Submitted:
21 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Quorum sensing (QS) is a sophisticated bacterial cell-to-cell communication mechanism that allows bacteria to sense population density through the secretion and detection of diffusible small molecular signals. This process leads to the coordinated expression of specific genes at the transcriptional level. Over time, continuous research has elucidated the genetic elements and regulatory principles of QS. Recently, synthetic biology has leveraged these insights to construct genetic circuits incorporating QS components, enabling both intra-species and inter-species artificial communication. These QS-based genetic circuits hold significant potential for applications in biotechnology and biomedicine. This paper reviews several well-characterized microbial QS systems and their functional roles, while also introducing the application of QS-based genetic circuits in cellular communication across species. The paper further discusses the role of QS in the development of biological computing tools, population density regulation, and metabolic flux control, offering a forward-looking perspective on future advancements. For intra-species communication, the focus is on the use of QS systems in constructing biological computing tools, including toggle switches, biosensors, and logic gates in synthetic biology. These tools, designed on the QS mechanism, can more precisely coordinate cellular behavior by integrating biological control circuits to achieve spatial, temporal, and population-level regulation. In the context of inter-species communication, the introduction of QS systems plays a pivotal role in population density control and metabolic flow regulation. By recombining metabolic networks, QS enables the redistribution of metabolic flux in desired pathways, facilitating the regulation of population density and the co-culture of mixed strains. Moreover, combining QS with oscillator models has shown great potential in synchronizing microbial communities. In summary, in-depth research into QS mechanisms and their applications not only lays a solid foundation for understanding microbial ecological competition and dynamic balance, but also offers promising avenues for regulating pathogenic bacteria and developing innovative disease control strategies.
Keywords:
Quorum sensing
; Intra-species communication
; Inter-species communication
; Genetic circuits
; Synthetic biology
; Population density control
; Metabolic regulation
Background
The basic communication and simple information exchange between organisms is a sign of the existence of life. Previously, it was believed that only multicellular organisms had the ability to exchange information in any form, and very few bacteria could communicate through specific diffusible signaling molecules. With the continuous in-depth study of microbiology, it has been discovered that different “languages” can be used to communicate information between prokaryotes in the microcosm, and then make different responses to changing environments in a group model similar to multicellular organisms [1]. This kind of cell-to-cell communication engineering is called quorum sensing (QS), that is, bacteria can respond to changes in the cell density and species composition of the surrounding microbial community to change their behavior accordingly [2,3]. Quorum sensing involves the generation and release of extracellular signal molecules and the detection within the population. These signal molecules are called autoinducers (AI). The concentration of AI will continue to increase with the increase of cell density. After reaching a certain concentration, it can combine with the receptor protein in the cell to form a receptor protein complex, which ultimately changes the overall pattern of gene expression [4]. The process controlled by quorum sensing includes bioluminescence, synthesis of antibiotics, regulation of nitrogen fixation genes, expression of pathogenic genes, bacterial movement, and biofilm formation [5,6]. Using a single cell to carry out these processes is costly and inefficient, but using cell populations to carry out these processes will have relatively high efficiency [7]. Existing studies have shown that bacteria can use QS for specific intraspecies communication, or use QS to sense the number of bacteria in the surrounding population to regulate their own behavior and achieve interpopulation communication [8,9]. Analyzing the quorum sensing system and the cross-border information exchange and signal transduction mechanism mediated by it not only provides a new entry point for clarifying the pathogenic regulation mechanism of pathogens, elucidating the mechanism of microbial ecological competition and dynamic balance, but also for regulating microorganisms. The ecological system and the development of new disease prevention and control strategies have laid a solid foundation. Therefore, the author of this article will introduce in detail the main functions and molecular mechanisms of the quorum-sensing system and its application in biotechnology.
1. Quorum Sensing System of Natural Bacteria and Its Main Mechanisms
Bacteria can realize intra- and inter-species information exchange by synthesizing signal molecules by themselves so that they can perform functions that a single cell cannot complete like multicellular organisms [10]. Thus far, the autoinducer signal molecules discovered are basically divided into three categories: N-acylated homoserine lactones (N-acylated homoserine lactones, AHLs) signal molecules; amino acids and short peptides (autoinducing peptides, AIPs) ), these two types of autoinducers mainly assist in the realization of intraspecific communication; and the third signal molecule is furanosyl borate diester (AI-2 type signal molecule), which can assist in the realization of interspecific communication [11]. The composition of common quorum-sensing systems in bacteria and their regulatory functions can be seen in Table 1. According to the different functions of bacterial populations, bacterial quorum sensing systems can be divided into four types: quorum sensing systems that regulate bioluminescence, biofilm formation, spore formation, and virulence.
1.1. Quorum Sensing System That Regulates Bioluminescence: LuxI/R, AI-2 Quorum Sensing System
In the 1960s, some people studied the luminescence phenomenon of marine fish’s symbiotic bacteria Vibrio fischeri, and found that the luminescence phenomenon of Vibrio fischeri colonized the luminous organs of Hawaiian squid. The density changes are related [23]. When in seawater, the concentration of Vibrio fischeri is low, and the concentration of surrounding signal molecules is also low, and the entire bacterial population appears black. Bioluminescence will only occur when the bacterial density reaches a certain concentration, which will be more conducive to the predation and avoidance of natural enemies of marine fishes symbiotically with Vibrio fischeri [24]. As the other side of mutually beneficial symbiosis, the marine fish will also provide a superior living environment for Vibrio fischeri, so that it can use the rich nutrients in the host to achieve very high community density [25,26]. At present, the LuxI/R quorum sensing system of Vibrio fischeri is used as a classic model for quorum sensing research on Gram-negative bacteria (Figure 1). The bioluminescence gene lux gene cluster of Vibrio fischeri is composed of 8 genes (luxA~E, luxG, luxI, and luxR) [27]. Its quorum sensing system includes two operons luxICDABE and luxR. The operon luxICDABE is responsible for encoding the self-deductor synthase gene (luxI) and luminescence gene (luxCDABEG). LuxI synthesis signal molecule N-3-oxo-hexanoyl homoserine lactone (3-oxo-C6-HSL, OHHL). The operon luxR is responsible for encoding the signal molecule receptor protein LuxR. In the case of low cell density, luxI performs transcription at a basic level. As the cell density increases, OHHL accumulates in the medium, and after reaching the threshold level, it will bind to the autoinducer domain of LuxR to form a complex. Then, the LuxR-OHHL complex will bind to the lux promoter region (lux box) upstream of luxI, starting the expression of the luciferase subunit gene luxICDABEG [28], and realize the luminescence of the Vibrio fischeri population. At the same time, a positive feedback loop will be formed to further express luxI and produce more OHHL [4,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52].
In addition to LuxI/R quorum sensing system, the marine bacteria Vibrio harveyi has both gram-negative and positive quorum sensing regulatory systems to jointly regulate bioluminescence phenomena [53]. Like other Gram-negative bacteria, Vibrio hawaii can produce the signal molecule AHL as an autoinducer (AI-1), but its signal sensing and transmission components are similar to the two-component protein of Gram-positive bacteria [54]. Vibrio hawaii also produces the second set of autoinducer system AI-2, which is different from homoserine lactone autoinducers in other bacteria. It is synthesized by LuxS protein and is generally used as a signal molecule for information exchange between bacterial species [55]. The specific regulation method of the quorum sensing system of Vibrio hawaii is as follows: When the cell density is low, due to the lack of AI-1 and AI-2, LuxN and LuxQ as phosphokinases will phosphorylate LuxN, and then phosphorylate LuxO. The latter LuxO becomes an active repressor protein, which indirectly inhibits the transcription of the luciferase gene luxCDABE, so Vibrio hawaii does not produce fluorescence when the cell density is low. When the bacterial population density reaches a certain threshold, the autologous inducers AI-1 and AI-2 will interact with LuxN and LuxPQ, respectively. After it has phosphatase activity, it transmits information to the phosphotransferase LuxU, which dephosphorylates LuxO into an inactive repressor protein, releases the inhibitory effect on luxCDABE, and makes Vibrio hawaii luminescence [56]. Similar to this mechanism, Vibrio cholerae also uses phosphorylation-dephosphorylation, a complex quorum sensing system to regulate gene expression [57].
1.2. Quorum Sensing System Formed by Biofilm: Las/Rhl/Quinolone Quorum Sensing System
Pseudomonas aeruginosa is an important pathogenic bacteria that cause cancer, AIDS, and cystic fibrosis (CF). It can be isolated from soil and water and is mediated by homoserine lactones. The QS system regulates the process of its life activities [58,59]. The reason Pseudomonas aeruginosa has become a typically conditioned pathogen is mainly that it can release virulence factors, including exoproteases, lipases, and exotoxins, among which most of the virulence factors are regulated by quorum sensing [60]. In addition, Pseudomonas aeruginosa often adheres to the surface of human tissues, and by secreting some lipid proteins and fibrin, it wraps itself to form a highly organized membrane-like polymer, that is, a biofilm, which resists the human immune system. And the treatment of traditional antibiotics [61]. Quorum sensing system will affect the resistance of Pseudomonas aeruginosa by regulating the formation of biofilm structure, bacterial adhesion, and its dissociation from biofilm and recolonization. Studies have found that after Pseudomonas aeruginosa colonies form a biofilm, the drug resistance will be significantly improved [62], the mechanism is likely to be that the biofilm will prevent antibacterial drugs from entering the membrane, especially for the deeper bacteria in the membrane.
The quorum-sensing system of Pseudomonas aeruginosa includes Las and Rhl signaling systems based on acylated homoserine lactone [63] and 2-alkyl-4(1H)-quinolone [2-alkyl-4(1H)-quinolone, AHQ]’s quinolone signaling system, etc. [64], they together constitute a very complex cascade control system. A large number of genetic studies have previously revealed the two quorum sensing systems LasR/I and RhlR/I. The autoinducer molecules of the Las and Rhl systems are the AHL homologue 3-oxododecanoyl homoserine lactone [N-(3-oxododecanoyl)-L-homoserine lactone, 3-oxo-C12-HSL] and butyl homoserine lactone (N-butanoyl homoserine lactone, C4-HSL).
The Las and Rhl systems are composed of transcriptional activators lasR and rhlR and signal syntheses lasI and rhlI, respectively. Through the action of these two quorum sensing systems, bacteria can express different virulence factors at different stages of infection to improve toxicity [65]. Quinolone contains more autoinducer molecules, 2-heptyl-3-hydroxy-4-quinolone (PQS), and its precursor 2-heptyl-3-hydroxy-4-quinoline (HHQ) is two of them. In the Las system, when the cell population density is low, LasI will produce lower levels of 3-oxo-C12-HSL. As the population density increases and reaches a certain threshold, 3-oxo-C12-HSL will combine with LasR. The complex LasR-3-oxo-C12-HSL they form will activate many genes for transcription, including lasB, toxA, rhlR, and lasI. When the complex activates lasI, a positive feedback regulation pathway will be produced. At the same time, 3-oxo-C12-HSL itself also has immunomodulatory activity, and 3-oxo-C12-HSL alone is related to the virulence of Pseudomonas aeruginosa. When the complex activates rhlR, a cascade effect of Las and Rhl quorum sensing systems will occur, and it will control the activity of RhlR at a later stage of transcription. Subsequently, RhlR activates the expression of genes related to the production of various secondary metabolites (such as hydrogen cyanide). At the same time, the RhlI/R system can feedback control the expression of the LasI/R system. In addition, the signal molecule PQS of the quinolone quorum sensing system also has a cascade regulatory relationship with the Las and Rhl systems. The synthesis of PQS is positively regulated by LasI and negatively regulated by RhlI, while the PQS system can positively regulate the expression of RhlI and LasI [66]. These three systems regulate each other to make bacteria better adapt to the environment, and the quinolone signaling system has more control over the pathogenicity and environmental adaptability of bacteria [67].
1.3. Quorum Sensing System for Sporulation: CSF Quorum Sensing System
Bacillus subtilis bacteria, a single cell that is rod-shaped, has no capsule, has flagella around it, and can produce spores. Because of its widespread existence in nature, it has the characteristics of high-temperature resistance, acid and alkali resistance, and extrusion resistance. At present, the bacteria are mainly used as enzyme-producing bacteria in industry, agriculture, medicine and health, and food health [68,69]. There are mainly two signal molecules involved in the Bacillus subtilis quorum-sensing system: ComX pheromone, and competence and sporulation factor (competence and sporulation factor, CSF). ComX encodes a Pre-ComX leader peptide containing 55 amino acid residues, and Pre-ComX is then secreted out of the cell membrane after being processed and modified by the isoprene converting enzyme ComQ [70]. The activity of the pheromone is controlled by the ComP/ComA two-component regulation system. When the bacterial population density reaches the threshold, the pheromone will bind to the related receptor ComP on the membrane to activate the enzyme and promote its autophosphorylation; at the same time, phosphorylation modified ComP (ComP~P) will activate the phosphorylated ComA (ComAP). ComAP will directly activate the expression of related downstream genes, such as activating the srfA promoter and induce the differentiation of subgroups of Bacillus subtilis into a competent state [71,72]. CSF is coded by the gene PhrC, which is a species-specific quorum-sensing molecule that enables different strains of Bacillus subtilis to communicate through quorum sensing. Under the action of oligopeptide permease (Opp), extracellular CSF is transported into the cell [73]. When the intracellular concentration is low, CSF can promote the development of competence. It binds to the phosphatase RapC and inhibits its activity, which increases the level of phosphorylated ComA and promotes the expression of ComS. With the deterioration of environmental conditions and the consumption of nutrients, intracellular CSF will promote the differentiation of bacterial subgroups into the sporulation stage after reaching a certain threshold concentration [74].
1.4. Virulence-Producing Group Induction System: Agr Group Induction System
Staphylococcus aureus is a common human pathogen and the pathogen of many acute and chronic infections. It secretes a large number of toxic factors such as toxins, hemolysin, and phenol-soluble regulatory proteins. Ji et al. 1995 [75] found that the expression of Staphylococcus aureus pathogenic genes is affected by bacterial cell density. Under the regulation of Agr quorum sensing mechanism, autoinducer peptides are secreted out of the cell as signal molecules and drive the expression of disease-causing genes by responding to a series of downstream signal molecules. The core agr operon of the Agr quorum-sensing system includes the precursor peptide synthetase AgrD of AIP, AgrB responsible for encoding processing proteins, AgrC encoding a histidine kinase sensor, and AgrA, a response regulator that induces transcription. In this quorum-sensing system, AgrD will maintain a low expression level under normal conditions, and the signal molecule AIP is obtained under the processing of AgrB and is continuously secreted outside the cell. When the concentration of extracellular AIP reaches a certain threshold, AgrC will be activated to cause autophosphorylation and phosphorylation of the reaction regulator AgrA. The phosphorylated AgrAP will bind to the promoter region upstream of agrABCD to form a positive feedback and start the expression of agrABCD gene and other genes controlled by the Agr regulatory system [76], which will ultimately achieve the expression regulation of the target gene.
2. A Synthetic Biology Quorum Sensing Gene Circuit Involved in the Dynamic Regulation of Intracellular Communication
In the microbial population, bacteria will be affected by the interaction of the same or different kinds of cells adjacent to it and the external environment. They have long-term symbiosis and coexistence, forming a complex signal molecular communication system through continuous evolution. Bacteria generate and induce chemical signal molecules through the quorum-sensing regulation system, and synchronously regulate the expression of a group of genes in a species complexity and population density-dependent manner so that bacterial populations can interact and contain, and finally achieve mutual adaptation [77,78]. At present, the research on quorum sensing systems has been relatively thorough, and the mechanism of action has gradually become clear. It is widely used to characterize complex intracellular response mechanisms and construct different quorum-sensing gene circuits, which has a profound impact on the development of synthetic biology. The quorum-sensing system of bacteria is mainly divided into two categories: intra-cell communication and inter-cell communication. The current research and review of quorum-sensing intra-cell communication are relatively concentrated on biofilm formation and quorum sensing quenching. As well as environmental monitoring, there are relatively few aspects such as building biological computing tools, regulating population density, and regulating metabolic flow. Therefore, the following will first focus on the main research results of toggle switches, biosensors, and logic gates in synthetic biology in quorum sensing intraspecies communication in recent years.
2.1. Toggle Switch
The main purpose of metabolic engineering is to use multigene recombination technology to improve the yield and production efficiency of target products. The traditional methods of modification in this field are mainly gene knockout and overexpression, but gene knockout may cause the growth of the bacteria to decline [79,80]. Therefore, it is necessary to activate and deactivate the expression of specific genes in time. Toggle switches are a useful tool to solve this problem. They are a large class of metabolic control switches that can be applied to synthetic gene circuits to increase the yield of target products [81]. Our laboratory Gu et al. [82] increased the shikimate yield of wild-type E. coli BW25113 to 1.73 g/L by inhibiting protein inactivation, blocking the competitive pathway, and overexpressing key enzymes. Then a toggle switch that can regulate the expression of aroK was constructed. It was found that the expression of aroK in the recombinant strain P-9 would maintain a high level during the growth phase, and then the expression level could be reduced by adding an optimal concentration of the inducer within an appropriate time. Finally, it was found that the recombinant strain P-9 could produce 13.15 g/L shikimic acid without adding any aromatic compounds, which proved the successful construction of auxotrophic E. coli that synthesizes shikimate.
Now, researchers are trying to construct a series of new gene circuits by combining toggle switches with quorum sensing systems to achieve the purpose of transformation and achieve interspecies communication among bacterial populations. Studies have shown that the quorum-sensing communication system not only participates in the expression and regulation of microbial pathogenic genes but also directly affects the resistance of microorganisms [83,84]. Salmonella, which secretes anticancer proteins, has an inhibitory effect on tumors, but non-specific expression can cause damage to healthy tissues. Therefore, Swofford et al. [85] to construct a tumor-sensitive gene expression switch and avoid harm to healthy tissues, integrated the Lux quorum-sensing system and a GFP fluorescent reporter gene into non-pathogenic Salmonella. The QS system and constitutive Salmonella were co-injected into tumor-bearing mice to quantify the protein expression in mice. At the same time, immunofluorescence technology was used to quantitatively locate the bacteria and fluorescent protein GFP in the tumor, and it was found that the newly constructed switch can be used as a very useful cancer treatment tool. It can target the tumor well and at the same time treat healthy tissues. No damage.
Soma et al. [86] combined the lux promoter with a positive feedback loop in E. coli, and constructed a synthetic Lux system as a cell density adjustable sensor. The researchers used the system to trigger the metabolic toggle switch (MTS) and constructed QS-MTS by integrating the two. Using QS-MTS at the ideal cell density, the metabolic flow is redirected from the TCA cycle to the production of isopropanol, thereby significantly increasing the production of isopropanol.
Engineered strains that can achieve self-regulation and redirection of metabolic flow can maximize the yield of the target product and reduce the need for human monitoring of the fermentation process. Therefore, researchers invested a lot of effort to construct a gene device independent of pathways, induction, and intervention. Gupta et al. [87] designed the QS mechanism to regulate the pfk-1 gene (this gene determines the carbon flux of glycolysis and cell growth), which can determine the best point to turn off gene expression according to the required time and cell density. When AHL is not present, the transcription regulator EsaR170V will bind to PesaS. With the continuous increase of cell density, the gradual accumulation of AHL will reduce the activity of EsaR170V and turn off the expression of pfk-1 gene. Therefore, most of the glucose will be transferred to the production route to increase myoinositol (MI) production.
In most cases, researchers use a single, common strategy to dynamically regulate metabolic flux pathways. Doong et al. [88] combined pathway-independent strategies and pathway-specific strategies to construct a control mechanism involving two dynamic orthogonal adjustable strategies. This mechanism can achieve hierarchical regulation and independently regulate the expression of two different enzymes, thereby increasing the production of D-glucuronic acid. After reaching the AHL concentration threshold, the pathway-independent strategy based on the QS system will convert the utilization of glucose from glycolysis to the production of D-glucuronic acid, thereby switching the cell from “growth” mode to “production” mode; the specific strategy is to use inositol as an intermediate metabolite sensor to achieve dynamic autonomous control, thereby increasing the production of D-glucuronic acid.
Currently, most reports on the QS system rely on down-regulation rather than up-regulation of gene expression, which greatly limits its potential as a molecular switch for regulating metabolic flux. Cui et al. [89] constructed a dual-function modular Phr60-Rap60-Spo0A QS system based on two natural initiators PabrB (Spo0A-P down-regulated) and PspoiiA (Spo0A-P up-regulated). By changing the position, number, and sequence of Spo0A-P binding sites, a promoter library with up-regulation and down-regulation capabilities was constructed. Then, the dual-function modular system was applied to the production of menaquinone-7 (MK-7) in Bacillus subtilis 168, and the production of MK-7 was significantly increased by 40 times after regulation by this dynamic pathway. Our laboratory Gu et al. [90] also constructed a dual-function dynamic switch based on the Esa-QS system, which can synchronize the up-regulation and down-regulation of genes at different times and intervals. The dual-function dynamic switch was applied to the synthesis of 5-aminolevulinic acid (5-ALA) and poly-β-hydroxybutyrate (PHB), and it was found that the yields were increased by 6 times and 12 times, respectively.
The above studies have combined the QS system with the metabolic toggle switch to activate the synthetic circuit after reaching the cell density threshold, thereby realizing the transformation of the self-induced metabolic state. As the application of this mechanism in metabolic engineering continues to mature, the combination of engineered microbial communities and QS-induced metabolic switches will continue to make new contributions to synthetic biology.
2.2. Biosensor
As people’s interest in building new biological elements that can regulate cell function grows stronger, people have begun to explore innovative tools in the field of biocomputing. Biosensors have received widespread attention due to their ease of use and low cost, and their excellent performance in providing real-time data under relevant physiological environmental conditions [91,92]. For example, a regulation mode that does not require the involvement of protein transcription factors but directly senses intracellular signal molecules or changes in the extracellular environment by nucleic acids, namely, riboswitches. It combines the aptamer domain and the expression domain, which can dynamically regulate gene expression by sensing the concentration of the substrate [93]. However, due to the lack of a correct understanding of the binding methods of ligands and aptamers and suitable screening methods, there are certain difficulties in the transformation of natural ribosomes. Our laboratory Pang et al. [94] proposed a strategy for the evolution and screening of N-acetylneuraminic acid (NeuAc) riboswitches in vivo. A high-threshold riboswitch was selected to optimize some pathways and key enzymes. After experimenting with glucose as the only carbon source, it was found that the highest yield of N-acetylneuraminic acid could reach 14.32 g/L.
Since then, many studies have found that the combination of biosensors and quorum sensing systems can be used as a new way of microbial modification to realize the transformation of the self-induced metabolic state. For example, genetically engineered whole-cell biosensors, which are powerful tools for detecting specific substances in bacterial proteins and pathways, have been widely used in synthetic biology research, including the detection of quorum sensing molecules (QSMs) [95,96]. Raut et al. [97] constructed and optimized the AI-2 whole-cell sensor system based on Vibrio harveyi BB170 strain. When AI-2 enters the cell, it binds to LuxP. When LuxP binds to AI-2, its conformation changes, which in turn initiates the transcription of luxCDABE, promotes the expression of luciferase and its substrates, and ultimately leads to bioluminescence, dependent on luciferin. The light signal produced by the enzyme is proportional to the AI-2 in the cellular environment. Therefore, the system can be applied to detect AI-2 levels in saliva, feces, and intestinal samples of patients with inflammatory bowel disease (IBD), providing an efficient, sensitive, and convenient method for the early detection and diagnosis of inflammatory bowel disease. The whole-cell biosensor based on the quorum sensing mechanism has played a great role in medical diagnosis and detection, and provided powerful help in biological control and ecological environment protection. For example, mercury pollution. Mercury is widely used in the chemical manufacturing industry or the electronics manufacturing field, but it has strong biological toxicity. Therefore, it is essential to develop a cost-effective, easy-to-operate, and environmentally friendly biological detection method for heavy metal mercury. Cai et al. [98] combined the positive feedback system based on quorum sensing with the mercury-specific operon MerR to construct a mercury-specific whole-cell biosensor containing an engineered Hg2+ tunable MerR protein. The Mercury-specific operon containing MerR protein controls the expression of LuxI, and the LuxR-AHL complex formed will activate Prlux and express RFP in turn. At the same time, three strategies QS1, QS2, and QS3 were designed to verify the amplification effect of the actual output signal of the biosensor, and it was found that the whole-cell probe containing a positive feedback system has a high sensitivity to mercury ions. Moreover, fitting the fluorescence signal to the classic Hill equation, it is found that the response of the constructed probe to mercury ions is close to the ultra-sensitive curve. In addition, Thapa et al. [99] used the principle of bioluminescence to develop a photobiosensor to detect the presence of pollutants in the environment, combining fiber optic technology and photodiodes to detect luminescence and convert the light signal into a measurable electric signal. The device can regulate the luminescence process through the Lux operon based on the quorum sensing mechanism so that it has a certain specificity for detecting heavy metals. When there are heavy metals in the environment, the luminescence phenomenon of bacteria will be weakened, but when barium is monitored, it is found that the luminescence phenomenon is both increasing and decreasing. That is to say, the constructed photobiosensor can be very good for the environment. Qualitative and quantitative detection of pollutants.
In the field of synthetic biology, many bioengineering systems and biological components are used to design clinical testing tools that are practical and easy to operate. However, only a few studies have proved the basic functionality of these testing tools in the clinic [100,101], and have not considered the complexity of clinical test samples and how to optimize the clinical testing tools. Therefore, researchers began to use cell-free protein synthesis systems as biosensors for pathogen diagnosis and treatment [102]. Wen et al. [103] constructed a modular DNA-encoded sensor with a cell-free protein expression system, which can be used to detect Pseudomonas aeruginosa infection markers in sputum samples of patients with cystic fibrosis. After optimizing the cell-free system, the experiment found that the biosensing system can quantitatively detect the quorum-sensing signal molecule 3-oxo-C12-HSL in sputum samples at the nanomolar level, which helps to improve the speed of clinical diagnosis.
2.3. Logic Gate
In recent years, with the rapid development of biotechnology, especially synthetic biology, a variety of logic gate control systems with complex gene circuits have been constructed in microorganisms to achieve precise control on space, time, and population scales. Positive feedback circuits and negative feedback circuits are ubiquitous in physical systems and biological systems [104]. The similarities and complexity between these two types of systems have attracted many researchers to develop biomolecular computing systems, in addition to studying toggle switches and biosensors, they have also achieved fruitful research results in the combination of logic gates and other biological control circuits [105]. Logic gate, also known as the “basic unit of digital logic circuit”, is a circuit that performs logical operations such as “or”, “and”, “not”, “NOR”, and “AND”. As early as 2002, some researchers constructed an AND gate system that initiated the line response by adding signal molecules from an external source [106,107]. In this logic gate, external sources add IPTG and ATC as the inducer input, and after the response, the expression of GFP is activated as the output. Since the reporter genes of most gene lines are only fluorescent proteins, they are basically used for signal detection. Therefore, afterwards, more researchers are committed to combining quorum sensing systems such as Lux, Las, and Rhl with gene circuits in microorganisms, building various logic gates based on the QS system, to better coordinate cells at the population level behavior, and promote the application in many aspects such as biomedicine and environmental control. Hu et al. [108] constructed an AND gate system based on Lux quorum sensing module in Shewanella oneidensis (MR-1) mtrA gene knockout mutant, and realized logic gates in microbial fuel cells (MFCs). In the application, they first integrated the IPTG response module, QS module, and output module (reporter gene or target gene mtrA) to form an AND gate system to control bacterial extracellular electron transfer. When the concentration of Lux-AHL reaches the threshold, it will combine with LuxR to promote the activation of Lux promoter. At the same time, with the addition of IPTG, the inhibition of the tac promoter by Lacl protein will be lifted. When the two input signals of this AND gate system respond simultaneously, extracellular electrons begin to transfer.
In the use of quorum sensing systems to design and gate dynamic control circuits, multiple operons have been widely used to regulate the behavior of a single promoter. However, when the cell recognizes two foreign signals at the same time, it may also initiate the expression of genes in the system. Therefore, it is possible to combine internal and external signal regulation to simultaneously drive gene expression. When an inducer is added exogenously, the cell can only initiate gene expression when it is in a specific physiological state. Shong et al. [109] have constructed and characterized two synthetic AND gate promoters, which require a group sensing signal and an exogenously added inducer to initiate gene expression. Both LEE and TTE gene expression require two signal molecules. When only one of them is present, the gene expression above the background level cannot be observed. The researchers assessed the ability of the promoter to function in a QS-dependent manner by adding endogenous AHL products, and observed that only in the presence of exogenous IPTG or ATC, gene expression would increase with the increase of cell density. Therefore, the AND gate regulatory system constructed by the researchers can be used as a model of a new regulatory system, integrating the QS system with other gene circuits to achieve dynamic changes in gene expression in metabolic engineering applications.
It has now been found that the QS system has a wide range of applications in metabolic engineering and can dynamically regulate the expression of target production genes. Since the physiological state of cells affects metabolic regulation, He et al. [110] in our laboratory designed a self-inducing AND gate control system that can respond to the system by simultaneously sensing cell concentration and cell physiological state. First, the random mutation of LuxR and the optimization of the luxRI promoter are performed, and a series of variable QS systems that can sense different cell concentrations are obtained. At the same time, by detecting the fluorescence intensity during cell growth, the corresponding stationary phase sensing system is selected. Finally, the newly constructed system was applied to PHB production, and it was found that PHB output could be increased by 1 to 2 times. The synthetic logic gate constructed by the researcher can be used as a tool of metabolic engineering, as a general dynamic induction system, which can sense complex signals without specific induction factors.
In synthetic biology, each biological element is treated as a Lego unit, and synthetic gene networks that perform different functions are obtained through modification and combination. In the past 10 years, toggle switches, biosensors, and logic gates designed based on the background of synthetic biology have been combined with some biological control circuits to achieve greater results in food, medicine and environmental reform, and are used to prevent and control humans. Microbial diseases and overcoming the problem of microbial drug resistance provide useful references and promote the cross-penetration and joint development of microbiology, genetics, plant protection, and other disciplines.
3. Synthetic Biological Quorum Sensing Gene Circuit Involved in the Dynamic Regulation of Inter-Specific Cell Communication
With the rapid development and convergence of synthetic biology and microbiology, gene synthesis circuits designed based on quorum sensing modules have rapidly developed into research hotspots in the field of microbiology in recent years. The specificity of the quorum sensing system of bacteria can ensure that bacteria recognize themselves in a complex environment and accurately realize information exchange. However, because bacteria do not exist independently in nature, they influence each other, and there are both symbiotic and competitive relationships. Therefore, to better adapt to the complex and changeable environment, bacteria can initiate the expression of specific genes through interspecies communication, thereby controlling the state of the entire community and improving the viability of the entire population [111]. Therefore, the application prospects of quorum sensing systems in inter-species cell communication will be reviewed in detail below.
3.1. Population Density Control
Metabolic engineering aims to design based on the existing cellular metabolic network and redistribute the metabolic flux of the required pathway through reorganization, thereby achieving high-value drugs, nutrients, and industrial chemicals [112]. In the process of using the substrate to produce the ideal target product, the yield cannot be maximized by relying only on the natural metabolic pathway. Therefore, to reconstruct the carbon metabolism pathway and increase the yield, Wang et al. [113] constructed an EP-bifido pathway in E. coli by combining the glycolysis pathway, the pentose phosphate pathway and the “bifidus shunt”. Applying this approach to the production of PHB, mevalonic acid, and fatty acids, it was found that it can reduce carbon dioxide emissions and increase the molar conversion rate of the product. But An et al. [114] and Goo et al. [115] proved that the nutrients in the microbial growth environment are usually limited, and the relatively crowded environment is not conducive to the growth and metabolism of microbes. Therefore, regulating population density is of great significance for increasing the yield of metabolites. Patidar et al. [116] took Platymonas sp and Oliver from the Bermuda high seas as the research objects, and used a two-stage culture mode for co-cultivation to evaluate the production of biofuels under different stress conditions. Based on the co-cultivation model, olive fungus and its metabolites on the high seas of Bermuda can promote the growth of flat algae, thereby increasing lipid production. HHQ and PQS mediate mutually beneficial symbiosis by increasing the cell abundance of olive bacteria on the high seas of Bermuda and forming a biofilm on the biological surface. However, when the concentrations of HHQ and PQS are higher, they can inhibit the growth of Platymona.
The gut microbiota plays a key role in establishing and maintaining host health. Kim et al. [117] used quorum sensing to reconstruct an information transmission system between natural E. coli and attenuated Salmonella typhimurium. Researchers functionalize one species to produce inducible signals, while another species can use gene integrated circuits to detect and record signals. This newly constructed signal transmission system successfully realized the intraspecies and interspecies signal transmission of microorganisms in the intestinal tract of mice and provided a basis for further understanding of the interactions between bacteria in other environments.
Brexó et al. [118] found that lactic acid bacteria (LAB) and wild yeast are inevitably produced during the production of bioethanol from sugarcane. Moreover, when lactic acid bacteria and yeast coexist in the fermenter, organic acids and other extracellular metabolites are produced, resulting in a decrease in ethanol production. The interaction between cells in the fermentation process will promote the communication between bacteria and yeast, and they are likely to coexist through group induction. Therefore, a better understanding of the mechanism of coexistence based on quorum sensing can generate a series of related theoretical and technical supports, and then make certain contributions to the biofuel industry.
Nowadays, many microbial ecologists try to construct some small synthetic ecosystems to explore the complexity of regulation mechanisms between populations. Scott et al. [119] combined two orthogonal QS mechanisms and population control mechanisms to control the population density of competitive Salmonella typhimurium. Lux-QS and Rpa-QS can combine with two lysis genes to form SLCs in the two bacteria for regulation. The results of the study show that when Lux-QS and Rpa-QS are combined with the lysis gene and then co-cultured, the initial population ratio between them is 100:1, and the population ratio can reach 1:1 after 10 h. However, when there is no lysis gene, the two strains will be taken over by Lux-QS strain after co-cultivation. That is, the integration of two orthogonal QS mechanisms to form an orthogonal decomposition system has great advantages for the co-cultivation of stable competing strains.
3.2. Oscillator
In a large-scale gene regulatory network, only a small number of regulatory factors can be used to regulate a relatively large number of genes to coordinate complex cell behaviors. Many metabolic activities, such as population control, hormone secretion, circadian rhythm, etc., are closely related to synchronous oscillation [120,121,122]. Therefore, many researchers are very interested in building oscillator models, and building synthetic gene oscillators (SGOs) has become one of the main directions of synthetic regulation research. McMillen et al. [123] combined the relaxation oscillator composed of PRE promoter, gene X (cii) and gene Y (ftsh) with Lux-QS mechanism in E. coli, and used the intercellular signal transduction mechanism to couple the oscillator and Induce synchronous oscillation. The protein Ftsh can degrade cii, and the complex of AHL and LuxR can activate the transcription of cii. When the intracellular AHL concentration reaches the threshold, it can bind to another LuxR in other cells to regulate its cii level. By modeling the system, it is also proved that the newly constructed mechanism will indeed achieve synchronization behavior across cell populations.
Although the application of oscillators in synthetic biology has made great progress, it is relatively difficult to find a stable gene circuit in a complex environment. This complex environment can cause considerable cell-to-cell differences in the genetic circuit, which may hinder function at the colony level. Prindle et al. [124,125] integrated the genetic relaxation oscillator, the gas-phase oxidative reduction signal between populations, and the Lux-QS system to form a genetic “biological mixture” coupled between different colonies. When it does not contain arsenite, ArsR will inhibit the expression of LuxR, so there is no fluorescence and oscillation. However, when the arsenite content is sufficient, this inhibitory effect will be lifted, and the LuxR-AHL complex will promote the expression and oscillation of fluorescent genes, and finally achieve the synchronization of thousands of oscillating populations on the centimeter scale.
In addition to the previous problems in the biotechnological application of synthetic biology based on oscillators, another challenge is to create a synergistic system of microorganisms based on population-level behavior that can use cell signaling mechanisms to regulate gene expression in multiple cell types. Chen et al. [126] constructed a microbial consortium composed of two different cell types, one of which is an “activation” strain and the other is a “repression” strain. These strains will produce two orthogonal cell signaling molecules that regulate gene expression in the dual-strain synthetic route. The activating factor produces Rhl-AHL, which promotes the target gene transfer of the two strains, while the inhibitor produces Cin-AHL, which inhibits the transcription of the two strains mediated by the LacI protein. In addition, there is a negative feedback loop in which the AiiA protein can degrade the two AHLs. These two strains will only vibrate at the population level when they are co-cultured. The ability to regulate population level dynamics through the use of genetic engineering of multi-cooperative strains points out the direction for the construction of complex tissues and organs with multiple cell types.
The use of cell signal transduction mechanisms to regulate gene expression of multiple cell types has been successful, but if synthetic multicellular systems are to be used in complex environments such as soil or intestinal microbiota, or combined with materials science, then through cell coordinated expression of genes across time and space will be essential. Kim et al. [127] constructed a two-strain synthetic flora, these two strains will produce two orthogonal quorum sensing molecules, thus forming a regulatory network that can connect positive feedback and negative feedback. When co-cultured in a microfluidic device, these two strains will oscillate. In a large group, it is found that the temporary coordinated oscillation of the entire group depends on the existence of an inherent positive feedback circuit, which amplifies and transmits intercellular signals. That is, the newly constructed multi-cell system can realize the coordinated expression of genes by using the short, short-distance coupling between the constituent cells.
The above studies show that the combination of the QS system and the oscillator model has great potential in regulating the synchronization of microbial complexes. It is conducive to the practical application of metabolic engineering, especially the development and optimization of production pathways for high-value metabolites. In the future, the design and construction of this part need to be further optimized to achieve more precise control.
3.3. Co-Cultivation of Mixed Strains
The continuous updates and improvements of synthetic biology and metabolic engineering components provide a favorable basis for the construction of more complex and efficient microbial synthesis systems, but may be limited by the resources and environment available in a single cell. To solve this problem, researchers have proposed a strategy, namely co-cultivation and fermentation, to overcome these limitations by coordinated distribution of substances between subgroups [128].
Dinh et al. [129] constructed a growth regulation circuit based on quorum sensing, which provides an additional parameter for adjusting the composition of the co-culture in the fermentation process. This line down-regulates the growth rate by reducing glycolytic flux and increasing cell density. The relationship between cell density and growth rate down-regulation can be adjusted by changing the expression level of AHL synthase to obtain different co-cultures under the same inoculation ratio dynamic. Applying this line to the co-culture production process of naringin, the titer can be increased by 60% by changing the strain inoculation ratio. In addition, it has been proved that after the growth control circuit is combined with the communication module, the cell density of one subspecies can regulate the transcription of the other module, thereby coordinating the behavior between cells and increasing the naringin titer.
Many Proteus can use quorum sensing to regulate the production of public products such as antibacterial agents and proteases, which can be shared in a microbial community. Evans et al. [130] constructed a laboratory model to study the importance of population regulation of antimicrobial agents in interspecies competition. In this model, both Burkholderia spp. and Spatholobus purpureus use antibacterial agents regulated by the QS mechanism to inhibit the production of other species. The researchers proved that in the co-cultivation model, Pollinin inhibited the emergence of quorum sensing defect mutants, and the increase in the production of antimicrobial agents made the population of purple flower color bacteria more competitive, that is, competition between species strengthen cooperative behavior by restricting mutants with quorum sensing defects. In addition, since the Pseudomonas aeruginosa QS system participates in the regulation of the production of a variety of public products, it can be a good model for studying microbial cooperation. Smalley et al. [131] co-cultured Pseudomonas aeruginosa and Bifidobacterium multivorum in the laboratory, and studied three kinds of Pseudomonas aeruginosa QS system-regulated antibacterial agents hydrogen cyanide, rhamnose ester and The importance of phenazine in biological competition. Through research, it is found that when Pseudomonas aeruginosa and Bifidobacterium polyphaga are co-cultured, Pseudomonas aeruginosa will be dominant, and these three antibacterial agents will jointly promote the competitiveness of Pseudomonas aeruginosa. Both of these bacteria require the Pseudomonas aeruginosa population to regulate proteases to promote growth. Therefore, QS-regulated antibacterial agents are important for Pseudomonas aeruginosa and can prevent the use of public proteases by bifidobacteria.
By summarizing the three aspects of QS mechanism in controlling population density, building oscillator models, and co-cultivating mixed strains, we have a further understanding of bacterial quorum sensing communication systems. It will help to create new theories and new methods for the regulation and control of microbial quorum sensing communication systems in the future, and regulate certain functions by artificially interfering or promoting the microbial quorum sensing system.
Conclusion and Outlook
Quorum sensing system in intercellular communication of microorganisms is one of the major discoveries in the field of microbiology research in the past 20 years, and there is a relatively comprehensive understanding and research on it. Microorganisms are widely present in various communities, and they can use some intercellular communication systems including complex signal molecules, signal sensors, and signal transduction mechanisms to quickly adapt to the ever-changing environment. This article reviews in detail the quorum sensing system that regulates bioluminescence, biofilm formation, sporulation, and virulence. It greatly improves the understanding of different quorum sensing systems in various microorganisms and helps in-depth study of the interaction between various regulatory network relationships. The microbial quorum sensing system in nature is very complex and diverse. There are still many signal molecules that have not been identified, and the signal pathways and regulation mechanisms are not clear. Therefore, in the era when small molecule separation and identification and protein structure analysis technologies are gradually mature, more energy should be devoted to the research and development of diversified and efficient quorum sensing systems to help reveal the mechanism of interaction between microorganisms and make it gain a competitive advantage, which greatly enhances the survival probability of them in the environment. The QS system has had a great impact on the social behavior of bacterial populations, making it better to adapt to changing environments. These studies have greatly enhanced people’s understanding of the application of quorum sensing systems in bacterial intra- and inter-cell communication, and further demonstrated the importance and complexity of studying microbial quorum sensing systems (Figure2). At present, many researches focus on the formation of biofilms, environmental testing, and disease control. There are relatively few studies on the construction of biocomputing tools and the regulation of population density. Therefore, this review explains in detail the dynamic regulation applications of bacteria based on quorum sensing mechanisms within and between species in recent years. It focuses on the construction of biological computing tools that combine quorum sensing systems with various basic regulatory circuits, and based on quorum sensing. The genetic elements of the company have improved in terms of controlling population density and regulating metabolic flow, striving to simplify the fermentation process, reduce costs, achieve precise regulation, and actively move closer to the direction of industrialization.
Actively developing synthetic biology transformation technology based on quorum sensing is of great scientific significance and application value for clarifying the information exchange between microorganisms and the communication system between pathogens and hosts: Firstly, understand the principles and mechanisms of the basic quorum sensing system of bacteria, research on bacterial quorum sensing communication systems and pathogen-host transboundary communication systems will help researchers establish a relatively complete microbial quorum sensing research system. Not only can provide some basic and scientific theoretical guidance, but also help researchers develop new ideas and new methods based on quorum sensing research. Secondly, the microbial communication system based on quorum sensing will participate in the regulation of various living habits and various physiological processes of bacteria. The QS system plays a good role in the communication between pathogens and between pathogens and hosts, making it the prevention and treatment of biological diseases. And new breakthroughs in the treatment of animal and plant diseases also provide a useful reference for the development of new green and safe agriculture and sound medical services. In addition, quorum sensing gene circuits have been widely used in the fields of industry, agriculture, medicine, and other fields. In various fields, new genetic elements and synthetic circuits are constructed through synthetic biology methods, which realizes the reorganization of metabolic flow and the increase of yield, and promotes microorganisms. The cross-integration and common development of various disciplines such as science, genetics, and botany. However, there are still some deficiencies or unclear points in the current research. For example, there is a certain leakage of dynamic regulation based on quorum sensing, which cannot achieve rigorous regulation of target gene expression; orthogonality between different regulatory systems and related interspecific cells. The applications of communication control of metabolic flow and pulse generators also need further research. In the future, it is believed that the strategy of optimizing dynamic circuits based on quorum sensing systems will have more room for development in the field of biochemical engineering.
References
- Geethanjali, et al., Quorum sensing: A molecular cell communication in bacterial cells. Journal of Biomedical Sciences, 2019. 5(2): p. 23-34.
- Bassler, B.L., et al., Intercellular signalling in Vibrio harveyi: sequence and function of genes regulating expression of luminescence. Mol Microbiol, 1993. 9(4): p. 773-86.
- Fuqua, W.C., S.C. Winans, and E.P. Greenberg, Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J Bacteriol, 1994. 176(2): p. 269-75.
- Galloway, W.R., et al., Quorum sensing in Gram-negative bacteria: small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem Rev, 2011. 111(1): p. 28-67.
- Choudhary, S. and C. Schmidt-Dannert, Applications of quorum sensing in biotechnology. Appl Microbiol Biotechnol, 2010. 86(5): p. 1267-79.
- Mangwani, N., et al., Bacterial quorum sensing: functional features and potential applications in biotechnology. J Mol Microbiol Biotechnol, 2012. 22(4): p. 215-27.
- Bassler, B.L. and R. Losick, Bacterially speaking. Cell, 2006. 125(2): p. 237-46.
- Clinton, A. and K.P. Rumbaugh, Interspecies and interkingdom signaling via quorum signals. Israel Journal of Chemistry, 2016. 56(5): p. 265-272.
- Straight, P.D. and R. Kolter, Interspecies chemical communication in bacterial development. Annu Rev Microbiol, 2009. 63: p. 99-118.
- Goo, E., et al., Control of bacterial metabolism by quorum sensing. Trends Microbiol, 2015. 23(9): p. 567-76.
- Ryan, R.P. and J.M. Dow, Diffusible signals and interspecies communication in bacteria. Microbiology (Reading), 2008. 154(Pt 7): p. 1845-1858.
- Ng, W.-L. and B.L. Bassler, Bacterial quorum-sensing network architectures. Annual review of genetics, 2009. 43(1): p. 197-222.
- Waters, C.M. and B.L. Bassler, Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol., 2005. 21(1): p. 319-346.
- Jakobsen, T.H., et al., Targeting quorum sensing in Pseudomonas aeruginosa biofilms: current and emerging inhibitors. Future microbiology, 2013. 8(7): p. 901-921.
- Neelapu, N.R.R., T. Dutta, and S. Challa, Quorum sensing and its role in Agrobacterium mediated gene transfer. Implication of quorum sensing system in biofilm formation and virulence, 2018: p. 259-275.
- Atkinson, S., et al., Quorum sensing and the lifestyle of Yersinia. Current issues in molecular biology, 2006. 8(1): p. 1-10.
- Wopperer, J., et al., A quorum-quenching approach to investigate the conservation of quorum-sensing-regulated functions within the Burkholderia cepacia complex. Applied and environmental microbiology, 2006. 72(2): p. 1579-1587.
- Barnard, A.M., et al., Quorum sensing, virulence and secondary metabolite production in plant soft-rotting bacteria. Philosophical Transactions of the Royal Society B: Biological Sciences, 2007. 362(1483): p. 1165-1183.
- Liu, X., et al., Characterisation of two quorum sensing systems in the endophytic Serratia plymuthica strain G3: differential control of motility and biofilm formation according to life-style. BMC microbiology, 2011. 11: p. 1-12.
- Carlier, A.L., Regulation of surface polysaccharides in Pantoea stewartii subsp. stewartii. 2008: University of Connecticut.
- Puskas, A., et al., A quorum-sensing system in the free-living photosynthetic bacterium Rhodobacter sphaeroides. Journal of bacteriology, 1997. 179(23): p. 7530-7537.
- Chu, W., et al., Role of the quorum-sensing system in biofilm formation and virulence of Aeromonas hydrophila. African Journal of Microbiology Research, 2011. 5(32): p. 5819-5825.
- Patel, K., et al., Spatially propagating activation of quorum sensing in Vibrio fischeri and the transition to low population density. Phys Rev E, 2020. 101(6-1): p. 062421.
- Nealson, K.H., T. Platt, and J.W. Hastings, Cellular control of the synthesis and activity of the bacterial luminescent system. J Bacteriol, 1970. 104(1): p. 313-22.
- Whiteley, M., S.P. Diggle, and E.P. Greenberg, Progress in and promise of bacterial quorum sensing research. Nature, 2017. 551(7680): p. 313-320.
- Whiteley, M., S.P. Diggle, and E.P. Greenberg, Corrigendum: Progress in and promise of bacterial quorum sensing research. Nature, 2018. 555(7694): p. 126.
- Swartzman, A., et al., A new Vibrio fischeri lux gene precedes a bidirectional termination site for the lux operon. J Bacteriol, 1990. 172(12): p. 6797-802.
- Stevens, A.M., K.M. Dolan, and E.P. Greenberg, Synergistic binding of the Vibrio fischeri LuxR transcriptional activator domain and RNA polymerase to the lux promoter region. Proc Natl Acad Sci U S A, 1994. 91(26): p. 12619-23.
- Liang, X., et al., Bifunctional Doscadenamides Activate Quorum Sensing in Gram-Negative Bacteria and Synergize with TRAIL to Induce Apoptosis in Cancer Cells. J Nat Prod, 2021.
- Chbib, C., Impact of the structure-activity relationship of AHL analogues on quorum sensing in Gram-negative bacteria. Bioorg Med Chem, 2020. 28(3): p. 115282.
- Nguyen, P.D.T., et al., Quorum sensing between Gram-negative bacteria responsible for methane production in a complex waste sewage sludge consortium. Appl Microbiol Biotechnol, 2019. 103(3): p. 1485-1495.
- Qais, F.A., M.S. Khan, and I. Ahmad, Broad-spectrum quorum sensing and biofilm inhibition by green tea against gram-negative pathogenic bacteria: Deciphering the role of phytocompounds through molecular modelling. Microb Pathog, 2019. 126: p. 379-392.
- Manner, S. and A. Fallarero, Screening of Natural Product Derivatives Identifies Two Structurally Related Flavonoids as Potent Quorum Sensing Inhibitors against Gram-Negative Bacteria. Int J Mol Sci, 2018. 19(5).
- Lipa, P., M. Koziel, and M. Janczarek, [Quorum sensing in Gram-negative bacteria: signal molecules, inhibitors and their potential therapeutic application]. Postepy Biochem, 2017. 63(4): p. 242-260.
- Melkina, O.E., Goryanin, II, and G.B. Zavilgelsky, [Histone-like protein H-NS as a negative regulator of quorum sensing systems in gram-negative bacteria]. Genetika, 2017. 53(2): p. 165-72.
- Lemfack, M.C., et al., Novel volatiles of skin-borne bacteria inhibit the growth of Gram-positive bacteria and affect quorum-sensing controlled phenotypes of Gram-negative bacteria. Syst Appl Microbiol, 2016. 39(8): p. 503-515.
- Kong, F.D., et al., Metabolites with Gram-negative bacteria quorum sensing inhibitory activity from the marine animal endogenic fungus Penicillium sp. SCS-KFD08. Arch Pharm Res, 2017. 40(1): p. 25-31.
- Chen, Z. and J. Xiang, [Advances in the research of LuxR family protein in quorum-sensing system of gram-negative bacteria]. Zhonghua Shao Shang Za Zhi, 2016. 32(9): p. 536-8.
- Papenfort, K. and B.L. Bassler, Quorum sensing signal-response systems in Gram-negative bacteria. Nat Rev Microbiol, 2016. 14(9): p. 576-88.
- Wang, D., et al., The Comparison of the Combined Toxicity between Gram-negative and Gram-positive Bacteria: a Case Study of Antibiotics and Quorum-sensing Inhibitors. Mol Inform, 2016. 35(2): p. 54-61.
- Stockli, M., et al., Coprinopsis cinerea intracellular lactonases hydrolyze quorum sensing molecules of Gram-negative bacteria. Fungal Genet Biol, 2017. 102: p. 49-62.
- Wang, D., et al., Mechanism-based QSAR Models for the Toxicity of Quorum Sensing Inhibitors to Gram-negative and Gram-positive Bacteria. Bull Environ Contam Toxicol, 2016. 97(1): p. 145-50.
- Wang, T., et al., Prediction of mixture toxicity from the hormesis of a singl`e chemical: A case study of combinations of antibiotics and quorum-sensing inhibitors with gram-negative bacteria. Chemosphere, 2016. 150: p. 159-167.
- Zhang, W. and C. Li, Exploiting Quorum Sensing Interfering Strategies in Gram-Negative Bacteria for the Enhancement of Environmental Applications. Front Microbiol, 2015. 6: p. 1535.
- Husain, F.M., et al., Sub-MICs of Mentha piperita essential oil and menthol inhibits AHL mediated quorum sensing and biofilm of Gram-negative bacteria. Front Microbiol, 2015. 6: p. 420.
- Morohoshi, T., et al., Inhibition of quorum sensing in gram-negative bacteria by alkylamine-modified cyclodextrins. J Biosci Bioeng, 2013. 116(2): p. 175-9.
- Husain, F.M. and I. Ahmad, Doxycycline interferes with quorum sensing-mediated virulence factors and biofilm formation in gram-negative bacteria. World J Microbiol Biotechnol, 2013. 29(6): p. 949-57.
- Galloway, W.R., et al., Applications of small molecule activators and inhibitors of quorum sensing in Gram-negative bacteria. Trends Microbiol, 2012. 20(9): p. 449-58.
- Camps, J., et al., Paraoxonases as potential antibiofilm agents: their relationship with quorum-sensing signals in Gram-negative bacteria. Antimicrob Agents Chemother, 2011. 55(4): p. 1325-31.
- Deng, Y., et al., Listening to a new language: DSF-based quorum sensing in Gram-negative bacteria. Chem Rev, 2011. 111(1): p. 160-73.
- Myszka, K. and K. Czaczyk, [Quorum sensing mechanism as a factor regulating virulence of Gram-negative bacteria]. Postepy Hig Med Dosw (Online), 2010. 64: p. 582-9.
- Chai, H., et al., Functional properties of synthetic N-acyl-L-homoserine lactone analogs of quorum-sensing gram-negative bacteria on the growth of human oral squamous carcinoma cells. Invest New Drugs, 2012. 30(1): p. 157-63.
- Mok, K.C., N.S. Wingreen, and B.L. Bassler, Vibrio harveyi quorum sensing: a coincidence detector for two autoinducers controls gene expression. EMBO J, 2003. 22(4): p. 870-81.
- Chen, X., et al., Structural identification of a bacterial quorum-sensing signal containing boron. Nature, 2002. 415(6871): p. 545-9.
- Waters, C.M. and B.L. Bassler, The Vibrio harveyi quorum-sensing system uses shared regulatory components to discriminate between multiple autoinducers. Genes Dev, 2006. 20(19): p. 2754-67.
- Freeman, J.A. and B.L. Bassler, Sequence and function of LuxU: a two-component phosphorelay protein that regulates quorum sensing in Vibrio harveyi. J Bacteriol, 1999. 181(3): p. 899-906.
- Zhu, J., et al., Quorum-sensing regulators control virulence gene expression in Vibrio cholerae. Proc Natl Acad Sci U S A, 2002. 99(5): p. 3129-34.
- Li, J., et al., Quorum sensing in Escherichia coli is signaled by AI-2/LsrR: effects on small RNA and biofilm architecture. Journal of bacteriology, 2007. 189(16): p. 6011-6020.
- Williams, P. and M. Camara, Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: a tale of regulatory networks and multifunctional signal molecules. Curr Opin Microbiol, 2009. 12(2): p. 182-91.
- Fuqua, C. and E.P. Greenberg, Self perception in bacteria: quorum sensing with acylated homoserine lactones. Curr Opin Microbiol, 1998. 1(2): p. 183-9.
- Zeng Lili, S.D., Niu Peiguang, Research progress on the quorum sensing system of Pseudomonas aeruginosa and its inhibitors. Antibiotics, 2014. 39(9): p. 701-705, 714.
- Singh, B.N., et al., Lagerstroemia speciosa fruit extract modulates quorum sensing-controlled virulence factor production and biofilm formation in Pseudomonas aeruginosa. Microbiology (Reading), 2012. 158(Pt 2): p. 529-538.
- Diggle, S.P., et al., 4-quinolone signalling in Pseudomonas aeruginosa: old molecules, new perspectives. Int J Med Microbiol, 2006. 296(2-3): p. 83-91.
- Diggle, S.P., et al., The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem Biol, 2007. 14(1): p. 87-96.
- Song, S.-S., et al., Plant responds to bacterial quorum-sensing system. Chinese Journal of Coll Biology, 2005.
- Parsek, M.R. and E.P. Greenberg, Acyl-homoserine lactone quorum sensing in gram-negative bacteria: a signaling mechanism involved in associations with higher organisms. Proc Natl Acad Sci U S A, 2000. 97(16): p. 8789-93.
- Schuster, M. and E.P. Greenberg, A network of networks: quorum-sensing gene regulation in Pseudomonas aeruginosa. Int J Med Microbiol, 2006. 296(2-3): p. 73-81.
- Jia, L., Applied characters and Status of Bacillus subtilis [J]. Meat Research, 2009. 11: p. 18-21.
- Ming, H., D. Lina, and T. Qing, Advances in Application Research of Bacillus subtilis [J]. Journal of Anhui Agricultural Sciences, 2008. 27.
- Ansaldi, M., et al., Specific activation of the Bacillus quorum-sensing systems by isoprenylated pheromone variants. Mol Microbiol, 2002. 44(6): p. 1561-73.
- Comella, N. and A.D. Grossman, Conservation of genes and processes controlled by the quorum response in bacteria: characterization of genes controlled by the quorum-sensing transcription factor ComA in Bacillus subtilis. Mol Microbiol, 2005. 57(4): p. 1159-74.
- Roggiani, M. and D. Dubnau, ComA, a phosphorylated response regulator protein of Bacillus subtilis, binds to the promoter region of srfA. J Bacteriol, 1993. 175(10): p. 3182-7.
- Bacon Schneider, K., T.M. Palmer, and A.D. Grossman, Characterization of comQ and comX, two genes required for production of ComX pheromone in Bacillus subtilis. J Bacteriol, 2002. 184(2): p. 410-9.
- Koroglu, T.E., et al., Global regulatory systems operating in Bacilysin biosynthesis in Bacillus subtilis. J Mol Microbiol Biotechnol, 2011. 20(3): p. 144-55.
- Ji, G., R.C. Beavis, and R.P. Novick, Cell density control of staphylococcal virulence mediated by an octapeptide pheromone. Proc Natl Acad Sci U S A, 1995. 92(26): p. 12055-9.
- Mayville, P., et al., Structure-activity analysis of synthetic autoinducing thiolactone peptides from Staphylococcus aureus responsible for virulence. Proceedings of the National Academy of Sciences, 1999. 96(4): p. 1218-1223.
- Case, R.J., M. Labbate, and S. Kjelleberg, AHL-driven quorum-sensing circuits: their frequency and function among the Proteobacteria. The ISME journal, 2008. 2(4): p. 345-349.
- Srivastava, D. and C.M. Waters, A tangled web: regulatory connections between quorum sensing and cyclic di-GMP. Journal of bacteriology, 2012. 194(17): p. 4485-4493.
- Herring, C.D., J.D. Glasner, and F.R. Blattner, Gene replacement without selection: regulated suppression of amber mutations in Escherichia coli. Gene, 2003. 311: p. 153-163.
- Kim, J.Y. and H.J. Cha, Down-regulation of acetate pathway through antisense strategy in Escherichia coli: Improved foreign protein production. Biotechnology and bioengineering, 2003. 83(7): p. 841-853.
- PANG Qingxiao, L.Q., QI Qingsheng, Application of switch for synthetic biology in metabolic engineering. Biotechnology Bulletin, 2017. 33(1): p. 58-63.
- Gu, P., et al., Tunable switch mediated shikimate biosynthesis in an engineered non-auxotrophic Escherichia coli. Sci Rep, 2016. 6: p. 29745.
- Chen, Y.F., et al., Quorum sensing and microbial drug resistance. Yi Chuan, 2016. 38(10): p. 881-893.
- LIANG Zhibin, C.Y., CHEN Yufan, et al., RND family efflux pump and its relationship with microbial quorum sensing system. Genetics, 2016. 38(10): p. 894-901.
- Swofford, C.A., N. Van Dessel, and N.S. Forbes, Quorum-sensing Salmonella selectively trigger protein expression within tumors. Proceedings of the National Academy of Sciences, 2015. 112(11): p. 3457-3462.
- Soma, Y. and T. Hanai, Self-induced metabolic state switching by a tunable cell density sensor for microbial isopropanol production. Metab Eng, 2015. 30: p. 7-15.
- Gupta, A., et al., Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit. Nature biotechnology, 2017. 35(3): p. 273.
- Doong, S.J., A. Gupta, and K.L.J. Prather, Layered dynamic regulation for improving metabolic pathway productivity in Escherichia coli. Proc Natl Acad Sci U S A, 2018. 115(12): p. 2964-2969.
- Cui, S., et al., Engineering a Bifunctional Phr60-Rap60-Spo0A Quorum-Sensing Molecular Switch for Dynamic Fine-Tuning of Menaquinone-7 Synthesis in Bacillus subtilis. ACS Synth Biol, 2019. 8(8): p. 1826-1837.
- Gu, F., et al., Quorum Sensing-Based Dual-Function Switch and Its Application in Solving Two Key Metabolic Engineering Problems. ACS Synth Biol, 2020. 9(2): p. 209-217.
- Kalia, V.C., Quorum sensing vs quorum quenching: a battle with no end in sight. 2015: Springer.
- Miller, C. and J. Gilmore, Detection of Quorum-Sensing Molecules for Pathogenic Molecules Using Cell-Based and Cell-Free Biosensors. Antibiotics (Basel), 2020. 9(5).
- Winkler, W., A. Nahvi, and R.R. Breaker, Thiamine derivatives bind messenger RNAs directly to regulate bacterial gene expression. Nature, 2002. 419(6910): p. 952-6.
- Pang, Q., et al., In vivo evolutionary engineering of riboswitch with high-threshold for N-acetylneuraminic acid production. Metab Eng, 2020. 59: p. 36-43.
- Nivens, D.E., et al., Bioluminescent bioreporter integrated circuits: potentially small, rugged and inexpensive whole-cell biosensors for remote environmental monitoring. J Appl Microbiol, 2004. 96(1): p. 33-46.
- Rawson, D.M., A.J. Willmer, and A.P. Turner, Whole-cell biosensors for environmental monitoring. Biosensors, 1989. 4(5): p. 299-311.
- Raut, N., P. Pasini, and S. Daunert, Deciphering bacterial universal language by detecting the quorum sensing signal, autoinducer-2, with a whole-cell sensing system. Anal Chem, 2013. 85(20): p. 9604-9.
- Cai, S., et al., Engineering highly sensitive whole-cell mercury biosensors based on positive feedback loops from quorum-sensing systems. Analyst, 2018. 143(3): p. 630-634.
- Thapa, A., et al. Development of a biosensor using photobacterium Spps. For the detection of environmental pollutants. in 2017 2nd International Conference on Bio-engineering for Smart Technologies (BioSMART). 2017. IEEE.
- Courbet, A., et al., Detection of pathological biomarkers in human clinical samples via amplifying genetic switches and logic gates. Sci Transl Med, 2015. 7(289): p. 289ra83.
- Pardee, K., et al., Rapid, low-cost detection of Zika virus using programmable biomolecular components. Cell, 2016. 165(5): p. 1255-1266.
- Soltani, M., et al., Reengineering cell-free protein synthesis as a biosensor: Biosensing with transcription, translation, and protein-folding. Biochemical Engineering Journal, 2018. 138: p. 165-171.
- Wen, K.Y., et al., A Cell-Free Biosensor for Detecting Quorum Sensing Molecules in P. aeruginosa-Infected Respiratory Samples. ACS Synth Biol, 2017. 6(12): p. 2293-2301.
- Nandagopal, N. and M.B. Elowitz, Synthetic biology: integrated gene circuits. science, 2011. 333(6047): p. 1244-1248.
- Shong, J. and C.H. Collins, Quorum sensing-modulated AND-gate promoters control gene expression in response to a combination of endogenous and exogenous signals. ACS synthetic biology, 2014. 3(4): p. 238-246.
- Din, M.O., et al., Interfacing gene circuits with microelectronics through engineered population dynamics. Science advances, 2020. 6(21): p. eaaz8344.
- Hasty, J., D. McMillen, and J.J. Collins, Engineered gene circuits. Nature, 2002. 420(6912): p. 224-230.
- Hu, Y., et al., Programming the quorum sensing-based AND gate in Shewanella oneidensis for logic gated-microbial fuel cells. Chemical Communications, 2015. 51(20): p. 4184-4187.
- Shong, J. and C.H. Collins, Quorum sensing-modulated AND-gate promoters control gene expression in response to a combination of endogenous and exogenous signals. ACS Synth Biol, 2014. 3(4): p. 238-46.
- He, X., et al., Autoinduced AND Gate Controls Metabolic Pathway Dynamically in Response to Microbial Communities and Cell Physiological State. ACS Synth Biol, 2017. 6(3): p. 463-470.
- Garg, N., G. Manchanda, and A. Kumar, Bacterial quorum sensing: circuits and applications. Antonie Van Leeuwenhoek, 2014. 105(2): p. 289-305.
- Schuster, S., T. Dandekar, and D.A. Fell, Detection of elementary flux modes in biochemical networks: a promising tool for pathway analysis and metabolic engineering. Trends Biotechnol, 1999. 17(2): p. 53-60.
- Wang, Q., et al., Engineering an in vivo EP-bifido pathway in Escherichia coli for high-yield acetyl-CoA generation with low CO2 emission. Metab Eng, 2019. 51: p. 79-87.
- An, J.H., et al., Bacterial quorum sensing and metabolic slowing in a cooperative population. Proc Natl Acad Sci U S A, 2014. 111(41): p. 14912-7.
- Goo, E., et al., Bacterial quorum sensing, cooperativity, and anticipation of stationary-phase stress. Proc Natl Acad Sci U S A, 2012. 109(48): p. 19775-80.
- Patidar, S.K., et al., Pelagibaca bermudensis promotes biofuel competence of Tetraselmis striata in a broad range of abiotic stressors: dynamics of quorum-sensing precursors and strategic improvement in lipid productivity. Biotechnology for biofuels, 2018. 11(1): p. 1-16.
- Kim, S., et al., Quorum sensing can be repurposed to promote information transfer between bacteria in the mammalian gut. ACS synthetic biology, 2018. 7(9): p. 2270-2281.
- Brexó, R.P. and A.d.S. Sant’Ana, Microbial interactions during sugar cane must fermentation for bioethanol production: does quorum sensing play a role? Critical reviews in biotechnology, 2018. 38(2): p. 231-244.
- Scott, S.R., et al., A stabilized microbial ecosystem of self-limiting bacteria using synthetic quorum-regulated lysis. Nature microbiology, 2017. 2(8): p. 1-9.
- Mondragon-Palomino, O., et al., Entrainment of a population of synthetic genetic oscillators. Science, 2011. 333(6047): p. 1315-1319.
- Potvin-Trottier, L., et al., Synchronous long-term oscillations in a synthetic gene circuit. Nature, 2016. 538(7626): p. 514-517.
- Tu, B.P. and S.L. McKnight, Metabolic cycles as an underlying basis of biological oscillations. Nature reviews Molecular cell biology, 2006. 7(9): p. 696-701.
- McMillen, D., et al., Synchronizing genetic relaxation oscillators by intercell signaling. Proc Natl Acad Sci U S A, 2002. 99(2): p. 679-84.
- Hasty, J., Sensing array of radically coupled genetic biopixels. 2012, Wiley Online Library.
- Prindle, A., et al., A sensing array of radically coupled genetic ‘biopixels’. Nature, 2012. 481(7379): p. 39-44.
- Chen, Y., et al., SYNTHETIC BIOLOGY. Emergent genetic oscillations in a synthetic microbial consortium. Science, 2015. 349(6251): p. 986-9.
- Kim, J.K., et al., Long-range temporal coordination of gene expression in synthetic microbial consortia. Nat Chem Biol, 2019. 15(11): p. 1102-1109.
- Jagavati, S., et al., Cellulase production by co-culture of Trichoderma sp. and Aspergillus sp. under submerged fermentation. Dyn Biochem Process Biotechnol Mol Biol, 2012. 6(1): p. 79-83.
- Dinh, C.V., X. Chen, and K.L.J. Prather, Development of a Quorum-Sensing Based Circuit for Control of Coculture Population Composition in a Naringenin Production System. ACS Synth Biol, 2020. 9(3): p. 590-597.
- Evans, K.C., et al., Quorum-sensing control of antibiotic resistance stabilizes cooperation in Chromobacterium violaceum. ISME J, 2018. 12(5): p. 1263-1272.
- Smalley, N.E., et al., Quorum Sensing Protects Pseudomonas aeruginosa against Cheating by Other Species in a Laboratory Coculture Model. J Bacteriol, 2015. 197(19): p. 3154-9.
Figure 1.
Schematic of LuxI/LuxR quorum sensing system (Vibrio fischeri as an example).
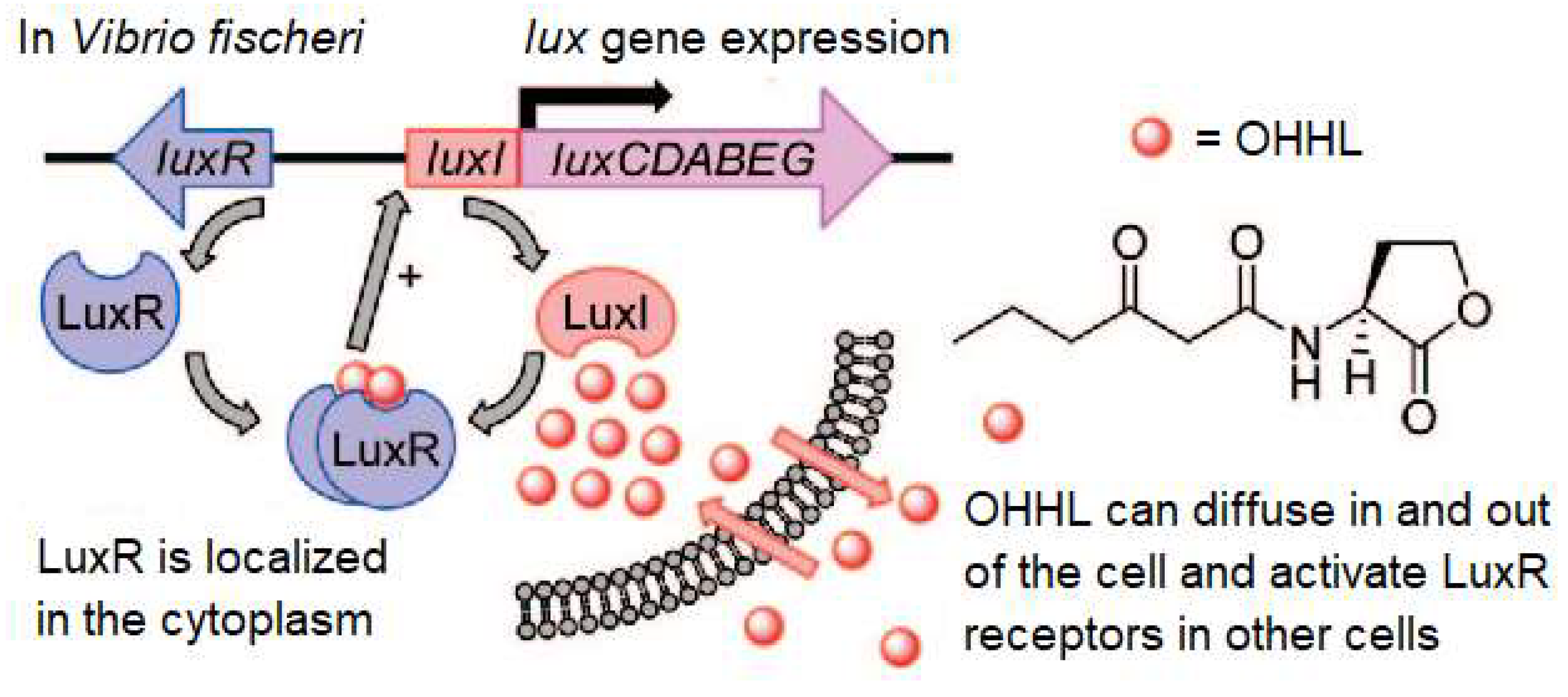
Figure 2.
Graphical representation of QS system control strategies covered in this review.
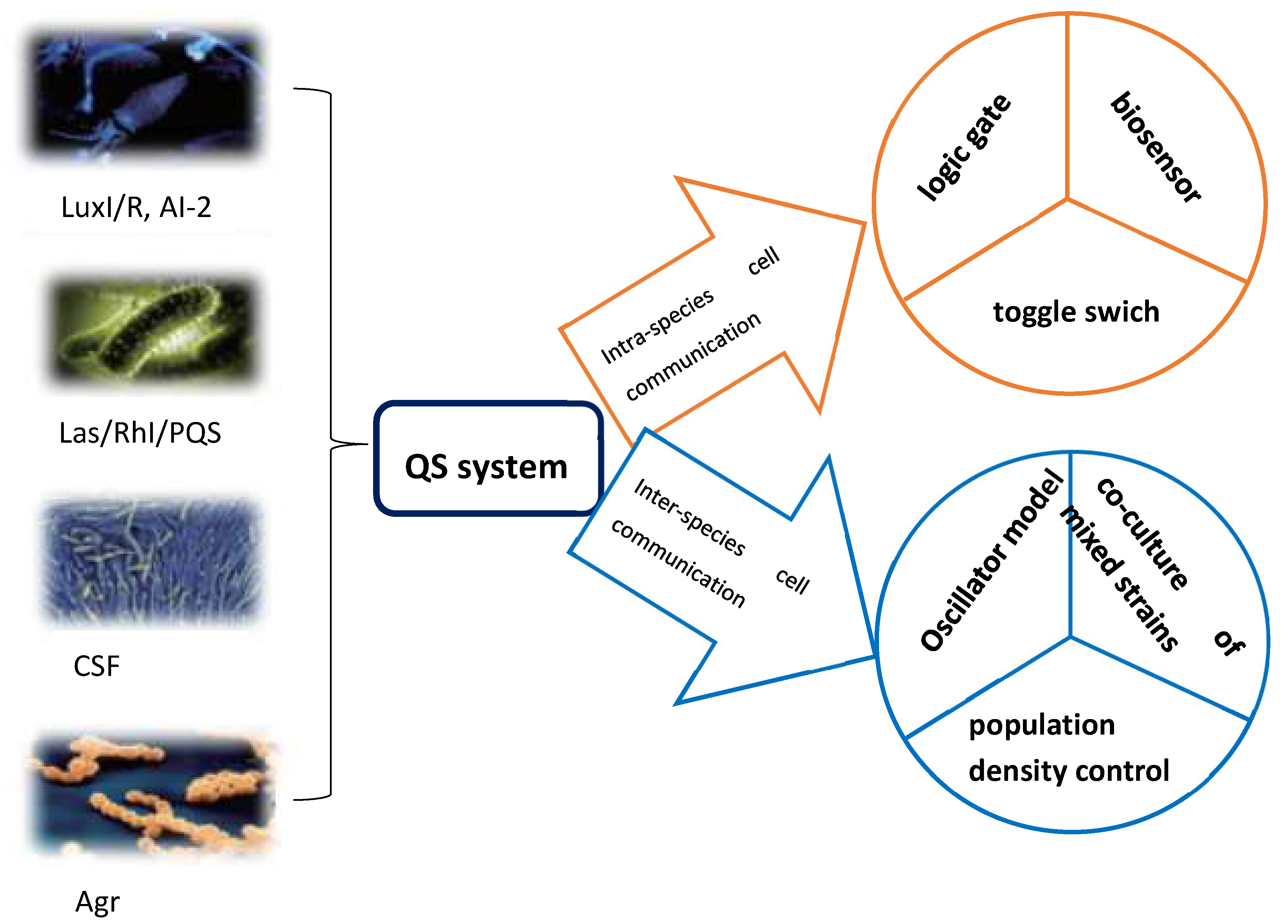

Table 1.
Typical QS systems and functions in bacteria.
| Bacterial species | Regulatory gene | Signal molecule(s) | Functions | References |
|---|---|---|---|---|
| Vibrio fischeri | luxI/luxR, luxS-luxP/Q | 3-oxo-C6-HSL AI-2 |
Bioluminescence, motility | [12,13] |
| Pseudomonas aeruginosa | lasI/lasR, rhlI/rhlR, pqsABCDE | 3-oxo-C12-HSL C4-HSL HHQ/PQS |
Virulence, motility and biofilms formation | [14] |
| Agrobacterium tumefaciens | traI/traR | 3-oxo-C8-HSL | Plasmid conjugation | [15] |
| Yersinia pseudotuberculosis | ypsI/ypsR, ytbI/ytbR | C6-HSL, 3-oxo-C6-HSL, C8-HSL | Motility, clumping and biofilms formation | [16] |
| Burkholderia cepacia | cepI/cepR | C8-HSL | Protease synthesis | [17] |
| Erwinia carotovora | expI/expR, carI/carR | 3-oxo-C6-HSL | Antibiotic synthesis | [18] |
| Serratia plymuthica | splI/splR, spsI/spsR, sptR | 3-oxo-C6-HSL, C6-HSL, C4-HSL | Biofilms formation | [19] |
| Erwinia stewartii | esaI/esaR | 3-oxo-C6-HSL | Virulence | [20] |
| Rhizobium leguminosarum | rhiI/ rhiR | C6-HSL, C8-HSL 3-hydroxy-7-cis-C14-HSL |
Expression of rhizosphere genes | [20] |
| Rhodobacter sphaeroides | cerI/cerR | 7-cis-C14-HSL | Evacuated bacterial accumulation | [21] |
| Aeromonas hydrophila | ahyI/ahyR | C4-HSL | Biofilms formation, exoproteases | [22] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



