
Submitted:
26 August 2024
Posted:
28 August 2024
You are already at the latest version
Abstract
Fungal trunk disease (FTD) poses a significant threat to hazelnut (Corylus avellana L.) production worldwide. In Chile, the fungus Diplodia mutila, from the Botryosphaeriaceae family, has been frequently identified causing this disease in the Maule and Ñuble Regions. However, control measures for D. mutila remain limited. This research aimed to evaluate the effectiveness of chemical and biological fungicides against D. mutila under in vitro, controlled pot experiment, and field conditions. An in vitro screening of 30 fungicides was conducted. The effectiveness was assessed by measuring the length of vascular lesions in hazelnut branches inoculated with D. mutila mycelium disks under controlled and field conditions. Field trials were conducted in a hazelnut orchard in Ñiquén, Ñuble Region, Chile. Results showed that three biological and five chemical fungicides were selected in vitro with > 31% inhibition after 14 days. In pot experiments, all fungicides reduced necrotic lesions on branches by 32 to 61%. In field experiments, the most effective systemic fungicides were fluopyram/tebuconazole, fluxapyroxad/pyraclostrobin, and tebuconazole, while the effectiveness of antagonists Pseudomonas protegens ChC7 and Bacillus subtilis QST713 varied with seasonal temperatures. Effective conventional and biological fungicides against D. mutila could be integrated into disease management programs to protect hazelnut wounds from infections.
Keywords:
Corylus avellana L.
; fungal trunk disease
; fungicides
; biocontrol agents
; Pseudomonas protegens ChC7
; Bacillus subtilis QST 713
1. Introduction
The cultivated area of European hazelnut (Corylus avellana L.) has increased significantly in Chile over the last ten years, reaching 36,375 hectares [1] which represents 9,7% of the total land surface used for fruit production, ranking hazelnut as the fifth most important fruit crop in the country. Several diseases caused by bacteria and fungi have become more recurrent in this nut fruit crop. On a national scale, there are reports of bacterial diseases such as crown gall caused by Agrobacterium tumefaciens; bacterial canker caused by Pseudomonas avellanae, P. syringae pv. syringae, and P. syringae pv. coryli; and bacterial blight caused by Xanthomonas arboricola pv. corylina; while reports of fungal diseases include root rot caused by Armillaria mellea; and wood canker by Dothiorella vidmadera (=Diplodia coryli) [2]. In recent years, a higher incidence of diseases affecting the woody tissues of the plant has been reported in hazelnut and other important fruit species in Chile, such as walnut, kiwi, and grapes [3,4,5]. Furthermore, environmental conditions and climate change are recognized predisposing factors for this type of diseases [6].
Different pathogens causing wood damage in hazelnut have been reported in Italy [7,8], the United States [9] and Chile [10,11,12,13,14], with symptoms such as cankers, vascular discoloration, and regressive death of trunk, branches, twig or the entire plant. This syndrome has been called fungal trunk disease (FTD) in hazelnut and other species [8]. These wood pathogens include members of the Botryosphaeriaceae family, with around 300 species, some of which are recognized for affecting numerous woody hosts in temperate and tropical regions [15]. Botryosphaeria pathogens enter through natural openings and wounds resulting from management practices in the orchard, where lesions are evident in the lateral sections of affected branches, typically expanding along the xylem [16,17]. Within this family, species of the genera Diplodia and Dothiorella have been mainly described in hazelnut [9,11,14,18]. In Chile, Diplodia coryli Fuckel, reclassified as Dothiorella vidamera W.M. Pitt, J.R. Úrbez-Torres & Trouillas sp. nov. [19], has been reported causing greyish discoloration in wood and regressive death in five-year-old plants of cv. Barcelona [11], while vascular necrosis caused by Diplodia mutila (Fr.) Mont (teleomorph Botryosphaeria stevensii Shoemaker, [1964]) was first reported in hazelnut orchards in Oregon, USA [9], and recently in the Maule and Ñuble Regions, Chile [14]. This fungal pathogen has also been described causing vascular discoloration, cankers on branches and trunk, and death of small branches in apple trees [20], grapevines [21], walnut trees [22], and Araucaria araucana [23] in Chile. Despite of the impact of plant pathogens such as D. mutila, research has mainly focused on agricultural practices such as selection of healthy plants, pruning of infected tissues, and tool disinfections, while there are no studies addressing control strategies for these pathogens in hazelnut [2,24].
Fungicides such as fluazinam, carbendazim, methyl thiophanate, fludioxonil, pyraclostrobin, myclobutanil, penconazole, tebuconazole have shown high efficacy on isolates of D. mutila from grapevines under in vitro conditions [25]. However, the main buyer of European hazelnuts produced in Chile maintains strict restrictions on the use of chemical fungicides [26], and thus biological control agents (BCAs) can play an important role in the control of D. mutila. However, few studies have evaluated the direct effect of BCAs on D. mutila. There is evidence that fungi such Acremonium mucronatum significantly reduced mycelial growth of D. mutila in vitro and on branches of Quercus trees [27], while other studies on the biocontrol activity of Trichoderma harzianum Th-1 INTA showed parasitism on D. mutila and a marked inhibitory effect on conidia germination and pycnidium formation under in vitro conditions [28]. On the other hand, isolates of Pseudomonas putida JRSK-39, P. koreensis JRSK-45, and Bacillus halotolerans JFM-64, which are rhizobacteria isolated from grapevine, have showed significant reductions in mycelial growth of D. mutila [29], while research on P. aurofaciens has demonstrated a reduction in necrotic lesions caused by D. mutila on stems of Fraxinus excelsior plants [30]. In Chile, it has been described that the aging of hazelnut orchards results in a higher detection of fungal phytopathogen species causing cankers, vascular necrosis, and cambium death [13,14].
Therefore, the objective of this study was to assess the effectiveness of chemical and biological fungicides against D. mutila under in vitro, controlled and field conditions.
2. Results
2.1. Selection of Active Ingredients of Chemical and Biological Fungicides
After 48 h of incubation, nine products (43%) inhibited the growth of D. mutila between 76 to 99% (+++) (Table 1). However, fifteen of the products evaluated were discarded after 120 h of incubation, while only thiophanate-methyl, fluxapyroxad/pyraclostrobin, fluopyram/tebuconazole, fluazinam, tebuconazole, and prochloraz showed inhibition of the fungal colony between 31% and 75% (++). After 14 days, fungal growth was reactivated in most products, while fungal inhibition was maintained in PDA plates treated with prochloraz, fluazinam, tebuconazole, fluxapyroxad/pyraclostrobin, and fluopyram/tebuconazole.
Among the antagonist bacteria, significant differences were observed (Kruskal-Wallis, p = 0.02). Bacillus subtilis QST713 showed the greatest inhibition of D. mutila colony growth (52%), followed by Pseudomonas protegens ChC7, while Pantoea sp. AP113 and P. protegens Ca6 had lower efficacy, with 6.3% and 38%, respectively (Table 2; Figure 1).
In the evaluation of the degree of antagonism, the biological fungicide based on a consortium of Bionectria ochroleuca Mitique, Trichoderma gamsii Volqui, Hypocrea virens Ñire [Mamull® WG. Bio Insumos Nativa SpA, Chile] outperformed D. mutila and covered at least two-thirds of the surface of the medium (Class 2), classifying it as an antagonist agent [31]. On the contrary, the biological product based on Trichoderma spp. and Bacillus spp. [Puelche – VTO. Bio Insumos Nativa SpA, Chile] allowed the colonization of less than a third of the surface of the medium (Class 4), which is not considered antagonistic (Figure 1).
2.2. Assessment of Effectiveness of Chemical and Biological Fungicides against Diplodia mutila in Hazelnut Plants under Pot-Controlled Conditions
The average length of necrotic lesions (mm) for each treatment against D. mutila is shown in Table 3. No interaction was observed between treatment and cultivar factors (p = 0.91), and there were no differences among cultivars (p = 0.87), but significant differences were observed among the treatments applied to the holes inoculated with the fungus (p < 0.01; C.V. = 9.5). Compared to the untreated control, all chemical and biological fungicides significantly reduced the necrosis associated with the development of the fungus and the death of internal branch tissues, with a reduction ranging between 32% and 61%. The most effective treatments were fluxapyroxad/pyraclostrobin, fluazinam, the consortium of B. ochroleuca Mitique, T. gamsii Volqui, and H. virens Ñire, and fluopyram/tebuconazole, with no significant differences among them. Additionally, the antagonists B. subtilis QST 713 and P. protegens ChC7 showed no statistical differences compared to fluopyram/tebuconazole and prochloraz.
2.3. Assessment of Effectiveness of Chemical and Biological Fungicides against Diplodia mutila under Field Conditions
Pearson’s analysis determined that there was no correlation (p > 0.05) between branch diameter (mm) and length of vascular necrosis (mm) caused by the fungus in the two seasons under study (r = 0.20 [p = 0.32] for the 2020-21 season and r = 0.05 [p = 0.80] for the 2021-22 season).
During the 2020-21 season (Table 4), no interaction was detected between the factors of chemical treatments and inoculation time (p = 0.27; C.V. = 6.34). Furthermore, there were no significant differences in the length of vascular necrosis caused by D. mutila in branches treated with chemical fungicides (p = 0.82). However, significant differences were detected among the average lengths of vascular necrosis at each inoculation time (p > 0.01), indicating that branches inoculated with D. mutila on the same day as application presented greater necrotic lesions.
There was no interaction between the factors of biological treatments and inoculation time (p = 0.41; C.V. = 4.56). Differences were observed among lesions in branches treated with biological control agents (p > 0.01; Figure 2), where in the presence of the antagonist P. protegens ChC7, a 29.2% reduction in necrosis was observed compared to the control. There were no differences between inoculation times (p = 0.79).
In the 2021-22 season (Table 5), there was an interaction between the factors of chemical treatments and inoculation time (p < 0.01; C.V. = 2.75). Additionally, each factor showed significant differences (p < 0.01). Except for fluazinam, chemical fungicides significantly reduced the length of the lesion caused by D. mutila when inoculated on the same day as treatment application, ranging from 24 to 37% compared to the control. However, in the inoculation conducted 24 hours after treatment application, only the systemic fungicides tebuconazole, fluxapyroxad/pyraclostrobin, and fluopyram/tebuconazole showed significant differences, ranging from 29 to 38% compared to the control. Additionally, significant differences were observed among inoculation times (p < 0.01), with the length of vascular necrosis being lower in all treatments of the first inoculation compared to the second inoculation.
There was no interaction between the factors of biological treatments and inoculation time (p = 0.19; C.V. = 3.89). Differences were observed among lesions in branches treated with biological control agents (p < 0.01; Figure 3), where in the presence of the antagonist B. subtilis QST 713 and the consortium of fungi (B. ochroleuca Mitique, T. gamsii Volqui and H. virens Ñire), reductions of 38 and 40%, respectively, were observed. Additionally, there were differences between inoculation times (p < 0.01), with a 33% increase in necrotic lesions in the second inoculation.
Based on the number of catkins and glomerules evaluated, the inoculated and treated branches did not affect the fruiting potential of the species when evaluated as variation in the density of male and female flowers (Table S2).
3. Discussion
Diplodia mutila is a phytopathogenic fungus primarily associated with wood diseases in agricultural crops and forestry species of interest [9,14,22,23,32,33,34,35]. This study represents the first investigation into the effectiveness of chemical and biological fungicides in reducing the damage caused by this fungus in European hazelnut. Of the nineteen chemical fungicides evaluated in vitro, using a quick screening test by using recommended commercial doses in other fruit crops, only six qualitatively reduced the development of D. mutila when applied at commercial doses on the culture medium. In particular, methyl thiophanate was highly toxic on D. mutila, which agrees with the findings of Mondello et al. (2018) [25] and Sosa et al. (2022) [28]. However, due to the European Commission’s decision not to renew approval for this active substance based on the latest report [36], which highlighted insufficient information regarding its safe use in the environment and with regard to mammals, methyl thiophanate was excluded from the plant trials in this research.
Inhibition rates demonstrated the antagonistic effect of Chilean strains of Pseudomonas protegens on D. mutila. Among these strains, P. protegens ChC7 was the most effective and has been available in the local market since 2022 (TANIRI® WP, Bio Insumos Nativa SpA., Chile) due to its ability to induce plant resistance [37,38]. Previous research has shown that these P. protegens strains, isolated from the wheat rhizosphere, exhibit antagonistic activity against other phytopathogenic species affecting important crops in Chile [37,39,40]. This activity is associated with the synthesis of secondary metabolites, including antibiotics such as 2,4-diacetylphloroglucinol (2,4-DAPG), pyrrolnitrin, and pyoluteorin, which are produced by genes phl, prn, and plt [37,39,40,41]. Additionally, this bacterial species can induce plant resistance [38,42]. In a study on the control of necrotic wood fungi, soil rhizobacteria isolates from grapevines showed effective results in inhibiting the mycelial growth of D. mutila compared to various Pseudomonas species, using a methodology similar to that used in this study [29]. Furthermore, Huang et al. (2022) [43] determined the efficacy of P. protegens in reducing mycelial growth and postharvest rot caused by a Botryosphaeriaceae fungus like Botryosphaeria dothidea, the causal agent of apple ring rot. In our study, Bacillus subtilis QST 713 exhibited the highest inhibitory effect on the mycelial growth of D. mutila among the tested antagonistic bacteria in vitro. This Bacillus strain is widely used as a biological control agent (BCA) due to its broad-spectrum inhibitory activity against several phytopathogens through the production of antibiotics and antifungal enzymes [44], which provides evidence of its inhibitory effect in vitro on the mycelial growth of D. mutila. Moreover, it can induce systemic acquired resistance (SAR) in plants [45].
The consortium of B. ochroleuca Mitique, T. gamsii Volqui and H. virens Ñire (anamorph: Trichoderma virens) showed the highest efficacy in the in vitro experiments, although there are few published reports on its use against wood-infecting pathogens. Silva-Valderrama et al. (2021) [46] demonstrated that this consortium restricted over 50% of the mycelium area of D. seriata, Neofusicoccum parvum, and Phaeomoniella chlamydospora obtained from grapevines on PDA plates. Additionally, Mondello et al. (2018) [25] showed that an isolate of T. gamsii was effective in inhibiting the teleomorph of D. mutila in in vitro studies, which is associated with possible antibiosis and mycoparasitism [47]. These findings agree with the in vitro results obtained in the present study, highlighting the potential of BCAs like Trichoderma for the control of pathogens causing wood necrosis in European hazelnut.
The controlled pot experiment validated the efficacy of chemical fungicides against D. mutila observed in the in vitro assays based in a quick screening test, as all five products effectively reduced necrotic lesions on the branches of inoculated nursery plants. The manufacturer’s recommended dose for chemical fungicides considers concentrations that are non-phytotoxic to a variety of crops, including vegetables, industrial crops, and fruit and forest trees (Table S3). Therefore, the dose used for fruit trees was applied. The concentration of the active ingredient (mg L-1 or ppm) in the commercial dose varied for each fungicide, reaching concentrations within the treated and subsequently inoculated hole wound with the pathogen ranging from 0.05 mg (tebuconazole) to 0.14 mg (prochloraz) (Table S4). However, the observed high efficacy of the commercial doses of the five chemical fungicides and the three biofungicides against D. mutila needs validation for their efficacy against other wood-necrotizing fungal species such as D. coryli [11], Diaporthe australafricana [48], and Neofusicoccum parvum [12], which have been reported affecting European hazelnut in Chile.
In general, there is limited information on the epidemiology and etiology of D. mutila infection in European hazelnut. According to a comprehensive study and review of vine disease management conducted by Gramaje et al. (2018) [49], Botryosphaeria dieback caused by botryosphaeria fungi is transmitted through airborne spore dispersal, with infection primarily occurring at cuts from annual pruning or any open wounds, followed by colonization and necrosis of vascular tissues. A monitoring study of D. mutila conidia conducted over two years in grapevines in France determined that conidia release was associated with rain episodes, and high-risk infection periods occurred from early to mid-fall, mid-spring, and late summer, with low conidia detection during winter [50]. Mycelial growth of D. mutila in vitro occurs within the range of 5°C to 35°C, with optimal growth between 20°C and 30°C and an optimum temperature of 25°C [51]. Considering temperature variability during the month of treatment application in the field (October 2020), the environmental conditions recorded 30 days after plant inoculation had an average temperature of 12.8°C, with an average minimum of 5.2°C (Range: 0.8 – 10.7 °C) and an average maximum of 20.4°C (Range: 16.2 – 26.8°C) (Table S5). During the 208 days of the field experiment in the 2020-21 season, temperatures above 20°C were observed for 41 days. In the second season, over the 160-day period of the experiment, temperatures above 20°C were recorded for 60 days, with a 30-day period post-inoculation having an average temperature of 15.5°C and an average minimum of 7.5°C (Range: 2.1 – 11.7 °C) and an average maximum of 23°C (Range: 15.3 – 29.3°C). Temperature variations between the two seasons could explain the differences in the length of necrotic lesions observed on branches inoculated with D. mutila, suggesting that longer necrotic lesions in the vascular tissue of branches inoculated in the 2021-22 season were related to a more extended period of favorable temperatures for mycelial development. Moreover, regardless of the time in which the tissue remained infected with this pathogen, temperature would be a determining factor in the level of damage experienced by hazelnut tissues. On the other hand, the conditions in the second season favored the development of the fungus and allowed for significant differences between the treatments based on chemical and biological fungicides. Precipitation and relative humidity conditions did not vary significantly between the seasons and would not have influenced the observed necrosis levels for each treatment (Table S5). This information provides insights into environmental conditions favoring pathogen infection and, combined with data on D. mutila spread, can help decision-making regarding fungicide application to protect the plants.
The results from both pot and field experiments, particularly in the second season, confirm the efficacy of traditional chemical fungicides in mitigating the damage inflicted by D. mutila on hazelnut branches. Furthermore, active ingredients with systemic properties led to a notable decrease in necrotic lesions compared to contact-based products. In the case of branches inoculated 24 hours post-fungicide application, tebuconazole, fluxapyroxad/pyraclostrobin, and fluopyram/tebuconazole demonstrated prolonged activity in the plant wound, resulting in a more pronounced reduction in pathogen-induced damage.
In the pot experiments, antagonistic organisms proved to be highly effective in disease control (30 to 55% efficacy). However, there was variability in the performance of biological treatments between the seasons in the field trials. Furthermore, it was evident that the impact of biological treatments persisted in both direct and deferred inoculations, revealing that beneficial microorganisms could reproduce in the tissues of hazelnut plants, achieving effective control of D. mutila. Only P. protegens ChC7 demonstrated efficacy in the 2020-21 season, while B. subtilis QST 713 and the consortium of fungi B. ochroleuca Mitique, T. gamsii Volqui and H. virens Ñire showed positive results in the 2021-22 season. P. protegens is a bacterium isolated from wheat roots, and thus it grows at lower temperatures [37,39], which could explain its higher efficacy during the colder temperatures of the first season. In contrast, B. subtilis QST 713 and Trichoderma spp. or Hyprocrea spp. require higher temperatures (25-55°C; 25-30°C; 25-30°C, respectively) for optimal antagonistic activity [36,52,53], which could account for their greater efficacy in controlling D. mutila during the second season.
The results of the floral density analysis (Table S2) demonstrate that the application of treatments did not affect the productive potential of the plant. These findings suggest that no damage is generated in the wood that could affect the floral development of both catkins and clusters in the early stages of infection caused by D. mutila. This might occur because floral induction in hazelnut occurs between 4-5 months before flowering, specifically during the months of December and January [54]. Given that the fungus was inoculated in early spring, the period of greatest damage caused by this pathogen does not necessarily coincide with the period of greatest susceptibility to negative effects on floral development. Another explanation could be associated with the source-sink relationship, as it has been determined that branches or floral shoots of hazelnut do not exhibit autonomy in their carbohydrate source-sink capacity and are essential for floral development [55]. Therefore, even though the damage caused by the fungus resulted in injury at the phloem level of the branch, it does not imply an interruption in the direct flow of carbohydrates for the development of new flowers for the following season.
Considering that the main market for the consumption and export of Chilean hazelnuts is Europe, the use of active substances for pest and disease control faces higher restrictions aimed at minimizing environmental impact. Therefore, antagonists (Pseudomonas protegens or Bacillus spp.) and active ingredients with systemic action (fluopyram/tebuconazole, fluxapyroxad/pyraclostrobin, and tebuconazole) would be effective tools in reducing damage caused by D. mutila. However, it is essential to conduct accurate monitoring of environmental conditions during crop production seasons, particularly rainfall and temperatures above 20°C, as they favor the spread and development of this fungus. Similarly, environmental factors would influence the selection of antagonistic microorganisms to be included in a phytosanitary management plan that promotes their reproduction and control activity. The use of registered fungicides and BCAs is crucial for establishing integrated disease control and maintaining quality and safety in hazelnut production.
4. Materials and Methods
4.1. Phytopathogenic Fungus
The phytopathogenic fungus Diplodia mutila isolate F096 was selected due to its high aggressiveness on hazelnut (Corylus avellana L.) [14]. This isolate was collected from wood exhibiting vascular necrosis in 2018 from hazelnut orchards in the Ñuble Region, Chile. The phytopathogen was stored in the microorganism collection of the Plant Pathology Laboratory at the Faculty of Agronomy, Universidad de Concepción, Chillán Campus.
4.2. In Vitro Pathogen Control Assays
Thirty treatments were used to assess the efficacy in controlling D. mutila under in vitro conditions. The evaluated treatments included nineteen chemical fungicides with different modes of action, three natural extracts and salts, and eight biological control agents (BCAs), consisting of three commercial biopesticides and five bacterial microorganisms from the collection of the Plant Pathology Laboratory at the Faculty of Agronomy (Table S1).
4.3. Selection of Treatments with Fungicidal Activity against Diplodia mutila
Solutions were prepared using the commercial doses or concentrations suggested by the manufacturer for each fungicide (Table S1). For the chemical fungicides, a 500 µL aliquot of each product solution was evenly spread on a Petri dish (80 mm diameter) containing 10 mL of potato dextrose agar (PDA) medium. After one hour, once the medium surface was dry, 5 mm diameter discs of actively growing D. mutila mycelium were placed to assess the ability to inhibit fungal mycelial growth. The assessment was conducted at 48 and 120 h after incubation using a rating scale with five levels of mycelium growth on the PDA medium: (-) = Normal fungal growth, (+) = Moderate fungal growth (1 to 30% less than normal fungal growth), (++) = Mild fungal growth (31 to 75% less fungal growth), (+++) = Limited fungal growth (76 to 99%), and (++++) = 100% inhibition of fungal growth (Figure S1). Three repetitions were performed for each treatment. The plates were kept in an incubation chamber and incubated in the dark at 25°C for two weeks.
For bacterial and fungal antagonists, commercial doses of bioproducts and suspensions of bacteria grown in King B (KB) broth from the laboratory collection were applied to the PDA medium using a 10 µL bacteriological inoculation loop. Two straight parallel lines were extended 15 mm from the 5 mm diameter mycelium disc of the target fungus, placed at the centre of the plate. Three repetitions were performed for each treatment. The plates were maintained in an incubation chamber at 25°C in darkness for one week. Pseudomonas protegens bacterial strains were grown by taking 150 µL of the strain sample and cultured in KB broth at 25°C for 48 h in a shaking incubator at 150 rpm. The bacterial concentration was determined by counting colony forming units (CFUs) in serial dilutions.
For the biological treatments based on antagonistic bacteria, fungal growth was determined by measuring the fungal colony mycelium diameter (mm) with a steel ruler (Hand®, 200 mm), while the action of products with Trichoderma species was evaluated using the antagonism classification proposed by Bell et al. (1982) [31], with the following growth patterns: Class 1 = Trichoderma completely overgrows the pathogen and covers the entire medium surface; Class 2 = Trichoderma overgrows at least two-thirds of the medium surface; Class 3 = Trichoderma and the pathogen each colonize approximately one-half of the medium surface (more than one-third and less than two-thirds), and neither organism dominates the other; Class 4 = The pathogen colonizes at least two-thirds of the medium surface and appears to withstand encroachment by Trichoderma invasion, occupying the entire medium surface; Class 5 = The pathogen completely dominates Trichoderma, overgrows it, and occupies the entire medium surface. Trichoderma was considered a good antagonist if the average score for a given comparison was less than or equal to 2, but not very antagonistic if the number was greater than or equal to 3.
4.4. Assessment of Effectiveness of Chemical and Biological Fungicides against D. mutila in Hazelnut Plants under Controlled Conditions
The pot experiment to assess the preventive control of D. mutila with the selected fungicide treatments was conducted using two years old hazelnut plants of cv. Barcelona and Tonda di Giffoni. Plants with a central trunk axis of approximately 150 cm in height were maintained in individual plastic pots of 0.03 m3 with a substrate of peat moss:perlite (3:1).
A 6.5 mm diameter hole was drilled in the trunk of each plant using a drill (Bauker®, model SD-GS1041, China) to a depth of 4 mm. Subsequently, 300 µL of the chemical fungicide or BCA treatment selected from the in vitro assays was applied to each hole using a micropipette. After 20 min, the treated hole was inoculated with a 5 mm diameter mycelium disc of D. mutila, and additionally, a PDA disc was inoculated as a control in a separate hole. The inoculated wounds were sealed with plastic film throughout the experiment to prevent dehydration. Plants were maintained under a 50% shaded Raschel mesh cover and irrigated three times weekly for five months. The treatments were distributed in a randomized complete block design with a factorial arrangement, and the experiment had seven replications.
At the end of the pot experiment, the central axes of each plant were removed, and each branch or stem segment was cut vertically through the inoculation point to measure the length of the necrotic lesion (mm) and assess the effect of each treatment on D. mutila compared to the water control. Additionally, fungi were isolated from the lesions by cutting pieces of the advancing necrosis zone and disinfecting the surface for 1 min in a 2.5% sodium hypochlorite solution, followed by two washes with sterile distilled water (SDW) for 1 min. Each branch piece was placed on plates with PDA medium containing streptomycin (200 mg L-1). The plates were incubated in a growth chamber at 25°C for 10 days in darkness. Fungi were identified based on their morphological characteristics observed using an optical microscope (Motic® BA310E).
4.5. Assessment of Effectiveness of Chemical and Biological Fungicides against D. mutila under Field Conditions
4.5.1. Study Area
The experiment was conducted in an eight-year-old hazelnut orchard of cv. Tonda di Giffoni over two consecutive seasons (2020-21 and 2021-22). The orchards were located in "Campo San Gregorio," Ñiquén, Ñuble Region, Chile (36°18’25’’S; 71°49’32’’W), owned by Frutícola AgriChile S.A. The orchards were established in a multiple-leader trained system with a planting framework of 5 m between rows and 3 m between plants on the row, under drip irrigation and fertirrigation. Soils belonged to the Tuiquilemu series (Inceptisols) [orchard in the 2020-21 season] and the Mirador series (Alfisols) [orchard in the 2021-22 season], with loamy and loamy-clayey textures, respectively [56]. The orchards were managed with a fertilization program, irrigation, insect control, and weed control implemented by the company. The trials were established on September 30, 2020 [first season] and October 13, 2021 [second season]. The experiment used a factorial arrangement in a completely randomized block design with four replications. Treatments were applied to five trees per plot, arranged in two rows separated by an untreated row to prevent drift. Evaluation was focused on the central tree in each treated row to ensure no contamination from adjacent treatments due to potential wind drift.
4.5.2. Fungal Inoculations in Hazelnut Trees
Five healthy branches per tree were selected, and a hole approximately 4 mm deep and 6.5 mm in diameter was made using an electric drill (Bauker ®, SD-GS1041, China)[57,58]. The trees were sprayed with the chemical and biological fungicides (Table 6) selected from the previous experiments, using an experimental adjustable static piston sprayer. An average volume of 2.5 L per plant for each application (1000 L per hectare) was used, and the trees were sprayed until leaf runoff. One hour after the treatments were applied, the fungus was inoculated into the holes of the branches using 5 mm diameter discs of actively growing D. mutila mycelium, ensuring contact with the internal tissues of the wood (vascular cambium). For each treatment, a branch was inoculated with a sterile PDA disc as a control. Finally, the holes were sealed with plastic film to protect the lesion from external contaminants and improve moisture conditions for phytopathogen growth. The inoculation methodology was repeated the following day on two additional branches per tree that had already been drilled and sprayed with the treatments the previous day.
Additionally, seven days after fungicide application, the diameter of the inoculation zone of each branch was measured with a Peltier caliper (Ubermann, 6” stainless, United States) to determine its influence as a covariate on the length of the vascular lesion.
The inoculated branches were removed from the orchards after seven months in the first season and after five months in the second season for evaluation. A vertical cut was made through the vascular tissue that passed through the inoculation point on the branch to measure the length of the necrotic lesion from end to end. Subsequently, the edge of the necrotic area observed in the inoculated branches was cut, and the surface was disinfected for 1 min in a 2.5% sodium hypochlorite solution, followed by two washes for 1 min with SDW. Isolations were carried out by placing a piece of necrotic tissue on PDA. The plates were incubated in the dark at 25°C for 10 days. D. mutila was identified through morphological observations under an optical microscope. For each plant, an inoculated branch was selected to assess the number of male (catkins) and female (glomerulus) flowers, which were compared with uninoculated branches within each experimental plot. The evaluation was conducted at BBCH 55 (male flowers) and BBCH 600 (female flowers) in the first season, while in the second season, male flowers were evaluated at BBCH 57 and female flowers at BBCH 600 [59].
Throughout the experiment, the environmental conditions of the area were monitored monthly, using data from the meteorological station of Frutícola Agrichile S.A., San Gregorio, Ñiquén, Ñuble Region (Table S5).
4.6. Data Analysis
The results of the evaluations conducted in the in vitro, pot, and field experiments were subjected to normality and homogeneity of variance analysis using the Shapiro-Wilk and Levene tests, respectively. If assumptions were fulfilled, the data were analyzed by analysis of variance (ANOVA); otherwise, the non-parametric Kruskal-Wallis test was used. Means were compared using the Fisher’s Least Significant Difference (LSD) test (α = 0.05). All the statistical analyses were conducted using Infostat software (Universidad de Córdoba, Argentina) version 2022 [60].
5. Conclusions
Bacillus subtilis QST 713, Pseudomonas protegens ChC7, the consortium of Bionectria ochroleuca Mitique, Trichoderma gamsii Volqui, and Hypocrea virens Ñire were selected as effective BCAs to inhibit the development of Diplodia mutila mycelium in the in vitro assays. The application of the selected chemical fungicides and BCAs on the wounds subsequently inoculated with D. mutila mycelium in hazelnut wood resulted in significant reductions in wood necrosis in nursery plants, whereas field trials demonstrated that systemic fungicides fluopyram/tebuconazole, fluxapyroxad/pyraclostrobin, and tebuconazole and the three selected products based on BCAs were effective in reducing necrosis in inoculated wounds in branches, but BCA efficacy against D. mutila varied between seasons. The chemical and biological fungicides assessed in this research are options of control strategies to reduce the incidence of D. mutila in hazelnut production, and thus they may be included in future integrated disease management programs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, data curation, formal analysis, investigation, methodology, validation, visualization, writing—original draft, and writing—review and editing, V.R.; methodology, J.S.M. and B.R.; writing—review and editing, R.M.B. and E.S.; project administration, M.J.L., T.D.G. and M.M.; conceptualization, funding acquisition, formal analysis, investigation, methodology, project administration, resources, supervision, validation, and writing—review and editing, E.M.E. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Integrated Disease Management Program of the Laboratory of Phytopathology of the Faculty of Agronomy, Universidad de Concepción, Project: “Wood decay pathogens affecting hazelnut in Chile (Project I and II)” Research Project Ferrero Hazelnut Company (HCo) and Universidad de Concepción.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Centro de Información de Recursos Naturales (CIREN) Catastro Frutícola Región Metropolitana 2023 2023.
- Latorre, B. Compendio de las enfermedades de las plantas; Ediciones Universidad Católica de Chile: Santiago, Chile, 2018; ISBN 978-956-14-2297-1. [Google Scholar]
- Díaz, G.A.; Zoffoli, J.P.; Ferrada, E.E.; Lolas, M. Identification and Pathogenicity of Diplodia, Neofusicoccum, Cadophora, and Diaporthe Species Associated with Cordon Dieback in Kiwifruit Cultivar Hayward in Central Chile. Plant Disease 2021, 105, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Larach, A.; Torres, C.; Riquelme, N.; Valenzuela, M.; Salgado, E.; Seeger, M.; Besoain, X. Yield Loss Estimation and Pathogen Identification from Botryosphaeria Dieback in Vineyards of Central Chile over Two Growing Seasons. Phytopathologia Mediterranea 2020, 59, 537–548. [Google Scholar] [CrossRef]
- Luna, I.; Besoain, X.; Saa, S.; Peach-Fine, E.; Cadiz Morales, F.; Riquelme, N.; Larach, A.; Morales, J.; Ezcurra, E.; Ashworth, V.E.T.M.; et al. Identity and Pathogenicity of Botryosphaeriaceae and Diaporthaceae from Juglans Regia in Chile. Phytopathol. Mediterr. 2022, 61, 79–94. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Fabi, A.; Varvaro, L. Summer Heat and Low Soil Organic Matter Influence Severity of Hazelnut Cytospora Canker. Phytopathology® 2014, 104, 387–395. [Google Scholar] [CrossRef]
- Belisario, A.; Maccaroni, M.; Coramusi, A. First Report of Twig Canker of Hazelnut Caused by Fusarium Lateritium in Italy. Plant Disease 2005, 89, 106–106. [Google Scholar] [CrossRef]
- Martino, I.; Monchiero, M.; Gullino, M.L.; Guarnaccia, V. Characterization and Pathogenicity of Fungal Species Associated with Hazelnut Trunk Diseases in North-Western Italy. J Plant Pathol 2024. [Google Scholar] [CrossRef]
- Wiman, N.G.; Webber, J.B.; Wiseman, M.; Merlet, L. Identity and Pathogenicity of Some Fungi Associated with Hazelnut (Corylus Avellana L.) Trunk Cankers in Oregon. PLoS ONE 2019, 14, e0223500. [Google Scholar] [CrossRef]
- Guerrero Contreras, J.; Galdames Gutierrez, R.; Ogass Contreras, K.; Pérez Fuentealba, S. First Report of Diaporthe Foeniculina Causing Black Tip and Necrotic Spot on Hazelnut Kernel in Chile. Plant Disease 2020, 104, 975. [Google Scholar] [CrossRef]
- Guerrero, J.A.; Pérez, S.M. First Report of Shoot Blight and Canker Caused by Diplodia Coryli in Hazelnut Trees in Chile. Plant Disease 2013, 97, 144–144. [Google Scholar] [CrossRef]
- Lolas, M.A.; Latorre, B.A.; Ferrada, E.; Grinbergs, D.; Chilian, J.; Ortega-Farias, S.; Campillay-Llanos, W.; Díaz, G.A. Occurrence of Neofusicoccum Parvum Causing Canker and Branch Dieback of European Hazelnut in Maule Region, Chile. Plant Disease, 1539. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.; San Martín, J.; Ruiz, B.; Rojas, K.; Lisperguer, M.J.; De Gregorio, T. Fungi Associated with Wood Damage in Hazelnut ( Corylus Avellana L.) between the Maule and Araucanía Regions of Chile. Acta Hortic. [CrossRef]
- Moya-Elizondo, E.A.; Ruiz, B.E.; Gambaro, J.R.; San Martin, J.G.; Lisperguer, M.J.; De Gregorio, T. First Report of Diplodia Mutila Causing Wood Necrosis in European Hazelnut in Chile. Plant Disease 2023, 107, 565. [Google Scholar] [CrossRef]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in the Botryosphaeriales: Looking Back, Looking Forward. Fungal Biology 2017, 121, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Cankers and Other Diseases Caused by the Botryosphaeriaceae. In Infectious forest diseases; Gonthier, P., Nicolotti, G., Eds.; CABI: UK, 2013; ISBN 978-1-78064-040-2. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as Endophytes and Latent Pathogens of Woody Plants: Diversity, Ecology and Impact. Fungal Biology Reviews 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Alves, A.; Abdollahzadeh, J.; Phillips, A.J.L. Phylogeny, Morphology and Pathogenicity of Botryosphaeriaceae, Diatrypaceae and Gnomoniaceae Associated with Branch Diseases of Hazelnut in Sardinia (Italy). Eur J Plant Pathol 2016, 146, 259–279. [Google Scholar] [CrossRef]
- Pitt, W.M.; Úrbez-Torres, J.R.; Trouillas, F.P. Dothiorella Vidmadera, a Novel Species from Grapevines in Australia and Notes on Spencermartinsia. Fungal Diversity 2013, 61, 209–219. [Google Scholar] [CrossRef]
- Díaz, G.A.; Latorre, B.A.; Ferrada, E.; Lolas, M. Identification and Characterization of Diplodia Mutila, D. Seriata, Phacidiopycnis Washingtonensis and Phacidium Lacerum Obtained from Apple (Malus x Domestica) Fruit Rot in Maule Region, Chile. Eur J Plant Pathol 2019, 153, 1259–1273. [Google Scholar] [CrossRef]
- Díaz, G.A.; Auger, J.; Besoain, X.; Bordeu, E.; Latorre, B.A. Prevalence and Pathogenicity of Fungi Associated with Grapevine Trunk Diseases in Chilean Vineyards. Cienc. Inv. Agr. 2013, 40, 327–339. [Google Scholar] [CrossRef]
- Díaz, G.A.; Latorre, B.A.; Ferrada, E.; Gutiérrez, M.; Bravo, F.; Lolas, M. First Report of Diplodia Mutila Causing Branch Dieback of English Walnut Cv. Chandler in the Maule Region, Chile. Plant Disease 2018, 102, 1451. [Google Scholar] [CrossRef]
- Besoain, X.; Guajardo, J.; Camps, R. First Report of Diplodia Mutila Causing Gummy Canker in Araucaria Araucana in Chile. Plant Disease 2017, 101, 1328–1328. [Google Scholar] [CrossRef]
- Silvestri, C.; Bacchetta, L.; Bellincontro, A.; Cristofori, V. Advances in Cultivar Choice, Hazelnut Orchard Management, and Nut Storage to Enhance Product Quality and Safety: An Overview. J Sci Food Agric 2021, 101, 27–43. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Disease 2018, 102, 1189–1217. [Google Scholar] [CrossRef]
- European Comission Active Substances Considered Approved According to Regulation (EC) No. 1107/2009 2023.
- Ragazzi, A.; Moricca, S.; Vagniluca, S.; Dellavalle, I. Antagonism of Acremonium Mucronatum towards Diplodia Mutila in Tests in Vitro and in Situ. European Journal of Forest Pathology 1996, 26, 235–243. [Google Scholar] [CrossRef]
- Sosa, M.C.; Lutz, M.C.; Lódolo, X.; Basso, C.N. In Vitro and in Vivo Activity of Chemical Fungicides and a Biofungicide for the Control of Wood Diseases Caused by Botryosphaeriales Fungi in Apple and Pear. International Journal of Pest Management 2022, 68, 328–338. [Google Scholar] [CrossRef]
- Kenfaoui, J.; Lahlali, R.; Laasli, S.-E.; Lahmamsi, H.; Goura, K.; Radouane, N.; Taoussi, M.; Fardi, M.; Tahiri, A.; Barka, E.A.; et al. Unlocking the Potential of Rhizobacteria in Moroccan Vineyard Soils: Biocontrol of Grapevine Trunk Diseases and Plant Growth Promotion. Biological Control 2023, 186, 105338. [Google Scholar] [CrossRef]
- Przybyl, K. Effect of Pseudomonas Spp. on Inoculation of Young Plants of Fraxinus Excelsior Stem with Diplodia Mutila. Dendrobiology 2003. [Google Scholar]
- Bell, D.K.; Wells, H.D.; Markham, C.R. In Vitro Antagonism of Trichoderma Species Against Six Fungal Plant Pathogens. Phytopathology 1982, 72, 379. [Google Scholar] [CrossRef]
- Carlucci, A.; Cibelli, F.; Lops, F.; Raimondo, M.L. Characterization of Botryosphaeriaceae Species as Causal Agents of Trunk Diseases on Grapevines. Plant Disease 2015, 99, 1678–1688. [Google Scholar] [CrossRef] [PubMed]
- Valencia, A.L.; Gil, P.M.; Latorre, B.A.; Rosales, I.M. Characterization and Pathogenicity of Botryosphaeriaceae Species Obtained from Avocado Trees with Branch Canker and Dieback and from Avocado Fruit with Stem End Rot in Chile. Plant Disease 2019, 103, 996–1005. [Google Scholar] [CrossRef]
- Díaz, G.A.; Valdez, A.; Halleen, F.; Ferrada, E.; Lolas, M.; Latorre, B.A. Characterization and Pathogenicity of Diplodia, Lasiodiplodia, and Neofusicoccum Species Causing Botryosphaeria Canker and Dieback of Apple Trees in Central Chile. Plant Disease 2022, 106, 925–937. [Google Scholar] [CrossRef]
- Lee, C.A.; Rooney-Latham, S.; Brown, A.A.; McCormick, M.; Baston, D. Pathogenicity of Three Botryosphaeriaceae Fungi, Diplodia Scrobiculata, Diplodia Mutila, and Dothiorella Californica, Isolated from Coast Redwood ( Sequoia Sempervirens ) in California. Forest Pathology 2022, 52, e12764. [Google Scholar] [CrossRef]
- Anastassiadou, M; Arena, M.; Auteri, D.; Brancato, A.; Bura, L.; Carrasco Cabrera, L.; Chaideftou, E.; Chiusolo, A.; Crivellente, F.; et al.; European Food Safety Authority (EFSA) Peer Review of the Pesticide Risk Assessment of the Active Substance Bacillus Amyloliquefaciens Strain QST 713 (Formerly Bacillus Subtilis Strain QST 713). EFS2 2021, 19. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.; Cattan, N.; Ruiz, B.; San Martin, J. Ability of Chilean Bacteria to Control Pseudomonas Syringae Pv. Actinidiae in Kiwifruit by Antibiosis and Induction of Resistance. J Plant Pathol 2019, 101, 849–883. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.; Lindow, S.; Sands, D.; Reifschneider, F.; Vanneste, J.; Lopes, C.; Rossato, M. The Plant Health, a View from the Plant Bacteriology; Moya-Elizondo, E.; Chillán, Chile., 2019; ISBN 978-956-401-337-4.
- Castro Tapia, M.P.; Madariaga Burrows, R.P.; Ruiz Sepúlveda, B.; Vargas Concha, M.; Vera Palma, C.; Moya-Elizondo, E.A. Antagonistic Activity of Chilean Strains of Pseudomonas Protegens Against Fungi Causing Crown and Root Rot of Wheat (Triticum Aestivum L.). Front. Plant Sci. 2020, 11, 951. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.A.; Cattan, N.C.; Arismendi, N.L.; Doussoulin, H.A. Determination of 2, 4-Diacetylphloroglucinol (2, 4-DAPG) and Phenazine-Producing Pseudomonas Spp. in Wheat Crops in Southern Chile. In Proceedings of the Phytopathology; AMER PHYTOPATHOLOGICAL SOC 3340 PILOT KNOB ROAD, ST PAUL, MN 55121 USA, 2013; Vol. 103, pp. 100–100.
- Ramette, A.; Frapolli, M.; Saux, M.F.-L.; Gruffaz, C.; Meyer, J.-M.; Défago, G.; Sutra, L.; Moënne-Loccoz, Y. Pseudomonas Protegens Sp. Nov., Widespread Plant-Protecting Bacteria Producing the Biocontrol Compounds 2,4-Diacetylphloroglucinol and Pyoluteorin. Systematic and Applied Microbiology 2011, 34, 180–188. [Google Scholar] [CrossRef]
- Weller, D.M.; Mavrodi, D.V.; Van Pelt, J.A.; Pieterse, C.M.J.; Van Loon, L.C.; Bakker, P.A.H.M. Induced Systemic Resistance in Arabidopsis Thaliana Against Pseudomonas Syringae Pv. Tomato by 2,4-Diacetylphloroglucinol-Producing Pseudomonas Fluorescens. Phytopathology® 2012, 102, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, J.; Li, J.; Shan, X.; Duan, Y. Endophytic Bacterium Pseudomonas Protegens Suppresses Mycelial Growth of Botryosphaeria Dothidea and Decreases Its Pathogenicity to Postharvest Fruits. Front. Microbiol. 2022, 13, 1069517. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, T.; Leni, G.; Bulla, G.; Giorni, P. Reduction of Mycotoxigenic Fungi Growth and Their Mycotoxin Production by Bacillus Subtilis QST 713. Toxins 2022, 14, 797. [Google Scholar] [CrossRef] [PubMed]
- Environmental Protection Agency (EPA) Bacillus Subtilis Strain QST 713 (PC Code 006479) Biopesticide Registration Action Document 2006.
- Silva-Valderrama, I.; Toapanta, D.; Miccono, M.D.L.A.; Lolas, M.; Díaz, G.A.; Cantu, D.; Castro, A. Biocontrol Potential of Grapevine Endophytic and Rhizospheric Fungi Against Trunk Pathogens. Front. Microbiol. 2021, 11, 614620. [Google Scholar] [CrossRef]
- Mesguida, O.; Haidar, R.; Yacoub, A.; Dreux-Zigha, A.; Berthon, J.-Y.; Guyoneaud, R.; Attard, E.; Rey, P. Microbial Biological Control of Fungi Associated with Grapevine Trunk Diseases: A Review of Strain Diversity, Modes of Action, and Advantages and Limits of Current Strategies. JoF 2023, 9, 638. [Google Scholar] [CrossRef]
- Guerrero, J.; Pérez, S. First Report of Diaporthe Australafricana -Caused Stem Canker and Dieback in European Hazelnut ( Corylus Avellana L.) in Chile. Plant Disease 2013, 97, 1657–1657. [Google Scholar] [CrossRef]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk Diseases With Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Disease 2018, 102, 12–39. [Google Scholar] [CrossRef]
- Kuntzmann, P.; Villaume, S.; Bertsch, C. Conidia Dispersal of Diplodia Species in a French Vineyard. Phytopathologia Mediterranea 2009, 48, 150–154. [Google Scholar]
- Chen, S.; Morgan, D.P.; Hasey, J.K.; Anderson, K.; Michailides, T.J. Phylogeny, Morphology, Distribution, and Pathogenicity of Botryosphaeriaceae and Diaporthaceae from English Walnut in California. Plant Disease 2014, 98, 636–652. [Google Scholar] [CrossRef]
- Chaverri, P.; Samuels, G.J.; Stewart, E.L. Hypocrea Virens Sp. Nov., the Teleomorph of Trichoderma Virens. Mycologia 2001, 93, 1113–1124. [Google Scholar] [CrossRef]
- Rinu, K.; Sati, P.; Pandey, A. Trichoderma Gamsii (NFCCI 2177): A Newly Isolated Endophytic, Psychrotolerant, Plant Growth Promoting, and Antagonistic Fungal Strain. J. Basic Microbiol. 2014, 54, 408–417. [Google Scholar] [CrossRef]
- Germain, E. THE REPRODUCTION OF HAZELNUT (CORYLUS AVELLANA L.): A REVIEW. Acta Hortic. 1994, 195–210. [CrossRef]
- Pasqualotto, G.; Carraro, V.; De Gregorio, T.; Huerta, E.S.; Anfodillo, T. Girdling of Fruit-Bearing Branches of Corylus Avellana Reduces Seed Mass While Defoliation Does Not. Scientia Horticulturae 2019, 255, 37–43. [Google Scholar] [CrossRef]
- Stolpe, N. Descripciones de Los Principales Suelos de La VIII Región de Chile; Publicaciones - Departamento de Suelos y Recursos Naturales; Trama Impresores S.A: Chillán, Chile.; Vol. 1;
- Aćimović, S.G.; Harmon, C.L.; Bec, S.; Wyka, S.; Broders, K.; Doccola, J.J. First Report of Diplodia Corticola Causing Decline of Red Oak ( Quercus Rubra ) Trees in Maine. Plant Disease 2016, 100, 649. [Google Scholar] [CrossRef]
- Antony, S.; Billones-Baaijens, R.; Steel, C.C.; Stodart, B.J.; Savocchia, S. Pathogenicity and Progression of Botryosphaeriaceae Associated with Dieback in Walnut Orchards in Australia. Eur J Plant Pathol 2024, 168, 723–742. [Google Scholar] [CrossRef]
- Paradinas, A.; Ramade, L.; Mulot-Greffeuille, C.; Hamidi, R.; Thomas, M.; Toillon, J. Phenological Growth Stages of ‘Barcelona’ Hazelnut (Corylus Avellana L.) Described Using an Extended BBCH Scale. Scientia Horticulturae 2022, 296, 110902. [Google Scholar] [CrossRef]
- Balzarini, M.; Gonzalez, L.; Tablada, M.; Casanoves, F.; Di Rienzo, J.; Robledo, C. InfoStat. User’s Manual 2008.
Figure 1.
Pre-selection assay of antagonist microorganisms against Diplodia mutila after 7 days of incubation. (a) Control: D. mutila; (b) Trichoderma spp. and Bacillus spp. [Puelche-VTO, Bio Insumos Nativa SpA]; (c) Bionectria ochroleuca Mitique, Trichoderma gamsii Volqui, Hypocrea virens Ñire [Mamull, Bio Insumos Nativa SpA]; (d) Bacillus subtilis QST 713 [Serenade ASO, Bayer]; (e) Pantoea sp. AP113. (f) Pseudomonas protegens Ca2+Ca6+ChC7; (g) P. protegens Ca2; (h) P. protegens Ca6; and (i) P. protegens ChC7.
Figure 1.
Pre-selection assay of antagonist microorganisms against Diplodia mutila after 7 days of incubation. (a) Control: D. mutila; (b) Trichoderma spp. and Bacillus spp. [Puelche-VTO, Bio Insumos Nativa SpA]; (c) Bionectria ochroleuca Mitique, Trichoderma gamsii Volqui, Hypocrea virens Ñire [Mamull, Bio Insumos Nativa SpA]; (d) Bacillus subtilis QST 713 [Serenade ASO, Bayer]; (e) Pantoea sp. AP113. (f) Pseudomonas protegens Ca2+Ca6+ChC7; (g) P. protegens Ca2; (h) P. protegens Ca6; and (i) P. protegens ChC7.
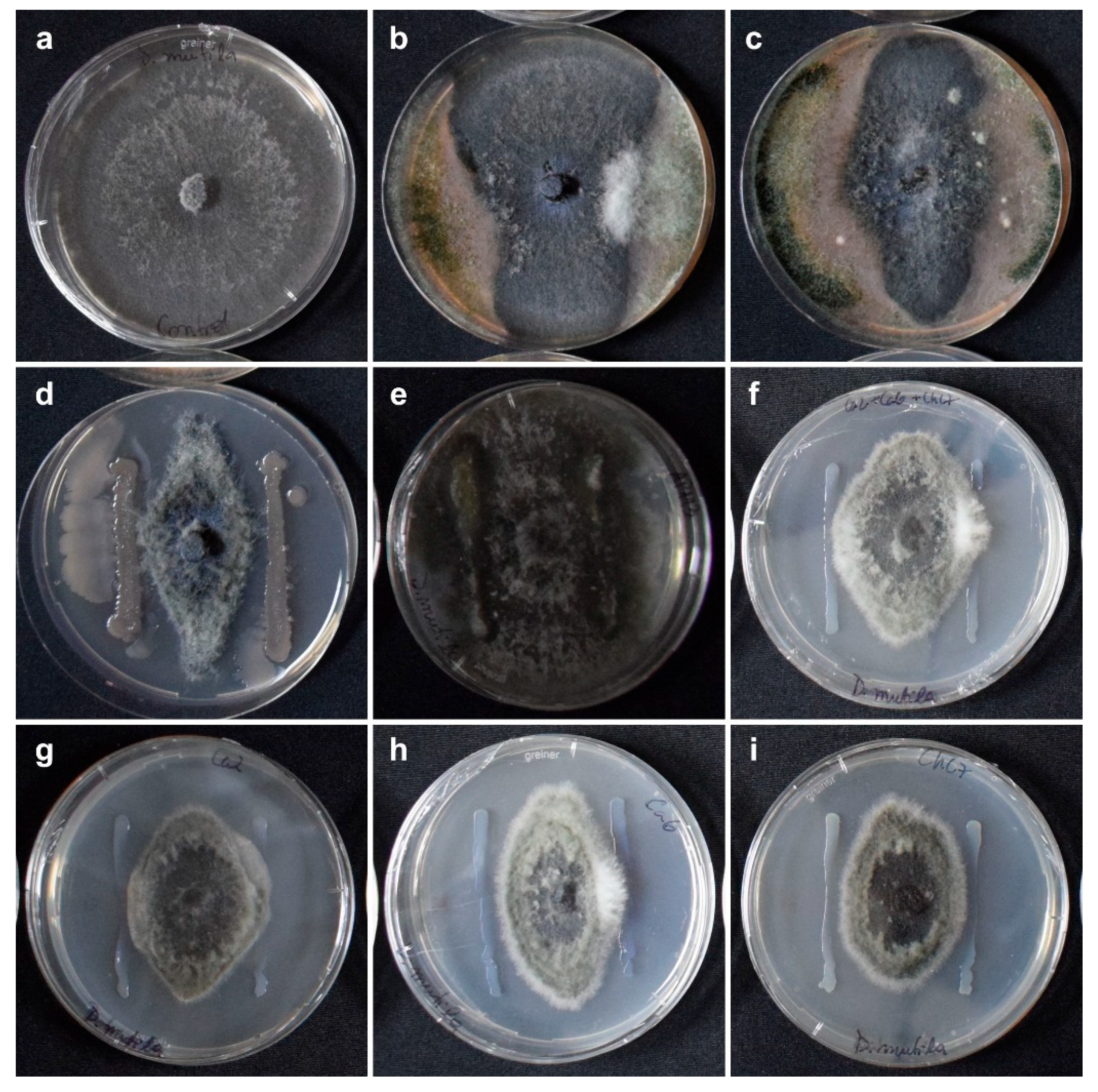
Figure 2.
Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni, inoculated the same day and 24 hours after the application of biological control agents, under field conditions during the 2020-2021 season. Bars with different letters indicate statistical differences between treatments according to Fisher’s LSD test (α = 0.05).
Figure 2.
Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni, inoculated the same day and 24 hours after the application of biological control agents, under field conditions during the 2020-2021 season. Bars with different letters indicate statistical differences between treatments according to Fisher’s LSD test (α = 0.05).
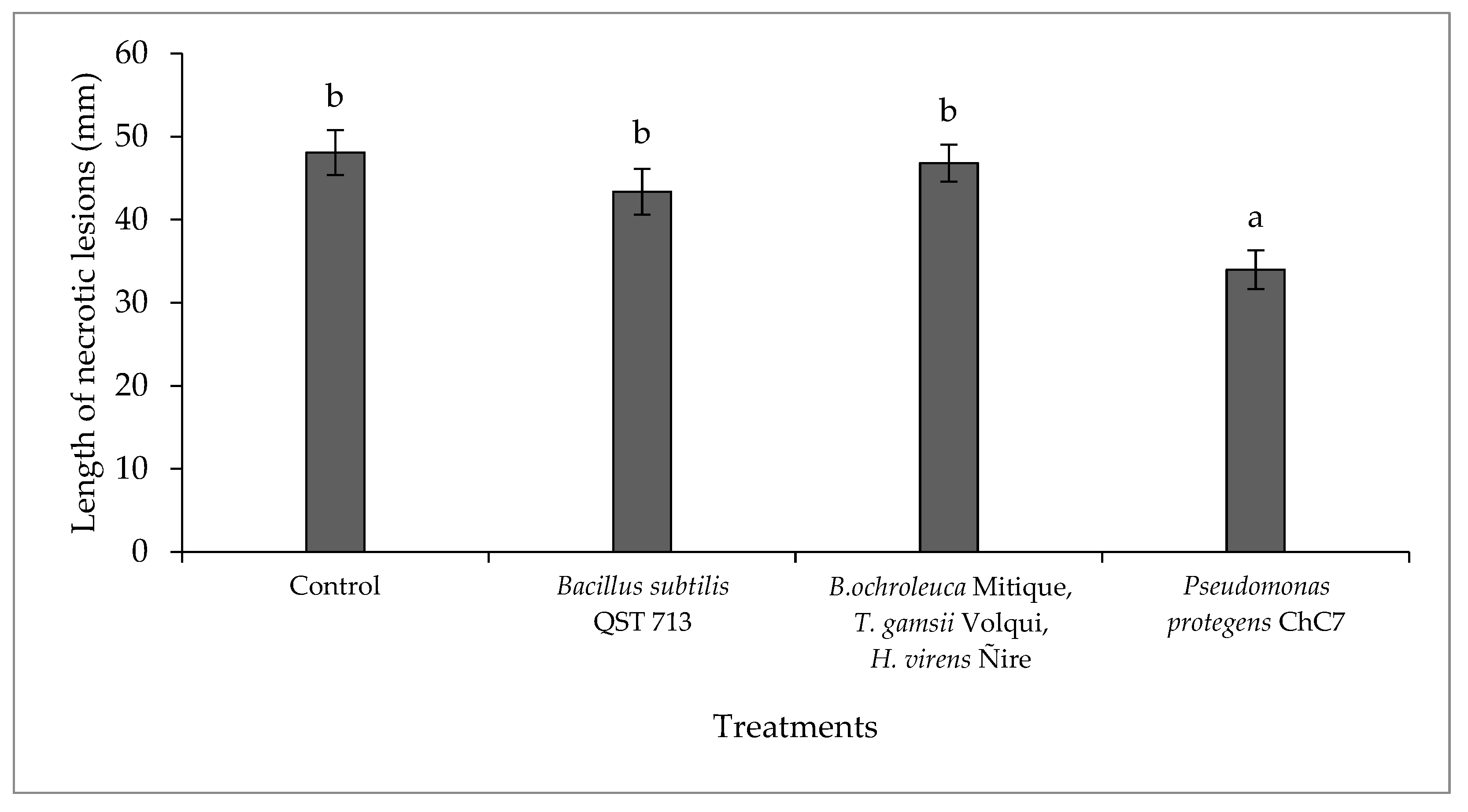
Figure 3.
A) Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni; B) inoculations on the same day (first inoculation) and 24 h later (second inoculation) after the application of biological control agents, under field conditions in the 2021-22 season. Bars with different letters indicate statistical differences between treatments (A) and inoculation times (B) according to Fisher’s LSD test (α = 0.05).
Figure 3.
A) Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni; B) inoculations on the same day (first inoculation) and 24 h later (second inoculation) after the application of biological control agents, under field conditions in the 2021-22 season. Bars with different letters indicate statistical differences between treatments (A) and inoculation times (B) according to Fisher’s LSD test (α = 0.05).
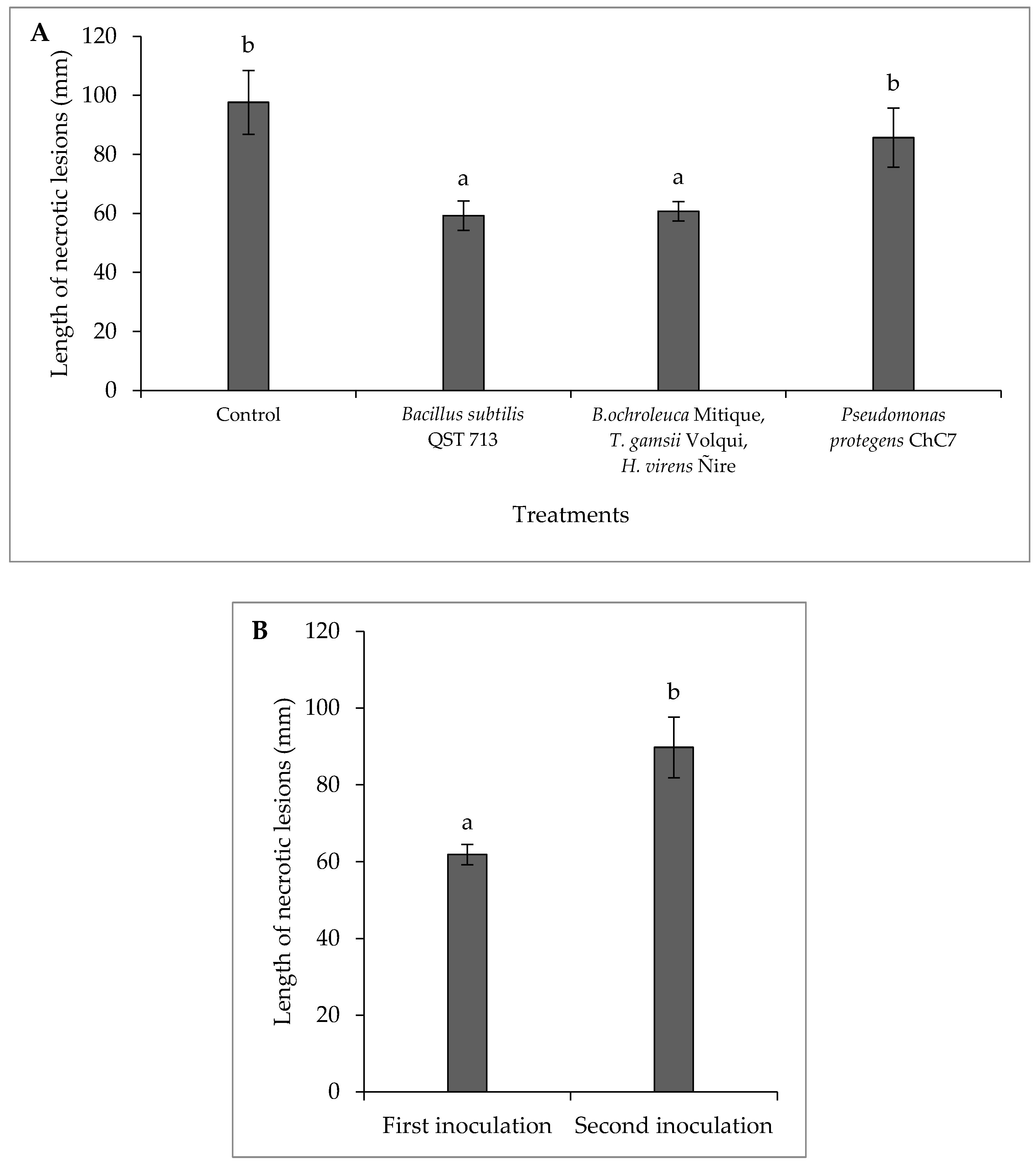

Table 1.
Degree of inhibition of in vitro mycelial growth of each fungicide on Diplodia mutila determined by mycelium development categories in the Petri dish at different incubation times.
Table 1.
Degree of inhibition of in vitro mycelial growth of each fungicide on Diplodia mutila determined by mycelium development categories in the Petri dish at different incubation times.
| Incubation time | |||
|---|---|---|---|
| Fungicide group / Active ingredient | 48 hours | 5 days | 14 days |
| (B) Methyl benzimidazole carbamate (MBC) fungicides | |||
| Methyl thiophanate | (++++)1 | (++++) | (++) |
| (C) Quinone outside inhibitor (QoI) Fungicides | |||
| Pyraclostrobin | (++) | (-) | (-) |
| Azoxystrobin/Difenoconazole | (++) | (+) | (-) |
| Trifloxystrobin | (+) | (-) | (-) |
| Kresoxim methyl/Miclobutanil | (++) | (-) | (-) |
| Kresoxim methyl/Tebuconazole | (+++) | (+) | (-) |
| (C) Succinate dehydrogenase inhibitor (SDHI) fungicides | |||
| Penthiopyrad | (++) | (-) | (-) |
| Fluxapyroxad/Pyraclostrobin | (+++) | (++) | (++) |
| Fluopyram/Tebuconazole | (+++) | (++) | (++) |
| (C) Not grouped | |||
| Fluazinam | (++++) | (+++) | (++) |
| (D) Anilinopyrimidine (AP) fungicides | |||
| Pyrimethanyl/Difenoconazole | (++) | (-) | (-) |
| (G) Demethylation inhibitor (DMI) fungicides (SBI: Class I) | |||
| Difenoconazole2 | (++) | (+) | (-) |
| Difenoconazole3 | (++) | (-) | (-) |
| Difenoconazole4 | (++) | (+) | (-) |
| Miclobutanil | (++) | (-) | (-) |
| Difenoconazole/Kresoxim methyl | (++) | (-) | (-) |
| Tebuconazole | (+++) | (++) | (++) |
| Prochloraz | (+++) | (+++) | (++) |
| (M) Dithiocarbamates and relatives (electrophiles) | |||
| Mancozeb | (++++) | (-) | (-) |
| Natural extracts & salts | |||
| Potassium Hydrogenicarbonate | (-) | (-) | (-) |
| Extracts of Quillaja saponaria and Yucca schidigera | (-) | (-) | (-) |
| Plant extracts and natural fatty acids | (+++) | (-) | (-) |
1 Mycelium development on PDA medium: (-) = Normal fungal growth, (+) = Moderate fungal growth (1 to 30% less than the normal fungal growth), (++) = Mild fungal growth (31 to 75% less fungal growth), (+++) = Limited fungal growth (76 to 99%), and (++++) = 100% inhibition of fungal growth.2 Difenoconazol 25 EC, Agrospec®; 3 Dominio 25 EC, Anasac; 4 Score® 250 EC, Syngenta.
Table 2.
Mycelial inhibition percentage from different bacterial strains with antagonistic activity against fungus Diplodia mutila after seven days of incubation under in vitro conditions. Different letters indicate significant differences between treatments according to the non-parametric Kruskal-Wallis test (p ≤ 0.05).
Table 2.
Mycelial inhibition percentage from different bacterial strains with antagonistic activity against fungus Diplodia mutila after seven days of incubation under in vitro conditions. Different letters indicate significant differences between treatments according to the non-parametric Kruskal-Wallis test (p ≤ 0.05).
| Bacterial strain | Percentage of mycelial inhibition | |
|---|---|---|
| Pseudomonas protegens Ca2 | 41.4 | abc |
| Pseudomonas protegens Ca6 | 37.9 | ab |
| Pseudomonas protegens ChC7 | 45.3 | bc |
| P. protegens strains Ca2+Ca6+ChC7 | 42.2 | abc |
| Pantoea sp. AP113 | 6.3 | a |
| Bacillus subtilis QST 713 | 51.9 | c |
| Interquartile range (IQR) | 28.2 | |
| P-value | 0.02 | |
Table 3.
Mean length values of necrotic lesions caused by Diplodia mutila on branches of hazelnut plants cv. Barcelona and Tonda di Giffoni (average of both cultivars), sprayed with different fungicides and inoculated immediately after application under controlled pot conditions. Different letters indicate statistical differences between treatments according to Fisher’s LSD test (α = 0.05).
Table 3.
Mean length values of necrotic lesions caused by Diplodia mutila on branches of hazelnut plants cv. Barcelona and Tonda di Giffoni (average of both cultivars), sprayed with different fungicides and inoculated immediately after application under controlled pot conditions. Different letters indicate statistical differences between treatments according to Fisher’s LSD test (α = 0.05).
| Treatments | Mean length (mm) of necrotic lesions ж in inoculated branches | |
|---|---|---|
| Water control | 50.9 | d |
| Bacillus subtilis QST 713 | 33.9 | bc |
|
Bionectria ochroleuca Mitique Trichoderma gamsii Volqui Hypocrea virens Ñire |
23.1 | ab |
| Pseudomonas protegens ChC7 | 26.9 | bc |
| Fluazinam | 23.9 | ab |
| Fluopyram/Tebuconazole | 25.7 | abc |
| Fluxapyroxad/Pyraclostrobin | 19.9 | a |
| Prochloraz | 34.6 | c |
| Tebuconazole | 31.1 | bc |
| Coefficient of variation | 9.5 | |
| P-value | <0.01 | |
Table 4.
Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni, with inoculations on the same day and 24 h after fungicide application, under field conditions in the 2020-21 season. Row with different letters indicate statistical differences between inoculation times according to Fisher’s LSD test (α = 0.05).
Table 4.
Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni, with inoculations on the same day and 24 h after fungicide application, under field conditions in the 2020-21 season. Row with different letters indicate statistical differences between inoculation times according to Fisher’s LSD test (α = 0.05).
| Mean length of necrotic lesions (mm) | ||
|---|---|---|
| Treatments | Inoculation on the day of fungicide application | Inoculation 24 h after fungicide application |
| Control | 48.1 | 45.4 |
| Fluazinam | 46.7 | 45.6 |
| Fluopyram/Tebuconazole | 60.7 | 36.0 |
| Fluxapyroxad/Pyraclostrobin | 45.0 | 32.5 |
| Prochloraz | 45.4 | 41.5 |
| Tebuconazole | 50.9 | 39.3 |
| Average | 49.5 B | 40.1 A |
| Coefficient of variation | 6.34 | |
| P-value Inoculation time | 0.008 | |
Table 5.
Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni with inoculations on the same day of applications and 24 h after fungicide application, under field conditions in the 2021-22 season. Different lowercase letters indicate significant differences between treatments, and different uppercase letters indicate significant differences between inoculation times within each treatment, according to Fisher’s LSD test (α = 0.05).
Table 5.
Mean length values of necrotic lesions caused by the fungus Diplodia mutila on branches of hazelnut cv. Tonda di Giffoni with inoculations on the same day of applications and 24 h after fungicide application, under field conditions in the 2021-22 season. Different lowercase letters indicate significant differences between treatments, and different uppercase letters indicate significant differences between inoculation times within each treatment, according to Fisher’s LSD test (α = 0.05).
| Mean length of necrosis (mm) | ||||||
|---|---|---|---|---|---|---|
| Treatments | Inoculation on the day of fungicide application | Inoculation 24 h after fungicide application | ||||
| Control | 71.8 | c | A | 123.8 | b | B |
| Fluazinam | 60.2 | bc | A | 108.6 | b | B |
| Fluopyram/Tebuconazole | 45.3 | a | A | 87.8 | a | B |
| Fluxapyroxad/Pyraclostrobin | 48.7 | a | A | 80.8 | a | B |
| Penthiopyrad | 45.5 | a | A | 121.8 | b | B |
| Prochloraz | 54.3 | ab | A | 113.0 | b | B |
| Tebuconazole | 47.9 | a | A | 76.9 | a | B |
| Average | 72.2 | 101.8 | ||||
| Coefficient of variation | 2.75 | |||||
| P-value T x I | < 0.01 | |||||
Table 6.
Chemical fungicides and biological control agents selected in the in vitro assays and further evaluated under controlled and field conditions.
Table 6.
Chemical fungicides and biological control agents selected in the in vitro assays and further evaluated under controlled and field conditions.
| Active ingredient | Formulation | Trade name | Manufacturer | Commercial dose |
|---|---|---|---|---|
| Fluazinam | 500 g L-1 SC | Shirlan ® 500 | Syngenta | 75 mL hL-1 (Apple trees) |
| Fluopyram/Tebuconazole | 200/200 g L-1 SC | Luna ® Experience 400 | Bayer | 40 mL hL-1 (Stone and pome trees) |
| Fluxapiroxad/Pyraclostrobin | 250/250 g L-1 SC | Elmus ® | BASF | 40 mL hL-1 (Stone and pome trees) |
| Penthiopyrad | 200 g L-1 SC | Fontelis ® | Corteva | 40 mL hL-1 (Apple trees) |
| Prochloraz | 400 g L-1 EC | Mirage 40% | Adama | 120 mL hL-1 (Wheat and barley) |
| Tebuconazole | 430 g L-1 SC | Tebuconazole 430 | Agrospec | 40 mL hL-1 (Stone trees) |
| Bacillus subtilis QST 713 | 13,68 g L-1 SC (1 x 109 cfu g-1) |
Serenade ® ASO | Bayer | 600 mL hL-1 (Stone and pome trees) |
|
Bionectria ochroleuca Mitique Trichoderma gamsii Volqui Hypocrea virens Ñire |
10/10/10 g kg-1 WG (3 x 108 cfu g-1) |
Mamull ® | Bio Insumos Nativa |
100 g hL-1 (Stone and pome trees) |
| Pseudomonas protegens ChC7 | 1 x 107 | N/D1 | N/D | 1000 mL hL-1 |
1N/D = Name not defined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



