Submitted:
27 September 2024
Posted:
30 September 2024
You are already at the latest version
Abstract
Salinity is a major abiotic stress affecting agricultural productivity. This research investigated the impact of mycorrhizal fungi (AMF), bacteria (PGPR), and their combination on pepper plants under salt stress (200 mM NaCl). Conducted in a controlled climate chamber, the study assessed plant height, stem diameter, leaf number, leaf area, leaf weight, shoot fresh and dry weights, SPAD values, leaf relative water content (RWC), leaf water potential, and mineral concentrations (Na, K, Mg, Ca, P, Cu, Mn, Fe, Zn). AMF application significantly increased plant height (39.50 cm) compared to PGPR (37.75 cm) and the control (37.25 cm). While salt stress reduced plant height by 34.90%, AMF and PGPR mitigated these effects, with AMF improving height by 21.65% under saline conditions. AMF and PGPR positively influenced plant diameter, leaf number, and fresh weight. Physiological measurements, including SPAD values, RWC, and osmotic potential, showed that AMF+PGPR applications were most effective in increasing RWC. Nutrient analysis revealed reduced calcium and magnesium levels due to salt stress. In conclusion, AMF and PGPR enhanced pepper plants' growth and physiological responses under salt stress, indicating their potential to improve agricultural practices in saline environments.
Keywords:
salinity
; mycorrhiza
; bacteria
; pepper
; abiotic stress
1. Introduction
Climate change is rapidly increasing its impact worldwide, significantly altering plant habitats. The factors causing adversities in the plant life cycle are termed "stress." Abiotic stress factors include drought, salinity, temperature fluctuations, excessive water, nutrient deficiencies or excesses, heavy metals, and air pollution. Biotic stress factors consist of pathogens, insects, and herbivores. These stresses complicate plant growth and development processes, resulting in reduced yield and quality of agricultural crops [1,2,3,4].
Salinity, an abiotic stress factor, ranks second after drought, affecting approximately 20% of usable agricultural land [5]. Alterations in climate conditions and improper agricultural practices contribute to the escalation of soil salinity [6,7,8]. When cultivated in saline soils, plants encounter numerous challenges due to salinity [9,10,11]. Salinity diminishes plants' capacity to absorb water, resulting in damage and potential fatality [12,13]. Additionally, it disrupts osmotic potential, exerting detrimental effects on germination, growth, physiology, and productivity [14,15]. Numerous studies have documented stunted plant growth across various plant species under saline conditions [16,17,18]. The decline in photosynthesis coupled with an increase in respiration under salt stress leads to reduced plant growth and a decline in dry matter production [19,20].
Salt stress results in decreased levels of photosynthetic pigments, antioxidant production, osmolyte accumulation, water uptake, root hydraulic conductivity, magnesium (Mg) levels, potassium/sodium (K+/Na+) and magnesium/ sodium (Mg2+/Na2+) ratios, as well as the uptake of calcium (Ca2+), phosphorus (P), and other nutrients. However, it is noted that reactive oxygen species (ROS) production such as hydrogen peroxide (H2O2), malondialdehyde (MDA), electrolyte leakage, and the influx of Na are increased [21,22,23]. To mitigate these adverse effects of salt stress, the utilization of biostimulants has experienced a rapid increase in recent years. Among the biostimulants materials employed to alleviate the detrimental impact of salt stress are vermicompost, seaweed fertilizer, tea waste, leonardite, zeolite, humic acid, fulvic acid, mycorrhiza, and bacteria [24,25].
In field conditions, plants are surrounded by diverse microorganisms, including bacteria and fungi, which contribute to enhancing plant growth and yield under various stress conditions [26,27,28,29,30]. Arbuscular mycorrhiza fungi (AMF) plays a crucial role in alleviating the toxicity of salt stress by normalizing the uptake mechanism of essential nutrients in plants. It enhances the water balance mechanism, leading to improved tolerance to salt stress [31,32]. Some literature suggests that growth regulators produced by mycorrhizal fungi mitigate the adverse effects on plants exposed to salt stress [26,33]. In studies, it has been reported that plants inoculated with AMF develop better than uninoculated plants under salt stress [32,34,35]. AMF application has been reported to be effective in alleviating the negative effects of salt stress in different plant species, such as tomato [36,37], pepper [38], cucumber [39], eggplant [40], zucchini [41], beans [42], maize [43], lettuce [44,45,46].
Plant Growth-Promoting Rhizobacteria (PGPR) microscopic organisms found in the rhizosphere, constitute the largest part of soil microorganisms. They play a vital role in agricultural production by promoting plant growth as biofertilizers. PGPR have been reported to exert various effects on plant growth, nutrient uptake, reducing the need for chemical fertilizer use, and promoting the production of indole-3-acetic acid (IAA) and cytokines in plants, as well as the promotion of gibberellins [47,48,49,50]. Research on the wide-ranging applications of rhizobacteria has been conducted by numerous researchers. These studies have focused on the effects of rhizobacteria, such as detoxification of heavy metals [18,51], degradation of pesticides [52,53], tolerance to salinity [54,55,56], biological control of plant diseases and pests [53,56,57], increased utilization of nutrients and minerals by plants [58,59], and support for plant growth through the production of phytohormones and enzymes [60,61]. It is known that the application of bacteria is effective in regulating the negative effects of stress conditions on plant growth and development, and supports the production of signal molecules. PGPR stimulate the production of the aminocyclopropane-1-carboxylate deaminase (ACCD) enzyme contribute positively to plant growth and development. This is particularly evident in its ability to reduce plant ethylene levels under various environmental stress conditions. It is also known that the application of PGPR is effective in increasing IAA, gibberellins (GA) and and zeatin (Zt) in the plant [62].
Pepper (Capsicum annuum) belongs to the genus Capsicum in the Solanaceae family. It is a cultivated plant, with a one-year life cycle in temperate climates and several years in tropical climates [38]. Pepper (Capsicum annuum L.) is a significant agricultural product, valued for its economic importance and the nutritional content of its fruits, which include vitamin C and antioxidants essential for human health [63]. Most pepper varieties exhibit moderate sensitivity to salt stress [38,64]. The optimum electrical conductivity (EC) value in pepper cultivation, which is moderately sensitive to salinity, is 1.5 dS m-1. Beyond this threshold, plant growth and yield experience a significant decrease, particularly at values above 5 dS m-1 [38]. In this study, the objective was to investigate the impact of bacterial, mycorrhizal, and combined bacterial+mycorrhizal applications on young pepper plants under salt stress.
This study distinguishes itself from previous research in the field through several noteworthy aspects and insights. Firstly, the investigation into the combined effects of arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) is a significant advancement. Many earlier studies typically focused on the isolated effects of either AMF or PGPR, often neglecting the potential synergistic benefits of their combined application. This study demonstrates that the combination of AMF and PGPR can enhance plant growth parameters, such as height and leaf number, under salt stress conditions, providing compelling evidence for the benefits of integrative approaches in plant stress management. Secondly, the study employs a comprehensive array of physiological assessments, including SPAD values and leaf relative water content (RWC). This multidimensional analysis allows for a deeper understanding of the mechanisms by which these microbial treatments mitigate the detrimental effects of salt stress. In contrast, much prior research has often focused solely on growth metrics without delving into the physiological underpinnings of plant responses. Moreover, the detailed examination of mineral nutrient uptake across various treatments reveals significant interactions that contribute to plant health. The study highlights the reductions in calcium and magnesium content under salt stress, which contrasts with previous findings that may not have fully addressed these critical aspects of nutrient dynamics. The study also emphasizes the importance of maintaining a favorable ion balance, showcasing how AMF and PGPR applications can influence sodium and chlorine uptake under saline conditions. Additionally, the statistical robustness of the findings, with clear distinctions between treatment effects across multiple parameters, enhances the credibility and generalizability of the results. This thorough analysis not only provides insights into the specific benefits of AMF and PGPR applications but also offers a broader perspective on developing effective strategies for managing plant stress in saline environments. In summary, this study significantly contributes to the existing literature by providing a more nuanced understanding of how microbial applications can interact synergistically to promote plant resilience against salt stress. Its comprehensive approach and focus on both growth and physiological responses set it apart from previous research, paving the way for future studies to explore integrated microbial strategies in sustainable agriculture.
2. Materials and Methods
The research was carried out in the climate chamber with 25/20°C (day/night) for 16 hours under light and 8 hours of dark, 65-70% relative humidity and 400 µmol m-2s-1 light intensity conditions. Plant material was the Tekin F1 pepper variety obtained from Yüksel Seed company.
Pepper seeds were planted in a medium containing peat:perlite (2:1) mixture. After there staying here for 37 days, the young plants were then transferred to pots with a volume of two liters, containing peat:perlite (2:1) mixture. Pepper plants were cultivated for 42 days according to the following distinct applications, with one plant per pot. Pepper plants were irrigated with the nutrient solution shown in Table 1 [65]. After transferring the pepper plants to pots, 15 days later, the salt, AMF, and PGPR applications were initiated. Salt stress application was started with 50 mM NaCl, then gradually increased to 200 mM NaCl as the final concentration in 3 days. The experiment was carried out according to the random plots experimental design with four replications. Each replication consisted of 8 plants. In the study, eight treatments were applied.
- Control (C)
- Arbuscular Mycorrhizal Fungus (AMF)
- Plant Growth-Promoting Rhizobacteria (PGPR)
- Arbuscular Mycorrhizal Fungus+ Plant Growth-Promoting Rhizobacteria (AMF+PGPR)
- Salt stress (S)
- Salt stress+Arbuscular Mycorrhizal Fungus (S+AMF)
- Salt stress+Plant Growth-Promoting Rhizobacteria (S+PGPR)
- Salt stress+Arbuscular Mycorrhizal Fungus+Plant Growth-Promoting Rhizobacteria (S+AMF+PGPR)
The AMF bio-fertilizer, known as 'Endo Roots Soluble' (ERS), is a blend comprising nine different mycorrhiza species: Glomus intraradices, Glomus aggregatum, Glomus mosseae, Glomus clarum, Glomus monosporus, Glomus deserticola, Glomus brasilianum, Glomus etunicatum, and Gigaspora margarita. In this experiment, the liquid bacterial bio-fertilizer, 'Medbio,' was utilized, consisting of four distinct bacterial species: Bacillus subtilis (1x109), Bacillus licheniformis (2x106), Bacillus megaterium (1x109), and Pseudomonas putita (1x1010). Pepper seedlings were inoculated only once during sowing, with approximately 1000 mycorrhizae spores per plant [65]. The PGPR application was administered once a week, in conjunction with a nutrient solution, at a dosage of 1 ml of Metbio per liter of nutrient solution. Additionally, PGPR was applied weekly as a foliar spray as dosage of 1 ml of Metbio per liter of water. Since the application of PGPR was applied, the equivalent amount of water was applied to the leaves in control and salt applications.
The amount of nutrient solution applied to the plants was determined according to the drainage rate (DR) that drained in the containers placed at the bottom of the pots. Treatment solutions and irrigation were applied so that the drainage rate was approximately 22 ± 2% [66]. Salt treatment had the same nutrient solution with the addition of 200 mM NaCl. The electrical conductivity (EC) of the control nutrient solution was between 1.8 and 2.4 dS m–1, and the EC of the saline nutrient solution was between 20.0 and 21.0 dS m–1.
DR = drainage solution (ml)/ applied nutrient solution (ml) × 100
2.1. Parameters Investigated in Experiment
Pepper plants were compared 51 days after transplanting (DAT) for the plant height, stem diameter, leaf number, stem weight, leaf weight, leaf area, strem dry weight, leaf dry weight, leaf relative water content, SPAD, leaf water potential, osmotic potential.
2.1.1. Plant Growth Parameters
The part of the plant from the root collar to the growing tip was measured in cm (± 0.5) with a meter as plant height [67]. Plant samples from each replicate were weighed in grams on a precision scale [65]. After removing all leaves and the root, the remaining part was weighed in grams on a precision scale [68]. Leaf and stem dry weight was determined in grams on a precision scale after the leaf samples were dried in an oven at 65℃ for 48 hours [65]. Readings were taken with a Minolta SPAD meter (Minolta 502, Osaka, Japan) to determine the shade of green that varies with the amount of chlorophyll in the fifth leaves of pepper plants [69,70]. The number of branches was determined by counting all the branches on the plant [68]. The number of leaves was determined by counting all the leaves on the plant [71]. Leaf area was measured with the leaf area meter (Li-3100, LICOR, Lincoln, NE, USA) in square centimeters [65].
2.1.2. Ion Leakage (%)
In order to determine the ion leakage of plants under treatment conditions, leaf discs with a diameter of 1 cm were taken from fully developed and one sample of leaves. The leaf discs were washed in distilled water and dried gently with blotting paper. Three leaf discs were placed in each test tube and the tubes were shaken on a shaker for 4 hours. The ion leakage in each sample was determined using an EC meter (WTW pH/Cond 3320, Weilheim, Germany), and considered as the first measurement (EC1). Leaf disks in the same solution were autoclaved and ion leakage was determined at room temperature and considered as the final measurement (EC2). Ion leakage was calculated using the following formula [72,73].
Ion leakage (%) = (EC1/EC2) × 100
2.1.3. Leaf Relative Water Content (RWC)
Developed leaves from each replication were used to determine the relative water content of the leaves. First, 1 cm discs were taken from the leaves and the fresh weight (FW) was determined by weighing them on a precision scale [74]. Turgor weight (TW) was determined by keeping these leaf discs in distilled water for 4 hours. After the turgor weight was determined, the leaf discs were dried at 70°C for 24 hours. Dry weight (DW) was determined after the dried leaf discs were weighed on a precision scale [75,76].
RWC (%) = [(FW-DW)/(TW-DR)] × 100
2.1.4. Determination of Leaf Osmotic Potential (MPa)
After taking 1 g of the leaves of pepper plants (3-4 leaves), the leaf was homogenized with 19 g of pure water in a porcelain mortar. The homogenized sample was passed through a micro-filter and 50 μl was used in Gonatec brand osmometer, and readings were taken on Cryoscopic brand and Osmomat 030 model osmometer on the basis of freezing point. After the readings were read and recorded as Osmol/ kg, these values were converted into MPa units [69,77].
2.1.5. Leaf Water Potential (MPa)
2.1.6. Leaf Elemental Analysis
Leaf samples were washed in pure water. The washed samples were placed in paper bags and dried at 70 °C. Dry leaf samples were ground for mineral element analysis. The 0.2 g of each sample was weighed and the samples were burned in small glass bottles in a muffle furnace for 6 hours (550°C). The burnt samples were extracted with 2 mL 3.3% HCI and 18 mL H2O. Calcium (Ca), magnesium (Mg), potassium (K), sodium (Na), copper (Cu), manganese (Mn), iron (Fe) and zinc (Zn) were determined by atomic absorption spectrophotometer (Varian FS220, Las Vegas, NV, USA) [67]. The chlorine (Cl) concentration in leaf samples was determined using titrimetric analysis with silver nitrate (AgNO3) with the Mohr method [78].
2.1.7. Statistical Analysis
This study was carried out with control, NaCl treatment, bacteria, mycorrhiza, and bacteria+mycorrhiza treatment, NaCl treatment+bacteria, NaCl treatment+mycorrhiza and NaCl treatment+bacteria+mycorrhiza treatment. In climate chamber experiments was used randomized plots experimental design. Data from the results were subjected to analysis using JMP. Mean significant difference were compared by least significant difference (LSD) multiple range test at p≤0.05 using JMP 8 Software package (JMP®, Version 8. SAS Institute Inc., Cary, NC, 1989-2019). Percentage change of experiments results were determineted using MS Excel.
3. Results
The AMF application resulted in the highest plant height (39.50 cm), while the PGPR application (37.75 cm) and the control application (37.25 cm) exhibited similar results (Table 2). Particularly, salt stress has been identified to have a detrimental effect on plant height, with the lowest measurements recorded under salt stress conditions. However, both AMF and PGPR, either individually or in combination, have mitigated the adverse effects of salt stress. Salt stress has led to a significant reduction in plant height, recording a 34.90% decrease compared to the control group. In contrast, under saline conditions, the application of AMF demonstrated a significant increase of 21.65%, whereas the PGPR application exhibited a 9.28% increase. The combined application of AMF and PGPR under salt stress conditions resulted in a notable 12.37% increase in plant height. The difference between applications in terms of plant diameter was found to be statistically significant. The plant diameters for AMF, PGPR, and control applications were measured as 6.52 mm, 6.50 mm, and 6.37 mm, respectively. While the lowest plant diameter was 4.53 mm in salt stress, AMF, PGPR and AMF+PGPR applications alleviated the negative effect of salt stress. Plant diameter decreased by 28.89% in salt stress compared to the control. Plant diameter increased by 12.58% in salt+AMF application, 12.36% in salt+PGPR application and 5.52% in salt+AMF+PGPR application compared to salt application. The number of leaves was found to be statistically significant among the applications. The highest number of plant leaves was 45.00 in control application and the lowest number was 10 leaves in salt application. Under salt stress conditions, the number of leaves was 16.75, 14.75 and 11.00 in AMF, PGPR and AMF+PGPR applications, respectively (Table 2). Salt stress caused a significant decrease in leaf number and caused a 77.78% decrease compared to the control. Compared to salt application, salt+AMF, salt+PGPR and salt+AMF+PGPR applications increased leaf number by 67.50%, 47.50% and 10.00%, respectively. The highest bifurcation among the applications was determined to be in PGPR application with 7.00 per plant. Salt stress had a negative effect on bifurcation. AMF, PGPR and AMF+PGPR increased bifurcation compared to salt stress. Plant fresh weight changed between 59.44 g and 11.81 g among the applications. The lowest plant fresh weight was in salt+AMF+PGPR application with 11.81 g and the highest was in PGPR application with 59.44 g (Table 2). Under salt stress, plant fresh weight decreased by 71.87% compared to the control. However, salt+AMF and salt+PGPR applications caused an increase of 37.69% and 3.09%, respectively, while salt+AMF+PGPR application caused a decrease of 13.23% compared to salt stress. Both AMF and PGPR alone were effective in ameliorating the negative effect of salt application on leaf weight. The difference between applications in terms of plant dry weight was statistically significant. The highest plant dry weight was 6.29 g in PGPR application and the lowest was 1.74 g in salt+AMF+PGPR application.
The highest leaf area was in PGPR, AMF, Control and AMF+PGPR applications with 1837.25, 1553.25, 1450.75 and 1436.25 cm2/plant, respectively. The lowest leaf area was in salt application with 382.75 cm2/plant and decreased by 73.62% (Table 2).
3.1. Physiological Parameters
The difference between applications in terms of SPAD value was found statistically significant. Compared to the control (45.69), SPAD value increased in salt+PGPR (57.63), salt+AMF (56.73), salt (54.44) and salt+AMF+PGPR (51.30) applications but decreased in AMF (45.65), AMF+PGPR (43.58) and PGPR (41.58) applications (Figure 1). Leaf relative water content (RWC) was lowest in salt application at 72.69% and highest in AMF+PGPR application at 93.58%. It was higher in AMF+PGPR, AMF and PGPR applications compared to the control (Figure 2). RWC decreased by 18.31% under salt stress compared to the control. Moreover, it was observed that salt+AMF, salt+PGPR, and salt+AMF+PGPR applications increased leaf relative water content by 19.82%, 3.74%, and 3.32%, respectively, compared to salt stress. The highest ion leakage was in salt application with 55.40% and the lowest was in AMF application with 8.24%. Under salt stress conditions, AMF+PGPR reduced the ion leakage increased by 426.12% in salt stress compared to control. The ion leakages were decreased in salt+AMF+PGPR, salt+AMF ve salt+PGPR applications by 22.47%, 35.18%, and 53.14%, respectively (Figure 3). The lowest osmotic potential was in salt application with -7.97 Ψs and the highest was in AMF+PGPR application with -3.11 Ψs (Figure 4). Leaf osmotic potential increased by 105.70% in salt stress compared to control anddecreased by 38.14%, 34.50% and 30.24% in salt+PGPR, salt+AMF+PGPR and salt+AMF applications compared to salt stress.
3.2. Mineral Nutrient Uptake
Calcium, Mg, K, Na and Cl contents between applications were found statistically significant. The highest Ca content was in the control application with 1.058% and the lowest was in the AMF+PGPR application with 0.699%. Under salt stress conditions, Ca content decreased in AMF, PGPR and AMF+PGPR applications. In other words, these applications were ineffective under salt stress conditions (Table 3). The magnesium content of pepper plants varied between 1.100% and 1.334%. Mg content increased in all applications except for AMF+PGPR compared to the control. Under salt stress conditions, the AMF, PGPR and AMF+PGPR applications decreased Mg content compared to the salt application (Table 3). Under salt stress conditions, K increased in the AMF+PGPR application compared to the salt stress, which increased leaf Na content. The lowest leaf Na was in the PGPR application (0.22%). Leaf Na content decreased in salt+AMF (0.23%), salt+PGPR (3.0%), and salt+AMF+PGPR (2.99%) applications compared to salt stress (3.05%). Salt stress increased leaf chlorine content. The lowest leaf Cl content was in the AMF application and decreased in salt+AMF (2.27%), salt+PGPR (2.53%) and salt+AMF+PGPR application (2.63%) compared to salt stress (2.69%). Copper varied between 12.25 ppm and 18.00 ppm. Cu content increased in the salt+AMF+PGPR, salt+AMF and salt+PGPR applications and decreased in the AMF+PGPR, AMF and PGPR applications compared to the control The highest Mn content was 218.67 ppm in the salt+PGPR application and the lowest was 87.68 ppm in the control. Mn content increased in the PGPR application (218.64 ppm), while it decreased in AMF (161.00 ppm) and the AMF+PGPR application (185.50 ppm) compared to salt stress (188.33 ppm). Mn content increased in other applications compared to the control application (Table 4). Salt stress increased the leaf Zn content. Leaf Zn content was lowest in the AMF (125.25 ppm). Salt+AMF (160.33 ppm), salt+PGPR (171.00 ppm) and salt+AMF+PGPR applications (174.67 ppm) increased Zn content compared to the control application (131.67 ppm). Fe content varied between 90.50 ppm and 101.75 ppm. Fe content of the control and salt applications were close to each other. While Fe content was 100.50 ppm in the control application, it was 97.50 ppm, 89.00 ppm and 86.75 ppm in AMF, PGPR and AMF+PGPR applications, respectively (Table 4). Under salt stress conditions, PGPR, AMF and AMF+PGPR applications were insufficient to increase Fe content. Compared to the control, leaf Fe content increased by 1.24%, 4.32% and 10.67% in salt, AMF+PGPR and PGPR applications, respectively. Leaf Fe content decreased by 3.19%, 4.67% and 11.06% in salt+PGPR, salt+AMF and salt+AMF+PGPR compared to the salt application, respectively.
4. Discussion
Salt stress negatively affects various biological processes in plants, including photosynthesis, cell division, and overall growth, primarily by disrupting the balance of cellular ions. This disruption often leads to reduced chlorophyll levels, limiting photosynthetic efficiency and stunting plant growth [79]. High salinity increases osmotic pressure, causing plants to close their stomata, which restricts CO2 intake and further diminishes photosynthesis. The buildup of sodium and chloride ions in saline conditions also hinders growth. Plants under salt stress commonly show reduced height, which is linked to lower chlorophyll content [80]. However, research indicates that arbuscular mycorrhizal fungi (AMF) can effectively counteract these negative effects. AMF can improve stomatal conductance, enhancing CO2 uptake and reducing water loss. Plants inoculated with AMF generally have higher chlorophyll levels, boosting their photosynthetic capacity and resilience to salt stress. These fungi help by colonizing plant roots, improving water uptake and root hydraulic conductivity even in low water conditions. They promote root development and increase root surface area, which enhances the absorption of nutrients and water [81,82]. According to [38], increasing salt concentration from 1.5 dS m⁻¹ to 6 dS m⁻¹ led to a significant 30% reduction in plant height. In bell pepper plants exposed to salt stress levels of 1.5, 7.0, and 12.5 dS m⁻¹, treatments with both bacteria and fungi resulted in the highest stem length at 1.5 dS m⁻¹. Salt stress has been shown to negatively affect pepper plant height [83], with [84] noting that bacterial and fungal treatments effectively maintained plant height as salt concentrations increased. Studies also indicate that salt stress reduces plant growth, with a 23.3% decrease in stem diameter when salt concentration increased from 1.5 dS m⁻¹ to 6 dS m⁻¹; however, stem diameters under moderate salinity were similar to those in non-saline conditions [38]. Significant differences in stem diameter were observed under salt stress compared to the control groups [83]. The application of arbuscular mycorrhizal fungi (AMF), plant growth-promoting rhizobacteria (PGPR), and their combination (AMF+PGPR) effectively reduced the negative impacts of salt stress on pepper plant diameter [38]. Salt stress also decreases the leaf count in various pepper varieties. AMF application was found to be more effective than PGPR or their combination in mitigating the negative effects of salt stress on leaf number [85,86,87]. Under salt stress, combining AMF and PGPR alleviated these adverse effects. Rhizobacteria under salt stress may help reduce ethylene levels and promote plant growth by producing IAA-derived metabolites through ACC deaminase, as suggested by recent research [89].
Salt stress triggers the production of 1-aminochloropropane-1-carboxylate (ACC) in plant tissues, leading to increased ethylene levels that negatively impact root and shoot development. However, applying arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) can significantly boost plant fresh weight under saline conditions [88]. Rhizobacteria that produce ACC deaminase can lower ethylene levels, thereby promoting plant growth through IAA-derived metabolites [89]. Salt stress adversely affects shoot weight, but mycorrhizal inoculation helps mitigate this effect [90]. Mycorrhizae notably increase the fresh weight of pepper plants. Under salt stress, combinations of salt+AMF, salt+PGPR, and salt+AMF+PGPR improved leaf area by 51.53%, 17.57%, and 6.92%, respectively. Salt stress also reduces leaf area in pepper plants, which impacts fresh and dry weights and hampers biomass accumulation [87,91]. Plants adapt to saline environments through complex physiological changes; high osmotic pressure limits water uptake, causing increased water loss and slowed development. Reducing leaf area can help mitigate salinity effects by minimizing water loss and improving the plant's ability to cope with stress. The largest leaf area of pepper plants (1302.77 cm²) was observed under control conditions, while the smallest (101.29 cm²) occurred at 100 mM salinity stress [83].
Salinity, caused by high levels of Na⁺ and Cl⁻ ions, lowers soil water potential, making it harder for plants to absorb water and disrupting root membrane permeability. This hyperosmotic stress reduces water content in plant cells, decreases leaf water potential, and lowers turgor pressure, ultimately hindering cell division and growth [92,93]. Salt stress leads to decreased relative water content (RWC) mainly due to reduced water uptake and potential damage to cell walls [94,95,96]. Studies have shown that RWC decreases in pepper plants under saline conditions [97,98,99,100]. However, mycorrhizal species like Glomus mosseae and G. intraradices can improve RWC under salt stress, with G. intraradices performing better than G. mosseae or non-mycorrhizal plants [101]. Mycorrhizal inoculation was found to positively affect RWC, leaf length, and width. Mycorrhizal plants maintained higher RWC, reaching 81.10% compared to 60.01% in non-mycorrhizal plants [102]. The hyphal network of mycorrhizae increases root surface area, enhancing water and nutrient uptake while providing protection against soil pathogens and stress conditions [103]. Plants treated with arbuscular mycorrhizal fungi (AMF) have shown better water retention under salt stress due to improved root absorption and expanded root systems [104]. Additionally, plant growth-promoting rhizobacteria (PGPR) increase antioxidant enzyme activity under salt stress, helping to reduce oxidative stress and maintain RWC [105]. According to [106], both AMF and PGPR enhance mineral uptake and root system efficiency in saline conditions, which can improve RWC and support cellular turgor pressure.
Under stress conditions, decreased stomatal conductance [107] and reduced enzyme activity [92] can lead to CO2 buildup between cells, indicating potential damage to photosynthetic organs [108]. Mycorrhizal inoculation in tomato plants improved photosynthesis and CO2 assimilation through non-stomatal means. Furthermore, mycorrhizae significantly increased photosynthetic pigment levels in tomato leaves grown in saline-alkaline soil, enhancing the efficiency of light energy utilization [109]. In pepper plants treated with mycorrhiza, higher photosynthesis rates were associated with a larger leaf area [110]. Mycorrhizal symbiosis enhances assimilate accumulation, improves mineral nutrition, and boosts fruit quality [111]. Compared to the control group, treatments with salt+PGPR, salt+AMF+PGPR, salt+AMF, and salt alone showed varying osmotic potential values. The application of AMF, PGPR, and their combination reduced osmotic potential under salt stress. Salinity also causes stomata to close, limiting CO2 diffusion and disrupting photosynthesis, which increases reactive oxygen species (ROS) [92,93]. Salt stress negatively affects photosynthesis by damaging pigments, thylakoid membrane proteins, membrane lipids, and enzymes [79,91]. Stomatal closure limits CO2 entry into chloroplasts, reducing intercellular CO2 levels and photosynthesis rates [79]. This decrease in photosynthesis is mainly due to lower water potential, leading to stomatal closure and restricted CO2 diffusion because of reduced mesophyll conductivity [91,112]. In addition to reduced stomatal permeability, external factors can also limit photosynthesis by affecting the enzymes involved in CO2 uptake [112,113]. High salt concentrations significantly decrease chlorophyll content compared to control conditions. However, using mycorrhizae and rhizosphere bacteria can help mitigate the negative effects of salt stress on photosynthetic pigments. The decrease in chlorophyll under salt stress is often due to disrupted membrane structures and insufficient nutrients for chlorophyll production [108]. Sodium ions can reduce magnesium uptake, which negatively affects chlorophyll synthesis. Mycorrhizae can enhance magnesium uptake, thereby increasing chlorophyll production in saline conditions [114]. Salt stress can damage chlorophyll and other photosynthetic pigments, reducing their levels [115,116,117]. Studies have shown that applying mycorrhiza [118,119], rhizosphere bacteria [120], or a combination of both [121,122] helps preserve chlorophyll content. Under salt stress, chlorophyll levels decrease because the absorption of essential elements for its synthesis is inhibited [123]. However, the presence of AMF can improve nutrient absorption and reduce stomatal limitations, thus enhancing photosynthetic capacity [124]. Research shows that AMF can positively affect photosynthesis in cotton seedlings under salt-alkali stress. AMF colonization improves nutrient uptake from the soil, alleviates salt toxicity, and increases the availability of essential elements for chlorophyll synthesis, leading to better chlorophyll production and metabolism in the leaves [125].
Plants use several biochemical and molecular strategies to cope with high salinity stress, including managing ion balance, producing osmoprotectants, synthesizing antioxidants, and adjusting stress-related gene expression [126]. Osmolytes, which are electrically neutral and highly soluble, help maintain cell turgor pressure by increasing osmotic pressure in the cytoplasm [127,128]. Under salt stress, the application of arbuscular mycorrhizal fungi (AMF), plant growth-promoting rhizobacteria (PGPR), and their combination reduces ion leakage compared to stress alone. Salt stress can cause membrane damage, leading to leakage of soluble substances and increased electrical conductivity in tissues. High levels of proline help preserve membrane and protein structures and enhance calcium accumulation, which maintains high water potential in plants [38,129]. Reactive oxygen species, which form as a general response to stress, can oxidize membrane lipids, damaging membrane structures and reducing selective permeability [130]. Increased membrane damage leads to greater electrolyte leakage, which affects plasma membrane integrity [131,132]. Mycorrhizal treatments have been shown to lower membrane permeability under various NaCl concentrations [122], and bacterial treatments have also reduced membrane permeability in beans under salt stress compared to untreated controls [133].
Salt stress did not significantly affect iron (Fe) content in plants. However, applying arbuscular mycorrhizal fungi (AMF), plant growth-promoting rhizobacteria (PGPR), or their combination led to a slight reduction in Fe levels compared to the control group. Despite this, these treatments effectively increased copper (Cu) uptake, with leaf Cu content rising by 22.03% for both salt+AMF and salt+PGPR treatments, and by 13.56% for salt+AMF+PGPR. Copper content in pepper leaves generally decreases under salt stress. Salt stress can both stimulate and inhibit the uptake of trace elements in plants. For instance, while salt stress increased leaf zinc (Zn) content, combining AMF and PGPR significantly boosted Zn levels under salt stress. Leaf manganese (Mn) content also increased by 114.79% in salt-stressed plants compared to controls. However, Mn content decreased in plants treated with salt+AMF and salt+AMF+PGPR, while it increased by 16.11% in plants treated with salt+PGPR [134]. Enhanced root architecture from AMF inoculation improves the uptake of water and minerals, especially under drought conditions. Studies have shown that AMF inoculation can significantly boost the absorption of essential minerals, including nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), iron (Fe), zinc (Zn), and copper (Cu). Salt stress can lead to increased uptake of some micronutrients, reflecting a plant's adaptive response to salinity. Mycorrhizal plants generally have higher nutrient levels under moderate salt stress compared to non-mycorrhizal plants, highlighting the importance of AMF in optimizing mineral nutrition under challenging conditions. This improved nutrient absorption is due to the deeper penetration of AMF hyphae into the soil, allowing for more effective nutrient uptake. Potassium content decreased by 4.67%, 14.30%, 15.17%, and 18.11% in plants treated with AMF+PGPR, AMF, PGPR, and salt, respectively, compared to the control. Under salt stress, PGPR and AMF+PGPR treatments increased potassium content by 0.64% and 5.01%, while AMF treatment resulted in a 0.64% decrease. This reduction in potassium (K⁺) is associated with the buildup of reactive oxygen species, particularly hydrogen peroxide (H₂O₂), which, along with increased abscisic acid (ABA) and higher levels of sodium (Na⁺) and chloride (Cl⁻) in shoots, causes stomatal closure [135]. Mycorrhizal inoculation has been shown to enhance the uptake of potassium, calcium, phosphorus, and nitrogen in pepper plants irrigated with seawater [119,136,137]. Additionally, applying mycorrhiza under salt stress has been reported to increase the content of essential mineral elements and facilitate sodium ion storage in vacuoles. Leaf magnesium (Mg) content significantly increased by 17.70%, 14.91%, 14.55%, and 5.08% in the salt, PGPR, AMF+PGPR, and AMF treatments, respectively, compared to the control. The decrease in chlorophyll content under salt stress is linked to membrane damage and insufficient nutrient uptake needed for chlorophyll production [108]. This drop in chlorophyll can be attributed to reduced magnesium uptake due to the antagonistic effect of sodium ions. Mycorrhizae have been shown to enhance magnesium uptake, thus supporting chlorophyll synthesis in saline conditions [114]. Calcium is vital for plant growth, membrane stability, osmotic balance, and intracellular signaling, helping plants withstand various stress conditions. It acts as a secondary messenger during stress, including salinity [138,139]. In this study, leaf calcium (Ca) content increased by 1.03%, 32.39%, and 38.57% with AMF, PGPR, and AMF+PGPR treatments, respectively, while it decreased by 7.55% under salt stress compared to the control. Arbuscular mycorrhizal fungi (AMF) improve the availability of mineral nutrients, particularly phosphorus, which interacts with calcium, magnesium, and zinc [140]. Previous research showed a decrease in calcium content from 1.83 in the control to 1.75 under salt stress [141]. Many studies have found that NaCl application reduces calcium levels in pepper plants [85,142,143,144,145].
The accumulation of sodium (Na⁺) and chloride (Cl⁻) ions in plants competes with essential nutrients like potassium (K⁺), calcium (Ca²⁺), nitrogen (N), phosphorus (P), and magnesium (Mg), leading to nutrient deficiencies and imbalances [146,147]. Excess Na⁺ in the cytosol, due to its ease of passage through cell membranes, can cause Na⁺ toxicity, disrupting cell turgor pressure, membrane potential, and integrity, and impairing K⁺ uptake, which is vital for many enzymatic activities. This results in K⁺ deficiency and disrupted metabolic processes. A high cytosolic K⁺/Na⁺ ratio reflects a plant's ability to withstand salinity, suggesting that adding K⁺ to the growth medium can help alleviate Na⁺ toxicity [138,139,148]. In this study, leaf sodium (Na) content increased dramatically by 950.86% under salt stress compared to the control. However, the application of salt+PGPR, salt+AMF+PGPR, and salt+AMF reduced leaf Na content by 8.53%, 1.89%, and 1.48%, respectively. Previous studies have consistently shown that salt stress raises Na levels in plants [149,150,151]. Notably, bacterial inoculation during salt stress can increase the K⁺ ratio in pepper plants [152]. High sodium (Na⁺) concentrations in the root zone can interfere with calcium (Ca²⁺) uptake and transport, leading to lower Ca²⁺/Na⁺ ratios. This imbalance not only negatively affects plant growth but also causes various morphological and anatomical changes. Excess Na⁺ in the cytosol can compete with Ca²⁺, potentially replacing Ca²⁺ in the membrane and compromising membrane integrity [91]. Research shows that salt stress raises sodium levels in both the leaves and roots of pepper plants while lowering calcium and potassium levels [85]. High chloride (Cl⁻) levels in the cytosol can also disrupt chloroplast function and inhibit photosynthesis, leading to the production of reactive oxygen species (ROS). These ROS can damage important components like chlorophyll, causing leaf discoloration and necrotic lesions [153]. In our study, leaf Cl content increased by 88.93% under salt stress compared to the control. However, the application of salt+AMF, salt+PGPR, and salt+AMF+PGPR reduced leaf Cl content by 15.72%, 5.77%, and 2.33%, respectively. Other research has shown that bacteria such as Stenotrophomonas maltophilia and Serratia marcescens can reduce leaf Cl content under 50 and 100 mM salt conditions [2]. Similarly, inoculating with Azospirillum brasilense and Pantoea dispersa under salt stress has also been reported to lower leaf Cl content [152]. Notably, the highest chloride levels were observed in hot pepper varieties subjected to salt treatment [85].
5. Conclusions
In conclusion, the application of arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) significantly enhanced the growth and physiological resilience of pepper plants under salt stress. AMF treatment was particularly effective in increasing plant height, while PGPR also contributed to improved growth parameters, including plant diameter and leaf number. Both treatments successfully mitigated the adverse effects of salt stress, leading to better chlorophyll content, higher relative water content, and reduced ion leakage. Additionally, nutrient uptake, particularly of calcium and magnesium, improved with AMF, PGPR, and their combination. These findings highlight the potential of AMF and PGPR as valuable biostimulants in saline environments, offering promising strategies for sustainable agriculture. Further research could elucidate the underlying mechanisms and broaden their application in combating soil salinity.
Author Contributions
Conceptualization, S.D.; methodology, S.D.; software, S.D.; validation, S.D.; formal analysis, S.D.; investigation, S.D.; resources, S.D.; data curation, S.D.; writing—original draft preparation, S.D.; writing—review and editing, S.D.; visualization, S.D.; supervision, S.D.; project administration, S.D.; funding acquisition, S.D.. The author has read and agreed to the published version of the manuscript.
Funding
This research was funded by Sultan DERE. The APC was funded by Sultan DERE.
Data Availability Statement
No new data were created, but the data can be made available upon request.
Acknowledgments
The author would like to acknowledge H. Yildiz Dasgan for their support in this work.
Conflicts of Interest
The author declares no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Aydın, F.; Sarptaş, H. İklim değişikliğinin bitki yetiştiriciliğine etkisi: model bitkiler ile Türkiye durumu. PAJES. 2018, 24, 512–521. [Google Scholar] [CrossRef]
- Alveroğlu, L. The effects of some bacteria (Serratia marcescens and Stenotrophomonas maltophilia) and potassium nitrate applications to salt tolerance in pepper plants. Yüksek Lisans Tezi, Eskişehir Osmangazi Üniversitesi Fen Bilimleri Enstitüsü, Eskişehir, 2014.
- Gül, V. Stres Terminolojisi. Bitkilerde Abiyotik Ve Biyotik Stres Yönetimi. Iksad Publications: 2022.
- Kopecká, R.; Kameniarová, M.; Černý, M.; Brzobohatý, B.; Novák, J. Abiotic stress in crop production. Int. J. Mol. Sci. 2023, 24(7), 6603. [Google Scholar] [CrossRef] [PubMed]
- El Sabagh, A.; Islam, M.S.; Skalicky, M.; Ali Raza, M.; Singh, K.; Anwar Hossain, M.; Arshad, A. Salinity stress in wheat (Triticum aestivum L.) in the changing climate: Adaptation and management strategies. Front. Agron. 2021, 3, 661932. [Google Scholar]
- Çolak, G.; Keser, Ö.; Çatak, E. Tuz stresine maruz bırakılan raphanus sativus l.'de embriyonik yaprak (kotiledon) gelişimleri. EJONS, 2020, 4(16), 987-997.
- Dhiman, J.; Prasher, S.O.; El Sayed, E.; Patel, R.M.; Nzediegwu, C.; Mawof, A. Effect of hydrogel based soil amendments on yield and growth of wastewater irrigated potato and spinach grown in a sandy soil. Environ. Technol. Innov. 2021, 23, 101730. [Google Scholar] [CrossRef]
- Khamidov, M.; Ishchanov, J.; Hamidov, A.; Dönmez, C.; Djumaboev, K. Assessment of soil salinity changes under the climate change in the Khorezm region, Uzbekistan. Int. J. Environ. Res. Public Health 2022, 19(14), 8794. [Google Scholar] [CrossRef]
- Ullah, A.; Bano, A.; Khan, N. Climate Change and Salinity Effects on Crops and Chemical Communication Between Plants and Plant Growth-Promoting Microorganisms Under Stress. Front. Sustain. Food Syst. 2021, 5, 618092. [Google Scholar] [CrossRef]
- Kurt, C.H.; Tunçtürk, M.; Tunçtürk, R. Tuz stresi koşullarında yetiştirilen soya (Glycine max L.) bitkisinde bazı fizyolojik ve biyokimyasal değişimler üzerine salisilik asit uygulamalarının etkileri. Ege Univ. Ziraat Fak. Derg. 2023, 60(1), 91-101.
- Gürsoy, M. An overview of the effects of salt stress on plant development. In Proceedings of the 9th International Zeugma Conference on Scientific Research, Zeugma, Turkey; 2023. [Google Scholar]
- Aslan, M. Ö. Karnitin uygulamalarının tuz stresi altında biber ve hıyar tohumlarının çimlenme ve çıkış performansları üzerine olan etkilerinin incelenmesi. MSc Thesis, Kahramanmaraş Sütçü İmam Üniversitesi, Fen Bilimleri Enstitüsü, Bahçe Bitkileri Anabilim Dalı, Kahramanmaraş, Turkey, 2022. [Google Scholar]
- Mohammed, A.A.; Söylemez, S.; Sarhan, T.Z. Effect of biofertilizers, seaweed extract and inorganic fertilizer on growth and yield of lettuce (Lactuca sativa var. longifolia L.). Harran Tarım ve Gıda Bilimleri Dergisi 2022, 26(1), 60-71.
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional Regulators of Plant Growth, Development, and Stress Responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-plant interactions: strategies ensuring plant growth promotion under drought and salinity stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.K.; Singh, P.P.; Kumar, A. Interaction of plant growth promoting bacteria with tomato under abiotic stress: a review. Agric. Ecosyst. Environ. 2018, 267, 129–140. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Luo, Y.M.; Freitas, H. Inoculation of endophytic bacteria on host and non-host plants – effects on plant growth and Ni uptake. J. Hazard. Mater. 2011, 195, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24(13), 2452. [Google Scholar] [CrossRef] [PubMed]
- Azeem, M.; Pirjan, K.; Qasim, M.; Mahmood, A.; Javed, T.; Muhammad, H.; Rahimi, M. Salinity stress improves antioxidant potential by modulating physio-biochemical responses in Moringa oleifera Lam. Sci. Rep. 2023, 13, 2895. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.; Pal, N.; Dey, S.; Mandal, A. Characterizations of surfactant synthesized from palm oil and its application in enhanced oil recovery. J. Taiwan Inst. Chem. Eng. 2017, 81, 343–355. [Google Scholar] [CrossRef]
- Naamala, J.; Subramanian, S.; Msimbira, L.A.; Smith, D.L. Effect of NaCl stress on exoproteome profiles of Bacillus amyloliquefaciens EB2003A and Lactobacillus helveticus EL2006H. Front. Microbiol. 2023, 14, 1206152. [Google Scholar] [CrossRef]
- Zhang, L.; Freschi, G.; Rouphael, Y.; Pascale, S.; Lucini, L. The differential modulation of secondary metabolism induced by a protein hydrolysate and a seaweed extract in tomato plants under salinity. Front. Plant Sci. 2023, 13. [Google Scholar] [CrossRef]
- Dere, S. Organic substances application to enhance abiotic stress tolerance in plants. Iksad Yayinevi: 2020; pp. 1.
- Alkaç, O.S.; Tuncel, M.E.; Dinçer, E.; Donat, A. The effect of compost, bacteria, and mycorrhiza applications on the phytochemical contents of edible tulip petals. Turkish J. Agric. Food Sci. Technol. 2023, 11(3), 470–477. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Inbaraj, M.P. Plant-microbe interactions in alleviating abiotic stress—A mini review. Front. Agron. 2021, 3, 667903. [Google Scholar] [CrossRef]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Mustafa, A. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci. 2021, 22(19), 10529. [Google Scholar] [CrossRef]
- Adedayo, A.A.; Babalola, O.O. Fungi That Promote Plant Growth in the Rhizosphere Boost Crop Growth. J. Fungus. 2023, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.A.; Islam, S.; Gani, G.; Dar, Z.M.; Masood, A.; Baligah, S.H. AM fungi as a potential biofertilizer for abiotic stress management. in arbuscular mycorrhizal fungi in agriculture-new insights. IntechOpen. 2023. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Ruíz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi: a review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Li, Z.; Wu, N.; Meng, S.; Wu, F.; Liu, T. Arbuscular mycorrhizal fungi (AMF) enhance the tolerance of Euonymus maackii Rupr. at a moderate level of salinity. PLoS One 2020, 15(4), e0231497. [Google Scholar]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: current understanding and new challenges. Front. Plant Sci. 2019, 470. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Chanratana, M.; Kim, K.; Seshadri, S.; Sa, T. Impact of arbuscular mycorrhizal fungi on photosynthesis, water status, and gas exchange of plants under salt stress—a meta-analysis. Front. Plant Sci. 2019, 10, 457. [Google Scholar] [CrossRef]
- Dastogeer, K.M.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant salinity tolerance conferred by arbuscular mycorrhizal fungi and associated mechanisms: a meta-analysis. Front. Plant Sci. 2020, 11, 588550. [Google Scholar] [CrossRef]
- Balliu, A.; Sallaku, G.; Rewald, B. AMF inoculation enhances growth and improves the nutrient uptake rates of transplanted, salt-stressed tomato seedlings. Sustainability. 2015, 7, 15967–15981. [Google Scholar] [CrossRef]
- Liu, M.Y.; Li, Q.S.; Ding, W.Y.; Dong, L.W.; Deng, M.; Chen, J.H.; Wu, Q.S. Arbuscular mycorrhizal fungi inoculation impacts expression of aquaporins and salt overly sensitive genes and enhances tolerance of salt stress in tomato. Chem. Biol. Technol. Agric. 2023, 10(1), 5. [Google Scholar] [CrossRef]
- Altunlu, H. 2020. The Effects of mycorrhiza and rhizobacteria application on growth and some physiological parameters of pepper (Capsicum annuum L.) under salt stress. J. Agric. Fac. Ege Univ. 2020, 57, 501–510. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones, and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25(6), 1102–1114. [Google Scholar] [CrossRef]
- Mohammad, A.; Mittra, B. Effects of inoculation with stress-adapted arbuscular mycorrhizal fungus Glomus deserticola on growth of Solanum melongena L. and Sorghum sudanese Staph. seedlings under salinity and heavy metal stress conditions. Arch. Agron. Soil Sci. 2013, 59(2), 173–183. [Google Scholar]
- Abdulhadi, S.A.A. Effects of arbuscular mycorrhizal fungi on seedling development of edible seed pumpkin under salty soil conditions. Master’s Thesis, The Graduate School of Natural and Applied Science of Selçuk University, 2017; 35 pp.
- Sharma, N.; Aggarwal, A.; Yadav, K. Arbuscular mycorrhizal fungi enhance growth, physiological parameters and yield of salt stressed Phaseolus mungo (L.) Hepper. Eur. J. Environ. Sci. 2017, 7(1), 22-27.
- Wang, H.; An, T.; Huang, D.; Liu, R.; Xu, B.; Zhang, S.; Chen, Y. Arbuscular mycorrhizal symbioses alleviating salt stress in maize is associated with a decline in root-to-leaf gradient of Na+/K+ ratio. BMC Plant Biol. 2021, 21(1), 1–15. [Google Scholar] [CrossRef]
- Santander, C.; Sanhueza, M.; Olave, J.; Borie, F.; Valentine, A.; Cornejo, P. Arbuscular mycorrhizal colonization promotes the tolerance to salt stress in lettuce plants through an efficient modification of ionic balance. J. Soil Sci. Plant Nutr. 2019, 19, 321–331. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Ben-Laouane, R.; Slimani, A.; Boutasknit, A.; Anli, M.; Oufdou, K.; Meddich, A. Autochthonous biostimulants as a promising biological tool to promote lettuce growth and development under salinity conditions. Environ. Sci. Proc. 2022a, 16(1), 41. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Ben-Laouane, R.; Lahlali, R.; Anli, M.; Ikan, C.; Boutasknit, A.; Meddich, A. Application of indigenous rhizospheric microorganisms and local compost as enhancers of lettuce growth, development, and salt stress tolerance. Microorganisms 2022b, 10(8), 1625. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key players in sustainable agriculture by improving soil fertility, plant tolerance, and crop productivity. Microb. Cell Fact. 2014, 13, 1–10. [Google Scholar] [CrossRef]
- Etesami, H.; Emami, S.; Alikhani, H.A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects—a review. J. Soil Sci. Plant Nutr. 2017, 17(4), 897–911. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J. S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Smith, D.L. Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Gómez-Godínez, L.J.; Aguirre-Noyola, J.L.; Martínez-Romero, E.; Arteaga-Garibay, R.I.; Ireta-Moreno, J.; Ruvalcaba-Gómez, J.M. A Look at Plant-Growth-Promoting Bacteria. Plants 2023, 12(8), 1668. [Google Scholar] [CrossRef]
- Nadeem, N.; Asif, R.; Ayyub, S.; Salman, S.; Shafique, F.; Ali, Q.; Malik, A. Role of rhizobacteria in phytoremediation of heavy metals. Biol. Clin. Sci. Res. J. 2020, 2020(1).
- Ahemad, M.; Khan, M.S. Productivity of greengram in tebuconazole–stressed soil, by using a tolerant and plant growth-promoting Bradyrhizobium sp. MRM6 strain. Acta Physiol. Plant. 2012, 34, 245–254. [Google Scholar] [CrossRef]
- Armenova, N.; Tsigoriyna, L.; Arsov, A.; Petrov, K.; Petrova, P. Microbial detoxification of residual pesticides in fermented foods: current status and prospects. Foods. 2023, 12, 1163. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-tolerant plant growth-promoting rhizobacteria for enhancing crop productivity of saline soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant growth-promoting bacteria: biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 2020a, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Tirry, N.; Kouchou, A.; Laghmari, G.; Lemjereb, M.; Hnadi, H.; Amrani, K.; El Ghachtouli, N. Improved salinity tolerance of Medicago sativa and soil enzyme activities by PGPR. Biocatalysis Agric. Biotechnol. 2021, 31, 101914. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant associated rhizobacteria for biocontrol and plant growth enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef]
- Bhojiya, A.A.; Joshi, H.; Upadhyay, S.K.; Srivastava, A.K.; Pathak, V.V.; Pandey, V.C.; Jain, D. Screening and optimization of zinc removal potential in Pseudomonas aeruginosa-HMR1 and its plant growth-promoting attributes. Bull. Environ. Contam. Toxicol. 2022, 108(3), 468–477. [Google Scholar] [CrossRef]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms 2023, 11(4), 1088. [Google Scholar] [CrossRef]
- Bhanse, P.; Kumar, M.; Singh, L.; Awasthi, M.K.; Qureshi, A. Role of plant growth-promoting rhizobacteria in boosting the phytoremediation of stressed soils: Opportunities, challenges, and prospects. Chemosphere 2022, 303, 134954. [Google Scholar] [CrossRef]
- Vocciante, M.; Grifoni, M.; Fusini, D.; Petruzzelli, G.; Franchi, E. The role of plant growth-promoting rhizobacteria (PGPR) in mitigating plant’s environmental stresses. Appl. Sci. 2022, 12, 1231. [Google Scholar] [CrossRef]
- Kumar, N.; Tahmish, F.; Mishra, J.; Mishra, I.; Verma, S.; Verma, R.; Vermad, M.; Bhattacharya, A.; Verma, P.; Mishra, P.; Bharti, C. Halo-tolerant plant growth promoting rhizobacteria for improving productivity and remediation of saline soils. J. Adv. Res. 2020b, 26, 69–82. [Google Scholar] [CrossRef]
- Sarıyer, T.; Çiftci, H.N. Effect of different storage periods on quality properties in pepper (Capsicum annuum L. cv. Oskar F1). COMU J. Agric. Faculty 2022, 10(1), 137-142.
- López-Serrano, L.; Calatayud, Á.; López-Galarza, S.; Serrano, R.; Bueso, E. Uncovering salt tolerance mechanisms in pepper plants: a physiological and transcriptomic approach. BMC Plant Biol. 2021, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dasgan, H.Y.; Yilmaz, M.; Dere, S.; Ikiz, B.; Gruda, N.S. Bio-Fertilizers Reduced the Need for Mineral Fertilizers in Soilless-Grown Capia Pepper. Horticulturae 2023, 9(2), 188. [Google Scholar] [CrossRef]
- Schröder, F.G.; Lieth, J.H. Irrigation control in hydroponics. In Hydroponic Production of Vegetables and Ornamentals; Savvas, D., Passam, H., Eds.; Embryo Publications: Athens, Greece, 2002; pp. 263–298. [Google Scholar]
- Dere, S. Domateste besin özellikleri ve kalitenin kuraklığa dayanıklılıkla arttırılması. Doctoral Thesis, Çukurova Üniversitesi Fen Bilimleri Enstitüsü, Adana, 2019. [Google Scholar]
- Yilmaz, M. Topraksız biber yetiştiriciliğinde mikoriza ve bakteri biyogübreleri kullanılarak mineral gübrelerin azaltılması. MSc Thesis, Çukurova University, Adana, Turkey, 2020. [Google Scholar]
- Dasgan, H.Y.; Kuşvuran, Ş.; Abak, K.; Sarı, N. Screening and saving of local vegege tarımsal araştırma enstitüsübles for their resistance to drought and salinity. UNDP Project Final Report, 2010.
- Altuntas, O.; Dasgan, H.Y.; Akhoundnejad, Y.; Kutsal, I.K. Does Silicon Increase the tolerance of a sensıtıve pepper genotype to salt stress? Acta Sci. Pol. Hortorum Cultus. 2020, 19, 87–96. [Google Scholar] [CrossRef]
- Dere, S. Kuraklık stresi koşullarında bakteri uygulamasının domates bitkileri üzerine etkileri. Türk Doğa ve Fen Dergisi 2021, 10(1), 52-62.
- Bayram, M.; Dasgan, H.Y. (2020). Useful parameters and correlations for screening of tomatoes to salt tolerance under field conditions. Russ. Agric. Sci. 2020, 46, 451–458. [Google Scholar]
- Altuntas, O.; Dasgan, H.Y.; Akhoundnejad, Y. Silicon-induced salinity tolerance improves photosynthesis, leaf water status, membrane stability, and growth in pepper (Capsicum annuum L.). Hortic. Sci. 2018, 53, 1820–1826. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Mazrou, Y.S.; Hafez, Y.M. Silicon foliar application mitigates salt stress in sweet pepper plants by enhancing water status, photosynthesis, antioxidant enzyme activity, and fruit yield. Plants 2020, 9, 733. [Google Scholar] [CrossRef]
- Sadasivam, V.; Packiaraj, G.; Subiramani, S.; Govindarajan, S.; Kumar, G.P.; Kalamani, V.; Vemuri, L.; Narayanasamy, J. Evaluation of seagrass liquid extract on salt stress alleviation in tomato plants. Asian J. Plant Sci. 2017, 16, 172–183. [Google Scholar]
- Akhoundnejad, Y.; Baran, S. Boosting drought resistance in pepper (Capsicum annuum L.) with the aid of arbuscular mycorrhizal fungi and key phytohormones. Hortic. Sci. 2023, 58, 1358–1367. [Google Scholar]
- Coban, A.; Akhoundnejad, Y.; Dere, S.; Dasgan, H.Y. Impact of salt-tolerant rootstock on the enhancement of sensitive tomato plant responses to salinity. HortScience 2020, 55(1), 35–39. [Google Scholar] [CrossRef]
- Daşgan, H.Y.; Bayram, M.; Kuşvuran, S.; Çoban, G.A.; Akhoundnejad, Y. Screening of tomatoes for their resistance to salinity and drought stress. Glob. J. Biol. Agric. Health Sci. 2018, 8(24).
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7(6), Art. no: 132.
- Yavas, I.; Ilker, E. Changes in photosynthesis and phytohormone levels in plants exposed to environmental stress conditions. Agric. 2020, 267, 129–140. [Google Scholar]
- Abdel Latef, A.A.H.; Chaoxing, H. Does inoculation with Glomus mosseae improve salt tolerance in pepper plants? . J. Plant. Growth. Regul. 2014, 33, 644–653. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L. ) plants exposed to salinity. Environ. Exp. Bot. 2014, 98, 20–31. [Google Scholar] [CrossRef]
- Badem, A.; Söylemez, S. Effects of nitric oxide and silicon application on growth and productivity of pepper under salinity stress. J. King Saud Univ. Sci. 2022, 34(6), 102189. [Google Scholar] [CrossRef]
- Rayavarapu, V.B.; Padmavathi, T. Effect of Bacillus sp. and Glomus monosporum on growth and antioxidant activity of bell pepper (Capsicum annuum) under salinity stress. J. Glob. Agric. Ecol. 2016, 6(1), 57-67.
- Kadan, H.Y.; Üzal, Ö. Tuz stresi ve geri kazanım sürecinde biberin (Capsicum annuum L.) bitki gelişimi ve iyon alımındaki değişimler. J. Inst. Sci. Technol. 2020, 10(3), 1476-1485.
- Yaşar, F.; Yaşar, Ö. Tuz stresi altındaki çarliston biber çeşidinin gelişim performansı. ISPEC J. Agric. Sci. 2022, 6(4), 835–841. [Google Scholar]
- Kaymak, M.A. Grafen oksit uygulamalarının biber (Capsicum annuum L.)'de tuz stresine karşı etkileri. MSc Thesis, Atatürk Üniversitesi, Erzurum, Turkey, 2023. [Google Scholar]
- Trung, N.T.; Hieu, H.V.; Thuan, N.H. Screening of strong 1-Aminocyclopropane-1-Carboxylate deaminase producing bacteria for improving the salinity tolerance of cowpea. Appl. Microbiol.: Open Access.
- Safronova, V.I.; Stepanok, V.V.; Engqvist, G.L.; Alekseyev, Y.V.; Belimov, A.A. Root-associated bacteria containing 1-aminocyclopropane-1-carboxylate deaminase improve growth and nutrient uptake by pea genotypes cultivated in cadmium supplemented soil. Biol. Fertil. Soils 2006, 42(3), 267–272. [Google Scholar] [CrossRef]
- Al-Amri, S.M. Mitigation of salinity stress of pepper (Capsicum annuum l.) by arbuscular mycorrhizal fungus, glomus constrictum. Appl Ecol Environ Res. 2019. 17, 9965-9978. [CrossRef]
- Sevgi, B.; Leblebici, S. Tuz stresinin bitkiler üzerindeki etkileri ve geliştirilen tolerans mekanizmaları. Düzce Üniversitesi Bilim ve Teknoloji Dergisi 2023, 11(3), 1498-1516.
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25(2), 239–250. [Google Scholar] [CrossRef]
- Shahzad, B.; Fahad, S.; Tanveer, M.; Saud, S.; Khan, I.A. Plant responses and tolerance to salt stress. In Approaches for Enhancing Abiotic Stress Tolerance in Plants, 1st ed.; CRC Press: Florida, USA, 2019; pp. 61–78. [Google Scholar]
- Parvin, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Mohsin, S.M.; Fujita, M. Comparative physiological and biochemical changes in tomato (Solanum lycopersicum L.) under salt stress and recovery: Role of antioxidant defense and glyoxalase systems. Antioxidants 2019, 8, 350. [Google Scholar]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Díaz Vivancos, P.; Sánchez-Blanco, M.; Hernández, J. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Hafez, Y.M.; El-Afry, M.M.; Tantawy, D.S.; Alshaal, T. Effect of some osmoregulators on photosynthesis, lipid peroxidation, antioxidative capacity, and productivity of barley (Hordeum vulgare L. ) under water deficit stress. Environ. Sci. Pollut. Res. 2018, 25, 30199–30211. [Google Scholar] [CrossRef]
- Cicek, N.; Çakirlar, H. The effect of salinity on some physiological parameters in two maize cultivars. Bulg. J. Plant Physiol. 2002, 28(1-2), 66-74.
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47(1), 39–50. [Google Scholar] [CrossRef]
- El-Tayeb, M.A. Response of barley grains to the interactive effect of salinity and salicylic acid. Plant Growth Regul. 2005, 45, 215–224. [Google Scholar] [CrossRef]
- Aktas, H.; Abak, K.; Eker, S. Anti-oxidative responses of salt-tolerant and salt-sensitive pepper (Capsicum annuum L.) genotypes grown under salt stress. J. Hortic. Sci. Biotechnol. 2012, 87(4), 360-366.
- Çekiç, F.Ö.; Ünyayar, S.; Ortaş, İ. Effects of arbuscular mycorrhizal inoculation on biochemical parameters in Capsicum annuum grown under long term salt stress. Turkish J. Bot. 2012, 36, 63–72. [Google Scholar] [CrossRef]
- Basak, H.; Çimrin, K.M.; Turan, M.; Güneş, A.; Ozlu, E. Response of mycorrhiza-inoculated pepper and amino acids to salt treatment at different ratios. Commun. Soil Sci. Plant Anal. 2019, 50(3), 350–361. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32(2), 429–448. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104(7), 1263–1280. [Google Scholar] [CrossRef]
- Powles, S.B. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol. 1984, 35(1), 15–44. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza 2008, 18(6), 287–296. [Google Scholar] [CrossRef]
- Zhang, F.; Jia-Dong, H.E.; Qiu-Dan, N.I.; Qiang-Sheng, W.U.; Ying-Ning, Z.O.U. Enhancement of drought tolerance in trifoliate orange by mycorrhiza: changes in root sucrose and proline metabolisms. Notulae Botanicae Horti Agro-botanici Cluj-Napoca 2018, 46(1), 270–276. [CrossRef]
- Altuntas, O.; Dasgan, H.Y.; Akhoundnejad, Y.; Nas, Y. Unlocking the potential of pepper plants under salt stress: mycorrhizal effects on physiological parameters related to plant growth and gas exchange across tolerant and sensitive genotypes. Plants 2024, 13(10), 1380. [Google Scholar] [CrossRef]
- Nzanza, B.; Marais, D.; Soundy, P. Effect of arbuscular mycorrhizal fungal inoculation and biochar amendment on growth and yield of tomato. Int. J. Agric. Biol. 2012, 14, 965–969. [Google Scholar]
- Flexas, J.; Bota, J.; Galmes, J.; Medrano, H.; Ribas-Carbó, M. Keeping a positive carbon balance under adverse conditions: responses of photosynthesis and respiration to water stress. Physiologia Plantarum 2006, 127(3), 343–352. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany 2009, 103(4), 551–560. [Google Scholar] [CrossRef] [PubMed]
- Giri, B.; Mukerji, K.G. Mycorrhizal inoculant alleviates salt stress in Sesbania aegyptiaca and Sesbania grandiflora under field conditions: evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 2004, 14(5), 307–312. [Google Scholar] [CrossRef] [PubMed]
- Kusvuran, S.; Dasgan, H.Y.; Abak, K. Responses of okra genotypes to drought stress: VII. Vegetable. In Agricultura Sempozium, 2008, pp. 329-333.
- İzci, B. Pamukta (Gossypium hirsutum L.) farklı tuz konsantrasyonlarının in vitro koşullarda fotosentetik pigmentler üzerine etkisi. Alinteri J. Agric. Sci. 2009, 17(2), 7-13.
- Demirel, K.; Genç, L.; Mendeş, M.; Saçan, M.; Kızıl, Ü. Estimation of growth curve parameters for pepper (Capsicum annuum cv. Kapija) under deficit irrigation conditions. Ege Üniv. Ziraat Fak. Derg. 2012, 49(1), 37-43.
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Tullio, M.; Rivera, C.M.; Rea, E. Alleviation of salt stress by arbuscular mycorrhizal in zucchini plants grown at low and high phosphorus concentration. Biol. Fertil. Soils 2008, 44(3), 501–509. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Sönmez, Ö.; Aydemir, S.; Tuna, A.L.; Cullu, M.A. The influence of arbuscular mycorrhizal colonisation on key growth parameters and fruit yield of pepper plants grown at high salinity. Sci. Hortic. 2009, 121(1), 1–6. [Google Scholar] [CrossRef]
- Karlıdağ, H.; Yıldırım, E.; Turan, M.; Pehluvan, M.; Dönmez, F. Plant growth-promoting rhizobacteria mitigate deleterious effects of salt stress on strawberry plants (Fragaria × ananassa). HortScience 2013, 48(5), 563–567. [Google Scholar] [CrossRef]
- Badizi, B.M.; Zarandi, M.M. Some physiological and growth parameters of Pistachio vera L. under coinoculation with endomycorrhizae and Bacillus subtilis in response to salinity. Bull. Env. Pharmacol. Life Sci. 2016, 1, 70–77. [Google Scholar]
- Koç, A.; Balcı, G.; Ertürk, Y.; Keles, H.; Bakoglu, N.; Ercisli, S. Influence of arbuscular mycorrhizae and plant growth promoting rhizobacteria on proline, membrane permeability and growth of strawberry (Fragaria x ananassa) under salt stress. J. Appl. Bot. Food Qual. 2016, 89, 77–89. [Google Scholar] [CrossRef]
- Abdel Latef, A.A. Changes of antioxidative enzymes in salinity tolerance among different wheat cultivars. Cereal Res. Commun. 2010, 38, 43–55. [Google Scholar] [CrossRef]
- Haro, H.; Sanon, K.B.; Le Roux, C.; Duponnois, R.; Traoré, A.S. Improvement of cowpea productivity by rhizobial and mycorrhizal inoculation in Burkina Faso. Symbiosis 2018, 74(2), 107–120. [Google Scholar] [CrossRef]
- Ma, L.; Liu, X.; Lv, W.; Yang, Y. Molecular mechanisms of plant responses to salt stress. Front. Plant Sci. 2022, 13, 934877. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global plant-responding mechanisms to salt stress: Physiological and molecular levels and implications in biotechnology. Crit. Rev. Biotechnol. 2015, 35(4), 425–437. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Saxena, J.; Nema, R.; Singh, D.; Gupta, A. Phytochemistry of medicinal plants. J. Pharmacogn. Phytochem. 2013, 1(6), 168–182. [Google Scholar]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251(1), 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Gomaa, E. Effect of plant growth promoting Bacillus subtilis and Pseudomonas fluorescens on growth and pigment composition of radish plants (Raphanus sativus) under NaCl stress. Photosynthetica 2012, 50, 263–272. [Google Scholar] [CrossRef]
- Ekinci, M.; Yildirim, E.; Turan, M. Ameliorating effects of hydrogen sulfide on growth, physiological and biochemical characteristics of eggplant seedlings under salt stress. South Afr. J. Bot. 2021, 143, 79–89. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. Plant Physiol. Biochem. 2018a, 132, 375–384. [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.A.; Ali, H.M.; Alayafi, A.A.; et al. Serratia liquefaciens KM4 improves salt stress tolerance in maize by regulating redox potential, ion homeostasis, leaf gas exchange and stress-related gene expression. Int. J. Mol. Sci. 2018b, 19, 3310. [Google Scholar] [CrossRef]
- Ahmad, M.; Zahir, Z.A.; Asgha, H.N.; Arshad, M. The combined application of rhizobial strains and plant growth promoting rhizobacteria improves growth and productivity of mung bean (Vigna radiata L.) under salt-stressed conditions. Ann. Microbiol. 2012, 62, 1321–1330. [Google Scholar]
- Bora, M. Değişik vejetasyon dönemlerine kadar uygulanan farklı tuz konsantrasyonlarının biberde meydana getirdiği fizyolojik, morfolojik ve kimyasal değişikliklerin belirlenmesi. Yüksek Lisans Tezi, Namık Kemal Üniversitesi Fen Bilimleri Enstitüsü, Tekirdağ, 2015.
- Hedrich, R.; Shabala, S. Stomata in a saline world. Curr. Opin. Plant Biol. 2018, 46, 87–95. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, S.; Mishra, S. Influence of arbuscular mycorrhizal (AM) fungi and salinity on seedling growth, solute accumulation and mycorrhizal dependency of Jatropha curcas L. J. Plant Growth Regul. 2010, 29, 297–306. [Google Scholar] [CrossRef]
- Scharnagl, K.; Sanchez, V.; Von Wettberg, E. The impact of salinity on mycorrhizal colonization of a rare legume, Galactia smallii, in South Florida pine rocklands. BMC Res. Notes 2018, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal. Behav. 2010, 5(3), 233–238. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A new insight of salt stress signaling in plants. Molecules Cells 2016, 39(6), Art. no: 447.
- Al-Karaki, G.N.; Hammad, R.; Rusan, M. Response of two tomato cultivars differing in salt tolerance to inoculation with mycorrhizal fungi under salt stress. Mycorrhiza 2001, 11, 43–47. [Google Scholar] [CrossRef]
- Tuna, A.L.; Eroğlu, B. Tuz stresi altındaki biber (Capsicum annuum L.) bitkisinde bazı organik ve inorganik bileşiklerin antioksidatif sisteme etkileri. Anadolu J. Agric. Sci. 2017, 32(1), 121-131. [CrossRef]
- Güneş, A.; Inal, A.; Alpaslan, M. Effect of salinity on stomatal resistance, proline, and mineral composition of pepper. J. Plant Nutr. 1996, 19(2), 389–396. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Martínez, V.; Carvajal, M. Osmotic adjustment, water relations and gas exchange in pepper plants grown under NaCl or KCl. Environ. Exp. Bot. 2004, 52(2), 161–174. [Google Scholar] [CrossRef]
- Lycoskoufis, I.H.; Savvas, D.; Mavrogianopoulos, G. Growth, gas exchange, and nutrient status in pepper (Capsicum annuum L.) grown in recirculating nutrient solution as affected by salinity imposed to half of the root system. Sci. Hortic. 2005, 106(2), 147-161. [CrossRef]
- Arslan, M.G. Tuz Stresi Altındaki Acı ve Çarliston Biber Çeşitlerinin Genç-Orta ve Yaşlı Yapraklarında İyon Hareketleri. MSc Thesis, Van Yüzüncü Yıl Üniversitesi, Fen Bilimleri Enstitüsü, Bahçe Bitkileri Anabilim Dalı, Van, Turkey, 2022. [Google Scholar]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant salt stress: Adaptive responses, tolerance mechanisms and bioengineering for salt tolerance. Bot. Rev. 2016, 82(4), 371–406. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22(6), 4056–4075. [Google Scholar] [CrossRef]
- Wu, H. Plant salt tolerance and Na⁺ sensing and transport. Crop J. 2018, 6(3), 215–225. [Google Scholar] [CrossRef]
- Kusvuran, S. Relationships between physiological mechanisms of tolerance to drought and salinity in melons. Ph.D. Thesis, Department of Horticulture, Institute of Natural and Applied Sciences, Çukurova University, Adana, Turkey, 2010; p. 356. [Google Scholar]
- Esin, F. Bazı çilek çeşitlerinde NaCl uygulamasının bitki gelişimi ve iyon içeriği üzerine etkisi. Yüksek Lisans Tezi, Yüzüncü Yıl Üniversitesi Fen Bilimleri Enstitüsü, Van, 2007.
- Kusvuran, Ş.; Ellialtıoğlu, Ş.; Abak, K.; Yaşar, F. Bazı kavun (Cucumis sp.) genotiplerinin tuz stresine tepkileri. Ankara Üniversitesi Biyoteknoloji Enstitüsü, 2002; Proje No: 2002-58, Ankara.
- Del Amor, F.M.; Cuadra-Crespo, F. Plant growth-promoting bacteria as a tool to improve salinity tolerance in sweet pepper. Funct. Plant Biol. 2011, 39(1), 82–90. [Google Scholar] [CrossRef]
- Geilfus, C.M. Chloride: From nutrient to toxicant. Plant Cell Physiol. 2018, 59(6), 1132–1140. [Google Scholar] [CrossRef]
Figure 1.
Effects of the treatments on SPAD values of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p < 0.05).
Figure 1.
Effects of the treatments on SPAD values of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p < 0.05).
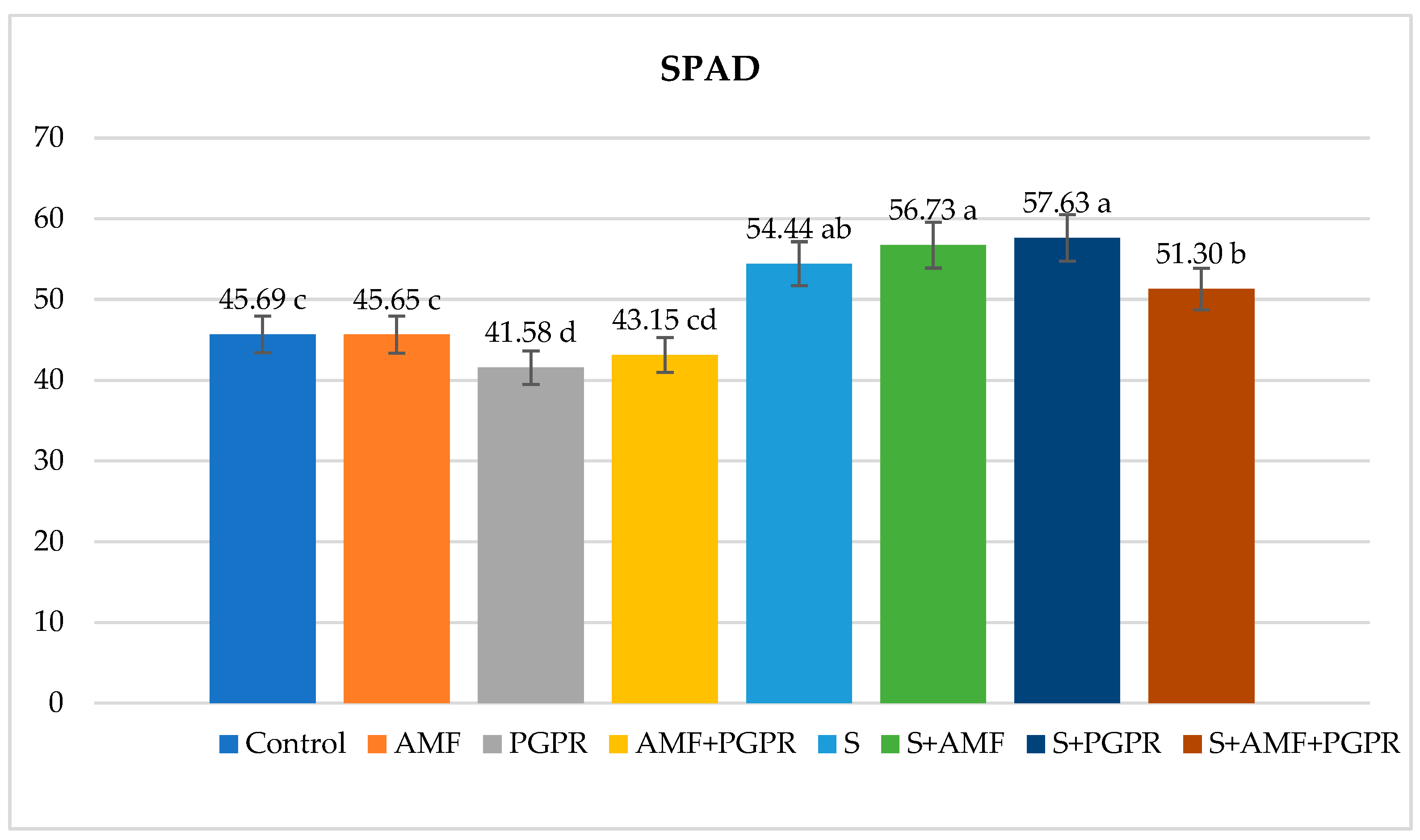
Figure 2.
Effects of the treatments on RWC of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
Figure 2.
Effects of the treatments on RWC of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
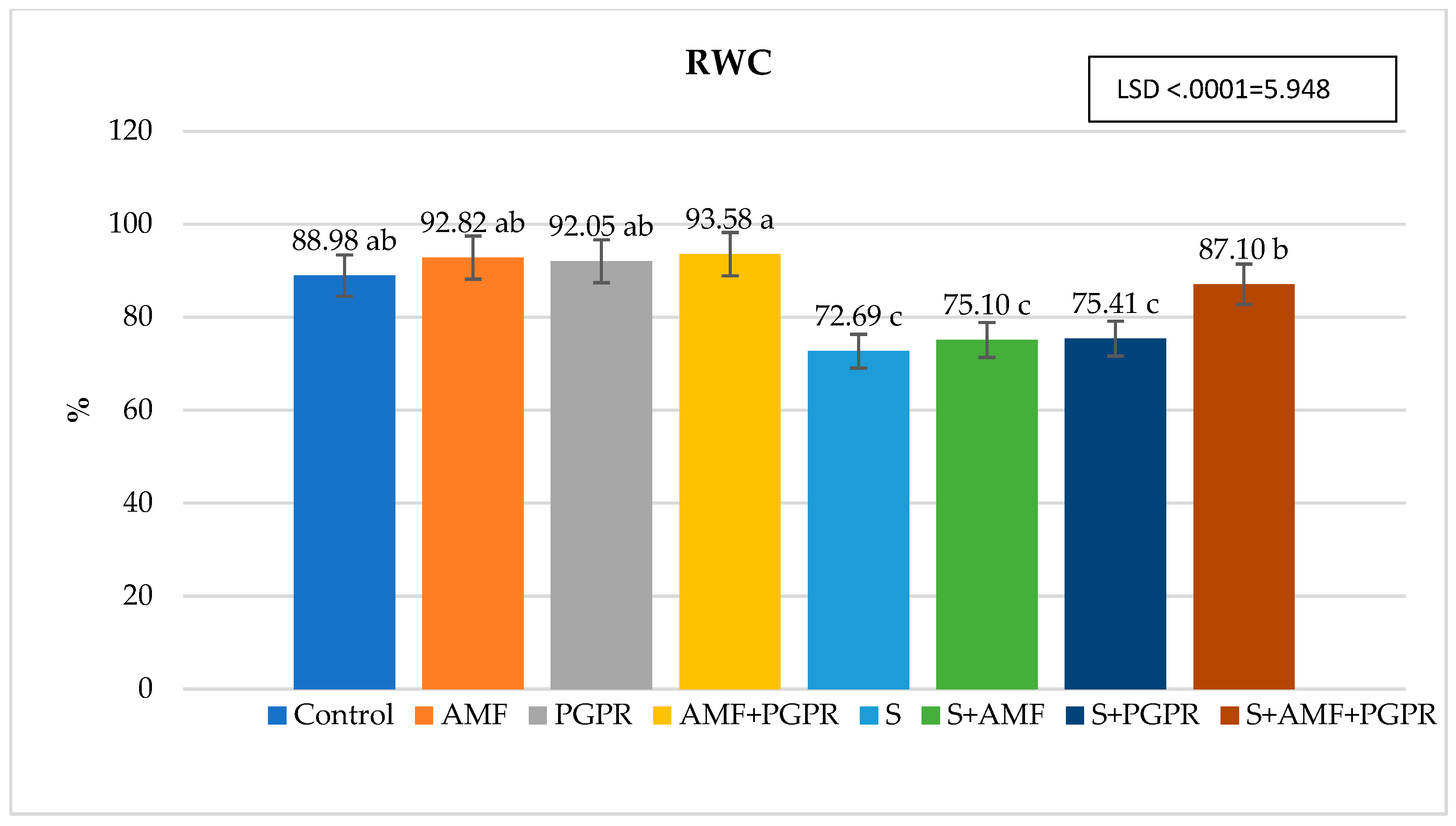
Figure 3.
Effects of the treatments on ion leakage of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
Figure 3.
Effects of the treatments on ion leakage of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
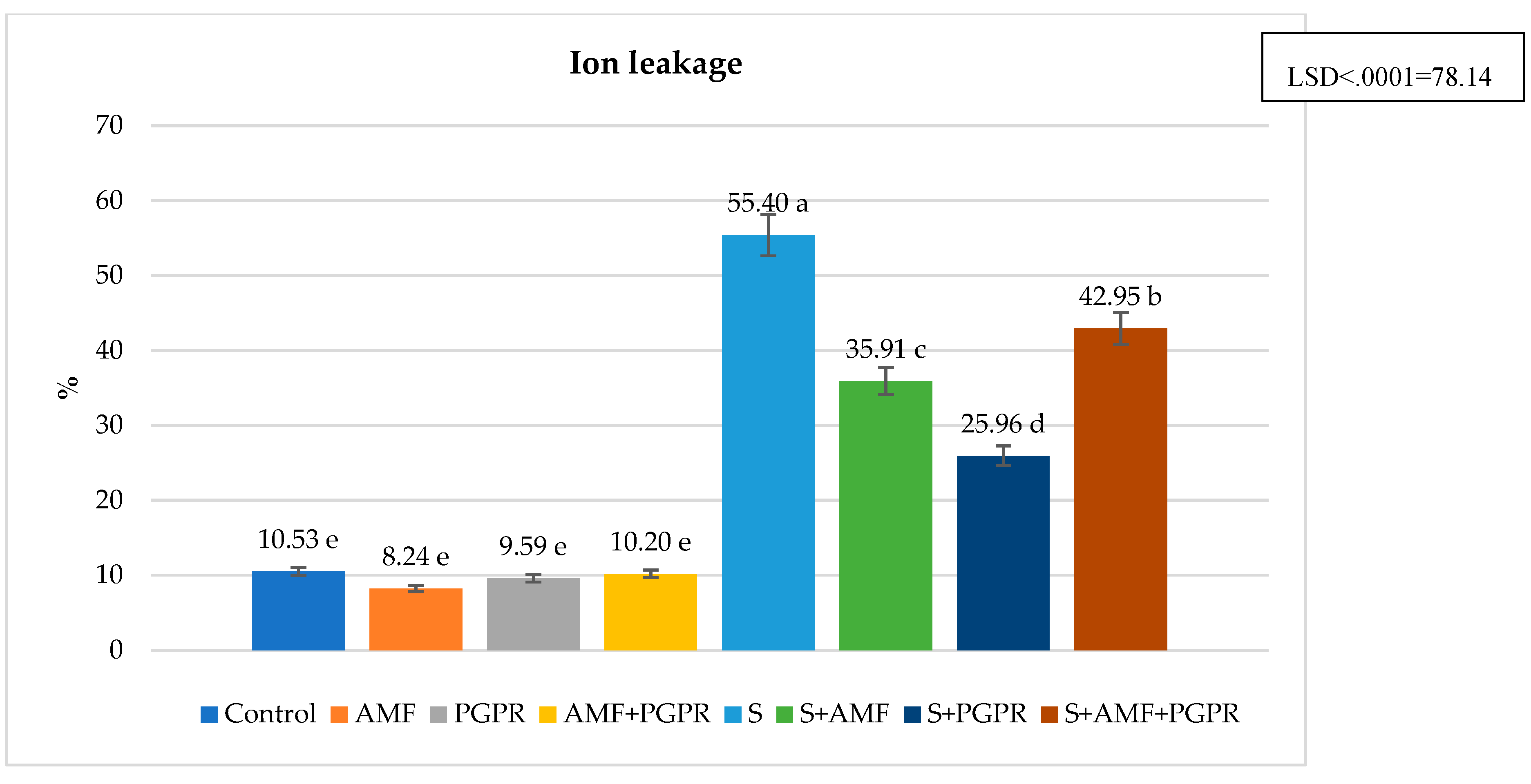
Figure 4.
Effects of C, AMF, PGPR, AMF+PGPR, S, S+AMF, S+PGPR and S+AMF+PGPR on osmotic potential of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
Figure 4.
Effects of C, AMF, PGPR, AMF+PGPR, S, S+AMF, S+PGPR and S+AMF+PGPR on osmotic potential of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
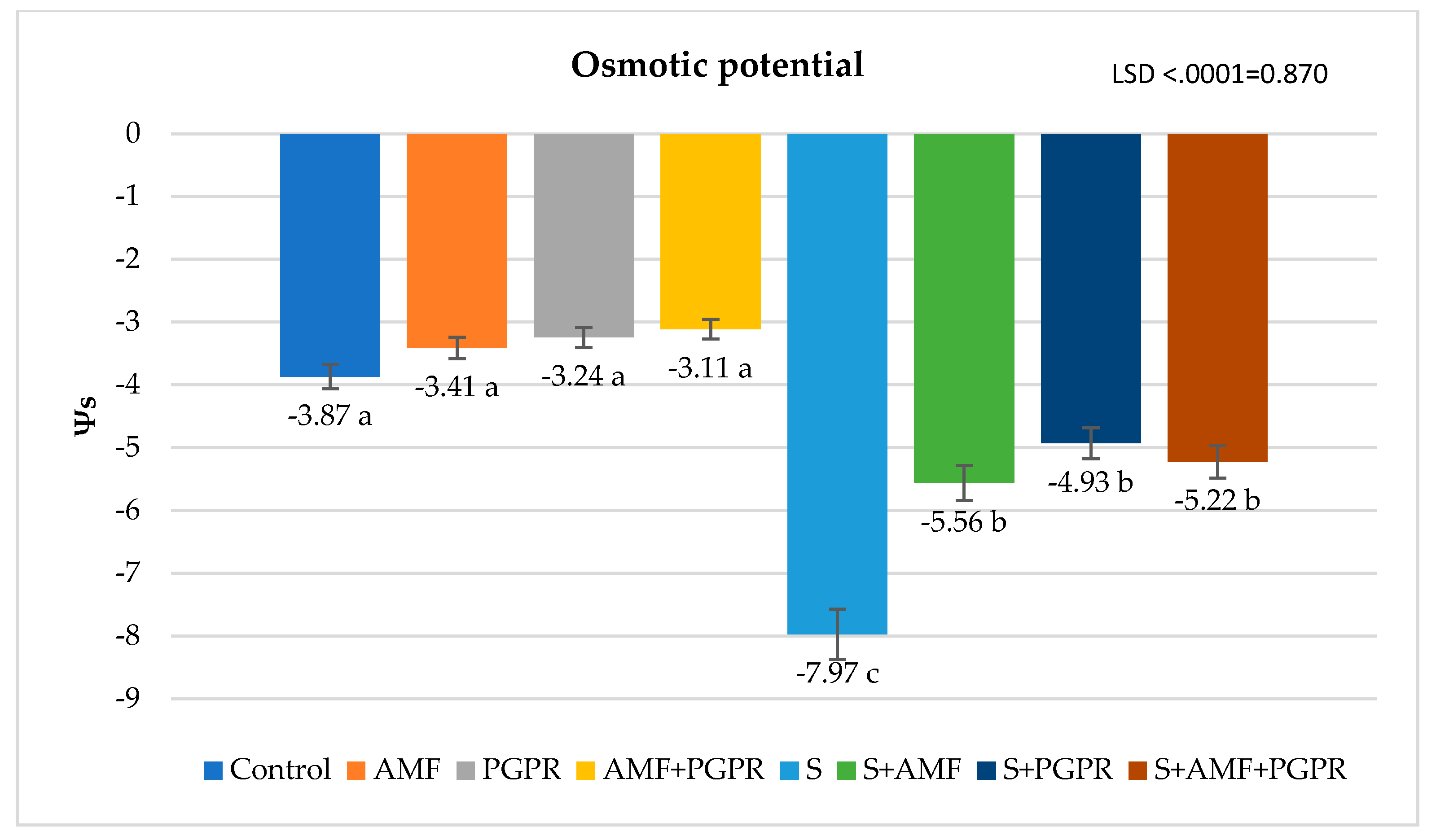

Table 1.
The nutrient solution used in the control treatment (100% mineral fertilization) (mg L-1).
| N | P | K | Ca | Mg | Fe | Mn | Zn | B | Cu | Mo |
|---|---|---|---|---|---|---|---|---|---|---|
| 100-239 | 40-81 | 96-370 | 150-250 | 50-92 | 5-10 | 1.97 | 0.25 | 0.7 | 0.07 | 0.05 |
Table 2.
Effects of the treatments on growth parameters of pepper plant. All data represent three replicates.
Table 2.
Effects of the treatments on growth parameters of pepper plant. All data represent three replicates.
| Application | Plant height (cm) | Plant diameter (mm) | Number of leaf per plant (number) | Number of bifurcations per plant (number) | Plant fresh weight (g/plant) | Leaf area (cm2/plant) |
|---|---|---|---|---|---|---|
| Control | 37.25 ab | 6.37 a | 45.00 a | 5.25 bc | 48.38 ab | 1450.75 b |
| AMF | 39.50 a | 6.52 a | 37.25 b | 5.75 ab | 47.94 ab | 1553.25 ab |
| PGPR | 37.75 ab | 6.50 a | 37.25 b | 7.00 a | 59.44 a | 1837.25 a |
| AMF+PGPR | 36.25 b | 5.44 b | 34.25 b | 6.00 ab | 39.11 b | 1436.25 b |
| Salt | 24.25 d | 4.53 d | 10.00 d | 2.00 e | 13.61 c | 382.75 c |
| Salt+AMF | 29.5 c | 5.10 bc | 16.75 c | 4.00 cd | 18.74 c | 580.00 c |
| Salt+PGPR | 26.5 cd | 5.09 bc | 14.75 cd | 4.50 bcd | 14.03 c | 450.00 c |
| Salt+AMF+PGPR | 27.25 cd | 4.78 cd | 11.00 d | 3.00 de | 11.81 c | 409.25 c |
| LSD0.05 | <.0001 | <.0001 | <.0001 | <.0001 | <.0001 | <.0001 |
| p | 3.017 | 0.402 | 4.963 | 1.616 | 13.677 | 361.422 |
1 There is no significant difference between the means with the same letter in the table at p<0.05.
Table 3.
Effects of the treatments on macro nutrient concentration (%) of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
Table 3.
Effects of the treatments on macro nutrient concentration (%) of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
| Application | Ca | Mg | K | Na | Cl |
|---|---|---|---|---|---|
| Control | 1.058 a | 1.126 de | 9.506 a | 0.29 c | 1.42 d |
| AMF | 0.966 b | 1.184 bcde | 9.023 bc | 0.23 c | 1.30 d |
| PGPR | 0.712 c | 1.139 cde | 9.231 ab | 0.22 c | 1.66 c |
| AMF+PGPR | 0.699 c | 1.100 e | 8.568 cd | 0.31 c | 1.49 cd |
| Salt | 0.983 ab | 1.334 a | 7.777 e | 3.05 a | 2.69 a |
| Salt+AMF | 0.978 b | 1.236 abcd | 7.726 e | 2.79 b | 2.27 b |
| Salt+PGPR | 0.937 b | 1.305 ab | 7.832 e | 3.00 a | 2.53 a |
| Salt+AMF+PGPR | 0.973 b | 1.263 abc | 8.174 de | 2.99 a | 2.63 a |
| LSD0.05 | <.0001 | 0.0067 | <.0001 | 0.82 | 0.68 |
| P | 0.080 | 0.128 | 0.487 | 2.99 | 2.63 |
Table 4.
Effects of the treatments on micro nutrient concentration (mg/kg) of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
Table 4.
Effects of the treatments on micro nutrient concentration (mg/kg) of pepper leaf. All data represent three replicates. There is no significant difference between means with the same letter in the same histogram (p<0.05).
| Application | Cu | Mn | Fe | Zn |
|---|---|---|---|---|
| Control | 14.75 bc | 87.68 f | 100.50 a | 131.67 cd |
| AMF | 13.50 cd | 101.33 ef | 97.50 ab | 125.25 d |
| PGPR | 13.50 cd | 119.67 d | 89.00 cd | 138.50 c |
| AMF+PGPR | 12.25 d | 109.33 de | 86.75 d | 129.33 d |
| Salt | 14.75 bc | 188.33 b | 101.75 a | 171.67 a |
| Salt+AMF | 18.00 a | 161.00 c | 97.00 abc | 160.33 b |
| Salt+PGPR | 18.00 a | 218.67 a | 98.50 ab | 171.00 a |
| Salt+AMF+PGPR | 16.75 ab | 185.50 b | 90.50 bcd | 174.67 a |
| LSD0.05 | 0.0003 | <.0001 | 0.0048 | <.0001 |
| P | 2.431 | 14.788 | 8.073 | 8.460 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



