Submitted:
25 September 2024
Posted:
25 September 2024
You are already at the latest version
Abstract
Pinosylvin and its methylated derivatives, pinosylvin monomethyl ether (PME) and dihydropinosylvin monomethyl ether (DPME), are representative phytoalexins found in Pinus species. Both PME and DPME have strong nematicidal activity against pine wood nematodes (PWNs). In this study, we investigated the resistance of Pinus densiflora to PWN after trunk injection or soil drenching with a fungal elicitor prepared from Penicillium chrysogenum. Fungal elicitor treatment by soil drenching resulted in increased accumulation of PME and DPME in the needles of the plants. When the pine trees without fungal elicitor pretreatment were inoculated with PWNs, 67.9% died after five months, but only 33.3% died among the plants pretreated with the fungal elicitor. Injection of the fungal elicitor into the trunks of P. densiflora trees also strongly increased the synthesis of PMEs. The concentration of PME in the leaves of P. densiflora reached a maximum (155.4 µg/g DW) after 4 weeks of fungal elicitor injection. The second injection of the elicitor after the first injection of the fungal elicitor also increased the accumulation of PME. To investigate PWN resistance in P. densiflora after two months of trunk injection of elicitor, the downward movement of PWNs from the top of branches to the bottom was monitored after PWNs were inoculated on the holes of the upper internodal segments of branches. The number and mobility of PWNs that moved from the bottom of branches were highly reduced in fungal elicitor-injected pine plants. This result suggests an eco-friendly approach for eliciting-induced PWN resistance in PWN-susceptible P. densiflora via fungal elicitor treatment.

Keywords:
Fungal elicitor
; Pine wood nematode
; Pine wilt disease
; Korean red pine
; Elicitor-induced PWN resistance
1. Introduction
Pine wood nematodes (Bursaphelenchus xylophilus; PWNs) are responsible for causing pine wilt disease (PWD), which is a serious and often fatal condition for pine trees. Pine species native to Eastern Asia and Western European countries are highly susceptible to PWD, which causes severe damage to forest ecosystems and significant economic losses in the forest product industry [5].
The current method for controlling PWD mainly relies on insecticide application via foliar sprays on pine plants to eliminate pine sawyer beetles (Monochamu spp.), which are insect vectors of PWNs [11,14]. Trunk injection of nematicides in pine plants is the most frequently used protocol for preventing PWN proliferation and decreasing the longevity and feeding activity of insect vectors [12,24]. Abamectin (Avermectin B1) and emamectin benzoate are the two most widely used nematicides and are members of the macrocyclic lactone family [12]. However, the application of toxic chemicals has raised concerns for nonharmful insects living in forest environments [1].
Many attempts have been made to develop beneficial eco-friendly compounds with nematicidal activity against PWNs [3]. Biological control methods for pine wilt disease aim to manage the spread of the nematode and reduce its impact on pine populations. Certain fungi and bacteria are known to kill PWNs; thus, these microorganisms can be used as biological control agents to reduce the population of PWNs [26,29]. Chitosan is frequently used as a potential natural compound to manage plant diseases [9,23]. Nunes da Silva et al. [21] evaluated the application of different molecular weight chitosans as a soil amendment to control PWD in maritime pine (Pinus pinaster, very susceptible to the disease) and stone pine (Pinus pinea, less susceptible). Elicitation with methyl jasmonate, salicylic acid, and benzo(1,2,3)-thiadiazole-7-carbothioic acid-S-methyl ester via foliar spraying increases tolerance to B. xylophilus [18]. The addition of this biofertilizer (mixed with diazotrophic bacteria and a chitosan-producing fungus, Cunninghamella elegans) to soils confers resistance against the PWN in P. pinaster [21].
Phytoalexins are important secondary metabolites in plant immunity, as they play crucial roles in defending plants against various pathogens. Pinosylvin and its methylated derivatives, pinosylvin monomethyl ether (PME) and dihydropinosylvin monomethyl ether (DPME), are representative phytoalexins found in pine species. Suga et al. [25] reported that pinosylvin, DPME, and PME are the most toxic compounds against PWNs among the extracted compounds from the Pinaceae family. Although pinosylvin and its methylated derivatives are very toxic to PWNs, these compounds are not detectable or exist in low amounts in the fresh tissue of branches and needles of pine plants [10,22,28], except for nonliving heartwood tissues [7]. The production of pinosylvin stilbenoids in sapwood and needles in pine is activated only by abiotic and biotic stresses [6,8]. In P. strobus, which is highly resistant to PWD, the accumulation of pinosylvin stilbenoids (PME and DPME) is increased upon PWN infection [10]. In contrast, in both Korean red pine (P. densiflora) and Koran pine (Pinus koraiensis), which are susceptible to PWD, there is no change in these substances upon PWN infection [10]. On the basis of these results, [10] proposed that PWD resistance in P. strobus is related to the ability to synthesize PME and DPME substances upon PWN infection.
To date, no studies have been conducted to increase the synthesis of pinosylvin stilbene through biotic or abiotic stress treatments in pine plants susceptible to PWD. However, increased accumulation of PME and DPME has been reported when calli and cultured cells induced from PWN-susceptible pine species were treated with fungi or fungus-derived elicitors. In the cell culture of P. sylvestris, treatment with fungal mycelium extracts from Lophodermium seditiosum strongly increased the accumulation of PME hundreds of times [17]. Kim et al. [16] reported that PME and DPME accumulation was highly increased in calli of P. koraiensis when the callus was treated with fungus-derived elicitors. The above results show that even for PWN-susceptible pine species such as P. koraiensis and P. sylvestris, the cultured cells recognized and quickly responded to the fungal elicitor, resulting in increased synthesis of pinosylvin stilbenes. Therefore, it was hypothesized that fungal elicitor treatment of PWD-susceptible pine plants might increase the accumulation of pinosylvin stilbenoid substances, which may confer PWD resistance upon infection with the nematode.
Trunk injection or soil drenching is frequently used for the delivery of pesticides, insecticides, or nutrients into the stems of woody plants. There are no reports of the trunk injection of elicitors for the prevention of PWD. Moreover, no studies have investigated the correlation between PWD resistance and pinosylvin stilbene accumulation by applying biotic and abiotic stimuli to pine plants. Therefore, in this study, pine trees were treated with fungus-derived elicitors to increase the synthesis of pinosylvin stilbenes, and then, we investigated the possible acquisition of PWN resistance in PWN-susceptible P. densiflora.
2. Materials and Methods
2.1. Preparation of Fungal Elicitors and PWNs
Penicillium chrysogenum KCTC 6052 was provided by KCTC, Biological Resources Center of Korea Research Institute of Bioscience and Biotechnology (KRIBB). To manufacture an elicitor using P. chrysogenum, the fungi were grown in potato dextrose agar (PDA) medium purchased from Sigma‒Aldrich, agar (20 g/L) was added to make a solid medium, and the pH of the medium was adjusted to 5.0 before sterilization. For suspension culture of the fungus, a small amount (1 mL) of the mixed fungal culture medium was inoculated into a 500 ml flask containing 100 ml of PDA liquid medium, and the flasks were agitated on a gyratory shaker at 100 rpm. After 2 weeks of culture, the fungal suspension medium was filtered through filter paper (Whatman No. 1) and filtered again through a 0.45 µm membrane filter. The filtered solution was then autoclaved at 120 °C for 15 minutes and used as an elicitor.
PWNs (Bursaphelenchus xylophilus) were propagated on filamentous fungi (Botrytis cinerea), which were subsequently grown on potato dextrose agar (PDA) media under dark conditions. The propagated PWNs were isolated via the Baerman funnel method [31].
2.2. Fungal Elicitor Treatment of Pine Trees by Soil Drenching
The fungal culture mixture was diluted by adding water to a concentration of 20% (v/v). The fungal culture mixture (500 ml) was applied via soil drenching to a 1 L soil pot where two-year-old Korean red pine trees were planted. As a control, the same amount of PDA medium was diluted with water. Pine leaves were harvested 1 week, 2 weeks, 3 weeks, and 4 weeks after the fungal elicitor was applied to the soil pot and used as materials for analyzing the contents of DPME and PME via GC/MS. To investigate the effects of multiple fungal elicitor treatments, the same amount of fungal culture mixture (500 ml) was subjected to soil drenching after 8 weeks of the first fungal elicitor treatment. Pine leaves were harvested 2 and 4 weeks after the additional fungal elicitor treatment to analyze the contents of DPME and PME.
2.3. PWD Resistance of Pine Plants after Soil Drenching Treatment with Fungal Elicitors
To inoculate the PWNs in pine trees, a part of the bark of the pine trunk is cut with a knife, cotton is inserted into the wounded bark, and then, a culture medium containing approximately 500 PWNs is inoculated there. After inoculation, the samples were sealed with parafilm. In the case of pine trees not treated with fungal elicitors as a control group, the bark was wounded in the same way, and the same number of nematodes were inoculated. The progress of PWD was monitored after PWN inoculation, and the survival rate of the pine trees was subsequently examined after 5 months. More than 15 inoculated pine trees were included in each of the control and treatment groups, and 3 repetitions were performed.
2.4. Trunk Injection of Fungal Elicitors on Pine Trees
The fungal culture medium was diluted by adding water to a concentration of 20% (v/v). Fungal culture medium (20 ml) was injected into the drilled holes of the trunks of 10-year-old Korean red pine trees. As a control, the same amount of PDA medium diluted with water was injected into the trunk. Pine leaves were harvested 1 week, 2 weeks, 3 weeks, and 4 weeks after the fungal elicitor injection was applied to the pine plants, which were subsequently used as materials for analyzing the contents of DPME and PME.
2.5. Investigation of PWN Movement in Branches of P. densiflora Plants after Trunk Injection of Fungal Elicitors
Internodes of control and fungal elicitor-treated pine (Pinus densiflora) plants were cut to 100 mm to make nodal segments, and then, a hole was drilled at the top, approximately 500 PWNs were inoculated there, and the samples were sealed with parafilm. The branch segments were placed into a 50 mL Falcon tube. At this time, 5 mL of water was added to a tube in which the lower parts of the segments were submerged in water. After that, the cap of the falcon tube was closed and erected, and after 24 hours, the number and viability (movement) of PWNs in the water removed from the bottom of the branch segments were determined via optical microscopy. The branch segments were sampled from three independent pine plants, and the analysis of the number and viability of PWNs that were removed from the bottom of branch segments was performed on more than 5 segments per pine plant.
2.6. GC‒MS Analysis
To measure the contents of the DPME and PME compounds via gas chromatography‒mass spectrometry (GC‒MS), the pine leaves were air-dried in an oven at 50 °C. Each sample was pulverized to make a powder, and then 100 mg was weighed, immersed in 100% methanol (1 ml), and sonicated at a frequency of 20 kHz for 30 min at 40 °C. Then, the supernatant obtained by centrifugation (15000 × g for 10 min) was filtered through a 0.45 μm membrane to remove impurities. Filtered aliquots were injected into a GC (Agilent 7890A) coupled to an MSD system (Agilent 5975C) equipped with an HP-5MS capillary column (30 m × 0.25 mm, film thickness 0.25 mm). The injection temperature was 250 °C, and the column temperature was 70 °C for 4 min, 220 °C at 5 °C min-1, heated to 320 °C at 4 °C min-1, and held at 320 °C for 5 min. The carrier gas was He at a flow rate of 1.2 ml min-1. The interface temperature was 300 °C with split/nonsplit implantation (10:1), the temperature of the ionization chamber was 250 °C, and ionization was performed by electron bombardment at 70 eV. The standard compounds of DPME and PME used for GC‒MS analysis were purchased from Sigma‒Aldrich Co. The experiment was repeated three times.
2.7. Statistical Analysis
The quantitative data are expressed as the means ± standard errors (SEs). All the statistical analyses were performed with SPSS software (SPSS Science, Chicago, IL, USA). Statistical differences among means were calculated via one-way ANOVA followed by Duncan’s post hoc test.
3. Results
3.1. DPME and PME Accumulation in P. densiflora Trees after Soil Drenching Treatment with Fungal Elicitors
P. chrysogenum was cultured on the surface of solid PDA media for two weeks (Figure 1a) and then transferred to PDA liquid media for suspension culture for two weeks (Figure 1b). A fungal elicitor (Figure 1c) was prepared from sterilized filtered liquid medium after two weeks of culture of P. chrysogenum. PWN-susceptible Korean red pine (P. densiflora) plants were treated with the fungal elicitor by soil drenching. To investigate the metabolic changes in P. densiflora plants after fungal elicitor treatment, the accumulation of secondary metabolites in the needle extracts of the pine plants was analyzed via GC. The selected ion monitoring (SIM) of the GCs revealed that pinosylvin stilbenoids (PME and DPME) clearly increased in the leaves after fungal elicitor treatment (Figure 2a-d), peaked after 4 weeks of elicitor treatment and declined after 8 weeks. The identification of DPME and PME was confirmed by mutually comparing the analysis results of the peaks of standard compounds (Figure 1e). The peaks of DPME and PME in the control group were very small (Figure 2a). The accumulation of DPME and PME in the leaves of the control plants was 1.6 ± 0.2 μg/g DW and 14.2 ± 1.5 μg/g DW, respectively (Figure 2f). After 4 weeks of elicitor treatment, the DPME and PME contents increased to 12.7 ± 1.3 μg/g DW and 156.2 ± 13.2 μg/g DW, respectively (Figure 2f). Thus, the DPME content increased 7.9-fold, and the PME content increased 11-fold compared with those of the control group after 4 weeks of elicitor treatment.
To investigate the accumulation of DPME and PME in needles by repetitive fungal elicitor treatment, the fungal elicitor was additionally treated by soil drenching after 8 weeks of the first fungal elicitor application (Figure 2g-i). The accumulation of DPME and PME increased again after 4 and 8 weeks of additional soil drenching treatment with fungal elicitors (Figure 2g-H). Analysis of DPME and PME in needles revealed that the additional treatment with the fungal elicitor resulted in higher levels of DPME and PME than the initial treatment with the fungal elicitor (Figure 2i). After 4 weeks of additional fungal elicitor treatment, the DPME and PME contents increased to 23.5 ± 7.5 μg/g DW and 266.3 ± 23.5 μg/g DW, respectively (Figure 2f). Thus, the DPME content increased 13.8-fold, and the PME content increased 20.9-fold compared with those of the control group after 4 weeks of additional fungal elicitor treatment.
3.2. Investigation of PWD after Soil Drenching with Fungal Elicitors
The fungal culture was applied by soil drenching, the nematodes were inoculated into pine trunks two weeks later, and the progression of PWD was examined consecutively until five months later. In the soil drenching treatment with fungal culture medium only, 100% of the pine plants maintained healthy green leaves after 5 months (Figure 3a, d). In contrast, after the inoculation of PWNs without fungal culture medium, 33.3% of the plants maintained healthy green leaves, but the other plants presented severely wilted leaves with symptoms of PWD (Figure 3b, d). Among the PWN-infected plants subjected to the fungal culture medium, 67.9% had green leaves without PWD symptoms until after five months (Figure 3c, d).
3.3. DPME and PME Accumulation in P. densiflora Trees after Trunk Injection of Fungal Elicitors
The accumulation of DPME and PME in pine leaves after 1, 2, 3, and 4 weeks of trunk injection of the fungal elicitor was analyzed by GC. The GC SIM peaks of DPME and PME in the control group were very small but greatly increased after trunk injection of the fungal elicitor, and the highest peak was observed after 4 weeks of elicitor treatment (Figure 4a-e). The accumulation of DPME and PME in the leaves of untreated control plants was 1.7 ± 0.6 μg/g DW and 12.7 ± 1.5 μg/g DW, respectively (Figure 4e). The DPME and PME contents were 13.2 ± 1.3 μg/g DW and 155.4 ± 13.3 μg/g DW, respectively, after 4 weeks of fungal elicitor injection (Figure 4e). Compared with those of the control group, the DPME content increased 7.8-fold, and the PME content increased 12.2-fold after 4 weeks of elicitor injection.
3.4. The Vitality of PWNs Moving from the Top to the Bottom of a Pine Branch Segment
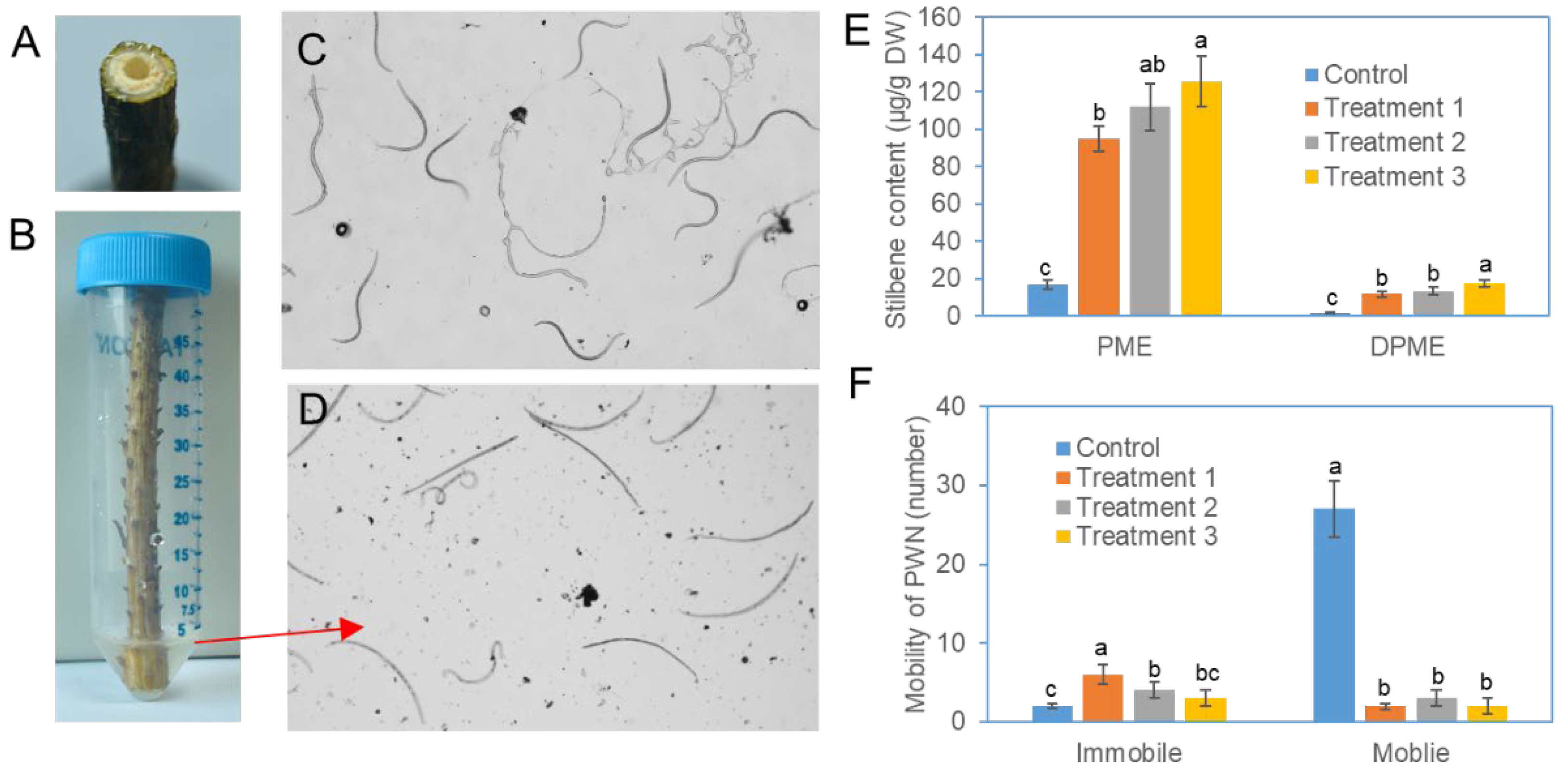
Internodal segments (10 mm long) were prepared from branches of control and fungal elicitor-treated pine (P. densiflora) trees. PWNs were inoculated on a drilled hole at the distal top of each segment (Figure 5a). The number and viability of the PWNs that moved from the bottom of the branch segments into the water were examined with an optical microscope (Figure 5b). When the PWNs were removed from the branches of a pine tree that was not treated with the fungal elicitor, the PWNs showed motion with an actively pendulating wavy shape, as shown in the camera picture (Figure 5c). On the other hand, most of the PWNs that moved from the branches of the pine trees treated with fungal elicitors had a straight shape, and although some young larvae moved, their motility was significantly weakened (Figure 5d). Compared with those in the control plants, the contents of DPME and PME in the three pine plants subjected to trunk injection of fungal elicitors significantly increased to 95–126 μg/g DW compared with 16.7 μg/g DW in the control plants (Figure 5e). In the control group, a total of 29 PWNs were counted in the water after 24 h (Figure 5e). On the other hand, the number of PWNs that moved into the water was highly reduced in the branches treated with the fungal elicitor (Figure 5e).
4. Discussion
In this study, we investigated whether fungal elicitor treatment affects PME and DPME accumulation in Korean red pine trees and whether this change is associated with resistance to PWD. The fungal elicitor used was the medium filtrate after P. chrysogenum liquid culture, which has already been shown to strongly increase the synthesis of PME and DPME in Korean pine cell cultures through the treatment of the medium filtrate of P. chrysogenum [16]. Among the fungal elicitors derived from 7 different fungal species, Penicillium, Trichoderma and Aspergillus, a fungal elicitor prepared from P. chrysogenum was the most effective elicitor for increasing the production of DPME and PME in P. strobus cells [30]. The fungal elicitors were either inoculated into the soil where the pine trees were planted or directly injected into the trunks of the pine trees and then analyzed for changes in the contents of PME and DPME for 4 weeks. The results revealed that both soil drenching and tree injection of the fungal elicitor significantly increased the accumulation of PME and DPME. PME is highly toxic to PWNs, resulting in 100% killing of PWNs in 24 hours at concentrations as low as 10 ppm according to an in vitro assay [25]. Therefore, we speculated that increasing PME by fungal elicitor treatment of Korean red pine, which is highly susceptible to PWD, may slow the progression of PWD caused by PME toxicity. To test this hypothesis, we inoculated PWNs into Korean Red pine trees after two weeks of fungal elicitor soil treatment and monitored the development of PWD symptoms for five months. The results revealed that 78% of the fungal elicitor-untreated pine plants presented symptoms of PWD, whereas only 33% of the pine trees pretreated with the elicitor presented symptoms of PWD, confirming that pretreatment with the fungal elicitor was highly effective in reducing PWD. This result supports our previous hypothesis that fungal elicitor treatment of P. densiflora plants results in increased pinosylvin stilbenoid accumulation, which may confer toxicity to PWNs.
Since trunk injection of fungal elicitors into pine trees resulted in a significant increase in the PME and DPME contents, we investigated the effects of trunk elicitor injection of fungal elicitors on the motility and activity of PWNs after PWN infection. Inoculation of the PWNs was performed on the top of the internodal segments of branches of 10-year-old pine plants after two weeks of trunk injection of fungal cultures. We examined the migration and mobility of nematodes to the bottom of branches from the top. The results revealed that the movement of PWNs downward to the branch was significantly inhibited because the number of nematodes was significantly reduced in branches pretreated with the fungal elicitor. Moreover, the movement of PWNs that moved from the branches was highly weakened, and these PWNs began to lose movement within 24 hrs. These results suggest that the increased accumulation of PME and DPME in pine trees after injection of fungal cultures may directly affect the movement of inoculated PWNs. Thus, trunk injection of fungal elicitors has the potential to provide a protocol for controlling PWD disease in P. densiflora plants.
Currently, aerial spraying and trunk injection of abamectin-based insecticides constitute a general way to eliminate PWN vectors, pine sawyer beetles [19], and control PWD in pine plants [12,24]. However, it is widely believed that these methods cause severe damage to the biological ecosystem of forests [15]. Therefore, many researchers are actively developing eco-friendly control technologies as alternatives to chemical pesticide applications [3]. Among them, representative control substances for PWD in pine plants often utilize microorganisms (fungi, bacteria, and actinomycetes) [3]. On the other hand, chitosan has been reported to have a preventive effect on PWD when it is sprayed on pine trees [21]. Treatments with methyl jasmonate, salicylic acid, and methyl salicylate have also been reported to be protective against PWNs [13,18]. Although neither of these studies confirmed the association between PWN protection and pinosylvin stilbenoid accumulation, it is possible that elicitor treatment may be associated with pinosylvin stilbenoid synthesis.
In P. strobus, which is highly resistant to PWD, pinosylvin stilbenoid (PME and DPME) accumulation is highly increased upon PWN infection. However, both Korean red pine (Korean red pine (P. densiflora) and Korean pine (P. koraiensis), which are susceptible to PWD, do not exhibit changes in these substances upon PWN infection [10]. Thus, the ability of PWNs to accumulate pinosylvin stilbenoids upon infection is likely related to PWD resistance in P. strobus plants because pinosylvin stilbenoids (PME and DPME) are highly toxic to PWNs [10,25]. Pinosylvin stilbenoids are produced from the precursor phenylalanine to PME or DPME [4]. Transcriptional regulators play key roles in plant defense mechanisms [2]. In a previously reported study, the lack of an increase in pinosylvin stilbenoids upon PWN infection in PWD-susceptible Korean Red pine and Korean pine [10] was attributed to a defect in the role of transcriptional regulators in controlling the synthesis of pinosylvin stilbenoids. Therefore, the differences in pinosylvin stilbenoid accumulation between strobe pine and Korean pine upon PWN infection are likely due to differences in the role of transcriptional regulators that control the synthesis of these substances. In recent years, much progress has been made in understanding the mechanisms of disease resistance associated with plant disease immunity [27]. In the case of pine plants, terpenoids, flavonoids, and stilbenoids have been interpreted as the main secondary metabolites associated with immunity in PWD resistance [20]. Therefore, analyzing the role of transcriptional regulators that control the synthesis of these compounds in pine tree species may provide important clues for understanding PWD immunity and for their use in breeding for the future development of PWD-resistant pine trees.
5. Conclusions
We found that the treatment of P. densiflora plants with fungal elicitors via either soil-drenching treatment or trunk injection significantly increased the synthesis of pinosylvin stilbenes. In addition, pretreatment with fungal elicitors strongly increased the survival of P. densiflora plants after PWN infection. This result suggests that the increased PWD resistance of P. densiflora plants after infection with PWNs may be due to the enhanced immune response caused by fungal elicitor pretreatment. The enhanced accumulation of pinosylvin stilbene by fungal elicitor treatment is interpreted as an important defense reaction against PWNs. It is suggested that fungal elicitor pretreatment can be utilized as an environmentally friendly PWN control technology in pine plants. The increased immunity of Asian and European pine trees against PWNs caused by fungal elicitor pretreatment could be an important approach for reducing the degree of damage caused by PWD and could be used to understand the molecular events of increased immunity against PWNs in the future.
Author Contributions
Y.E.C. designed this research. Y.B.M., Y.R.K. and J.Y.H. conducted the experiments and analysed the data. Y.B.M. and Y.E.C. wrote the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy restrictions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bai, S.H.; Ogbourne, S. Eco-toxicological effects of the avermectin family with a focus on abamectin and ivermectin. Chemosphere 2016, 154, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.P.; Liu, S.; Somssich, I.E. Transcriptional events defining plant immune responses. Curr. Opin. Plant Biol. 2017, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dou, G.; Yan, D.H. Research progress on biocontrol of Pine Wilt Disease by microorganisms. Forests 2022, 13, 1047. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V. Regulation of stilbene biosynthesis in plants. Planta 2017, 246, 597–623. [Google Scholar] [CrossRef]
- Futai, K. Pine wood nematode, Bursaphelenchus Xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef]
- Gehlert, R.; Schöppner, A.; Kindl, H. Stilbene synthase from seedlings of Pinus sylvestris: Purification and induction in response to fungal infection. Mol. Plant Microbe Interact. 1990, 3, 444–449. [Google Scholar] [CrossRef]
- Harju, A.M.; Venäläinen, M. Measuring the decay resistance of Scots pine heartwood indirectly by the Folin-Ciocalteu assay. Can. J. For. Res. 2006, 36, 1797–1804. [Google Scholar] [CrossRef]
- Harju, A.M.; Venäläinen, M.; Laakso, T.; Saranpää, P. Wounding response in xylem of scots pine seedlings shows wide genetic variation and connection with the constitutive defence of heartwood. Tree Physiol. 2009, 29, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Hassan, O.; Chang, T. Chitosan for eco-friendly control of plant disease. Asian J. Plant Pathol. 2017, 11, 53–70. [Google Scholar] [CrossRef]
- Hwang, H.S.; Han, J.Y.; Choi, Y.E. Enhanced accumulation of pinosylvin stilbenes and related gene expression in Pinus strobus after infection of pine wood nematode. Tree Physiol. 2021, 41, 1972–1987. [Google Scholar] [CrossRef]
- Ikeda, T. Integrated pest management of Japanese pinewilt disease. Eur. J. Forest Pathol. 1984, 14, 398–414. [Google Scholar] [CrossRef]
- James, R.; Tisserat, N.; Todd, T. Prevention of pine wilt of scots pine (Pinus sylvestris) with systemic abamectin injections. Arboric Urban For. 2006, 32, 195–201. [Google Scholar] [CrossRef]
- Jeon, H.W.; Park, A.R.; Sung, M.; Kim, N.; Mannaa, M.; Han, G.; et al. Systemic acquired resistance-mediated control of pine wilt disease by foliar application with methyl salicylate. Front Plant Sci. 2022, 12, 812414. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.K.; Lee, U.G.; Cha, D.; Kim, D.S.; Jung, C. Can insecticide applications used to kill vector insects prevent pine wilt disease? Pest Manag. Sci. 2021, 77, 4923–4929. [Google Scholar] [CrossRef]
- Kim, B.N.; Kim, J.H.; Ahn, J.Y.; Kim, S.; Cho, B.K.; Kim, Y.H.; Min, J. A short review of the pinewood nematode, Bursaphelenchus xylophilus. J. Toxicol Environ Health Sci. 2020, 12, 297–304. [Google Scholar] [CrossRef]
- Kim, Y.R.; Han, J.Y.; Choi, Y.E. Production of nematicidal pinosylvin stilbenes in cell suspension cultures of Pinus koraiensis by fungal elicitation. Forests 2022, 11, 2933. [Google Scholar] [CrossRef]
- Lange, B.M.; Trost, M.; Heller, W.; Langebartels, C.; Sandermann, H. Elicitor-Induced formation of free and cell-wall-bound stilbenes in cell-suspension cultures of Scots pine (Pinus sylvestris L.). Planta 1994, 194, 143–148. [Google Scholar] [CrossRef]
- López-Villamor, A.; da Silva, M.; Vasconcelos, M.W. Evaluation of plant elicitation with methyljasmonate, salicylic acid and benzo (1, 2, 3)-thiadiazole-7-carbothioic acid-S-methyl ester for the sustainable management of the pine wilt disease. Tree Physiol. 2022, 42, 2596–2613. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M. and Sattelle, D.B. Neonicotinoids: insecticides acting on insect nicotinic acetylcholine receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Modesto, I.; Mendes, A.; Carrasquinho, I.; Miguel, C.M. Molecular defense response of pine trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus. Cells 2022, 11, 3208. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Santos, C.S.; Cruz, A.; López-Villamor, A.; Vasconcelos, M.W. Chitosan increases Pinus pinaster tolerance to the pinewood nematode (Bursaphelenchus xylophilus) by promoting plant antioxidative metabolism. Sci. Rep. 2021, 11, 3781. [Google Scholar] [CrossRef] [PubMed]
- Rosemann, D.; Heller, W.; Sandermann, H. Biochemical plant responses to ozone: II. Induction of stilbene biosynthesis in Scots pine (Pinus sylvestris L.) Seedlings. Plant Physiol. 1991, 97, 1280–1286. [Google Scholar] [CrossRef]
- Saberi-Riseh, R.; Hassanisaadi, M.; Vatankhah, M.; Babaki, S.A.; Barka, E.A. Chitosan as potential natural compound to manage plant diseases, Int. J. Biol. Macromol. 2022, 220, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Sousa, E.; Naves, P.; Vieira, M. Pevention of pine wilt disease induced by Bursaphelenchus xylophilus and Monochamus galloprovincialis by trunk injection of emamectin benzoate. Phytoparasitica 2013, 41, 143–148. [Google Scholar] [CrossRef]
- Suga, T.; Ohta, S.; Munesada, K.; Ide, N.; Kurokawa, M.; Shimizu, M.; Ohta, E. Endogenous pine wood nematicidal substances in pines, Pinus Massoniana, P. Strobus and P. Palustris. Phytochtmistry 1993, 33, 1395–1401. [Google Scholar] [CrossRef]
- Tian, B.; Li, N.; Lian, L.; Liu, J.; Yang, J.; Zhang, K.Q. Cloning, expression and deletion of the cuticle-degrading protease BLG4 from nematophagous bacterium Brevibacillus laterosporus G4. Arch. Microbiol. 2006, 186, 297–305. [Google Scholar] [CrossRef]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Current Opinion in Plant Biology 2010, 13, 459–465. [Google Scholar] [CrossRef]
- Willför, S.; Hemming, J.; Reunanen, M.; Holmbom, B. Phenolic and lipophilic extractives in Scots pine knots and stemwood. Holzforschung 2003, 57, 359–372. [Google Scholar] [CrossRef]
- Yin, C.; Wang, Y.; Zhang, Y.; Wang, H.; Duan, B.; Tao, R.; Gao, J.; Sung, C. Hypothesized mechanism of biocontrol against pine wilt disease by the nematophagous fungus Esteya vermicola. Eur. J. Plant Pathol. 2020, 156, 811–818. [Google Scholar] [CrossRef]
- Kim, Y. R.; Moon, Y.B.; Choi, H.B.; Han, J.Y.; Choi, H.S.; Shim, S.R.; Choi, Y.E. MYB4 is the best candidate transcription factor involved in pinosylvin stilbene biosynthesis in Pinus strobus L. cells by fungal elicitor treatment. Annals of Forest Sci. 2024, 81, 18–37. [Google Scholar] [CrossRef]
- Hooper, D.J. Extraction of nematodes from plant material. In Laboratory Methods for Work with Plant and Soil Nematodes; Reference Book, No., 402, *!!! REPLACE !!!*, Southey, J.F., Eds.; Ministry of Agriculture, Fisheries and Food, Her Majesty’s Stationery Office: London, UK, 1986; pp. 51–58. [Google Scholar]
Figure 1.
Photos of fungal elicitors prepared from the liquid media of P. chrysogenum after 2 weeks of suspension culture in PDA liquid media. (a) Photo of P. chrysogenum mycelia. (b) Photo of P. chrysogenum mycelia after 2 weeks of flask culture. (c) Photo of filtered fungal culture medium for elicitor treatment.
Figure 1.
Photos of fungal elicitors prepared from the liquid media of P. chrysogenum after 2 weeks of suspension culture in PDA liquid media. (a) Photo of P. chrysogenum mycelia. (b) Photo of P. chrysogenum mycelia after 2 weeks of flask culture. (c) Photo of filtered fungal culture medium for elicitor treatment.
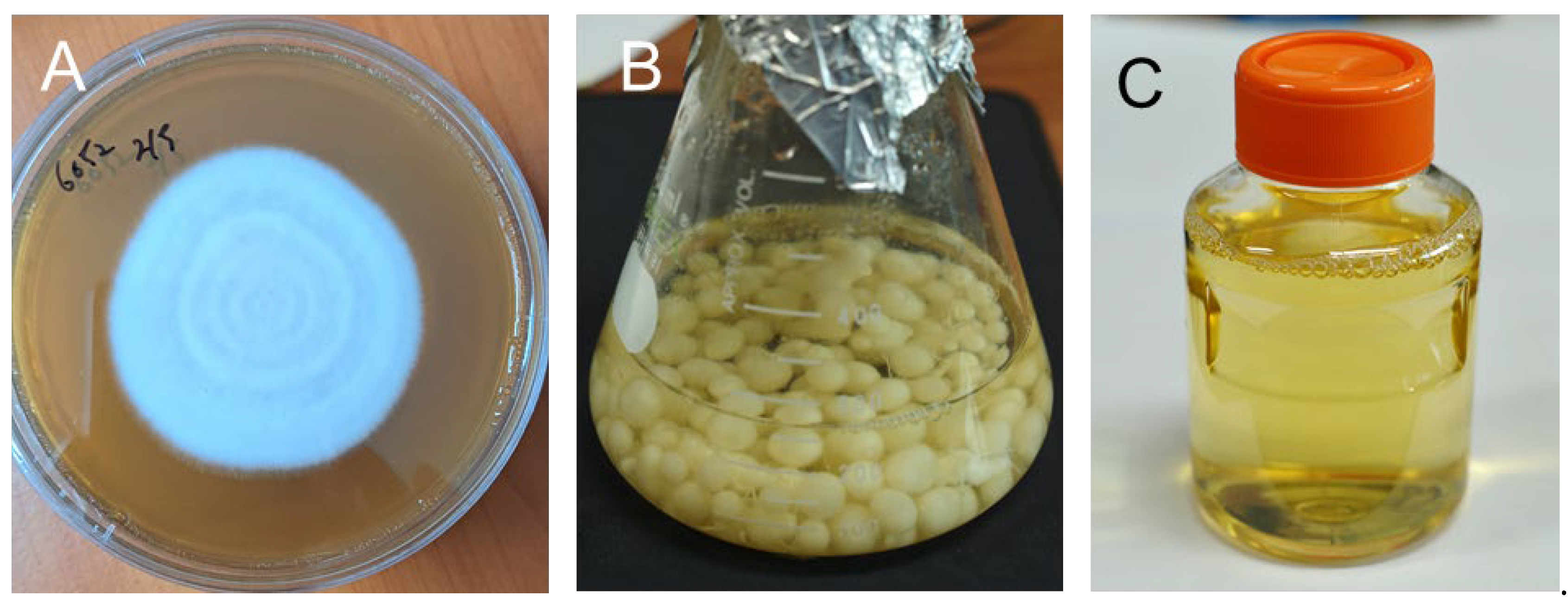
Figure 2.
GC/MS analysis of DPME and PME production in needles of P. densiflora after the soil drenching treatment with fungal elicitors prepared from P. chrysogenus. (a) GC chromatograms of extracts from needles without elicitor treatment. (b-d) GC chromatograms of extracts from needles after 2 weeks (b), 4 weeks (c), and 8 weeks of elicitor treatment (d). (e) GC chromatograms of authentic standards of DPME and PME. (f) Contents of DPME and PME in needles after elicitor treatment. (g) GC chromatograms of authentic standards of DPME and PME. (g-h) GC chromatograms of extracts from needles after 2 weeks (g) and 4 weeks (h) of additional elicitor treatment. The elicitor treatment was performed 8 weeks after the first elicitor treatment. (i) GC chromatograms of authentic standards of DPME and PME. (j) Contents of DPME and PME in needles after additional elicitor treatment. Different letters above the bars indicate significantly different values (P<0.05), which were calculated via one-way ANOVA followed by Duncan’s post hoc analysis.
Figure 2.
GC/MS analysis of DPME and PME production in needles of P. densiflora after the soil drenching treatment with fungal elicitors prepared from P. chrysogenus. (a) GC chromatograms of extracts from needles without elicitor treatment. (b-d) GC chromatograms of extracts from needles after 2 weeks (b), 4 weeks (c), and 8 weeks of elicitor treatment (d). (e) GC chromatograms of authentic standards of DPME and PME. (f) Contents of DPME and PME in needles after elicitor treatment. (g) GC chromatograms of authentic standards of DPME and PME. (g-h) GC chromatograms of extracts from needles after 2 weeks (g) and 4 weeks (h) of additional elicitor treatment. The elicitor treatment was performed 8 weeks after the first elicitor treatment. (i) GC chromatograms of authentic standards of DPME and PME. (j) Contents of DPME and PME in needles after additional elicitor treatment. Different letters above the bars indicate significantly different values (P<0.05), which were calculated via one-way ANOVA followed by Duncan’s post hoc analysis.
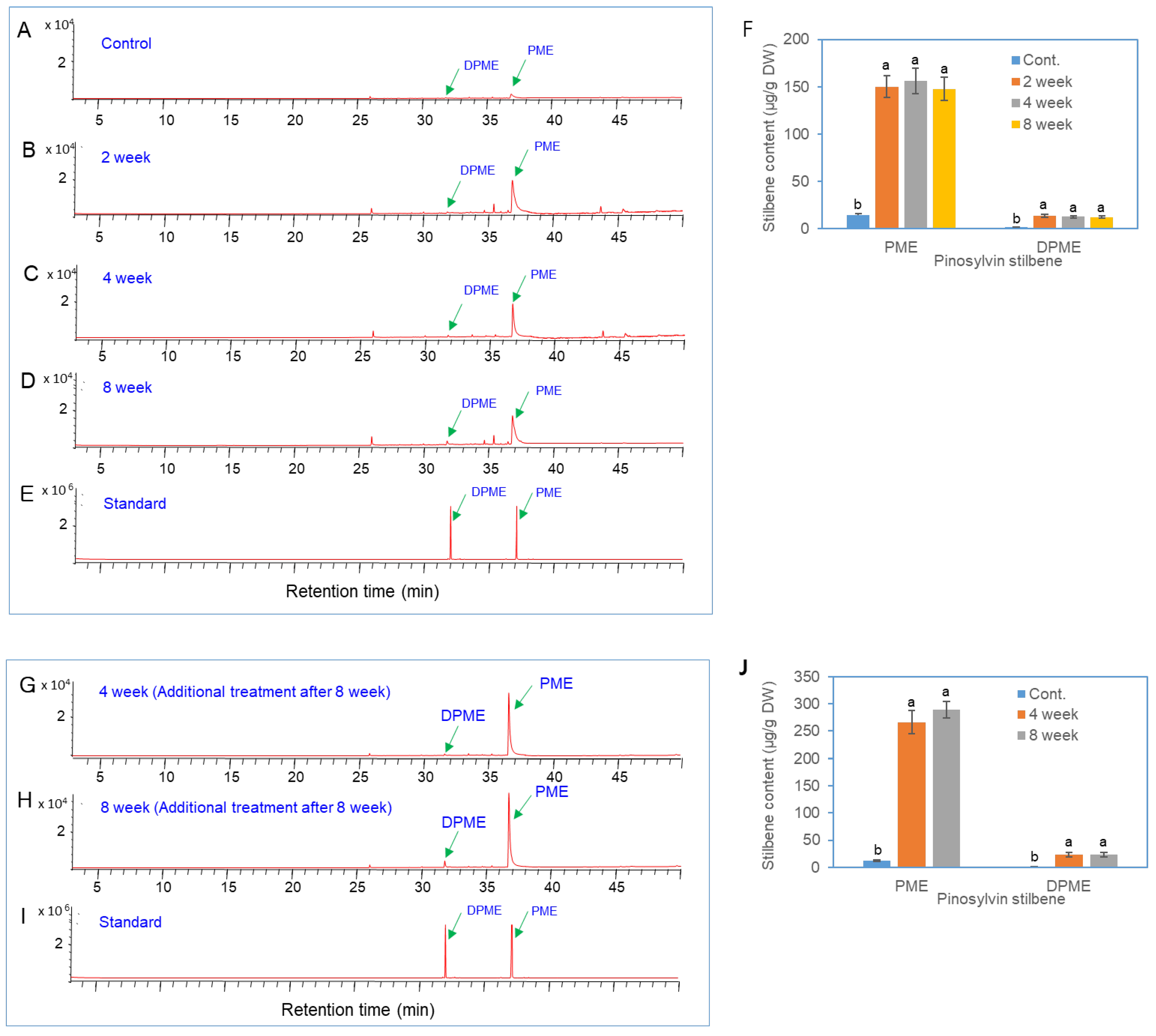
Figure 3.
Photographs and survival of PWN-infected P. densiflora plants after soil drenching treatment with fungal elicitors. (a) Control plants without PWN infection and elicitor treatment. (b) Pine trees without elicitor pretreatment and then infested with PWN 2 weeks later. (c) Pine trees pretreated with elicitor for two weeks and then infested with PWN 2 weeks later. (d) Effects of fungal elicitor pretreatment on the survival of pine plants after infection with PWNs. Images and data were acquired after 5 months of PWN infection.
Figure 3.
Photographs and survival of PWN-infected P. densiflora plants after soil drenching treatment with fungal elicitors. (a) Control plants without PWN infection and elicitor treatment. (b) Pine trees without elicitor pretreatment and then infested with PWN 2 weeks later. (c) Pine trees pretreated with elicitor for two weeks and then infested with PWN 2 weeks later. (d) Effects of fungal elicitor pretreatment on the survival of pine plants after infection with PWNs. Images and data were acquired after 5 months of PWN infection.
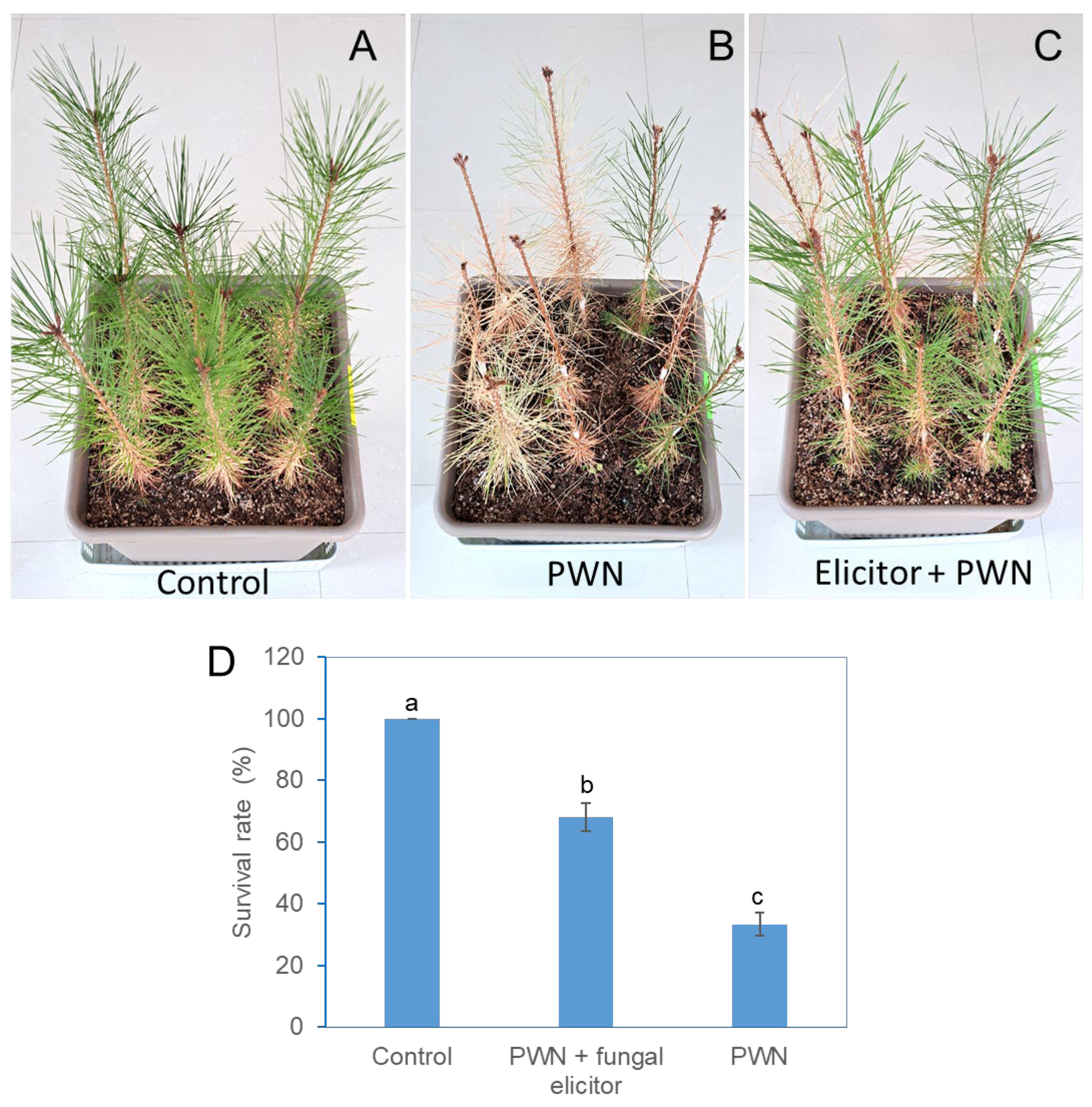
Figure 4.
GC/MS analysis of DPME and PME production in needles of P. densiflora after trunk injection of fungal elicitors prepared from P. chrysogenus. (a) GC chromatogram of extracts from needles without trunk injection of fungal elicitor. (b-e) GC chromatograms of extracts from needles after 1 week (b), 2 weeks (c), 3 weeks (d), and 4 weeks (e) of trunk injection of elicitor. (f) GC chromatograms of authentic standards of DPME and PME. (g) Contents of DPME and PME in needles after trunk injection of fungal elicitors. Different letters above the bars indicate significantly different values (P<0.05), which were calculated via one-way ANOVA followed by Duncan’s post hoc analysis.
Figure 4.
GC/MS analysis of DPME and PME production in needles of P. densiflora after trunk injection of fungal elicitors prepared from P. chrysogenus. (a) GC chromatogram of extracts from needles without trunk injection of fungal elicitor. (b-e) GC chromatograms of extracts from needles after 1 week (b), 2 weeks (c), 3 weeks (d), and 4 weeks (e) of trunk injection of elicitor. (f) GC chromatograms of authentic standards of DPME and PME. (g) Contents of DPME and PME in needles after trunk injection of fungal elicitors. Different letters above the bars indicate significantly different values (P<0.05), which were calculated via one-way ANOVA followed by Duncan’s post hoc analysis.
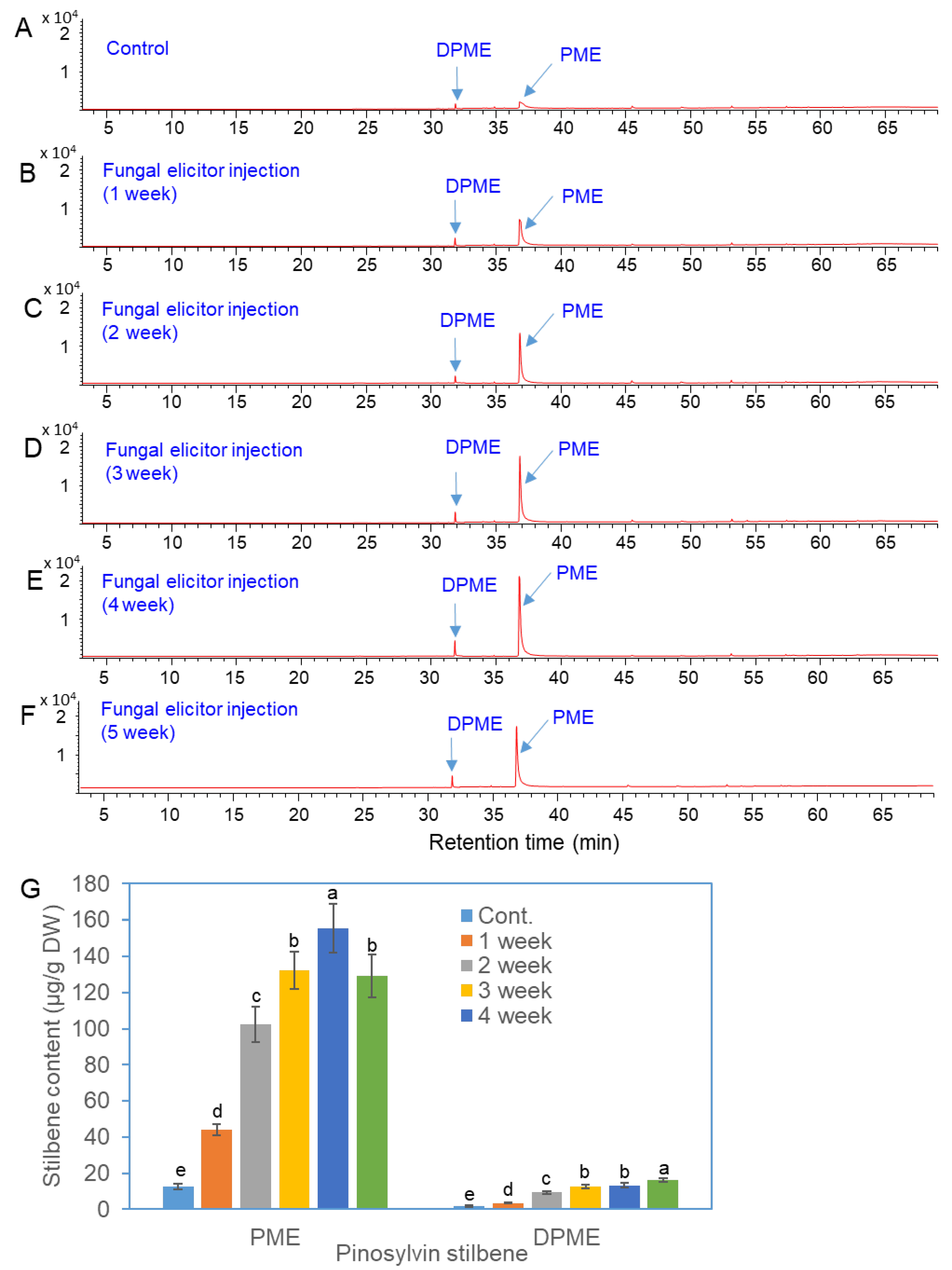
Figure 5.
Effects of elicitor trunk injection on PWN mobility in the branches of P. densiflora plants. (a) Photographs of PWN infection on the hole of the distal excised portion of the branches. (b) PWN-infected branch segment in a 50 ml Falcon tube. The proximal excised portion of a branch immersed in water. (c) Photograph of PWNs moving from a branch excised from a plant without elicitor trunk injection. (d) Photograph of PWNs removed from a branch excised from plants with elicitor trunk injection. (e) Contents of DPME and PME in needles of control and three independent pine plants after trunk injection of fungal elicitors. (f) Number and mobility of PWNs removed from branch segments of control plants and fungal elicitor-treated plants by trunk injection. Different letters above the bars indicate significantly different values (P<0.05), which were calculated via one-way ANOVA followed by Duncan’s post hoc analysis.
Figure 5.
Effects of elicitor trunk injection on PWN mobility in the branches of P. densiflora plants. (a) Photographs of PWN infection on the hole of the distal excised portion of the branches. (b) PWN-infected branch segment in a 50 ml Falcon tube. The proximal excised portion of a branch immersed in water. (c) Photograph of PWNs moving from a branch excised from a plant without elicitor trunk injection. (d) Photograph of PWNs removed from a branch excised from plants with elicitor trunk injection. (e) Contents of DPME and PME in needles of control and three independent pine plants after trunk injection of fungal elicitors. (f) Number and mobility of PWNs removed from branch segments of control plants and fungal elicitor-treated plants by trunk injection. Different letters above the bars indicate significantly different values (P<0.05), which were calculated via one-way ANOVA followed by Duncan’s post hoc analysis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



