
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Côte d’Ivoire has the main cashew tree culture area worldwide, but the cashew apple produced is still underutilized although its potential for industry. In the present study, the aim was to isolate and identify acetic acid bacterial strains, and to assess their potential use for cashew apple-based vinegar production. Vinegar mother from fermented juice was used to isolate acetic acid bacteria on a standard glucose-based medium. Physiological and biochemical tests followed by 16S rRNA gene analysis and phylogeny were used for isolate characterization. Moreover, their acetic acid production capacity was assessed. As results, five (05) strains of the Acetobacter genus were isolated. Phenotypic and phylogenetic analysis revealed that four of them, namely OYA2, OYA6, OYA9, and OYA10, belong to A. tropicalis / A. senegalensis species complex with 99,7% or 100% similarity. The last strain, OYA7, match to A. syzygii (99,7%). All the isolates were resistant to 15° alcohol (v/v) and grew well between pH 5.0 and 6.5. Their optimal growth temperatures varied between 27°C and 37°C, and only isolate OYA6 grows up to 40°C. They produced vinegar with yield (Yp/s , g/g) varying from 0.82 to 0.92, and acetic acid content (g/L) of 80.67 ± 2.1, 70.26 ± 0.9, 70.11 ± 1.7, 68.70 ± 1.5 and 67.22 ± 0.40 were obtained with OYA6, OYA7, OYA2, OYA10, and OYA9, respectively. Thus, the isolate OYA6 appeared as the best candidate for vinegar production, owing to its superior yield and thermotolerance ability that need to be further explored for industry use purpose.
Keywords:
Acetobacter
; indigenous microbiome
; vinegar
; acetic acid
; Cashew apple valorization
1. Introduction
Acetic acid bacteria (AAB) are generally known as vinegar-producing micro-organisms. They occur naturally on fruits and vegetables at the fermentable stage. They belong to the Acetobacteraceae family of the Alphaproteobacteria class [1]. The genus type of the family, namely Acetobacter, encompasses 34 species validly published to date, (https://lpsn.dsmz.de/genus/acetobacter); accessed on 17 August 2024) (Figure A1). AAB are a group of Gram-negative or Gram-variable bacteria, obligatorily aerobic, ellipsoidal or rod-shaped, non-spore-forming, and peritrich or polar flagellates. In general, they have a rapid oxidative fermentation capacity to oxidize ethanol to acetic acid and to oxidize a wide range of sugars and sugar alcohols to the corresponding organic acids in the presence of oxygen, although some are very low producers [2,3].
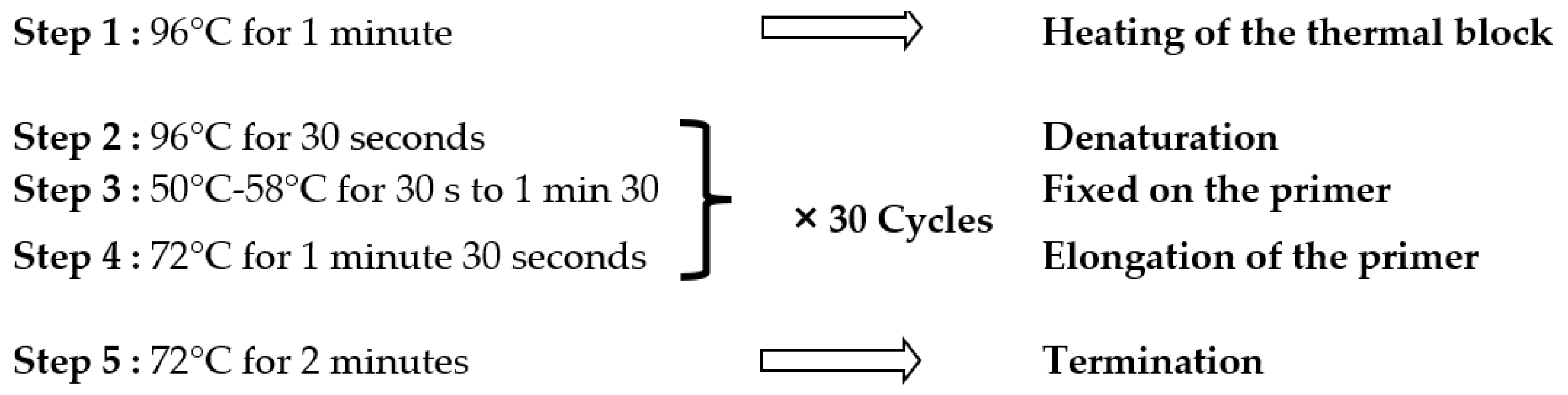
Figure 1.
PCR programming phase.
Vinegar is a precious additive and food supplement that has been used around the world for thousands of years [4]. According to the FAO [5] and the European Union standard (NF EN 13188-October 2000)[6], vinegar is produced from agricultural materials containing starch and/or sugar by the process of double fermentation, alcoholic and acetic. Acetic acid is the principal aromatic compound in vinegar, and techniques have been developed to improve vinegar production. In general, these improvements increase the rate at which ethanol is converted to acetic acid by bacteria of the genus Acetobacter. The temperature at which they grow and metabolize varies between 25°C and 30°C [7]. Temperature control is of particular concern, as ethanol oxidation is an exothermic reaction. An increase of 2 to 3°C causes deterioration in the rate of acetification and even inefficiency in vinegar production [8]. In tropical countries, an important selection parameter is the strain's thermotolerance, which, combined with the interest in vinegar quality, has led to a growing search for potential strains. The thermotolerant strains are used to avoid a low rate of acetification or fermentation stoppage [9]. Moreover, the use of strains also capable of high acid production is a desirable asset. To date, several acetic acid bacteria have been selected from tropical countries in African. For example in Senegal (West Africa), Ndoye et al. [10] selected two strains of Acetobacter tropicalis and A. pasteurianus for their ability to grow at 40°C and 45°C, respectively, and proposed them for the production of an artisanal spirit vinegar. These thermotolerant isolates, especially those from A. tropicalis species, are being used for industrial vinegar production in the sub-Saharan Africa [11]. A similar experiment was carried out in Ghana for isolating indigenous Acetobacter using traditional heap fermentations of cocoa beans, leading to the characterization and the taxonomic description of Acetobacter ghanensis sp. nov [12]. Thus, the bioprospection and selection of indigenous Acetobacter strains constitute a promising way for fostering the production of acetic acid and vinegar using domestic biomass [13,14].
Acetic acid bacteria could be difficult to isolate, due to the “viable but unculturable state” phenomenon [15]. It is also very difficult to correctly identify acetic acid bacteria down to the species level based on their physiological and biochemical characteristics [3]. Thus, for proper identification of AAB, the use of molecular methods and phylogenetic reconstruction is recommended [16]. As for their cultivation, the culture medium for acetic acid bacteria is generally composed of glucose, ethanol, peptone, yeast extract, calcium carbonate, and agar [17]. When the composition of carbon sources is altered, selective isolation of acetic acid bacteria can be expected [18]. For example, strains of Asaia bogorensis and Asaia siamensis were first isolated using sorbitol or dulcitol instead of glucose [19]. Thus, several carbon sources are used as enrichment cultures to efficiently isolate acetic acid bacteria to date [20], using diverse “non-conventional” and/or underutilized products such as mangoes [10], cocoa beans [12], papaya and sugarcane [21], in addition to standard fermented products like beer, vinegar and cider [16]. The cashew apple fruit is classified among the underutilized products in low-technological nations in Africa despite having high nutritional content [22]. In Côte d’Ivoire, the country which is the largest nut producer worldwide ( 1.300.000 tons of raw cashew nuts in 2024), the majority of apple fruit (an estimated of 7.776.870 tons) was left to rot in the field after removal of the raw nut [23,24,25,26]. This situation leads to important greenhouse gas (CH4 and CO2) emissions being released as a result of the non-controlled decomposition process [26]. Thus, there is a need of prospects for sustainable development of the cashew industry [23], and in the last decade, the Ivorian government has elaborated ambitious objectives to increase the percentage of products processed domestically [26]. In this context, this study aims to isolate and identify acetic acid bacteria from cashew apples, and to assess their potential to produce cashew apple-based vinegar.
2. Materials and Methods
2.1. Bacterial Strains and Isolation Technics
Ripe cashew apples were randomly collected from cashew tree in Yamoussoukro (6.74 N 5.38 W), the political capital of Côte d’Ivoire . The apples were washed and stored in a vat for 10 days. The apples were then pressed and 2L of the liquid collected was fermented at 30°C for 4 weeks. A vinegar mother solution was formed on the fermenting liquid. From this mother solution, 100 mL were collected and a decimal dilution was made up to 10-9. About 200 µl of the diluted solution were used to inoculate each agar plate (Ø 100 mm Petri dish) and all agar plates inoculated with solution from 10-3 to 10-5 dillutions were incubated at 30°C for 5 days. After the cells growth, single strain purity was verified by several replicates on nutrient agar [27].
2.2. Culture Medium
A medium based on glucose, yeast extract, calcium carbonate, penicillin and mycostatin (GYC) was used for isolation and culture. GYC culture medium consisted of Yeast extract 1%; Glucose 7.5%, Calcium carbonate 3%; Agar-agar 2% and Penicillin 0.5%. It was sterilized at 121°C for 15 min in an autoclave (HV 50 Hirayama, Japan), cooled to 45°C, and then 5% Mycostatin was added. Each Petri dish was filled with ca. 15 mL of prepared GYC medium for subsequent utilization.
2.3. Biochemical and Cultural Identification of Isolated Strains
Biochemical and cultural identification tests were carried-out to confirm that the strains isolated belonged to the Acetobacter genus. For cultural identification, we noted the shape, size, and color of the colonies. Conventional biochemical tests, including Gram stain, catalase, and oxidase tests, were performed according to the guidelines of Bergey's Manual of Systematic Bacteriology [28].
2.4. Molecular Identification
2.4.1. PCR Amplification of 16S rDNA and PCR Programming
Presumptive Acetobacter strains were subjected to genomic DNA amplification sequence analysis with primers 16Sd (5'-GCTGGCATGCTTAACACAACAT-3') and 16Sr (5'- GGAGGTGATCCAGCCGCAGGT-3') [29]. DNA was extracted from fresh cells following standard methods [30]. Briefly, for each isolate, a single colony was picked from a fresh culture on GYC medium and resuspended in 50 µl sterile water. After a brief vortex, the mixture was incubated at 95 °C for 15 min, centrifugate and the supernatant containing the release DNA was used as a template for the analysis. As for the PCR, the total reaction volume per strain was 50 µl (Table 1). For each amplification reaction, a microtube containing only the DNA-free reaction medium was used as a negative control.
PCR reaction conditions, carried-out in a thermocycler (Eppendorf, France) using a standard PCR program (Figure 1). PCR products were visualized by electrophoresis at 100V for 45 min on a 1% agarose gel in 1X TBE (Tris Borate EDTA) buffer. The molecular weight of reaction products was estimated with reference to a 1Kb molecular weight marker (100 bp to 12,000 bp) (Thermo ScientificTM SM0313, Thermofischer, France) to verify the expected size of amplified 16S rDNA which is around 1450 bp with the primers used [30].
2.4.2. 16S rDNA Sequencing
The PCR product was purified and sequenced using the Sanger method in the Eurofins laboratory (France). The sequences obtained were then assembled into double stranded DNA using the Bio Edit version 7.25 program. The sequences obtained were compared with those deposited in GenBank, using the BLAST algorithm (NCBI). BlastN with NCBI/GenBank online standard databases [nucleotide collection (nr/nt) and Whole-genome shotgun contigs (wgs)] was used for the identification of the isolates. The presumptive taxonomic affiliations obtained were further confirmed using (i) phylogenetic reconstructions and (ii) similarity level calculations between the novel sequences and the sequences of the selected reference strains [31]. All 16S rDNA gene sequences generated for the 05 isolates, and used in this study were deposited in GenBank under the accession numbers PP359644 to PP359648.
2.4.3. Phylogenetic Analysis and Similarity Distance Calculations
The phylogenetic analysis was inferred with the MEGA software version 7.0.26 [32]. Prior to the phylogenetic reconstruction, we carried-out a multiple sequence alignment by aligning the 16S rDNA gene sequences from the five isolates with those of the reference type strains collected in GenBank (Table 2). Briefly, 16S edited sequences were aligned with MUSCLE as implemented in MEGA software v.7, and a subsequent phylogenetic tree was reconstructed using the neighbor-joining (NJ) and maximum likelihood (ML) methods [33,34], with a Bootstrap value of 1000 [35]. The best-fit nucleotide substitution model of the aligned sequences was selected according to the Bayesian information criterion (BIC) [36] in MEGA software v.7. Finally, a pairwise similarity distance was calculated to infer the genetic similarity level between the sequences of isolated bacteria from Côte d’Ivoire and the selected reference sequences [37,38].
2.5. Selection of the Strains According to Physiological and Biochemical Characteristics
The selection of high-performance acetic acid bacteria strains comes down to their ability to withstand high levels of temperature, pH, and ethanol [46]. Strain kinetics are performed to determine growth optima. Acetobacter isolates harvested from fresh colonies grown on plates were used for the experiment. For each isolate, selected colonies were used to inoculate 10 mL of GYP broth (Glucose, Yeast Extract, and Peptone) and incubate for 7 days. Cell growth was estimated by measuring optical density (OD) at 600 nm using a spectrophotometer [47]. Bacterial growth rate was represented by µmax, the number of cells produced per unit of time. The details of each test are provided below:
- Thermo-tolerance test: Cultures were incubated at temperatures of 30 °C, 35 °C, 37 °C, and 40 °C.
- Effect of pH: Bacteria were grown at pH 3, 5, 6, 6.5 and 7
- Ethanol tolerance test: Bacteria were grown in GYP medium supplemented with at 5°, 10° and 15° alcohol (v/v).
2.6. Acetic Acid Production
The Acetobacter strains isolated and identified were used as a ferment to produce cashew apple vinegar. Acetobacter strains were grown on YG broth (yeast extract, glucose) to an OD600 of 0.7. Then 10 g/L were used to inoculate cashew apple alcohol to carry-out acetic fermentation. Batch fermentation (semi-continuous) was carried out in 500 mL Erlenmeyer flasks with 150mL alcoholic solution, at room temperature of 30°C ± 3 and pH 6 (Figure 2). Samples were taken at defined time intervals and analyzed.
2.7. Estimation of Acetic Acid
Acetic acid concentration was determined using the titratable acidity method described by AOAC (1990). A sample (1 mL) was pipetted into the 250 mL Erlenmeyer flask, and 20 mL distilled water and 5 drops of phenolphthalein were added to the flask. The sample solution was titrated with 0.1 N sodium hydroxide until a pink color was obtained. The acetic acid content was calculated using the following formula:
Where V = volume of final 0.1 N NaOH solution,
N = normality of NaOH solution,
60.05 = molar mass of acetic acid (g/mol)
W = weight of vinegar sample.
2.8. Fermentation Parameters
Yields were calculated at the end of fermentation. Volumetric acetic acid productivity (Qp , g/L/h) was calculated as the ratio between acetic acid concentration at the end of the run (Pf , g/L) and fermentation time (t,h).
Pf : (g/L) Acetic acid concentration at end of fermentation; t (h): Fermentation time.
Yield of acetic acid on ethanol consumed (Yp/s , g/g):
Where E0 and Ef g/L Initial and final ethanol concentration.
Ethanol conversion was calculated using this equation,
The efficiency of conversion of ethanol to acetic acid was estimated by the following relationship:
Where Yth is the theoretical value of Yp/s (1.36 g/g).
2.9. Statistical Analysis
The data collected were analyzed using STATISTICA 7.1 software. Mean ± SD were calculated and analysis of variance was considered significant when the p-value is inferior to α=5%. Tests for significant difference in mean values were performed using Duncan's multiple range test (p < 0.05).
3. Results and Discussion
Acetic acid bacteria (AAB) with good acid-producing potential can be found in cashew apple juices in different tropical countries [27]. The isolates characterized so far are intensively tested for acetic acid production and tolerance against different stresses, including ethanol, but their identification rely often on physiological and biochemical technics [27]. The genetic identity of the acetic acid bacteria isolated from the microbiome of cashew apple in tropical areas is still unknown. In Côte d’Ivoire, vinegar production trial with cashew apple have been explored, but the Acetobacter strains used have been isolated from various sources such as palm wine and fermented cocoa beans [48]. Thus, the present work which aims to isolate and assess the functional features of acetic acid bacteria using native fermented cashew apple juices provides the first comprehensive analysis of cashew apple microbiome in Côte d’Ivoire. The study used a multi-approach to analyse five (05) indigenous acetic acid bacteria from Ivorian cashew apples, including a biochemical and microscopic traits assessment, a molecular identification using 16S rRNA gene sequences, the test of tolerance of isolates against ethanol, temperature, and pH stresses as well as the assessment of their potential use as vinegar producers in laboratory conditons.
3.1. Isolation, Screening, and Identification of the Strains for Production of Acetic
3.1.1. Biochemical and Cultural Identification of Isolated Strains
After 5 days of incubation at 30°C, colonies showing a halo due to acidification of the GYC medium were selected [14,49] (Figure 3). In consequence, isolation results showed the presumptive presence of 05 acetic acid bacteria on GYC medium. When visualized, OYA2, OYA6, OYA10, OYA7, and OYA9 have the same brown color, but they have two types of shape, which consist of regular (e.g. OYA2) and irregular (e.g. OYA9) shapes (Figure 3).
Microscopic examination revealed that the bacteria are motile, the cells are long, filamentous and have pink fusiform swellings, making them Gram-negative (Figure 4). For Cire et al. [50], microscopic examination of acetic acid bacterial colonies isolated in mango alcohol in Senegal, in west-Africa, showed the same shape and Gram staining.
Colonies are colorless after a few seconds in the presence of N-dimethyl paraphenylene diamine oxalate. This means that isolated strains are oxidase-negative. Also, the catalase test performed on the bacteria enabled us to distinguish catalase-positive bacteria. All the morphological and biochemical characteristics of strains isolated from the Ivorian fermented cashew apple juice are presented in Table 3.
Acetic acid bacteria are difficult to identify down to species level on the basis of physiological and biochemical characteristics [3]. There are several reasons for this difficulty, including the occurrence of spontaneous mutations and related phenotypic changes [51]. Mutation causes a loss of several biochemical traits, such as ethanol oxidation, non-production of cellulose, non-production of brown pigment. For these reasons, an accurate identification of acetic acid bacteria required a sequencing of molecular markers such as the 16S rRNA coding gene.
3.1.2. Molecular Identification
The BlastN analysis allowed us to identify our isolates as Acetobacter strains. These taxonomic affiliations were further confirmed using a combination of a phylogenetic analysis and a similarity level calculation between the sequences of our isolates and those of the selected reference type strains (Table A1). As preliminary analyses showed the Ivorian isolates to belong to Acetobacter syzygii species, as well as to Acetobacter tropicalis and Acetobacter senegalensis species (Figure A1), subsequent phylogenetic analysis included the 16S sequence of all the 12 types strains closely related to these two lineages and available at the time of writing (August 13th, 2024). An additional three type strains of Acetobacter genus representative of the remaining lineages were also selected, making a total number of 15 Acetobacter reference strains for the final phylogenetic analysis (Figure 5). The analyses showed that strain OYA7 match to Acetobacter syzygii and shares 99.7% of similarity value with its type strain NBRC 16604T, while all the remaining isolates (i.e. OYA2, OYA6, OYA9 and OYA10) are genetically related to both the A. tropicalis and A. senegalensis species (Figure 5).
The analysis showed that the 16S sequences of isolates OYA10, OYA9 and OYA2 are 100% identical to Acetobacter tropicalis strain LMG 19825T & A. senegalensis strain LMG 23690T, (hereinafter referred to as A. tropicalis / A. senegalensis species complex) while the isolate OYA6 is 99.7% similar to the type species of these two species (Figure 5; Table A1). It is worth mentioning that the 16S rRNA gene sequence retrieved from the whole genome sequence (wgs) of A. senegalensis LMG 23690T (Accession number LHZU01000060.1) was used in this analysis instead of the 1386 bp-sequence archived under GenBank accession AY883036 (NR_043252) (deposited by Ndoye et al. in 2007 [8], which contained too much degenerate nucleotides (e.g. K, W, N) and may have quality concerns. With the data retrieved from the wgs, A. tropicalis type strain LMG 19825T & A. senegalensis type strain LMG 23690T shared 100% identical 16S rRNA gene sequence. Similar findings have been reported in previous studies for the 16S rRNA gene sequence of these two type species (e.g. [42,45]). The same trend of results was also obtained for two other couples of Acetobacter species, namely Acetobacter malorum LMG 1746T (LHZC01000018.1) and Acetobacter cerevisiae LMG 1625T (LHZA01000074.1), as well as Acetobacter sacchari VTH-Ai14T (LC103252) and Acetobacter nitrogenifigens DSM 23921T (BAPG01000028.1) (Figure 5), confirming the limited taxonomic resolution of the 16S rRNA gene to resolve the phylogeny of AAB species [16,52]. Although it is still recognized as universal molecular marker for identifying bacterial isolates, the 16S rRNA gene sequences (partial and full-length) were sometimes found to be too conserved for accurately differentiating many bacteria species, including soil and plant-associated bacteria [44,52,53,54]. As for the AAB species, the use of protein-encoding genes, such as dnaK, groEL and hsp60, has been shown to provide reliable delineations & phylogenies, to reflect overall genome similarities, and to match (digital) DNA–DNA hybridization (DDH) values [16,44,52]. These metrics could be applied to the 05 isolates for further taxonomic analysis. These analyses could help to determine whether these bacteria isolated from fermented cashew apple juice in Côte d’Ivoire, mainly OYA6 and OYA7, may represent novel lineages of acetic acid bacteria.
It is interesting to note that, by Blast analysis, the Ivorian isolates from the A. tropicalis / A. senegalensis species complex (OYA2, OYA6, OYA9 and OYA10) were found to be closely related to many thermotolerant AAB isolated from fermented plant beverages such as Acetobacter strain FF82 (Accession number LC462264.1) and Acetobacter strain FF35 (Accession number LC462263.1). The Ivorian isolates are also genetically similar to many other strains isolated from different substrates such as palm wine in Nigeria (e.g. Acetobacter strain AFF6; Accession number MT158803.1 for OYA6), sugarcane juice in India (e.g. Acetobacter strain MA; Accession number MG279707.1 for OYA2, OYA9 and OYA10) or cocoa beans in fermentation (e.g. Acetobacter strain At1 ; Accession number MN909151.1 for OYA6). The isolate OYA7, which is genetically related to A. syzgii species, appeared also highly similar (99,7%) to diverse Acetobacter strains isolated from food-derived sources such as a kefir grain (e.g. Acetobacter strain GM1 ; Accession number KX987248). Moreover, Acetobacter senegalensis type strain LMG 23690T which was also isolated from the same geographical region in Africa, i.e. the tropical sub-Saharan Africa, as the isolates OYA2, OYA6 OYA7, OYA9, and OYA10, was shown to be a thermotolerant acetic acid bacterium [8]. This strain was isolated in Senegal more than a decade and used nowadays for industrial vinegar production [11]. All these functional characteristics of the published strains that were found to be closely related to OYA2, OYA6, OYA7, OYA9 and OYA10 prompt us to perform various phenotypic analyses of our Acetobacter isolates to assess their ability to serve as good thermotolerant candidates for vinegar production. The data obtained are reported in the next section.
3.1.3. Physiological Characterization of Isolated Acetic Acid Bacteria Strains
The 05 strains isolated and identified as acetic acid bacteria were physiologically characterized to determine the effect of temperature, pH, and alcohol during their growth.
- Effect of temperature
Figure 6 below shows the effect of temperature on the acetic acid bacteria strains growth. It shows that the optimum growth temperature for the 05 strains ranges from 27°C to 37°C. Only strain OYA6 grows up to 40°C. Acetic acid bacteria are mesophilic, with optimum growth temperatures between 25 and 30°C. Above this optimum temperature, essential enzymes such as alcohol dehydrogenase and aldehyde dehydrogenase can be denatured, causing deterioration in the rate of acetification and even inefficiency in vinegar production [55]. However, the isolation of acetic acid bacteria able to grow at high temperatures will be important for the vinegar industry, in order to limit losses linked to bioreactor cooling, particularly in tropical countries [56].
- Effect of pH
The optimum pH for growth was between 5 and 7 (Figure 7). These results are comparable to those of Cire et al. [50], who reported a pH of 5-6.5 as the optimum condition for acetic acid bacteria growth. It should be noted that the optimum pH for growth of AAB lies between 5-6.5 [57,58]. In our work conditions, no growth was observed at pH 3 for any of the five (05) strains. On the other hand, strong growth was observed at pH 6.5 for all the isolates, but the Acetobacter OYA6 strain showed the strongest growth. These results are congruent with the growth partern expected from Acetobacter strains in general, although growths at lower or higher pH values have been also reported [59,60].
- Effect of alcohol
The 05 strains were grown at 5°, 10° and 15° alcohol (v/v) (Figure 8). These results are comparable to those of Sharafi et al. [49], who reported the isolation of bacterial strains able to grow at 5-11% v/v ethanol. This result confirmed the growth of AAB in the presence of low ethanol concentration [61,62] and showed that these strains are resistant to alcohol. This could be of interest to the vinegar industry in the search for ethanol-resistant acetic acid bacteria. Ethanol is the main substrate for acetic acid production. When there is a high concentration of ethanol, it can cause stress for acetic acid bacteria, inhibiting their growth and reducing acetic productivity [63]. However, obtaining high acetic acid concentrations requires the presence of acetic acid bacteria capable of tolerating high ethanol concentrations. In addition, an acetification process using high ethanol concentrations could improve the quality of the vinegar produced [64]
3.2. Acetic acid Fermentation
Acetic fermentation parameters were monitored and recorded in Table 4. All strains of acetic acid bacteria isolated from were fermented cashew juice from Côte d’Ivoire were found to be capable of producing acetic acid [65]. This could be explained by the fact that ethanol was oxidized by acetic acid bacteria to produce acetic acid [66]. Acetic acid production evolved progressively over time until day 24 (or 576 hours) of fermentation, with maximum values of 80.67 g.L−1 with strain OYA6, and values of 70.26 ± 0.9 g/L; 70.11 ± 1.69 g/L; 68.70 ± 1.51 g/L and 67.22 ± 0.37 g/L, respectively for strains OYA7, OYA2, OYA10, and OYA9 (Table 4). Statistical analysis reveals a significant difference in acetic acid production by Acetobacter OYA6 compared to strains OYA2, OYA7, OYA9, and OYA10. This is because Acetobacter OYA6 also grew well in the fermentation medium. In the present study, Acetobacter OYA6 had good physiological properties regarding the temperature, pH, and ethanol stresses. The experiments showed that the type of aeration are important factors in acetic acid production by Acetobacter strains, as reported before [49]. We showed that the use of semi-continuous fermentation could contribute to a high final acidity. It should be noted that in both continuous and semi-continuous modes, acetification cycles are achieved by renewal of the initially alcoholic medium inoculated with acetic acid bacteria. This hypothesis is supported by different authors [67] who claim that semi-continuous or continuous fermentation is this fermentation mode used in industry to improve yield. Moreover, Acourene et al. [68] have shown that every 36 hours of continuous acetic fermentation can produce date vinegar grading between 7.2 and 8.5 degrees acetic with an hourly productivity of between 0.458 - 0.488 g/L/hour. Successive analysis by acidity determination in our study reveals the moment of alcohol exhaustion in the medium. Furthermore, the lowest residual alcohol content is attributed to OYA6 (5.18 ± 0.13 g/L). All fermentations obtained had a residual alcohol content of less than 15 g/L standard recommended by current regulations [5]. The kinetics of acetic fermentation revealed the trend in acetic acid concentration. It can be seen that all curves show the same trend, reflecting the oxidation of ethanol by Acetobacter isolates as a function of time. These results corroborate those of authors who have carried-out acetic fermentation kinetics [47,49]. The five strains used in this study show better growth and ethanol oxidation capacity. With 92.6 g/L initial ethanol, no overoxidation was observed during acetic fermentation, as overoxidation is unfavorable to vinegar production. According to Saeki et al. [69], overoxidation does not occur when oxidizable ethanol and other carbon sources remain in the culture medium, or when the acetic acid concentration exceeds 4.5%. Thus, overoxidation can be avoided with a high initial ethanol concentration, and with ethanol-tolerant Acetobacter strains [56]. Taken togther, we can conclude that our strains have different features suitable for fruit vinegar production.
4. Conclusions
The search for acetic acid bacteria (AAB) in cashew apple juice in this study aimed at isolating and characterizing indigenous microorganisms capable of vinegar production in Côte d’Ivoire. The five (05) Acetobacter isolates are affiliated to A. syzgii (OYA7) and A. tropicalis / A. senegalensis species complex (isolates OYA2, OYA6, OYA9 & OYA10), and proved to be promising, as they can produce vinegar with, at least, acetic acid content of 67 g/L. These indigenous isolates are resistant to alcohol up to 15° alcohol (v/v) and their optimal growth occurs in acidic conditions, between pH ranging from 5 to 6.5. The results obtained also indicate that the five isolates can grow at moderate temperatures in the range of 27°C to 37°C. Interestingly, only strain OYA6 is capable to grow at 40°C and is also able to produce vinegar with 80 g/L of acetic acid; thus sharing similar features with the reference strain A. senegalensis LMG 23690T (Table 5). This thermotolerant strain of AAB was isolated from mango fruits in Senegal more than a decade and used for industrial vinegar production in developing countries, mainly in West Africa [11]. Taken together, the isolate OYA6 appeared as the best candidate that could be used for producing a cashew apple-based vinegar in the Ivorian industrial context. To make this objective effective, additional tests needs to be carried-out in future experiments, including the viability test of these indigenous Acetobacter isolates under different stress conditions as well as a fermentation test in an industrial controlled environment (e.g. bioreactor) as required [11,70].
Author Contributions
Conceptualization, Y.A.O. and K.R.F.; methodology, Y.A.O., I.A.C, D.M.A. and A.C.I.Y.; software, K.R.F and Y.A.O.; validation, K.R.F. and D.S.; formal analysis, Y.A.O.; investigation, D.S. and K.R.F.; resources, A.Z.; K.B.Y.; E.A. and D.S.; data curation, Y.A.O. and K.R.F; writing—original draft preparation, Y.A.O.; writing—review and editing, K.R.F.; visualization, A.Z., E.A. and K.B.Y.; supervision, K.R.F., A.Z., E.A. and K.B.Y.; project administration, D.S.; funding acquisition, D.S. and K.B.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the African Center of Excellence for Waste Recovery (CEA-VALOPRO) from INP-HB (Côte d’Ivoire), which is funded by the French Development Agency (AFD), grant number : CCI 167901 T. The APC was funded by the same agency.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within this article.
Acknowledgments
Authors would like to acknowledge the Cashew Value Chain Competitiveness Project, also known as PPCA (Projet de Promotion de la Compétitivité de la chaine de valeur de l’Anacarde - Côte d’Ivoire) for their valuable technical supports when we started this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chuvochina, M.; Mussig, A.J.; Chaumeil, P.-A.; Skarshewski, A.; Rinke, C.; Parks, D.H.; Hugenholtz, P. Proposal of Names for 329 Higher Rank Taxa Defined in the Genome Taxonomy Database under Two Prokaryotic Codes. FEMS Microbiol. Lett. 2023, 370, fnad071. [CrossRef]
- Kersters, K.; Lisdiyanti, P.; Komagata, K.; Swings, J. The Family Acetobacteraceae: The Genera Acetobacter, Acidomonas, Asaia, Gluconacetobacter, Gluconobacter, and Kozakia. In The Prokaryotes: Volume 5: Proteobacteria: Alpha and Beta Subclasses; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, 2006; pp. 163–200 ISBN 978-0-387-30745-9.
- Sengun, I.Y.; Karabiyikli, S. Importance of Acetic Acid Bacteria in Food Industry. Food. Control. 2011, 22, 647–656. [CrossRef]
- Shafiei, R.; Leprince, P.; Sombolestani, A.S.; Thonart, P.; Delvigne, F. Effect of Sequential Acclimation to Various Carbon Sources on the Proteome of Acetobacter Senegalensis LMG 23690T and Its Tolerance to Downstream Process Stresses. Front. Microbiol. 2019, 10, 608. [CrossRef]
- FAO-WHO Food Standards Programme. ROME. https://www.fao.org/fao-who-codexalimentarius/en/ (accessed on 14 February 2024).
- AFNOR NF EN 13188-Octobre 2000 : Vinaigre Produit Fabriqué à Partir de Liquides d’origine Agricole. Définitions, Prescriptions, Marquage. - Recherche Google Available online: https://www.boutique.afnor.org/fr-fr/norme/nf-en-13188/vinaigre-produit-fabrique-a-partir-de-liquides-dorigine-agricole-definition/fa036606/17808 (accessed on 7 March 2024).
- Joyeux, A.; Lafon-Lafourcade, S.; Ribéreau-Gayon, P. Evolution of Acetic Acid Bacteria During Fermentation and Storage of Wine. Appl. Environ. Microbiol. 1984, 48, 153–156.
- Ndoye, B.; Cleenwerck, I.; Engelbeen, K.; Dubois-Dauphin, R.; Guiro, A.T.; Van Trappen, S.; Willems, A.; Thonart, P. Acetobacter Senegalensis Sp. Nov., a Thermotolerant Acetic Acid Bacterium Isolated in Senegal (Sub-Saharan Africa) from Mango Fruit (Mangifera Indica L.). Int. J. Syst. Evol. Microbiol. 2007, 57, 1576–1581. [CrossRef]
- Konate, M.; Akpa, E.; Bernadette, G.; Koffi, L.; Honore, O.; Niamke, S. Banana Vinegars Production Using Thermotolerant Acetobacter Pasteurianus Isolated From Ivorian Palm Wine. J. Food Res. 2015, 4, p92. [CrossRef]
- Ndoye, B.; Lebecque, S.; Dubois-Dauphin, R.; Tounkara, L.; Guiro, A.-T.; Kere, C.; Diawara, B.; Thonart, P. Thermoresistant Properties of Acetic Acids Bacteria Isolated from Tropical Products of Sub-Saharan Africa and Destined to Industrial Vinegar. Enzyme Microb. Technol. 2006, 39, 916–923. [CrossRef]
- Ndoye, B.; Shafiei, R.; Sanaei, N.S.; Cleenwerck, I.; Somda, M.K.; Dicko, M.H.; Tounkara, L.S.; Guiro, A.T.; Delvigne, F.; Thonart, P. Acetobacter Senegalensis Isolated from Mango Fruits: Its Polyphasic Characterization and Adaptation to Protect against Stressors in the Industrial Production of Vinegar: A Review. J. Appl. Microbiol. 2022, 132, 4130–4149. [CrossRef]
- Cleenwerck, I.; Camu, N.; Engelbeen, K.; De Winter, T.; Vandemeulebroecke, K.; De Vos, P.; De Vuyst, L. Acetobacter Ghanensis Sp. Nov., a Novel Acetic Acid Bacterium Isolated from Traditional Heap Fermentations of Ghanaian Cocoa Beans. Int. J. Syst. Evol. Microbiol. 2007, 57, 1647–1652. [CrossRef]
- Ouattara, A.; Somda, M.; SINA, H.; Mousse, W.; Christine, N. tcha; C.A.T, O.; Traore, A.; Baba-Moussa, L.; Ouattara, A. Screening and Molecular Identification of Indigenous Strains of Acetic Acid Bacteria Isolated from Mango Biotopes in Burkina Faso. Afr. J. Food Sci. Tech. 2021, 12, 1–7. [CrossRef]
- Lee, D.-H.; Kim, S.-H.; Lee, C.-Y.; Jo, H.-W.; Lee, W.-H.; Kim, E.-H.; Choi, B.-K.; Huh, C.-K. Screening of Acetic Acid Bacteria Isolated from Various Sources for Use in Kombucha Production. Fermentation. 2023, 10, 18. [CrossRef]
- Vegas, C.; Mateo, E.; González, A.; Jara, C.; Guillamon, J.; Poblet, M.; Torija, M.; Mas, A. Population Dynamics of Acetic Acid Bacteria during Traditional Wine Vinegar Production. Int. J. Food Microbiol. 2010, 138, 130–136. [CrossRef]
- Sombolestani, A.S.; Cleenwerck, I.; Cnockaert, M.; Borremans, W.; Wieme, A.D.; De Vuyst, L.; Vandamme, P. Novel Acetic Acid Bacteria from Cider Fermentations: Acetobacter Conturbans Sp. Nov. and Acetobacter Fallax Sp. Nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 6163–6171. [CrossRef]
- Yamada, Y.; Hosono, R.; Lisdyanti, P.; Widyastuti, Y.; Saono, S.; Uchimura, T.; Komagata, K. Identification of Acetic Acid Bacteria Isolated from Indonesian Sources, Especially of Isolates Classified in the Genus Gluconobacter. J. Gen. Appl. Microbiol. 1999, 45, 23–28. [CrossRef]
- Komagata, K.; Iino, T.; Yamada, Y. The Family Acetobacteraceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin, Heidelberg, 2014; pp. 3–78 ISBN 978-3-642-30197-1.
- Katsura, K.; Kawasaki, H.; Potacharoen, W.; Saono, S.; Seki, T.; Yamada, Y.; Uchimura, T.; Komagata, K. Asaia Siamensis Sp. Nov., an Acetic Acid Bacterium in the Alpha-Proteobacteria. Int. J. Syst. Evol. Microbiol. 2001, 51, 559–563. [CrossRef]
- Suzuki, R.; Zhang, Y.; Iino, T.; Kosako, Y.; Komagata, K.; Uchimura, T. Asaia Astilbes Sp. Nov., Asaia Platycodi Sp. Nov., and Asaia Prunellae Sp. Nov., Novel Acetic Acid Bacteria Solated from Flowers in Japan. J. Gen. Appl. Microbiol. 2010, 56, 339–346. [CrossRef]
- Lino, T.; Suzuki, R.; Kosako, Y.; Ohkuma, M.; Komagata, K.; Uchimura, T. Acetobacter Okinawensis Sp. Nov., Acetobacter Papayae Sp. Nov., and Acetobacter Persicus Sp. Nov.; Novel Acetic Acid Bacteria Isolated from Stems of Sugarcane, Fruits, and a Flower in Japan. J. Gen. Appl. Microbiol. 2012, 58, 235–243. [CrossRef]
- Aluko, A.; Makule, E.; Kassim, N. Effect of Clarification on Physicochemical Properties and Nutrient Retention of Pressed and Blended Cashew Apple Juice. Food Sci. Nutr. 2023, 11. [CrossRef]
- Doudjo, S.; Dornier, M.; Abreu, F.; Assidjo, E.; Yao, B.; Reynes, R. The Cashew ( Anacardium Occidentale ) Industry in Côte d’Ivoire: Analysis and Prospects for Development. Fruits. 2011, 66, 237–245. [CrossRef]
- Gnagne, A.; Soro, D.; Ouattara, Y.; Koui, E.; Koffi, E. A Literature Review of Cashew Apple Processing. Afr. J. Food Agric. Nutr. Dev. 2023, 23, 22452–22469. [CrossRef]
- Yaya Anianhou, O.; Soro, D.; Kakou, E.; Koné, K.; Assidjo, E.; Mabia, G.; Kamagate, M. Synthesis and Characterisation of Saccharomyces Cerevisiae Immobilised Cells from Cashew Apple Bagasse. Revue des Bioressources. 2023, 18, 3736–3749. [CrossRef]
- Guero, M.; Drion, B.; Karsch, P. Study of the Biomass Potential in Côte d’Ivoire 2021.
- Usharani, G.; Rani, Mrs.G.U. Isolation and Identification of an Acetobacter Isolate from Cashew Apple as a Potential Strain for Vinegar Production. Int. J. Recent Sci. Res. 2011, 3, 080–084.
- Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; 1st ed.; Wiley, 2015; ISBN 978-1-118-96060-8.
- Ruiz, A.; Poblet, M.; Mas, A.; Guillamon, J. Identification of Acetic Acid Bacteria by RFLP of PCR-Amplified 16S rDNA and 16S-23S rDNA Intergenic Spacer. Int. J. Syst. Evol. Microbiol. 2000, 50, 1981–1987. [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 1991; pp. 283–293 ISBN 978-3-642-83964-1.
- Gnangui, S.; Fossou, R.; Ebou, A.; Amon, R.E.; Koua, D.; Kouadjo, G.; Cowan, D.; Zézé, A. The Rhizobial Microbiome from the Tropical Savannah Zones in Northern Côte d’Ivoire. Microorganisms. 2021, 9, 1842. [CrossRef]
- Kumar, G.; Arya, P.; Gupta, K.; Randhawa, V.; Acharya, V.; Singh, A. Comparative Phylogenetic Analysis and Transcriptional Profiling of MADS-Box Gene Family Identified DAM and FLC-like Genes in Apple (Malus x Domestica). Sci. Rep. 2016, 6, 20695. [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 24, 189–204. [CrossRef]
- Rogers, J.; Swofford, D. A Fast Method for Approximating Maximum Likelihoods of Phylogenetic Trees from Nucleotide Sequences. Syst. Biol. 1998, 47, 77–89. [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evol. 1985, 39, 783. [CrossRef]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464.
- Rashid, H.; Young, J.P.; Everall, I.; Clercx, P.; Willems, A.; Braun, M.; Wink, M. Average Nucleotide Identity of Genome Sequences Supports the Description of Rhizobium Lentis Sp. Nov., Rhizobium Bangladeshense Sp. Nov. and Rhizobium Binae Sp. Nov. from Lentil (Lens Culinaris) Nodules. Int. J. Syst. Evol. Microbiol. 2015, 65. [CrossRef]
- Fossou, R.; Ziegler, D.; Zézé, A.; Barja, F.; Perret, X. Two Major Clades of Bradyrhizobia Dominate Symbiotic Interactions with Pigeonpea in Fields of Côte d’Ivoire. Front. microbiol. 2016, 7. [CrossRef]
- Cleenwerck, I.; Vandemeulebroecke, K.; Janssens, D.; Swings, J. Re-Examination of the Genus Acetobacter, with Descriptions of Acetobacter Cerevisiae Sp. Nov. and Acetobacter Malorum Sp. Nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1551–1558. [CrossRef]
- Lisdiyanti, P.; Kawasaki, H.; Seki, T.; Yamada, Y.; Uchimura, T.; Komagata, K. Identification of Acetobacter Strains Isolated from Indonesian Sources, and Proposals of Acetobacter Syzygii Sp. Nov., Acetobacter Cibinongensis Sp. Nov., and Acetobacter Orientalis Sp. Nov. J. Gen. Appl. Microbiol. 2001, 47, 119–131. [CrossRef]
- Lisdiyanti, P.; Kawasaki, H.; Seki, T.; Yamada, Y.; Uchimura, T.; Komagata, K. Systematic Study of the Genus Acetobacter with Descriptions of Acetobacter Indonesiensis Sp. Nov., Acetobacter Tropicalis Sp. Nov., Acetobacter Orleanensis (Henneberg 1906) Comb. Nov., Acetobacter Lovaniensis (Frateur 1950) Comb. Nov., and Acetobacter Estunensis (Carr 1958) Comb. Nov. J. Gen. Appl. Microbiol. 2000, 46, 147–165. [CrossRef]
- Spitaels, F.; Li, L.; Wieme, A.; Balzarini, T.; Cleenwerck, I.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. Acetobacter Lambici Sp. Nov., Isolated from Fermenting Lambic Beer. Int. J. Syst. Evol. Microbiol. 2014, 64, 1083–1089. [CrossRef]
- Dutta, D.; Gachhui, R. Novel Nitrogen-Fixing Acetobacter Nitrogenifigens Sp. Nov., Isolated from Kombucha Tea. Int. J. Syst. Evol. Microbiol. 2006, 56, 1899–1903. [CrossRef]
- Vu, H.T.L.; Yukphan, P.; Bui, V.T.T.; Charoenyingcharoen, P.; Malimas, S.; Nguyen, L.K.; Muramatsu, Y.; Tanaka, N.; Tanasupawat, S.; Le, B.T.; et al. Acetobacter Sacchari Sp. Nov., for a Plant Growth-Promoting Acetic Acid Bacterium Isolated in Vietnam. Ann. Microbiol. 2019, 69, 1155–1163. [CrossRef]
- Pitiwittayakul, N.; Yukphan, P.; Chaipitakchonlatarn, W.; Yamada, Y.; Theeragool, G. Acetobacter Thailandicus Sp. Nov., for a Strain Isolated in Thailand. Ann. Microbiol. 2015, 65, 1855–1863. [CrossRef]
- Wang, B.; Rutherfurd-Markwick, K.; Zhang, X.-X.; Mutukumira, A.N. Isolation and Characterisation of Dominant Acetic Acid Bacteria and Yeast Isolated from Kombucha Samples at Point of Sale in New Zealand. Curr. Res. Nutr. Food Sci. 2022, 5, 835–844. [CrossRef]
- Mounir, M.; Shafiei, R.; Zarmehrkhorshid, R.; Hamouda, A.; Ismaili Alaoui, M.; Thonart, P. Simultaneous Production of Acetic and Gluconic Acids by a Thermotolerant Acetobacter Strain during Acetous Fermentation in a Bioreactor. J. Biosci. Bioeng. 2016, 121, 166–171. [CrossRef]
- N’Guessan, Y.D.; Akpa, E.E.; Samaci, lamine; Ouattara, H.; Akoa, E.E.; Ahonzo-Niamké, S.L. Vinegar Production Trial from Cashew Apple (Anacardium Occidentale) Using Thermotolerant Acetobacter Strains with High Acetic Acid Yield in Non-Optimized Small Scale Conditions. Microbiol. Nat. 2020, 1, 107–119. [CrossRef]
- Sharafi, S.; Rasooli, I.; Beheshti-Maal, K. Isolation, Characterization and Optimization of Indigenous Acetic Acid Bacteria and Evaluation of Their Preservation Methods. Iran. J. Microbiol. 2010, 2, 38–45.
- Cire Kourouma, M.; Mbengue, M.; Sarr, K.; Toure Kane, C. Isolement, identification et caractérisation de souches de bactéries acétiques a partir d’un alcool de mangue fermente. Int. J. Adv. Res. 2021, 9, 271–281. [CrossRef]
- Heo, J.; Sombolestani, A.S.; Laureys, D.; De Clippeleer, J.; Won, M.; Vandamme, P.; Kwon, S.-W. Acetobacter Vaccinii Sp. Nov., a Novel Acetic Acid Bacterium Isolated from Blueberry Fruit (Vaccinium Corymbosum L.). Int. J. Syst. Evol. Microbiol. 2022, 72. [CrossRef]
- Huang, C.-H.; Chang, M.-T.; Huang, L.; Chu, W.-S. Molecular Discrimination and Identification of Acetobacter Genus Based on the Partial Heat Shock Protein 60 Gene (Hsp60) Sequences. J. Sci. Food Agric. 2014, 94. [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [CrossRef]
- Kwon, S.-W.; Park, J.-Y.; Kim, J.-S.; Kang, J.W.; Cho, Y.-H.; Lim, C.-K.; Parker, M.; Lee, G.-B. Phylogenetic Analysis of the Genera Bradyrhizobium, Mesorhizobium, Rhizobium and Sinorhizobium on the Basis of 16S rRNA Gene and Internally Transcribed Spacer Region Sequences. Int. J. Syst. Evol. Microbiol. 2005, 55, 263–270. [CrossRef]
- Tesfaye, W.; Morales, M.L.; Garcı́a-Parrilla, M.C.; Troncoso, A.M. Wine Vinegar: Technology, Authenticity and Quality Evaluation. Trends. Food. Sci. Tech. 2002, 1, 12–21. [CrossRef]
- Chen, Y.; Bai, Y.; Li, D.; Wang, C.; Xu, N.; Hu, Y. Improvement of the Flavor and Quality of Watermelon Vinegar by High Ethanol Fermentation Using Ethanol-Tolerant Acetic Acid Bacteria. Int. J. Food Eng. 2017, 13. [CrossRef]
- Drysdale, G.S.; Fleet, G.H. Acetic Acid Bacteria in Winemaking: A Review. Am. J. Enol. Vitic. 1988, 39, 143–154. [CrossRef]
- Kyu-Ho, B.; Chae-Won, K.; Chul-Ho, K. Isolation of an Acetic Acid Bacterium Acetobacter Pasteurianus CK-1 and Its Fermentation Characteristics. J. Life. Sci. 2022, 32. [CrossRef]
- Mamlouk, D.; Gullo, M. Acetic Acid Bacteria: Physiology and Carbon Sources Oxidation. Indian J. Microbiol. 2013, 53, 377–384. [CrossRef]
- Soumahoro, S.; Goualie, B.G.; Adom, J.N.; Ouattara, H.G.; Koua, G.; Doue, G.G.; Niamke, S.L. Effects of Culture Conditions on Acetic Acid Production by Bacteria Isolated from Ivoirian Fermenting Cocoa (Theobroma Cacao L.) Beans. J. App. Bioscience. 2016, 95, 8981. [CrossRef]
- El-Askri, T.; Yatim, M.; Sehli, Y.; Rahou, A.; Belhaj, A.; Castro-Mejías, R.; Durán-Guerrero, E.; Hafidi, M.; Rachid, Z. Screening and Characterization of New Acetobacter Fabarum and Acetobacter Pasteurianus Strains with High Ethanol–Thermo Tolerance and the Optimization of Acetic Acid Production. Microorganisms. 2022, 10, 1741. [CrossRef]
- Lisdiyanti, P.; Kawasaki, H.; Seki, T.; Yamada, Y.; Uchimura, T.; Komagata, K. Systematic Study of the Genus Acetobacter with Descriptions of Acetobacter Indonesiensis Sp. Nov., Acetobacter Tropicalis Sp. Nov., Acetobacter Orleanensis (Henneberg 1906) Comb. Nov., Acetobacter Lovaniensis (Frateur 1950) Comb. Nov., and Acetobacter Estunensis (Carr 1958) Comb. Nov. J. Gen. Appl. Microbiol. 2000, 46, 147–165. [CrossRef]
- Gullo, M.; Giudici, P. Acetic Acid Bacteria in Traditional Balsamic Vinegar: Phenotypic Traits Relevant for Starter Cultures Selection. Int. J. Food Microbiol. 2008, 125, 46–53. [CrossRef]
- Wei, K.; Cao, X.; Li, X.; Wang, C.; Hou, L. Genome Shuffling to Improve Fermentation Properties of Acetic Acid Bacterium by the Improvement of Ethanol Tolerance. Int. J. of Food. Sci. Tech. 2012, 47, 2184–2189. [CrossRef]
- Raspor, P.; Goranovic, D. Biotechnological Applications of Acetic Acid Bacteria. Crit. Rev. Biotechnol. 2008, 28, 101–124. [CrossRef]
- Cleenwerck, I.; De Vos, P. Polyphasic Taxonomy of Acetic Acid Bacteria: An Overview of the Currently Applied Methodology. Int. J. Syst. Evol. Microbiol. 2008, 125, 2–14. [CrossRef]
- Hussenet, C. Instrumentation, modélisation et automatisation de fermenteurs levuriers à destination oenologique. Génie des procédés, L’UNIVERSITE PARIS-SACLAY: FRANCE, 2017.
- Acourene, S.; Ammouche, A.; Djaafri, K. Valorisation des rebuts de dattes par la production de la levure boulangère, de l’alcool et du vinaigre. Sciences & Technologie. C, Biotechnologies 2008, 38–45.
- Saeki, A.; TaniguchiI, M.; Matsushita, K.; Toyama, H.; Theeragool, G.; Lotong, N.; Adachi, O. Microbiological Aspects of Acetate Oxidation by Acetic Acid Bacteria, Unfavorable Phenomena in Vinegar Fermentation. Biosci. Biotechnol. Biochem. 1997, 61, 317–323. [CrossRef]
- Shafiei, R.; Delvigne, F.; Thonart, P. Flow-Cytometric Assessment of Damages to Acetobacter Senegalensis during Freeze-Drying Process and Storage. Acetic Acid Bacteria 2013, 2, 10. [CrossRef]
Figure 2.
Acetic fermentation process.
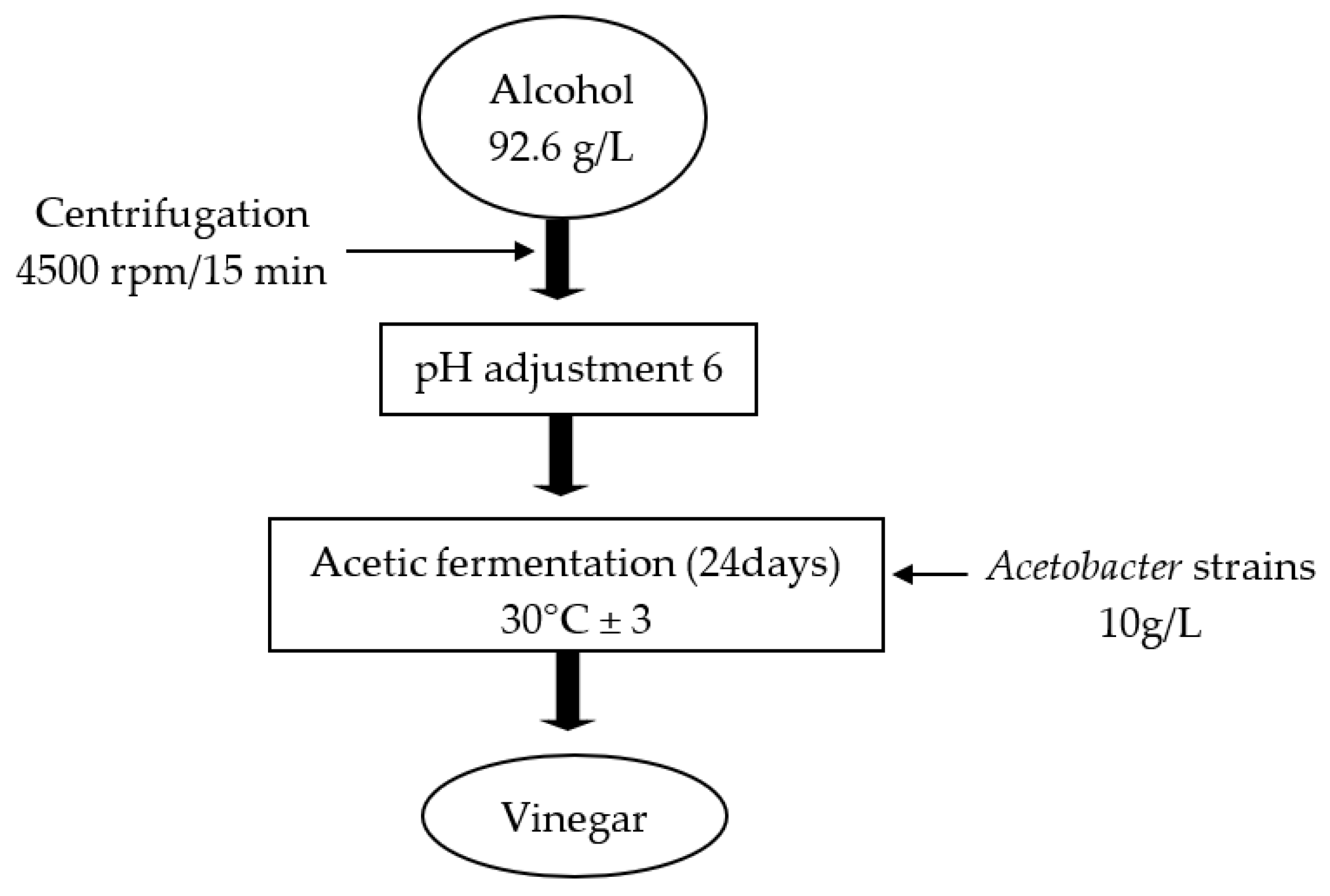
Figure 3.
Appearance of bacteria isolated with regular (A) and irregular (B) shape.
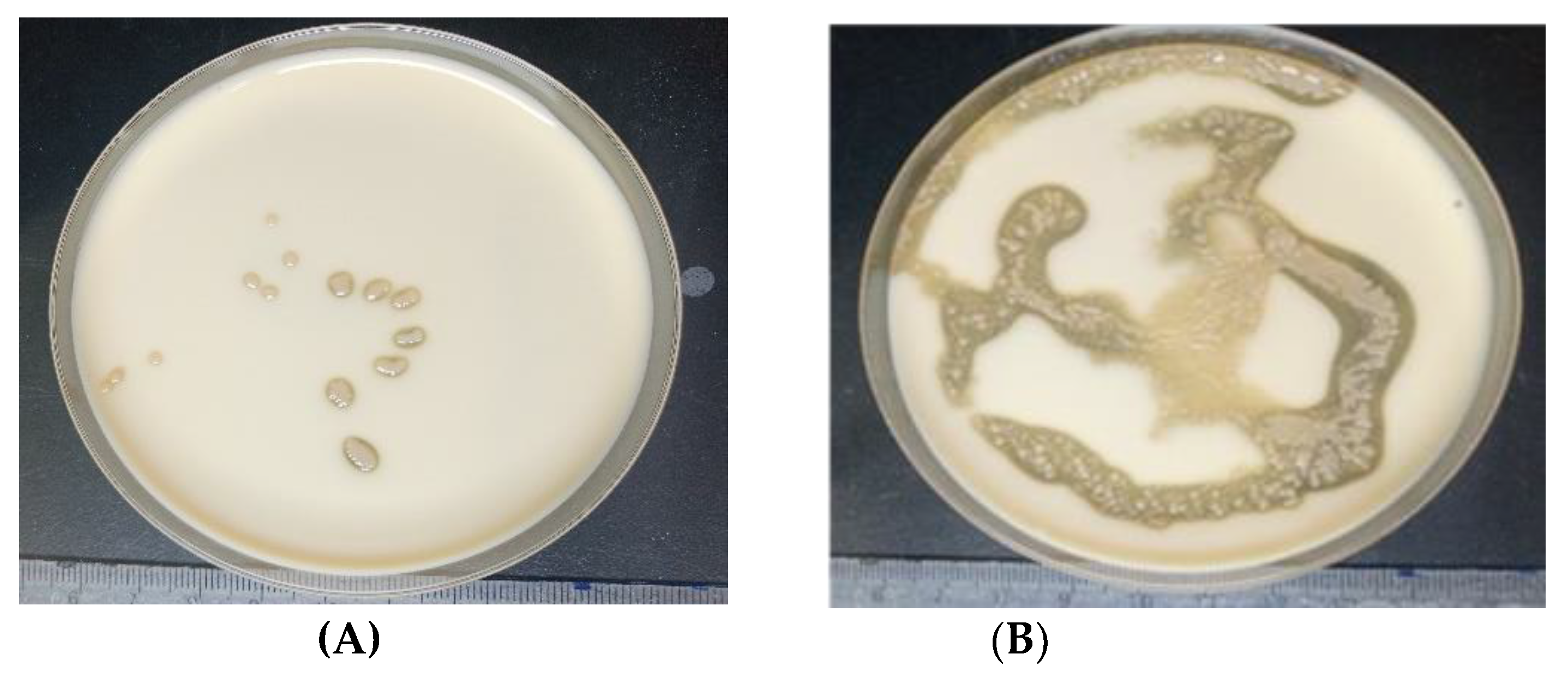
Figure 4.
Gram-negative bacilli observed after gram staining.
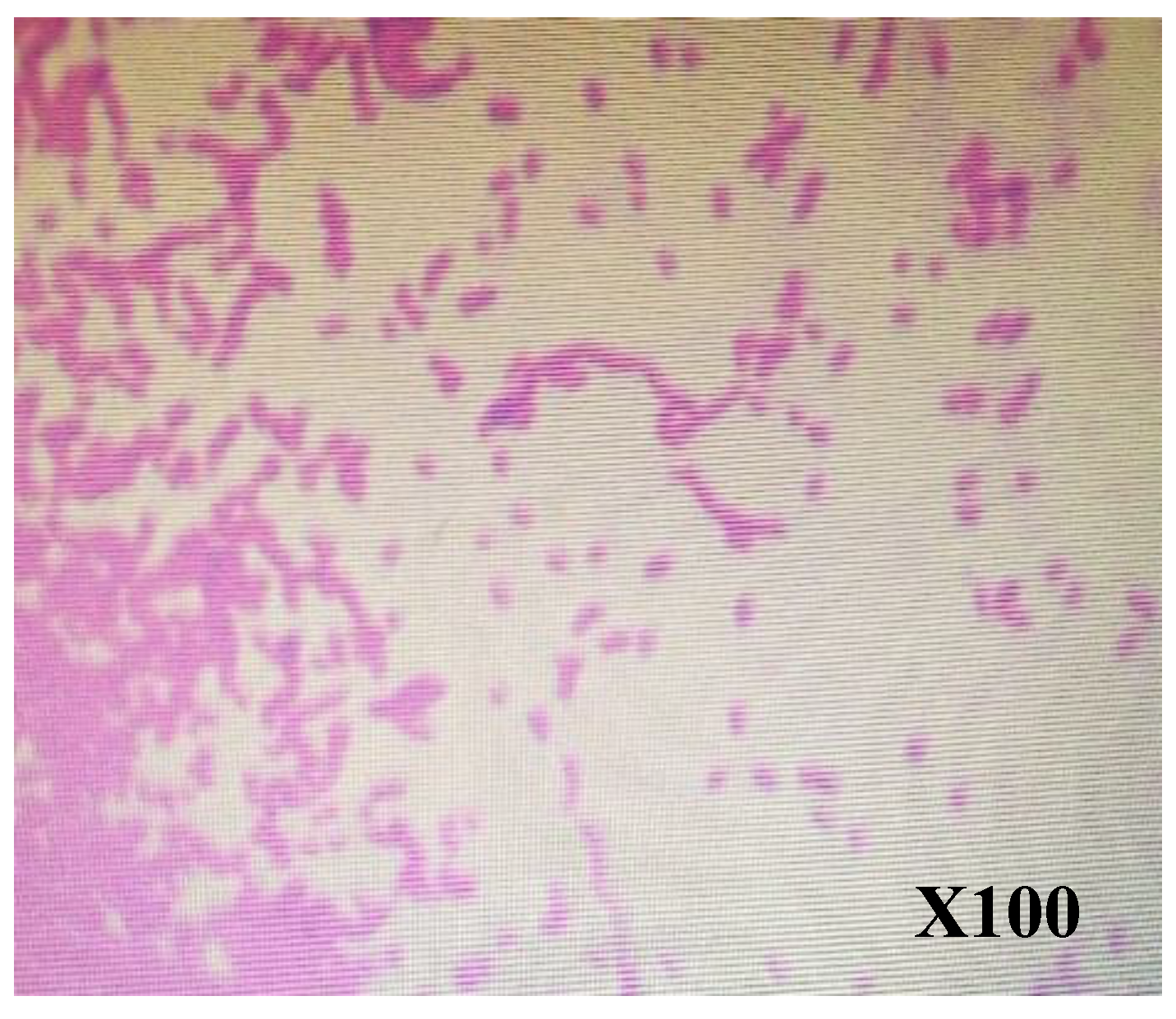
Figure 5.
Maximum Likelihood (ML) phylogenetic tree based on 16 S rRNA gene sequences of Acetobacter strains isolated from Cash apple in Côte d’Ivoire and related species of the genus. All the sequences were aligned with MUSCLE and analysis used the T92 +G model, a total of 967 positions, 1000 replicates with bootstrap values higher than 70% shown at branch nodes. The sequence accession numbers are shown within parentheses next to the species name. Scale bar indicates the number of substitutions per site. Asaia bogorensis NBRC 16594T was used as an outgroup.
Figure 5.
Maximum Likelihood (ML) phylogenetic tree based on 16 S rRNA gene sequences of Acetobacter strains isolated from Cash apple in Côte d’Ivoire and related species of the genus. All the sequences were aligned with MUSCLE and analysis used the T92 +G model, a total of 967 positions, 1000 replicates with bootstrap values higher than 70% shown at branch nodes. The sequence accession numbers are shown within parentheses next to the species name. Scale bar indicates the number of substitutions per site. Asaia bogorensis NBRC 16594T was used as an outgroup.
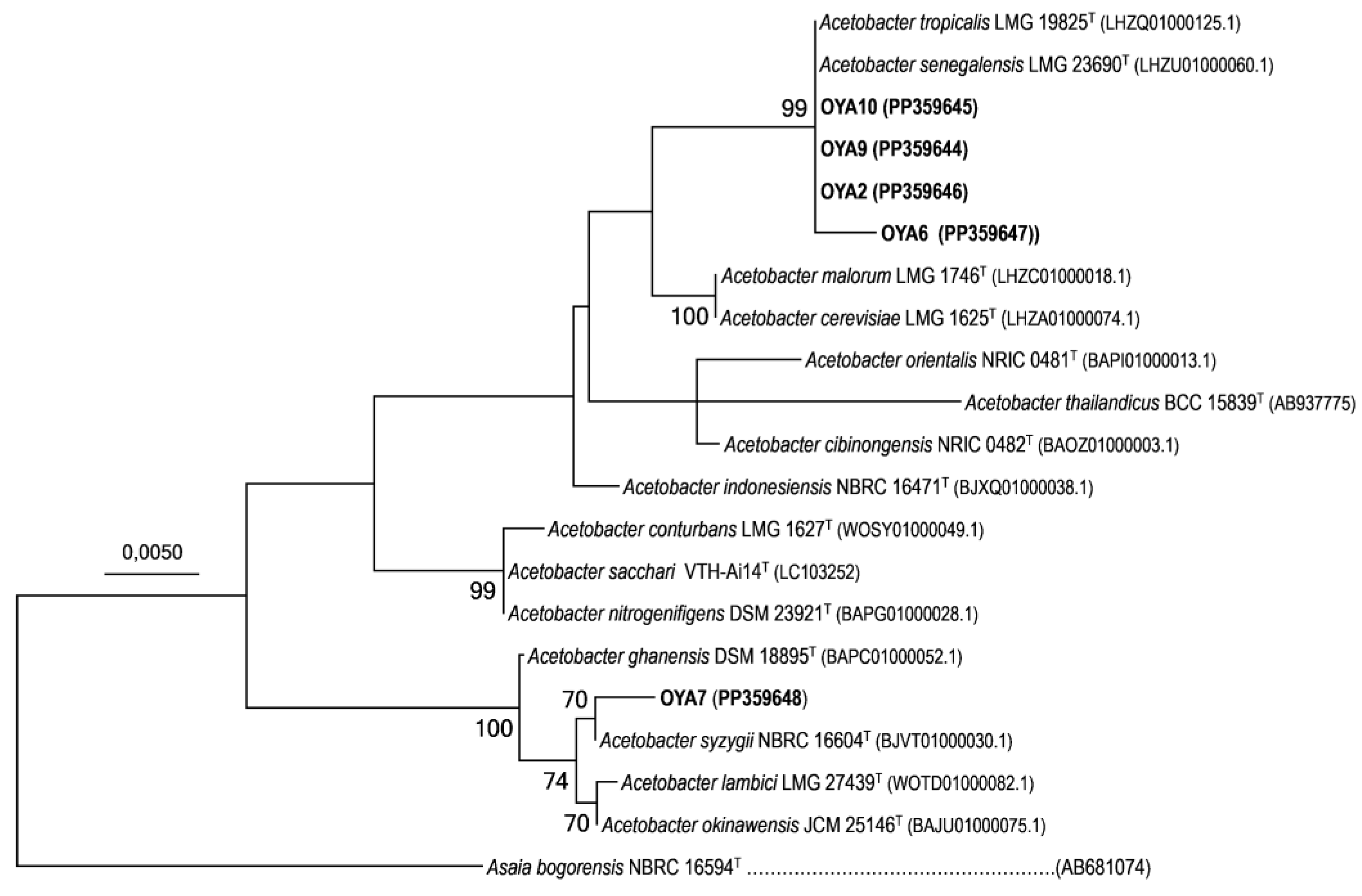
Figure 6.
Effect of temperature on isolates growth.
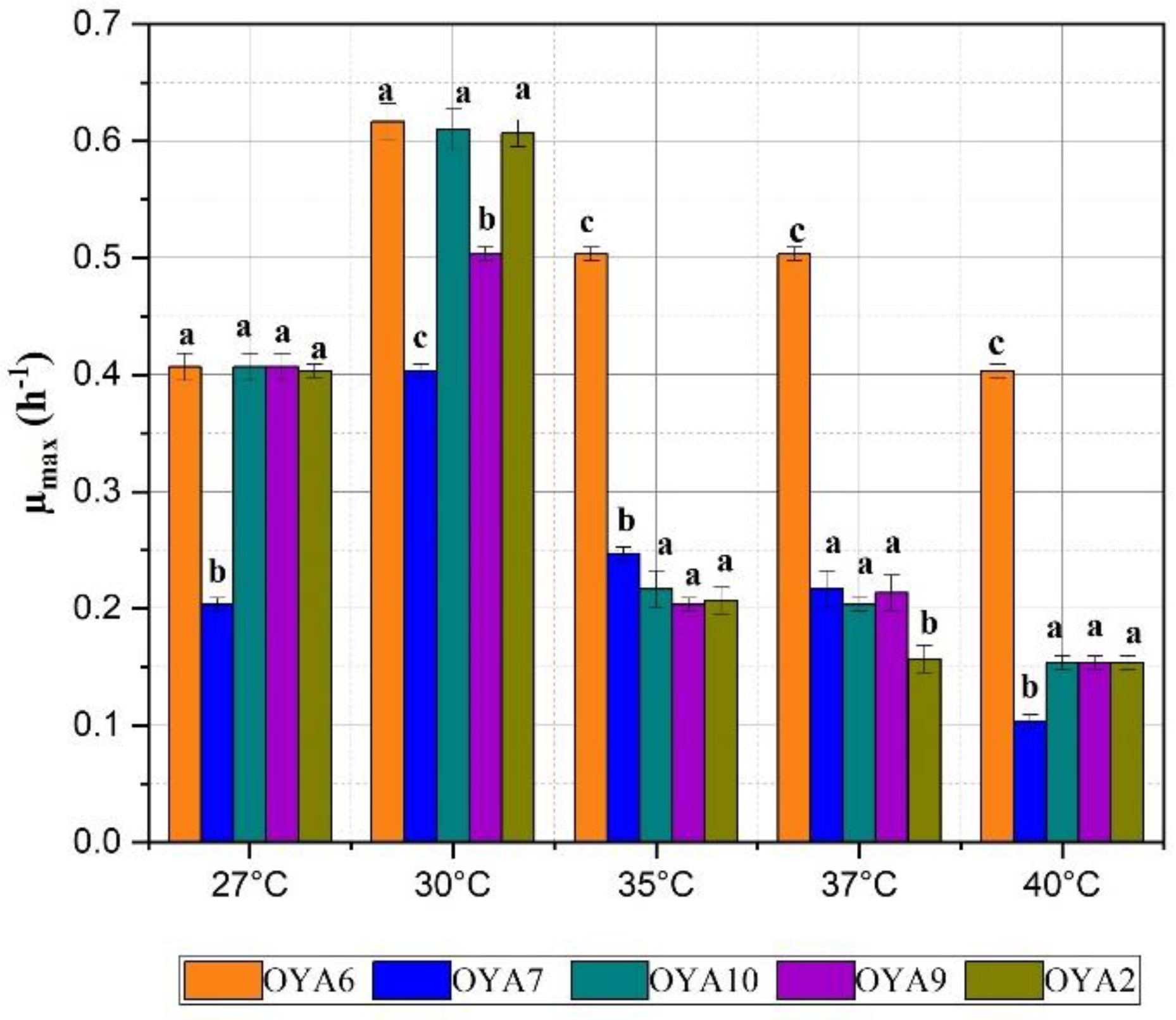
Figure 7.
Effect of pH on isolates growth.
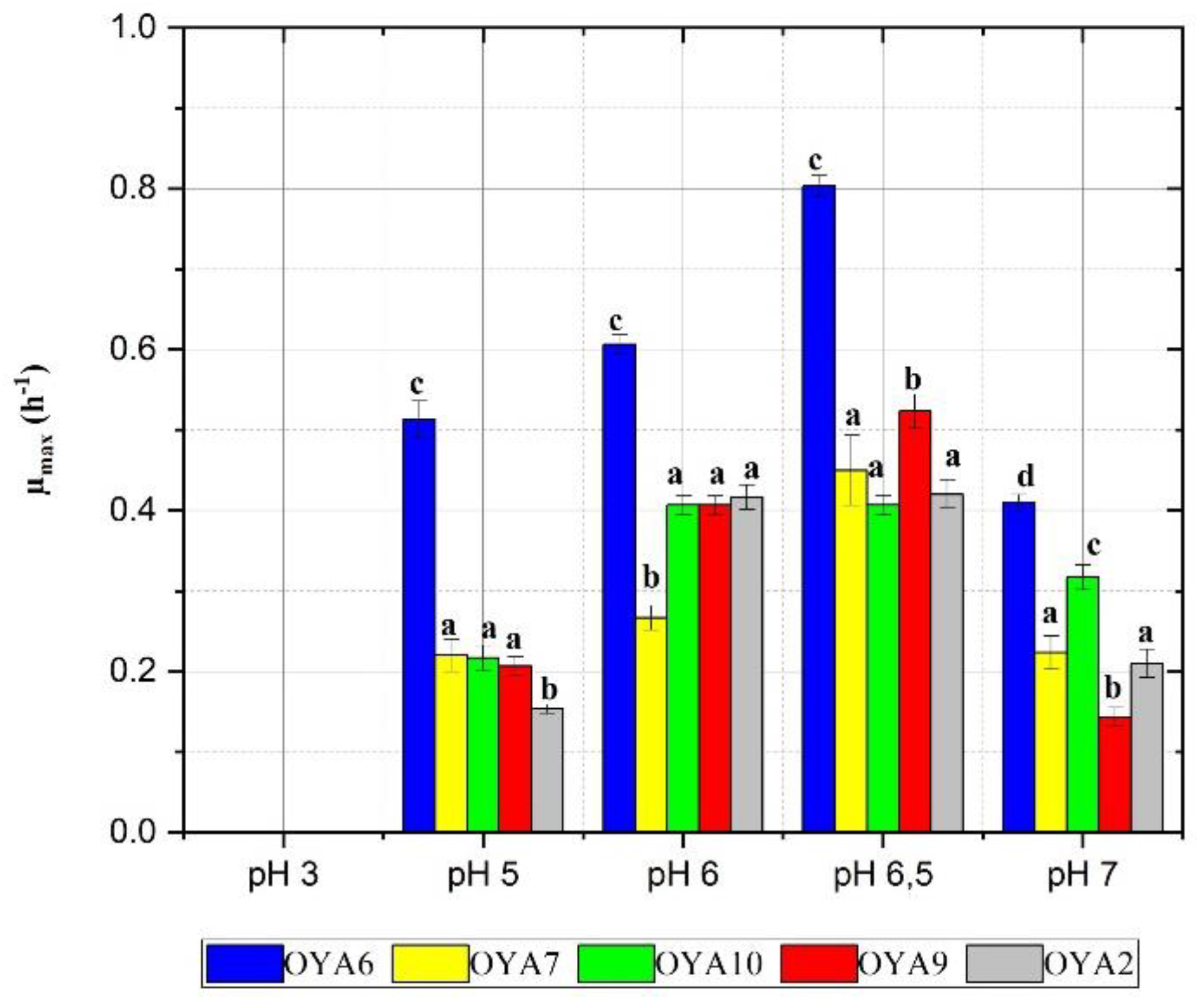
Figure 8.
Effect of ethanol on isolates growth.
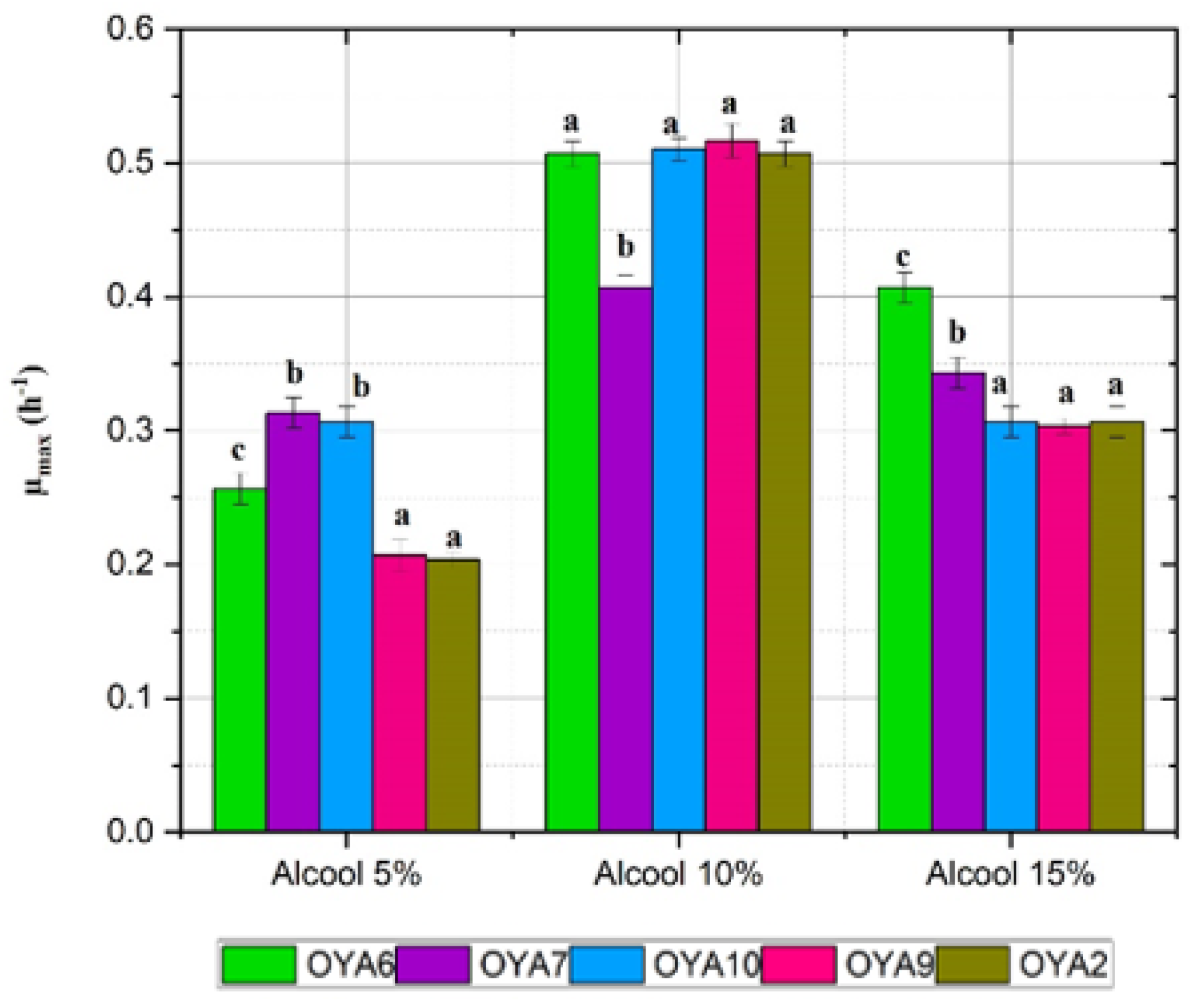

Table 1.
PCR reaction medium composition.
| Concentration | Sample Volume | |
|---|---|---|
| 2X oneTaq standard buffer | 2 X | 25 µl |
| Amorce 1 (avant) | 10 µM | 5 µl |
| Amorce 2 (inverse) | 10 µM | 5 µl |
| ADN | Ca. 10 ng/µl | 3 µl |
| Distilled sterile water | - | 12 µl |
| Total reaction volume for each amplification | 50 µl | |
Table 2.
List of the 15 reference strains of Acetobacter and corresponding GenBank accessions used in this study. Their genome sequences were used as data for species of the Acetobacter found to be genetically related to the acetic acid bacteria isolated from the Ivorian cashew apples. They were mainly used for the phylogeny reconstruction and for the genetic similarity value calculations.
Table 2.
List of the 15 reference strains of Acetobacter and corresponding GenBank accessions used in this study. Their genome sequences were used as data for species of the Acetobacter found to be genetically related to the acetic acid bacteria isolated from the Ivorian cashew apples. They were mainly used for the phylogeny reconstruction and for the genetic similarity value calculations.
| Reference Type Strain | Isolation Source & Country | 16S Data1 | Genome Data | Reference | |
|---|---|---|---|---|---|
| 1 | A. cerevisiae LMG 1625T | Beer (ale) in storage; Canada |
AJ419843 | LHZA01000074 | Cleenwerck et al.[39] |
| 2 | A. cibinongensis NRIC 0482T | montana (fruit); Indonesia |
AB681085 | BAOZ01000003 | Lisdiyanti et al.[40] |
| 3 | A. conturbans LMG 1627T | Laboratory-scale cider fermentations, UK | MN744433 | WOSY01000049 | Sombolestani et al. [16] |
| 4 | A. ghanensis 18895T | Traditional heap fermentations of cocoa beans; Ghana | EF030713 | BAPC01000052 | Cleenwerck et al. [12] |
| 5 | A. indonesiensis 16471T | Fruit of Annona muricata; Indonesia | AB032356 | BJXQ01000038 | Lisdiyant et al.[41] |
| 6 | A. lambici LMG 27439T | fermenting lambic beer ; Belgium | HF969863 | WOTD01000082 | Spitaels et al.[42] |
| 7 | A. malorum LMG 1746T | Rotting apple ; Belgium | AJ419844 | LHZC01000018 | Cleenwerck et al.[39] |
| 8 | A. nitrogenifigens DSM 23921T | Kombucha tea ; India | AY669513 | BAPG01000028 | Dutta & Gachhui.[43] |
| 9 | A. okinawensis JCM 25146T | Sugarcane (stem); Japon | AB665068 | BAJU01000075 | Lino et al.[21] |
| 10 | A. orientalis NRIC 0481T | Canna (flower) ; Indonesia |
AB681086 | BAPI01000013 | Lisdiyanti et al.[40] |
| 11 | A. sacchari VTH-Ai14T | stems of sugarcane; Thailand |
LC103252 | LC103252 | Vu et al.[44] |
| 12 | A. senegalensis LMG 23690T | mango fruit ; Senegal | AY883036 | LHZU01000060 | Ndoye et al.[8] |
| 13 | A. syzygii NBRC 16604T | Malay rose apple (fruit); Indonesia | AB681084 | BJVT01000030 | Lisdiyanti et al.[40] |
| 14 | A. thailandicus BCC 15839T | flower of the blue trumpet vine; Thailand | AB937775 | Not available | Pitiwittayakul et al.[45] |
| 15 | A. tropicalis LMG 19825T | Coconut juice; Indonesia | AB032354 | LHZQ01000125 | Lisdiyanti et al.[40] |
1 For the sequences analyses, we preferentially used the 16S sequence retrieved from the whole-genome shotgun contigs (wgs) of each strain (when possible), for quality concerns. Some type strains include more than one ambiguous positions (N, W…) per 100 bp count in their 16S rDNA data archived in GenBank, under Nucleotide collection (nr/nt) and appeared inapropriate for the analysis. For example, the 16S (nr/nt) sequence of A. senegalensis LMG 23690T (AY883036), which is underlined in Table 2, diverged too much (> 2%; ca. 30 divergent nucleotides, including 5 gaps) from the 16S sequences retrieved from the genome of the same type strain obtained from two independent sources (cf. GenBank 16S locus_tag LHZU01000060 & 16S locus_tag BAPN01000029).
Table 3.
Biochemical and cultural characteristics of isolated strains.
| Test | Morphology | Color | Motility | Gram Staining | Catalase | Oxidase |
|---|---|---|---|---|---|---|
| OYA10 | Irregular | Brown | + | - | + | - |
| OYA2 | Irregular | Brown | + | - | + | - |
| OYA9 | regular | Brown | + | - | + | - |
| OYA6 | Irregular | Brown | + | - | + | - |
| OYA7 | Irregular | Brown | + | - | + | - |
Table 4.
Acetic fermentation parameters with isolated acetic acid bacteria strains.
| Strain of Acetic Acid Bacteria | Fermentation Time (h) |
Initial Alcohol (g/L) |
Final Alcohol (g/L) |
Acetic Acid Produced (g/L) |
Qp (g/L/h) |
Yp⁄s (g/g) |
Conversion (%) |
η (%) |
|---|---|---|---|---|---|---|---|---|
| OYA6 | 576 | 92.6 ± 3.43 a | 5.18 ± 0.13 a | 80.67 ± 2.1 c | 0.14 | 0.92 | 94.41 | 67.85 |
| OYA7 | 576 | 92.6 ± 3.43 a | 10.34 ± 0.26 b | 70.26 ± 0.9 a | 0.12 | 0.85 | 88.83 | 62.8 |
| OYA10 | 576 | 92.6 ± 3.43 a | 10.49 ± 0.55 b | 68.7 ± 1.51 ab | 0.12 | 0.84 | 88.67 | 61.52 |
| OYA9 | 576 | 92.6 ± 3.43 a | 10.61 ± 1.69 b | 67.22 ± 0.37 b | 0.12 | 0.82 | 88.54 | 60.28 |
| OYA2 | 576 | 92.6 ± 3.43 a | 11.2 ± 1.15 b | 70.11 ± 1.69 a | 0.12 | 0.86 | 88.54 | 62.88 |
All values are mean ± SD (n = 3); Means with different superscript letters in the same row are significantly different at p < 0.05 by Duncan’s multiple range test. a > b > c >d.
Table 5.
Main futures of the 05 Acetobacter strains isolated from cashew apple in Yamoussoukro, Côte d’Ivoire, together with the reference strain A. senegalensis LMG 23690T.
Table 5.
Main futures of the 05 Acetobacter strains isolated from cashew apple in Yamoussoukro, Côte d’Ivoire, together with the reference strain A. senegalensis LMG 23690T.
| Isolate / Strain | 16S rDNA-Based Identification | 16S Genbank Accession | Stress at 40°C (Temperature) | Stress at 10° & 15° (v/v) of Alcohol | Acetic Acid Produced (g/L) | Yp⁄s (g/g) |
|
|---|---|---|---|---|---|---|---|
| 1 | OYA2 | A. tropicalis & A. senegalensis (100%) | PP359646 | doesn't grow | resistant | 70.11 ± 1.69 | 0.86 |
| 2 | OYA6 | A. tropicalis (99.7%) | PP359647 | still grow | resistant | 80.67 ± 2.1 | 0.92 |
| 3 | OYA7 | A. syzygii (99.7%) | PP359648 | doesn't grow | resistant | 70.26 ± 0.9 | 0.85 |
| 4 | OYA9 | A. tropicalis & A. senegalensis (100%) | PP359644 | doesn't grow | resistant | 67.22 ± 0.37 | 0.82 |
| 5 | OYA10 | A. tropicalis & A. senegalensis (100%) | PP359645 | doesn't grow | resistant | 68.7 ± 1.51 | 0.84 |
| 6 | CWBI-B4181 (=LMG 23690T =DSM 18889T) | A. senegalensis | AY883036 | still grow | resistant | - | Up to 0.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



