Scholar] [CrossRef]
- Yang, J.; Jones, M.S.; Ramos, R.I.; Chan, A.A.; Lee, A.F.; Foshag, L.J.; Sieling, P.A.; Faries, M.B.; Lee, D.J. Insights into Local Tumor Microenvironment Immune Factors Associated with Regression of Cutaneous Melanoma Metastases by Mycobacterium bovis Bacille Calmette-Guerin. Frontiers in oncology 2017, 7, 61. [Google Scholar] [CrossRef]
- Gandhi, N.M.; Morales, A.; Lamm, D.L. Bacillus Calmette-Guerin immunotherapy for genitourinary cancer. BJU international 2013, 112, 288–297. [Google Scholar] [CrossRef]
- Morra, M.E.; Kien, N.D.; Elmaraezy, A.; Abdelaziz, O.A.M.; Elsayed, A.L.; Halhouli, O.; Montasr, A.M.; Vu, T.L.; Ho, C.; Foly, A.S.; et al. Early vaccination protects against childhood leukemia: A systematic review and meta-analysis. Scientific reports 2017, 7, 15986. [Google Scholar] [CrossRef]
- Usher, N.T.; Chang, S.; Howard, R.S.; Martinez, A.; Harrison, L.H.; Santosham, M.; Aronson, N.E. Association of BCG Vaccination in Childhood With Subsequent Cancer Diagnoses: A 60-Year Follow-up of a Clinical Trial. JAMA network open 2019, 2, e1912014. [Google Scholar] [CrossRef]
- Sui, Y.; Berzofsky, J.A. Trained immunity inducers in cancer immunotherapy. Frontiers in immunology 2024, 15, 1427443. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune network 2018, 18, e27. [Google Scholar] [CrossRef] [PubMed]
- van Puffelen, J.H.; Novakovic, B.; van Emst, L.; Kooper, D.; Zuiverloon, T.C.M.; Oldenhof, U.T.H.; Witjes, J.A.; Galesloot, T.E.; Vrieling, A.; Aben, K.K.H.; et al. Intravesical BCG in patients with non-muscle invasive bladder cancer induces trained immunity and decreases respiratory infections. Journal for immunotherapy of cancer 2023, 11. [Google Scholar] [CrossRef]
- Udagawa, M.; Kudo-Saito, C.; Hasegawa, G.; Yano, K.; Yamamoto, A.; Yaguchi, M.; Toda, M.; Azuma, I.; Iwai, T.; Kawakami, Y. Enhancement of immunologic tumor regression by intratumoral administration of dendritic cells in combination with cryoablative tumor pretreatment and Bacillus Calmette-Guerin cell wall skeleton stimulation. Clinical cancer research : an official journal of the American Association for Cancer Research 2006, 12, 7465–7475. [Google Scholar] [CrossRef]
- Yao, K.; Wang, W.; Li, H.; Lin, J.; Tan, W.; Chen, Y.; Guo, L.; Lin, D.; Chen, T.; Zhou, J.; et al. Bacillus Calmette Guerin (BCG) activates lymphocyte to promote autophagy and apoptosis of gastric cancer MGC-803 cell. Cellular and molecular biology 2018, 64, 11–16. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, C.; Cruces, K.P.; Riestra Ayora, J.; Martin-Sanz, E.; Sanz-Fernandez, R. BCG immune activation reduces growth and angiogenesis in an in vitro model of head and neck squamous cell carcinoma. Vaccine 2017, 35, 6395–6403. [Google Scholar] [CrossRef]
- Zhang, Y.; Morgan, M.J.; Chen, K.; Choksi, S.; Liu, Z.G. Induction of autophagy is essential for monocyte-macrophage differentiation. Blood 2012, 119, 2895–2905. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Shi, F.; Zhou, Z.; Sun, F.; Sun, M.H.; Sun, Q.; Chen, L.; Li, D.; Jiang, C.Y.; Zhao, R.Z.; et al. M1 macrophage mediated increased reactive oxygen species (ROS) influence wound healing via the MAPK signaling in vitro and in vivo. Toxicology and applied pharmacology 2019, 366, 83–95. [Google Scholar] [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Frontiers in immunology 2020, 11, 583084. [Google Scholar] [CrossRef]
- Ahmadi-Dehlaghi, F.; Mohammadi, P.; Valipour, E.; Pournaghi, P.; Kiani, S.; Mansouri, K. Autophagy: A challengeable paradox in cancer treatment. Cancer medicine 2023, 12, 11542–11569. [Google Scholar] [CrossRef]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. International journal of molecular sciences 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Deleyto-Seldas, N.; Efeyan, A. The mTOR-Autophagy Axis and the Control of Metabolism. Frontiers in cell and developmental biology 2021, 9, 655731. [Google Scholar] [CrossRef] [PubMed]
- Puente, C.; Hendrickson, R.C.; Jiang, X. Nutrient-regulated Phosphorylation of ATG13 Inhibits Starvation-induced Autophagy. The Journal of biological chemistry 2016, 291, 6026–6035. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell 2004, 119, 753–766. [Google Scholar] [CrossRef]
- Gonzalez-Orozco, M.; Strong, E.J.; Paroha, R.; Lee, S. Reversing BCG-mediated autophagy inhibition and mycobacterial survival to improve vaccine efficacy. BMC immunology 2022, 23, 43. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Bakhru, P.; Saikolappan, S.; Das, K.; Soudani, E.; Singh, C.R.; Estrella, J.L.; Zhang, D.; Pasare, C.; Ma, Y.; et al. An autophagy-inducing and TLR-2 activating BCG vaccine induces a robust protection against tuberculosis in mice. NPJ vaccines 2019, 4, 34. [Google Scholar] [CrossRef]
- Aceves-Sanchez, M.J.; Barrios-Payan, J.A.; Segura-Cerda, C.A.; Flores-Valdez, M.A.; Mata-Espinosa, D.; Pedroza-Roldan, C.; Yadav, R.; Saini, D.K.; de la Cruz, M.A.; Ares, M.A.; et al. BCG∆BCG1419c and BCG differ in induction of autophagy, c-di-GMP content, proteome, and progression of lung pathology in Mycobacterium tuberculosis HN878-infected male BALB/c mice. Vaccine 2023, 41, 3824–3835. [Google Scholar] [CrossRef]
- Iivanainen, S.; Koivunen, J.P. Possibilities of Improving the Clinical Value of Immune Checkpoint Inhibitor Therapies in Cancer Care by Optimizing Patient Selection. International journal of molecular sciences 2020, 21. [Google Scholar] [CrossRef]
- Valero, C.; Lee, M.; Hoen, D.; Zehir, A.; Berger, M.F.; Seshan, V.E.; Chan, T.A.; Morris, L.G.T. Response Rates to Anti-PD-1 Immunotherapy in Microsatellite-Stable Solid Tumors With 10 or More Mutations per Megabase. JAMA oncology 2021, 7, 739–743. [Google Scholar] [CrossRef]
- Cardillo, F.; Bonfim, M.; da Silva Vasconcelos Sousa, P.; Mengel, J.; Ribeiro Castello-Branco, L.R.; Pinho, R.T. Bacillus Calmette-Guerin Immunotherapy for Cancer. Vaccines 2021, 9. [Google Scholar] [CrossRef]
- Ylosmaki, E.; Fusciello, M.; Martins, B.; Feola, S.; Hamdan, F.; Chiaro, J.; Ylosmaki, L.; Vaughan, M.J.; Viitala, T.; Kulkarni, P.S.; et al. Novel personalized cancer vaccine platform based on Bacillus Calmette-Guerin. Journal for immunotherapy of cancer 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Yim, K.; Melnick, K.; Mott, S.L.; Carvalho, F.L.F.; Zafar, A.; Clinton, T.N.; Mossanen, M.; Steele, G.S.; Hirsch, M.; Rizzo, N.; et al. Sequential intravesical gemcitabine/docetaxel provides a durable remission in recurrent high-risk NMIBC following BCG therapy. Urologic oncology 2023, 41, 458 e451-458 e457. [Google Scholar] [CrossRef] [PubMed]
- Abou Chakra, M.; Packiam, V.T.; Duquesne, I.; Peyromaure, M.; McElree, I.M.; O’Donnell, M.A. Combination intravesical chemotherapy for non-muscle invasive bladder cancer (NMIBC) as first-line or rescue therapy: where do we stand now? Expert opinion on pharmacotherapy 2024, 25, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Rentsch, C.A.; Bosshard, P.; Mayor, G.; Rieken, M.; Puschel, H.; Wirth, G.; Cathomas, R.; Parzmair, G.P.; Grode, L.; Eisele, B.; et al. Results of the phase I open label clinical trial SAKK 06/14 assessing safety of intravesical instillation of VPM1002BC, a recombinant mycobacterium Bacillus Calmette Guerin (BCG), in patients with non-muscle invasive bladder cancer and previous failure of conventional BCG therapy. Oncoimmunology 2020, 9, 1748981. [Google Scholar] [CrossRef]
- https://newtbvaccines.org/vaccine/vpm1002/. Available online: https://newtbvaccines.org/vaccine/vpm1002/ (accessed on 26 Octomber).
- Rentsch, C.A.; Thalmann, G.N.; Lucca, I.; Kwiatkowski, M.; Wirth, G.J.; Strebel, R.T.; Engeler, D.; Pedrazzini, A.; Huttenbrink, C.; Schultze-Seemann, W.; et al. A Phase 1/2 Single-arm Clinical Trial of Recombinant Bacillus Calmette-Guerin (BCG) VPM1002BC Immunotherapy in Non-muscle-invasive Bladder Cancer Recurrence After Conventional BCG Therapy: SAKK 06/14. European urology oncology 2022, 5, 195–202. [Google Scholar] [CrossRef]
- Kuhtreiber, W.M.; Tran, L.; Kim, T.; Dybala, M.; Nguyen, B.; Plager, S.; Huang, D.; Janes, S.; Defusco, A.; Baum, D.; et al. Long-term reduction in hyperglycemia in advanced type 1 diabetes: the value of induced aerobic glycolysis with BCG vaccinations. NPJ vaccines 2018, 3, 23. [Google Scholar] [CrossRef]
- Dias, H.F.; Mochizuki, Y.; Kuhtreiber, W.M.; Takahashi, H.; Zheng, H.; Faustman, D.L. Bacille Calmette Guerin (BCG) and prevention of types 1 and 2 diabetes: Results of two observational studies. PloS one 2023, 18, e0276423. [Google Scholar] [CrossRef]
- https://www.faustmanlab.org/publications/.
- Haahr, S.; Michelsen, S.W.; Andersson, M.; Bjorn-Mortensen, K.; Soborg, B.; Wohlfahrt, J.; Melbye, M.; Koch, A. Non-specific effects of BCG vaccination on morbidity among children in Greenland: a population-based cohort study. International journal of epidemiology 2016, 45, 2122–2130. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Ravn, H.; Birk, N.M.; Kjaergaard, J.; Nissen, T.N.; Pihl, G.T.; Thostesen, L.M.; Greisen, G.; Jeppesen, D.L.; Kofoed, P.E.; et al. BCG Vaccination at Birth and Rate of Hospitalization for Infection Until 15 Months of Age in Danish Children: A Randomized Clinical Multicenter Trial. Journal of the Pediatric Infectious Diseases Society 2019, 8, 213–220. [Google Scholar] [CrossRef]
- de Castro, M.J.; Pardo-Seco, J.; Martinon-Torres, F. Nonspecific (Heterologous) Protection of Neonatal BCG Vaccination Against Hospitalization Due to Respiratory Infection and Sepsis. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America 2015, 60, 1611–1619. [Google Scholar] [CrossRef]
- Schaltz-Buchholzer, F.; Biering-Sorensen, S.; Lund, N.; Monteiro, I.; Umbasse, P.; Fisker, A.B.; Andersen, A.; Rodrigues, A.; Aaby, P.; Benn, C.S. Early BCG Vaccination, Hospitalizations, and Hospital Deaths: Analysis of a Secondary Outcome in 3 Randomized Trials from Guinea-Bissau. The Journal of infectious diseases 2019, 219, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Nankabirwa, V.; Tumwine, J.K.; Mugaba, P.M.; Tylleskar, T.; Sommerfelt, H.; Group, P.-E.S. Child survival and BCG vaccination: a community based prospective cohort study in Uganda. BMC public health 2015, 15, 175. [Google Scholar] [CrossRef]
- Gofrit, O.N.; Bercovier, H.; Klein, B.Y.; Cohen, I.R.; Ben-Hur, T.; Greenblatt, C.L. Can immunization with Bacillus Calmette-Guerin (BCG) protect against Alzheimer’s disease? Medical hypotheses 2019, 123, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Klinger, D.; Hill, B.L.; Barda, N.; Halperin, E.; Gofrit, O.N.; Greenblatt, C.L.; Rappoport, N.; Linial, M.; Bercovier, H. Bladder Cancer Immunotherapy by BCG Is Associated with a Significantly Reduced Risk of Alzheimer’s Disease and Parkinson’s Disease. Vaccines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Makrakis, D.; Holt, S.K.; Bernick, C.; Grivas, P.; Gore, J.L.; Wright, J.L. Intravesical BCG and Incidence of Alzheimer Disease in Patients With Bladder Cancer: Results From an Administrative Dataset. Alzheimer disease and associated disorders 2022, 36, 307–311. [Google Scholar] [CrossRef]
- Singh, A.K.; Praharaj, M.; Lombardo, K.A.; Yoshida, T.; Matoso, A.; Baras, A.S.; Zhao, L.; Srikrishna, G.; Huang, J.; Prasad, P.; et al. Re-engineered BCG overexpressing cyclic di-AMP augments trained immunity and exhibits improved efficacy against bladder cancer. Nature communications 2022, 13, 878. [Google Scholar] [CrossRef]
- Du, J.; Xu, R. RORalpha, a potential tumor suppressor and therapeutic target of breast cancer. International journal of molecular sciences 2012, 13, 15755–15766. [Google Scholar] [CrossRef]
- Vinogradova, T.V.; Chernov, I.P.; Monastyrskaya, G.S.; Kondratyeva, L.G.; Sverdlov, E.D. Cancer Stem Cells: Plasticity Works against Therapy. Acta naturae 2015, 7, 46–55. [Google Scholar] [CrossRef]
- Huang, Y.; Hong, W.; Wei, X. The molecular mechanisms and therapeutic strategies of EMT in tumor progression and metastasis. Journal of hematology & oncology 2022, 15, 129. [Google Scholar] [CrossRef]
- Fontana, R.; Mestre-Farrera, A.; Yang, J. Update on Epithelial-Mesenchymal Plasticity in Cancer Progression. Annual review of pathology 2024, 19, 133–156. [Google Scholar] [CrossRef]
- Xiong, G.; Xu, R. Retinoid orphan nuclear receptor alpha (RORalpha) suppresses the epithelial-mesenchymal transition (EMT) by directly repressing Snail transcription. The Journal of biological chemistry 2022, 298, 102059. [Google Scholar] [CrossRef] [PubMed]
- Kilic, G.; Matzaraki, V.; Bulut, O.; Baydemir, I.; Ferreira, A.V.; Rabold, K.; Moorlag, S.; Koeken, V.; de Bree, L.C.J.; Mourits, V.P.; et al. RORalpha negatively regulates BCG-induced trained immunity. Cellular immunology 2024, 403-404, 104862. [Google Scholar] [CrossRef] [PubMed]
- Zarenezhad, E.; Kanaan, M.H.G.; Abdollah, S.S.; Vakil, M.K.; Marzi, M.; Mazarzaei, A.; Ghasemian, A. Metallic Nanoparticles: Their Potential Role in Breast Cancer Immunotherapy via Trained Immunity Provocation. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, W.; Zheng, P.; Yang, Y.; Liu, Q.; Hu, Y.; He, J.; Long, Q.; Ma, Y. A “trained immunity” inducer-adjuvanted nanovaccine reverses the growth of established tumors in mice. Journal of nanobiotechnology 2023, 21, 74. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, Y.; Milling, L.E.; Chang, J.Y.H.; Santollani, L.; Sheen, A.; Lutz, E.A.; Tabet, A.; Stinson, J.; Ni, K.; Rodrigues, K.A.; et al. Intratumourally injected alum-tethered cytokines elicit potent and safer local and systemic anticancer immunity. Nature biomedical engineering 2022, 6, 129–143. [Google Scholar] [CrossRef]
- Lutz, E.A.; Agarwal, Y.; Momin, N.; Cowles, S.C.; Palmeri, J.R.; Duong, E.; Hornet, V.; Sheen, A.; Lax, B.M.; Rothschilds, A.M.; et al. Alum-anchored intratumoral retention improves the tolerability and antitumor efficacy of type I interferon therapies. Proceedings of the National Academy of Sciences of the United States of America 2022, 119, e2205983119. [Google Scholar] [CrossRef]
- Nascimento, L.V.; Santos, C.C.; Leite, L.C.; Nascimento, I.P. Characterisation of alternative expression vectors for recombinant Bacillus Calmette-Guerin as live bacterial delivery systems. Memorias do Instituto Oswaldo Cruz 2020, 115, e190347. [Google Scholar] [CrossRef]
- Editorial. The ‘war on cancer’ isn’t yet won. Nature 2022, 601, 297. [Google Scholar] [CrossRef]
- Alekseenko, I.; Zhukova, L.; Kondratyeva, L.; Buzdin, A.; Chernov, I.; Sverdlov, E. Tumor Cell Communications as Promising Supramolecular Targets for Cancer Chemotherapy: A Possible Strategy. International journal of molecular sciences 2024, 25. [Google Scholar] [CrossRef]
Figure 1.
Simplified scheme of BCG action. A) Signaling pathways mediating interactions with BCG. B) TRIM increase response diagram.
Figure 1.
Simplified scheme of BCG action. A) Signaling pathways mediating interactions with BCG. B) TRIM increase response diagram.
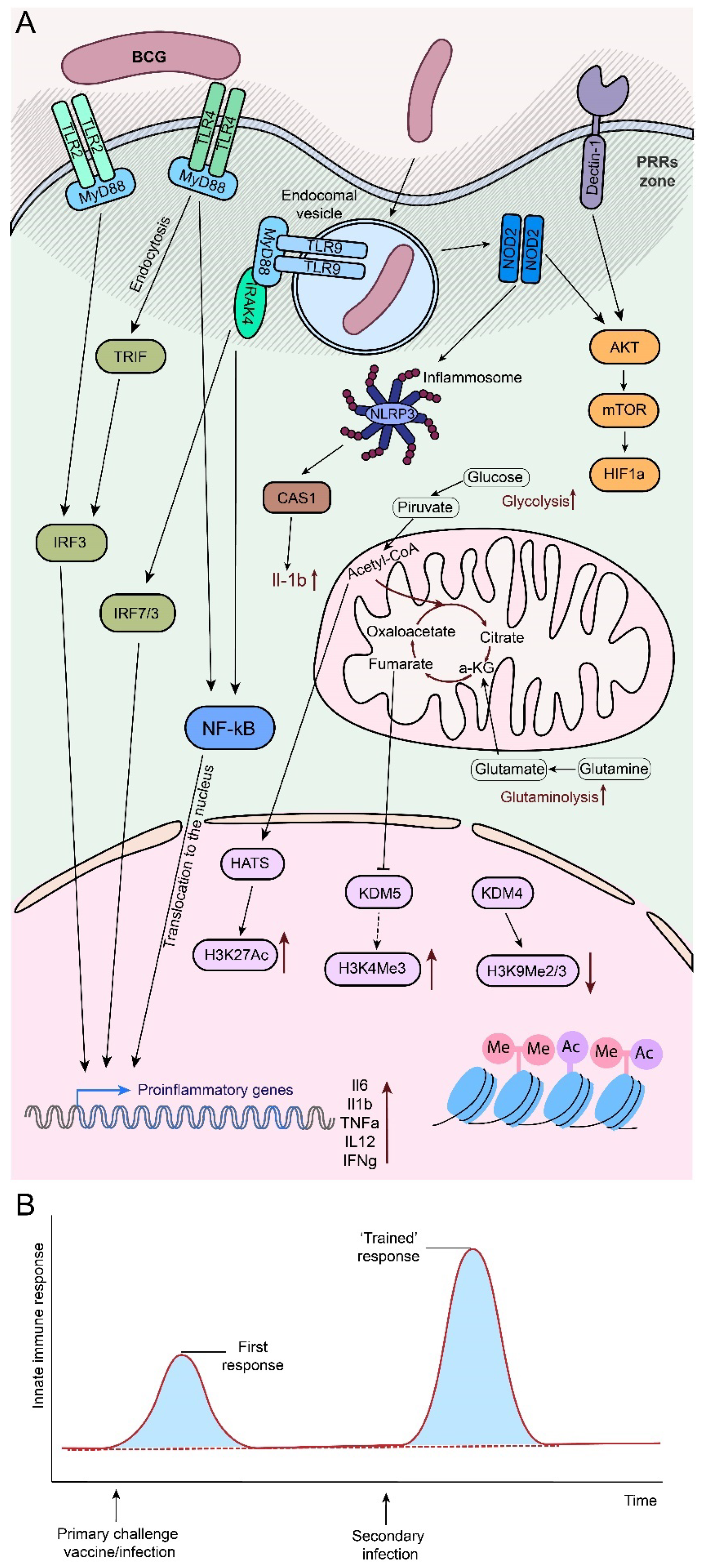
Figure 2.
Distribution of actual clinical trials with BCG. The distribution by conditions of current clinical trials using BCG is shown. The number of such studies (n=) and percentage of the total are given. A total of 62 CTs were analyzed. The diagram on the right shows the distribution by BCG use options for the cancer group.
Figure 2.
Distribution of actual clinical trials with BCG. The distribution by conditions of current clinical trials using BCG is shown. The number of such studies (n=) and percentage of the total are given. A total of 62 CTs were analyzed. The diagram on the right shows the distribution by BCG use options for the cancer group.
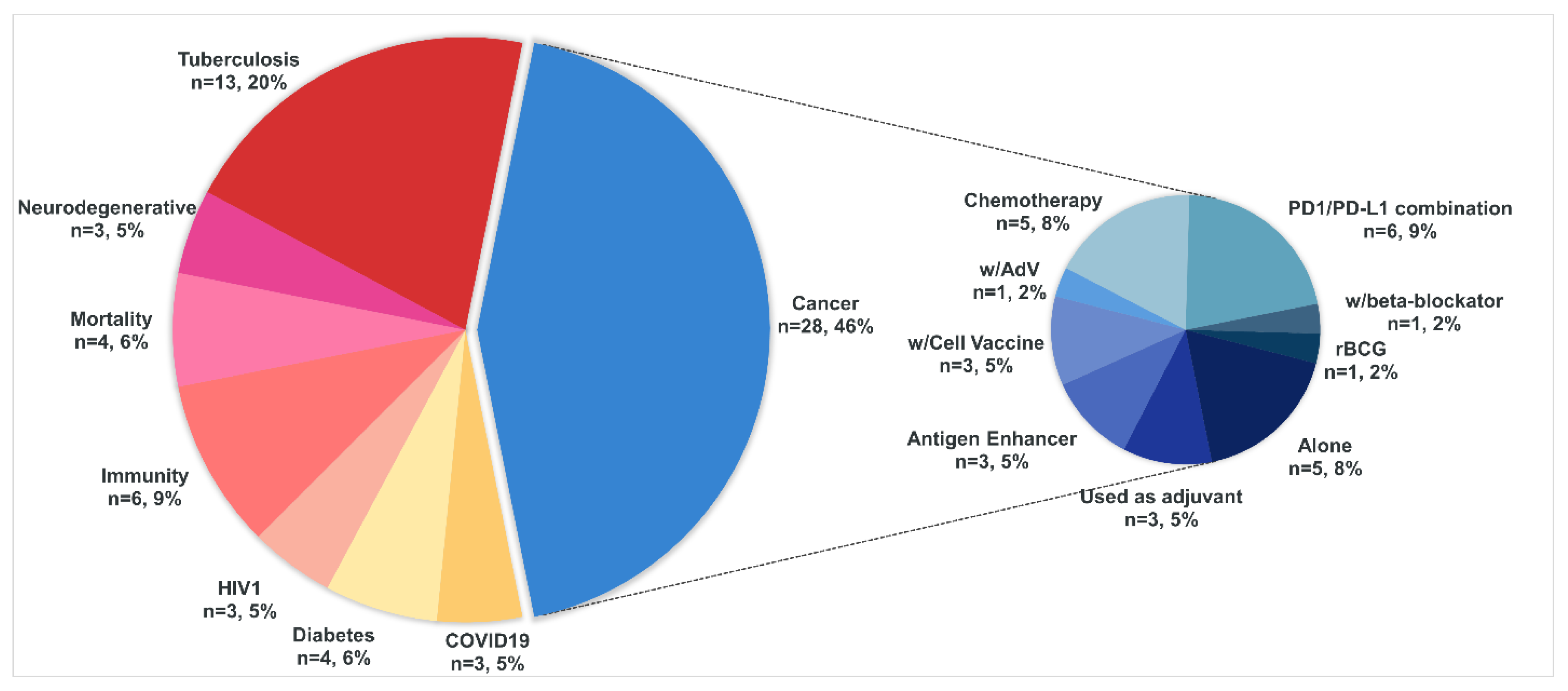

Table 1.
The main mechanisms triggered by BCG/TRIM effectors and their impact on immune cells.
| Effector | In immunity | In Cancer |
|---|---|---|
| GM-CSF | APC differentiation and expansion, Monocytes activation (↑HLA-DR; ↑CD86; ↑TNFα; ↑IL-1β) [56], differentiation to DC, MΦ [57,58] | Dual role in cancer progression [59,60] Myeloid DCs differentiation, enhancing Th1 responses [61] Th9 cell differentiation, antitumor cytotoxic T lymphocyte responses ↑ [62] Excessive - exhaust immune cells, cancer growth ↑; insufficient - immune surveillance ↓ [60] |
| Il-1β | Main inflammation and fever activator [63] Autoinflammatory syndromes [64] IL-1β signaling activates NF-κB and MAPK pathways, inducing expression of various inflammatory genes [65] T cell polarization towards Th1 and Th17 subtypes [66] IFNs signaling ↑, immunity against some viral infections ↑ [67] |
Resolution of acute inflammation leading to the induction of adaptive anti-tumor responses; chronic inflammatory - risk of developing cancer ↑ [68] Chronic inflammation - tumor growth ↑, metastasis ↑ [66] In TME: angiogenesis ↑, tumor growth, metastasis ↑ [69,70] Suppresses anti-tumor immune responses, metastasis ↑ [71] Immunosuppressive monocytes polarization to IL-10 MΦ, CD8+ T inhibition [72] Angiogenesis ↑, antiapoptotic ↑, MDSC ↑ [66] |
| IFNs | Induce antiviral, antiproliferative, and immunomodulatory effects; enhance antigen presentation, [73]. Inflammatory cytokines production↑, anti-inflammatory ↓ [74,75] NK cytotoxic activity ↑, MΦ phagocytosis↑ and pro-inflammatory cytokines production↑; DCs maturation and activation, antigen presentation↑, T cells activation ↑ [76,77] CD4+T >> Th1; CD8+T activation↑, proliferation↑ [78,79,80] |
Persistent IFN expression can lead to immunosuppression in later stages of tumor progression [81]. Cancer-immunity cycle: antigen release, T cell activation, tumor cell killing [82]. Chronic stimulation - tumor resistance ↑ [83] IFNγ tumor angiogenesis↓; Treg apoptosis↑, proinflammatory M1 MΦ antitumor activity ↑ immunosuppression↑, T cell apoptosis↑, immune tolerance↑ [84] Cancer cell stemness ↑, metastasis ↑ [85,86]. EMT↑ [87]. >PD-L1&IDO↑ immune evasion↑ [88] |
| TNFα | Major pro-inflammatory cytokine [89,90] Activates various immune cells, including MΦs and NFs, pathogen phagocytosis ↑; inflammatory mediators ↑ [89],[91] Apoptosis ↑ [90,92] ILs ↑ (e.g., IL-1, IL-6); IFNs ↑ [89,93] Excessive activation – chronic inflammation – autoimmune diseases [89] |
Pro-inflammatory cancer permissive TME [91,94] Immunosuppressive TME [95] NF-κB activation - cancer survival ↑, apoptosis resistance ↑, invasion ↑, metastasis ↑ [96,97]. VEGF↑, angiogenesis↑ [98] |
| IL-6 | Pro-inflammatory [99] B cells activation and differentiation; Ig production; humoral immune response [100] Naive CD4+ T → Th17 cells; Treg activity modulation; prevent autoimmunity [101] C-reactive protein production [102] Hematopoiesis regulation, HSCs differentiation [103] Metabolic regulation, insulin sensitivity and lipid metabolism [104] |
Dual action in TME - drives malignancy/promotes anti-tumor adaptive immunity [105] Proliferation and survival of cancer cells↑, JAK/STAT3↑, cell growth↑, apoptosis↓ [106] Lymphangiogenesis ↑, VEGF↑, tumor growth↑, invasion ↑ [107] Immune escape↑, immunosuppressive TME↑, Tregs↑, MDSCs↑ [108] N-cadherin↑ E-cadherin↓ EMT↑, metastasis↑, migration↑, invasion↑ [109] |
| Th1 polarization | Th1 - immune response against intracellular pathogens, primarily through the production of cytokines like IFNγ and TNFα MΦ activation↑, phagocytosis↑, CD8+T&NK functionality↑, Th2 response↓ [110,111,112] IFNg secretion [113] |
Th1 - anti-tumor immunity: production IFNγ↑ > cytotoxic activity of CD8+ T/NK cells↑. MΦ activation↑, >M1 MΦ phenotype pro-inflammatory, phagocytosis↑, ROS↑, tumor growth↓, angiogenesis↓, tumor immune evasion↓ [114,115] |
| Acetate/ acetyl-CoA | Acetyl-CoA - H3K27ac acetylation [116] Metabolic supply for Tcells & MΦ, Tregs differentiation, Th2 functionality, Anti-inflammatory, inhibiting pro-inflammatory cytokine production, anti-inflammatory M2 MΦ↑ [117,118,119] |
Acetate increase the expressions of SNAI1 and ACSS2 under glucose limitation. ACSS2 knockdown > SNAI1↓, cell migration↓; ACSS2 overexpression > SNAI1↑, histone H3K27 acetylation (H3K27ac)↑, metastasis↑, migration↑ [116] ACSS2 catalyzes the conversion of acetate to acetyl-CoA [120] ACSS2 inhibition – impair tumor cell metabolism and potentiate antitumor immunity [120] Regulation of acetyl-CoA metabolic enzymes may support tumor growth and TME survival [121] |
Designations: ↑- activation; ↓ - inhibition; >> - differentiation; > - stimulation/activation; ACSS2 acetyl-CoA synthetase 2. EMT epithelial–mesenchymal transition; HSCs hematopoietic stem cells. IFNγ interferon-gamma. MDSC Myeloid-derived suppressor cells; MΦ macrophages; TME tumor microenvironment; TNFα tumor necrosis factor-alpha; ROS reactive oxygen species.
Table 2.
Various factors hindering the development of TRIM.
| Factor | In immunity | In Cancer |
|---|---|---|
| NOD2-deficienсy | Fail in infection resistance Fail in TRIM formation [123] |
Pro-tumorigenic MΦ, promote lung adenocarcinoma progression [123] |
| Butyrate | Histone deacetylase 3 (HDAC3) inhibition, HDAC3 deacetylates H3K27ac and H3K9ac – active promoter/open chromatin marks [124] | Therapeutic target in cancer [125] |
| Fumarate | Inhibitory effect to the activity of KDM5, induce enrichment of H3K4me3 on promoters [126] |
Fumarate hydratase loss is possibly pro-tumorigenic [126] Fumarate accumulation and cause hereditary leiomyomatosis and renal cell cancer, supporting its oncometabolite role [127,128] |
| Ascorbate | Antioxidative and Anti-Inflammatory [129] Abrogate TRIM inhibiting HIF1a [130] |
One of the most controversial drugs, despite many articles about antitumor action, it is suggested that there is not sufficient evidence base. Thus, at the time (Apr 5, 2024), the FDA has not approved the use of IV vitamin C as a treatment for cancer [131] Facilitates tumour cell death through the generation of ROS and ferroptosis [132] Potential anti-tumor capabilities [133] Boosts DNA damage and cell death in melanoma cells [134] High concentrations triggering cell death and low amounts promoting tumor growth [135] |
| Rapamycin, 2-deoxyglucose (2-DG) or metformin | Inhibition of the mTOR pathway or glycolysis [136] | Rapamycin anti-cancer role discussion [137,138]. Metformin and 2DG exhibit multiple metabolic and immunomodulatory anti-cancer effects, suppressed proliferation or PD-L1 expression [139] |
Table 3.
Clinical trials with rBCG strains.
| Name | Modification | Studies | Refs |
|---|---|---|---|
| VPM1002 | Recombinant, urease C-deficient, listeriolysin expressing BCG vaccine strain generated in order to induce a broader immune response against mycobacterial antigens. Consequently, VPM1002 was formulated as VPM1002BC for intravesical immunotherapy. | NCT04387409, P3 NCT04351685, P3 NCT03152903, P2 |
[227] |
| VPM1002BC | Recombinant live BCG strain in which the urease C gene is deleted by insertion of the gene encoding the haemolysin listeriolysin (LLO). | NCT02371447,P1/2 NCT04630730; P2 |
[228] [226] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



