
Submitted:
18 October 2024
Posted:
21 October 2024
You are already at the latest version
Abstract
Reactive oxygen species (ROS) are essential molecules involved in intercellular communication, signal transduction and metabolic processes. Abiotic stresses cause accumulation of excess ROS in plant cells. The issues of regulation of antioxidant protection of plants by natural and synthetic compounds with antioxidant activity still remain one of the important and relevant areas of fundamental and applied research. Glutathione (GSH) plays an important role in stress resistance and redox homeostasis of the cell and effectively protects the cell from stress-induced generation of ROS. An increase in the GSH content in plant cells can contribute to an increase in plant resistance to various types of stressors. However, an increase in the GSH content in plants can negatively affect their growth and development. We have shown that growing Nicotiana tabacum in the presence of tetrapeptide AEDL contributes to an increase in the GSH content by 3.24 times. At the same time, the tobacco plant was more developed, especially its root system. A scheme of the mechanism of regulation of the redox balance in the stem cell niche and the participation of the peptide AEDL and GSH in the regulation of the fate of stem cells was proposed.
Keywords:
GSH
; peptide AEDL
; ROS
; stem cells
1. Introduction
Redox metabolism in plant cells inevitably includes the formation and accumulation of highly toxic reactive oxygen species (ROS) [1,2,3]. As a result of unfavorable biotic and abiotic conditions, the level of ROS in plants increases. Conditions leading to damage to cellular organelles and cell membranes caused by ROS are called oxidative stress [4]. ROS accumulation most often occurs near electron transport chains. In plants, the accumulation of excess ROS occurs near the thylakoid membranes of chloroplasts, in which the process of photosynthesis takes place [5] and the internal membranes of mitochondria, which carry out the respiratory process [6]. Constant exposure to oxidative stress is accompanied by the destruction of RNA and DNA, leading to lipid oxidation [1,7]. Plants have developed a complex enzymatic and non-enzymatic antioxidant system (AOS), which maintains the concentration of ROS and prevents their accumulation of ROS [8,9]. These two systems work together to control ROS levels. The presence of a more powerful AOS system may be one of the mechanisms of plant stress resistance to external adverse effects [9].
Glutathione (GSH), a γ-glutamylcysteinylglycine tripeptide, is an important component involved in many cellular processes in plants [10]. GSH is an unusual peptide that forms a peptide bond between the amino group of cysteine and the carboxyl group of the glutamic acid side chain. GSH is an essential molecule, however, there is still no complete clarity on this issue. The main function of this small molecule is due to its antioxidant properties. GSH reduces ROS and is itself converted into the oxidized form of GSSG. Under control conditions, without stress, the GSH/GSSG ratio reaches 20:1. Under stress conditions, this ratio changes. The ratio of the reduced form of glutathione to its oxidized form GSH/GSSG shows the level of oxidative stress, which is one of the most important parameters of the cell state. The redox potential depends on the GSH concentration. Even if the GSH/GSSG ratio remains unchanged, but the GSH concentration increases, this will lead to a decrease in potential [11].
It was found by immunoprecipitation that GSH is localized in the nucleus and cytosol during the G1 phase cell cycle [12]. This localization of GSH is assumed to be dynamic. Translocation of GSH from the cytosol to the nucleus during the G1 phase is accompanied by cytosolic oxidation and accumulation of ROS in the cytosol [13].
In plants, GSH effectively protects the cell from stress-induced formation of reactive oxygen species [14]. GSH can react chemically with ROS [15]. The relationship between peroxide and GSH status has been proven [16]. At a low peroxide content, the GSH/GSSG ratio is almost equal to 1. However, an increase in the peroxide concentration is accompanied by the conversion of GSH into the oxidized form of GSSG and a change in the GSH/GSSG ratio. From these data, it can be concluded that GSH is a good marker for oxidative stress caused by increased peroxide production. The chemical reaction of GSH peroxide reduction proceeds slowly [17], but the presence of peroxidases accelerates this process. Heme peroxidases (GPx) are divided into three classes. Class II GPCs are found in fungi, and class III GPCs are found only in plants [18]. GPCs, also known as guaiacol-type peroxidases, are encoded by several genes and are localized in the apoplast and vacuole. GPCs may be involved in the formation of ROS [19].
Being one of the main components of the antioxidant system, GSH is oxidized by reactive oxygen species and thereby preventing increased oxidation of cellular components. A distinctive feature of GSH from other primary and secondary metabolites, which can also react with ROS, is the rapid reduction of its oxidized form [14]. GSH is contained in the cell in millimolar concentrations. This high level of GSH and the rapid rate of reduction of its oxidized form GSSG provide it with an irreplaceable role in the redox homeostasis of the cell. Reduction of the oxidized form of GSSG is carried out by the enzyme glutathione reductase (GR) [20]. GR is localized in chloroplasts and mitochondria [21,22,23], but another form has been found in the cytosol and peroxisome [24]. The enzyme is constantly active, however, under oxidative stress its level can increase even more. However, no increase in stress tolerance was observed in several plant species when GR was overexpressed [25].
As a result of various negative environmental influences, plant development processes slow down [26]. Using Arabidopsis mutants, GSH was shown to be involved in the regulation of plant growth and adaptation to abiotic and biotic environments [27,28]. A significant increase in the redox potential leads to damage to compartments sensitive to redox effects, resulting in growth arrest and/or even death [29]. It is known that in the root quiescent center (QC), responsible for the elongation of root cells, a high oxidative status is maintained [30]. The redox potential of glutathione in these cell types is relatively high and the GSH:GSSG ratio content of the vacuole or endoplasmic reticulum is low [31]. However, along with partial cell death as a result of exposure to high redox potential, high redox potential in resting cells influences the processes that determine cell fate [32] and associated responses to abiotic stress [33].
Another function of GSH related to protecting plants from stress is its participation in the detoxification process. GSH is a substrate for the synthesis of phytochelatins, which are a polymeric form of glutathione [34], which is capable of binding heavy metals [35,36], and is also involved in the detoxification of xenobiots together with glutathione-S- transferase (GST) [8], which catalyzes the formation of a covalent bond between the sulfur atom of the cysteine residue of GSH with an electrophilic compound [37].
The GST family in plants exhibits diverse biochemical and physiological functions [38]. It has been shown that some GSTs can perform antioxidant functions, others have peroxidase activity and are induced by H2O2 and, therefore, can be considered markers of increased intracellular H2O2 content [39].
The formation of disulfide bonds with various proteins ensures the participation of GSH in various signaling processes. Changes in the antioxidant activity of glutathione are accompanied by changes in its participation in cellular signaling pathways and interaction with various GSH-dependent enzymes [40].
GSH synthesis occurs in two ATP-dependent reactions. The first stage involves the synthesis of γ-glutamylcysteine (γGC) from glutamate and cysteine, which is catalyzed by the enzyme γ-glutamylcysteine synthetase or γ-glutamylcysteine ligase (GSH1, γ-GCL) [41]. In the second step, glutathione synthetase (GS or GSH2) catalyzes the formation of GSH from γGC and glycine. In plants, GSH synthesis occurs in plastids and cytosol [42,43]. Mutations in γ-glutamylcysteine ligase (GCL) in Arabidopsis have been shown to weaken plant defense mechanisms against abiotic stresses [44].
GSH plays an important role in plant development. Increasing the GSH content in plants would significantly activate the mechanisms of antioxidant and adaptive systems. In addition, GSH is widely used in pharmaceutical practice as an antioxidant and also as a food additive. Therefore, increasing the GSH content in plants may also have commercial value. One of the most common approaches to increase GSH content is to produce transgenic plants. Basically, transgenic plants with increased expression of GCL were used. Overexpression of γ-ECS in tobacco chloroplasts resulted in a significant increase in GSH in tobacco leaves [45] and in Arabidopsis resulted in an approximately twofold increase in GSH content [46]. A significant increase in GSH was achieved by overexpression of bifunctional γ-ECS/GSH-S from Streptococcus [47]. However, research has shown that significant success has not been achieved using transgenes due to the complex control of GSH content in cells [48].
Another way to increase GSH content may be to increase the sulfur (S) content in the nutrient medium, since GSH is a scavenger of non-protein sulfur. Indeed, GSH has been associated with changes in resistance to certain stresses as a result of changes in sulfur availability [49]. Stresses that increase oxidation have been reported to upregulate S uptake. For example, ozone exposure increases cysteine and GSH levels, respectively [50].
Peptides are small molecules that have been found in all plant organs [51]. Depending on their localization, peptides have specific functional activity. They participate in the regulation of cell differentiation, growth, development and plant defense, in addition, peptides participate in intercellular communications and long-distance signaling [52,53,54]. The number of functionally characterized peptide hormones exceeds the number of classical plant hormones. One of the largest and best-studied families of peptide hormones is CLE (CLV3 / ESR), consisting of 12-13 amino acids [55]. Peptides CLE have a wide range of functional activities, including control of the activity of apical meristems of shoots and roots and cambium, differentiation of vascular tissues, formation of lateral roots and nodules, early embryogenesis, stomatal development and response to several environmental factors: water availability and changes in the composition of soil nitrogen. Stem cells are important precursor cells in plants that can divide and specialize into other cell types, such as leaves and flowers. Control of stem cell differentiation and continuous replenishment of the stem cell pool are essential for normal plant development. It is proposed that two negative feedback loops control stem cells in Arabidopsis. One loop involves the peptide CLV3, the other involves CLE40 [56].
Previously, it was shown that the short tetrapeptide AEDL stimulates the development of the root system in Nicotiana tabacumх [57]. It was suggested that the peptide AEDL acts similarly to the peptide CLV3. The FITC-labeled peptide is localized predominantly in the elongation zone and slightly in the meristem zone. This localization of the peptide AEDL suggests its binding to the hydrophobic leucine-rich motif of the receptor CLV1, thereby preventing its penetration into the stem cell niche [58,59]. Binding of the peptide AEDL to the receptor CLV1 leads to activation of the receptor complex, thereby limiting the stem cell population and activating the process of stem cell differentiation. At the same time, the formation of a complex between the peptide AEDL and the receptor CLV1 prevents the penetration of the peptide CLV3 into the meristem from the QC zone. By preventing the penetration of the peptide CVL3 into the QC zone, the process of activation of the Wuschel-like homeobox (WUS) transcription factors and activation of the process of stem cell differentiation occurs.
Since the peptide AEDL is involved in the regulation of the proliferation-differentiation process, and this process depends on the redox balance, the aim of this study was to determine the relationship between the peptide AEDL and one of the most important participants in the redox balance - GSH.
2. Results
2.1. Plant Materials
Salt stress is the most common abiotic stress that negatively affects the growth and development of most crop plants [60]. High salt concentrations have a negative effect mainly due to disruption of the ionic and osmotic balance in the cell. In saline soils, high levels of sodium ions lead to delayed plant growth and even death. There are many approaches to reduce the negative effects of salt stress. We have previously shown that growing N.tabacum in the presence of the peptide AEDL prevents the negative effects of 150 mM NaCl. It should be noted that the peptide AEDL contributes to a noticeable activation of plant development, especially the root system, compared to the control tobacco (Figure 1).
2.2. Expression of RGF1 Gene
Plant growth regulatory factors (RGFs) are specific transcription factors and participate in the regulation of plant root system development. [61,62,63]. RGFs are a whole family of the peptides, but the most studied is RGF1.
Peptide AEDL stimulates gene activity in RGF1 roots by 2.68 times compared to the control N.tabacum. Addition of 150 mM NaCl leads to a 1.9-fold decrease in RGF1 gene activity. Although the presence of AEDL increases RGF1 gene expression by 1.75 times, its activity does not reach the gene activity in the control N.tabacum sample.
2.3. ROS Content
ROS perform both signaling and regulatory functions in plant cells [64]. They are formed in various cell compartments: in chloroplasts, mitochondria, peroxisomes, plasma membrane, cytosol, and cell membrane [65]. Abiotic stresses, including salt stress, lead to oxidative stress and, consequently, to an increase in ROS content [66]. ROS content was determined using the Carboxy-H2DFFDA marker. ROS production was detected in all root tissues, but with different fluorescence intensities in different root zones. Since not all root zones were equally stained for ROS, we assessed the distribution of cells with elevated ROS levels in different root zones by combining images taken using phase-contrast microscopy and a fluorescent label in one focal plane. In the control samples of N.tabacum, as well as in the samples grown in the presence of AEDL, only a slight staining of ROS production was observed (Figure 3A, 3B). In tobacco grown in the presence of 150 mM NaCl, an increase in fluorescence intensity was observed; the highest intensity of ROS production was found in the meristem zone. The dye was practically not identified in the root cap, division zone, and elongation zone. Intense staining of fluorescence was observed in the absorption zone. Moreover, in this zone, the cells of the peripheral root tissues - the epidermis and cortex - were stained, while the tissues of the central cylinder were almost not stained (Figure 3C, 3D).
In tobacco roots grown under the influence of NaCl, the ROS marker Carboxy-H2DFFDA was observed in all zones: in the root cup cells, differentiation, elongation and in the absorption zones, and the staining was more intense in the elongation zone, which indicates an increase in the content of ROS production in these cells/zones and activation of oxidative stress. In all other zones, the staining was approximately the same. In the peripheral root tissues, the cortex cells were brightly stained, the epidermis cells were less bright, and the cells of the central cylinder were very weak. When growing N.tabacum in the presence of the peptide AEDL, the ROS marker was not identified in the root cap cells and differentiation zone, a slight fluorescent glow was observed in the elongation and absorption zones. Fluorescence in these zones was most clearly determined in the epidermal cells and to a lesser extent in the cortex cells. When tobacco grown in the presence of the peptide AEDL was exposed to 150 mM NaCl, the ROS content increased compared to tobacco grown without NaCl. The ROS marker fluorescence intensity accumulated in the division and elongation zones (Figure 3). More intense fluorescence of the peripheral root tissues was noted in the cortex and epidermis and minimal in the central cylinder.
Thus, the effect of sodium chloride on control tobacco plants differs from the effect of sodium chloride in combination with the peptide AEDL. A distinctive feature of sodium chloride treatment in the presence of the peptide AEDL is a decrease in the proportion of cells stained with the ROS marker in the differentiation zone of tobacco roots. Thus, if the direct effect of NaCl on tobacco root cells leads to an increase in the number of cells with an increased ROS content in the differentiation and elongation zones (by 10 and 18.2 times, respectively), then in the presence of AEDL, the number of cells with an increased ROS pool in these zones increases significantly less (by 5.8 times). Consequently, the peptide AEDL increases the resistance of cells in the differentiation and absorption zones to stress conditions under salinity (Figure 3). The presence of the peptide AEDL during Nicotiana tabacum cultivation protects root tissues, especially epidermal cells.
2.3.1. H2O2 content
The peroxide content is one of the markers of damage to plant tissues when exposed to stress factors (Table 1).
Based on the obtained data, it can be concluded that 150 mM NaCl leads to an increase in the H2O2 content in the roots of N.tabacum by 1.47 times. Although the H2O2 content in the leaves is significantly lower than in the roots by 4.45 times, an increase in the concentration of sodium chloride leads to an increase in the H2O2 content in the leaves by 2.48 times. The peptide AEDL reduces the amount of H2O2 in the roots of N.tabacum by 1.35 times and slightly affects the H2O2 content in the leaves, reducing its content by 1.08 times. In addition, the peptide AEDL partially neutralizes the negative effect of NaCl, reducing the formation of H2O2 in the roots by 1.1 times and more significantly in the leaves by 1.52 times.
2.4. Antioxidant Activity
Inhibition of the 2,2-diphenyl-1-picrylhydrazyl (DPPH) oxidation process is an indicator of the activity of the antioxidant system. Table 2 presents data on the antioxidant activity (AOA) of N.tabacum grown under different conditions and when exposed to 150 mM NaCl.
The AOA in the roots of N.tabacum is 1.64 times higher than in the leaves. Salt stress leads to a decrease in AOA in the roots of N.tabacum by 3.18 times, and in the leaves only by 1.57 times. Growing N.tabacum in the presence of the peptide AEDL increases AOA in the roots by 3.18 times, and in the leaves by 1.57 times. The presence of AEDL leads to an increase in tobacco resistance to the action of 150 mM NaCl and an increase in antioxidant activity. At the same time, AOA in the roots of N.tabacum is even slightly higher than in the roots of the control tobacco. The peptide AEDL contributes to an increase in antioxidant activity in the roots of tobacco after exposure to NaCl by 3.32 times, and in the leaves by 1.49 times.
2.4.1. Expression of MnSOD and Сu/ZnSOD Genes
The main participants in the antioxidant system are enzymes - superoxide dismutase (SOD) [67]. SODs represent a large family of enzymes that differ in their metal cofactors. The main function of SOD enzymes is to convert superoxide ion (O2.-) into hydrogen peroxide. Depending on the cofactor, these enzymes differ in their localization.
The expression of MnSOD and Cu/ZnSOD genes in tobacco roots in the presence of the peptide AEDL increases by 1.85 times and 1.27 times, respectively (Figure 4). An exception is the expression of the MnSOD gene in tobacco leaves: the peptide AEDL does not change its level. Addition of 150 mM NaCl leads to a 1.9-fold decrease in the expression level of the Cu/ZnSOD gene in N.tabacum roots and practically does not change the activity of the MnSOD gene. In tobacco leaves, sodium chloride even leads to an increase in the expression of the Cu/ZnSOD gene.
2.4.2. GSH Content
The GSH content was determined in the roots and leaves of Nicotiana tabacum grown on MS medium without and in the presence of the short peptide AEDL (Table 3).
In N.tabacum roots, the GSH content exceeds its content in leaves by 2.86 times. However, the GSH content in N.tabacum roots can be increased by 3.24 times when growing the plant in the presence of the short peptide AEDL. An increase in GSH concentration was also observed in N.tabacum leaves, although much less, only by 1.36 times. A high concentration of nitric chloride in the nutrient medium leads to a decrease in the GSH content in tobacco roots by 1.18 times. It is interesting to note that the decrease in GSH concentration in tobacco leaves under the influence of NaCl is more significant, by 2.15 times. The inverse relationship is observed in tobacco grown in the presence of the peptide AEDL under the influence of NaCl: in leaves, the GSH content decreases by 1.27 times, and in roots - by 1.83. It should be noted that although the GSH content in tobacco roots grown in the presence of AEDL decreases under the influence of NaCl, it is not so significant and even exceeds the GSH level in the roots of the control tobacco sample by 1.72 times.
2.4.3. Biosynthesis of GSH, Expression of GSH1 and GSH2 Genes
It is known that the biosynthesis of glutathione occurs in two stages: the synthesis of γ-glutamylcysteine (γGC) from glutamate and cysteine, which is catalyzed by the enzyme γ-glutamylcysteine synthetase or γ-glutamylcysteine ligase (GSH1, γ-GCL). In the second stage, glutathione synthetase (GS or GSH2) catalyzes the formation of GSH from γGC and glycine [42].
It is known that glutathione biosynthesis occurs in two stages: the synthesis of γ-glutamylcysteine (γGC) from glutamate and cysteine, which is catalyzed by the enzyme γ-glutamylcysteine synthetase or γ-glutamylcysteine ligase (GSH1, γ-GCL). In the second stage, glutathione synthetase (GS or GSH2) catalyzes the formation of GSH from γGC and glycine [42]. When growing N. tabacum in the presence of AEDL, the expression of the GSH1 and GSH2 genes increases by 1.4 and 1.47 times, respectively (Figure 5). In leaves, the expression of the GSH1 gene in tobacco in the presence of AEDL increases by 1.29 times, and the GSH2 gene only by 1.08 times. The presence of 150 mM NaCl in the nutrient medium leads to a decrease in the activity of the GSH1 and GSH2 genes, especially in the leaves. If the decrease in the activity of the GSH1 gene in the roots was only 1.14 times, then in the leaves it was 1.83 times. The decrease in the activity of the GSH2 gene under the influence of NaCl is not as dramatic as that of the GSH1 gene, only in the leaves the expression level fell 1.29 times. Growing N. tabacum in the presence of AEDL leads to an increase in the expression level of the GSH1 gene in the roots by 2.68 times, which is even higher than without NaCl by 1.67 times. In tobacco leaves grown under the same conditions, the peptide AEDL also increases the expression of the GSH1 gene. Although the expression level increases by 2.28 times compared to tobacco leaves grown in the presence of NaCl without the peptide AEDL, the expression level of the GSH1 gene remains almost at the same level as in tobacco leaves grown in the presence of the peptide AEDL without NaCl.
Addition of NaCl to the nutrient medium slightly reduces the expression of the GSH2 gene in tobacco roots by 1.05 times and more significantly in leaves by 1.29 times. However, the presence of the peptide AEDL leads to an increase in the activity of the GSH2 gene in the roots by 1.83 times and especially in the leaves - by 2.48 times. It should be noted that the peptide AEDL stimulates the activity of the GSH2 gene both in the roots and in the leaves of tobacco even more intensely in the presence of NaCl than without it.
The addition of 150 mM NaCl had a negative effect on GSH content (Table 3). The expression activity of the GSH1 and GSH2 genes was significantly reduced in both the roots and leaves of N.tabacum and its mutant. It is interesting to note that the highest expression of the GSH1 and GSH2 genes was in N.abacumt grown in the presence of AEDL (even higher than in the mutant), it increased by 2.68 times and 1.83 times, respectively. In N.tabacum roots, a slight decrease in GSH content by 1.18 times was observed. But in tobacco grown in the presence of AEDL, the GSH content, although decreased with the addition of 150 mM NaCl, still remained increased by 2.03 times compared to the control Nt under salt stress. The GSH content in the roots of the tobacco mutant decreases more significantly compared to N.tabacum by 2.95 times, and even in the presence of AEDL the concentration was reduced by 1.41 times.
2.4.4. Expression of GR and GST Genes
GSH binds to ROS, turning into the oxidized form of GSSG,
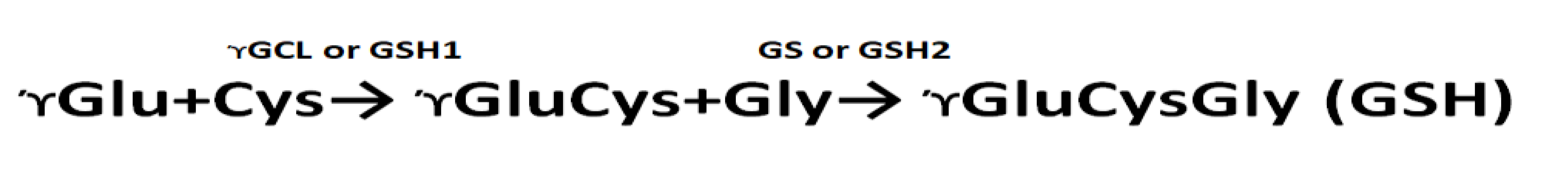
which is reduced by the enzyme glutathione reductase (GR) 20
Thus, the ratio of the reduced and oxidized forms of GSH:GSSG and, accordingly, the reduced form of GSH, depends on the activity of the GR enzyme.
The GR gene activity in tobacco roots is almost independent of the action of sodium chloride, but depends on the presence of the peptide AEDL and increases by 1.41 times. GST is an enzyme that, together with GSH, participates in detoxification. This is especially important under the influence of both abiotic and biotic stresses. The GST gene activity increases in the roots of tobacco grown in the presence of the peptide AEDL by 1.36 times. It should be noted that in tobacco leaves grown in the presence of the peptide AEDL, the expression of the GR and GST genes remains virtually unchanged. However, salt stress leads to a sharp increase in the expression of these genes by 4.26 and 4.39 times, respectively.
3. Discussion
Plants are constantly exposed to various environmental influences. These influences can be both long-term and short-term and vary in intensity. Under the influence of stress factors, plants either adapt to them or die [60]. Plants have developed a whole complex of counteraction to stressors. Depending on the protective mechanisms, plants under the influence of stress factors either acclimatize or die. Salt stress has a negative effect on the development of Nicotiana tabacum, its growth slows down, especially the root system. The short peptide AEDL promotes the development of the root system of Nicotiana tabacum and reduces the negative impact of sodium chloride.
Abiotic stresses, including salt stress, lead to the accumulation of excess ROS. Tobacco grown in the presence of the peptide AEDL accumulates ROS under the influence of NaCl to a lesser extent. Moreover, the distribution of ROS by zones in the roots of tobacco grown under different conditions is different. A distinctive feature of Nicotiana tabacum grown in the presence of AEDL is that the main amount of excess ROS accumulates in the zones of elongation and differentiation and is practically absent in the meristem and cap zones.
The peptide hormone RGF1 belongs to the Root Meristem Growth Factors family and is a secreted peptide of 13 amino acids [61,62,63]. The main function of peptides RGF is to regulate the development of the root system in plants. RGF family peptides are predominantly expressed in the root meristem in the stem cell region and participate in the formation of the root stem cell niche [68]. The RGF1 signaling cascade through receptors regulates the formation of the PLETHORA (PLT) gradient, which is known as the master regulator of root formation [68]. It has been shown that RGF1 is involved in the distribution of ROS along root development zones [69]. RGF1 can transmit a signal through ROS, controlling the size of the meristematic zone. It was found that after RGF1 treatment, the O2•– level in the meristematic zone increases, while the H2O2 content in the elongation and differentiation zones decreases [69].
Based on the obtained data, it follows that the peptide AEDL stimulates the synthesis of the peptide RGF1, the amount of which increases more than 2.5 times. It can be assumed that superoxide ion can accumulate in the RGF1-PLT signaling pathway, which can lead to PLT oxidation. However, it was found that in the roots of N.tabacum in the presence of the peptide AEDL, the expression of the MnSOD and Cu / ZnSOD genes increases, which promote the conversion of superoxide ion into H2O2. However, we showed that in tobacco roots the content of H2O2 in the presence of the peptide AEDL decreases almost 1.5 times. This fact may indicate the active participation of GSH in the neutralization of excess H2O2, especially since an increase in the concentration of GSH in tobacco roots in the presence of the peptide AEDL was found by more than 3 times.
Glutathione -γ-glutamylcysteinylglycine (GSH), a small molecule, has proven to be an important molecule without which plants cannot develop normally [10]. The reasons why this small molecule is essential are not fully understood, but it can be concluded that GSH performs functions in plant development that cannot be performed by other thiols or antioxidants. Known functions of GSH include roles in biosynthetic pathways, detoxification, antioxidant biochemistry, and redox homeostasis. Since ROS, especially H2O2, accumulate in plants under various abiotic stresses, many researchers are interested in increasing GSH levels in plants. The main strategies for increasing GSH content are the use of transgenic plants. However, it has been observed that elevated GSH levels do not always have a beneficial effect on plant development.
Motivated by the important role of GSH in plant function, many efforts have been made to increase its content in several plant species. The main strategic approaches to increase GSH content have largely relied on ectopic expression of GCL. Using transgenic plants, GSH content in plants could be increased two to six times [70]. However, many experiments did not achieve the expected results [71]. Overexpression of chloroplast γ-ECS in tobacco was accompanied by an increase in GSH levels, however, there was an increase in oxidation and tissue damage [45]. Another group of researchers found that one of the chloroplast lines with multiple insertions exhibited symptoms of early leaf senescence [72]. Another study showed that the overexpressor experienced a decrease in biomass and photosynthesis [73]. Other authors reported that tobacco overexpressors with very high GSH content did not show significant impairment in functional activity, and they consider these tobacco lines to be interesting subjects for further research [15]. Transgenic tobacco plants expressing a more complex StGCL-GS construct were reported to exhibit extreme GSH accumulation (up to 12 μM) in leaves, more than 20-30 times the GSH content of wild-type plants [15]. Surprisingly, this dramatically increased GSH production does not affect plant growth while increasing plant tolerance to abiotic stress. In addition, plants expressing StGCL-GS provide a new, economical source for GSH production that is competitive with existing yeast-based systems [47].
To increase the GSH content, we used the peptide AEDL. When growing tobacco in the presence of the peptide,AEDL the GSH content reliably increases in the roots by 3.24 times, and in the leaves by 1.36 times. It should be noted that with this option for increasing the GSH content, the plants feel comfortable, their root system is more developed compared to the control option, and the leaves have a larger area.
In tobacco grown in the presence of AEDL, the activity of γ-glutamylcysteine ligase increases only 1.47-fold, while GSH2 expression increases 1.4-fold. The second stage is probably limiting for GSH synthesis. This fact is important for the regulation of GSH content in plants; its accumulation can have negative consequences for normal plant development.
GSH is involved in H2O2 detoxification in complex with glutathione peroxidase (GP) and GST. Interestingly, GP expression levels were so low that they were not discussed in this study. Probably, the GSH-GST complex played a major role in H2O2 detoxification and detoxification. This fact is confirmed by the increase in GR expression, which is designed to reduce the oxidized form of GSSH to GSH. When growing N.tabacum in the presence of the peptide AEDL, a decrease in ROS formation is observed compared to control samples. Low concentrations of the peptide AEDL increase the expression of both Cu/ZnSOD and MnSOD, as well as the genes responsible for GSH biosynthesis - GSH1 and GSH2, which leads to an increase in AOA.
For a long time it was believed, that the accumulation of ROS has a negative effect, leading to disruption of plant development, tissue damage and, depending on the degree of negative impact, even death. ROS trigger signaling in response to stress, and excess ROS are neutralized by antioxidants to prevent oxidative damage to cells. However, in our experiment, the level of ROS and H2O2 in mutant tobacco is lower than in wild tobacco, and a significant slowdown in growth is observed in it. Recently, evidence has accumulated that the redox balance determines the fate of stem cells [74]. Plant growth and development depend on the maintenance and continued differentiation of stem cells located in the central zone (CZ) of the apical meristem in roots (RAM) and shoots (SAM). The processes of proliferation and differentiation of stem cells are strictly controlled by signaling molecules, peptides and transcription factors [74]. Stem cell fate is determined by a negative feedback mechanism between the homeodomain transcription factor WUSCHEL (WUS), which is expressed in a small subset of organizing center (OC) cells, and the secreted peptide CLAVATA3 (CLV3), which negatively regulates WUS expression. Downregulation or loss of WUS function causes plant stem cell shrinkage or death. On the other hand, the peptide CLV3 binds to the CLV1 receptor, a leucine-rich kinase receptor that is located at the boundary of the stem cell niche and interferes with the transit of CLV3 and the suppression of WUS activity.
Recently, it was shown that to regulate the processes of proliferation and differentiation of stem cells, a redox balance is necessary and its main participants are H2O2 and O2.- [56]. Regulation of the redox balance also occurs through a negative feedback loop mechanism. It was found that SOD is localized in the peripheral zone (PZ) and is involved in the conversion of O2.- to H2O2. The accumulation of H2O2 leads to suppression of WUS expression and an increase in the process of stem cell differentiation. Increasing the content of O2.- in the CZ leads to an increase in the stem cell niche by activating WUS expression. Thus, self-maintenance of the ROS balance in stem cells occurs. There is no evidence of the participation of GSH in the regulation of the redox balance in stem cells, however, we believe that GSH is also integrated into this process.
Increased expression of Cu/ZnSOD and MnSOD in tobacco grown in the presence of the peptide AEDL leads to active neutralization of O2.-, converting them into H2O2. A decrease in the O2.- content in stem cells is accompanied by suppression of WUS activity and a decrease in the stem cell pool and, consequently, an increase in cellular differentiation. On the other hand, an increase in the GSH content and activation of the GR and GST genes involved in the detoxification of H2O2 together with GSH leads to a decrease in the H2O2 content and, accordingly, to an increase in stem cell proliferation. Although N.tabacum grown in the presence of the peptide AEDL also shows a decrease in the ROS content and activation of the Cu/ZnSOD and MnSOD genes, the root system becomes more developed compared to the control samples. These results suggest that the peptide is capable of participating in the regulation of the redox balance in stem cells. We suggested the following mechanism of regulation of ROS balance (Figure 6).
AEDL stimulates GSH synthesis activity. As a tripeptide, GSH can bind to the CLV1 receptor, which helps prevent its penetration into the PZ and neutralize H2O2. An increase in H2O2 suppresses WUS expression, activating the process of stem cell differentiation. Previously, we proposed a mechanism for regulating the growth of root cells and their elongation with the participation of the peptide AEDL in Nicotiana tabacum [58]. The peptide AEDL binds to the CLV1 receptor, preventing the CLV3 peptide from binding to it and its penetration into the stem cell niche and, thereby, suppressing WUS expression. Here we propose that by binding to the receptor CLV1, the peptide AEDL interferes with GSH binding, resulting in O2.- remaining high and WUS activity promoting an forming of the stem cell niche. Thus, an additional negative feedback loop is formed involving GSH and the peptide AEDL, regulating the redox balance in the stem cell niche. According to the presented scheme, it can be assumed that high concentrations of glutathione can lead to disruption of this balance and, as a consequence, to significant changes in the process of plant development.
4. Materials and Methods
4.1. Plant Material
Seeds of tobacco (Nicotiana tabacum L.) cultivar Samsun were placed in flasks containing hormone-free Murashige–Skoog (MS) agar medium. Next, tobacco seedlings were cut and planted in test tubes with liquid MS medium with or without 10–7 M AEDL or 150 mM NaCl. Experiments were carried out in four replicates. .
4.2. Fluorescence Microscopy
To determine ROS by the fluorescent method, root tips (4–5 mm) of seedlings were incubated in 25–50 nM carboxy-H2DFFDA (Thermo Fisher Scientific, USA) according to our method [57]. The samples were analyzed under an Olympus BX51 fluorescent microscope (Japan) with a 10X objective at a wavelength of 490 nm. Images were obtained using a Color View digital camera (Germany).
4.3. Biochemical Analysis
Antioxidant activity (AOA) was determined by the decrease in the coloration of the 5x10-5M alcohol solution 2,2-diphenyl-1-picrylhydrazyl (DPPH). Absorbance was measured at λ= 517 nm. AOA was calculated using the formula: (Ao-A/Ao)x100% [75]. The concentration of peroxide in aqueous solutions of plant material was determined by the reduction in coloration of a 0.02 M solution of KMnO4. Absorbance was measured at λ=480 nm [76]. The glutathione (GSH} content in mM was determined by the Elman method by the appearance of color after the addition of 0.01 M alcohol solution. Absorption Absorbance was measured at λ= 412 nm [77].
4.4. Total RNA Isolation and Gene Expression Analysis
Using a standard RNA isolation kit-Extran RNA Syntol (Russia), total RNAs were isolated from wheat roots and shoots grown under different conditions. cDNAs were synthesized by reverse transcription according to the standard method (Syntol, Moscow, Russia).
RT-PCR using SYBR Green I (Syntol) was performed in a CFX 96 Real-Time thermal cycler (BioRad,USA). Information on the structure of the FeSOD, MnSOD, GSH1, GSH2, GR and GST genes in N. tabacum was obtained from NCBI. Primers for the genes were synthesized by Syntol. Each RT-PCR reaction was performed in three repeats.
4.5. Statistical Methods
Statistical processing of experimental data was carried out using analysis of variance using the ANOVA program and t-Student test (DPS software) with significant differences at p<0.05. The least significant difference method was used to test significance. Values are presented as means ± standard deviations of triplicate biological replicates
5. Conclusions
It is noted that the peptide AEDL stimulates plant growth, especially the root system. The central dormant zone responsible for plant development has a high oxidative level, which regulates the fate of stem cells. It is assumed that GSH and the peptide AEDL form an additional negative feedback loop, participate in the regulation of the redox balance in the stem cell niche and the regulation of the fate of stem cells. It was found that AEDL activates GSH biosynthesis. From the presented scheme it follows that high concentrations of GSH can lead to disruption of this balance and, as a consequence, to significant changes in the process of plant development. However, the peptide AEDL, controlling the binding of GSH to the CVL1 receptor, prevents the penetration of excess GSH into the meristem zone and thereby prevents a decrease in the redox balance.
Author Contributions
N.V.K. performed light and fluorescent microscopy, evaluated data, wrote and finalized the manuscript L.I.F. designed and performed experiment, PCR, prepared figures, evaluated data, wrote and finalized the manuscript;.
Funding
The reported study was supported by FGUM-0431-2022-0003 of the Ministry of Science and Higher Education of the Russian Federation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem., 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot., 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Rohman, M.M.; Talukder, M.Z.A.; Hossain, M.G.; Uddin, M.S.; Amiruzzaman, M.; Biswas, A.; Ahsan, A.F.M.S.; Chowdhury, M.A.Z. Saline sensitivity leads to oxidative stress and increases the antioxidants in presence of proline and betaine in maize (Zea mays L. ) inbred. Plant Omics. J., 2016, 9, 35–47. [Google Scholar]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Stem Cell Dev. Biol., 2018, 80, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The roles of mitochondrial reactive oxygen species in cellular signaling and stress response in plants. Plant Physiol., 2016, 171, 1551–1559. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Del Rio, L. A. ROS and RNS in plant physiology: an overview. Journal of Experimental Botany, 2015, 66, 2827–2837. [Google Scholar] [CrossRef]
- Dorion, S.; Ouellet, J.C.; Rivoal, J. Glutathione metabolism in plants under stress: beyond reactive oxygen species detoxification. Metabolites, 11.
- Navrot, N.; Finnie, C.; Svensson, B.; Hägglund, P. Plant redox proteomics. J Proteomics, 2011, 12, 1450–1462. [Google Scholar] [CrossRef]
- Guo, W.; Ou, S.; Lu, Q. ; Zhong, Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity, Plant Physiol. Biochem., 2006, 44, 828-836. , 44.
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Jenni, Han Y. ; Neukermans, J.; Marquez –Garcia B.; Foyer C.H. Glutathione in plants: an integrated overview. Plant, Cell & Environment, 2012, 35, 454–484. [Google Scholar]
- Cairns, N.G.; Pasternak, M.; Wachter, A.; Cobbett, C.S.; Meyer, A.J. Maturation of Arabidopsis seeds is dependent on glutathione biosynthesis within the embryo. Plant Physiology, 2006, 141, 446–455. [Google Scholar] [CrossRef]
- Zechmann, B.; Mauch, F.; Sticher, L.; Müller, M. Subcellular immunocytochemical analysis detects the highest concentrations of glutathione in mitochondria and not in plastids. Journal of Experimental Botany, 4017. [Google Scholar]
- Diaz-Vivancos, P.; Wolff, T.; Markovic, J.; Pallard, O.F.V.; Foyer, C.H. A nuclear glutathione cycle within the cell cycle. Biochemical Journal, 2010, 431, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Arisi, A.; Jouanin, L.; Kunert, K.; Rennenberg, H.; Foyer, C. Glutathione: biosynthesis, metabolism and relationship to stress tolerance explored in transformed plants. J. Exp. Bot., 1998, 49, 623–647. [Google Scholar] [CrossRef]
- Chaouch, S.; Queval, G.; Vanderauwera, S.; Mhamdi, A.; Vandorpe, M.; Langlois-Meurinne, M.; Van Breusegem, F.; Saindrenan, P.; Noctor. G. Peroxisomal hydrogen peroxide is coupled to biotic defense responses by isochorismate synthase 1 in a daylength-related manner. Plant Physiology, 2010, 153, 1692–1705.
- Queval, G.; Thominet, D.; Vanacker, H.; Miginiac-Maslow, M.; Gakière, B.; Noctor, G. H2O2-activated up-regulation of glutathione in Arabidopsis involves induction of genes encoding enzymes involved in cysteine synthesis in the chloroplast. Molecular Plant, 2009, 2, 344–356.
- Mhamdi, A.; Hager, J.; Chaouch, S. Arabidopsis GLUTATHIONE REDUCTASE 1 plays a crucial role in leaf responses to intracellular H2O2 and in ensuring appropriate gene expression through both salicylic acid and jasmonic acid signaling pathways. Plant Physiology, 2010, 153, 1144–1160.
- Zámocky, M.; Furtmüller, P.G.; Obinger, C. Evolution of structure and function of Class I peroxidases. Archives of Biochemistry and Biophysics, 2010, 500, 45–57.
- Cosio, C.; Dunand, C. Specific functions of individual class III peroxidase genes. Journal of Experimental Botany, 2009, 60, 391–408.
- Ding, S.; Jiang, R.; Lu, Q.; Wen, X.; Lu, C. Glutathione reductase 2 maintains the function of photosystem II in Arabidopsis under excess light. Biochim Biophys Acta Bioenerg, 2016, 6, 665–67. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta, 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Müller-Schüssele, S.J.; Wang, R.; Gütle, D.D.; Romer, J.; Rodriguez-Franco, M.; Scholz, M.; Buchert, F.; Lüth, V.M.; Kopriva, S.; Dörmann, P.; Schwarzländer, M.; Reski, R.; Hippler, M.; Meyer, A.J. Chloroplasts require glutathione reductase to balance reactive oxygen species and maintain efficient photosynthesis. Plant J., 2020, 103, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Marty, L.; Bausewein, D.; Müller, C.; Bangash, S.A.K.; Moseler, A.; Schwarzländer, M.; Müller-Schüssele, S.J.; Zechmann, B.; Riondet, C.; Balk, J.; Wirtz, M.; Hell, R.; Reichheld, J.-P.; Meyer, A.J. Arabidopsis glutathione reductase 2 is indispensable in plastids, while mitochondrial glutathione is safeguarded by additional reduction and transport systems. New Phytol., 2019, 224, 1569–1584. [Google Scholar] [CrossRef] [PubMed]
- Kataya, A.M.R.; Reumann, S. Arabidopsis glutathione reductase 1 is dually targeted to peroxisomes and the cytosol. Plant Signaling & Behavior, 2010, 5, 171–175. [Google Scholar]
- Ding, S.H.; Lu, Q.T.; Zhang, Y.; Yang, Z.P.; Wen, X.G.; Zhang, L.X.; Lu, C.M. Enhanced sensitivity to oxidative stress in transgenic tobacco plants with decreased glutathione reductase activity leads to a decrease in ascorbate pool and ascorbate redox state. Plant Molecular Biology, 2009, 69, 577–592. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res., 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- ausz, M.; Sircelj, H.; Grill D. The glutathione system as a stress marker in plant ecophysiology: is a stress-response concept valid? Journal of Experimental Botany, 2004, 55, 1955–1962.
- Ogawa, K. Glutathione-associated regulation of plant growth and stress responses Antioxid Redox Signal, 2005, 7, 973-81.
- Kranner, I.; Birtic, S.; Anderson, K.M.; Pritchard, H.W. Glutathione half-cell reduction potential: a universal stress marker and modulator of programmed cell death. Free Radical Biology & Medicine, 2165. [Google Scholar]
- Kranner, I.; Beckett, R.P.; Wornik, S.; Zorn, M.; Pfeifhofer, H.W. Revival of a resurrection plant correlates with its antioxidant status. The Plant Journal,.
- Enyedi, B.; Várnai, P.; Geiszt, M. Redox state of the endoplasmic reticulum is controlled by Ero1L-alpha and intraluminal calcium. Antioxidants & Redox Signaling, 2010, 13, 721–729. [Google Scholar]
- Jiang, K.; Feldman, L. Positioning of the auxin maximum affects the character of cells occupying the root stem cell niche. Plant Signaling and Behavior, 2010, 5, 1–3.
- Dinneny, J.R.; Long, T.A.; Wang, J.Y.; Mace, D.; Pointer, S.; Barron, C.; Brady. S.M.; Schiefelbein, J.; Benfey, P.N. Cell identity mediates the response of Arabidopsis roots to abiotic stress. Science, 2008, 32, 942–945.
- Cazalé, A.C.; Clemens, S. Arabidopsis thaliana expresses a second functional phytochelatin synthase. FEBS Letters, 2001, 507, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: roles in heavy metal detoxifaction and homeostasis. Annual Review of Plant Biology, 2002, 53, 159–182.
- Alfenito, M.R.; Souer, E.; Goodman, C.D.; Buell, R.; Mol, J.; Koes, R.; Walbot, V. Functional complementation of anthocyanin sequestration in the vacuole by widely divergent glutathione S-transferases. Plant Cell, 1998, 10, 1135–1149. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Edwards, R. (2010) Glutathione S-transferases. The Arabidopsis Book, 2010, 8, e0131. [Google Scholar] [CrossRef]
- Dixon, D.P.; Davis, B.G.; Edwards, R. Functional divergence in the glutathione transferase superfamily in plants. Identification of two classes with putative functions in redox homeostasis in Arabidopsis thaliana. Journal of Biological Chemistry, 2002, 277, 30859–30869.
- Queval, G.; Thominet, D.; Vanacker, H.; Miginiac-Maslow, M.; Gakière, B.; Noctor, G. H2O2-activated up-regulation of glutathione in Arabidopsis involves induction of genes encoding enzymes involved in cysteine synthesis in the chloroplast. Molecular Plant, 2009, 2, 344–356.
- Csiszár, J.; Horváth, E.; Bela, K.; Gallé, Á. Glutathione-related enzyme system: Glutathione reductase (GR), glutathione transferases (GSTs) and glutathione reroxidases (GPXs) In: Gupta DK, Palma JM, Corpas FJ, editors. Redox state as a central regulator of plant-cell stress responses. Cham: Springer International Publishing, 2016, 137–158.
- Wachter, A.; Wolf, S.; Steininger, H.; Bogs, J.; Rausch, T. Differential targeting of GSH1 and GSH2 is achieved by multiple transcription initiation: implications for the compartmentation of glutathione biosynthesis in the Brassicaceae. Plant J., 2005, 41, 15–30. [Google Scholar] [CrossRef]
- Noctor, G.; Gomez, L.; Vanacker, H.; Foyer, C.H. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. J. Exp. Bot., 2002, 53, 1283–1304. [Google Scholar] [CrossRef]
- Parisy, V.; Poinssot, B.; Owsianowski, L.; Buchala, A.; Glazebrook, J.; Mauch, F. Identification of PAD2 as a γ-glutamylcysteine synthetase highlights the importance of glutathione in disease resistance of Arabidopsis. Plant J., 2007, 49, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Creissen, G.; Firmin, J.; Fryer, M.; Kular, B.; Leyland, N.; Reynolds, H.; Pastori, G.; Wellburn, F.; Baker, N.; Wellburn, A.; Mullineaux, P. Elevated glutathione biosynthetic capacity in the chloroplasts of transgenic tobacco paradoxically causes increased oxidative stress. The Plant Cell, 1999, 11, 1277–1291. [Google Scholar] [CrossRef]
- Xiang, C.; Werner, B.L.; Christensen, E.M.; Oliver, D.J. The biological functions of glutathione revisited in Arabidopsis transgenic plants with altered glutathione levels. Plant Physiology, 2001, 126, 564–574.
- Liedschulte, V.; Wachter, A.; Zhigang, A.; Rausch, T. Exploiting plants for glutathione (GSH) production: uncoupling GSH synthesis from cellular controls results in unprecedented GSH accumulation. Journal of Plant Biotechnology, 2010, 8, 807–820.
- Rennenberg, H.; Herschbach, C.; Haberer, K.; Kopriva, S. Sulfur metabolism in plants: are trees different? Plant Biol. (Stuttg.), 2007, 9, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Höller, K.; Király, L.; Künstler, A.; Müller, M.; Gullner, G.; Fattinger, M.; Zechmann, B. Enhanced glutathione metabolism is correlated with sulfur-induced resistance in tobacco mosiac virus-infected genetically susceptible Nicotiana tabacum plants. Molecular Plant-Microbe Interactions, 1459. [Google Scholar]
- Bick, J.A.; Setterdahl, A.T.; Knaff, D.B.; Chen, Y.; Pitcher, L.H.; Zilinskas, B.A.; Leustek, T. Regulation of the plant-type 5′-adenylyl sulfate reductase by oxidative stress. Biochemistry, 9040. [Google Scholar]
- Fedoreyeva, L.I. Molecular Mechanisms of Regulation of Root Development by Plant Peptides. Plants (Basel) 2023, 12, 1320. [Google Scholar] [CrossRef]
- Albert, M. Peptides as triggers of plant defence. J. Exp. Bot. 2013, 64, 5269–5279. [Google Scholar] [CrossRef]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B. , Sussman, M.R. A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Grienenberger, E.; Fletcher, J.C. Polypeptide signaling molecules in plant development. Curr. Opin. Plant Biol. 2015, 23, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Fiers, M.; Golemiec, E; van der Schors, R. ; van der Geest, L.; Li, K. W.; Stiekema, W. J.; Liu C.-M. The CLAVATA3/ESR motif of CLAVATA3 is functionally independent from the nonconserved flanking sequences. Plant Physiol 2006, 141, 1284–1292. [Google Scholar] [CrossRef]
- Schlegel, J.; Denay, G.; Wink, R.; Gustavo Pinto, K.; Stahl, Y.; Schmid, J.; Blümke, P.; Simon, R. G.W. Control of Arabidopsis shoot stem cell homeostasis by two antagonistic CLE peptide signalling pathways eLife. 2021, 10, e70934.
- Fedoreyeva,L. I.; Baranova, E.N.;Chaban, I.A.; Dilovarova, T.A.; Vanyushin,B.F.; Kononenko,N.V. Elongating Effect of the Peptide AEDL on the Root of Nicotiana tabacum under Salinity. Plants (Basel), 2022, 11, 1352–1367. [Google Scholar]
- Poliushkevich, L.O.; Gancheva, M.S.; Dodueva, I.E.; Lutova, L.A. Receptors of CLE peptides in plants. Russ. J. Plant Physiol. 2020, 67, 1–16. [Google Scholar] [CrossRef]
- Dolzblasz, A.; Nardmann, J.; Clerici, E.; Causier, B.; van der Graaff, E.; Chen, J.; Devies, B.; Werr, W.; Laux, N. Stem cell regulation by Arabidopsis WOX genes. Mol Plant. 2016, 9, 1028–1039. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu Rev Plant Biol., 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Meng, L; Buchanan, B. B.; Feldman, L.J.; Luan, S. CLE-like (CLEL) peptides control the pattern of root growth and lateral root development in Arabidopsis. Proc Natl Acad Sci USA 2012, 109, 1760–1765. [Google Scholar] [CrossRef]
- Whitford, R.; Fernandez, A.; Tejos, R.; Pérez, A.C.; Kleine-Vehn, J.; Vanneste, S.; Drozdzecki, A.; Leitner, J.; Abas, L.; Aerts, M.; .Hoogewijs, K.; Baster, P.; De Groodt, R.; Lin, Y.-C.; Storme, V.; Van de Peer, Y.; Beeckman, T.; Madder, A.; Devreese, B; Luschnig, C. ; Friml, J.; Hilson, P. GOLVEN secretory peptides regulate auxin carrier turnover during plant gravitropic responses. Dev Cell 2012, 22, 678–685. [Google Scholar] [CrossRef]
- Fernandez, A.; Drozdzecki, A.; Hoogewijs, K.; Nguyen, A.; Beeckman, T.; Madder, A.; Hilson, A. Transcriptional and functional classification of the GOLVEN/ROOT GROWTH FACTOR/CLE-like signaling peptides reveals their role in lateral root and hair formation. Plant Physiol 2013, 161, 954–970. [Google Scholar] [CrossRef]
- Mittler, R. : Vanderauwera, S.: Suzuki, N.: Miller, G.: Tognett,i V.B.: Vandepoele, K.: Gollery, M.: Shulaev, V.: Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 2011. 16, 300–309. [Google Scholar] [CrossRef]
- Foyer, C.H. , Noctor G. Redox signaling in plants. Antioxid. Redox Signal. 2013, 18, 2087–2090. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Zhou, W.; Wei, L.; Xu, J.; Zhai, Q.; Jiang, H.; Chen, R.; Chen, Q.; Sun, J.; Chu, J.; Zhu, L. , Liu, C.-M.; Li, C. Arabidopsis Tyrosylprotein sulfotransferase acts in the auxin/PLETHORA pathway in regulating postembryonic maintenance of the root stem cell niche. Plant Cell. 2010, 22, 3692–3709. [Google Scholar]
- Nakayama, T.; Shinohara, H.; Tanaka, M.; Baba, K.; Ogawa-Ohnishi, M.; Matsubayashi, Y. A peptide hormone required for Casparian strip diffusion barrier formation in Arabidopsis roots. Science 2017, 355, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Strohm, S.; Jouanin, L.; Kunert, K.J.; Foyer, C.H.; Rennenberg, H. Synthesis of glutathione in leaves of transgenic poplar overexpressing γ−glutamylcysteine synthetase. Plant Physiology, 1996, 112, 1071–1078.
- Rennenberg, H.; Herschbach, C.; Haberer, K.; Kopriva, S. Sulfur metabolism in plants: are trees different? Plant Biol. (Stuttg.), 2007, 9, 620–637. 9.
- Herschbach, C.; Rizzini, L.; Mult, S.; Hartmann, T.; Busch, F.; Peuke, A.D.; Kopriva, S.; Ensminger , I. Over-expression of bacterial γ-glutamylcysteine synthetase (GSH1) in plastids affects photosynthesis, growth and sulphur metabolism in poplar (Populus tremula × Populus alba) dependent on the resulting γ-glutamylcysteine and glutathione levels. Plant, Cell & Environment, 2009, 33, 1138–1151.
- Ivanova, L.A.; Ronzhina, D.A.; Ivanov, L.A.; Stroukova, L.V.; Peuke, A.D.; Rennenberg, H. Overexpression of gsh1 in the cytosol affects the photosynthetic apparatus and improves the performance of transgenic poplars on contaminated soil. Plant Biology, 2011, 13, 649–659.
- Zeng, J.; Dong, Z.; Wu, H.; Tian, Z.; Zhao, Z. Redox regulation of plant stem cell fate EMBO J. , 2017, 36, 2844–2855. [Google Scholar]
- Adesanwo, J.K.; Makinde, O.O.; Obafemi, C.A. Phytochemical analysis and antioxidant activity of methanol extract and betulinic acid isolated from the roots of Tetracera potatoria. J. of Pharmacy Research, 2013, 6, 903 907.
- Zalutskaya, Zh.M.; Skryabina, U.S.; Ermilova, E. V. Hydrogen peroxide generation and transcription regulation of antioxidant enzyme expression Chlamydomonas reinbardtii under hypothermia. Plant Physiology (Rus), 2019, 66, 104–111. [Google Scholar]
- Ellman, G. L. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 1959, 82, 70–81. [Google Scholar] [CrossRef]
Figure 1.
Nicotiana tabacum, grown in different conditions: 1- control; 2- 150 mM NaCl; 3- AEDL; 4- AEDL+ 150 mM NaCl.
Figure 1.
Nicotiana tabacum, grown in different conditions: 1- control; 2- 150 mM NaCl; 3- AEDL; 4- AEDL+ 150 mM NaCl.

Figure 2.
Expression RGF1 gene in root Nicotiana tabacum. Significant differences were defined as p<0.05.
Figure 2.
Expression RGF1 gene in root Nicotiana tabacum. Significant differences were defined as p<0.05.
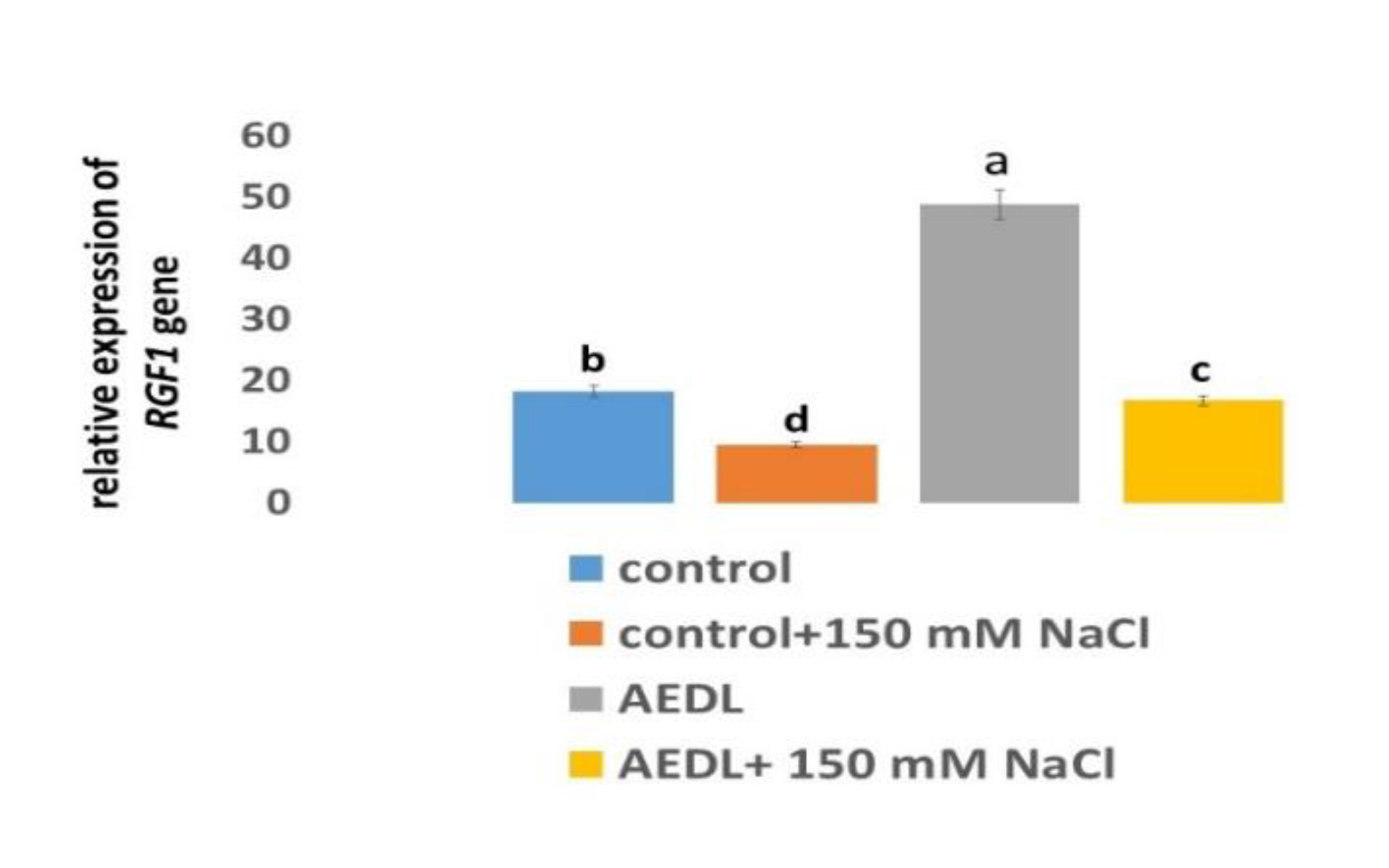
Figure 3.
A - Distribution of ROS+ and ROS - cells in the root zones N. tabacum. control-a,a’; 150 mM NaCl- b,b’; AEDL- c,c’; AEDF+150 mM NaCl – d,d’. DZ - differentiation zone, EZ - elongation zone, MZ-meristem zone. Bar 400 µm. B -The intensity of ROS fluorescence in N. tabacum, grown in different conditions. C - Distribution of fluorescence intensity in the root zones. D - Distribution of fluorescence intensity in the root tissues. Significant differences were determined p < 0.05.
Figure 3.
A - Distribution of ROS+ and ROS - cells in the root zones N. tabacum. control-a,a’; 150 mM NaCl- b,b’; AEDL- c,c’; AEDF+150 mM NaCl – d,d’. DZ - differentiation zone, EZ - elongation zone, MZ-meristem zone. Bar 400 µm. B -The intensity of ROS fluorescence in N. tabacum, grown in different conditions. C - Distribution of fluorescence intensity in the root zones. D - Distribution of fluorescence intensity in the root tissues. Significant differences were determined p < 0.05.
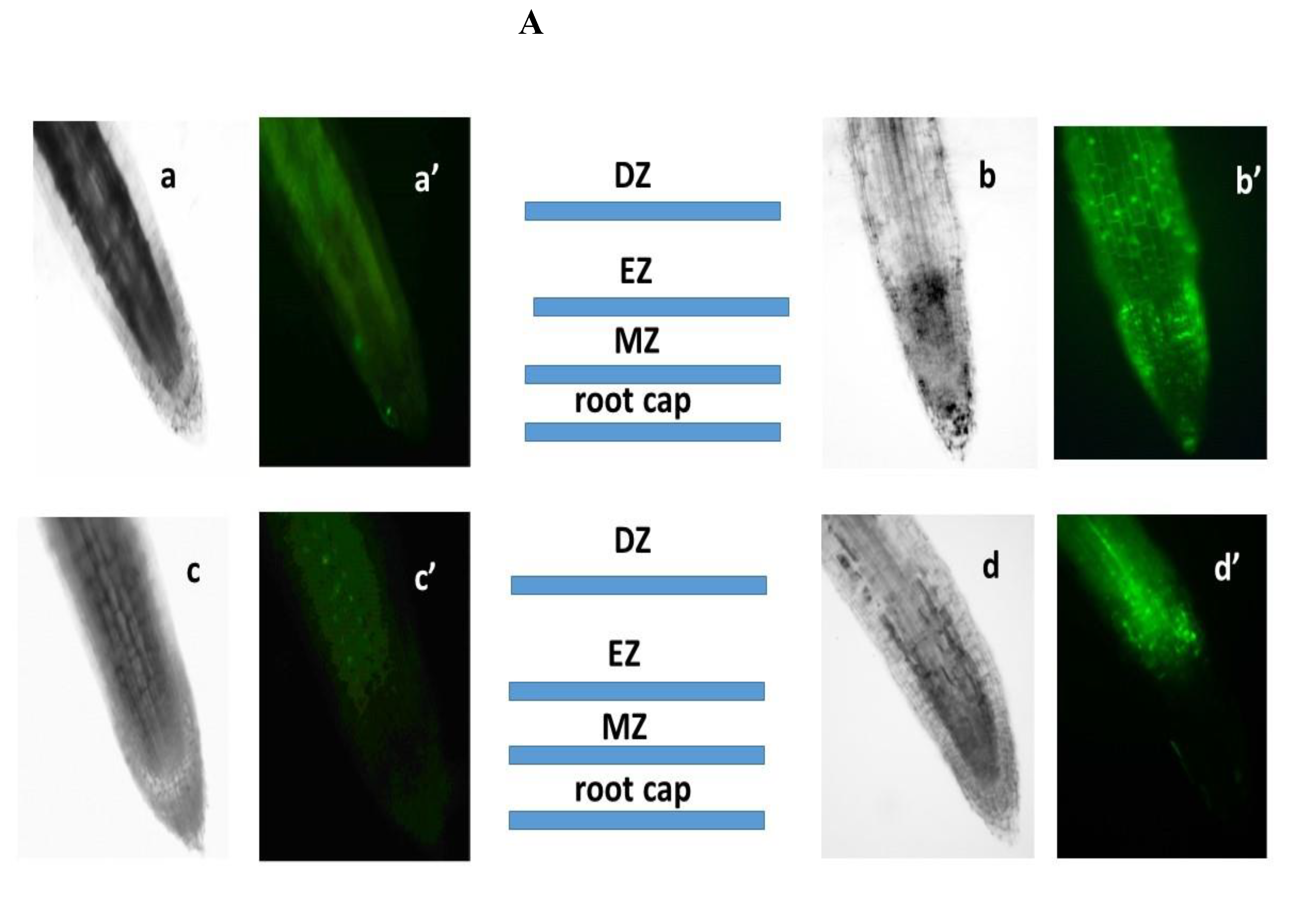
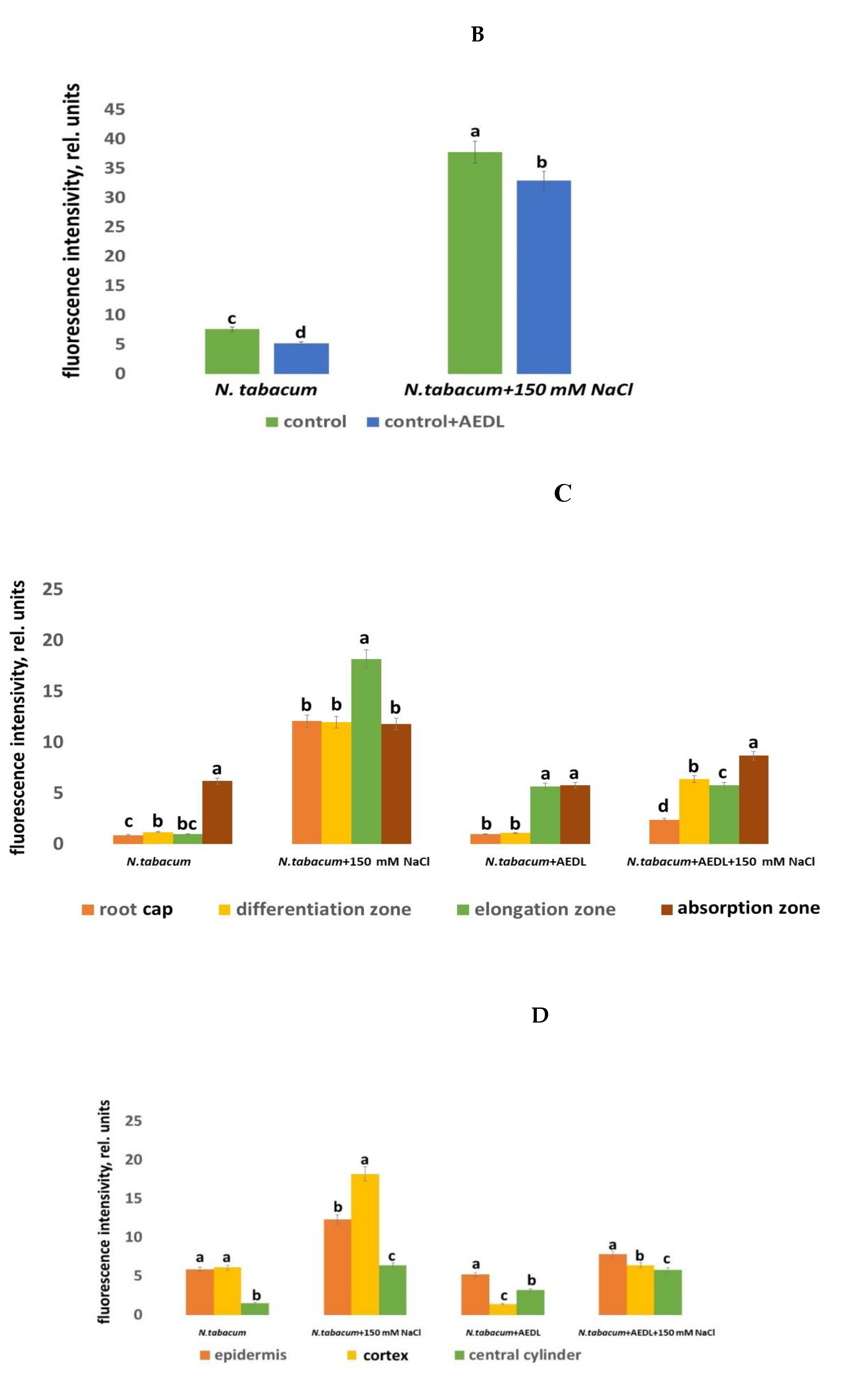
Figure 4.
Expression of MnSOD and Cu/ZnSOD genes in N. tabacum in roots (1) and leaves (2), grown in different conditions A,C – control; B,D – 150 mM NaCl. Significant differences were defined as p<0.05.
Figure 4.
Expression of MnSOD and Cu/ZnSOD genes in N. tabacum in roots (1) and leaves (2), grown in different conditions A,C – control; B,D – 150 mM NaCl. Significant differences were defined as p<0.05.
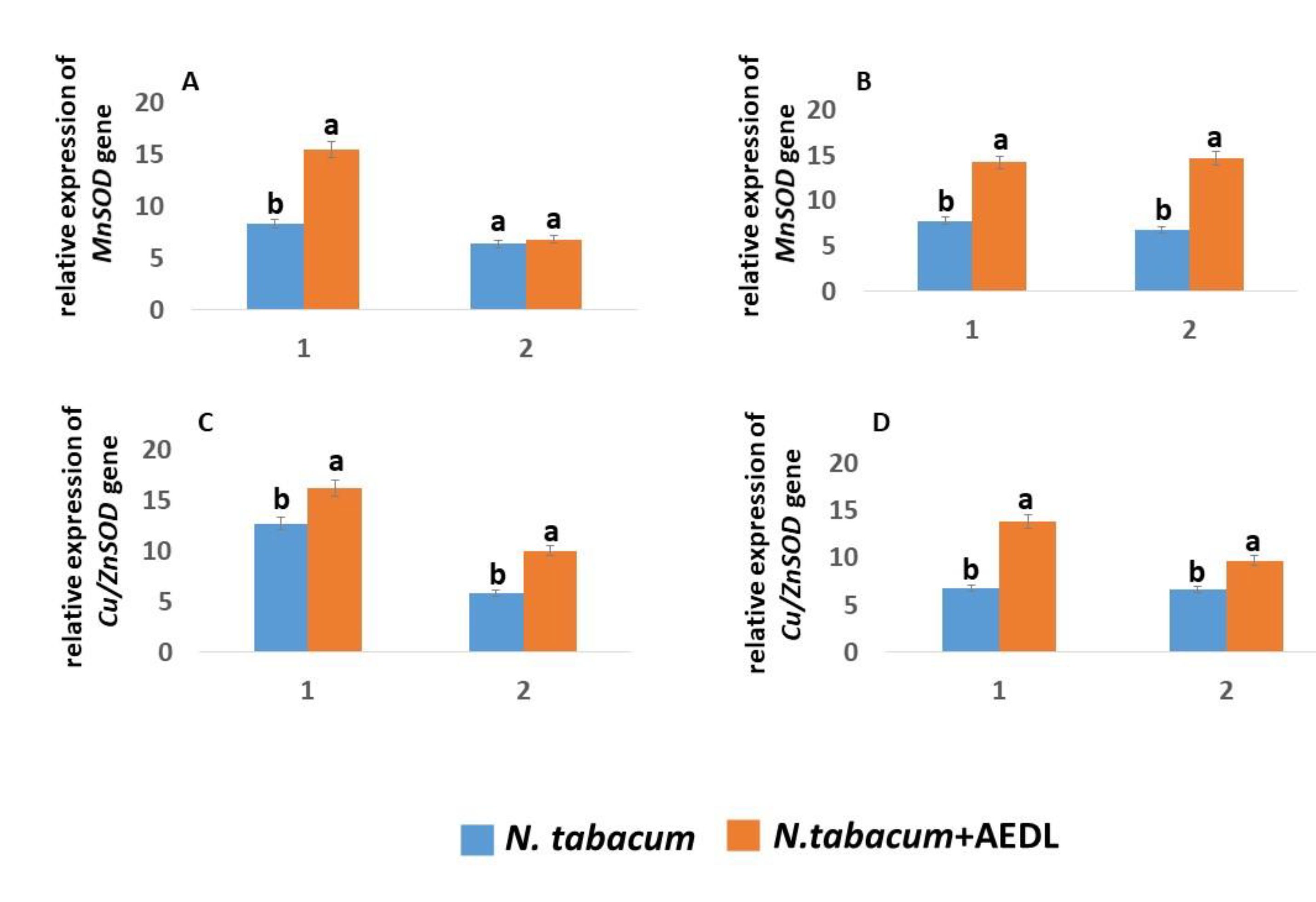
Figure 5.
Expression of the GSH1 and GSH2 genes in N. tabacum in roots (1) and leaves (2), grown in different conditions A,C – control; B,D – 150 mM NaCl. Significant differences were defined as p<0.05.
Figure 5.
Expression of the GSH1 and GSH2 genes in N. tabacum in roots (1) and leaves (2), grown in different conditions A,C – control; B,D – 150 mM NaCl. Significant differences were defined as p<0.05.
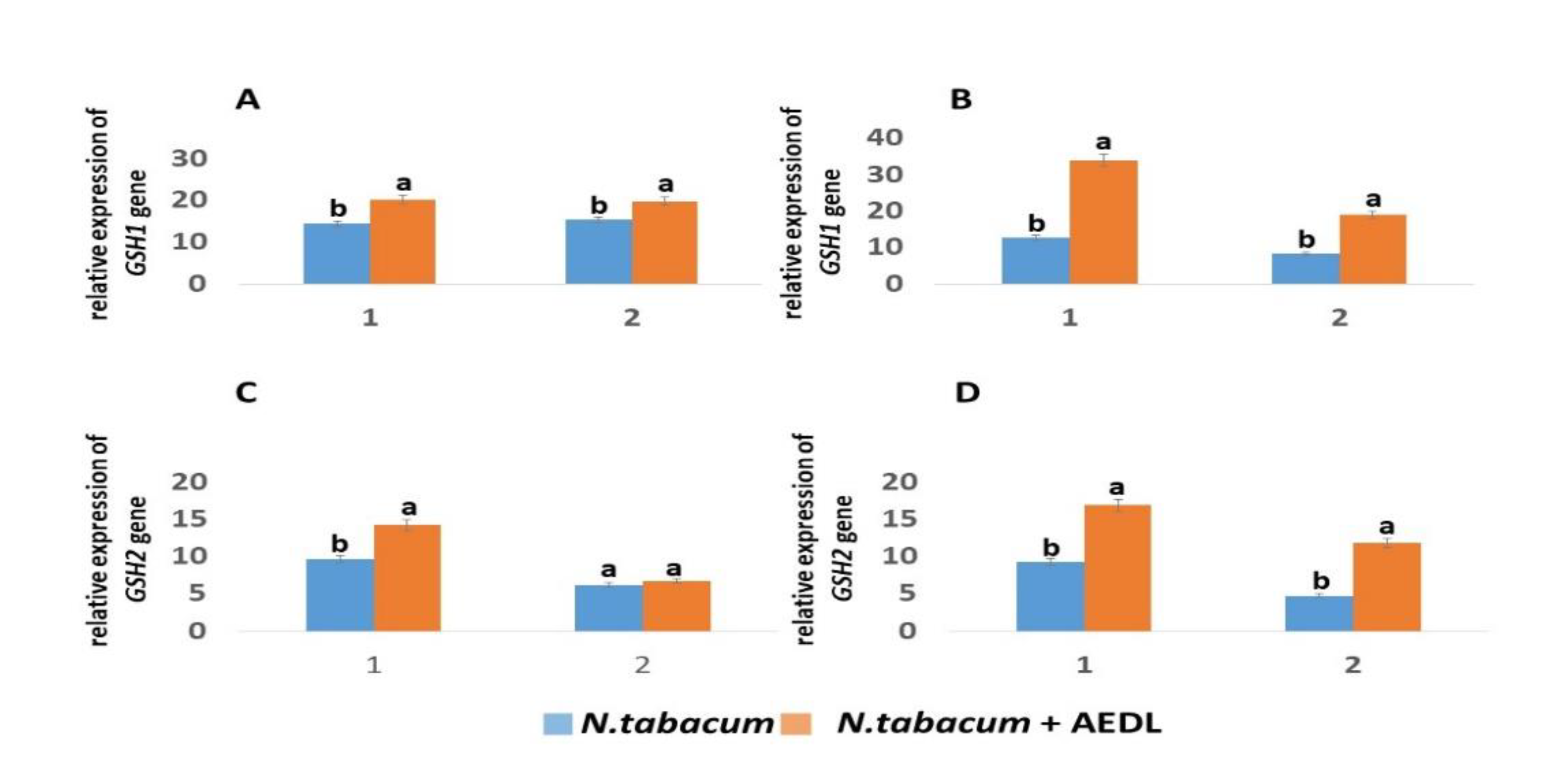
Figure 5.
Expression of the GR and GST genes in N. tabacum in roots (1) and leaves (2), grown in different conditions A,C – control; B,D – 150 mM NaCl. Significant differences were defined as p<0.05.
Figure 5.
Expression of the GR and GST genes in N. tabacum in roots (1) and leaves (2), grown in different conditions A,C – control; B,D – 150 mM NaCl. Significant differences were defined as p<0.05.
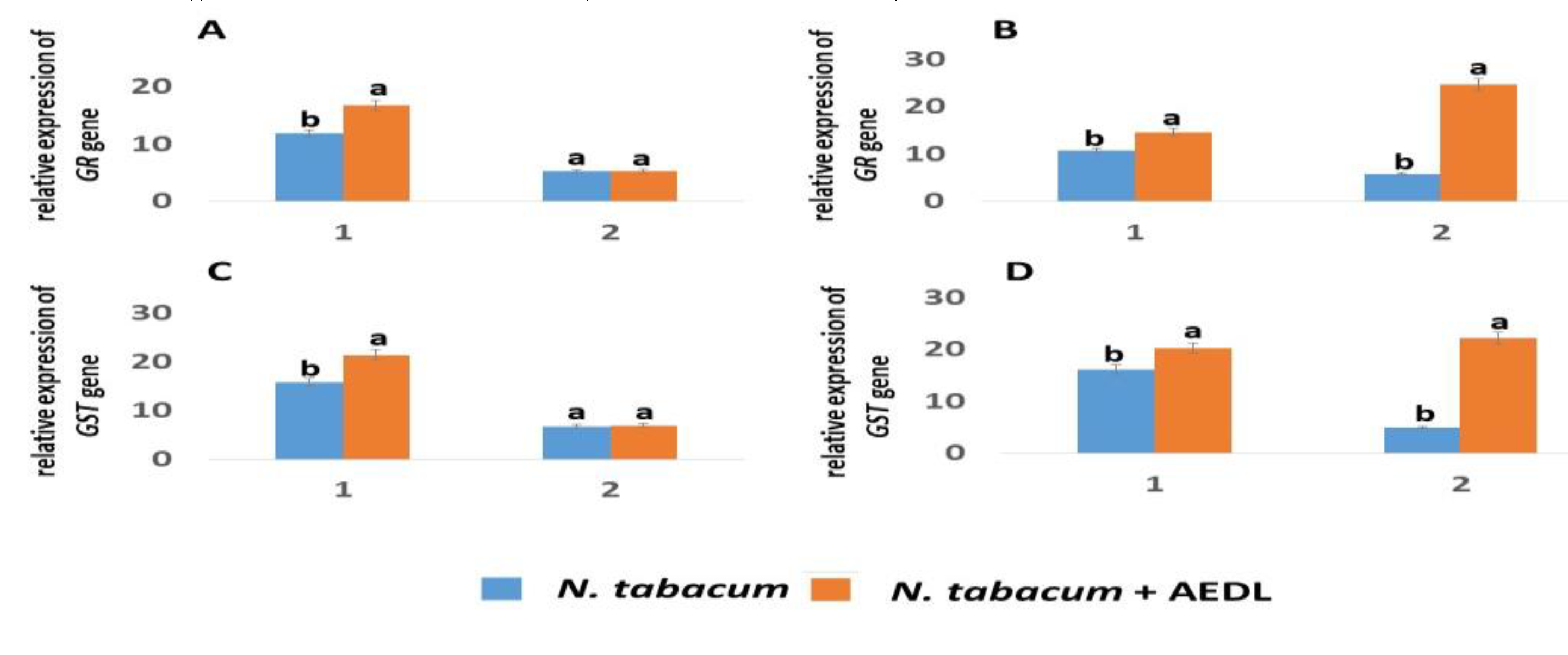
Figure 6.
Peptide AEDL and GSH form a negative feedback stem cell fate loop. .
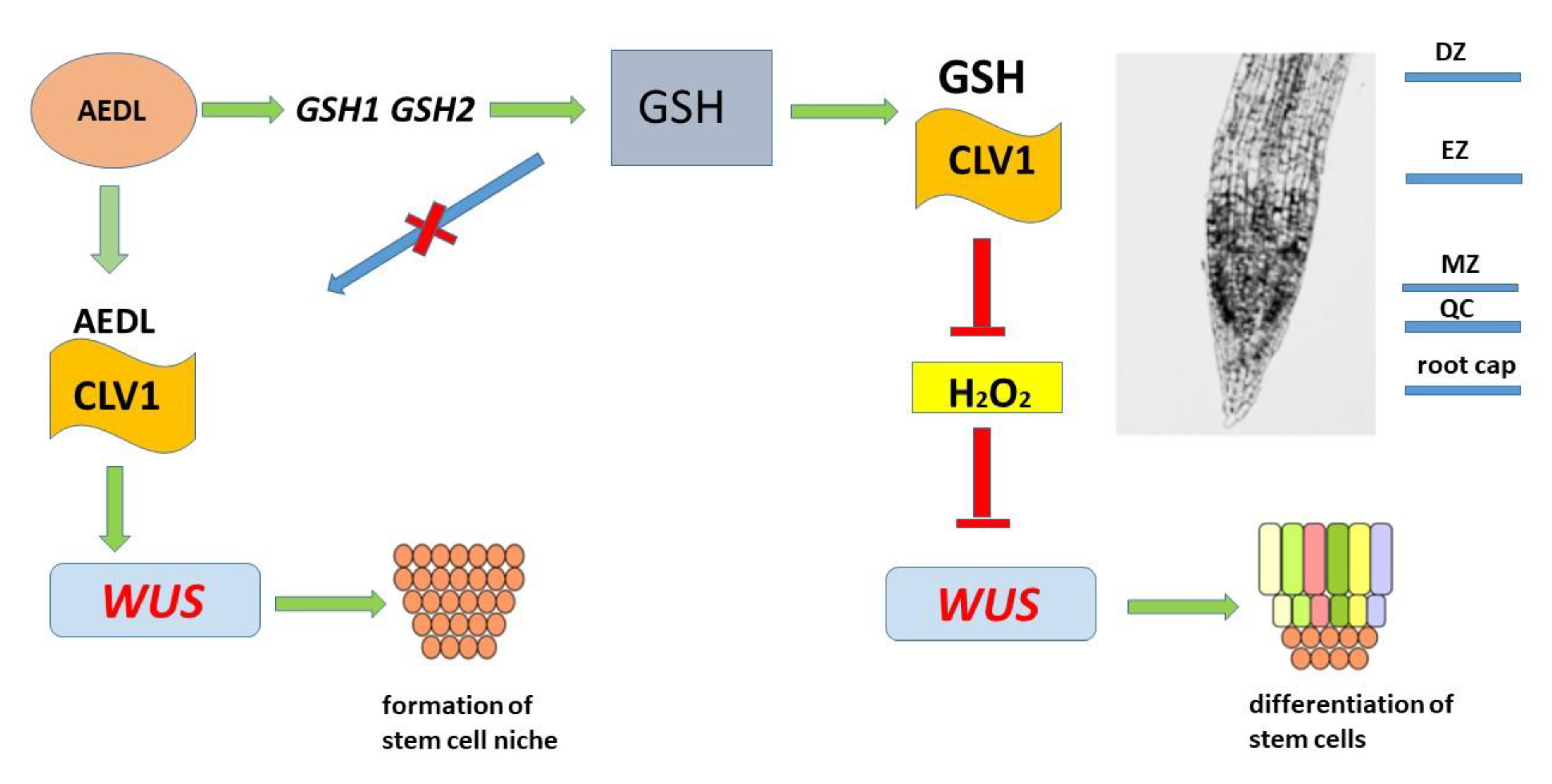

Table 1.
H2O2 content in N. tabacum, grown in different conditions.
| Varieties growth condition H2O2 mkg/g | |||||||
| N.tabacum control | 12.1±0.60 с | ||||||
| root | +AEDL | 8.93±0.45 d | |||||
| +NaCl | 17.8±0.89 a | ||||||
| +AEDL+NACL | 16.1±0.80 b | ||||||
| N.tabacum | control | 2.72±0.14 c | |||||
| shoot | +AEDL | 2.51±0.12 d | |||||
| +NaCl | 7.98±0.40 a | ||||||
| +AEDL+NACL | 5.23±0.26 b | ||||||
Data were expressed as mean ± standard deviation (SD; n = 5), and significant differences were determined p < 0.05.
Table 2.
AOA in Nicotiana tabacum, grown in different conditions.
| Varieties growth condition | AOA | |||||||
| %, inhibitory | ||||||||
| N.tabacum control | 37.91±1.89 b | |||||||
| root | +AEDL | 53.70±2.68 a | ||||||
| +NaCl | 11.90±0.59 c | |||||||
| +AEDL+NACL | 38.10±1.90 b | |||||||
| N.tabacum | control | 23.13±1.16 b | ||||||
| shoot | +AEDL | 28.40±1.42 a | ||||||
| +NaCl | 14.71±0.73 d | |||||||
| +AEDL+NACL | 21.96±1,10 c |
Data were expressed as mean ± standard deviation (SD; n =5), and significant differences were determined p < 0.05.
Table 3.
GSH content in N. tabacum grown in different conditions.
| varieties growth condition GSH, mM/g | |||||||
| N.tabacum control | 0.80±0.04 c | ||||||
| root | +AEDL | 2.59±0.13 a | |||||
| +NaCl | 0.68±0.03 d | ||||||
| +AEDL+NACL | 1.38±0.07 b | ||||||
| N.tabacum | control | 0.28±0.01 b | |||||
| shoot | +AEDL | 0.38±0.,02 a | |||||
| +NaCl | 0.13±0.01 c | ||||||
| +AEDL+NACL | 0.30±0.01 b | ||||||
Data were expressed as mean ± standard deviation (SD; n = 5), and significant differences were determined p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



