
Submitted:
25 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
The invasion of the golden mussel has resulted in considerable environmental and socioeconomic alterations, which present a considerable threat to the native biodiversity and sustainability of the region. It is of the utmost importance to gain an understanding of the distribution and biological characteristics of this Asian mussel, as well as its interaction with human activities, in order to develop effective strategies for mitigating and preventing its further spread. This study examines the dispersal route and incidence of golden mussels, tracing their movement from initial populations in Argentina to their arrival in the São Francisco River Basin (SFR). The presence of the mussel was confirmed through an integrative assessment, which included shell taxonomic analyses and mitochondrial DNA signatures. This identified populations located 7.5 km from the river's mouth, in proximity to the Atlantic Ocean, in areas such as shrimp farms, artisanal ports, and marinas. The analysis of mitochondrial DNA revealed the presence of South American-specific and shared ancestral haplotypes in the SFR, Grande River, and Argentina. These findings indicate that intracontinental colonisation towards the northeast of South America originated from Asian populations that entered South America via Argentina. The absence of Asian-specific signatures in the RSF, combined with its geomorphological structure, which is unsuitable for large ports or transoceanic vessels, supports the hypothesis of intercontinental dispersal of Limnoperna fortunei.
Keywords:
bioinvasion
; exotic invasive species
; watersheds
; human facilitation
; intercontinental dispersal
; bridgehead effects
; mitochondrial DNA signatures
; invasion genetics
; gene flow
1. Introduction
When introduced to a new area, mussels can become an environmental issue [1,2]. These species are often transported via ballast water [3,4], and their introduction can lead to the extinction of local species [5]. For instance, Mytella charruana (d’Orbigny, 1842) = Mytella strigata (Hanley, 1843) has been found in estuarine environments in North America and Asia [6,7,8], while Limnoperna fortunei (Dunker, 1857) has been found in freshwater environments in South America (SA) [9,10]. According to [10], Brazil reportedly spent at least USD 105.53 billion over 35 years (1984–2019) on mitigation, with an average annual expenditure of USD 3.02 (±9.8) billion. Invaders caused damages and losses amounting to USD 104.33 billion, while stakeholders invested only USD 1.19 billion in their prevention, control, or eradication [10].
Limnoperna fortunei, also known as the golden mussel, is native to China and Southeastern Asia and is an invasive species in different ecosystems worldwide [11,12,13]. It can be found in mussel banks (juveniles and adults) with highly dense populations [14,15] adhering to natural and artificial substrates [16,17] in environments presenting low salinity (<3 PSU) water [18,19,20,21] and temperatures ranging from 0 °C to 33 °C [11,22,23]. Its sexual maturation occurs at 3 to 4 months when the shell length of these individuals reaches 6 mm [24,25].
The dispersion and introduction of the golden mussel are often linked to pelagic larvae in ballast water discharged by ships in SA in 1990 [12]. The presence of L. fortunei in SA has resulted in economic and environmental losses [26,27]. Brazil’s expenses totaled USD 9.97 million and were solely related to the control, damage repair, prevention, and research on L. fortunei [10]. The Itaipu Hydroelectric Power Plant (HPP) is an example of this impact, stopping operation for three days to clean golden mussels from the machinery, resulting in a loss of approximately USD 750,000 [28].
Human activities are a significant factor in the dispersion of this species [29,30,31]. The proliferation of L. fortunei severely affects drainage systems and flooded lake reservoirs used for power generation [32] and fishery production [17]. The species is successfully dispersed upstream in South American rivers so frequently that its current frontier lies on the margin of the Amazon basin [33]. It has already been identified in the São Francisco River (SFR) basin [17,34] and a large portion of the Rio de La Plata basin [35]. The species advances an average of 240 km/year upstream in the middle and lower Paraná River [36] and 30 km/year in the Uruguay River basin [37]. The golden mussel has been shown to affect biological diversity, leading to imbalances in the food chain and alterations in the structure and dynamics of species living in limnic environments [32,38]. Darrigran et al. [24] found that the impact of golden mussels on native populations is due to their attachment to the structures of other organisms, such as mollusks and crustaceans.
Shell morphology (conchology) and molecular tools are currently utilized to identify golden mussels [39,40,41]. Molecular tools have become increasingly crucial in bioinvasion studies [42]. They aid mitigation efforts and provide baseline data on the structure of golden mussel populations [28,40,43,44,45,46,47,48]. Research on the population structure of the golden mussel indicates that the initial L. fortunei propagules were introduced to Argentina and Brazil via ballast water from Japan and Korea [44,45]. To analyze the evolutionary processes related to each animal model, Cox1 barcode haplotypes are necessary for population genetics [49,50,51], and fragmentation standards of 600 to 800 bp should be maintained [50]. The absence of these conditions can lead to the emergence of artifacts that affect the comparison or identification of haplotypes within a given species [50]. Haplotypes of L. fortunei Cox1 barcode have been identified in Asia and SA [28,43,44,45,46,47,48]. Reports indicate that exclusive haplotypes derived from Asia [48] share ancestral haplotypes (Lfm03) from Asia. Additionally, exclusive haplotypes (Lfm04) from SA populations have been observed [28,44,45,46].
This study aims to (1) identify and discuss an intercontinental dispersal route for golden mussels, tracing their movement from initial populations in Argentina to their arrival in the estuarine region of the São Francisco River; (2) use integrated conchological and molecular approaches (DNA footprints) to identify populations established in this newly invaded region; and (3) discuss and list the human factors facilitating their spread in this river basin in South America.
2. Materials and Methods
2.1. São Francisco River
The São Francisco River (SFR) basin spans 2863 km and covers 639,219 km2, equivalent to 7.5% of Brazil’s national territory (Figure 1), and is located in the northeast area of South America (SA). The basin comprises four physiographic sectors: Upper, Middle, Sub-Middle, and Lower. Junqueira [52] describes the end portion of the basin as the Lower São Francisco River (LSFR). In the early 1960s, the course of the Piumhi River, a tributary of the Grande River (GR) at the head of the Paraná River (PR) basin, was reversed to the Upper sector of the SFR basin [53]. The LSFR basin covers a total area of 30,337 km2 and extends for 274 km (5% of the basin). It includes the area between the power plant complex in Paulo Afonso County (Bahia State) and its mouth in the Atlantic Ocean (see Text S1, Figures S1 and S2). Studies have shown that coastal stretches along the LSFR, specifically the SFR Delta, are undergoing erosion (see Text S1). The LSFR has a longitudinal gradient and fauna closer to an estuarine environment than an estuary. The reduction and regulation of the flow resulting from the various dams along the course of the SFR, especially the Xingó dam located just 180 km from the mouth, probably caused the salt wedge intrusion [54]. The alterations to the ichthyofauna assessed in [54] illustrate the changes caused by river damming in this region. Notably, this region has no reports of intercontinental port activity (see Text S1).
2.2. Collected Specimens
Golden mussels were first observed in the LSFR during ichthyological studies [54]. The mollusks were found in April 2017 and November 2018 as bycatch collected through manual fish trawling (dimension: 30 m × 2.8 m; 5 mm mesh between nodes) carried out every month from May 2017 to April 2018 in shallow areas of the LSFR during spring tide. Apnea diving was subsequently used in an active search near specific points at the mouth of the SFR in the counties of Brejo Grande and Ilha das Flores (Figure 1 and Table S1).
Specimens were collected at these locations for molecular analysis on natural and artificial substrates. The points searched were at landing ports for artisanal fishing and marinas (see Figures S2 and S3). The rate of advance (RA) of the golden mussel in the watersheds of SA was calculated using the distance obtained from Google Earth and the number of months between mussel observation dates described in publications, multiplied by 12. The first month of the year is considered to be the month not reported in the references (see Table S1). One hundred specimens were manually collected at each point where the populations had established themselves. Salinity was measured using an Atago® manual refractometer (Ribeirão Preto - SP, Brazil). The specimens were fixed in ethanol and refrigerated at -20 °C for preservation. Molecular identification was performed using adductor muscle tissues.
2.3. Identification and Biometry
Ninety specimens (VIF—Village Ilha das Flores: 30 specimens; BGM—Brejo Grande Marine: 30 specimens; MRS—Mouth of the São Francisco River: 30 specimens) were identified using the conchological criteria suggested in [41]. Pictures of the collected specimens were taken using a Leica® M205 C stereomicroscope (Heerbrugg - Kanton St. Gallen, Switzerland) equipped with an MC170 HD real-time digital camera (Leica®) and LAS 4.8.0 software (Leica®). The specimen size (umbo-ventral axis) was measured using a digital caliper (see Figure S3), modified from [55]. The relative frequencies of the class (shell size) of the three population samples of L. fortunei were analyzed according to Sturges’s rule in the Lower São Francisco River basin. In all, 56 specimens were genetically identified from the three localities sampled (VIF—Village Ilha das Flores: 18 specimens; BGM—Brejo Grande Marine: 19 specimens; MRS—Mouth of the São Francisco River: 19 specimens).
2.4. Identification of DNA Footprints and Dispersal Route
A subset of fifty-six specimens was used for DNA total extraction using the phenol-chloroform protocol [56] of the specimens identified based on morphology. The mitochondrial gene Cox1 fragment was amplified using the primers [58] described. Polymerase chain reaction (PCR) was performed according to a protocol defined in [49]. The PCR product was purified, and all specimen fragments (both strands) were sequenced using the automatic ABI PRISM TM 377 DNA Sequencer by Macrogen (www.Macrogen.com; Seoul, North Korea). After sequencing, all electropherograms were checked manually in MEGA 10.2.2 software [101]. Finally, consensus strings were generated; the location in the mitogenome (KP756905) of L. fortunei was annotated in MEGA 10.2.2 software [103] and compared with the sequences available in GenBank (www.ncbi.nlm.nih.gov/nuccore/). The BLASTn tool (www.ncbi.nlm.nih.gov/BLAST/) was used to confirm the identity of haplotypes (Figure 2, see Table S2), according to [44,47]. Genetic distances were estimated using the Kimura 2-parameter evolutionary model (K2P) [103]. A neighbor-joining (NJ) tree [105] was infered using this model for graphical representation. Clade support was assessed by 1000 bootstrap pseudoreplicates using MEGA 10.2.2 [105] following [109]. Given the criticism of using K2P as a surrogate model [107] proposed in [109], we used p-distance. The best molecular evolution model (HKY + G) with Akaike Information Criterion corrected (AICc) was infered using jModelTest 1.0 software [109]. This analysis was performed on a web interface (https://bioinfo.mnhn.fr/abi/public/abgd/) using the simple distance model (p-distance) with a gap 1.5. The maximum intraspecific divergence (P) value was set between 0.001 and 0.1, and the number of recursive steps is the default (10). MrBayes v.3.2.6 [111] was used to construct the tree. As the models’ differences were minimal regarding estimated genetic divergence and tree topology, we represented the HKY + G molecular evolution model. Visualization and editing of clades were performed in FigTree v1.4.2 [111]. A haplotype network was reconstructed using PopART 1.7 [112] with the TSC method [113] to elucidate relationships between species and populations.
Assessment of the dispersal route of the golden mussel was conducted based on studies that provided or published Cox1 sequences, summarizing a total of 2160 specimens (735 from Asia and 1425 from SA). By comparing the available sequences and the haplotypes described in the literature, a standardization of the nomenclature was carried out, as presented in Table S2. Thus, the analysis of the golden mussel’s dispersal route was conducted by comparing the presence of haplotypes and the collection sites of the populations (see Table S2).
3. Results
3.1. Mussel Occurrence
The occurrence of golden mussels was reported at two different times (April 2017 and November 2018) at three locations in the LSFR basin (VIF—Village Ilha das Flores, BGM—Brejo Grande Marine; MRS—Mouth of the São Francisco River). These sites were the most easterly in SA. The occurrence of the mussel in the LSFR extended its distribution by 500 km in the largest exclusively Brazilian hydrographic basin. The occurrence of golden mussels in estuaries near (7.5 km) the SFR estuary had never been reported in northeastern SA (Figure 1). This expanded distribution resulted in a species advance of approximately 375 km/year downstream of the LSFR (see Table S1). Living mussels were collected from low-salinity collection sites such as estuarine (MRS: 4PSU) and freshwater (VIF and BGM) beaches associated with artisanal fishermen, landing ports, and drainage channels used for fish and white shrimp Litopeneus vanammei farming (see Figures S1 and S2). Live bivalves were collected from the underwater areas of marinas, artisanal fishing ports, and drainage channels used for fish farming. They were attached to unconsolidated (coarse sand and gravel), artificial (bricks and cement fragments), and naturally consolidated substrates (rocks). Around the natural consolidated substrate, it was possible to see golden mussels associated with native estuarine species (Crassotrea sp. and Vitta virginea) able to survive in low-salinity estuarine environments associated with river outlets. In Ilha das Flores (VIF), clusters of mussels were recorded, some of which were collected and observed next to aquatic macrophytes (see Figures S1–S3). Population densities were not assessed at these sites.
3.2. Mussel Identification and DNA Footprints
The shells of the collected specimens were formed by two equal valves with a triangular contour and an elongated base. The subterminal umbo; smooth, simple joint ligament; and very thin, straight, and toothless hinge (see Figure S3) confirmed the identification of L. fortunei in [39] and [41]. Voucher specimens were deposited in the mollusk collections of the Universidade Estadual de Ponta Grossa in the Coleção de Moluscos (UEPG-CMo: CMo 1120; CMo 1121; CMo 1122) and the Museu de Ciências Naturais of the Universidade Federal do Paraná (MCN-UFPR: MCN.Z 777). Individuals with shell lengths ranging from 5.25 to 23.79 mm (mean 9.81 ± 4.57 mm) were observed at the site (Table 1) in VIF, where it was possible to observe the presence of the largest individuals, indicating banks with adult specimens and an older and better-established population compared to the other two populations (BGM and MSR, see Figure S2). These two populations are located closer to the São Francisco River Delta.
Here, we present for the first time the compilation and annotation of all cytochrome oxidase (Cox1) gene fragment sequences in a mitogenome of L. fortunei (Figure 2) and the comparison of golden mussel Cox1 haplotype nomenclatures. Partial sequences of Cox1 from the collected samples (Figure 3, see Table S2) resulted in length alignments of 457 (1 sequence), 530 (1 sequence), and 555 (54 sequences) bp, located between positions 8761 and 9315 of the L. fortunei mitogenome (KP756905). Four of the forty-four haplotypes identified (in 2160 specimens of the 88 populations) in L. fortunei (Table 2) had 99.70% to 100% similarity in the three newly studied populations (VIF: Lfm02, Lfm03, and Lfm4; BGM: Lfm02, Lfm03, and Lfm4; MRS: Lfm01, Lfm02, Lfm03, and Lfm4) and were deposited in GenBank and BOLDSystem (references XX000000.0 to XX000000.0). Of the haplotypes identified (Table 2), two are exclusive to South America (Lfm01 and Lfm4), while two (Lfm02, and Lfm03) are found in South America (Paraná and São Francisco basins) and Asia (Taiwan, Japan, and China).
4. Discussion
4.1. Cox1 Barcode Annotation and Haplotypes in the Golden Mussel
The annotation of Cox1 fragments in the mitogenome of L. fortunei indicated that their position affects haplotype identification. The position and length of these Cox1 fragments in golden mussels are not the same as the polymorphic regions observed in haplotypes described in this gene in [44] and [47]. The sequences described in the literature and deposited in Cox1 gene databases range from 382 to 1509 bp in length and are located in different positions in the L. fortunei mitogenome (Figure 2). To avoid artifacts in the analysis of the dispersal route, it was essential to verify whether there is overlap across all Cox1 barcode loci when describing a haplotype for the specimen referenced in the studies. However, some polymorphic sites that could indicate a specific haplotype were not evaluated (Figure 2 and Figure 3, see Table S2). Therefore, it was crucial to emphasize the importance of accessing reference sequences [41,44,45,46,47,59,65] to analyze and identify L. fortunei haplotypes. If a new haplotype is described, it must be made available to the scientific community.
4.2. Identified Footprints and Possible Origins
Integrated molecular and morphological identification played a crucial role in confirming the L. fortunei in estuarine and freshwater regions of the LSFR. The exclusive haplotype found in this site (Lfm01 and Lfm04) is consistent with SA populations [28,45], providing evidence of the species’ successful invasion of new hydrographic environments. In a previous study [28], it was observed that the exclusive haplotype (Lfm39) was present in SA populations in the SFR and PR basins (Figure 3, see Tables S2 and S3).
Native distribution of L. fortunei is probably limited to the Pearl River basin in China [11,13]. This invasive species can be found in several Asian (Japan and Korea) and SA regions [30,45,47,66]. Ghabooli et al. [45] analyzed the genetic structure of 24 L. fortunei populations (10 from Asia and 14 from SA) living in invaded and native areas based on Cox1 gene sequences (~510 bp), and eight microsatellite markers indicated that bivalve populations living in Asia had greater diversity (23 haplotypes) than those living in SA (18 haplotypes), suggesting that Asia has been subject to a more significant number of introduction events for invasive populations. The authors also reported the fine genetic structure of L. fortunei on both continents, suggesting strong post-introduction selection and stochastic events inherent to the species’ biology. These events contribute to its population structure and the occurrence of nine known Cox1 haplotypes exclusive to SA (Lfm01, Lfm04, Lfm07, Lfm10, Lfm13, Lfm16-18, Lfm39). In the LSFR, two haplotypes exclusive to SA (Lfm01, Lfm04) were identified (Figure 3, see Table S2).
The Cox1 haplotypes (457 to 555 bp) observed in specimens collected from the LSFR (Table 2), Sobradinho, Barra Bonita HPP, Porto Primavera HPP, and Porto Hidroviário Bariri [28] exhibit features that are characteristic of populations from SA (Lfm02, Lfm03, Lfm11, Lfm15, Lfm36, and Lfm38), Taiwan, Korea, Japan, and China [28,44,45,46,47]. Based on the haplotypes analyzed in the LSFR, it can be inferred that the observed population in VIF (Lfm02, Lfm03, and Lfm04), BGM (Lfm02, Lfm03, and Lfm04), and MRS (Lfm01, Lfm02, Lfm03, and Lfm04) originated from populations located further south in the American continent. Ferreira et al. [67] observed similar conditions when comparing mussel populations from five different Brazilian reservoirs using the double digest restriction-site-associated DNA sequencing (ddRAD-seq) protocol for the golden mussel. These authors indicated the connectivity between basins and absent geographic structure [67], in agreement with the data presented here for the Cox1 gene. Exclusive SA haplotypes in the SFR and PR basins support the hypothesis of upward colonization toward hydrographic basins in northeastern Brazil (Figure 1, Table 2, and see Table S2). It is essential to emphasize the lack of port areas that are navigable by large vessels at the mouth of the SFR (Figure 1; [54,68]). The presence of the golden mussel after thirty years (from 1991 to 2020) in Argentina confirms the theory of the intracontinental spread of the mussel in SA [20]. This fact virtually excludes golden mussel reintroduction due to maritime cabotage (Figure 1), as supported in [69,70] and [35] (Figure 1, see Figure S2). The absence of golden mussels in coastal watersheds in southeast and northeast Brazil port areas, as confirmed in [30] and [28], supports the theory of intracontinental dispersal.
Furthermore, the specimens genotyped from the LSFR have haplotypes Lfm02 and Lfm03 (Figure 1, Figure 3, see Table S2). These data support the hypothesis that Asian specimens [71] initially invaded SA from Argentina [12,14]. The dispersion process of the golden mussel is indicated by the flow of the targeted gene of haplotypes from Asia to SA (Table 2, see Table S2). There were no exclusive haplotypes from SA in Asia (Table 2), as reported in [28,44,45,46,47,48]. This may be related to the export of grains such as soybeans and sugar from SA to Asia [72]. According to [73], some grain-loaded ships do not require ballast water to achieve stability. However, vessels arriving from Asia require ballast water to achieve stability; this feature facilitates the directional genetic flow from Asia to SA (Figure 1, see Table S2). Studies have shown that M. strigata, another invasive mussel worldwide, has invaded different areas of America and Asia [7,8]. Notably, L. fortunei and M. strigata can coexist in low salinities [19,74]. However, it is essential to note that only the golden mussel exhibits a directional flow from its native to the invaded area, as demonstrated by various studies [7,8,44,45,46].
4.3. An Intracontinental Route to Northeastern Brazil
The reversal of the course of the Piumhi River (a tributary of the GR at the head of the PR basin) into the SFR basin in the early 1960s [53] is thought to have facilitated the invasion of the golden mussel, as well as the connectivity of these basins (Figure 1). Invasion risk factors were observed in the studied environment (see Figure S1). This anthropogenic change, which eliminated the natural barrier between these basins, allowed the introduction of species and the genetic flow of fish (Hypostomus regani and Astyanax “bimaculatus group”) from the PR basin to the SFR basin [75,76]. The Capitólio dam interfaces the two river basins [53]. The proximity of these two basins, changes in water flow (Moreira Filho, personal communication), and the transit of ships between the basins may have contributed to the invasion of the golden mussel toward the northeastern region of SA. The tributaries of the GR are not navigable by commercial vessels, despite other dispersal vectors such as small recreational and fishing boats, fish farming, and the use of sand recovered from mussel-infested areas, such as the Tietê River and other sites in the PR basin [35]. Recently, the presence of the golden mussel in the communication channels of these two interconnected hydrographic basins has been observed in the SFR [34].
In addition, the presence of haplotypes in SA (Lfm01 to Lfm04, Lfm15, and Lfm38) close to the Piumhi River (observed at two sites in the GR tributary in [28]) indicates the likely connection between the two basins (Figure 1, Table 2, see Table S2). This evidence reinforces the concerns of the expert committee (participants in the National Plan for the Prevention, Control, and Monitoring of the Golden Mussel in Brazil) about the role of water transport from the SFR to the Tocantins River basins as a vector for the invasion of golden mussel larvae and adults [27]. There may be ways of introducing these mussels into the SFR basin other than the likely dispersal jumps from southeastern to northeastern Brazil in freshwater fish transport tanks [34,67,77].
4.4. Golden Mussel and Human Activities in Northeastern Brazil
Invasion risk factors were observed in the studied environment (see Figure S2C and S3); therefore, they need to be considered at this point to implement plans focused on mitigating the occurrence of this invasive species in the LSFR. Monitoring and early detection of invasive species are prerequisites for developing bioinvasion prevention and management plans [78]. The mussel species live in different freshwater environments such as streams, rivers, dams, lakes, coastal lagoons, lagoons in low-salinity scenarios, and river deltas [27,79]. Thus, the risk posed by each dispersal vector observed in the watershed is related to economic activities [30], social arrangements, local practices, and habits, which may differ between hydrographic systems and result in different invasion routes or corridors [80].
Golden mussel occurrence in inland regions closer to the coast suggests the vectors of recurrent dispersal in northeastern Brazil. Similar to the process observed in Japan [47], the introduction of mussels into northeastern Brazil is also associated with aquaculture [81]. Furlan-Murari et al. [82] observed golden mussel banks in tilapia (Oreochromis niloticus) aquaculture tank nets used in southern/southeastern Brazilian reservoirs. Curimatã (Prochilodus argenteus), tambaqui (Colossoma macropomum), tilapia, and common carp (Ciprinus carpio) stand out among the fish species farmed in freshwater tanks dug in the LSFR. These tanks enable the subsistence of about 2000 regional families [83].
The LSFR presented small golden mussel beds in low-salinity water channels associated with ponds dug to cultivate the gray shrimp species L. vanammei (see Figures S2 and S3). The northeast region represents the central Brazilian pole for gray shrimp farming in marine and freshwater environments, thus contributing to the social and economic development of the region [84,85]. The presence of L. fortunei can affect this production chain, as mussel fouling can affect pumping systems, reduce water flow in tanks, and intensify disease transmission processes. Campos et al. [86] developed a model to predict the distribution of golden mussels based on air temperature and rainfall. The scenarios generated have shown a high potential for L. fortunei to invade aquatic environments in Central America, North America, Europe, Africa, and Oceania, as well as the expansion of golden mussel occurrence areas in Asia and SA. According to the authors, the Amazon, Tocantins, Araguaia, and SFR river basins have tremendous potential for invasion in Brazil. Barbosa et al. [17] confirmed the occurrence of golden mussels in 2015 at two sites near the Sobradinho Hydroelectric Plant and another at the crossing (transposition system) of the SFR (Table 1). Similar observations were made in the Mato da Onça Reserve, close to the Xingó Hydroelectric Plant [87,88]. The current identification of golden mussel populations in freshwater (VIF and BGM) and estuarine (MRS) areas of the LSFR (average salinity 14.67 PSU, with a maximum of 26.43 PSU and a minimum of 3.59 PSU; [54,89]) in 2017 confirmed the predictions of [86] regarding the expanded distribution of golden mussels in the LSFR basin. Similar conditions have been observed for Lagoa dos Patos and the La Plata River [15,20]. However, the results of the present study are 10 years ahead of the projections made by some predictive models [77,86].
The dispersal rate of the golden mussel observed in the SFR basin is higher than that recorded for the Paraná basin (see Table S1), which may be explained by the initial colonization of the golden mussel in the upper SFR. The three populations identified in the LSFR showed a mean shell length of reproductive-age specimens (Table 1, see Figure S2). In SA, sexual maturity varies seasonally, starting at a 5–6 mm shell length in winter-spring and 7–10 mm in autumn [24]. The downstream dispersal of planktonic larvae [38] or juvenile/adult individuals associated with floating substrates, such as macrophytes, observed in the LSFR ([87], Figure 1, see Figures S1–S3) is a favorable and opposite condition to the upstream distribution of this invasive species in other hydrographic basins, such as the Paraná-Paraguay system, reported in Japan [47].
5. Conclusions
The present study confirmed the presence of L. fortunei populations in different LSFR localities and their range’s rapid expansion. These findings indicate an intracontinental colonization route toward northeastern SA. The data analyzed indicated that the populations of golden mussels living in the LSFR originated from populations established in other SA locations; these originated from Asian populations that entered SA via Argentina and reached the study area through human vectors. It is therefore necessary to conduct further studies focusing on monitoring possible vectors of introduction and spread and the occurrence of golden mussels in river basins located in the north and northeast regions of Brazil. In addition, measures to educate and raise awareness (especially of water users) of the problem can allow for the most efficient management. The situation in the Tocantins and Araguaia River basins is similar to that observed in the SFR, and the presence of the gold mussel in these basins is a reality.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. Rate of advance (RA) of the golden mussel in watersheds of South America, and Table S2. Sampling details and haplotype codes for the mitochondrial cytochrome c oxidase subunit I (Cox1) gene of the golden mussel Limnoperna fortunei. https://docs.google.com/document/d/1_D9itXHi2z90mEK9pIrHiOShhEBKyzz1C4IPzUtqK24/edit; Figure S1. Example of factors that may have contributed to the dispersion of the golden mussel in the Lower São Francisco River basin, Figure S2. Limnoperna fortunei present in different substrates in the Lower São Francisco River basin, Figure S3. Biometry and Conchiology of Limnoperna fortunei obtained in the Lower São Francisco River, and https://docs.google.com/document/d/1RO7Wi9TukBv0LRz9M_zBxM7YEPNZi10EKGAjgDClqp0/edit; Text S1. The Lower São Francisco River (LSFR) region; Text S2. Methodology. https://docs.google.com/document/d/1lO3uAcnviMT2L-B1tkWBBDLil_BEUyO1JUUdLSinHKA/edit.
Author Contributions
Conceptualization, A.L.F.J., R.B.-N., S.W.C., and R.F.A.; methodology, A.L.F.J., R.B.-N., V.M.-R., and S.W.C.; software, A.L.F.J. and R.B.-N.; formal analysis, A.L.F.J. and R.B.-N.; resources, A.L.F.J., R.B.-N., V.M.-R., L.R., M.C.A., P.D.B., S.W.C., and R.F.A.; writing—original draft preparation, A.L.F.J. and R.B.-N.; writing—review and editing, V.M.-R., L.R., M.C.A., P.D.B. S.W.C., and R.F.A.; visualization, S.W.C. and R.F.A.; supervision, S.W.C. and R.F.A.; funding acquisition, M.C.A., S.W.C., and R.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
A.L.F.J. was awarded a Ph.D. scholarship by the Brazilian National Council for Scientific and Technological Development (CNPq—142155/2019-5). This research was funded by CNPq grant number 308748/2021-2, Fundação Araucária NAPI-Bioinformática grant number 033/2021 and INCT/ADAPTA grant number 465540/2014-7. A.L.F.J. received a grant from CNPq.
Institutional Review Board Statement
The study was conducted in accordance with Portaria ICMBio 748/2022 and approved by the Instituto Chico Mendes de Conservação da Biodiversidade through the Biodiversity Authorization and Information System (ICMBio/SISBIO): license number 6625. In Brazil, the animal study protocol does not apply to studies involving invertebrate animals; it is only required for studies involving vertebrate animals or humans.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data produced by the research are being submitted to public repositories.
Acknowledgments
To the entire technical, administrative, and managerial team of Ponta Grossa State University (UEPG) and the Foundation for Support of Institutional, Scientific, and Technological Development of UEPG (FAUEPG), and, in particular, to Karina Aparecida Soares, Acir da Cruz Camargo, Amélia Harmatiuk De Oliveira, Professor Giovani Marino Favero, and Sinvaldo Baglie. All individuals included in this section have consented to the acknowledgement.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- McKindsey, C.W.; Landry, T.; O’Beirn, F.X.; Davies, I.M. Bivalve aquaculture and exotic species: A review of ecological considerations and management issues. J. Shellfish Res. 2007, 26, 281–294. [CrossRef]
- Joyce, P.W.; Kregting, L.; Dick, J.T. Relative impacts of the invasive Pacific oyster, Crassostrea gigas, over the native blue mussel, Mytilus edulis, are mediated by flow velocity and food concentration. NeoBiota 2019, 45, 19–37. [CrossRef]
- Silva, J.S.V.; Fernandes, F.C.; Souza, R.C.C.L.; Larsen, K.T.S.; Danelon, O.M. (Eds.) Água de Lastro e Bioinvasão; Interciência: Rio de Janeiro, Brazil, 2004; pp. 33–38.
- Carlton, J.T. Marine Bioinvasions: The alteration of marine ecosystems by non-indigenous species. Oceanography 1996, 9, 36–43.
- Boltovskoy, D.; Correa, N.M.; Burlakova, L.E.; Karatayev, A.Y.; Thuesen, E.V.; Sylvester, F.; Paolucci, E.M. Traits and impacts of introduced species: A quantitative review of meta-analyses. Hydrobiologia 2020, 18, 2225–2258. [CrossRef]
- Stenyakina, A.; Walters, L.J.; Hoffman, E.A.; Calestani, C. Food Availability and Sex Reversal in Mytella charruana, an Introduced Bivalve in the Southeastern United States. Mol. Reprod. Dev. Inc. Gamete Res. 2010, 77, 222–230.
- Calazans, C.S.H.; Walters, L.J.; Fernandes, F.C.; Ferreira, C.E.; Hoffman, E.A. Genetic structure provides insights into the geographic origins and temporal change in the invasive charru mussel (Sururu) in the southeastern United States. PLoS ONE 2017, 12, e0180619. [CrossRef]
- Lim, J.Y.; Tay, T.S.; Lim, C.S.; Lee, S.S.; Teo, S.M.; Tan, K.S. Mytella strigata (Bivalvia: Mytilidae): An alien mussel recently introduced to Singapore and spreading rapidly. Molluscan Res. 2018, 38, 170–186.
- Giglio, M.L.; Dreher Mansur, M.C.; Damborenea, C.; Penchaszadeh, P.E.; Darrigran, G. Reproductive pattern of the aggressive invader Limnoperna fortunei (Bivalvia, Mytilidae) in South America. Invertebr. Reprod. Dev. 2016, 60, 175–184.
- Adelino, J.R.P.; Heringer, G.; Diagne, C.; Courchamp, F.; Faria, L.D.B.; Zenni, R.D. The economic costs of biological invasions in Brazil: A first assessment. NeoBiota 2021, 67, 349–374. [CrossRef]
- Morton, B. Some aspects of the biology and functional morphology of the organs of feeding and digestion of Limnoperna fortunei (Dunker) (Bivalvia: Mytilacea). Malacologia 1973, 12, 265–281.
- Pastorino, G.; Darrigran, G.; Martín, S.M.; Lunaschi, L. Limnoperna fortunei (Dunker, 1857) (Mytilidae), nuevo bivalvo invasor en aguas del Río de la Plata. Neotropica 1993, 39, 101–102.
- Xu, M. Distribution and spread of Limnoperna fortunei in China. In Limnoperna Fortunei; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 313–320.
- Mansur, M.C.D.; Santos, C.P.; Darrigran, G.; Heydrich, I.; Callil, C.T.; Cardoso, F.R. Primeiros dados quali-quantitativos do mexilhão-dourado, Limnoperna fortunei (Dunker), no Delta do Jacuí, no Lago Guaíba e na Laguna dos Patos, Rio Grande do Sul, Brasil e alguns aspectos de sua invasão no novo ambiente. Rev. Bras. Zool. 2003, 20, 75–84.
- Barbosa, F.G.; Melo, A.S. Modelo preditivo de sobrevivência do Mexilhão Dourado (Limnoperna fortunei) em relação a variações de salinidade na Laguna dos Patos, RS, Brasil. Biota Neotrop. 2009, 9, 407–412.
- Santos, F.J.M.; Peña, A.P.; Luz, V.L.F. Considerações biogeográficas sobre a herpetofauna do submédio e da foz do rio São Francisco, Brasil. Estudos 2008, 35, 57–78.
- Barbosa, N.P.U.; Silva, F.A.; De Oliveira, M.D.; Neto, M.A.S.; De Carvalho, M.D.; Cardoso, A.V. Limnoperna fortunei (Dunker, 1857) (Mollusca, Bivalvia, Mytilidae): First record in the São Francisco River basin, Brazil. Check List 2016, 12, 1846.
- Deaton, L.E.; Derby, J.G.; Subhedar, N.; Greenberg, M.J. Osmoregulation and salinity tolerance in two species of bivalve mollusc: Limnoperna fortunei and Mytilopsis leucophaeta. J. Exp. Mar. Biol. Ecol. 1989, 133, 67–79. [CrossRef]
- Angonesi, L.G.; Rosa, N.G.; Bemvenuti, C.E. Tolerance to salinities shocks of the invasive mussel Limnoperna fortunei under experimental conditions. Iheringia Série Zool. 2008, 98, 66–69.
- Sylvester, F.; Cataldo, D.H.; Notaro, C.; Boltovskoy, D. Fluctuating salinity improves survival of the invasive freshwater golden mussel at high salinity: Implications for the introduction of aquatic species through estuarine ports. Biol. Invasions 2013, 15, 1355–1366. [CrossRef]
- Lucía, M.; Darrigran, G.; Gutierrez Gregoric, D.E. The most problematic freshwater invasive species in South America, Limnoperna fortunei (Dunker, 1857), and its status after 30 years of invasion. Aquat. Sci. 2023, 85, 5. [CrossRef]
- Darrigran, G. Potential impact of filter-feeding invaders on temperate inland freshwater environments. Biol. Invasions 2002, 4, 145–156. [CrossRef]
- Karatayev, A.Y.; Boltovskoy, D.; Burlakova, L.E.; Padilla, D.K. Parallels and contrasts between Limnoperna fortunei and dreissena species. In Limnoperna Fortunei: The Ecology, Distribution and Control of a Swiftly Spreading Invasive Fouling Mussel; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 261–297.
- Darrigran, G.; Penchaszadeh, P.; Damborenea, C. The reproductive cycle of Limnoperna fortunei (Dunker, 1857) (Mytilidae) from a Neotropical Temperate Locality. J. Shellfish Res. 1999, 18, 361–365.
- Karatayev, A.Y.; Boltovskoy, D.; Padilla, D.K.; Burlakova, L.E. The invasive bivalves dreissena polymorpha and Limnoperna fortunei: Parallels, contrasts, potential spread and invasion impacts. J. Shellfish Res. 2007, 26, 205–213. [CrossRef]
- Darrigran, G.; Damborenea, C. A South American bioinvasion case history: Limnoperna fortunei (Dunker, 1857), the golden mussel. Amer. Malac. Bull. 2005, 20, 105–112.
- Ministério do Meio Ambiente—MMA. Diagnóstico Sobre a Invasão do Mexilhão-Dourado (Limnoperna fortunei) no Brasil; Ministério do Meio Ambiente—Instituto do Meio Ambiente e dos Recursos Naturais Renováveis—IBAMA: Brasília, Brazil, 2017; p. 159.
- Ludwig, S.; Sari, E.H.; Paixão, H.; Montresor, L.C.; Araújo, J.; Brito, C.F.; Martinez, C.B. High connectivity and migration potentiate the invasion of Limnoperna fortunei (Mollusca: Mytilidae) in South America. Hydrobiologia 2021, 848, 499–513. [CrossRef]
- Oliveira, M.D.; Ayroza, D.M.R.; Castellani, D.; Campos, M.C.S.; Mansur, M.C.D. O mexilhão dourado nos tanques-rede das pisciculturas das Regiões Sudeste e Centro-Oeste. Panor. Aqüicultura 2014, 24, 22–29.
- Darrigran, G.; Agudo-Padrón, I.; Baez, P.; Belz, C.; Cardoso, F.; Carranza, A.; Damborenea, C. Non-native mollusks throughout South America: Emergent patterns in an understudied continent. Biol. Invasions 2020, 22, 853–871. [CrossRef]
- Mansano, C.F.M.; Pereira, B.C.A.; Gonçalves, G.S.; Nascimento, T.M.T. Sustainable alternative for the use of invasive species of golden mussel (Limnoperna fortunei) in the feeding of Nile tilapia (Oreochromis niloticus). Lat. Am. J. Aquat. Res. 2023, 51, 692–702. [CrossRef]
- Darrigran, G.; Damborenea, C. Ecosystem Engineering Impact of Limnoperna fortunei in South America. Zool. Sci. 2011, 28, 1–7. [CrossRef]
- Oliveira, M.; Hamilton, S.; Jacobi, C. Forecasting the expansion of the invasive golden mussel Limnoperna fortunei in Brazilian and North American rivers based on its occurrence in the Paraguay River and Pantanal wetland of Brazil. Aquat. Invasions 2010, 5, 59–73. [CrossRef]
- Santos, A.M.E.; Theodoro Junior, N.; Souza, R.F.M. Ocorrência do mexilhão-dourado (Limnoperna fortunei, Dunker 1857) no Canal do Sertão, Delmiro Gouveia-AL, Brasil. Rev. Gestão Água Am. Lat. 2022, 19, e18. [CrossRef]
- Oliveira, M.D.; Campos, M.C.S.; Paolucci, E.M.; Mansur, M.C.D.; Hamilton, S.K. Colonization and Spread of Limnoperna fortunei in South America. In Limnoperna Fortunei; Boltovskoy, D., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 333–355. [CrossRef]
- Darrigran, G.; de Drago, I.E. Distribución de Limnoperna fortunei (Dunker, 1857) (Mytilidae), en La Cuenca Del Plata, Region Neotropical. Medio Ambiente 2000, 13, 75–79.
- Boltovskoy, D.; Correa, N.; Cataldo, D.; Sylvester, F. Dispersion and Ecological Impact of the Invasive Freshwater Bivalve Limnoperna fortunei in the Río de la Plata Watershed and Beyond. Biol. Invasions 2006, 8, 947–963. [CrossRef]
- Boltovskoy, D. Limnoperna fortunei: The Ecology, Distribution and Control of a Swiftly Spreading Invasive Fouling Mussel; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; p. 476.
- Morton, B. The Biology and Anatomy of Limnoperna fortunei, a Significant Freshwater Bioinvader: Blueprints for Success. In Limnoperna fortunei; Boltovskoy, D., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 3–41. [CrossRef]
- Pie, M.R.; Boeger, W.A.; Patella, L.; Falleiros, R.M. A fast and accurate molecular method for the detection of larvae of the golden mussel Limnoperna fortunei (Mollusca: Mytilidae) in plankton samples. J. Molluscan Stud. 2006, 72, 218–219. [CrossRef]
- Mansur, M.C.D. Bivalves invasores límnicos: Morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp. In Moluscos Límnicos Invasores No Brasil: Biologia, Prevencão, Controle; Redes Editora: Porto Alegre, Brazil, 2012; pp. 61–74.
- Vidigal, T.H.D.A.; Coscarelli, D.; Monstresor, L.C. Molecular studies in Brazilian malacology: Tools, trends and perspectives. Lundiana 2013, 11, 47–63. [CrossRef]
- Tominaga, A.; Goka, K.; Kimura, T.; Ito, K. Genetic structure of Japanese introduced populations of the golden mussel, Limnoperna fortunei, and the estimation of their range expansion process. Biodiversity 2009, 10, 61–66. [CrossRef]
- Zhan, A.; Perepelizin, P.V.; Ghabooli, S.; Paolucci, E.; Sylvester, F.; Sardiña, P.; Cristescu, M.E.; MacIsaac, H.J. Scale-dependent post-establishment spread and genetic diversity in an invading mollusc in South America: Invasion genetics of Limnoperna fortunei. Divers. Distrib. 2012, 18, 1042–1055. [CrossRef]
- Ghabooli, S.; Zhan, A.; Sardiña, P.; Paolucci, E.; Sylvester, F.; Perepelizin, P.V.; Cristescu, M.E. Genetic diversity in introduced golden mussel populations corresponds to vector activity. PLoS ONE 2013, 8, e59328. [CrossRef]
- Paolucci, E.M.; Sardiña, P.; Sylvester, F.; Perepelizin, P.V.; Zhan, A.; Ghabooli, S.; Cristescu, M.E.; Oliveira, M.D.; MacIsaac, H.J. Morphological and genetic variability in an alien invasive mussel across an environmental gradient in South America. Limnol. Oceanogr. 2014, 59, 400–412. [CrossRef]
- Nakano, D.; Baba, T.; Endo, N.; Nagayama, S.; Fujinaga, A.; Uchida, A.; Shiragane, A.; Urabe, M.; Kobayashi, T. Invasion, dispersion, population persistence and ecological impacts of a freshwater mussel (Limnoperna fortunei) in the Honshu Island of Japan. Biol. Invasions 2015, 17, 743–759. [CrossRef]
- Borges, P.D.; Ludwig, S.; Boeger, W.A. Testing hypotheses on the origin and dispersion of Limnoperna fortunei (Bivalvia, Mytilidae) in the Iguassu River (Paraná, Brazil): Molecular markers in larvae and adults. Limnology 2017, 18, 31–39. [CrossRef]
- Ovenden, J. Mitochondrial DNA and marine stock assessment: A review. Mar. Freshw. Res. 1990, 41, 835. [CrossRef]
- Rubinoff, D. Utility of mitochondrial DNA barcodes in species conservation: DNA barcodes and conservation. Conserv. Biol. 2006, 20, 1026–1033. [CrossRef]
- Pečnikar, F.Ž.; Buzan, E.V. 20 years since the introduction of DNA barcoding: From theory to application. J. Appl. Genet. 2014, 55, 43–52. [CrossRef]
- Junqueira, R.A.C. Mapeamento Temático de Uso da Terra no Baixo São Francisco. In Projeto de Gerenciamento Integrado das Atividades Desenvolvidas em Terra na Bacia do São Francisco; CODEVASP, Brasilia, DF, Brazil, 2002; pp. 1-35. https://cdn.agenciapeixevivo.org.br/.
- Moreira Filho, O.; Buckup, P.A. A poorly known case of watershed transposition between the São Francisco and upper Paraná river basins. Neotrop. Ichthyol. 2005, 3, 449–452. [CrossRef]
- Bot Neto, R.L.; Cattani, A.P.; Spach, H.L.; Contente, R.F.; Cardoso, O.R.; Marion, C.; Schwarz Júnior, R. Patterns in composition and occurrence of the fish fauna in shallow areas of the São Francisco River mouth. Biota Neotrop. 2023, 23, e20221387. [CrossRef]
- Christo, S.W.; Ferreira-Junior, A.L.; Absher, T.M. Aspectos reprodutivos de mexilhões (Bivalvia, Mollusca) no complexo estuarino de Paranaguá, Paraná, Brasil. Bol. Do Inst. Pesca 2016, 42, 924–936. [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Extraction and Purification of Plasmid DNA. In Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 1989; pp. 21–152.
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299.
- Alves, F.A.; Beasley, C.R.; Hoeh, W.R.; da Rocha, R.M.; Simone, L.R.; Tagliaro, C.H. Detection of mitochondrial DNA heteroplasmy suggests a doubly uniparental inheritance pattern in the mussel Mytella charruana. Rev. Bras. Biociências 2012, 10, 176.
- Endo, N.; Sato, K.; Nogata, Y. Molecular based method for the detection and quantification of larvae of the golden mussel Limnoperna fortunei using real-time PCR. Plankton Benthos Res. 2009, 4, 125–128. [CrossRef]
- Vermulm Junior, H.; Giamas, M.T.D. Ocorrência do mexilhão-dourado Limnoperna fortunei (Dunker, 1857) (Mollusca; Bivalvia; Mytilidae), no trato digestivo do “Armal” Pterodoras granulosus (Valenciennes, 1821) (Siluriformes; Doradidae), do Rio Paraná, São Paulo, Brasil. Bol. Do Inst. De Pesca 2008, 34, 175–179.
- Canzi, C.; Fialho, N.S.; Bueno, G.W. Monitoramento e ocorrência do mexilhão dourado (Limnoperna fortunei) na hidrelétrica da Itaipu binacional, Paraná (BR). Rev. Ibero-Am. Ciências Ambient. 2014, 5, 117–122. [CrossRef]
- Andrade, J.; Cordeiro, N.I.; Montressor, L.C.; Luz, D.M.; Luz, R.C.; Martinez, C.B.; Vidigal, T.H. Effect of temperature on behavior, glycogen content, and mortality in Limnoperna fortunei (Dunker, 1857) (Bivalvia: Mytilidae). J. Limnol. 2018, 77, 189–198. [CrossRef]
- Vieira, J.P.; Lopes, M.N. Size-selective predation of the catfish Pimelodus pintado (Siluriformes: Pimelodidae) on the golden mussel Limnoperna fortunei (Bivalvia: Mytilidae). Zoologia 2013, 30, 43–48. [CrossRef]
- Lopes, M.; Vieira, J. Predadores potenciais para o controle do mexilhão-dourado. In Moluscos Límnicos Invasores No Brasil: Biologia, Prevenção, Controle; Redes Editora: Porto Alegre, Brazil, 2012; pp. 357–363.
- Uliano-Silva, M.; Americo, J.A.; Costa, I.; Schomaker-Bastos, A.; de Freitas Rebelo, M.; Prosdocimi, F. The complete mitochondrial genome of the golden mussel Limnoperna fortunei and comparative mitogenomics of Mytilidae. Gene 2016, 577, 202–208. [CrossRef]
- Pessotto, M.A.; Nogueira, M.G. More than two decades after the introduction of Limnoperna fortunei (Dunker 1857) in La Plata Basin. Braz. J. Biol. 2018, 78, 773–784. [CrossRef]
- Ferreira, J.G.; Soares-Souza, G.B.; Americo, J.A.; Dumaresq, A.; Rebelo, M.F. Population structure of the invasive golden mussel (Limnoperna fortunei) on reservoirs from five Brazilian drainage basins. bioRxiv. [CrossRef]
- Correia, I.O.; da Silva Andrade, A.C.; de Rezende, P.S. Erosão costeira e faixas de proteção no delta do rio São Francisco. Quat. Environ. Geosci. 2023, 14, 26–42.
- Silva, W.F. Determinação da carga de material em suspensão no rio São Francisco: Ano hidrológico 2007. Universidade Federal de Alagoas: Maceió, Brazil, 2009; p. 48.
- de Castro, C.N.; Pereira, C.N. Revitalização da Bacia Hidrográfica do Rio São Francisco: Histórico. In Diagnóstico e Desafios; Instituto de Pesquisa Econômica Aplicada: Brasília, Brazil, 2019; p. 366.
- Darrigran, G.; Pastorino, G. The recent introduction of a freshwater asiatic bivalve Limnoperna fortunei (Mytilidae) into South America. Veliger 1995, 32, 171–175.
- Giraudo, M.E. Dependent development in South America: China and the soybean nexus. J. Agrar. Chang. 2020, 20, 60–78. [CrossRef]
- Costa Fernandes, F.; Mansur, M.C.D.; Pereira, D.; de Godoy Fernandes, L.V.; Campos, S.C.; Danelon, O.M. Abordagem Conceitual dos Moluscos Invasores nos Ecossistemas Límnicos Brasileiros. In Moluscos Límnicos Invasores No Brasil: Biologia, Prevenção, Controle; Redes Editora: Porto Alegre, Brazil, 2012; pp. 19–23.
- Yuan, W.; Walters, L.J.; Schneider, K.R.; Hoffman, E.A. Exploring the survival threshold: A study of salinity tolerance of the nonnative mussel Mytella charruana. J. Shellfish Res. 2010, 29, 415–422. [CrossRef]
- Peres, W.A.M.; Bertollo, L.A.C.; Buckup, P.A.; Blanco, D.R.; Kantek, D.L.Z.; Moreira-Filho, O. Invasion, dispersion and hybridization of fish associated to river transposition: Karyotypic evidence in “Astyanax bimaculatus group” (Characiformes: Characidae). Rev. Fish Biol. Fish. 2012, 22, 519–526. [CrossRef]
- de O. Mendes-Neto, E.; Vicari, M.R.; Artoni, R.; Moreira-Filho, O. Description of karyotype in Hypostomus regani (Ihering, 1905) (Teleostei, Loricariidae) from the Piumhi river in Brazil with comments on karyotype variation found in Hypostomus. Comp. Cytogenet. 2011, 5, 133–142. [CrossRef]
- Barbosa, N.P.U.; Ferreira, J.A.; Nascimento, C.A.R.; Silva, F.A.; Carvalho, V.A.; Xavier, E.R.S.; Ramon, L.; Almeida, A.C.; Carvalho, M.D.; Cardoso, A.V. Prediction of future risk of invasion by Limnoperna fortunei (Dunker, 1857) (Mollusca, Bivalvia, Mytilidae) in Brazil with cellular automata. Ecol. Indic. 2018, 92, 30–39. [CrossRef]
- Davidson, A.; Fusaro, A.; Sturtevant, R.; Kashian, D. Development of a risk assessment framework to predict invasive species establishment for multiple taxonomic groups and vectors of introduction. Manag. Biol. Invasions 2017, 8, 25–36. [CrossRef]
- Correa, N.; Sardiña, P.; Perepelizin, P.V.; Boltovskoy, D. Limnoperna fortunei colonies: Structure, Distribution and Dynamics. In Limnoperna Fortunei; Boltovskoy, D., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 119–143.
- Darrigran, G.; Damborenea, C.; Drago, E.C.; Ezcurra de Drago, I.; Paira, A.; Archuby, F. Invasion process of Limnoperna fortunei (Bivalvia: Mytilidae): The case of Uruguay River and emissaries of the Esteros del Iberá Wetland, Argentina. Zoologia 2012, 29, 531–539. [CrossRef]
- Ito, K. Distribution and spread of Limnoperna fortunei in Japan. In Limnoperna Fortunei—The Ecology, Distribution and Control of a Swiftly Spreading Invasive Fouling Mussel; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 321–332.
- Furlan-Murari, P.J.; Ruas, C.D.F.; Ruas, E.A.; Benício, L.M.; Urrea-Rojas, A.M.; Poveda-Parra, A.R.; Lopera-Barrero, N.M. Structure and genetic variability of golden mussel (Limnoperna fortunei) populations from Brazilian reservoirs. Ecol. Evol. 2019, 9, 2706–2714. [CrossRef]
- Ribeiro-Neto, T.F.; Silva, A.H.G.D.; Guimarães, I.M.; Gomes, M.V.T. Piscicultura familiar extensiva no baixo São Francisco, estado de Sergipe, Brasil. Acta Fish. Aquat. Resour. 2016, 4, 62–69. [CrossRef]
- de Farias Costa, E.; Sampaio, Y. Direct and indirect job generation in the farmed shrimp production chain 1. Aquac. Econ. Manag. 2004, 8, 143–155. [CrossRef]
- Sá, T.D.; Sousa, R.R.d.; Rocha, Í.R.C.B.; Lima, G.C.D.; Costa, F.H.F. Brackish Shrimp Farming in Northeastern Brazil: The Environmental and Socio-Economic Impacts and Sustainability. Nat. Resour. 2013, 4, 538–550. [CrossRef]
- Campos, M.C.S.; Andrade, A.F.A.; Kunzmann, B.; Galvão, D.D.; Silva, F.A.; Cardoso, A.V.; Carvalho, M.D.; Mota, H.R. Modelling of the potential distribution of Limnoperna fortunei (Dunker, 1857) on a global scale. Aquat. Invasions 2014, 9, 253–265.
- CTSSBSF. 2016a. Available online: https://issuu.com/canoadocs/docs/alertamexdou-01-2016 (accessed on 16 September 2024).
- CTSSBSF. 2016b. Available online: https://issuu.com/canoadocs/docs/alertamexdou-02-2016 (accessed on 16 September 2024).
- Fonseca, S.L.; Magalhães, A.A.; Campos V.P.; Medeiros Y.D. Effect of the reduction of the outflow restriction discharge from the Xingó dam in water salinity in the lower stretch of the São Francisco River. Revista Brasileira de Recursos Hídricos Brazilian Journal of Water Resources 2020, 25, e4. [CrossRef]
- Campos, S.C.; Danelon, O.M. Abordagem conceitual dos moluscos invasores nos ecossistemas límnicos brasileiros. In Moluscos Límnicos Invasores No Brasil: Biologia, Prevenção, Controle; Redes Editora: Porto Alegre, Brazil, 2012; pp. 19–23.
- Pereira, C.N.; Souza, J.A.; Almeida, F.G. Aspectos econômicos da região do São Francisco. In Revitalização da Bacia Hidrográfica do rio São Francisco: Histórico, Diagnóstico e Desafios; Castro, C.N.d., Pereira, C.N., Eds.; Instituto de Pesquisa Econômica Aplicada: Brasília, Brazil, 2003; pp. 100–120.
- Diegues, A.C. An Inventory of Brazilian Wetlands; IUCN: Gland, Switzerland, 1994; 216p.
- Sato, Y.; Godinho, H.P. Peixes da bacia do São Francisco. In Estudos Ecológicos de Comunidades de Peixes Tropicais; Edusp: São Paulo, Brazil, 1999; pp. 401–413.
- Sato, Y.; Godinho, H.P. Migratory fishes of the São Francisco River. In Migratory Fishes of South America: Biology, Fisheries and Conservation Status; IDRC: Ottawa, ON, Canada, 2004; p. 380.
- Barbosa, L.M.; de Lima, C.C.U.; Santos, R.C.D.L.; de Carvalho, J.B.; Santos, C.F.; Albuquerque, A.L. As variações morfológicas do campo de dunas ativas entre Pontal do Peba e a foz do rio São Francisco (AL). IX Congresso da Associação Brasileira de Estudos do Quaternário / II Congresso do Quaternário de Países de Línguas Ibéricas / II Congresso sobre Planejamento e Gestão da Zona Costeira dos Países de Expressão Portuguesa, 2003, 1, 1-3.
- Bittencourt, A.C.S.P.; Martin, L.; Dominguez, J.M.L.; Ferreira, Y.D.A. Evolução paleogeográfica quaternária da costa do Estado de Sergipe e da costa sul do Estado de Alagoas. Rev. Bras. Geociênc. 1983, 13, 93–97.
- Bittencourt, A.C.D.S.P.; Dominguez, J.M.L.; Fontes, L.C.S.; Sousa, D.L.; Silva, I.R.; Da Silva, F.R. Wave refraction, river damming, and episodes of severe shoreline erosion: The São Francisco river mouth, Northeastern Brazil. J. Coast. Res. 2007, 23, 930–938.
- Medeiros, P.P.; dos Santos, M.M.; Cavalcante, G.H.; De Souza, W.F.L.; da Silva, W.F. Características ambientais do Baixo São Francisco (AL/SE): Efeitos de barragens no transporte de materiais na interface continente-oceano. Geochim. Bras. 2014, 28, 65.
- Fontes, A.L. Processos erosivos na desembocadura do Rio São Francisco. In Boletim de Resumos, VIII Congresso da ABEQUA; ABEQUA: São Paulo, Brazil, 2001; pp. 66–67.
- Vieira, C.L.; Gonçalves, V.; Vieira, R.C.; Beserra, M.L. Plano de Manejo Para Área de Proteção Ambiental de Piaçabuçu; Ministério do Meio Ambiente: Brasília, Brazil, 2010.
- Barreto, S.A.; Rodrigues, T.K. Usos e conflitos na Reserva Biológica de Santa Isabel no trecho da zona costeira do Grupo de Bacias Costeiras 01-Sergipe; Seminários Espaços Costeiros: Salvador-Ba, Brazil, 2016.
- Santos, E.A.P.D.; Landim, M.F.; Oliveira, E.V.D.S.; Silva, A.C.C.D.D. Conservação da zona costeira e áreas protegidas: A Reserva Biológica de Santa Isabel (Sergipe) como estudo de caso. Rev. Ambiental 2017,15(3), 41-57.
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791.
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [CrossRef]
- Collins, R.A.; Boykin, L.M.; Cruickshank, R.H.; Armstrong, K.F. Barcoding’s next top model: An evaluation of nucleotide substitution models for specimen identification. Methods Ecol. Evol. 2012, 3, 457–465. [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [CrossRef]
- Rambaut, A.; Drummond, A. FigTree: Tree Figure Drawing Tool, v1.4.2; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2012.
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [CrossRef]
Figure 1.
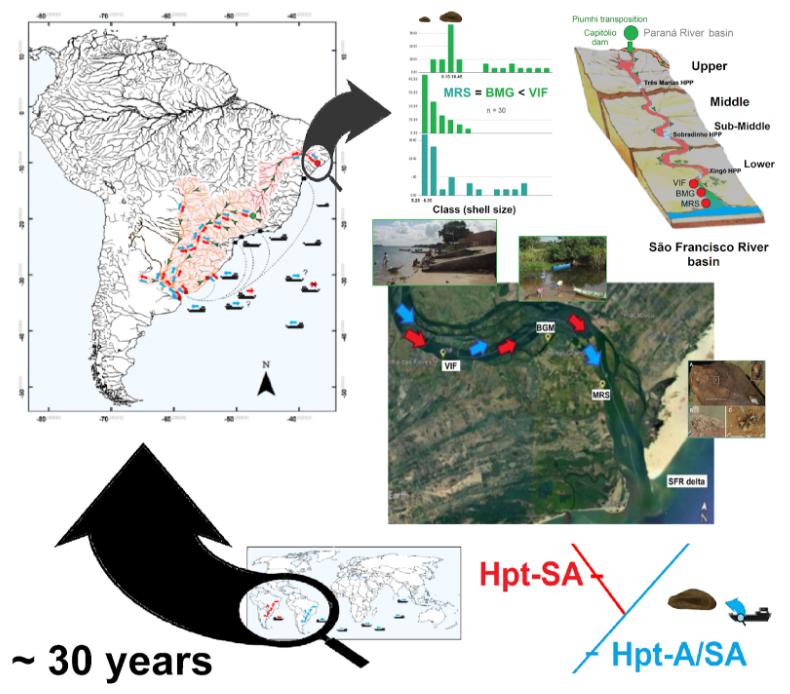
Occurrence of the golden mussel (red-shaded areas) in ecoregions (according to [29,31]), distribution of haplotypes, and new occurrence of Limnoperna fortunei in South America (SA), with emphasis on its presence in the hydrographic basins of the Paraná River (along the orange line) and the São Francisco River (along the red line). Blue arrows indicate the occurrence of the Asian ancestral haplotypes (shares of Asia and SA) (Hpt-A/SA); red arrows indicate the South American-derived haplotypes (exclusively SA) (Hpt-SA); green arrows indicate the river direction; the green circle indicates the region of the Capitólio dam and the Piumhi watershed transposition, see [53,76]; vessels: bulk cargo transport; red squares indicate ports with golden mussel occurrence; black squares indicate ports without occurrence; lines indicate cargo cabotage routes between ports with (continuous) and without mussels (dotted); X—doubts about the flow of the common haplotype in the sea routes with the absence of Hpt-SA to Asian populations; ?—doubts about the flow of Hpt-A/SA in the sea routes. (A) Zoom of the dispersion of the golden mussel (Limnoperna fortunei) in the Lower São Francisco River (LSFR) basin. The red polygon (A) indicates the region classified as the LSFR; the yellow polygon (A,B) indicates the site with a public warning of the presence of the golden mussel in 2016 [87,88]; yellow balls with a star (A–C) indicate confirmed occurrences of the golden mussel in the LSFR basin. Locations: Xingó HPP/Xingó dam: Hydroelectric Plant Xingó; VIF: Village Ilha das Flores; BGM: Brejo Grande Marine; MRS: Mouth of the São Francisco River. Distances: end of yellow polygon (Mato da Onça Reserve)-VIF (124,973.7 m); VIF-BGM (7499.5 m); VIF-MRS (11,494.6 m); BGM-MRS (4909.1 m).
Figure 1.
Occurrence of the golden mussel (red-shaded areas) in ecoregions (according to [29,31]), distribution of haplotypes, and new occurrence of Limnoperna fortunei in South America (SA), with emphasis on its presence in the hydrographic basins of the Paraná River (along the orange line) and the São Francisco River (along the red line). Blue arrows indicate the occurrence of the Asian ancestral haplotypes (shares of Asia and SA) (Hpt-A/SA); red arrows indicate the South American-derived haplotypes (exclusively SA) (Hpt-SA); green arrows indicate the river direction; the green circle indicates the region of the Capitólio dam and the Piumhi watershed transposition, see [53,76]; vessels: bulk cargo transport; red squares indicate ports with golden mussel occurrence; black squares indicate ports without occurrence; lines indicate cargo cabotage routes between ports with (continuous) and without mussels (dotted); X—doubts about the flow of the common haplotype in the sea routes with the absence of Hpt-SA to Asian populations; ?—doubts about the flow of Hpt-A/SA in the sea routes. (A) Zoom of the dispersion of the golden mussel (Limnoperna fortunei) in the Lower São Francisco River (LSFR) basin. The red polygon (A) indicates the region classified as the LSFR; the yellow polygon (A,B) indicates the site with a public warning of the presence of the golden mussel in 2016 [87,88]; yellow balls with a star (A–C) indicate confirmed occurrences of the golden mussel in the LSFR basin. Locations: Xingó HPP/Xingó dam: Hydroelectric Plant Xingó; VIF: Village Ilha das Flores; BGM: Brejo Grande Marine; MRS: Mouth of the São Francisco River. Distances: end of yellow polygon (Mato da Onça Reserve)-VIF (124,973.7 m); VIF-BGM (7499.5 m); VIF-MRS (11,494.6 m); BGM-MRS (4909.1 m).
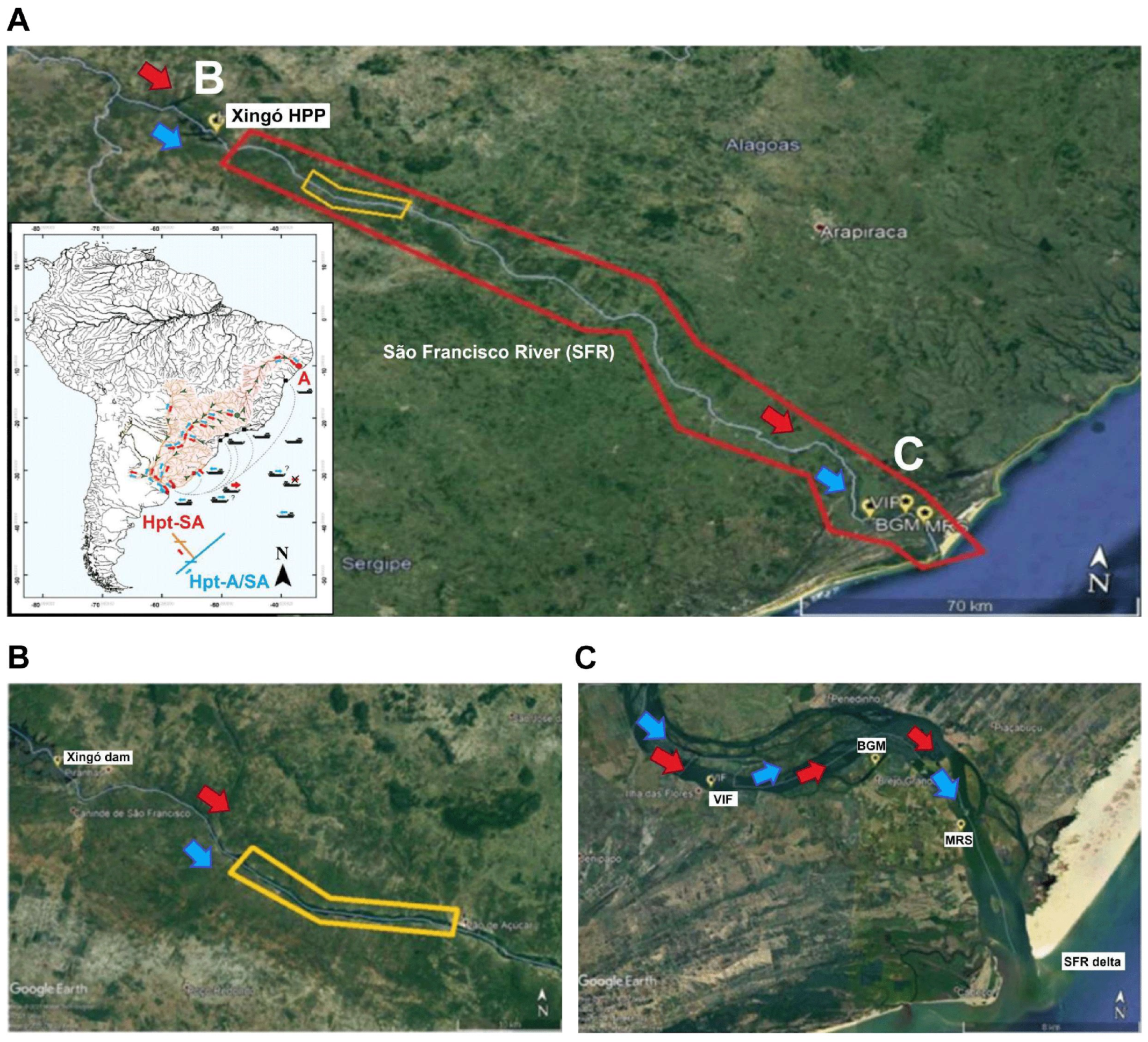
Figure 2.
Position of the sample barcode (Cox1) from the Lower São Francisco River and the sequence barcodes available from the Genbank and BOLD systems of the Limnoperna fortunei mitogenome (KP756905). bp: base pairs; Haplotypes: K01-21 (see [46]) and Lfm01-Lf43 (see [28,44,45,46]); Genbank and BOLD systems accessions: KP756905 [65], AB520611–AB520627, HQ843794–HQ843810, JX177086–JX177102 [28,44,45,46], AB828679–AB828682 [47], FW369972, DQ264395 [40], AB498011 [59].
Figure 2.
Position of the sample barcode (Cox1) from the Lower São Francisco River and the sequence barcodes available from the Genbank and BOLD systems of the Limnoperna fortunei mitogenome (KP756905). bp: base pairs; Haplotypes: K01-21 (see [46]) and Lfm01-Lf43 (see [28,44,45,46]); Genbank and BOLD systems accessions: KP756905 [65], AB520611–AB520627, HQ843794–HQ843810, JX177086–JX177102 [28,44,45,46], AB828679–AB828682 [47], FW369972, DQ264395 [40], AB498011 [59].
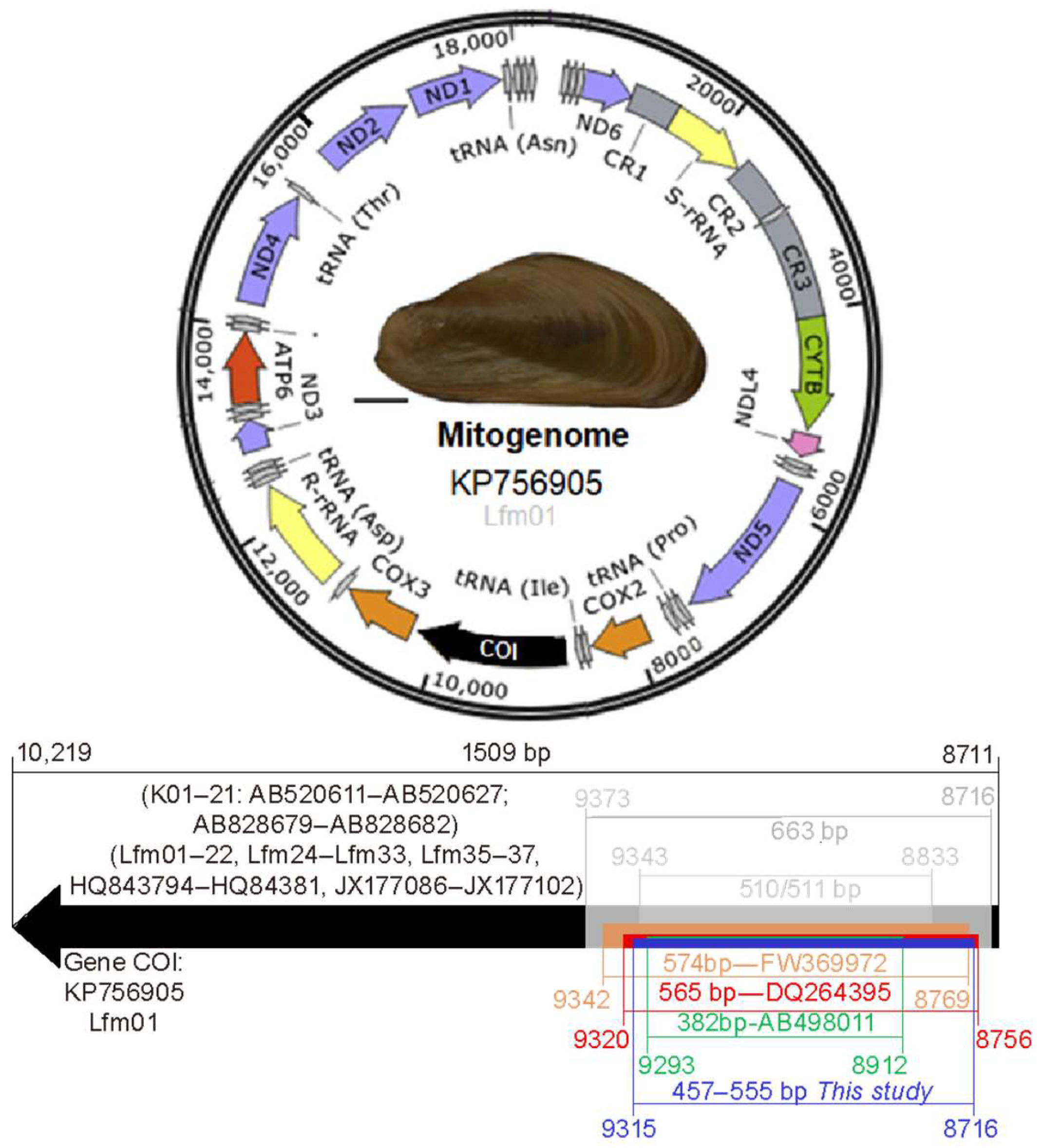
Figure 3.
Haplotype TSC network for the mitochondrial cytochrome c oxidase subunit I (Cox1) gene in the golden mussel exclusive (black and red circles) and shared (blue circles) in Asia and South America. Black circles—exclusive haplotypes of Asia; blue circles—haplotypes present in Asia and South America (identified in the Lower São Francisco River—LSFR); red circles—exclusive haplotypes of South America (identified in the LSFR); white circles—haplotypes not identified your continental origin; gray circles—outgroup Limnoperna siamensis.
Figure 3.
Haplotype TSC network for the mitochondrial cytochrome c oxidase subunit I (Cox1) gene in the golden mussel exclusive (black and red circles) and shared (blue circles) in Asia and South America. Black circles—exclusive haplotypes of Asia; blue circles—haplotypes present in Asia and South America (identified in the Lower São Francisco River—LSFR); red circles—exclusive haplotypes of South America (identified in the LSFR); white circles—haplotypes not identified your continental origin; gray circles—outgroup Limnoperna siamensis.
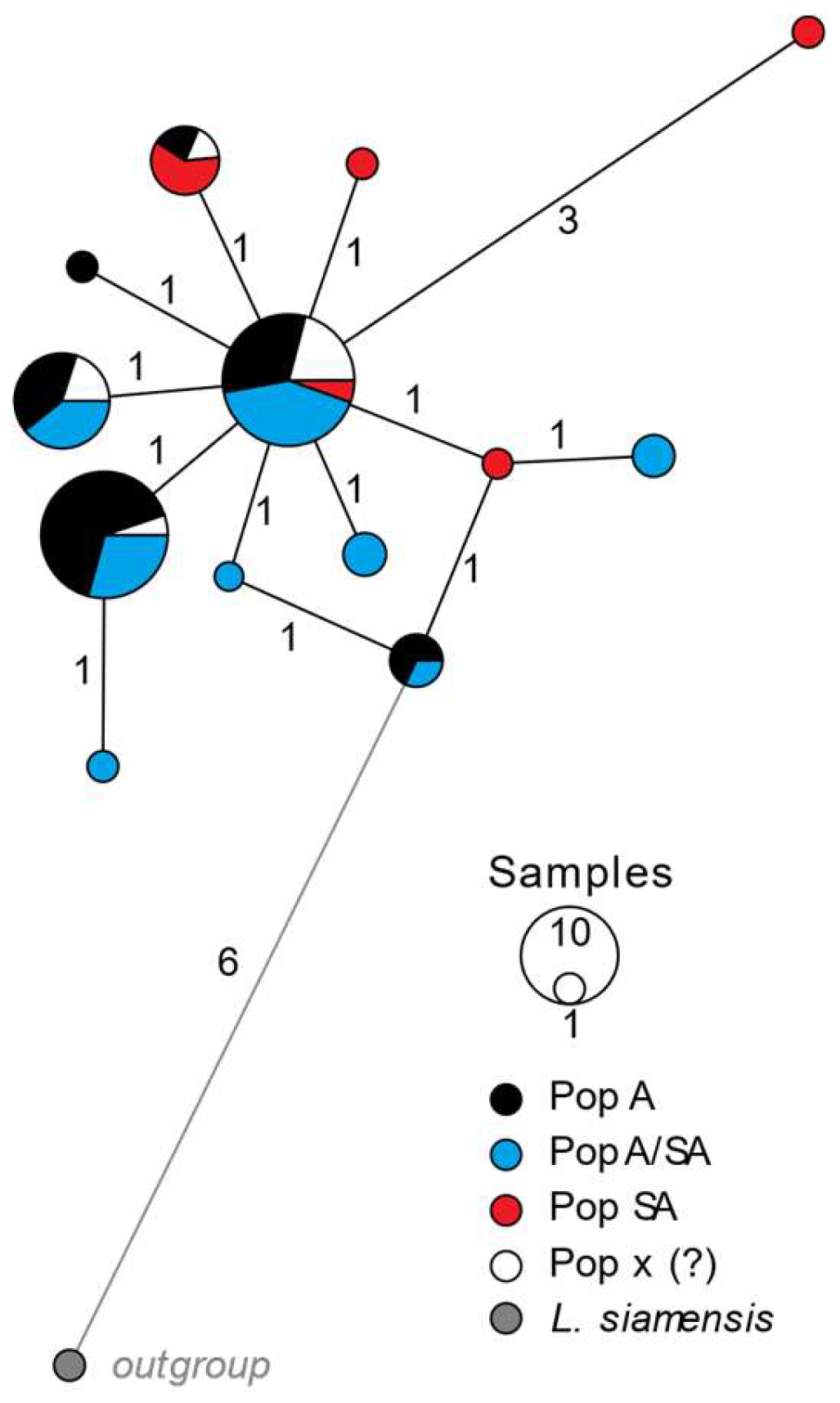

Table 1.
Biometrics of Limnoperna fortunei shells found in Brazil. n—number of mussels collected. LA—mean shell length; SD—standard deviation; LR—shell length range. T.S.—this study.
Table 1.
Biometrics of Limnoperna fortunei shells found in Brazil. n—number of mussels collected. LA—mean shell length; SD—standard deviation; LR—shell length range. T.S.—this study.
| Location | n | LA ± SD (mm) | LR (mm) a | References |
|---|---|---|---|---|
| São Francisco River ts (−8.54°; −39.456°) | 5 | - - c | 15.00 to 30.00 | [16] |
| Sobradinho reservoir (−9.409; −40.817) | 5 | - - c | 15.00 to 30.00 | [16] |
| Sobradinho hydroelectric (−9.433; −40.828) | 5 | - - c | 15.00 to 30.00 | [16] |
| Ilha da Flores (−10.435°; −36.531°) | 30 | 12.84 ± 4.95 * | 7.44 to 23.79 | T.S. |
| Brejo Grande marine (−10.421°; −36.464°) | 30 | 7.42 ± 1.82 * | 5.36 to 11.75 | T.S. |
| São Francisco River mouth (−10.445°; −36.426°) | 30 | 9.37 ± 4.45 * | 5.25 to 20.14 | T.S. |
| Porto Primavera hydroelectric (−22.474°; −52.474°) | 61 | - - c | 4.00 to 27.00 | [60] b |
| Itaipu reservoir—Paraná river (−24°; −54°) | - - c | - - c | 6.00 to 35.00 | [61] |
| Paraná River (−25.437°; −54.512°) | 800 | - - c | 21.00 to 27.00 | [62] |
| Mirim Lake (−31.799°; −52.382° and −32.116°; −52.582°) | 7789 | - - c | 3.00 to 32.00 | [63] d |
| Mirim Lake (−31.799°; −52.382° and −32.116°;−52.582°) | 147 | - - c | 3.00 to 14.00 | [63] e |
| São Gonçalo channel (−32.148°; −52.625°) | 7776 | - - c | 4.00 to 32.00 | [64] d |
ts—transposition system; * total average for the site (9.81 ± 4.57 mm); a variation in shell length; b inside the stomach of Pterodoras granulosus; c not cited by the authors; d depth from 3 to 6 m; e inside the stomach of Pimelodus pintado.
Table 2.
Haplotype Codes for the mitochondrial cytochrome c oxidase subunit I (Cox1) gene in the golden mussel Limnoperna fortunei. ID—identification of population; np, number of populations; $$ haplotype identity, according to [44] and [47]; black—exclusive haplotype of Asia (A); blue—haplotype present in A and South America (A/SA); red—exclusive haplotype of South America (SA); bold italic—haplotypes identified in this study (T.S.); bold—haplotypes observed continentally in SA; ** not described by the authors or haplotypes with another acronym (=100% similarity); ** sequences (<99.6% similarities).
Table 2.
Haplotype Codes for the mitochondrial cytochrome c oxidase subunit I (Cox1) gene in the golden mussel Limnoperna fortunei. ID—identification of population; np, number of populations; $$ haplotype identity, according to [44] and [47]; black—exclusive haplotype of Asia (A); blue—haplotype present in A and South America (A/SA); red—exclusive haplotype of South America (SA); bold italic—haplotypes identified in this study (T.S.); bold—haplotypes observed continentally in SA; ** not described by the authors or haplotypes with another acronym (=100% similarity); ** sequences (<99.6% similarities).
| Distribution | Country | HAPLOTYPES | Ref. | ||||
|---|---|---|---|---|---|---|---|
| ID | np | Codes $$ | |||||
| A/SA | Taiwan | TW1–TW2 | 2 | Lfm03, Lfm08 | [45] | ||
| A | TW1 | 1 | Lfm19 | ||||
| A/SA | Japan | JP1–JP5; JP7–JP29 | 28 | Lfm03 (=K07,K13 *), Lfm05 (=K09 *), Lfm09 (=K01 *), Lfm11 (=K05, K11, K17, K21 *), Lfm12 (=K12 *), Lfm15, Lfm27 (=K02 *), Lfm28 | [10,11,45,47,59,65] | ||
| A | JP1–JP29 | 29 | Lfm20 * (=K03 *), Lfm21 (=K04, K06 *), Lfm28–Lfm29 (=K10 *), Lfm37(=K08 *), K18 * | ||||
| A/SA | Korea | KR | 1 | Lfm11 | [45] | ||
| A | KR | 1 | Lfm21, Lfm26 | ||||
| A/SA | China | CH1–CH5 | 5 | Lfm02, Lfm03, Lfm06, Lfm11-12, Lfm15, Lfm27, Lfm36, Lfm38, Lfm41-42 | [28,45] | ||
| A | CH1–CH5 | 5 | Lfm21, Lfm24–25, Lfm30-33, Lfm35, Lfm37, Lfm43 | ||||
| SA | Brazil | SOB, VIF, BGM, MRS, CO, COR,VOL,IGA, JUP, POR, BAR, RB, IT1, IT2 *, IRB, MAC, BAL, POA1, SO | 19 | Lfm01, Lfm04, Lfm07, Lfm10, Lfm39 | [28,40] **, [44,45,46,65] *, T.S. |
||
| A/SA | SOB, VIF, BGM, MRS, CO, COR, VOL, IGA, JUP, ROS, POR, BAR, BON, RB, IT1, IT2 *, IT3, CX1 *, CX2, OS *, PQ, MAC, BAL, AMA, DMA, POA2, PEL, SO | 28 | Lfm02, Lfm03, Lfm05, Lfm06, Lfm08-09, Lfm11, Lfm15, Lfm36, Lfm38, Lfm41, Lfm42 | [28,44,45,46], T.S. |
|||
| SA | Uruguay | SG–SL | 2 | Lfm01, Lfm04, Lfm10, | [44,46] | ||
| A/SA | SG–SL | 2 | Lfm03, Lfm05, Lfm09, Lfm11–12, Lfm27 | ||||
| SA | Argentina | YR, YD, PR, FE, CA-1, CA-2, PU, SA, UR, RT, EC, CR, TI, SF, BA, QU, PL, MA | 18 | Lfm01, Lfm04, Lfm07, Lfm13, Lfm16-18 | [44,46] | ||
| A/SA | YR, YD, PR, FE, CA-1, CA-2, PU, SA, UR, RT, EC, CR, TI, SF, BA, QU, PL, PLA, MA | 19 | Lfm02, Lfm03, Lfm05, Lfm06, Lfm08-09, Lfm11, Lfm14, Lfm15 | ||||
| Total populations observed: 88 (37 in Asia and 51 in South America). | |||||||
| Total specimens observed: 2160 (735 in Asia and 1425 in South America). | |||||||
10 GenBank: FW369972(574 bp); 11 GenBank: AB828679 (658 bp); T.S. (~555 bp).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



