Submitted:
29 August 2024
Posted:
02 September 2024
You are already at the latest version
Abstract
The Aspergillus genus comprises fungi that are widely distributed in nature. The Terrei section is particularly important due to its functions in recycling organic matter. Aspergillus hortae is a species from this section that has been isolated from clinical samples, but its role as a pathogenic agent is unclear. In this study, three clinical A. hortae isolates from Colombia, initially identified as A. terreus, were characterised by micro and macro-morphological analysis at 26°C and 37°C. A bioinformatic pipeline was used for molecular typing, which included analysis of whole genome sequences for accurate species identification. Susceptibility assays were also conducted using the microdilution broth method (EUCAST). It was confirmed that this species is intrinsically resistant to Amphotericin B with a minimum inhibitory concentration (MIC) of 2-4 mg/L, and is also thermotolerant. We obtained the in vitro susceptibility profiles for three azole drugs (MIC, 0.25-1 mg/L). The three isolates showed phylogenetic divergence from the reference genome strain. A mutation (M769K) in the MelA ortholog of a hypopigmented isolate, which had the lowest MIC value for AmB (2 mg/L), was identified. This study presents the morphological characterization, molecular typing through whole-genome analysis and identification of susceptibility profiles to azoles and amphotericin B of three clinical isolates of A. hortae from Colombia.
Keywords:
Aspergillus hortae
; Genome
; Pathogenic fungi
; antifungals susceptibility
1. Introduction
Aspergillus section Terrei represents an important taxonomic group of the Aspergillus genus due to its biological diversity and functions in natural ecosystems. Species belonging to section Terrei are saprophytic, frequently occurring in soil and organic matter [1]. Furthermore, they are utilized in the industry as producers of secondary metabolites, including drugs and other bioactive compounds [2,3]. However, some isolates from this section are often isolated in clinical samples and linked with diseases in both human and animals [4]. One of the species described within the section Terrei is Aspergillus hortae, which is phylogenetically close to A. terreus [5]. A. hortae has been briefly mentioned in the literature as a human pathogenic species [4,6] and has been isolated from clinical samples such as ear secretions [5]. Currently, knowledge about the diversity of this fungus remains limited. As an organism with pathogenic capacity, it is necessary to acquire additional knowledge regarding its phenotypic and genotypic characteristics, as well as its geographical distribution.
The wide genotypic diversity present within the Aspergillus genus makes the section/species that the patient is exposed to a key parameter when selecting an appropriate antifungal treatment. This is because Aspergillus spp. do not have homogeneous susceptibility profiles [7]. In the section Terrei, for example, an intrinsic resistance to amphotericin B has been reported [4]. Nonetheless, the molecular and evolutionary mechanisms behind this resistance remain unclear. The issue of resistance to antifungal treatment is on the rise, with azoles in particular posing a worldwide challenge [8,9]. It is urgent to characterise the pathogenic species of the genus Aspergillus and understand their susceptibility profiles to different antifungal drugs.
Currently, whole-genome sequence approaches have expanded the knowledge about the genetic characteristics of pathogens [10]. These approaches enable simultaneous species identification, prediction of drug resistance, and discovery of genetic variants [11,12]. Despite the enormous risk that antifungal-resistant isolates from the genus Aspergillus can pose to human health, whole-genome sequences of this genus are still limited, particularly in non-fumigatus Aspergillus species [11]. The aim of this study was to provide a morphological characterization, perform molecular typing through whole-genome analysis, and identify susceptibility profiles to azoles and amphotericin B of three clinical isolates of A. hortae from Colombia.
2. Materials and Methods
Fungal isolates: Three clinical isolates of Aspergillus section Terrei were obtained from the Fungi Collection at the Corporación para Investigaciones Biológicas (CIB, Medellin; Colombia). Upon receiving the isolates, they were identified at the species level by microscopic identification technique based on morphological features [1]. These isolates were initially identified and reported as A. terreus, and were named as MC7, MC8, and MC10. Cottony colonies with a beige to cinnamon brown colour were observed with. Prior to this work, in Colombia A. terreus was the only species within the section Terrei reported in clinical samples.
Phenotypic characterization: After obtaining a pure culture, the isolates were inoculated at three points using a micropipette and an inoculum size of 1 μl per spot into Petri dishes containing Malt Extract Agar (MEA), Potato Dextrose Agar (PDA), Sabouraud Agar (SB) incubated at 25°C, and other dishes with Czapek Yeast Agar (CYA) , were incubated at 26°C and 37°C. After 7 days of incubation, the macroscopic characters of the colonies on each medium and the microscopic characteristics on MEA were evaluated. Macroscopic characteristics including colony diameters and texture, obverse and reverse colony colours, degree of sporulation, production of soluble pigments, and exudates were determined. After 10 days of incubation, microscopic characters such as the shape of conidial heads, the presence or absence of metulae between vesicle and phialides, colour of stipes, and the dimension, shape, and texture of stipes, vesicles, metulae, phialides, and conidia were evaluated. Each isolate was morphologically characterised using standardised and recommended methods for laboratories working with Aspergillus spp., as described by Samson et al. (2014) [1]. Photographic records of the colonies were obtained using a digital camera, and images of the microscopic structures were captured using the Leica ICC50 W microscope. The colony diameters were analysed in SPSS 24 software through a Klustal-Wallis test.
Antifungal Susceptibility Test: The antifungal susceptibility test was carried out via a 2X microdilution broth method following EUCAST instructions [13]. The antifungals evaluated in this study were Amphotericin B (AmB), Itraconazole (ITC), Voriconazole (VRC), as well as Posaconazole (POS). To test antifungal susceptibility, a suspension of each isolate was prepared in RPMI 1640 broth medium. The four antifungals mentioned above were diluted in dimethyl sulfoxide (DMSO). The strains were tested against 10 concentrations (0.03-8 mg/L) in 96-well plates by 2x dilution and incubated at 35°C for 48 h.
DNA Extraction: Isolates were cultured in Brain Heart Infusion (BHI) at 30°C, at 120 rpm. The biomass was collected during the exponential growth phase after 96 hours of incubation. Cell lysis was performed by mechanical disruption using liquid nitrogen. Genomic DNA was obtained using phenol/chloroform extraction, and RNA was eliminated by treatment with 10 μg of RNase A (Thermo Fisher Scientific, USA) for 120 min at 37°C [14]. DNA quality was evaluated using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA) to determine concentration and purity, using the default setting (1 OD = 50 mg/mL dsDNA). In addition, the integrity of the DNA recovered from isolates was evaluated through 1% agarose gel electrophoresis.
Whole Genome Sequencing: Library preparation was carried out using Illumina Nextera DNA Library Preparation Kit (Illumina Inc. San Diego, CA, USA), with 500 ng of DNA per sample. Next Generation Sequencing was performed using the Illumina Xten (Inc. BGI Hong Kong), generating 150-bp paired-end sequencing. The samples were run on one sequencing line on the Illumina platform, generating around 7 million paired-end reads per isolate and producing an average genome coverage of 30X. Raw data is available on the Sequence Read Archive (SRA) website from National Center for Biotechnology Information (NCBI) (Bioproject PRJNA975750), Biosamples codes SAMN35344990, SAMN35344991, SAMN35344992.
Pre-assembly Analysis: FastQC v0.11.8 program was used to analyse the FASTQ quality code of the short paired-end raw reads [15], Trimmomatic v0.39 was employed to filter out low-quality (Q<30) sequences and adapters [16].
Genome Assembly: The SPAdes 3.10 software [17] with the BayesHammer module for error correction [18] was used to process data. De novo assembly of the short reads (2x150) was performed, and iterative k-mer lengths (21, 33, 55, 77 bp) were used to take advantage of the paired-end reads. The draft genome assembly's quality was evaluated using QUAST 5.2.0 [19], comparing the metrics with representative genomes. Three parameters were utilized to verify the quality of each genome: the average coverage of paired-end reads, histograms of the distribution of the percentage of Guanine-Cytosine (% GC), and sequence alignments using the genes of the A. hortae IBT 26384 as a reference. The genomes available were downloaded from the JGI MycoCosm database (https://genome.jgi.doe.gov/programs/fungi/index.jsf).
Gene homology Analysis: The Aspergillus terreus model was used for ab initio gene prediction using Augustus v3.0.1 [20]. The predicted protein sets were then compared using OrthoFinder v2.0.9 pipeline [21] to analyse sequence homology with the representative genomes of Aspergillus section Terrei reported in the databases (Table S1).
Species Identification by barcoding: To identify the species of Aspergillus isolates, ITS, CaM, and BenA sequences were identified in the assembly using a local blastn search. The query reference sequences from GenBank®, A. terreus BenA (EF669520.1), A. hortae CaM (KP987054.1), and A. hortae ITS (OL711861.1), were used. A web blastn search was then performed on these sequences in the Nucleotide Collection (nr/nt) of NCBI and EMBL-bank databases with default settings (Figure S1).
Phylogenetic Analysis: The resulting sequences were aligned with reference sequences from Aspergillus section Terrei (Table S1) using the ClustalW v-2.1 program. For phylogenetic reconstruction, the Maximum Likelihood (ML) method [22] was employed with IQtree v1.4.4 software, and the best nucleotide substitution model was estimated. Phylogenetic analysis was conducted using individual genes and a concatenated matrix with BenA and CaM markers. The Bootstrap method with 1000 replications was used to evaluate the internal branches.
Whole-genome single-nucleotide polymorphism (SNP) calling and phylogenetic analysis of A. terreus clade
We download and available Illumina reads of species from this clade (SRA NCBI database) (Table S2). Each of the 10 Illumina data sets was independently aligned to the A. terreus reference genome using BWA version 0.5.9 [23] with default settings. SNPs and indels were called with Pilon version 1.4 using the haploid ploidy default setting. Variant call format (VCF) files were filtered using VCFtools version 0.1.1 (minimum depth 4). Alignments were constructed from SNP matrices extracted from the VCF files.
3. Results
3.1. Morphological Analysis
The colonies grown on CYA and MEA culture media at 26°C exhibit compact, columnar-shaped heads. The conidiophores are short, hyaline, and have smooth walls. A. hortae vesicles take the form of half-heads with extensive cylindrical metulae, from which the phialides emerge. The conidia are round, hyaline, and have smooth walls. When grown on CYA and MEA culture media at 37°C, the colonies display a dense cottony appearance with a beige to cinnamon brown colour. As the colony matures, a dark center with a lighter periphery becomes evident. The isolated MCA7 strain shows lighter colony pigmentation and a lower minimum inhibitory concentration (MIC). These morphological characteristics are consistent with what has been described in the literature for Aspergillus section Terrei (Figure 1).
After incubating the colonies at both 26°C and 37°C for five days in darkness , it was observed higher biomass production in all cultures at 37°C. The cultures grown at 26°C had a lower colony diameter (56.67 ± 5.60 mm), with significantly better growth (p<0.05) in all evaluated media at 37°C (86.92 ± 9.07 mm). Additionally, the highest colony diameter was observed in CYA at 37°C (98.7 ± 10.69 mm) while the lowest was observed in MEA at 26°C (51.67 ± 3.51 mm) (Figure 2).
3.2. Antifungal Susceptibility Testing
The three A. hortae isolates exhibited high MIC values upon AmB exposure (2-4 mg/L), with MCA7 displaying the lowest MIC value (2 mg/L), whereas the other isolates showed twice the MIC value (4 mg/L). In contrast, we observed the lowest MIC values with POS (0.25 mg/L), which were consistent across all three isolates. Similarly, the MIC values with ITC were also consistent at 1 mg/L. However, the MIC values with VRC were varied, with two isolates (MCA7 and MCA8) demonstrating a result of 0.5 mg/L and MCA10 having twice this value (1 mg/L) (Table 1).
3.3. Genome Assembly and Analysis
To compare genome structures and gene contents between Aspergilli, we sequenced, assembled, and predicted the proteins of the three A. hortae isolates included in this study. In the assemblies, we found characteristics like those reported for the A. hortae reference genome, including a genome size ranging from 29.86 - 31.90 Mb and a GC content of 52.05 - 52.19% (Table S1). The ab initio predictions showed a range of 13,120 - 13,300 predicted protein-coding genes, with a high proportion (>99%) of homologs found in the genus Aspergillus.
Using the local blastn tool, the sequences of the markers (ITS, CaM, BenA) were found within the assemblies, which are commonly used for phylogenetic analysis and identification of the Aspergillus genus [1]. Only one copy of these genes was found in each of the assemblies. Subsequently, a web search was conducted to compare these sequences with the databases, leading to the successful identification of the species as A. hortae/A. terreus. The best hit with BenA was with A. hortae species (100% identity and 99.81% coverage), while with ITS and CaM, we obtained similar hits for A. terreus species as well. The 99.81% coverage is for the gene. This result showed that the isolates had a high probability of being classified as A. hortae, and it was confirmed after phylogenetic analysis.
The alignment of three de novo assemblies from Colombian isolates to the reference genome (Asphor1_AssemblyScaffolds) showed conserved synteny when comparing the Locally Collinear Blocks (LCB) generated in mauve. However, in the alignment, LBC in the reverse panel (R) is observed in the three new assemblies, and these are relatively conserved in them, mainly in MCA8 and MCA10 isolates (Figure 2). After homology analysis using Orthofinder, we identified the orthologs associated with melanin production in A. terreus, MelA (XP_001212741), and TyrP (XP_001212742.1). These two proteins are conserved in the Terrei and Flavipedes sections, and TyrP orthologs were found in almost all evaluated species, while MelA orthologs were found only in 13 species (Figure 2A). MelA possesses a conserved Thioesterase domain of type I polyketide synthase (EntF), however, during protein alignment, a K769M point mutation was observed in the hypopigmented isolate MCA7. The Methionine at position 769 is conserved in all species, while MCA7 is the only isolate with a change in this amino acid (Figure 2B) (Figure 3 #hypopigmented isolated).
When the whole genome is aligned to A. hortae, TyrP and MelA are located contiguously within a LCB in the alignment of the four A. hortae genome assemblies. However, the two highly pigmented isolates (MCA8 and MCA10) showed the LBC with melanin-associated genes in an inverted orientation (Figure 2C). Finally, we zoomed into the MelA locus, where the nucleotide substitution t2307a (XM_001212741.1) was found in the MCA7 isolate.
3.4. Phylogenetic Analysis
Our best phylogenetic reconstruction was achieved using concatenated sequences of BenA and CaM in a partitioned matrix of 59 OTUs (Table S4) and 1344 characters. Modelfinder was used to determine the best nucleotide substitution model for each partition (Table S2). The three isolates were grouped with reference isolates from A. hortae, creating a monophyletic clade (Bs=88%) with IBT 6271, IBT 6271, HEGP06, PSL01, and SAT02. The reference genome strain IBT 26384 is observed as an outgroup in another monophyletic clade (Bs=94%) with strains IBT 16744 and CMV004A9. The A. hortae species is closely related to the clade formed by A. terreus and A. citrinoterreus species (Figure 4). The phylogenetic reconstruction using sequences from ITS marker, does not provide clear genotypic differentiation between A. hortae and other species from Aspergillus section Terrei (Supplementary Figure S2). In the phylogenetic tree of the clade A. terreus, the three Colombian isolates of A. hortae cluster with the reference strain IBT 26384 and are positioned as a sister species to A. terreus, with A. pseudoterreus serving as the outgroup.
4. Discussion
In this study, we described the first report of susceptibility profiles of A. hortae isolated in Colombia. Upon analysing the MIC of the isolates MCA-7, MCA-8 and MCA-10, and comparing them with the cut-off established in EUCAST, we confirmed that these isolates exhibit intrinsic resistance to AmB. As previously reported. Isolates from different species of Aspergillus section Terrei, including A. hortae, display a high tolerance to this antifungal in vitro. Consequently, patients do not respond when tAmB is employed to treat Aspergillosis caused by members of this section [4,24]. However, the AmB susceptibility profiles of the isolates were not uniform; MCA7 exhibited higher susceptibility to AmB than the other two isolates. Imbert et al, in 2018 also reported two A. hortae French isolates with low AmB MIC values (0.25 mg/L) [6], indicating genetic variability within this species.
The AmB resistant phenotype has been linked to changes in the cell membrane of the fungus alterations in the expression of membrane transporters, or modifications in the structure of the drug target [25]. Nevertheless, the exact resistance mechanisms in these species are still unclear. Further investigation of the genomes of a high number of species from this section, as well as the phylogenetically related sections Fumigati and Flavi, whose susceptibility profiles were heterogeneous, could aid in the elucidation of this matter [26,27].
The fact that we found isolates with heterogeneous AmB resistance and pigmentation levels indicates genetic diversity in this fungal species. Interestingly, the isolate with the lowest pigmentation had the lowest MIC value with AmB, and harboured a mutation in a melanin pathway gene. In Cryptococcus spp., high melanin pigmentation isolates are associated with decreasing AmB susceptibility [28]. Similarly, in W. dermatitidis, strains with PSK1 gene knocked out displayed increased susceptibility to both AmB and VRC [29]. This suggests that AmB susceptibility in A. hortae could also be associated with melanin production. Geib et al, 2016 demonstrated in A. terreus that the pigment produced by MelA and TyrP protects conidia from biotic and abiotic stress factors [30]. However, the role of the melanin in the AmB resistance was not analysed. A potential area for further research is an investigation into the susceptibility of MelA and TyrP knockout strains from section Terrei to AmB susceptibility. This study could help to understand the importance of this pigment in the antifungal resistance.
The three isolates were susceptible to the three evaluated azole drugs (ITC, POS, VRC). ITC and POS showed the lowest MIC values, in agreement with previous studies on this fungal species [8,24]. Meanwhile, VRC displayed a heterogeneous profile. This patron could be considered as a caution sign. Despite the fact that all isolates evaluated are classified as susceptible to this drug, its VRC resistance has been observed in other species that exhibit the same profile [8]. The isolated (MCA10) with VRC higher MIC value also had differences in the LCB profiles with respect to the other isolates (MCA7 and MCA8), when their draft assemblies were aligned to the reference genome. Intra-chromosomal rearrangements are a molecular mechanism recently associated with antifungal resistance evolution in C. auris [31].
Here, we reported three new genome assemblies from clinical isolates of A. hortae, which exhibit significant phylogenetic divergence from the reference genome strain (IBT 26384, a clinical isolate from Brazil). It is noteworthy that this is the first time that genomic variability has been found in this Aspergillus species. Steenwyk et al. (2022), using WGS analysis, demonstrated an extensive misidentification of species and low novel lineages detection in the genus Aspergillus when only a few barcode markers were used [23]. Despite the phenotypic differences, the phylogenetic reconstruction has revealed that the three isolates are clonal. We suggest that these phenotypic differences may be due to a potential variation in chromosome structure in A. hortae, although the finding requires analysis using long-read sequencing technologies that have enhanced precision and resolution in the identification of structural variations [32].
At the moment, the whole genome data for this species continues to be limited for phylogenomic reconstruction to examine the genetic diversity within this species. The species tree generated after homology analysis from section Terrei only showed that the three Colombian isolates are related to the only available A. hortae reference genome, whereas the BenA-CaM phylogeny with 10 isolates showed evidence of genetic divergence from this isolate. Further research with more isolates sequenced at genome level could be conducted to elucidate the true evolutive history and genetic diversity of A. hortae.
On the other hand, the ITS barcode marker has been used as a panfungal molecular test in the diagnostic laboratory [33,34]; however, it showed very low-resolution power in the A. terreus/A. hortae/A. citrinoterreus clade. It may have overestimated the epidemiology of A. terreus impeded our understanding of the actual impact on health of new cryptic species, such as A. hortae. The epidemiology of Aspergillosis has changed in recent years, with the emergence of new species, which could be attributed more to the improvement of typing techniques that have allowed for accurate classification [23,35]. Salem-Bango et al. (2023) propose the use of WGS in specialized laboratories to accurately identify Aspergillus species [12]. We agree that the adoption of WGS could be an effective solution for correct species identification in Aspergillus Section Terrei, and we also suggest using at least the BenA as a second barcode marker for more precise identification of species within the section.
A. hortae is a thermotolerant fungal species, a critical characteristic for microbial pathogens to thrive in human and animal hosts. This characteristic demonstrates how this species has evolved to adapt to the stress of growth in the host [36,37], and is also linked to virulence factors in other fungal pathogens, particularly in A. fumigatus [38], which tolerates up to 60°C, the upper temperature limit for eukaryotic organisms [39]. Lacker et al. (2019) also demonstrated that A. hortae at 37 °C exhibited the highest growth rates, and some isolates had high virulence potential in G. mellonella larvae [40]. These findings indicate that A. hortae could be an emergent human pathogenic fungal species. Furthermore, there is significant phenotypic and genetic variability among the isolates of this species, requiring further exploration.
5. Conclusions
This study presents the whole genome sequences, phenotypic characteristics, and susceptibility profiles to antifungals of three thermotolerant clinical isolates of A. hortae in Colombia, representing the country's first such documentation. Such studies aim to expand our understanding of pathogens and their reactions to current drug therapies, with the ultimate goal of enabling safer and more efficient dosage strategies for patient treatment.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: Bioinformatic workflow for species identification; Figure S2: Phylogenetic tree of Aspergillus section Terrei using sequences from the ITS marker; Table S1: Reference Proteomes used in OrthoFinder; Table S2: Assembly metrics MCA-7, MCA-8 y MCA-10; Table S3: List of best-fit models per partition for phylogenetic reconstruction; Table S4: Aspergillus section Terrei SRA codes for phylogenomic reconstruction.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, O.M.G and S.M.C; methodology, O.M.G. and A.M. L.; formal analysis, S.M.C, M.Q.T., A.M. L and O.M.G ; investigation, S.M.C, M.Q.T. and O.M.G.; resources, O.H.R, A.L.R, J.G.M and A.M.G.; data curation, O.M.G, M.C.Z and S.T.; writing—original draft preparation, S.M.C, M.Q.T. and O.M.G; writing—review and editing, O.H.R, S.T., M.C.Z., J.G.M, C.L.B. and A.M.G .; visualization, X.X.; supervision, O.H.R, J.G.M, C.L.B, A.L.R. and A.M.G.; project administration, A.M.L. and O.H.R; funding acquisition, O.H.R. and A.M.G. All authors have read and agreed to the published version of the manuscript.”.
Funding
This study was financed in part by To Ministerio de Ciencia, Tecnologia e Innovación by financial support (Grant code 221384467683) from Colombia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
It was no necessary because in this work, personal information of patients was not used.
Data Availability Statement
The sequencing data will be made available when the manuscript is publicly available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, Identification and Nomenclature of the Genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- Sallam, L.A.R.; El-Refai, A.-M.H.; Hamdy, A.-H.A.; El-Minofi, H.A.; Abdel-Salam, I.S. Role of Some Fermentation Parameters on Cyclosporin A Production by a New Isolate of Aspergillus Terreus. J. Gen. Appl. Microbiol. 2003, 49, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Alberts, A.W.; Chen, J.; Kuron, G.; Hunt, V.; Huff, J.; Hoffman, C.; Rothrock, J.; Lopez, M.; Joshua, H.; Harris, E.; et al. Mevinolin: A Highly Potent Competitive Inhibitor of Hydroxymethylglutaryl-Coenzyme A Reductase and a Cholesterol-Lowering Agent. Proc. Natl. Acad. Sci. U. S. A. 1980, 77, 3957–3961. [Google Scholar] [CrossRef]
- Risslegger, B.; Zoran, T.; Lackner, M.; Aigner, M.; Sánchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. A Prospective International Aspergillus Terreus Survey: An EFISG, ISHAM and ECMM Joint Study. Clin. Microbiol. Infect. 2017, 23, 776.e1–e776.e5. [Google Scholar] [CrossRef]
- Samson, R.A.; Peterson, S.W.; Frisvad, J.C.; Varga, J. New Species in Aspergillus Section Terrei. Stud. Mycol. 2011, 69, 39–55. [Google Scholar] [CrossRef]
- Imbert, S.; Normand, A.C.; Ranque, S.; Costa, J.M.; Guitard, J.; Accoceberry, I.; Bonnal, C.; Fekkar, A.; Bourgeois, N.; Houzé, S.; et al. Species Identification and In Vitro Antifungal Susceptibility of Aspergillus Terreus Species Complex Clinical Isolates from a French Multicenter Study. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Gautier, M.; Normand, A.-C.; Ranque, S. Previously Unknown Species of Aspergillus. Clin. Microbiol. Infect. 2016, 22, 662–669. [Google Scholar] [CrossRef]
- Zoran, T.; Sartori, B.; Sappl, L.; Aigner, M.; Sánchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. Azole-Resistance in Aspergillus Terreus and Related Species: An Emerging Problem or a Rare Phenomenon? Front. Microbiol. 2018, 9, 516. [Google Scholar] [CrossRef]
- Friedman, D.Z.P.; Schwartz, I.S. Emerging Fungal Infections: New Patients, New Patterns, and New Pathogens. J Fungi (Basel) 2019, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative Genomics Reveals High Biological Diversity and Specific Adaptations in the Industrially and Medically Important Fungal Genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef]
- Nargesi, S.; Valadan, R.; Abastabar, M.; Kaboli, S.; Thekkiniath, J.; Hedayati, M.T. A Whole Genome Sequencing-Based Approach to Track down Genomic Variants in Itraconazole-Resistant Species of Aspergillus from Iran. J Fungi (Basel) 2022, 8, 1091. [Google Scholar] [CrossRef] [PubMed]
- Salem-Bango, Z.; Price, T.K.; Chan, J.L.; Chandrasekaran, S.; Garner, O.B.; Yang, S. Fungal Whole-Genome Sequencing for Species Identification: From Test Development to Clinical Utilization. J Fungi (Basel) 2023, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Subcommittee on Antifungal Susceptibility Testing of the ESCMID European Committee for Antimicrobial Susceptibility Testing EUCAST Technical Note on the Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Conidia-Forming Moulds. Clin. Microbiol. Infect. 2008, 14, 982–984.
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual. 2001, 2001. [Google Scholar]
- Andrews, S.; Lindenbaum, P.; Howard, B.; Ewels, P. FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Nikolenko, S.I.; Korobeynikov, A.I.; Alekseyev, M.A. BayesHammer: Bayesian Clustering for Error Correction in Single-Cell Sequencing. BMC Genomics 2013, 14 Suppl 1, S7. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Stanke, M.; Schöffmann, O.; Morgenstern, B.; Waack, S. Gene Prediction in Eukaryotes with a Generalized Hidden Markov Model That Uses Hints from External Sources. BMC Bioinformatics 2006, 7, 62. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Steenwyk, J.L.; Balamurugan, C.; Raja, H.A.; Gonçalves, C.; Li, N.; Martin, F.; Berman, J.; Oberlies, N.H.; Gibbons, J.G.; Goldman, G.H.; et al. Phylogenomics Reveals Extensive Misidentification of Fungal Strains from the Genus Aspergillus. bioRxiv, 2022; 2022.11.22.517304. [Google Scholar]
- Kathuria, S.; Sharma, C.; Singh, P.K.; Agarwal, P.; Agarwal, K.; Hagen, F.; Meis, J.F.; Chowdhary, A. Molecular Epidemiology and in-Vitro Antifungal Susceptibility of Aspergillus Terreus Species Complex Isolates in Delhi, India: Evidence of Genetic Diversity by Amplified Fragment Length Polymorphism and Microsatellite Typing. PLoS One 2015, 10, e0118997. [Google Scholar] [CrossRef]
- Blum, G.; Hörtnagl, C.; Jukic, E.; Erbeznik, T.; Pümpel, T.; Dietrich, H.; Nagl, M.; Speth, C.; Rambach, G.; Lass-Flörl, C. New Insight into Amphotericin B Resistance in Aspergillus Terreus. Antimicrob. Agents Chemother. 2013, 57, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Ashu, E.E.; Korfanty, G.A.; Samarasinghe, H.; Pum, N.; You, M.; Yamamura, D.; Xu, J. Widespread Amphotericin B-Resistant Strains of Aspergillus fumigatus in Hamilton, Canada. Infect. Drug Resist. 2018, 11, 1549–1555. [Google Scholar] [CrossRef] [PubMed]
- Van Der Linden, J.W.M.; Warris, A.; Verweij, P.E. Aspergillus Species Intrinsically Resistant to Antifungal Agents. Med. Mycol. 2011, 49 Suppl 1, S82–S89. [Google Scholar] [CrossRef]
- Ikeda, R.; Sugita, T.; Jacobson, E.S.; Shinoda, T. Effects of Melanin upon Susceptibility of Cryptococcus to Antifungals. Microbiol. Immunol. 2003, 47, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Paolo, W.F., Jr.; Dadachova, E.; Mandal, P.; Casadevall, A.; Szaniszlo, P.J.; Nosanchuk, J.D. Effects of Disrupting the Polyketide Synthase Gene WdPKS1 in Wangiella [Exophiala] Dermatitidis on Melanin Production and Resistance to Killing by Antifungal Compounds, Enzymatic Degradation, and Extremes in Temperature. BMC Microbiol. 2006, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Geib, E.; Gressler, M.; Viediernikova, I.; Hillmann, F.; Jacobsen, I.D.; Nietzsche, S.; Hertweck, C.; Brock, M. A Non-Canonical Melanin Biosynthesis Pathway Protects Aspergillus terreus Conidia from Environmental Stress. Cell Chem Biol 2016, 23, 587–597. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Welsh, R.M.; Shea, T.; Batra, D.; Gade, L.; Howard, D.; Rowe, L.A.; Meis, J.F.; Litvintseva, A.P.; Cuomo, C.A. Clade-Specific Chromosomal Rearrangements and Loss of Subtelomeric Adhesins in Candida auris. Genetics 2021, 218. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, S.; Cao, S.; Liu, Y.; Cui, Z.; Wang, Y.; Guo, H. Long-Read Sequencing Settings for Efficient Structural Variation Detection Based on Comprehensive Evaluation. BMC Bioinformatics 2021, 22, 552. [Google Scholar] [CrossRef]
- Muñoz-Cadavid, C.; Rudd, S.; Zaki, S.R.; Patel, M.; Moser, S.A.; Brandt, M.E.; Gómez, B.L. Improving Molecular Detection of Fungal DNA in Formalin-Fixed Paraffin-Embedded Tissues: Comparison of Five Tissue DNA Extraction Methods Using Panfungal PCR. J. Clin. Microbiol. 2010, 48, 2147–2153. [Google Scholar] [CrossRef] [PubMed]
- Valero, C.; de la Cruz-Villar, L.; Zaragoza, Ó.; Buitrago, M.J. New Panfungal Real-Time PCR Assay for Diagnosis of Invasive Fungal Infections. J. Clin. Microbiol. 2016, 54, 2910–2918. [Google Scholar] [CrossRef] [PubMed]
- Balajee, S.A.; Houbraken, J.; Verweij, P.E.; Hong, S.-B.; Yaghuchi, T.; Varga, J.; Samson, R.A. Aspergillus Species Identification in the Clinical Setting. Stud. Mycol. 2007, 59, 39–46. [Google Scholar] [CrossRef]
- McCusker, J.H.; Clemons, K.V.; Stevens, D.A.; Davis, R.W. Saccharomyces cerevisiae Virulence Phenotype as Determined with CD-1 Mice Is Associated with the Ability to Grow at 42 Degrees C and Form Pseudohyphae. Infect. Immun. 1994, 62, 5447–5455. [Google Scholar] [CrossRef]
- Clemons, K.V.; McCusker, J.H.; Davis, R.W.; Stevens, D.A. Comparative Pathogenesis of Clinical and Nonclinical Isolates of Saccharomyces cerevisiae. J. Infect. Dis. 1994, 169, 859–867. [Google Scholar] [CrossRef]
- Bhabhra, R.; Askew, D.S. Thermotolerance and Virulence of Aspergillus fumigatus: Role of the Fungal Nucleolus. Med. Mycol. 2005, 43 (Suppl. 1), S87–S93. [Google Scholar] [CrossRef]
- Tansey, M.R.; Brock, T.D. The Upper Temperature Limit for Eukaryotic Organisms. Proc. Natl. Acad. Sci. U. S. A. 1972, 69, 2426–2428. [Google Scholar] [CrossRef]
- Lackner, M.; Obermair, J.; Naschberger, V.; Raschbichler, L.-M.; Kandelbauer, C.; Pallua, J.; Metzlaff, J.; Furxer, S.; Lass-Flörl, C.; Binder, U. Cryptic Species of Aspergillus Section Terrei Display Essential Physiological Features to Cause Infection and Are Similar in Their Virulence Potential in Galleria Mellonella. Virulence 2019, 10, 542–554. [Google Scholar] [CrossRef]
Figure 1.
Morphological characteristic of clinical isolates of A. hortae. MCA-7 (A), MCA-8 (B) and MCA-10 (C). Microscopic view Lactophenol cotton blue mount in 100X (top) and Macroscopic view of colonies in MEA (down). Scale bars = 10 μm.
Figure 1.
Morphological characteristic of clinical isolates of A. hortae. MCA-7 (A), MCA-8 (B) and MCA-10 (C). Microscopic view Lactophenol cotton blue mount in 100X (top) and Macroscopic view of colonies in MEA (down). Scale bars = 10 μm.
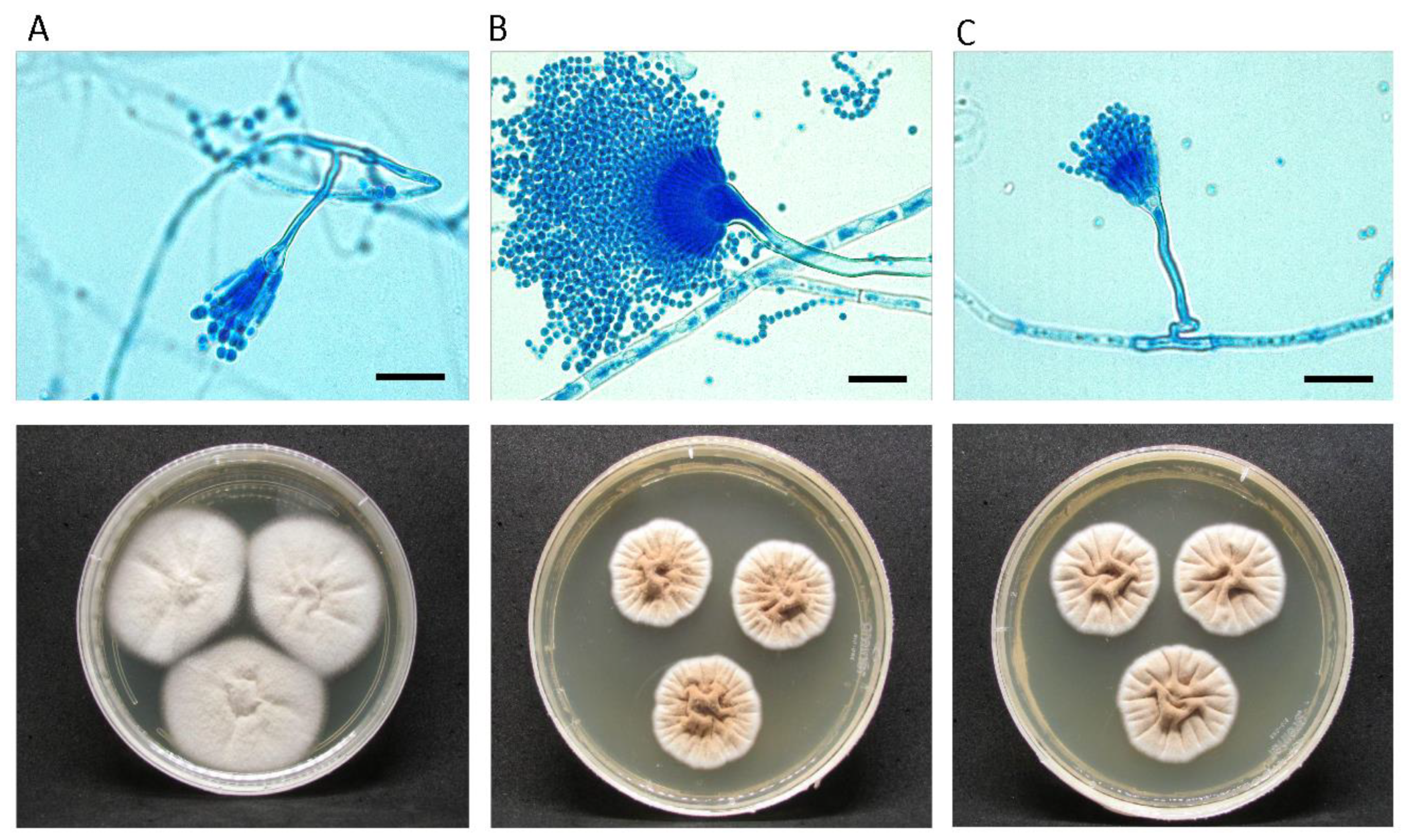
Figure 2.
Radial growth box plot of A. hortae clinical isolates in different culture media and temperatures.(t-test) *** statistical significance p<0.001.
Figure 2.
Radial growth box plot of A. hortae clinical isolates in different culture media and temperatures.(t-test) *** statistical significance p<0.001.
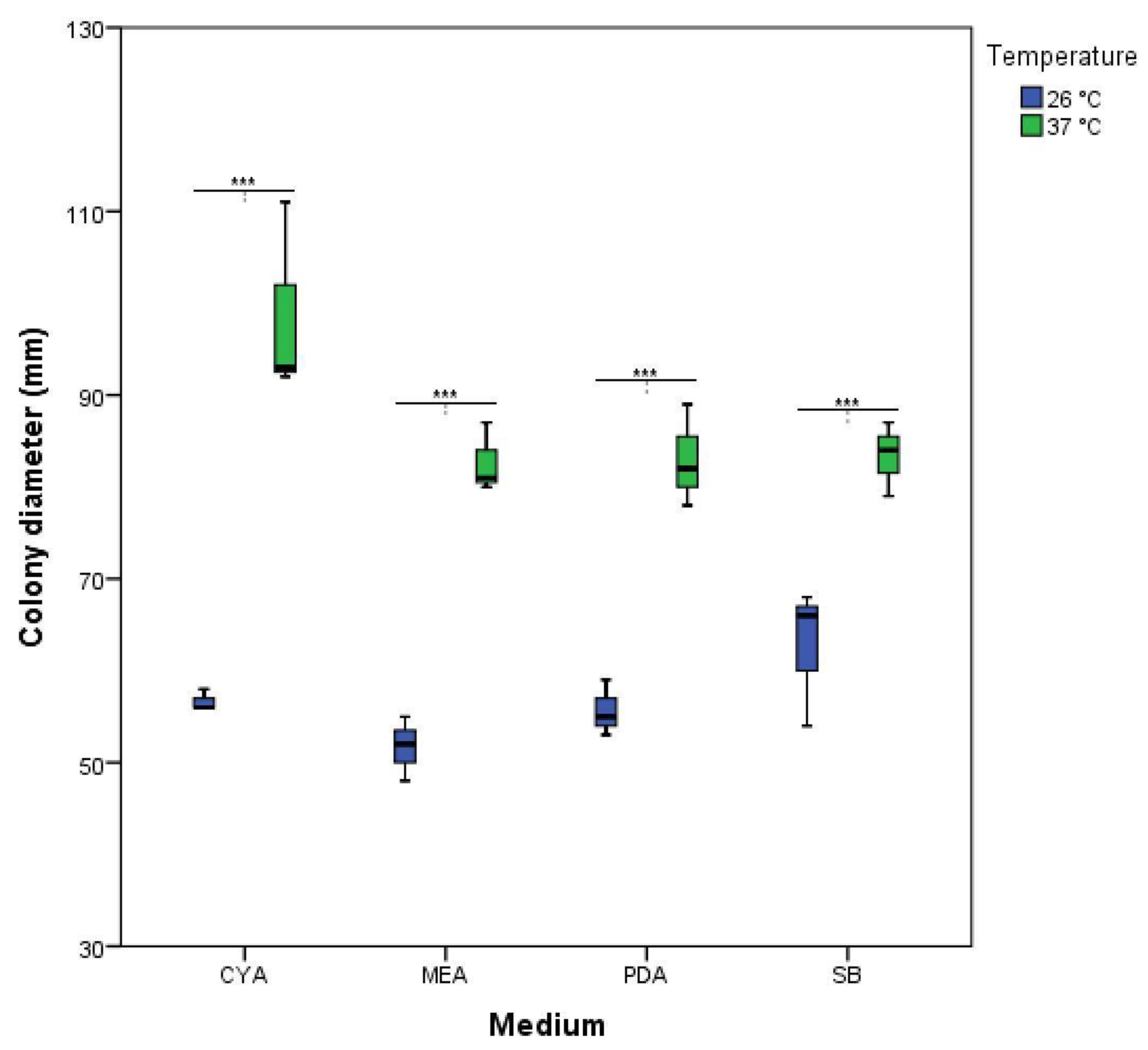
Figure 3.
Genomic variation analysis in A. hortae. (A) Neighbor joining phylogenetic tree of MelA in Terrei and Flavipedes sections. (B) Alignment of 12 MelA (XP_001212741) predicted orthologs from the Aspergillus section Terrei, highlighting a K679M substitution in the hypopigmented isolate MCA7. (C) Alignment of whole genome sequences of A. hortae. Three clinical isolates were aligned to the genome reference Asphor1_AssemblyScaffolds, and Mauve analysis reveals possible inverted regions (red R) in two of the clinical isolates. These regions contain the melanin associated genes (MelA and TyrP orthologs from A. terreus). (D) MelA locus exhibits a SNP, with an A to T substitution, in the MCA7 isolate.
Figure 3.
Genomic variation analysis in A. hortae. (A) Neighbor joining phylogenetic tree of MelA in Terrei and Flavipedes sections. (B) Alignment of 12 MelA (XP_001212741) predicted orthologs from the Aspergillus section Terrei, highlighting a K679M substitution in the hypopigmented isolate MCA7. (C) Alignment of whole genome sequences of A. hortae. Three clinical isolates were aligned to the genome reference Asphor1_AssemblyScaffolds, and Mauve analysis reveals possible inverted regions (red R) in two of the clinical isolates. These regions contain the melanin associated genes (MelA and TyrP orthologs from A. terreus). (D) MelA locus exhibits a SNP, with an A to T substitution, in the MCA7 isolate.
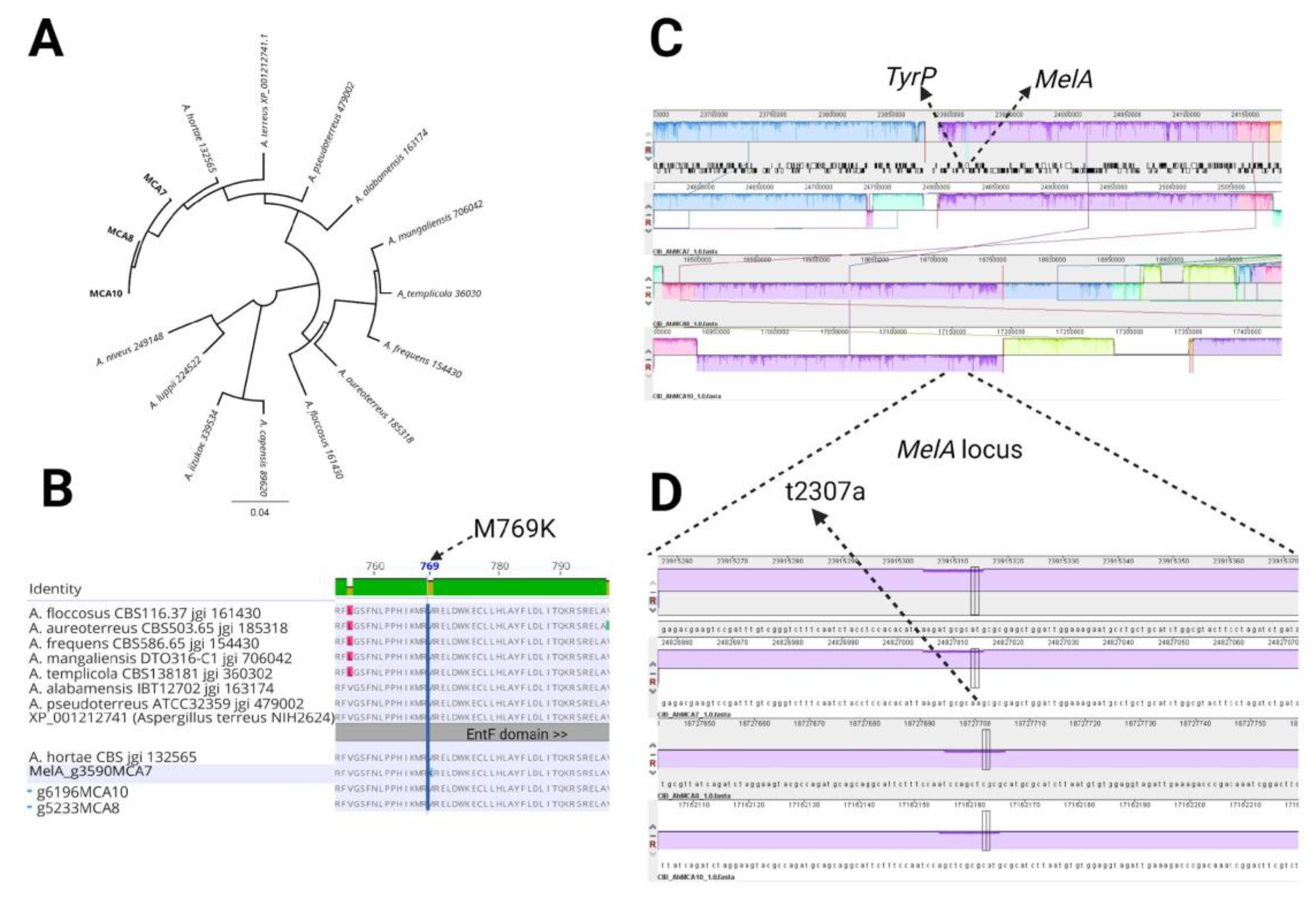
Figure 4.
Phylogenetic Analysis. Phylogenetic tree of Aspergillus section Terrei using concatenated sequences from the BenA-CaM markers, A. fumigatus Af293 CaM was utilized as an outgroup. (A). Phylogenomic tree of A. terreus clade based on 124,027 SNPs position (B). The trees were inferred using the Maximum Likelihood method. The numbers close to branches are % of supported bootstrap after 1,000 replications. Colombian clinical isolates of A. hortae are shown in bold.
Figure 4.
Phylogenetic Analysis. Phylogenetic tree of Aspergillus section Terrei using concatenated sequences from the BenA-CaM markers, A. fumigatus Af293 CaM was utilized as an outgroup. (A). Phylogenomic tree of A. terreus clade based on 124,027 SNPs position (B). The trees were inferred using the Maximum Likelihood method. The numbers close to branches are % of supported bootstrap after 1,000 replications. Colombian clinical isolates of A. hortae are shown in bold.


Table 1.
Antifungal susceptibility test of Aspergillus hortae isolates.
| VRC | ITC | POS | AmB | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | Isolate | MIC | INT | MIC | INT | MIC | INT | MIC | INT |
| A. hortae | MCA-7 | 0.5 | S | 1 | S | 0.25 | S | 2 | R |
| A. hortae | MCA-8 | 0.5 | S | 1 | S | 0.25 | S | 4 | R |
| A. hortae | MCA-10 | 1 | S | 1 | S | 0.25 | S | 4 | R |
VRC: Voriconazole. ITC: Itraconazole, POS: Posaconazole, AmB: Amphotericin B. S: Sensible. R: Resistant. MIC: Minimum Inhibitory Concentration (mg/L). INT: interpretation. VRC: Voriconazole. ITC: Itraconazole. POS: Posaconazole. AmB: Amphotericin B.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



