Submitted:
16 September 2024
Posted:
17 September 2024
You are already at the latest version
Abstract
Microalgae and their bioproducts hold significant potential across various applications, for instance, in wastewater remediation, CO2 fixation, and the synthesis of nutraceuticals, pharmaceuticals, and biofuels. The production of these organisms heavily relies upon environmental conditions, including factors like temperature, light, and nutrient supply, which can significantly impact growth. Furthermore, the limitations in nutrients or certain extreme environments can result in productivity increases. As such, microalgae growth systems have become a critical consideration for both research and industry. Both open and closed systems, such as raceway ponds and photobioreactors, respectively, have their own advantages and drawbacks in microalgae cultivation. However, for microalgae growth, photobioreactors may address most concerns, as the system has a lower risk of contamination and can deliver specific growth factors in a controlled manner. This review will summarize the growth and incubation factors of microalgae and the uses of bioreactors in both research and industrial capacities.
Keywords:
bioreactors
; photobioreactors
; microalgae
; cultivation factors
; incubation factors
1. Introduction
Microalgae are unicellular, photosynthetic organisms with a variety of potential applications. For instance, they can serve as a renewable energy source and are efficient producers of compounds commonly utilized in a variety of fields, such as pharmaceuticals and nutraceuticals, as depicted in Figure 1 [1,2,3,4]. Which has made microalgae highly sought after in research and industrial capacities, and to improve sustainability.
One such function of microalgae is in environmental betterment. Microalgae can be utilized to remediate wastewater while recovering useful products such as pigments and biogas [5]. This methodology, which utilizes wastewater in place of a traditional growth medium for the recovery process, encourages a circular bioeconomy by taking urban and industrial food wastewater and cultivating microalgae with the ability to reclaim useful products [5]. Furthermore, due to the organisms’ photosynthetic status, their consumption of carbon dioxide can be utilized for CO2 sequestering and simultaneous production of biomass [6]. The previously mentioned biomass productivity can include the synthesis of many appealing compounds.
Microalgae productivity encompasses the creation of lipids like polyunsaturated fatty acids (PUFAs) that can be used in the synthesis of biofuel or nutraceuticals [2,7]. For instance, Chlorella vulgaris can be the source of monounsaturated and saturated fatty acids that contribute to its potential in nutraceutical production [8]. Microalgae can also produce pigments like carotenoids, including compounds such as β-carotene, zeaxanthin, and astaxanthin, with high antioxidant capabilities [9,10,11]. As such, microalgae-based nutraceuticals have been synthesized to make use of microalgae extract’s healthy properties. For instance, compounds extracted from C. vulgaris exhibit a variety of health benefits, namely anti-obesity activity [2]. Furthermore, these bioproducts can be included in animal feed to improve the health of livestock. This has revealed it is possible to enhance the growth of domesticated chicken flocks under heat stress through the introduction of antioxidant-rich microalgae in their diet [12]. The cultivation of these photosynthetic organisms, thus, becomes a point of focus in obtaining these desirable microalgae bioproducts.
There are a variety of factors that impact microalgae growth and, occasionally, productivity. These include conditions such as light availability, temperature, pH, the rates of CO2 in air flow, nitrogen supply, and carbon source [13,14,15,16]. Under a 12-hour dark and 12-hour light period, the biomass and growth rate of Nannochloropsis salina is less compared to cultures grown under constant light [13]. However, under the former conditions, it was noted that cells were more efficient during the 12-hour light period [13]. When grown under constant light, a larger productivity rate was also observed in Nannochloropsis QU130, a strain that has adapted to the harsh climate of the Qatar desert, compared to cultures grown under light cycles [15]. Additionally, in the same strain, fluctuating temperatures increased cell size compared to constant temperatures [15]. Wherein observing cells cultivated under varied temperatures and continuous light resulted in the highest productivity [15].
pH is also a consideration when cultivating microalgae. As more CO2 is taken up by an increasing microalgae population, pH fluctuations occur, thus rendering it necessary to balance pH to keep the microalgae within a tolerable range [13]. The preferred pH for growth can vary depending on strain [17,18]. However, it has been observed that lipid productivity and accumulation are not significantly influenced by unideal pH environments in different microalgae strains [17,18].
In terms of airflow rates, 1L/h of 5% CO2 supplemented air facilitated faster growth for N. salina, as cultures reached a plateau or the stationary phase at about 10 days [13]. Whereas at the lower rate of 0.25 L/h, the stationary phase occurred at approximately 17 days [13]. By increasing both the nitrogen and CO2 sources, an approximately four-fold increase in biomass was observed [13]. Furthermore, in the nitrogen-deficient medium, 63% of N. salina dry weight was comprised of lipids [13]. Beyond air supply, the type of carbon source provided for microalgae cultures can also impact growth rates.
Some microalgae are mixotrophic and thus can utilize inorganic and organic forms of carbon and light for growth. An example is C. vulgaris [14]. In comparison to photoautotrophic growth, mixotrophy can increase productivity, which is dependent on microalgae species and other growth factors [14,19]. To elucidate the impact of mixotrophy in microalgae, a mathematical model was designed to observe the photoautotrophic, heterotrophic, and mixotrophic growth of C. vulgaris [14]. This revealed, the organism prefers photoautotrophic methods in a mixotrophic environment [14]. Therefore, to cater to specific growth and productivity rates, these growth factors must be adjusted accordingly.
Productivity can also be altered by culturing microalgae under stress conditions or extreme growth conditions. For instance, adjusting factors, like temperature, can increase the production of PUFAs [20]. Wherein, Nannochloropsis oculata and Isochrysis galbana were identified as microalgae species with high eicosapentaenoic acid (EPA) productivity at 20 °C and docosahexaenoic acid (DHA) productivity at 14 °C, respectively [20].
Light intensity can also be adjusted to increase growth rates. For instance, high light (360 µmol photons/(m2s)) increased growth approximately three-fold compared to low light conditions (6 µmol photons/(m2s)) for Nannochloropsis gaditana [21]. A similar result was observed in other microalgae species, where increases in light intensity invoked an approximately three-fold increase in fatty acid content in both Desmodesmus sp. and Scenedesmus obliquus after 15 days of cultivation [22]. Furthermore, under higher light intensity conditions, or 300 µE/m2s, the percentage of lipids within their biomass increased compared to the lower light intensity environment of 50 µE/m2s [22].
Nutritional deficiencies and unideal salt concentrations are also stress factors related to growth in microalgae [23]. Wherein under salinity stress, three strains of freshwater microalgae, Ankistrodesmus braunii, Ankistrodesmus falcatus, and Scenedesmus incrassatulus, have heightened lipid content present in their dry biomass [23]. With the addition of varying temperatures and nitrogen supply, the accumulation of lipids increased in Xanthonema hormidioides [24]. Where with a nitrogen concentration at 3 mM and temperature at 25 °C, lipid content within microalgal dry weight increased to its highest at 57.49% dry weight [24]. However, achieving stress conditions is not a requirement to ensure high biomass and productivity.
Microalgae growth can also be assisted by providing optimal growth conditions, which can result in an increase in growth and biomass productivity. This was described by Josephine et al. for C. vulgaris. Which revealed that specific temperature, pH, salinity, and light provide the ideal conditions for microalgae growth [25]. Thus, to provide optimal growth conditions, cultivation systems or devices were introduced to provide these growth factors for both research and industrial purposes alike.
The most common microalgae cultivation systems can be divided into two categories: open systems, such as ponds, and closed systems, like bioreactors [26,27]. Open systems are generally susceptible to contamination, whether it is bacterial or cross-contamination [28]. There are systems that prevent the latter, but still detect concentrations of the former, such as the twin-layer solid-state photobioreactor [28]. Photobioreactors (PBRs) are an example of a closed cultivation system with an additional light module, and thus tend to be preferable for microalgae cultivation. They do not require as much space as open raceway ponds, and have a low possibility of contamination [26]. However, for an open system, like a raceway pond, space requirements and the possibility of contamination are higher [26]. However, it is important to note that, generally, other factors, such as maintenance and setup costs, can be more reasonable [26]. One of the limiting factors for microalgae growth in open raceway ponds is light, as natural light tends to be the only source of this key cultivation factor [16]. Thus, it can only be distributed from the sun, which requires harvesting to occur more often to inhibit negative responses to light attenuation [16]. Furthermore, closed systems, like a bubble column photobioreactor, can cultivate higher biomass yield and nitrogen intake in microalgae compared to an open high-rate pond (HRP) system [29]. However, in an HRP, the net energy ratio or energy efficiency is higher, rendering it more favorable in that aspect [29]. Therefore, dependent on the purpose of the microalgae culture, there are many growth systems that can be tailored for a desirable result.
However, for research purposes, it may be preferable to utilize PBRs, as they can cover a variety of roles. The first role largely involves microalgae cultivation. PBRs allow the controlled delivery of a variety of growth factors, including mass transfer and agitation of the culture, which allows for the ability to adjust these factors for or to observe a certain outcome [30]. For instance, by taking advantage of the device’s ability to control growth conditions, PBRs have been used to observe the impact of various cultivation conditions on productivity in various microalgal species, such as C. vulgaris and Nannochloropsis oceanica [31,32,33,34]. They can also play an indirect role by cultivating microalgae for the design of a model to optimize the organism for biofuel production [32]. This paper will, therefore, delve into the importance of PBRs and their design in microalgae research and industry.
2. Types and Uses of Bioreactors
PBRs come in various designs and capacities (Table 1). For instance, a Fibonacci-type PBR was designed and scaled up to cultivate Dunaliella salina in an extreme solar environment, where growth factors like temperature and pH were controlled within optimal values for the microalgae [35]. This design scaled the culture to 1250 L and increased biomass concentrations three-fold compared to the culture grown in the same conditions in a raceway pond [35]. There are also tubular and panel PBRs, which can increase the efficiency of microalgae biomass productivity in the aerobic phase and biohydrogen production phase, respectively, for Chlamydomonas reinhardtii [36].
In an outdoor photobioreactor with a capacity of 50 L, C. vulgaris FSP-E was grown to optimize the production of protein in the microbial species to lower protein production costs for a fishmeal alternative [31]. A biomass productivity level of 268.1 mg/L/d and the protein productivity level of 155.4 mg/L/d were achieved with this cultivation system [31]. Analysis of the synthesized proteins validated the potential of utilizing the species as a feedstock for the production of a fishmeal alternative [31]. Beyond research applications, PBRs can be used in larger-scale capacities.
PBRs also have diverse uses for industrial purposes. For large-scale wastewater treatment and biofuel production, a floating offshore PBR was designed to address a couple of identifiable issues with biofuel feedstock cultivation [37]. This included scaling up the culture and its longevity [37]. One key element of this PBR’s wastewater remediation function is the cultivation of polycultures, which has aided in the stability, efficiency, and consistency of the culture and its products [37]. Beyond microalgae, wastewater can also be utilized for the cultivation of other microbes, including Zoogloea [38]. These microbes can produce polyhydroxyalkanoates (PHA), a biopolymer studied as an alternative for common plastics, but have not yet been widely adopted due to high production costs [38,49]. As such, a model was designed to predict microbe growth and PHA production in rice winery wastewater with varying organic loading rates (OLRs), which is related to microbe metabolism and productivity [38]. As an average OLR improved PHA productivity [38]. Beyond wastewater treatment, PBRs can also be involved in the mitigation of CO2.
When scaling up Tetraselmis sp. CTP4, tubular PBRs were tested for industrial-scale cultivation [39]. The productivity of photosynthetic microalgae is tied to CO2. As such, CO2 mitigation is a factor observed when considering the success of microalgae. Within a tubular PBR, CO2 mitigation efficiency reached 65%, contributing to the potential of the species for industrial-sized production [39]. Additionally, to improve biomass, this system can be used during the spring and summer months or modified to provide mixotrophic growth conditions [39]. In addition to industrial-scale PBR usage, microalgae applications in industrial capacities have been explored.
Prospective industrial applications of microalgae have been identified with the use of PBRs. This has been achieved through the production of PBR models to simulate microalgae growth and productivity in various climates [40]. Through studying Microchloropsis salina, such a model was created for open thin-layer cascade (TLC) PBRs by observing the species’ lipid productivity while utilizing two scalable TLC PBRs [40]. Through microalgae cultivation in the TLC PBRs and an accompanying model, the potential of M. salina for full-scale productivity of lipids on an industrial scale was revealed and confirmed [40]. However, these examples describe a fraction of bioreactors’ contribution to the research of microbes.
Bioreactors have diverse functions in microbial research, as they can also play an additional role in research by cultivating transgenic organisms. A genetically engineered strain of Streptococcus equi subsp. zooepidemicus was cultivated in a 3 L bioreactor to observe the strain’s ability to produce chondroitin, a precursor of chondroitin sulfate, and hyaluronic acid [44]. The latter two compounds are glycosaminoglycans, which are used in the cosmetic and pharmaceutical industries [44]. The inclusion of genes from Escherichia coli known to be associated with the synthesis of a chondroitin-like sugar, kfoC and kfoA, gave the recombinant S. equi subsp. zooepidemicus, the ability to produce both chondroitin and hyaluronic acid [44]. The 3 L bioreactors were utilized in the cultivation of the recombinant bacteria and, therefore, the production of desired compounds [44]. Bioreactors have also been involved in the larger-scale production of transgenic organisms. Saccharomyces cerevisiae was genetically engineered to produce high amounts of tyrosol and salidroside, used in the production of cosmetics, nutraceuticals, and pharmaceuticals [41]. In terms of bioreactor usage, a 5 L bioreactor was used to scale up the productivity of S. cerevisiae from shake flasks [41]. With this method, the yeast species was able to produce a high amount of the desired products [41].
A bioreactor was also utilized to increase lipid productivity, squalene, and DHA in Aurantiochytrium sp. T66 [46]. The latter of which has nutritional benefits for humans when incorporated into the diet [46,50,51]. Moreover, squalene is the precursor to human steroids, the production of which makes Aurantiochytrium sp. a good candidate for the industrial production of nutraceuticals, cosmetics, and pharmaceuticals [46]. After cultivation in a bioreactor, total lipid concentration was 5.90 g/L, where DHA made up 35.76%, and squalene yield increased by approximately 0.28 g/L in comparison to flask-cultivated microbes [46].
Bioreactors can also be used to cultivate plant cell lineswith the ability to produce compounds with various applications in cosmetics, nutraceuticals, and food coloring [45]. This plant biomass, from the red carrot R4G cell line, was observed to contain high concentrations of anthocyanins, which may point to its use as a food colorant [45]. Furthermore, the carrot cells’ biomass had a variety of health benefits in mouse cells, including anti-aging, anti-inflammatory, and antioxidant activity [45]. These results further identify the impact of bioreactors on research for the production of a variety of useful products, the diverse applications of which are noted in Figure 2.
When scaling up the production of microalgae with photobioreactors, several issues may arise (Table 2). Notably, keeping the process low-cost, culture collapse, and contamination [33,43]. However, to address the latter two, a two-stage process was created with C. vulgaris culture [33]. The first stage of this methodology requires the cultivation of the microalgae species in fermenters [33]. Stage two is inoculating the culture in a larger capacity flat panel PBR with the ability to culture 1000 L of microalgae [33]. These steps describe a heterotrophic method of scaling up [33]. By utilizing the autotrophic growth method, growing 1000 L of C. vulgaris would take approximately 35 days, whereas the previously mentioned method requires approximately five days [33]. Furthermore, the heterotrophic method essentially decreases the time and space required for scaled-up microalgae production in PBRs [33]. Therefore, this emphasizes the importance of identifying the ideal growth system for an intended purpose, whether that be for research or industry.
3. Photobioreactors for Energy
Currently, there is interest in increasing sustainability through the production of biofuels sourced from microbes like microalgae, illustrated in Figure 3. However, there are several challenges that have inhibited the usage of the product worldwide [42]. For instance, identifying the microbe species to be used as feedstock and determining its success in scaled-up systems can be difficult [42].
Biodiesel quality is correlated with the concentration of fatty acid type, which plays a role in the biofuel’s characteristics, such as cold-flow properties [48]. This makes it necessary to design an efficient method for microalgae cultivation to maximize ideal lipid concentrations for biodiesel production [48]. Which was performed through validation of Coelastrum sp. SM cultivated in an air-lift PBR as a feedstock for biofuel and fixation of CO2 [48]. Wherein analysis recorded that the highest lipid content reached 37.91% of dry weight and the highest carbohydrate content was 58.45% of dry weight. Furthermore, the culture had a CO2 fixation rate of 0.302 g/L*h, confirming the utility of the species in biodiesel production and CO2 fixation [48].
CO2 fixation has also been recorded in C. vulgaris cultivated in tubular PBRs [34]. The system was tested for its ability to scale up the microalgae culture to 100 L while utilizing sodium bicarbonate as a CO2 source [34]. As a result, lipid concentrations increased to about 26%, and CO2 fixation was approximately 0.925 g/L*d [34]. As the maximum concentration of sodium bicarbonate to completely increase lipid productivity has not yet been elucidated, we can only conclude that lipid productivity and sodium bicarbonate concentration are directly related, wherein an increase in the latter results in the increase of the former [34].
To produce biodiesel or biofuel, microalgae’s lipid productivity is a point of interest. In an open system, a raceway pond, Nannochloropsis sp. KMMCC 290 was cultivated to produce biodiesel [1]. In a flat-plate photobioreactor (FPP) under control conditions, or with a light intensity of 5,800 lux and continuous air supply with no carbon dioxide, lipid content was higher than in the raceway pond [1]. Whereas, under increased light intensity, gas exchange rates, and CO2 supply, lipid productivity was about 16.6-fold higher than in the raceway pond [1]. At the conclusion of this research, the productivity in different systems, such as a bubble column PBR and air-lift PBR, provided varying productivity rates, thus further pointing to the importance of identifying the optimal growth system for a desired result [1]. Another species, S. obliquus, was cultivated in a plastic-type flat panel PBR with a volume of 5 L [47]. When the nitrogen source for the culture was urea and light intensity reached 3000 lux with white-colored, fluorescent bulbs, the resulting culture’s dry biomass contained 40% lipids [47]. Furthermore, 66.6% of lipids were unsaturated fatty acids, which makes the species ideal for biofuel production [47]. The potential of microalgae species for alternative energy sources has also been realized through the production of models.
A computational fluid dynamic model was utilized to experimentally scale up a mutant cell line of C. reinhardtii by simulating varying sparger designs in a 120 L flat-plate photobioreactor [42]. The model predicted a biomass productivity increase of 18% in C. reinhardtii grown within the optimized closed PBR [42]. These studies have revealed the importance of PBRs in the synthesis of renewable energy sources utilizing the productivity of microalgae.
4. Incubation Factors for Photobioreactors
An effective photobioreactor design must take into consideration a variety of growth factors, such as hydrodynamics, light, growth kinetics, agitation, nutrient supply, and gas exchange (Table 3, Figure 4) [53].
Light is an important factor for microalgae growth. For example, a light intensity of 350 μmol/m2s, results in the highest productivity of carbohydrates at 48.11 gC/m3d for I. galbana [57].
Light intensity, gas exchange, and CO2 supply impact cell count in Nannochloropsis sp. KMMCC 290 cultures [1]. Increasing the growth factors previously listed can improve lipid content in cells [1]. Furthermore, light cycles can inhibit cell replication and stress cells, which induces an increase in lipid production [1]. Light as a growth factor, can be controlled with cultivation systems.
The thickness of a flat plate photobioreactor (FPP) impacts the light intensity received by the algae culture in the device [55]. The thinner the FPP, or the smaller the light path, the higher the lipid productivity and content in cells [55]. However, for a light path of 5 cm compared with the 10 cm light path, values such as growth rate, cell dry weight, and biomass productivity, were lower [55]. Thus, it is important to consider light intensity for the purpose of providing ideal growth conditions.
In addition to light intensity, cell density, growth medium, and light cycles also influence microalgae growth [56]. For Chlorococcum sp., saline water, 2500-3500 lux, and growth under 24 hours of light, resulted in optimal growth of the species on the fifth day [56]. However, this growth rate additionally relies on cell density, wherein low initial cell densities have a longer death phase after the 11th day of growth [56].
Temperature conditions are also an important consideration for microalgae cultivation. Under high-temperature stress, at 30 °C, lipid productivity improved in a mixed microalgae culture collected from the Nacharam Cheruvu in India [62]. Furthermore, analysis of lipid content noted a high concentration of saturated fatty acids due to stress phase growth under the mentioned temperature conditions [62]. At 25 °C, the growth rate was higher than hotter temperatures, for Chaetoceros sp. FIKU035 and Nannochloropsis sp. FIKU036 [54]. Whereas Tetraselmis suecica FIKU032 had a slightly higher growth rate at 30 °C, and could not be cultivated in higher temperatures [54]. Variability was also observed in biomass productivity and concentration [54]. Where these values were the highest for Nannochloropsis sp. FIKU036 cultivated at 25 °C, and the highest for Chaetoceros sp. FIKU035 and T. suecica FIKU032 at 30 °C [54].
Nutrient supply is highly involved with productivity in microalgae [60]. In freshwater with fertilizer, Tetradesmus almeriensis cultivated in a pilot-scale thin-layer cascade PBR had the most biomass productivity of 30.3 g/m2*day [60]. Whereas when the microalgae were grown in wastewater, protein and lipid productivity increased [60]. When Chlorella sp. GN1 was grown in nitrogen deprivation conditions, the lipid content increased in dry cell weight compared to phosphorous limitation or nitrogen and phosphorous sufficient conditions [55]. Under nitrogen-deprived conditions, lipid productivity reached 63.5 mg/L*day [55]. However, lipid concentration under nutrient-limited conditions was lower than Chlorella sp. GN1 was grown in nitrogen and phosphorous-sufficient conditions after eight days of growth [55]. Nutrient availability also impacts the productivity of other beneficial compounds in microalgae.
Fucoxanthin is a pigment with a variety of health benefits, including as an antioxidant, which has made the compound highly sought after to produce nutraceuticals and pharmaceuticals [3,4]. As such, methods to optimize the productivity of fucoxanthin have been studied [4]. This has revealed that providing high nitrogen levels in the initial f/2 growth medium for Phaeodactylum tricornutum can result in high productivity thus, increase biomass and concentrations of fucoxanthin [4]. Furthermore, high nitrogen media and low light conditions can increase fucoxanthin production [4]. This points to both growth factors attributing to the fucoxanthin production process in P. tricornutum [4].
Higher air flow correlated with higher cell density in Tisochrysis lutea cultivated in a bench-scale air-lift PBR [61]. Generally, for column-style photobioreactors, air supply has a positive correlation with gas exchange, as a result of each microalgae cell within the culture receiving more light exposure [61]. Furthermore, reducing hydrodynamic stress, caused by factors like shear stress, can further increase cell growth [61].
Different microalgae species may react to hydrodynamic stress differently [63]. Both shear rate and shear stress are values associated with calculating shear forces [64]. High shear stress and shear rate can damage cells, and microalgae’s reaction and resilience to such stress varies by strain [64]. However, shear stress near the walls of closed growth systems is necessary to inhibit the growth of a biofilm [65]. Biofilm formation can prevent the microalgae culture from receiving light [65]. Thus, C. vulgaris biofilm prevention and removal through wall shear forces was pursued through cultivation in a flat-panel PBR [65]. It was revealed that 0.2 Pa of wall shear stress prevents biofilm formation, whereas 6 Pa can disrupt an established biofilm, and 53 Pa is necessary to remove it [65].
Shear force can also be utilized to increase mass transfer, thus improving microalgae’s CO2 fixation [59]. This was achieved through applying shear force through water centrifugation to decrease the overall size of bubbles, or increase the bubbles’ surface area, within the microalgae culture [59]. This method increased biomass productivity by 50.7% [59]. However, high aeration creates high levels of shear stress, preventing biomass productivity of Arthrospira platensis in 2 L photobioreactors [66]. Thus, it is important to develop optimal growth settings to introduce reasonable levels of shear stress for the growing microalgae cultures.
Simulations of an air-lift photobioreactor predict that higher gas flow rates cause increases in photosynthetic efficiency [58]. This is due to the cells’ light exposure increasing as a result of introducing agitation [58]. However, this comes with a disclaimer, as high gas flow rates also cause high levels of shear stress, which decreases biomass [58]. Thus, for Porphyridium sp. the increased fluid mixing, a result of higher gas flow rates, also increases shear stress, causing cell damage,thus leading to decreased biomass productivity [58]. Additionally, challenges in microalgae cultivation come in the form of contamination.
Rotifers are predators of microalgae [67,68,69]. These organisms can decimate microalgae cultures in both open ponds and closed photobioreactors when introduced [68]. However, rotifer contamination is restricted in thin-layer cascade reactors, as their population density is limited in the system [68]. This notes that the impacts o[67,68,69f rotifer contamination vary in different growth systems [68].
The role of growth kinetic models is to convey how microalgae grow in specific environments within a time period [16,70]. As such, there are currently a variety of growth kinetic models available for microalgae species [16]. These models can point to the downsides of different growth methods, for instance, in open raceway ponds, growth kinetics have pointed toward the main issue of light attenuation [16]. One such model for an open raceway pond notes that systems around 30-35 cm are advantageous for outdoor growth, whereas values above this range are not [16].
One method to analyze cell growth kinetics involves maximum growth rate, a value calculated with the biomass concentration of the desired microalgae strain at a specific time period [66]. Additionally, cell productivity must also be calculated through the varied cell density over the cultivation period [66]. This method was utilized to calculate the growth kinetics of A. platensis in PBRs [66]. Growth rate kinetics can also be calculated with the Contois Equation, the differential of substrate concentration with consideration to time, and the differential of carbohydrate concentration with consideration to time [71]. This method can reveal the growth rate of microalgae cultivated using thin-layer photobioreactors in different mediums [71].
Furthermore, different growth kinetic models fit with experimental data [70]. For instance, the Gompertz model is more accurate than the Logistic model in determining the cultivation of Scenedesmus parvus in a PBR [70]. Both of these models utilize the same variables to calculate growth kinetics, such as initial and largest biomass, growth rate, and time, however, are arranged differently to gain varying results [70]. As a result, the more accurate model can be utilized to scale up cultures for industrial purposes in the future [70].
Growth kinetics can also model growth limitations reliant on dissolved inorganic carbon (DIC) concentration in the microalgae culture [72]. Wherein low levels of DIC, or limited carbon, can result in a decrease in growth kinetics [72]. However, this model has not been developed to consider the impact of mixotrophic growth on CO2 remediation by microalgae cultures [72]. Thus, growth kinetics can also assist with determining the ideal growth factors and system for microalgae cultivation.
5. Challenges and Perspectives
The cultivation of microalgae heavily relies upon growth conditions, wherein, the delivery of such factors can be dictated through the usage of cultivation systems [26,30]. These systems, categorized as either open or closed, differ greatly in their characteristics [26,27]. Generally, open systems have a higher risk of contamination from either other eukaryotic species or predators of microalgae, but can cater to larger volumes [26,28]. As such, for monoculture of smaller volumes, a PBR may be preferable, as the system provides the ability to deliver controlled growth conditions such as light and temperature. Of which these qualities in an open system, like a raceway pond, are more difficult to control [16,39]. Additionally, the usage of closed systems has a lower risk of contamination [26]. However, there are some recognizable challenges that PBRs face, which have been mainly tied to scaling up cultures [33,43]. Where production costs and limited cultivation volumes of PBRs pose issues [43]. Furthermore, large-volume outdoor PBRs are exposed to seasonal changes and natural light, which can vary drastically as time passes [35,39]. Thus, these challenges emphasize a need for more diverse PBR designs that can address some of the downsides of current models available. Presently, this has been pursued, as there are some PBR designs that are slightly more favorable for larger-scale microalgae production [33,35].
However, these models further confirm that there is a wide range of considerations when identifying or designing an ideal microalgae growth system. One must take into account the purpose of the final microalgae culture, whether that be for research or industrial purposes. The scale of the product and, if applicable, desired enhancements in productivity. This will assist in determining the type of microalgae growth system and the conditions required to induce desired biomass accumulation and productivity rates. For microalgae research, many of these considerations are addressed through the usage of PBRs that have allowed finer control of various growth factors.
Author Contributions
Z.D. developed the idea and outline of the article. S.D. made the figures and tables and wrote the manuscript. Z.D. critically revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by UHM UROP-Undergraduate Research Opportunities Program, USDA-NIFA HATCH project HAW05047-H, State of Hawaii Department of Agriculture, Center for Tropical and Subtropical Aquaculture through Grant No. 2020-38500-32559 and 2022-38500-38099 from the U.S. Department of Agriculture National Institute of Food and Agriculture.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Ju-Ling Chen (UROP, UHM), Kanak Pal, Ty Shitanaka, Cade Kane, and Rumesh Senthilnathan, for their help with this paper.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kwon, M. H., & Yeom, S. H. (2017). Evaluation of closed photobioreactor types and operation variables for enhancing lipid productivity of Nannochloropsis sp. KMMCC 290 for biodiesel production. Biotechnology and Bioprocess Engineering, 22(5), 604–611. [CrossRef]
- Regueiras, A., Huguet, Á., Conde, T., Couto, D., Domingues, P., Domingues, M. R., Costa, A. M., Silva, J. L. da, Vasconcelos, V., & Urbatzka, R. (2022). Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella vulgaris and Chlorococcum amblystomatis under Different Growth Conditions. Marine Drugs, 20(1), 9. [CrossRef]
- Arunkumar, K., Nalluri, M., Anjana, K., Mohan, G., & Raja, R. (2023). Fucoxanthin as antioxidant, anti-hyaluronidase and cytotoxic agent: Potential of brown seaweeds decoction for tea supplement. Journal of Food Measurement and Characterization, 17(4), 3980–3989. [CrossRef]
- Truong, T. Q., Park, Y. J., Winarto, J., Huynh, P. K., Moon, J., Choi, Y. B., Song, D.-G., Koo, S. Y., & Kim, S. M. (2024). Understanding the Impact of Nitrogen Availability: A Limiting Factor for Enhancing Fucoxanthin Productivity in Microalgae Cultivation. Marine Drugs, 22(2), 93. [CrossRef]
- Arashiro, L. T., Josa, I., Ferrer, I., Van Hulle, S. W. H., Rousseau, D. P. L., & Garfí, M. (2022). Life cycle assessment of microalgae systems for wastewater treatment and bioproducts recovery: Natural pigments, biofertilizer and biogas. Science of The Total Environment, 847, 157615. [CrossRef]
- Abraham, J., Prigiobbe, V., Abimbola, T., & Christodoulatos, C. (2023). Integrating biological and chemical CO2 sequestration using green microalgae for bioproducts generation. Frontiers in Climate, 4. [CrossRef]
- Xin, Y., Shen, C., She, Y., Chen, H., Wang, C., Wei, L., Yoon, K., Han, D., Hu, Q., & Xu, J. (2019). Biosynthesis of Triacylglycerol Molecules with a Tailored PUFA Profile in Industrial Microalgae. Molecular Plant, 12(4), 474–488. [CrossRef]
- Matos, J., Cardoso, C. L., Falé, P., Afonso, C. M., & Bandarra, N. M. (2020). Investigation of nutraceutical potential of the microalgae Chlorella vulgaris and Arthrospira platensis. International Journal of Food Science & Technology, 55(1), 303–312. [CrossRef]
- Shimidzu, N., Goto, M., & Miki, W. (1996). Carotenoids as Singlet Oxygen Quenchers in Marine Organisms. Fisheries Science, 62(1), 134–137. [CrossRef]
- Ahmed, F., Fanning, K., Netzel, M., Turner, W., Li, Y., & Schenk, P. M. (2014). Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chemistry, 165, 300–306. [CrossRef]
- Zhang, Z., Wang, B., Hu, Q., Sommerfeld, M., Li, Y., & Han, D. (2016). A New Paradigm for Producing Astaxanthin From the Unicellular Green Alga Haematococcus pluvialis. Biotechnology and Bioengineering, 113(10), 2088–2099. [CrossRef]
- Chaudhary, A., Mishra, P., Amaz, S. A., Mahato, P. L., Das, R., Jha, R., & Mishra, B. (2023). Dietary supplementation of microalgae mitigates the negative effects of heat stress in broilers. Poultry Science, 102(10), 102958. [CrossRef]
- Sforza, E., Bertucco, A., Morosinotto, T., & Giacometti, G. M. (2012). Photobioreactors for microalgal growth and oil production with Nannochloropsis salina: From lab-scale experiments to large-scale design. Chemical Engineering Research and Design, 90(9), 1151–1158. [CrossRef]
- Manhaeghe, D., Blomme, T., Van Hulle, S. W. H., & Rousseau, D. P. L. (2020). Experimental assessment and mathematical modelling of the growth of Chlorella vulgaris under photoautotrophic, heterotrophic and mixotrophic conditions. Water Research, 184, 116152. [CrossRef]
- Al Jabri, H., Taleb, A., Touchard, R., Saadaoui, I., Goetz, V., & Pruvost, J. (2021). Cultivating Microalgae in Desert Conditions: Evaluation of the Effect of Light-Temperature Summer Conditions on the Growth and Metabolism of Nannochloropsis QU130. Applied Sciences, 11(9), 3799. [CrossRef]
- Romagnoli, F., Weerasuriya-Arachchige, A. R. P. P., Paoli, R., Feofilovs, M., & Ievina, B. (2021). Growth Kinetic Model for Microalgae Cultivation in Open Raceway Ponds: A System Dynamics Tool. Environmental and Climate Technologies, 25(1), 1317–1336. [CrossRef]
- Bartley, M. L., Boeing, W. J., Dungan, B. N., Holguin, F. O., & Schaub, T. (2014). pH effects on growth and lipid accumulation of the biofuel microalgae Nannochloropsis salina and invading organisms. Journal of Applied Phycology, 26, 1431–1437. [CrossRef]
- Qiu, R., Gao, S., Lopez, P. A., & Ogden, K. L. (2017). Effects of pH on cell growth, lipid production and CO2 addition of microalgae Chlorella sorokiniana. Algal Research, 28, 192–199. [CrossRef]
- Patel, A., Matsakas, L., Hrůzová, K., Rova, U., & Christakopoulos, P. (2019). Biosynthesis of Nutraceutical Fatty Acids by the Oleaginous Marine Microalgae Phaeodactylum tricornutum Utilizing Hydrolysates from Organosolv-Pretreated Birch and Spruce Biomass. Marine Drugs, 17(2), 119. [CrossRef]
- Aussant, J., Guihéneuf, F., & Stengel, D. B. (2018). Impact of temperature on fatty acid composition and nutritional value in eight species of microalgae. Applied Microbiology & Biotechnology, 102(12), 5279–5297. [CrossRef]
- Perin, G. Perin, G., Cimetta, E., Monetti, F., Morosinotto, T., & Bezzo, F. (2016). Novel micro-photobioreactor design and monitoring method for assessing microalgae response to light intensity. Algal Research, 19, 69–76. [CrossRef]
- Nzayisenga, J. C., Farge, X., Groll, S. L., & Sellstedt, A. (2020). Effects of light intensity on growth and lipid production in microalgae grown in wastewater. Biotechnology for Biofuels, 13(1), 4. [CrossRef]
- Fawzy, M. A., El-Otify, A. M., Adam, M. S., & Moustafa, S. S. A. (2021). The impact of abiotic factors on the growth and lipid accumulation of some green microalgae for sustainable biodiesel production. Environmental Science and Pollution Research, 28(31), 42547–42561. [CrossRef]
- Gao, B., Hong, J., Chen, J., Zhang, H., Hu, R., & Zhang, C. (2023). The growth, lipid accumulation and adaptation mechanism in response to variation of temperature and nitrogen supply in psychrotrophic filamentous microalga Xanthonema hormidioides (Xanthophyceae). Biotechnology for Biofuels and Bioproducts, 16(1), 12. [CrossRef]
- Josephine, A., Kumar, T. S., Surendran, B., Rajakumar, S., Kirubagaran, R., & Dharani, G. (2022). Evaluating the effect of various environmental factors on the growth of the marine microalgae, Chlorella vulgaris. Frontiers in Marine Science, 9. [CrossRef]
- Narala, R. R., Garg, S., Sharma, K. K., Thomas-Hall, S. R., Deme, M., Li, Y., & Schenk, P. M. (2016). Comparison of Microalgae Cultivation in Photobioreactor, Open Raceway Pond, and a Two-Stage Hybrid System. Frontiers in Energy Research, 4. [CrossRef]
- Prado, L. O., Bolzani, H. R., Souza, H. H. S., Ruas, G., & Silva, G. H. R. (2023). Microalgal cultivation in open and closed systems under a tropical climate: A life cycle comparison. Journal of Cleaner Production, 422, 138631. [CrossRef]
- Naumann, T., Çebi, Z., Podola, B., & Melkonian, M. (2013). Growing microalgae as aquaculture feeds on twin-layers: A novel solid-state photobioreactor. Journal of Applied Phycology, 25(5), 1413–1420. [CrossRef]
- Magalhães, I. B., Ferreira, J., Castro, J. de S., Assis, L. R. de, & Calijuri, M. L. (2022). Agro-industrial wastewater-grown microalgae: A techno-environmental assessment of open and closed systems. Science of The Total Environment, 834, 155282. [CrossRef]
- Fernandes, B. D., Mota, A., Ferreira, A., Dragone, G., Teixeira, J. A., & Vicente, A. A. (2014). Characterization of split cylinder air-lift photobioreactors for efficient microalgae cultivation. Chemical Engineering Science, 117, 445–454. [CrossRef]
- Chen, C.-Y., Chang, Y.-H., & Chang, H.-Y. (2016). Outdoor cultivation of Chlorella vulgaris FSP-E in vertical tubular-type photobioreactors for microalgal protein production. Algal Research, 13, 264–270. [CrossRef]
- del Rio-Chanona, E. A., Liu, J., Wagner, J. L., Zhang, D., Meng, Y., Xue, S., & Shah, N. (2018). Dynamic modeling of green algae cultivation in a photobioreactor for sustainable biodiesel production. Biotechnology and Bioengineering, 115(2), 359–370. [CrossRef]
- Barros, A., Pereira, H., Campos, J., Marques, A., Varela, J., & Silva, J. (2019). Heterotrophy as a tool to overcome the long and costly autotrophic scale-up process for large scale production of microalgae. Scientific Reports, 9, 13935. [CrossRef]
- Ratomski, P., Hawrot-Paw, M., & Koniuszy, A. (2021). Utilisation of CO2 from Sodium Bicarbonate to Produce Chlorella vulgaris Biomass in Tubular Photobioreactors for Biofuel Purposes. Sustainability, 13(16), 9118. [CrossRef]
- Díaz, J. P., Inostroza, C., & Acién, F. G. (2021). Scale-up of a Fibonacci-Type Photobioreactor for the Production of Dunaliella salina. Applied Biochemistry and Biotechnology, 193(1), 188–204. [CrossRef]
- Oncel, S., & Kose, A. (2014). Comparison of tubular and panel type photobioreactors for biohydrogen production utilizing Chlamydomonas reinhardtii considering mixing time and light intensity. Bioresource Technology, 151, 265–270. [CrossRef]
- Novoveská, L., Zapata, A. K. M., Zabolotney, J. B., Atwood, M. C., & Sundstrom, E. R. (2016). Optimizing microalgae cultivation and wastewater treatment in large-scale offshore photobioreactors. Algal Research, 18, 86–94. [CrossRef]
- Fang, F., Xu, R.-Z., Huang, Y.-Q., Wang, S.-N., Zhang, L.-L., Dong, J.-Y., Xie, W.-M., Chen, X., & Cao, J.-S. (2019). Production of polyhydroxyalkanoates and enrichment of associated microbes in bioreactors fed with rice winery wastewater at various organic loading rates. Bioresource Technology, 292, 121978. [CrossRef]
- Pereira, H., Páramo, J., Silva, J., Marques, A., Barros, A., Maurício, D., Santos, T., Schulze, P., Barros, R., Gouveia, L., Barreira, L., & Varela, J. (2018). Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: From an agar plate to 100-m3 industrial photobioreactors. Scientific Reports, 8, 5112. [CrossRef]
- Schädler, T., Thurn, A.-L., Brück, T., & Weuster-Botz, D. (2021). Continuous Production of Lipids with Microchloropsis salina in Open Thin-Layer Cascade Photobioreactors on a Pilot Scale. Energies, 14(2), 500. [CrossRef]
- Liu, H., Tian, Y., Zhou, Y., Kan, Y., Wu, T., Xiao, W., & Luo, Y. (2021). Multi-modular engineering of Saccharomyces cerevisiae for high-titre production of tyrosol and salidroside. Microbial Biotechnology, 14(6), 2605–2616. [CrossRef]
- Ali, H., Solsvik, J., Wagner, J. L., Zhang, D., Hellgardt, K., & Park, C. W. (2019). CFD and kinetic-based modeling to optimize the sparger design of a large-scale photobioreactor for scaling up of biofuel production. Biotechnology and Bioengineering, 116(9), 2200–2211. [CrossRef]
- Bani, A., Fernandez, F. G. A., D’Imporzano, G., Parati, K., & Adani, F. (2021). Influence of photobioreactor set-up on the survival of microalgae inoculum. Bioresource Technology, 320, 124408. [CrossRef]
- Cimini, D., Iacono, I. D., Carlino, E., Finamore, R., Restaino, O. F., Diana, P., Bedini, E., & Schiraldi, C. (2017). Engineering S. equi subsp. Zooepidemicus towards concurrent production of hyaluronic acid and chondroitin biopolymers of biomedical interest. AMB Express, 7, 61. [CrossRef]
- Bianconi, M., Ceriotti, L., Cuzzocrea, S., Esposito, E., Pressi, G., Sgaravatti, E., Bertaiola, O., Guarnerio, C., Barbieri, E., Semenzato, A., Negri, S., Commisso, M., Avesani, L., & Guzzo, F. (2020). Red Carrot Cells Cultured in vitro Are Effective, Stable, and Safe Ingredients for Skin Care, Nutraceutical, and Food Applications. Frontiers in Bioengineering and Biotechnology, 8, 575079. [CrossRef]
- Patel, A., Rova, U., Christakopoulos, P., & Matsakas, L. (2019). Simultaneous production of DHA and squalene from Aurantiochytrium sp. grown on forest biomass hydrolysates. Biotechnology for Biofuels, 12(1), 255. [CrossRef]
- Abdel-Baset, A., Matter, I. A., & Ali, M. A. (2024). Enhanced Scenedesmus obliquus Cultivation in Plastic-Type Flat Panel Photobioreactor for Biodiesel Production. Sustainability, 16, 3148. [CrossRef]
- Mousavi, S., Najafpour, G. D., & Mohammadi, M. (2018). CO2 bio-fixation and biofuel production in an air-lift photobioreactor by an isolated strain of microalgae Coelastrum sp. SM under high CO2 concentrations. Environmental Science and Pollution Research, 25(30), 30139–30150. [CrossRef]
- Leong, Y. K., Show, P. L., Lin, H. C., Chang, C. K., Loh, H.-S., Lan, J. C.-W., & Ling, T. C. (2016). Preliminary integrated economic and environmental analysis of polyhydroxyalkanoates (PHAs) biosynthesis. Bioresources and Bioprocessing, 3(1), 41. [CrossRef]
- Swanson, D., Block, R., & Mousa, S. A. (2012). Omega-3 Fatty Acids EPA and DHA: Health Benefits Throughout Life. Advances in Nutrition, 3(1), 1-7. [CrossRef]
- Bernasconi, A. A., Wiest, M. M., Lavie, C. J., Milani, R. V., & Laukkanen, J. A. (2021). Effect of Omega-3 Dosage on Cardiovascular Outcomes: An Updated Meta-Analysis and Meta-Regression of Interventional Trials. Mayo Clinic Proceedings, 96(2), 304-313. [CrossRef]
- Gupta, P. L., Lee, S.-M., & Choi H.-J. (2015). A mini review: photobioreactors for large scale algal cultivation. World Journal of Microbiology and Biotechnology, 31(9), 1409-1417. [CrossRef]
- Li, M.-J., Tong, Z.-X., Zhou, Z.-J., Huang, D., & Wang, R.-L. (2019). A numerical model coupling bubble flow, light transfer, cell motion and growth kinetics for real timescale microalgae cultivation and its applications in flat plate photobioreactors. Algal Research, 44, 101727. [CrossRef]
- Chaisutyakorn, P., Praiboon, J., & Kaewsuralikhit, C. (2018). The effect of temperature on growth and lipid and fatty acid composition on marine microalgae used for biodiesel production. Journal of Applied Phycology, 30(1), 37–45. [CrossRef]
- Feng, P., Xu, Z., Qin, L., Alam, M. A., Wang, Z., & Zhu, S. (2020). Effects of different nitrogen sources and light paths of flat plate photobioreactors on the growth and lipid accumulation of Chlorella sp. GN1 outdoors. Bioresource Technology, 301, 122762. [CrossRef]
- Putri, D. S., Sari, D. A., Marianah, Astuti, S. P., & Wangiyana, I. G. A. S. (2021). Effect of medium type, light intensity, and photoperiod on the growth rate of microalgae Chlorococcum sp. local isolate. IOP Conference Series: Earth and Environmental Science, 913, 012071. [CrossRef]
- Guzmán-Palomino, A., Aguilera-Vázquez, L., Hernández-Escoto, H., García-Vite, P. M., & Martínez-Salazar, A. L. (2023). Dynamical Simulation, Sensitivity, and Productivity Analysis of a Light-Photoacclimation Model for Microalgae-Based Carbohydrate Production in Continuous Photobioreactors. Processes, 11(7), 1866. [CrossRef]
- Gao, X., Kong, B., & Vigil, R. D. (2018). Multiphysics simulation of algal growth in an air-lift photobioreactor: Effects of fluid mixing and shear stress. Bioresource Technology, 251, 75–83. [CrossRef]
- Cheng, J., Miao, Y., Guo, W., Song, Y., Tian, J., & Zhou, J. (2018). Reduced generation time and size of carbon dioxide bubbles in a volute aerator for improving Spirulina sp. growth. Bioresource Technology, 270, 352–358. [CrossRef]
- Villaró, S., Sánchez-Zurano, A., Ciardi, M., Alarcón, F. J., Clagnan, E., Adani, F., Morillas-España, A., Álvarez, C., & Lafarga, T. (2022). Production of microalgae using pilot-scale thin-layer cascade photobioreactors: Effect of water type on biomass composition. Biomass and Bioenergy, 163, 106534. [CrossRef]
- Falinski, K. A., Timmons, M. B., Callan, C., & Laidley, C. (2018). Response of Tisochrysis lutea [Prymnesiophycidae] to aeration conditions in a bench-scale photobioreactor. Journal of Applied Phycology, 30(4), 2203–2214. [CrossRef]
- Venkata Subhash, G., Rohit, M. V., Devi, M. P., Swamy, Y. V., & Venkata Mohan, S. (2014). Temperature induced stress influence on biodiesel productivity during mixotrophic microalgae cultivation with wastewater. Bioresource Technology, 169, 789–793. [CrossRef]
- Leupold, M., Hindersin, S., Gust, G., Kerner, M., & Hanelt, D. (2013). Influence of mixing and shear stress on Chlorella vulgaris, Scenedesmus obliquus, and Chlamydomonas reinhardtii. Journal of Applied Phycology, 25(2), 485–495. [CrossRef]
- Guler, B. A., Deniz, I., Demirel, Z., Oncel, S. S., & Imamoglu, E. (2020). Computational fluid dynamics modelling of stirred tank photobioreactor for Haematococcus pluvialis production: Hydrodynamics and mixing conditions. Algal Research, 47, 101854. [CrossRef]
- Belohlav, V., Zakova, T., Jirout, T., & Kratky, L. (2020). Effect of hydrodynamics on the formation and removal of microalgal biofilm in photobioreactors. Biosystems Engineering, 200, 315–327. [CrossRef]
- Hussin, A. A., To, S. W., Sani, M. H., Amin, M. F. M., & Kamaroddin, M. F. (2021). Optimisation and growth kinetic analysis of Microalgae, Arthrospira platensis in 2-L Photobioreactors. IOP Conference Series: Earth and Environmental Science, 842, 012036. [CrossRef]
- Wang, Y., Castillo-Keller, M., Eustance, E., & Sommerfeld, M. (2017). Early detection and quantification of zooplankton grazers in algal cultures by FlowCAM. Algal Research, 21, 98–102. [CrossRef]
- Deruyck, B., Nguyen, K. H. T., Decaestecker, E., & Muylaert, K. (2019). Modeling the impact of rotifer contamination on microalgal production in open pond, photobioreactor and thin layer cultivation systems. Algal Research, 38, 101398. [CrossRef]
- Martínez, C., Pessi, B. A., & Bernard, O. (2021). Dynamics and productivity of microalgae in presence of predators. IFAC-PapersOnLine, 54(3), 673–678. [CrossRef]
- Tan, K. M., Kassim, M. A., Ng, Z. J., & Lalung, J. (2020). Isolation and characterization of novel acidophilic microalgae from abandoned mining site area for carbohydrate biosynthesis and its kinetic growth study in photobioreactor. IOP Conference Series: Materials Science and Engineering, 716, 012011. [CrossRef]
- Yuarrina, W. P., Pradana, Y. S., Budiman, A., Majid, A. I., Indarto, & Suyono, E. A. (2018). Study of cultivation and growth rate kinetic for mixed cultures of local microalgae as third generation (G-3) bioethanol feedstock in thin layer photobioreactor. Journal of Physics: Conference Series, 1022(1), 012051. [CrossRef]
- Pruvost, J., Le Gouic, B., & Cornet, J.-F. (2022). Kinetic Modeling of CO2 Biofixation by Microalgae and Optimization of Carbon Supply in Various Photobioreactor Technologies. ACS Sustainable Chemistry & Engineering, 10(38), 12826–12842. [CrossRef]
Figure 1.
The applications of microalgae are diverse. As photosynthetic organisms, they can be used to sequester CO2. Furthermore, they can utilize wastewater as a growth medium while removing pollutants. During both processes, microalgae can simultaneously produce several important products. Of which have been involved in the creation of nutraceuticals, pharmaceuticals, cosmetics, animal feed, and biofuel. These microalgae-based products and remediation methodologies have important implications for sustainability in the future.
Figure 1.
The applications of microalgae are diverse. As photosynthetic organisms, they can be used to sequester CO2. Furthermore, they can utilize wastewater as a growth medium while removing pollutants. During both processes, microalgae can simultaneously produce several important products. Of which have been involved in the creation of nutraceuticals, pharmaceuticals, cosmetics, animal feed, and biofuel. These microalgae-based products and remediation methodologies have important implications for sustainability in the future.
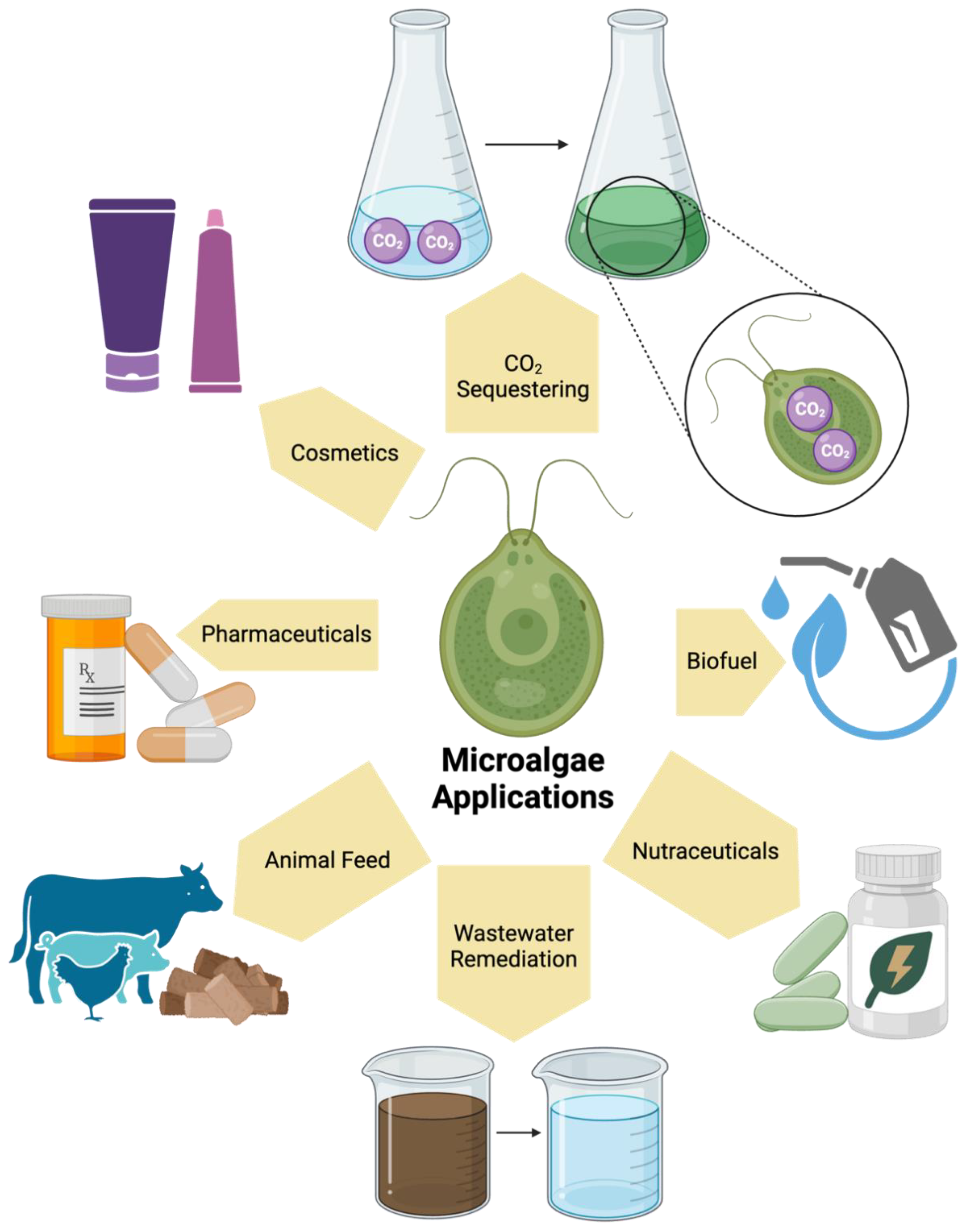
Figure 2.
One group of closed growth systems is bioreactors. These are systems with a variety of modules, such as agitation systems, temperature controls, and gas flow management. Thus, their ability to strictly control these parameters makes them an ideal system for microbe and plant cell line cultivation, and scaled growth. Through observation of the productivity of microbes or plant cell lines, bioreactors have assisted in the production of compounds such as fatty acids, pigments, and biohydrogen. Some of which have been used to synthesize products such as fish feed and biofuel. Furthermore, the growth of microbes in bioreactors, such as microalgae, has been used to study processes like wastewater remediation. All these applications have contributed to identifying the diverse functions of bioreactors for both research and industry purposes.
Figure 2.
One group of closed growth systems is bioreactors. These are systems with a variety of modules, such as agitation systems, temperature controls, and gas flow management. Thus, their ability to strictly control these parameters makes them an ideal system for microbe and plant cell line cultivation, and scaled growth. Through observation of the productivity of microbes or plant cell lines, bioreactors have assisted in the production of compounds such as fatty acids, pigments, and biohydrogen. Some of which have been used to synthesize products such as fish feed and biofuel. Furthermore, the growth of microbes in bioreactors, such as microalgae, has been used to study processes like wastewater remediation. All these applications have contributed to identifying the diverse functions of bioreactors for both research and industry purposes.
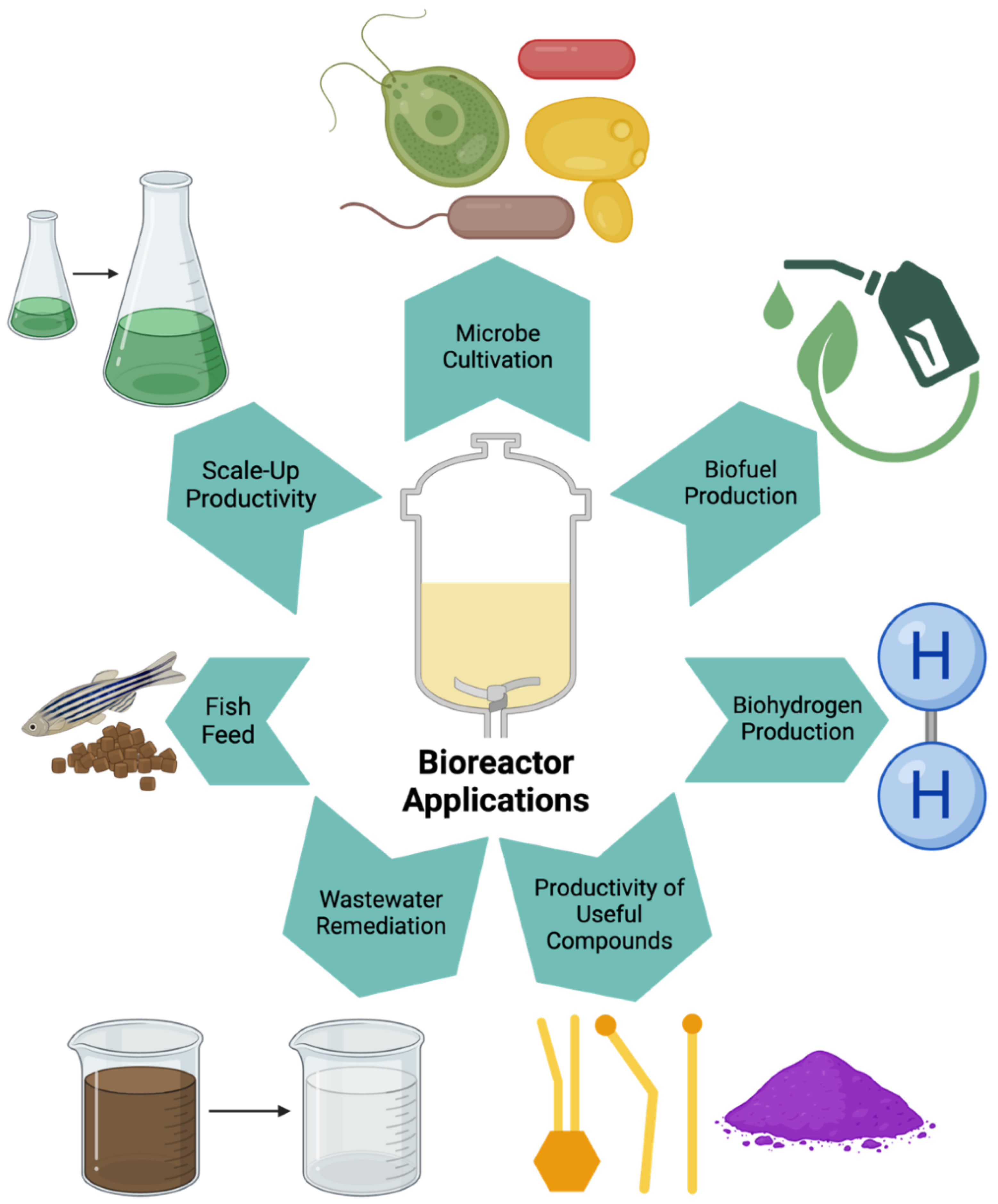
Figure 3.
Photobioreactors (PBRs) can control a variety of growth parameters necessary for microalgae cultivation. Thus, these systems have been utilized to identify ideal microalgae feedstocks for biofuel production, as specific ratios of certain fatty acid types are ideal for biofuel. Furthermore, the production of biofuel must eventually be performed on a larger scale. This has encouraged the design of PBRs for scaling-up productivity. Thus, PBRs have also contributed to the production and improvement of microalgae-based biofuel.
Figure 3.
Photobioreactors (PBRs) can control a variety of growth parameters necessary for microalgae cultivation. Thus, these systems have been utilized to identify ideal microalgae feedstocks for biofuel production, as specific ratios of certain fatty acid types are ideal for biofuel. Furthermore, the production of biofuel must eventually be performed on a larger scale. This has encouraged the design of PBRs for scaling-up productivity. Thus, PBRs have also contributed to the production and improvement of microalgae-based biofuel.
Figure 4.
Photobioreactor (PBR) designs can be tailored to microalgae growth conditions. These incubation factors exist to ensure microalgae cultured in PBRs were grown in the desired environment for specific outcomes, such as increased productivity rates. Growth factors include salinity, gas exchange, pH, nutrient sources, mixing, light, and temperature. Where varied conditions can result in microalgae stress, optimal growth, or even the destruction of the culture.
Figure 4.
Photobioreactor (PBR) designs can be tailored to microalgae growth conditions. These incubation factors exist to ensure microalgae cultured in PBRs were grown in the desired environment for specific outcomes, such as increased productivity rates. Growth factors include salinity, gas exchange, pH, nutrient sources, mixing, light, and temperature. Where varied conditions can result in microalgae stress, optimal growth, or even the destruction of the culture.
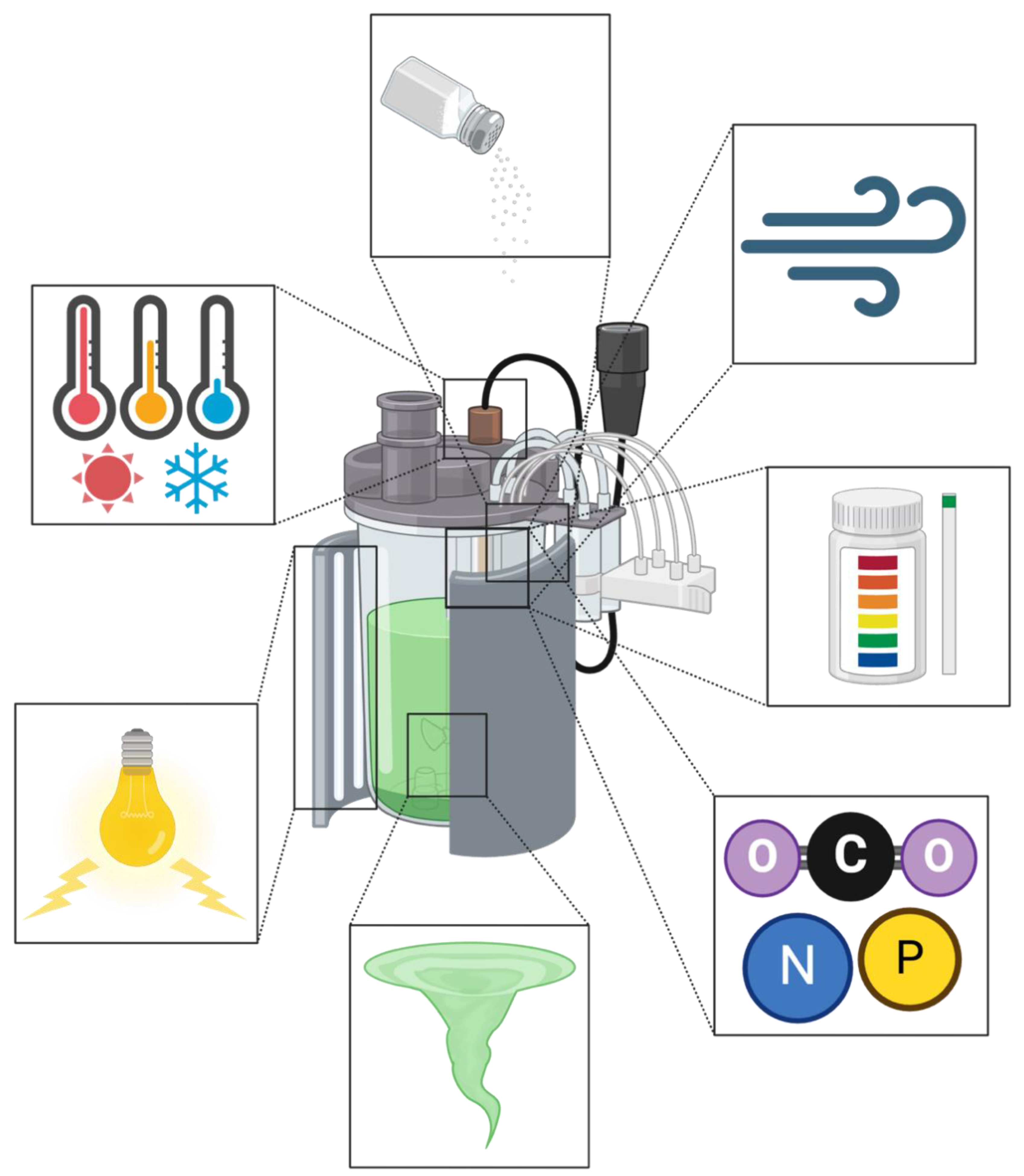

Table 1.
The diverse uses of bioreactors.
| Category of Use | Type of Bioreactor | Species Cultivated | Volume | Reference |
|---|---|---|---|---|
| Biohydrogen production | Tubular PBR | Chlamydomonas reinhardtii strain CC124 | 0.004 m3 | [36] |
| Panel PBR | 0.004 m3 | |||
| Fishmeal alternative production | Vertical tubular-type PBR | Chlorella vulgaris FSP-E | 50 L | [31] |
| Biofuel feedstock cultivation and wastewater remediation | Floating offshore PBR | Scenedesmus spp., Chlorella spp., Cryptomonas spp., Micractinium spp., Desmodesmus spp., Chlamydomonas spp., Euglena spp., Pandorina spp., Coelastrum spp., and Geitlerinema spp. | 4.18-20.91 m3 | [37] |
| Polyhydroxyalkanoate productivity in rice winery wastewater | Sequencing batch reactors | Zoogloea | 3L | [38] |
| Scale-up productivity | Fibonacci-type photobioreactor (PBR) | Dunaliella salina | 1250 L | [35] |
| Tubular PBR | Tetraselmis sp. CTP4 | 35 m3 | [39] | |
| 100 m3 | ||||
| 8 m2 thin-layer cascade PBR | Microchloropsis salina | 55 L | [40] | |
| 50 m2 thin-layer cascade PBR | 330 L | |||
| Bioreactor | Saccharomyces cerevisiae | 5 L | [41] | |
| Photobioreactor | Chlorella vulgaris | 100 m3 | [33] | |
| Pilot-scale flat-plate PBR | Chlamydomonas reinhardtii | 120 L | [42] | |
| Cultivation | Tubular reactor | Scenedesmus almeriensis | 3 m3 | [43] |
| Raceway reactor | 20 m3 | |||
| 4 m3 | ||||
| Thin-layer reactor | 1.5 m3 | |||
| Recombinant bacteria cultivation | Bioreactor | Streptococcus equi subsp. zooepidemicus | 3 L | [44] |
| Plant cell line cultivation | Bioreactor | red carrot R4G cell line | 50 L | [45] |
| Cultivation and productivity | Bioreactor | Aurantiochytrium sp. T66 | 1 L | [46] |
| Productivity | Tubular PBR | Chlorella vulgaris | 100 L | [34] |
| Fatty acid productivity | Plastic-type flat panel PBR | Scenedesmus obliquus | 5 L | [47] |
| CO2 biofixation and biofuel productivity | Air-lift PBR | Coelastrum sp. SM | 3.26 L | [48] |
| Lipid Productivity | Flat-plate PBR | Nannochloropsis sp. KMMCC 290 | 5 L | [1] |
| Bubble column PBR | ||||
| Air-lift PBR |
Table 2.
Summarization of challenges faced by current photobioreactors.
| Purpose | PBR Type | Challenge(s) | Reference(s) |
|---|---|---|---|
| Research | Fibonacci-type PBR | Smaller scaled versions of this design decreases illuminated surface area and increases the ratio of space to culture volume. |
[35] |
| Industry | Fibonacci-type PBR | Productivity varies with strains’ light requirements as large scale outside designs’ light source is solar. | [35] |
| Floating offshore PBR | In listed reference, this design was utilized to cultivate polyculture, thus replicability of data is uncertain. Additionally, relayed low lipid productivity rates. |
[37] | |
| Tubular PBR | For outdoor models, growth of culture is dependent on season. | [39] | |
| High lag phase/time compared to panel PBR. Higher pressure accumulation may result in lower productivity compared to panel PBR. | [36] | ||
| Vertical tubular-type PBR | This design can generate high shear stress. High aeration rate is not viable for large-scale growth. |
[31] | |
| Pilot-scale flat-plate PBR | This design can generate high shear stress. | [42] | |
| Research & Industry | Stir tank PBR | This design can generate high shear stress. | [1,52] |
| Horizontal tubular PBR | This design requires more space. Challenges also include gas transfer and heat transfer. |
[1,52] |
Table 3.
The incubation and growth factors for microalgae cultivation.
| Microalgae Species | Growth Factor Type | Growth Requirements |
Productivity | Reference | |
|---|---|---|---|---|---|
| Ankistrodesmus braunii | Salinity | 50 mM NaCl | After six days of cultivation, lipid content reached 34.4% dry weight | [23] | |
| Ankistrodesmus falcatus | Salinity | 100 mM NaCl | After 10 days of cultivation, lipid content reached 53% dry weight | ||
| Chaetoceros sp. FIKU035 | Temperature | 25 °C | Growth rate reached approximately 0.537 1/d | [54] | |
| Lipid productivity reached approximately 66.73 mg/L*d | |||||
| 30 °C | Biomass was about 777.93 mg/L and biomass productivity was approximately 388.97 mg/L*d | ||||
| Lipid productivity reached approximately 61.35 mg/L*d | |||||
| Chlamydomonas reinhardtii mutant | Gas Exchange | Airflow rate of 5.0-7.5 L/min | Increases biomass concentration by 18% | [42] | |
| Chlamydomonas reinhardtii strain CC124 | pH | 7.65 | Biomass productivity was approximately 31.8 mg/L*h in a tubular photobioreactor (PBR) | [36] | |
| Light Intensity | At tube: 150 µE/m2s At tank: 400 µE/m2s |
||||
| pH | 7.8 | Biohydrogen productivity was about 1.3 mL/L*h in a panel PBR | |||
| Light Intensity | 150 µE/m2s | ||||
| Chlorella sorokiniana DOE1412 | pH | 6 | Biomass productivity was approximately 0.140 g/L*day | [18] | |
| 7 | Average growth rate was approximately 0.353 g/L*day | ||||
| Chlorella sp. GN1 | Light Intensity | 5 cm light path, supplying higher light intensity than larger light paths | Lipid content was 53.5% | [55] | |
| Nitrogen Supply | 0.8 g/L urea, nitrogen concentration of approximately 3 mM | Biomass productivity rate was about 345 mg/L*d | |||
| Nitrogen Supply | Nitrogen deprived conditions: 0.01 g/L urea in growth medium | Lipid productivity was 63.5 mg/L*day | |||
| Lipid concentration comprised of 48.65% cells’ dry weight | |||||
| Phosphorous Supply | Phosphorous deprived conditions: 0.001 g/L K2HPO4*3H2O in growth medium | Lipid concentration comprised of 36.28% cells’ dry weight | |||
| Chlorella vulgaris | Light Intensity | 150 µE/m2s | After 8 days of cultivation, biomass productivity was 0.6 g/L | [22] | |
| CO2 Source | Sodium bicarbonate | Increase of lipid concentrations approximately 26% | [34] | ||
| CO2 fixation rate was about 0.925 g/L*d | |||||
| Temperature | 25 °C | Biomass reached 1.52 g/L | [25] | ||
| pH | 8.0 | ||||
| Salinity | 30 PSU | ||||
| Light | Blue light at 499-465 nm | ||||
| Chlorella vulgaris FSP-E | Nitrogen Supply | 18.6 mM urea concentration | Biomass productivity reached 268.1 mg/L/d and protein was produced at a rate of 155.4 mg/L/d | [31] | |
| Aeration Rate | 0.05 vvm | ||||
| Chlorococcum sp. | Light Intensity | 2500-3500 lux | After five days of cultivation, optimal growth rate was achieved | [56] | |
| Growth Medium | Saline water as water source for growth medium | After five days of cultivation, optimal growth rate was reached 323*104 cells/mL | |||
| Light Cycle | 24-hour light period | After nine days of cultivation, optimal growth rate was achieved | |||
| Initial Cell Density | - | Ideal cell density is dependent on growth conditions | |||
| Coelastrum sp. SM | CO2 Supply | 12% | Biomass productivity reached 0.267 g/L*d and CO2 biofixation rate was 0.302 g/L*h Lipid content was 37.91% of cell dry weight and carbohydrate content reached 58.45% of cell dry weight |
[48] | |
| Air flow | Approximately 0.06 VVM | ||||
| Light Intensity | 6900 lux | ||||
| Light Cycle | 12-hour light period, 12-hour dark period | ||||
| Desmodesmus sp. | Light Intensity data |
300 µE/m2s data |
After 8 days of cultivation, biomass productivity was 1.1 g/L | [22] | |
| After 15 days of cultivation, biomass productivity was 1.4 g/L | |||||
| After 8 days of cultivation, fatty acid content increased to 6.2% | |||||
| Dunaliella salina | Light Intensity | 600-995 μE/m2s | Biomass concentration and productivity was 0.96 g/L and 0.12 g/L*d, respectively | [35] | |
| Temperature | 18.2-22.5 °C | ||||
| pH | 7.5-8.5 | ||||
| Isochrysis galbana | Light Intensity | 350 μmol/m2s | Carbohydrate production of 48.11 gC/m3d | [57] | |
| Temperature | 14 °C | After 10 days of cultivation, docosahexaenoic acid (DHA) content was 19.55 mg/g of ash-free dry weight | [20] | ||
| After five days of cultivation, DHA productivity was 1.08 mg/L*d | |||||
| Nannochloropsis gaditana | Light Intensity | 360 µmol photons/(m2s) | Three-fold increase in growth compared to low light conditions | [21] | |
| Nannochloropsis oculata | Temperature | 20 °C | After five days of cultivation, eicosapentaenoic acid (EPA) productivity was 2.52 mg/L*d | [20] | |
| Nannochloropsis QU130 | Light Cycle | 24-hour light period at 500 µmolhν/m2s | Biomass productivity increased by 13.6%, to 33 g/m2*d | [15] | |
| Temperature | Fluctuating temperatures between 32-41 °C | ||||
| Larger cell size than continuous temperature conditions | |||||
| Nannochloropsis salina | Light Cycle | 24-hour light period | Growth rate reached 0.42 1/d | [13] | |
| Biomass concentration was about 0.77 g/L | |||||
| Gas Exchange | 1L/h of 5% CO2 supplemented air | After 10 days of cultivation, stationary phase was reached | |||
| Nitrogen Supply | Nitrogen-deprived medium with 0.075 g/L NaNO3 | Lipid concentration was 63% of cells’ dry weight | |||
| 1.5 g/L NaNO3 | Growth rate increased approximately 3.5-fold | ||||
| Gas Exchange | 5% CO2 supplemented air | ||||
| pH | 8 | Growth rate approximately 0.19 | [17] | ||
| Largest cell density after 21 days of cultivation was about 95.6*106 cells/mL | |||||
| 9 | Growth rate approximately 0.19 | ||||
| Largest cell density after 21 days of cultivation was about 92.8*106 cells/mL | |||||
| Nannochloropsis sp. FIKU036 | Temperature | 25 °C | Growth rate reached approximately 0.3311/d | [54] | |
| Biomass was about 885.35 mg/L and biomass productivity was approximately 293.05 mg/L*d | |||||
| Nannochloropsis sp. KMMCC 290 | Light Intensity | 11,600 lux | Cell concentration increased by 50%, with a final concentration of 0.51 g/L in air-lift photobioreactor (ALP) | [1] | |
| Lipid productivity increased by 47.7%, to 13.4*10-3 g/L/d in ALP | |||||
| Lipid productivity increased by 45.7%, to 18.8*10-3 g/L/d in flat-plate photobioreactor (FPP) | |||||
| Aeration Rate | 1.0 vvm or 5.0 L/min | ||||
| Cell concentration increased by 44.1%, with a final concentration of 0.49 g/L in ALP | |||||
| Lipid productivity was 13.4*10-3 g/L/d in ALP | |||||
| CO2 Feeding | 10% CO2 at 0.5 L/min for 2 hr every 12 hr intervals | Final cell concentration was 0.65 g/L for FPP | |||
| Lipid productivity reached 19.8*10-3 g/L/d for ALP and FPP | |||||
| Lipid content was 31.5% for ALP | |||||
| Phaeodactylum tricornutum | Nutrient Supply/Growth Medium | 8.82 mM nitrogen concentration in f/2 growth medium | After 10 days of cultivation, biomass concentration reached about 2.76 g/L | [4] | |
| Fucoxanthin content was around 2.18 mg/g of fresh weight | |||||
| Fucoxanthin productivity reached about 5.07 mg/L/d | |||||
| Increased fucoxanthin production to approximately 9.82 mg/L/d | |||||
| Light Intensity | 20 µmol/m2/s | ||||
| Porphyridium sp. | Air Flow | 0.16 cm/s | Dry biomass concentration reached approximately 5 g/L | [58] | |
| Scenedesmus incrassatulus | Salinity | 100 mM NaCl | After six days of cultivation, lipid content reached 37.7% dry weight | [23] | |
| Scenedesmus obliquus | Light Intensity |
150 µE/m2s | After 8 days of cultivation, biomass productivity was 0.8 g/L | [22] | |
| 300 µE/m2s | After 15 days of cultivation, biomass productivity was 1.2 g/L | ||||
| After 15 days of cultivation, fatty acid content increased to 11.6% | |||||
| Nitrogen Supply | Nitrogen source was urea | Cells’ dry biomass was composed of 40% lipids | [47] | ||
| Light Intensity | 3000 lux | ||||
| Spirulina sp. | Shear Force | Decreased bubble size (1.8 mm) and formation time (3.3 ms) in volute aerator | Average growth rate increased by 26.6% in comparison to using a strip aerator | [59] | |
| Biomass productivity increased by 50.7% in comparison to using a strip aerator | |||||
| Tetradesmus almeriensis | Nutrient Supply/Growth Medium | Freshwater with fertilizer | Biomass productivity was 30.3 g/m2*day | [60] | |
| Tetraselmis suecica FIKU032 | Temperature | 30 °C | Growth rate reached approximately 0.378 1/d | [54] | |
| Biomass was about 978.43 mg/L and biomass productivity was approximately 369.84 mg/L*d | |||||
| Tisochrysis lutea | Air Flow | 6.25 vvm | Specific net growth rate reached 3.8 L/min | [61] | |
| Xanthonema hormidioides | Temperature | 20 °C | After three days of cultivation, biomass productivity was 11.73 g/L | [24] | |
| Nitrogen Supply | 18 mM nitrogen concentration | ||||
| Temperature | 25 °C | After 18 days of cultivation, lipid content was 57.49% of cells’ dry weight | |||
| Nitrogen Supply | 3 mM nitrogen concentration | ||||
| Mixed microalgae culture sourced from the Nacharam Cheruvu in India | Temperature | 30 °C | Increase in total lipid productivity to 24.5% | [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



