
Submitted:
22 October 2024
Posted:
24 October 2024
You are already at the latest version
Abstract
Global demand for blueberries has surged in recent years, primarily due to their health benefits. However, blueberries postharvest losses, particularly firmness loss and decay, remain significant challenges. This study applied high concentrations of CO2 (15% and 20%) for 3 days at 1 °C to highbush (Duke) and rabbiteye (Ochlockonee) blueberries to assess their effects on maintaining quality, especially firmness, during low-temperature storage. Various quality parameters were evaluated, including titratable acidity, pH, total soluble solids, weight loss, and decay. The impact of gaseous treatments on firmness was analysed through mechanical parameters and the expression of genes related to cell wall integrity (XTH23, PL8, PG, PM3, EXP4, and VcGH5). The effectiveness of the treatments varied between species. High CO2 levels successfully reduced decay in both cultivars, but only the highbush variety (Duke) showed improvements in firmness. In Duke, CO2 treatments influenced the expression of genes associated to cell wall integrity, such as XTH23, PL8, and GH5. In contrast, the roles of PG and PME in firmness maintenance were minimal, showing no significant differences between treatments. While CO2 did not enhance firmness in Ochlockonee, it effectively reduced weight loss, highlighting the need for tailored postharvest strategies for different blueberry cultivars.
Keywords:
Vaccinium
; low temperature
; mechanical properties
; gene expression
; cell wall
1. Introduction
Blueberry (Vaccinium spp.) is a soft fruit popular among consumers due to its distinctive taste and high nutrient content, which includes phenolic acids and anthocyanins [1,2]. However, once harvested, the quality of blueberries is compromised due to their high respiration rate, softening and high susceptibility to fungal attack [3]. Although it is recommended to store blueberries at a temperature close to 0 °C to prolong their postharvest life, this condition alone is not sufficient to maintain quality as it can also promote softening, pedicel pitting, pericarp and pulp adhesion, and fungal decay [4]. While various studies have explored the development of technologies combined with low temperature to preserve the quality of soft berries postharvest, much of the focus has been on strawberries [5,6,7,8]. It is established that high CO2 concentrations, ranging from 15 to 20%, can mitigate fungal attack, lower respiration rates, and reduce softening in blueberries, thereby extending their shelf life [3]. Physical methods such as controlled atmosphere (CA) and modified atmosphere packaging (MAP) have been employed to extend the postharvest shelf life of blueberries [9,10,11]. Previous research has shown that short-term exposure (2-3 days) to 20% CO2 effectively maintains the quality of table grapes and strawberries stored at low temperature [6,12]. Although these short-term gaseous treatments may reduce costs compared to traditional CA by minimizing the duration of exposure to non-atmospheric conditions, their effectiveness in blueberries remains unstudied.
When assessing the efficacy of postharvest treatments on blueberries, firmness is one of the most important parameters to consider. On the other hand, firmness is a genetically controlled trait [13], so the genotype of a given blueberry is a factor influencing it. Therefore, changes in postharvest firmness can be very different between the most widely cultivated blueberry species in the world, the highbush (V. corymbosum L.) and the rabbiteye (V. ashei Reade) blueberries. Rivera et al. [14] found that mechanical parameters from penetration tests effectively assess blueberry firmness, revealing how postharvest controlled atmosphere conditions and genetic variations impact quality. The review highlighted that this method is valuable for distinguishing between berries, influenced by factors such as genetic differences [15], postharvest conditions [16,17] and maturity stage [18]. In earlier research, Rivera et al. [18] examined key mechanical properties related to moisture loss in two blueberry cultivars, Nui and Rahi, using double compression determined by the texture profile analysis (TPA) and puncture tests. Their findings indicated a strong correlation between water loss and hardness measured by TPA test, as well as the displacement at the berry skin break from the puncture test. Consequently, these mechanical parameters can serve as effective tools for quantifying changes in stored blueberries and tracking ripening progress in different cultivars. More recently, Sanchez-Ballesta et al. [19] used various mechanical parameters from a penetration test to measure the quality loss of 10 different commercial blueberries due to softening upon arrival at the retail market, just before consumer purchase, in terms of firmness. They found significant differences in the values of mechanical parameters associated with the skin-breaking (maximum skin-breaking force, distance required to break the skin, slope of the curve corresponding to skin penetration to break, and work required to break the berry skin).
The decrease in firmness is primarily caused by water loss and changes in the structure and composition of the cell wall [20]. During postharvest, fruit softening is closely linked to the disassembly of pectin and cellulose, which occurs gradually during storage. The degradation of cell wall components is carried out by the concerted action of several cell wall pectin-degrading enzymes, such as polygalacturonase (PG), cellulase (CL), β-galactosidase (β-Gal), and pectin methyl esterase (PME) [21,22,23]. Transgenic silencing of some of these genes reduced softening and increased the postharvest life of strawberries and apples [24,25]. Recently, FaPG1 knockout strawberry plants generated using the CRISPR/Cas9 system showed a reduced postharvest softening rate and fungal decay [26], suggesting that this gene plays a key role in maintaining firmness during postharvest.
In the case of blueberries, several studies have shown that the application of postharvest treatments helps to maintain fruit firmness and modulate the expression of genes encoding cell wall-modifying enzymes, with PG being one of the most studied. For instance, postharvest application of putrescine delayed fruit softening and inhibited VcPG1 expression levels [27]. Similarly, ethanol vapour delayed blueberry softening and downregulated PG gene expression [28]. However, the effect of the application of short-term gaseous treatments with high levels of CO2 on blueberry quality, particularly firmness, and the modulation of cell wall-modifying gene expression, remains unknown. Therefore, in this study, we have applied two gaseous treatments with different concentrations of CO2 (15% and 20%) for 3 days at 1 ºC to highbush (cv. Duke) and rabbiteye (cv. Ochlockonee) blueberries. We have evaluated their effectiveness in maintaining quality parameters and controlling total decay and water loss. Additionally, we have assessed how low temperature modified firmness and its preservation by the gaseous treatments by determining mechanical properties and the expression of genes related to cell wall-modifying enzymes, such as xyloglucan endotransglucosylase/hydrolase (XTH23), pectate lyase (PL8), PG, PME3, expansin (EXP4), and glycosyl hydrolase (GH5) during low-temperature storage for up to 29 days.
2. Results
2.1. Effect of Short-Term High CO2 Treatments and Storage at 1 ºC on Blueberry Quality and Weight Losses
The impact of short-term gaseous treatment on the quality parameters analysed depended on the cultivar (Duke and Ochlockonee) and the percentage of CO2 used. In Duke fruit, weight loss increased similarly for both treated and non-treated blueberries during cold storage. However, although low temperature increased weight loss in Ochlockonee fruit, the application of 20% of CO2 significantly reduced it by the end of cold storage compared to non-treated fruit (Figure 1).
The solid soluble content (SSC) of non-treated Duke fruit and those treated with 15% or 20% CO2 decreased significantly after 3 days at 0 °C (Table 1), with similar levels in all three cases. However, while the SSC content of fruit treated with 15% and 20% CO2 remained unchanged from day 3 until the end of storage, it increased in non-treated fruit to levels similar to those observed in freshly harvested fruit.
Titratable acidity (TA) increased in both, non-treated and CO2-treated Duke fruit on day 3. However, significant differences were observed between treated and non-treated Duke fruit at the end of the storage. In non-treated fruit, a decrease in TA was observed on day 29 compared to day 0 (Table 1). In treated fruit, TA varied according to the percentage of CO2. Duke blueberries treated with 15% CO2 showed an increase in TA on day 29 compared to freshly harvested fruit, whereas blueberries treated with 20% CO2 reached values similar to those observed on day 0. Since the maturation index (MI) is the ratio of SSC/TA, these changes affected this parameter, resulting in a significant decrease after 3 days in all conditions analysed. Non-treated Duke fruit showed a significant increase in MI compared to freshly harvested fruit, while fruit treated with 15% CO2 showed a decrease, and there was no change in fruit treated with 20% CO2. Regarding pH, this parameter increased significantly during storage of Duke fruit at 1 ºC under all conditions analysed, except for fruit treated with 15% CO2 after 3 days.
For Ochlockonee fruit, it should be noted that the SSC content of non-treated and 15% CO2-treated fruit increased significantly by the end of the storage compared to freshly harvested fruit (Table 2). However, no changes were observed in fruit treated with 20% CO2. Additionally, the TA of non-treated and 20% CO2-treated fruit increased during the storage at low temperature, whereas it decreased in fruit treated with 15% CO2. These changes in SSC and TA led to a decrease in MI in both non- and 20% CO2-treated fruit, while it increased in fruit treated with 15% of CO2. In addition, the pH increased significantly in non-treated and 15% CO2-treated Ochlockonee fruit stored at 1 ºC for up to 29 days compared to freshly harvested fruit, similar to Duke fruit, and remained unchanged in 20% CO2-treated fruit.
2.2. Mechanical Parameters
The study revealed significant differences in the shape of the force-distance curves (Figure 2) and the mechanical properties between CO2-treated and non-treated blueberries, as well as in the values of the various mechanical properties derived from them (Table 3 and Table 4). For example, the penetration force-distance curves obtained for Duke blueberries at the time of harvest and for non-treated and 20% CO2-treated fruit after 29 days of storage at 1 ºC are shown in Figure 2. The profiles show a noticeable peak force corresponding to the breaking of the skin or epidermis, followed by a large drop in force and then a continuous flow with much lower force than the maximum force when the probe penetrate the flesh. This fact suggests that skin strength has a greater effect on blueberry texture than the flesh. However, it should be noted that after 29 days of storage at low temperature, the slope and the distance or displacement at skin breaking show a decrease and an increase, respectively, compared to the values of the berries determined after the 20% CO2 treatment (Table 3). In addition, the maximum skin breaking force, as well as the slope at skin breaking and the penetration work of Duke blueberries, were also significantly lower in non-treated blueberries than in those treated with 15% and 20% CO2 for 29 days (Table 3).
After 3 and 14 days of storage, the skin-breaking force of Duke blueberries was lower for berries treated with both CO2 levels than for fruit stored in air (Table 3). However, after 29 days of storage at low temperature, berries stored in air were significantly softer than those treated with both CO2 levels. In contrast, for Ochlockonee blueberries, there was no significant difference in skin firmness between the berries at harvest and all the berries stored under the three different storage conditions and times (Table 4). Regarding the slope at skin breaking of Duke blueberries (Table 3), only the value corresponding to non-treated berries after 3 days of storage was significantly higher than that of freshly harvested berries and those stored for 3 days at both CO2 levels. As observed for breaking force, the lowest slope corresponded to non-treated blueberries for 29 days at 1 ºC. For Ochlockonee fruit (Table 4), the slope at skin breaking on day 3 of storage at 1 ºC was significantly higher for non-treated and 20% CO2-treated fruit than for freshly harvested blueberries. However, at the end of storage, no differences in this parameter were observed between fruit treated with both CO2 concentrations and untreated fruit.
Skin breaking distance values were the only parameter whose values increased with storage time in both treated and non-treated fruit of both cultivars compared to freshly harvested fruit (Table 3 and Table 4). However, while the increase occurred on day 29 in Duke, an increase was already observed on day 14 for Ochlockonee. It is also important to note the differences observed between the treated and non-treated samples analysed. In Duke, the highest increase was observed in the non-treated fruit stored for 29 days (Table 3). However, while the increase observed in the Ochlockonee fruit at 14 days did not show any significant differences between the different samples analysed, the fruit treated with 20% CO2 showed the smallest increase at 29 days (Table 4).
However, for both cultivars, very few significant differences in skin breaking work were observed both at harvest and during storage under the different conditions and time periods analysed (Table 3 and Table 4). Only non-treated Duke blueberries showed a lower penetration work value at 29 days than at harvest (Table 3).
Finally, a positive and significant correlation was found between force, slope, and work, while a negative and significant correlation was observed between slope and distance in both cultivars (data not shown).
2.3. Relative Expression of Genes Encoding Cell Wall-Modifying Enzymes
The temporal expression patterns of six genes encoding cell wall-modifying enzymes were analysed during low-temperature storage of non-treated and CO2-treated Duke and Ochlockonee blueberries using real-time PCR (Figure 3). Differences in gene expression were observed between Duke and Ochlockonee fruit.
An induction of XTH23 expression was observed after 14 days of storage of Duke fruit at 1 ºC, which was higher in both non-treated and 20% CO2-treated fruit. However, by day 29, expression decreased in the 20% CO2-treated fruit, while it continued to increase in the 15% treated and non-treated fruit, with the latter showing the highest accumulation. In Ochlockonee blueberries, expression increased in all the fruit analysed from day 3, with higher levels in 15% CO2-treated fruit at day 14 and in 20% CO2-treated fruit at day 29.
Regarding PL8 expression, it increased only in non-treated Duke fruit up to day 14, whereas in CO2-treated fruit expression increased only at the end of storage. The expression pattern was different in Ochlockonee fruit, where a decrease in expression was observed in fruit treated with 15% of CO2 compared to freshly harvested blueberries. However, there was no change in non-treated and 20% CO2-treated fruit, except on day 14 when a decrease was also observed.
In the case of PG, storage of non-treated Duke fruit at low temperature reduced its expression compared to freshly harvested blueberries. The application of high levels of CO2 resulted in a different pattern of expression at the end of the treatment, depending on the percentage applied. While 15% CO2 did not affect PG expression, the application of 20% increased the levels. However, in both cases, when the fruit were transferred to atmospheric conditions, a reduction or no change was observed compared to non-treated fruit. On the other hand, storage of non-treated Ochlockonee blueberries at low temperature temporarily increased PG accumulation. In this cultivar, the application of 15% CO2 did not modify the expression of PG until day 29, when a decrease was observed. However, short-term treatment with 20% CO2 significantly increased PG expression at day 3, similar to Duke fruit, with levels higher than those of non-treated fruit. Despite this increase, exposure of the 20% CO2-treated Ochlockonee fruit to atmospheric conditions reduced PG accumulation similarly to the 15% CO2-treated fruit.
The expression of PME3 showed an increase in CO2-treated Duke blueberries at the end (3 days) of both treatments, while no changes were observed on this day in non-treated fruit compared to freshly harvested one. However, from day 14 to the end of storage, the accumulation decreased sharply in all samples. In contrast, no changes in PME accumulation were observed in treated and non-treated Ochlockonee blueberries until day 29, when expression increased similarly in all samples.
Both cultivars showed a very similar expression pattern for EXP4. Notably, after 3 days, the expression was higher in fruit treated with 20% CO2 compared to the other samples, either because it increased (Duke) or remained the same (Ochlockonee). There was also a drastic decrease in EXP4 observed in all samples from day 14 to the end of storage.
Finally, GH5 also showed a similar expression in both cultivars. On day 3, no changes were observed in all samples analysed for both cultivars. However, on day 14, a strong increase in gene expression was observed in non-treated samples of both cultivars. Although expression also increased on this day in CO2-treated fruit, it was significantly lower. Additionally, GH5 expression continued to increase on day 29, although in this case, it was very similar in all analysed samples of both cultivars.
3. Discussion
The blueberry fruit undergoes a series of physiological and biochemical changes during postharvest storage at low temperature that lead to a loss of quality, primarily due to decreased firmness and fungal decay. Short-term treatments with 20% CO2 have been shown to be effective in maintaining quality attributes and reducing total decay in both table grapes and strawberries, as well as in maintaining firmness in strawberries [6,12,29]. However, the effect of these treatments on the postharvest quality preservation of blueberries remains unknown. Therefore, in the present study, as a first approximation, two concentrations of CO2 (15% and 20%) were tested and applied for 3 days at 1 ºC, with the aim of assessing their impact on the quality of blueberries stored for up to 29 days at low temperature.
Soluble solids, titratable acidity and pH are indicative parameters of fruit quality and significantly influence sensory characteristics. Differences in these parameters can be observed between cultivars of the two species at harvest, with Duke blueberries exhibiting lower SSC and higher TA than Ocklochonee (Table 1 and Table 2). Similar results have been observed in previous studies; for instance, Rodríguez Madrera et al. [30] indicated that Duke fruit had SSC of 9 and TA of 0.4%, while Ocklochonee fruit had SSC of 13.1 and TA of 0.2%. Similarly, Kim et al. [31] indicated that fully ripened Duke fruit had an SSC of 9.7 while the TA was 1.8. One of the first studies to describe the quality parameters that blueberries should meet to be considered of good quality indicated that they should contain more than 10% SSC, have TA values between 0.3 and 1.3% citric acid, an SSC/TA ratio of 10-33, and a pH between 2.25 and 4.25 [32]. In the case of the fruit analysed in this study, the values found in the freshly harvested fruits were generally within these ranges, although these parameters varied during storage depending on the cultivar and the treatment applied.
Changes in SSC, TA and pH were observed and varied according to cultivar, gaseous treatment applied, and duration of storage at 1 ºC. Low temperature storage is known to affect these quality attributes, generally increasing SSC and pH [10,33,34]. However, no consistent pattern has been reported for TA during low-temperature storage, with some authors reporting an increase [35], a decrease [36] or no change [34]. Our results align with these previous findings, showing that both SSC and pH increased in both cultivars at the end of storage at 1 ºC, while TA decreased in Duke fruit and increased in Ocklochonee fruit. The application of high levels of CO2 influenced these changes, with differences depending on the concentration applied. Notably, the short-term treatment with 20% of CO2 generally maintained levels more similar to those recorded in freshly harvested fruit. Furthermore, as expected, a negative correlation was found between TA and pH values, independent of the storage conditions, although it was only significant in Duke fruit (r = -0.710; p < 0.01). Regarding pH, Saftner et al. [37] examined the instrumental and sensory quality attributes of blueberry fruit from ten highbush cultivars, including Duke, and found significant positive correlations between pH values and both flavor intensity and overall eating quality. They noted that TA did not correlate with tartness scores or other flavor-related quality metrics. In our study, although no panel of trained tasters was used, trials conducted by ICTAN staff showed that the application of high levels of CO2 did not result in any unusual odours or flavours (data not shown). This could be because the pH of CO2-treated blueberries more closely resembles that of freshly harvested fruit, contrasting with the pH increase observed in non-treated blueberries of both cultivars.
Examining the changes in MI, defined as the SSC/TA ratio, which is commonly used as a measure of fruit sweetness and acceptability, it was observed that after 29 days at 1 °C, Duke fruit treated with 20% of CO2 had an MI similar to that of freshly harvested fruit. However, on the same day of sampling, storage in air significantly increased the ratios of SSC and TA, while the application of 15% CO2 decreased it. In the case of Ochlockonee fruit stored for 29 days at 1 °C, the short-term treatment with 15% of CO2 resulted in a significant increase in MI compared to freshly harvested fruit, whereas the application of 20% CO2 or storage under atmospheric conditions decreased it. Changes in the SSC/TA ratio during storage at low temperature or controlled atmosphere (CA) have been shown to be cultivar-dependent. Forney et al. [10] analysed the postharvest characteristics of five highbush blueberry cultivars and observed that after six weeks of storage in air at 1 °C, the MI decreased in the Jersey and Duke cultivars, but increased in Brigitta and Liberty or remained unchanged in Aurora. However, after storage in different CA conditions (10 kPa O2 + 10, 12.5, 15, 17.5, 20 kPa CO2), the SSC/TA ratio decreased in fruit of all cultivars stored in all atmospheres, except in Brigitta and Liberty stored in 15.0 kPa CO2, where it increased. Conversely, this ratio decreased in Lateblue blueberries stored in air or CA (3 kPa O2 + 11 kPa CO2) [35].
The application of short-term gaseous treatments was effective in controlling total decay in cultivars of both species. Similar results were obtained for blueberries stored under CA conditions. Forney et al. [10] indicated that CA storage of five highbush blueberry cultivars in atmospheres of 10 kPa O2 and 10–20 kPa CO2 inhibited fruit decay. However, CA storage of Lateblue blueberries under 3 kPa O2 and 11 kPa CO2 for 60 days was not effective controlling decay in comparison with non-treated fruit [35]. Our results showed differences between the concentrations applied, with the 20% CO2 treatment being the most effective in controlling decay in both cultivars.
Weight loss in blueberry fruit is an important quality factor and is mainly due to transpiration [38]. In this study, weight loss during cold storage was dependent on blueberry cultivar, storage time and the short-term gaseous treatment applied. The application of high levels of CO2 was not effective in controlling weight loss in Duke fruit, which reached similar levels in all samples by day 29. However, in Ocklochonee fruit, the application of the short-term treatment with 20% of CO2 reduced the weight loss after 29 days at 1 ºC compared to fruit treated with 15% CO2 and non-treated ones. Moreover, these differences were maintained after 2 days of shelf life at room temperature (data not shown). Previous studies have shown that short-term treatments of 3 days with 20% of CO2 at low temperature were effective in controlling water loss in strawberries and table grapes ([6,39,40]. In blueberries, it has been shown that CA storage with high CO2 levels reduced weight loss [10,41]. Duke blueberries showed the highest percentage of weight loss, as reported in several studies analysing different cultivars [10,42], which may relate to the observed differences in the efficacy of the gaseous treatments.
Previous research has shown that weight loss is the major cause of firmness changes during postharvest storage in blueberries [43]. Firmness is a critical quality parameter for fresh blueberries, significantly impacting consumer perception of quality and postharvest storage viability [14]. The investigation into mechanical properties uncovered notable distinctions between CO2-treated and non-treated blueberries across both cultivars. The application of both short-term gaseous treatments resulted in a higher maximum skin breaking force at the end of storage for Duke fruit. Although this force was lower than that of freshly harvested fruit, it was significantly higher than that for non-treated blueberries. Additionally, the slope at skin breaking in treated blueberries was greater than in non-treated blueberries, indicating less berry softening during postharvest storage, thereby confirming previous findings [44]. Moreover, the skin breaking distance values of the non-treated blueberries at the end of the storage period were greater than those of the treated blueberries. This observation suggests a lower loss of skin elasticity in the CO2-treated blueberries by the end of the storage period [14]. It has been reported that the distance at which the berry skin breaks, as determined by the penetration test, increases with greater water loss during storage [44]. Conversely, Chiabrando et al. [45] reported a decrease in the total work calculated from the TPA test during cold storage, which aligns with our results in non-treated fruit. In contrast, the application of 15 and 20% CO2 maintained the work until skin break even after 29 days of fruit storage at 1 ºC, similar to freshly harvested fruit. However, the firmness parameters of rabbiteye blueberries showed less significant differences between treated and non-treated fruit at any sampling point. In this sense, the values for maximum skin breaking force and skin breaking work showed no differences among all the analysed samples and the freshly harvested fruit. This suggests that short-term gaseous treatments do not significantly affect firmness in the Ochlockonee cultivar. This could be related to the fact that firmness is a genetically controlled trait, making the genotype of each blueberry a factor influencing firmness [13].
Fruit softening is linked to alterations in the composition and breakdown of the cell wall, primarily due to the disassembly of polysaccharides, which is driven by cell wall-modifying enzymes [46]. Previous studies have elucidated the key role of specific enzymes such as PG, PME and PL in the degradation of the cell wall in blueberries [47,48,49,50]. In this study, we have examined the relative expression of six genes (XTH23, PL8, PG, PME3, EXP4 and GH5) that encode enzymes involved in cell wall modification during the postharvest storage of blueberries. The levels of these transcripts varied depending on the cultivar and the short-term gaseous treatments applied. It is known that XTH plays a key role in cell wall modification by cleaving and rejoining xyloglucan, which is crucial for softening in various fruits [51,52]. For instance, overexpression of FvXTH9 and FvXTH6 in strawberries resulted in significantly softer fruit, requiring less force to penetrate the tissue compared to control fruit [53]. Similarly, overexpression of XTH8 from persimmon in tomato fruit led to reduced firmness [54]. Our results showed that low-temperature storage of Duke blueberries increased the expression of XTH23. In contrast, the application of high levels of CO2, which resulted in firmer fruit compared to non-treated ones, reduced the accumulation of the transcripts compared to non-treated fruit. However, in both treated and non-treated Ochlockonee fruit, where exposure to low temperatures decreased the firmness, XTH23 expression increased across all samples, with no major differences observed between them. Previous research has reported that the application of 18% CO2 to strawberries for 2 days resulted in firmer fruit along with the down-regulation of XTH30 and XTH15 expression compared to non-treated fruit. This suggests that the lower expression of XTH in CO2-treated fruit may be related to the maintenance of critical functions of xyloglucan in cell wall remodeling, which helps maintain turgor pressure [55].
Several studies have shown that the inhibition of pectate lyase genes in strawberries and tomatoes has resulted in an increase in fruit firmness [56,57]. In this context, our results indicated that the expression of PL8 in CO2-treated Duke fruit was significantly repressed until day 14, reaching levels comparable to those found in freshly harvested fruit. In contrast, PL8 expression increased in non-treated Duke fruit, which exhibited lower firmness. Notably, the expression pattern of this gene differed in the Ochlockonee cultivar, where both treated and non-treated fruit displayed more similar values for the analysed mechanical properties.
Fruit softening involves the solubilization and depolymerization of pectin, which is driven by changes in the activities of PG and PME [58]. PME demethylates pectin, converting methyl esters into free carboxylic acids, increasing substrate availability for PG and facilitating fruit softening. Recently, FaPG1 knockout strawberry plants generated using the CRISPR/Cas9 system exhibited a reduced rate of fruit softening, as well as increased susceptibility to Botrytis cinerea and water loss [26]. Furthermore, Wang et al. [59] demonstrated that exogenous ethylene stimulated blueberry softening and induced the expression of VcPG. However, our findings indicate that the expression of both PG and PME does not correlate with the loss of firmness observed in non-treated Duke fruit stored at low temperatures. This observation is supported by the significant induction of PME expression in fruit treated with 15 and 20% CO2 compared to freshly harvested and non-treated fruit. A similar pattern was noted for PG expression in fruit treated with 20% CO2. In the case of Ochlockonee fruit, a different firmness pattern was observed compared to Duke. A transient induction of PG was also observed at the end of the 20% CO2 treatment, while non-treated fruit displayed a higher expression of PG starting from day 14. Nevertheless, the mechanical properties evaluated did not show significant differences between treated and non-treated fruit.
Research indicates that expansins are linked to environmental stress tolerance such as drought, salt or heat, in various plant species [60,61] For instance, Han et al. [62] reported that overexpression of the wheat expansin gene TaEXPB23 enhanced oxidative stress tolerance in transgenic tobacco plants, which correlated with increased activities of cell wall-bound peroxidases. In our study, EXP4 exhibited a similar expression pattern in both blueberry cultivars, with higher expression levels after 3 days in fruit treated with 20% CO2, followed by those treated with 15%. In contrast, the non-treated fruit showed a decrease in EXP4 expression. This trend may indicate that the CO2-treated fruit possess greater tolerance to oxidative stress and the postharvest deterioration typically observed in fruit. Similar results were observed in strawberries treated with high levels of CO2 for two days and subsequently stored at low temperatures for up to 8 days, where firmer fruit exhibited higher EXP4 expression compared to non-treated fruit.
GHs are enzymes that modify plant cell wall polysaccharides and are regulated by both developmental stage and environmental stress. For example, storage of tomatoes at low temperature has been shown to delay ripening and senescence while preventing the induction of a glycoside hydrolase expression observed in fruit stored at room temperature [63]. Similarly, long term cold storage of peaches has been linked to increased expression of GH5 [64]. In our study, both blueberry cultivars exhibited a similar pattern of GH5 expression, with induction observed at day 14 in non-treated blueberries stored at low temperatures. In contrast, this expression was controlled in the treated fruit, regardless of the CO2 percentage applied. As with EXP4 expression, the changes in GH5 appear to be more closely related to the response of the fruit to low temperatures than to the mechanical properties analysed.
The analysis of variables using categorical principal component analysis (CATPCA) showed differences between the two cultivars (Supplementary Figures S1 and S2). In Duke blueberries, two distinct groupings were identified based on storage time. One group included samples from time 0 and 3 days, while the other comprised samples from medium- to long-term cold storage (14-29 days). Notably, the non-treated fruit stored for 29 days did not cluster with the others (Supplementary Figure S1), being the sample with the highest values for the MI, displacement at skin break, total decay and XTH23 gene expression. Thus, the positive impact of the 3-day gaseous treatment on the maintenance of fruit quality was evident, as these samples, treated and stored for 29 days at low temperatures, clustered with both treated and non-treated fruit from day 14. For the Ochlockonee cultivar, the CATPCA results showed groupings based on storage duration, with the samples treated with 15% of CO2 and stored for 29 days at 1 ºC being the only ones not grouped (Supplementary Figure S1). These samples also exhibited the highest values for MI and displacement at skin breakage. A significant positive correlation was also observed between changes in XTH23 expression and displacement at skin breakage values, mirroring findings in the Duke cultivar.
4. Materials and Methods
4.1. Plant Material and Postharvest Treatments
Organic highbush blueberries (Vaccinium corymbosum L. cv. Duke) and rabbiteye blueberries (Vaccinium ashei Reade cv. Ochlockonee) were harvested in June and September 2022, respectively, in Salas, Asturias, Spain (latitude: 43° 24' 35'' N, longitude: 6° 15' 38'' W, altitude: 243 m). Uniform, disease-free blueberries of both varieties, at commercial maturity, were hand-picked into 125-g solid board eco-friendly packaging punnets (Smurfit Kappa Polska) including a lid with a NatureFlex™ cellulose transparent film window (Futamura Chemical Co. Ltd, Nagoya, Japan). The dimensions of the punnets were 145 x 85 x 50 mm, and the weight was 16 g. Fruit were transported to ICTAN-CSIC (Madrid, Spain) on the same day of collection, and punnets were randomly divided in three batches, each containing 21 punnets, and stored in 1 m3 methacrylate cabins at 1 ± 0.5 °C with a relative humidity of 95%. Initially, 3 random punnets were separated that constitute the group of "time 0". One batch was stored under normal atmospheric conditions (non-treated fruit) for a total of 29 days. A second and third batch were stored with a gas mixture containing 15 kPa CO2 + 20 kPa O2 + 60 kPa N2 (15% CO2-treated fruit) or 20 kPa CO2 + 20 kPa O2 + 60 kPa N2 (20% CO2-treated fruit), respectively, during 3 days at 1 ºC and then transferred to air under the same conditions as the non-treated bunches for 26 days. Fruits from three punnets were sampled at different time points (0, 3, 8, 14, 21 and 29 days), frozen in liquid nitrogen, ground to a fine powder, and stored at -80 °C until further analysis.
4.2. Quality Assessments
Blueberry juice was extracted using a blender and analysed for SSC, TA and pH. SSC was determined using a digital and temperature-compensated refractometer Atago PR-101 (Atago, Co, Ltd., Japan). TA was determined by titration with 0.1N NaOH to pH 8.2 using the 862 Compact Titrosampler (Metrohm, Spain) and the results were expressed as % citric acid. The pH of the juice was measured on a pH meter with a glass electrode (micropH 2000, Crison, Spain).
Weight loss was determined by calculating the percentage of the fresh weight difference between the initial and final weights of the punnets on each sampling day, subtracting the weight of the punnet (16 g). The weight of the decayed blueberries was calculated by subtracting the weight of the healthy fruit from the total weight in each biological replicate. Total decay was expressed as the percentage of the decayed fruit with respect to the original weight.
4.3. Mechanical Properties.
Mechanical properties of blueberries were assessed using a TA.HDPlus texturometer (Stable Micro Systems, Ltd., Godalming, UK) fitted with a 5 kg load cell and operated via Texture Exponent Software (version 6.1.20.0). Each test involved placing a blueberry on a flat metal plate with its stem-root axis aligned parallel to the surface. A flat cylindrical stainless-steel probe (2 mm diameter, P/2) was used to penetrate the fruit at its equator to a depth of 50% of the equatorial diameter. The test speed was maintained at 1 mm/s, with a trigger force set at 0.020 N (2 g). Data was collected at a rate of 500 points per second. From the resulting force-distance curves, measurements were taken for maximum skin breaking force (N), the slope at the breaking point (N/mm), the displacement needed to break the skin (mm), and the work required for skin rupture (mJ), following the methodology described by Sanchez-Ballesta et al. [19]. Mechanical parameter values represent the average of 15 berries for Duke and 20 berries for Ochlockonee cultivars, based on availability.
4.4. Relative Gene Expression by Quantitative Real-Time RT-PCR (RT-qPCR)
RNA was extracted from three biological replicates of blueberry powder (0.4 g) following the protocol outlined by Yu et al. [65], with some modifications. cDNA synthesis was performed by reverse transcribing 0.7 μg of total RNA using the Maxima cDNA Kit with a dsDNase kit (Thermo Fisher Scientific, Waltham, USA), adhering to the manufacturer’s guidelines. Transcript levels were measured via RT-qPCR on an iCycler iQ thermal cycler (Bio-Rad), with quantification conducted using Real Time Detection System Software (version 2.0). The amplification reactions were prepared in a total volume of 12 μL, consisting of 6 μL of NZY qPCR Green Master Mix (2x) (NZYTech, Ltd), 1 μL of each primer (10 μM), and 1 μL of cDNA. The PCR profile included an initial denaturation at 95 °C for 10 minutes, followed by 40 amplification cycles comprising a 20-second denaturation at 95 °C and a 30-second annealing step at either 55 or 60 °C. Each biological replicate was analysed with two technical replicates per gene. The Blueberry Genome Database (Genome Data Base for Vaccinium, GDV; vaccinium.org, Vaccinium corymbosum cv. Draper v1.0 genome), provided the gene sequences for expression analysis quantified using the 2−ΔΔCT method, with EF1 serving as the housekeeping gene. Gene-specific primers for RT-qPCR (Supplementary Table 1) were designed using Primer 3 software [66]
4.5. Statistical Analysis
All descriptive analyses were conducted using IBM-SPSS Statistics software, specifically version 28.0.0 (IBM Corp. Armonk, NY, USA). The data were analysed using one-way analysis of variance (ANOVA), followed by Tukey´s post-hoc test (p < 0.05). SPSS can conduct principal component analysis using both quantitative (scalar) and qualitative (categorical) data, including CATPCA for categorical variables. In this approach, qualitative attributes, mechanical parameters and gene expression are considered unordered variables (multiple nominal), with each category represented by its corresponding number of states. All analyses were based on correlation matrices, and two dimensions were extracted to create scatterplots.
5. Conclusions
The efficacy of the gaseous treatments on the analysed blueberries depended on the species. Thus, while the application of high levels of CO2 (15% and 20%) at low temperature was effective in reducing decay in both cultivars, improvements in firmness were only observed in the highbush blueberry cv. Duke. In this cultivar, CO2 treatments modulated the expression of XTH23, PL8 and GH5, genes related to cell wall integrity. In contrast, the roles of PG and PME in maintaining firmness appeared minimal, with no significant differences between the two gaseous treatments. Although the application of high levels of CO2 did not affect the firmness of the rabbiteye blueberry cv. Ochlocknee, it effectively controlled fruit weight loss. Notably, a strong relationship was found between XTH23 expression, the skin-breaking displacement parameter, and maturation index values, regardless of the fruit´s postharvest response. These differences in response between species highlight the necessity of tailoring postharvest treatments to specific blueberry species/cultivars. Overall, the findings suggest that the application of short-term high CO2 treatments may be a viable strategy to prolong the postharvest life of highbush blueberries, thereby enhancing their quality.
Author Contributions
Conceptualization, M.T.S.-B. and I.R.; Methodology, J.D.T.-G., M.D.Á., B.H., M.I.E. C.M., I.R. and M.T.S.-B; Formal Analysis, J.D.T.-G., M.D.Á., I.R., and M.T.S.-B., Writing-Original Draft Preparation, J.D.T.-G., M.D.Á., B.H., I.R. and M.T.S.-B.; Writing-Review and Editing, I.R. and M.T.S.-B.; Visualization, M.I.E., C.M., I.R. and M.T.S.-B; Funding Acquisition, M.I.E., C.M., I.R. and M.T.S.-B; All authors have read and agreed to the published version of the manuscript.
Funding
This work is part of Project PID2020-113965RB-I00 funded by MICIU/AEI/10.13039/501100011033. J. D. Toledo-Guerrero is grateful to the Research Personnel Training Subprogram (FPI) of the Spanish Ministry of Science, Innovation and Universities (MICIU) (PRE2021-100846).
Data Availability Statement
Data is contained within the article or Supplementary Material.
Acknowledgments
We are grateful to the Ministry of Science and Innovation for the funding support. We thank “Arándanos La Peña S.L.” for the help we received in the harvesting of the fruit of this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tănase, E.E.; Popa, V.I.; Popa, M.E.; Geicu-Cristea, M.; Popescu, P.; Drăghici, M.; Miteluț, A.C. Identification of the most relevant quality parameters for berries-a review; Scientific Bulletin. Series F. Biotechnologies 2016, Vol XX, 222–233. [Google Scholar]
- Zorzi, M.; Gai, F.; Medana, C.; Aigotti, R.; Morello, S.; Peiretti, P.G. Bioactive compounds and antioxidant capacity of small berries. Foods 2020, 9, 623. [Google Scholar] [CrossRef] [PubMed]
- Horvitz, S. Postharvest Handling of Berries. In Postharvest Handling; Kahramanoglu, I., Ed.; IntechOpen, Ltd.: London, UK, 2017. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, C.; Cheng, S.; Wei, B.; Liu, X.; Ji, S. Changes in energy metabolism accompanying pitting in blueberries stored at low temperature. Food Chem. 2014, 164, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Ryu, J.A.; Liu, R.H.; Nock, J.F.; Watkins, C.B. Harvest maturity, storage temperature and relative humidity affect fruit quality, antioxidant contents and activity, and inhibition of cell proliferation of strawberry fruit. Postharvest Biol. Technol. 2008, 49, 201–209. [Google Scholar] [CrossRef]
- Blanch, M.; Sanchez-Ballesta, M.T.; Escribano, M.I.; Merodio, C. Water distribution and ionic balance in response to high CO2 treatments in strawberries (Fragaria vesca L. cv. Mara de Bois). Postharvest Biol. Technol. 2012, 73, 63–71. [Google Scholar] [CrossRef]
- Lafarga, T.; Colás-Medà, P.; Abadías, M.; Aguiló-Aguayo, I.; Bobo, G.; Viñas, I. Strategies to reduce microbial risk and improve quality of fresh and processed strawberries: A review. Innov. Food Sci. Emerg. Technol. 2019, 52, 197–212. [Google Scholar] [CrossRef]
- Liu, G.S.; Fu, D.Q.; Zhong, C.F.; Zong, J.; Zhou, J.H.; Chang, H.; Wang, B.G.; Wang, Y.X. Transcriptome combined with long non-coding RNA analysis reveals the potential molecular mechanism of high-CO2 treatment in delaying postharvest strawberry fruit ripening and senescence. Sci. Hortic. (Amsterdam). 2024, 323, 112505. [Google Scholar] [CrossRef]
- Falagán, N.; Miclo, T.; Terry, L.A. Graduated controlled atmosphere: A novel approach to increase “Duke” blueberry storage life. Front. Plant Sci. 2020, 11, 221. [Google Scholar] [CrossRef]
- Forney, C.F.; Jordan, M.A.; Pennell, K.M.; Fillmore, S. Controlled atmosphere storage impacts fruit quality and flavor chemistry of five cultivars of highbush blueberry (Vaccinium corymbosum). Postharvest Biol. Technol. 2022, 194, 112073. [Google Scholar] [CrossRef]
- Smrke, T.; Cvelbar Weber, N.; Razinger, J.; Medic, A.; Veberic, R.; Hudina, M.; Jakopic, J. Short-term storage in a modified atmosphere affects the chemical profile of blueberry (Vaccinium corymbosum L.) fruit. Horticulturae 2024, 10, 194. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Alvarez, I.; Escribano, M.I.; Merodio, C.; Romero, I. Effect of high CO2 levels and low temperature on stilbene biosynthesis pathway gene expression and stilbenes production in white, red and black table grape cultivars during postharvest storage. Plant Physiol. Biochem. 2020, 151, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Cappai, F.; Amadeu, R.R.; Benevenuto, J.; Cullen, R.; Garcia, A.; Grossman, A.; Ferrão, L.F. V.; Munoz, P. High-resolution linkage Map and QTL analyses of fruit firmness in autotetraploid blueberry. Front. Plant Sci. 2020, 11, 562171. [Google Scholar] [CrossRef] [PubMed]
- Rivera, S.; Giongo, L.; Cappai, F.; Kerckhoffs, H.; Sofkova-Bobcheva, S.; Hutchins, D.; East, A. Blueberry firmness - A review of the textural and mechanical properties used in quality evaluations. Postharvest Biol. Technol. 2022, 192, 112016. [Google Scholar] [CrossRef]
- Giongo, L.; Ajelli, M.; Pottorff, M.; Perkins-Veazie, P.; Iorizzo, M. Comparative multi-parameters approach to dissect texture subcomponents of highbush blueberry cultivars at harvest and postharvest. Postharvest Biol. Technol. 2022, 183, 111696. [Google Scholar] [CrossRef]
- Forney, C.F.; Jordan, M.A.; Nicholas, K.U.K.G. Effect of CO2 on physical, chemical, and quality changes in “Burlington” blueberries. In Proceedings of the Acta Horticulturae; 2003; 600. 587–593. [Google Scholar] [CrossRef]
- Giongo, L.; Poncetta, P.; Loretti, P.; Costa, F. Texture profiling of blueberries (Vaccinium spp.) during fruit development, ripening and storage. Postharvest Biol. Technol. 2013, 76, 34–39. [Google Scholar] [CrossRef]
- Rivera, S.; Kerckhoffs, H.; Sofkova-Bobcheva, S.; Hutchins, D.; East, A. Influence of harvest maturity and storage technology on mechanical properties of blueberries. Postharvest Biol. Technol. 2022, 191, 111961. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Marti-Anders, C.; Álvarez, M.D.; Escribano, M.I.; Merodio, C.; Romero, I. Are the blueberries we buy good quality? Comparative study of berries purchased from different outlets. Foods 2023, 12, 2621. [Google Scholar] [CrossRef]
- Olmedo, P.; Zepeda, B.; Rojas, B.; Silva-Sanzana, C.; Delgado-Rioseco, J.; Fernández, K.; Balic, I.; Arriagada, C.; Moreno, A.A.; Defilippi, B.G.; et al. Cell wall calcium and hemicellulose have a role in the fruit firmness during storage of blueberry (Vaccinium spp.). Plants 2021, 10, 1–13. [Google Scholar] [CrossRef]
- Wei, J.; Ma, F.; Shi, S.; Qi, X.; Zhu, X.; Yuan, J. Changes and postharvest regulation of activity and gene expression of enzymes related to cell wall degradation in ripening apple fruit. Postharvest Biol. Technol. 2010, 56, 147–154. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, Y.; Lin, H.; Lin, M.; Li, H.; Yuan, F.; Chen, Y.; Xiao, J. Effects of paper containing 1-MCP postharvest treatment on the disassembly of cell wall polysaccharides and softening in Younai plum fruit during storage. Food Chem. 2018, 264, 1–8. [Google Scholar] [CrossRef]
- Cappai, F.; Benevenuto, J.; Ferrão, L.F. V.; Munoz, P. Molecular and genetic bases of fruit firmness variation in blueberry—a review. Agronomy 2018, 8, 174. [Google Scholar] [CrossRef]
- Atkinson, R.G.; Sutherland, P.W.; Johnston, S.L.; Gunaseelan, K.; Hallett, I.C.; Mitra, D.; Brummell, D.A.; Schröder, R.; Johnston, J.W.; Schaffer, R.J. Down-regulation of POLYGALACTURONASE1 alters firmness, tensile strength and water loss in apple (Malus x domestica) fruit. BMC Plant Biol. 2012, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, C.; Blanco-Portales, R.; Barceló-Muñoz, M.; García-Gago, J.A.; Waldron, K.W.; Quesada, M.A.; Muñoz-Blanco, J.; Mercado, J.A. Antisense down-regulation of the strawberry β-galactosidase gene FaβGal4 increases cell wall galactose levels and reduces fruit softening. J. Exp. Bot. 2016, 67, 619–631. [Google Scholar] [CrossRef] [PubMed]
- López-Casado, G.; Sánchez-Raya, C.; Ric-Varas, P.D.; Paniagua, C.; Blanco-Portales, R.; Muñoz-Blanco, J.; Pose, S.; Matas, A.J.; Mercado, J.A. CRISPR/Cas9 editing of the polygalacturonase FaPG1 gene improves strawberry fruit firmness. Hortic. Res. 2023, 10, uhad011. [Google Scholar] [CrossRef]
- Song, X.; Dai, H.; Wang, S.; Ji, S.; Zhou, X.; Li, J.; Zhou, Q. Putrescine treatment delayed the softening of postharvest blueberry fruit by inhibiting the expression of cell wall metabolism key gene VcPG1. Plants 2022, 11, 1356. [Google Scholar] [CrossRef]
- Ji, Y.; Hu, W.; Xiu, Z.; Yang, X.; Guan, Y. Integrated transcriptomics-proteomics analysis reveals the regulatory network of ethanol vapor on softening of postharvest blueberry. LWT 2023, 180, 114649. [Google Scholar] [CrossRef]
- Romero, I.; Sanchez-Ballesta, M.T.; Maldonado, R.; Escribano, M.I.; Merodio, C. Expression of class I chitinase and β-1,3-glucanase genes and postharvest fungal decay control of table grapes by high CO2 pretreatment. Postharvest Biol. Technol. 2006, 41, 9–15. [Google Scholar] [CrossRef]
- Rodríguez Madrera, R.; Suárez Valles, B.; Campa Negrillo, A.; Ferreira Fernández, J.J. Physicochemical characterization of blueberry (Vaccinium spp.) juices from 55 cultivars grown in northern Spain. Acta Aliment. 2019, 48, 260–268. [Google Scholar] [CrossRef]
- Kim, J.G.; Kim, H.L.; Kim, S.J.; Park, K.S. Fruit quality, anthocyanin and total phenolic contents, and antioxidant activities of 45 blueberry cultivars grown in Suwon, Korea. J. Zhejiang Univ. Sci. B 2013, 14, 793–799. [Google Scholar] [CrossRef]
- Beaudry, R. Blueberry quality characteristics and how they can be optimized. In Annual Report of the Michigan State Horticultural Society, 122nd ed.; 1992; pp. 140–145. Available online: https://www.researchgate. 2857. [Google Scholar]
- Almenar, E.; Samsudin, H.; Auras, R.; Harte, B.; Rubino, M. Postharvest shelf life extension of blueberries using a biodegradable package. Food Chem. 2008, 110, 120–127. [Google Scholar] [CrossRef]
- Ji, Y.; Hu, W.; Jiang, A.; Xiu, Z.; Liao, J.; Yang, X.; Guan, Y.; Saren, G.; Feng, K. Effect of ethanol treatment on the quality and volatiles production of blueberries after harvest. J. Sci. Food Agric. 2019, 99, 6296–6306. [Google Scholar] [CrossRef] [PubMed]
- Chiabrando, V.; Giacalone, G. Shelf-life extension of highbush blueberry using 1-methylcyclopropene stored under air and controlled atmosphere. Food Chem. 2011, 126, 1812–1816. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.; Guerra, M.; Daniel, P.; Camelo, A.L.; Yommi, A. Quality changes of highbush blueberries fruit stored in CA with different CO2 levels. J. Food Sci. 2009, 74, S154–9. [Google Scholar] [CrossRef]
- Saftner, R.; Polashock, J.; Ehlenfeldt, M.; Vinyard, B. Instrumental and sensory quality characteristics of blueberry fruit from twelve cultivars. Postharvest Biol. Technol. 2008, 49, 19–26. [Google Scholar] [CrossRef]
- Moggia, C.; Graell, J.; Lara, I.; González, G.; Lobos, G.A. Firmness at harvest impacts postharvest fruit softening and internal browning development in mechanically damaged and non-damaged highbush blueberries (Vaccinium corymbosum L.). Front. Plant Sci. 2017, 8, 535. [Google Scholar] [CrossRef]
- Rosales, R.; Romero, I.; Fernandez-Caballero, C.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Low temperature and short-term high-CO2 treatment in postharvest storage of table grapes at two maturity stages: Effects on transcriptome profiling. Front. Plant Sci. 2016, 7, 1020. [Google Scholar] [CrossRef]
- Romero, I.; Domínguez, I.; Doménech-Carbó, A.; Gavara, R.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Effect of high levels of CO2 on the electrochemical behavior and the enzymatic and non-enzymatic antioxidant systems in black and white table grapes stored at 0 °C. J. Sci. Food Agric. 2019, 99, 6859–6867. [Google Scholar] [CrossRef] [PubMed]
- Schotsmans, W.; Molan, A.; MacKay, B. Controlled atmosphere storage of rabbiteye blueberries enhances postharvest quality aspects. Postharvest Biol. Technol. 2007, 44, 277–285. [Google Scholar] [CrossRef]
- Moggia, C.; Graell, J.; Lara, I.; Schmeda-Hirschmann, G.; Thomas-Valdés, S.; Lobos, G.A. Fruit characteristics and cuticle triterpenes as related to postharvest quality of highbush blueberries. Sci. Hortic. (Amsterdam). 2016, 211, 449–457. [Google Scholar] [CrossRef]
- Paniagua, A.C.; East, A.R.; Heyes, J.A. Interaction of temperature control deficiencies and atmosphere conditions during blueberry storage on quality outcomes. Postharvest Biol. Technol. 2014, 95, 50–59. [Google Scholar] [CrossRef]
- Rivera, S.; Kerckhoffs, H.; Sofkova-Bobcheva, S.; Hutchins, D.; East, A. Influence of water loss on mechanical properties of stored blueberries. Postharvest Biol. Technol. 2021, 176, 111498. [Google Scholar] [CrossRef]
- Chiabrando, V.; Giacalone, G.; Rolle, L. Mechanical behaviour and quality traits of highbush blueberry during postharvest storage. J. Sci. Food Agric. 2009, 89, 989–992. [Google Scholar] [CrossRef]
- Brummell, D.A.; Harpster, M.H. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol. Biol. 2001, 47, 311–339. [Google Scholar] [CrossRef] [PubMed]
- Chea, S.; Yu, D.J.; Park, J.; Oh, H.D.; Chung, S.W.; Lee, H.J. Fruit softening correlates with enzymatic and compositional changes in fruit cell wall during ripening in ‘Bluecrop’ highbush blueberries. Sci. Hortic. (Amsterdam). 2019, 245, 163–170. [Google Scholar] [CrossRef]
- Posé, S.; Paniagua, C.; Matas, A.J.; Gunning, A.P.; Morris, V.J.; Quesada, M.A.; Mercado, J.A. A nanostructural view of the cell wall disassembly process during fruit ripening and postharvest storage by atomic force microscopy. Trends Food Sci. Technol. 2019, 87, 47–58. [Google Scholar] [CrossRef]
- Sun, H.; Hao, D.; Tian, Y.; Huang, Y.; Wang, Y.; Qin, G.; Pei, J.; Abd El-Aty, A.M. Effect of chitosan/thyme oil coating and UV-C on the softening and ripening of postharvest blueberry fruits. Foods 2022, 11, 2795. [Google Scholar] [CrossRef]
- Liu, R.; Shang, F.; Niu, B.; Wu, W.; Han, Y.; Chen, H.; Gao, H. Melatonin treatment delays the softening of blueberry fruit by modulating cuticular wax metabolism and reducing cell wall degradation. Food Res. Int. 2023, 173, 113357. [Google Scholar] [CrossRef] [PubMed]
- Miedes, E.; Herbers, K.; Sonnewald, U.; Lorences, E.P. Overexpression of a cell wall enzyme reduces xyloglucan depolymerization and softening of transgenic tomato fruits. J. Agric. Food Chem. 2010, 58, 5708–5713. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bertomeu, J.; Miedes, E.; Lorences, E.P. Expression of xyloglucan endotransglucosylase/hydrolase (XTH) genes and XET activity in ethylene treated apple and tomato fruits. J. Plant Physiol. 2013, 170, 1194–1201. [Google Scholar] [CrossRef]
- Witasari, L.D.; Huang, F.C.; Hoffmann, T.; Rozhon, W.; Fry, S.C.; Schwab, W. Higher expression of the strawberry xyloglucan endotransglucosylase/hydrolase genes FvXTH9 and FvXTH6 accelerates fruit ripening. Plant J. 2019, 100, 1237–1253. [Google Scholar] [CrossRef]
- Han, Y.; Ban, Q.; Li, H.; Hou, Y.; Jin, M.; Han, S.; Rao, J. DkXTH8, a novel xyloglucan endotransglucosylase/hydrolase in persimmon, alters cell wall structure and promotes leaf senescence and fruit postharvest softening. Sci. Rep. 2016, 6, 39155. [Google Scholar] [CrossRef] [PubMed]
- del Olmo, I.; Romero, I.; Alvarez, M.D.; Tarradas, R.; Sanchez-Ballesta, M.T.; Escribano, M.I.; Merodio, C. Transcriptomic analysis of CO2-treated strawberries (Fragaria vesca) with enhanced resistance to softening and oxidative stress at consumption. Front. Plant Sci. 2022, 13, 983976. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Doménech, N.; Jiménez-Bemúdez, S.; Matas, A.J.; Rose, J.K.C.; Muñoz-Blanco, J.; Mercado, J.A.; Quesada, M.A. Antisense inhibition of a pectate lyase gene supports a role for pectin depolymerization in strawberry fruit softening. J. Exp. Bot. 2008, 59, 2769–2779. [Google Scholar] [CrossRef]
- Ren, H.; Zhao, Q.; Feng, Y.; Tang, P.; Wang, Y.; Jiang, J.; Hu, C.; Wang, Y.; Cui, B.; Xie, X.; et al. Gene-specific silencing of SlPL16, a pectate lyase coding gene, extends the shelf life of tomato fruit. Postharvest Biol. Technol. 2023, 201, 112368. [Google Scholar] [CrossRef]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit Softening: Revisiting the Role of Pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Q.; Zhou, X.; Zhang, F.; Ji, S. Ethylene plays an important role in the softening and sucrose metabolism of blueberries postharvest. Food Chem. 2020, 310, 125965. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.; Shi, Y.; Xu, J.; Huang, B. Transgenic tobacco plants overexpressing a grass PpEXP1 gene exhibit enhanced tolerance to heat stress. PLoS One 2014, 9, 100792. [Google Scholar] [CrossRef]
- Chen, Y.; Ren, Y.; Zhang, G.; An, J.; Yang, J.; Wang, Y.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improves oxidative stress tolerance in transgenic Arabidopsis plants. Plant Physiol. Biochem. 2018, 124, 190–198. [Google Scholar] [CrossRef]
- Han, Y.; Chen, Y.; Yin, S.; Zhang, M.; Wang, W. Over-expression of TaEXPB23, a wheat expansin gene, improves oxidative stress tolerance in transgenic tobacco plants. J. Plant Physiol. 2015, 173, 62–71. [Google Scholar] [CrossRef]
- Bai, C.; Wu, C.; Ma, L.; Fu, A.; Zheng, Y.; Han, J.; Li, C.; Yuan, S.; Zheng, S.; Gao, L.; et al. Transcriptomics and metabolomics analyses provide insights into postharvest ripening and senescence of tomato fruit under low temperature. Hortic. Plant J. 2023, 9, 109–121. [Google Scholar] [CrossRef]
- Tittarelli, A.; Santiago, M.; Morales, A.; Meisel, L.A.; Silva, H. Isolation and functional characterization of cold-regulated promoters, by digitally identifying peach fruit cold-induced genes from a large EST dataset. BMC Plant Biol. 2009, 9, 121. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Tang, H.; Zhang, Y.; Du, Z.; Yu, H.; Chen, Q. Comparison and Improvement of Different Methods of RNA Isolation from Strawberry (Fragria * ananassa). J. Agric. Sci. 2012, 4, 51–56. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 115. [Google Scholar] [CrossRef]
Figure 1.
Weight loss of non-treated, 15% and 20% CO2-treated Duke and Ochlockonee blueberries stored at 1 °C for 29 days. Values are the means of three replicate samples ± standard error. Different letters within each column indicate that the means are statistically different according to the Tukey-b test (p < 0.05).
Figure 1.
Weight loss of non-treated, 15% and 20% CO2-treated Duke and Ochlockonee blueberries stored at 1 °C for 29 days. Values are the means of three replicate samples ± standard error. Different letters within each column indicate that the means are statistically different according to the Tukey-b test (p < 0.05).
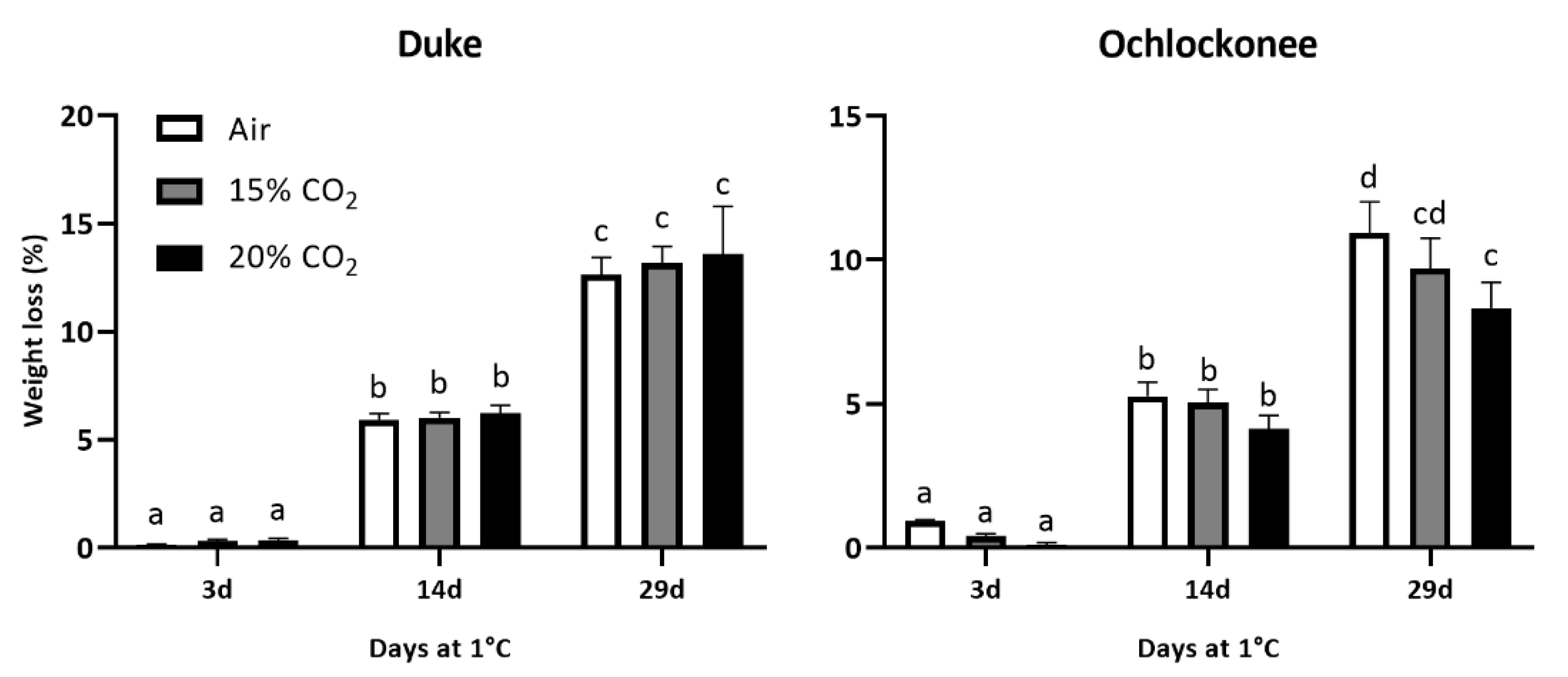
Figure 2.
Examples of penetration force-distance curves derived from the penetration test for Duke blueberries at the time of harvest and for non-treated and 20% CO2-treated fruit after 29 days of storage at low temperature.
Figure 2.
Examples of penetration force-distance curves derived from the penetration test for Duke blueberries at the time of harvest and for non-treated and 20% CO2-treated fruit after 29 days of storage at low temperature.
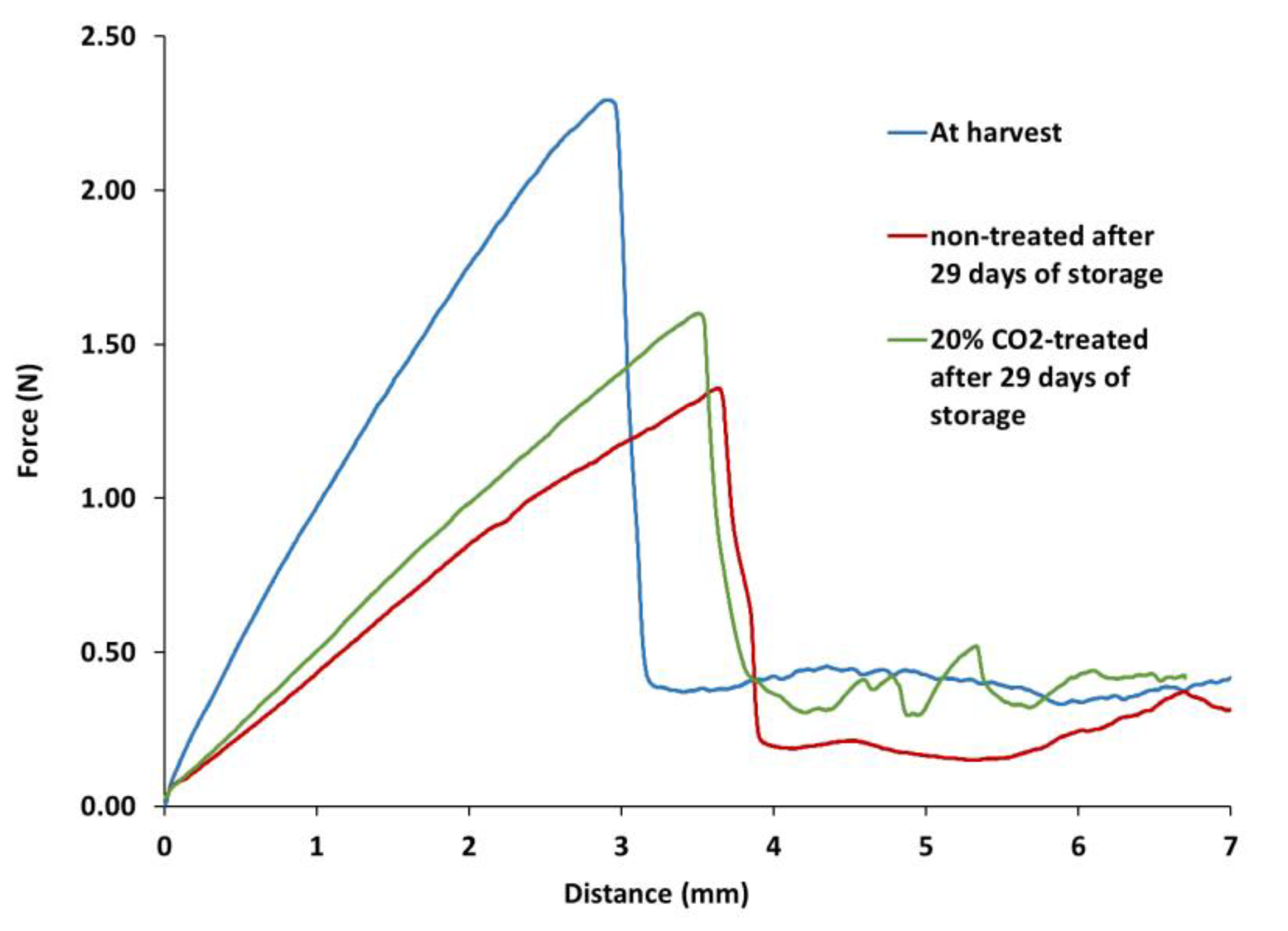
Figure 3.
Effect of low temperature and a 3-day treatment with 15% or 20% of CO2 on the expression of cell wall-related genes in Duke and Ochlockonee blueberries stored at 1 ºC for 29 days. Transcript levels were assessed by RT-qPCR and normalized using EF1 as a reference gene. Results were calculated relative to a calibrator sample (time 0) using the 2−ΔΔCt method. Values are the mean ± SD, n = 6. Different letters on bars indicate significant differences using the Tukey-b test (p < 0.05).
Figure 3.
Effect of low temperature and a 3-day treatment with 15% or 20% of CO2 on the expression of cell wall-related genes in Duke and Ochlockonee blueberries stored at 1 ºC for 29 days. Transcript levels were assessed by RT-qPCR and normalized using EF1 as a reference gene. Results were calculated relative to a calibrator sample (time 0) using the 2−ΔΔCt method. Values are the mean ± SD, n = 6. Different letters on bars indicate significant differences using the Tukey-b test (p < 0.05).
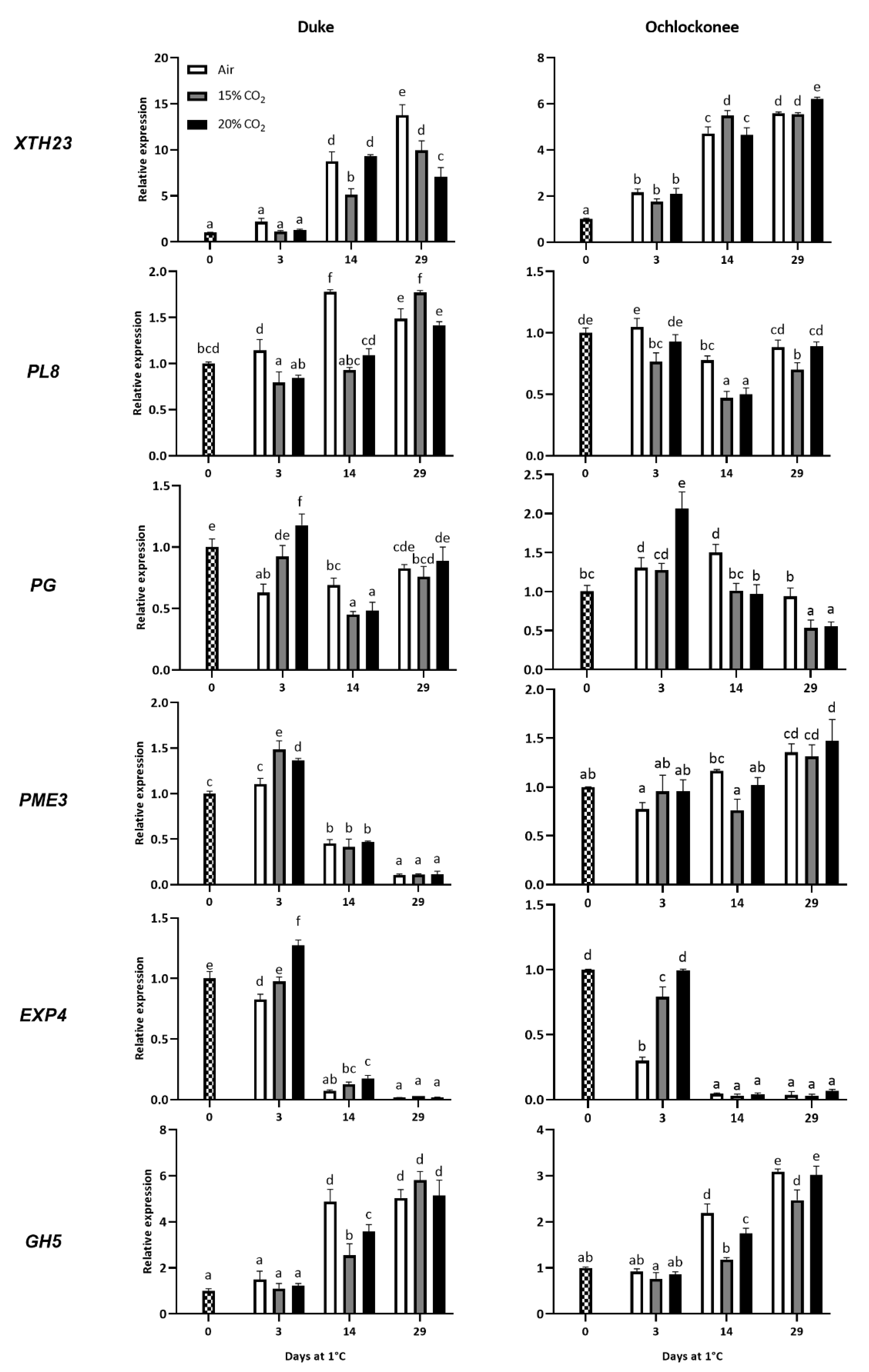

Table 1.
Soluble solids content (SSC), titratable acidity (TA), SSC/TA ratio (MI), pH and decay index of non-treated and 15% and 20% CO2-treated Duke blueberries stored at 1 °C for 29 days.
Table 1.
Soluble solids content (SSC), titratable acidity (TA), SSC/TA ratio (MI), pH and decay index of non-treated and 15% and 20% CO2-treated Duke blueberries stored at 1 °C for 29 days.
| Duke | ||||||
|---|---|---|---|---|---|---|
| SSC (⁰Brix) |
TA (% citric acid) |
MI (SSC/TA) |
pH | Decay index (%) |
||
| At harvest | 9.17 ± 0.38 b | 0.87 ± 0.01 b | 10.58 ± 0.51 c | 2.82 ± 0.01 a | 0000 ̶ | |
| Air | 3 d | 8.20 ± 0.17 a | 1.03 ± 0.02 e | 07.96 ± 0.26 a | 2.96 ± 0.06 bc | 0000 ̶ |
| 29 d | 9.60 ± 0.00 b | 0.66 ± 0.01 a | 14.47 ± 0.13 d | 3.24 ± 0.02 d | 5.70 ± 0.04 c | |
| 15% CO2 | 3 d | 8.37 ± 0.32 a | 1.06 ± 0.02 f | 07.92 ± 0.39 a | 2.89 ± 0.04 ab | 0000 ̶ |
| 29 d | 8.30 ± 0.10 a | 0.95 ± 0.01 d | 08.71 ± 0.15 ab | 2.95 ± 0.02 bc | 4.43 ± 0.01 b | |
| 20% CO2 | 3 d | 8.37 ± 0.25 a | 0.92 ± 0.01 c | 09.06 ± 0.32 b | 3.01 ± 0.02 c | 0000 ̶ |
| 29 d | 8.43 ± 0.31 a | 0.85 ± 0.00 b | 09.92 ± 0.36 c | 3.01 ± 0.01 c | 2.35 ± 0.02 a | |
Values are the means of three replicate samples ± standard error. Different letters within each column indicate that means are statistically different according to the Tukey-b test (p < 0.05).
Table 2.
Soluble solids content (SSC), titratable acidity (TA), SSC/TA ratio (MI), pH and decay index of non-treated and 15% and 20% CO2-treated Ochlockonee blueberries stored at 1 °C for 29 days.
Table 2.
Soluble solids content (SSC), titratable acidity (TA), SSC/TA ratio (MI), pH and decay index of non-treated and 15% and 20% CO2-treated Ochlockonee blueberries stored at 1 °C for 29 days.
| Ochlockonee | ||||||
|---|---|---|---|---|---|---|
| SSC (⁰Brix) |
TA (% citric acid) |
MI (SSC/TA) |
pH | Decay index (%) |
||
| At harvest | 12.67 ± 0.12 a | 0.34 ± 0.02 b | 37.70 ± 1.95 c | 2.81 ± 0.01 b | 0000 ̶ | |
| Air | 3 d | 12.70 ± 0.00 a | 0.45 ± 0.01 e | 28.44 ± 0.37 a | 2.85 ± 0.01 c | 0000 ̶ |
| 29 d | 13.87 ± 0.12 c | 0.44 ± 0.01 de | 31.53 ± 0.96 ab | 2.87 ± 0.02 cd | 2.03 ± 0.02 c | |
| 15% CO2 | 3 d | 13.30 ± 0.10 b | 0.32 ± 0.01 ab | 41.15 ± 1.00 c | 2.87 ± 0.01 cd | 0000 ̶ |
| 29 d | 13.33 ± 0.06 b | 0.30 ± 0.02 a | 45.03 ± 2.55 d | 2.89 ± 0.01 d | 1.62 ± 0.02 b | |
| 20% CO2 | 3 d | 13.20 ± 0.00 b | 0.39 ± 0.02 c | 34.20 ± 1.80 b | 2.76 ± 0.01 a | 0000 ̶ |
| 29 d | 12.70 ± 0.00 a | 0.41 ± 0.01 cd | 30.99 ± 0.76 ab | 2.79 ± 0.03 ab | 0.39 ± 0.01 a | |
Values are the means of three replicate samples ± standard error. Different letters within each column indicate that means are statistically different according to the Tukey-b test (p < 0.05).
Table 3.
Mechanical properties of the penetration test corresponding to non-treated and 15% and 20% CO2-treated Duke blueberries stored at 1 °C for 29 days.
Table 3.
Mechanical properties of the penetration test corresponding to non-treated and 15% and 20% CO2-treated Duke blueberries stored at 1 °C for 29 days.
| Duke | |||||
|---|---|---|---|---|---|
| Maximum Skin Breaking Force (N) |
Slope at Skin Breaking (N/mm) |
Displacement at Skin Breaking (mm) | Skin Breaking Work (mJ) |
||
| At harvest | 1.95±0.18 cd | 0.60±0.05 d | 3.23±0.28 a | 4.97±0.59 bcd | |
| Air | 3 d | 2.07±0.16 d | 0.68±0.06 e | 3.03±0.28 a | 5.39±0.54 d |
| 14 d | 1.91±0.16 cd | 0.63±0.06 de | 3.02±0.29 a | 5.17±0.49 bcd | |
| 29 d | 1.36±0.13 a | 0.29±0.03 a | 4.75±0.79 c | 4.25±0.63 a | |
| 15% CO2 | 3 d | 1.85±0.17 bc | 0.61±0.05 d | 3.02±0.25 a | 5.21±0.39 cd |
| 14 d | 1.69±0.21 b | 0.57±0.05 cd | 2.92±0.28 a | 4.77±0.47 abc | |
| 29 d | 1.69±0.16 b | 0.41±0.04 b | 4.09±0.40 b | 5.08±0.48 bcd | |
| 20% CO2 | 3 d | 1.68±0.13 b | 0.54±0.05 c | 3.04±0.27 a | 4.61±0.45 ab |
| 14 d | 1.77±0.16 bc | 0.54±0.05 c | 3.22±0.28 a | 4.88±0.46 bcd | |
| 29 d | 1.71±0.18 b | 0.40±0.04 b | 4.26±0.44 b | 5.08±0.52 bcd | |
Values are the means of fifteen replicate samples ± standard error. Different letters within each column indicate that means are statistically different according to the Tukey-b test (p < 0.05).
Table 4.
Mechanical properties of the penetration test corresponding to non-treated and 15% and 20% CO2-treated Ochlockonee blueberries stored at 1 °C for 29 days.
Table 4.
Mechanical properties of the penetration test corresponding to non-treated and 15% and 20% CO2-treated Ochlockonee blueberries stored at 1 °C for 29 days.
| Ochlockonee | |||||
|---|---|---|---|---|---|
| Maximum Skin Breaking Force (N) |
Slope at Skin Breaking (N/mm) |
Displacement at Skin Breaking (mm) | Skin Breaking Work (mJ) |
||
| At harvest | 1.43±0.12 ab | 0.49±0.04 c | 2.91±0.26 a | 3.92±0.40 ab | |
| Air | 3 d | 1.51±0.15 b | 0.56±0.05 d | 2.68±0.28 a | 4.27±0.41 b |
| 14 d | 1.45±0.17 ab | 0.41±0.03 b | 3.51±0.50 b | 3.91±0.52 ab | |
| 29 d | 1.41±0.19 ab | 0.38±0.06 ab | 3.78±0.59 bc | 3.75±0.61 ab | |
| 15% CO2 | 3 d | 1.35±0.13 ab | 0.54±0.05 cd | 2.49±0.24 a | 3.76±0.40 ab |
| 14 d | 1.45±0.19 ab | 0.41±0.06 b | 3.51±0.45 b | 3.76±0.59 ab | |
| 29 d | 1.34±0.23 a | 0.33±0.07 a | 4.15±0.74 c | 3.59±0.71 a | |
| 20% CO2 | 3 d | 1.50±0.15 ab | 0.55±0.05 d | 2.69±0.29 a | 4.23±0.39 b |
| 14 d | 1.39±0.17 ab | 0.39±0.05 ab | 3.56±0.38 b | 3.79±0.59 ab | |
| 29 d | 1.35±0.20 ab | 0.39±0.08 ab | 3.53±0.53 b | 3.61±0.61 a | |
Values are the means of twenty replicate samples ± standard error. Different letters within each column indicate that means are statistically different according to the Tukey-b test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



