
Submitted:
20 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Pigments are chemical compounds that impart color to materials via absorption, reflection, and refraction. Traditional sources of natural pigments include plant and insect tissues; however, microorganisms such as bacteria, yeasts, algae, and filamentous fungi also hold potential for pigment production. In this study, we investigated pigment production in 20 isolates from our in-house Actinobacteria strain collection, obtained from the banks of the Guaviare and Arauca Rivers in Colombia. Isolates were identified using 16S rRNA gene sequencing. We evaluated different culture media and determined their various bioactivities, including antioxidant, antibacterial, and cytotoxic properties. Promising pigmented hydroalcoholic extracts exhibiting the above-mentioned properties were identified and their chemical compositions were examined using LC-MS. Our findings highlighted a growth medium-dependent effect, as pigment production, coloration, and bioactivity outcomes were influenced by growth media. These results demonstrate the significant potential of Streptomyces isolates as sources of bioactive pigments for diverse applications.
Keywords:
Streptomyces
; Antibacterial
; Pigment
; Antioxidant
; Cytotoxic
1. Introduction
Pigments are colored compounds that should not be physically or chemically affected by the medium in which they are incorporated [1], and have been used since ancient times by civilizations such as China, India, and Egypt [2]. Currently, they are widely employed in various industries, including food, textiles, paints, cosmetics, pharmaceuticals, and plastics [3]. The global market for natural pigments in the food industry is estimated to reach $2.5 billion by 2025 [2].
Pigments can be classified, according to their origin, as natural or synthetic. Natural pigments are further categorized based on their specific origin (plant, mineral, microbial, or animal), color formation, chemical constitution, and application method. Examples of natural pigments include carotenoids, melanins, chlorophylls, flavonoids, anthocyanins, betalains and quinones. Synthetic pigments, on the other hand, are generally classified into cationic, anionic, and nonionic types, and by their applications as direct, acidic, basic, chrome mordants, azo, sulfur, reactive, and dispersed [4].
Despite their origins, pigments can be harmless or toxic to some degree. However, consumers often prefer natural pigments because they are considered safe, non-toxic, non-carcinogenic, and biodegradable [5]. In contrast, synthetic pigments can pose serious human health hazards and environmental risks [3,6] owing to their non-biodegradable nature [7]. They can cause adverse effects [8] such as decreased light penetration in water bodies [9] and increased chemical and biochemical oxygen demand (BOD and COD), and some are bioaccumulative, toxic, mutagenic, or carcinogenic [10] even at very low concentrations.
Natural pigment production is widespread among plants, microalgae, fungi, bacteria, and insects. Among bacterial groups, pigment production is particularly notable in actinobacteria [11], which is highly profitable and biotechnologically valuable [12,13]. Actinobacteria produce metabolites with unique chemical structures and diverse bioactivities, including antibiotics, biopesticides, phytohormones, antitumor compounds, antiviral agents, enzymes, enzyme inhibitors, anti-inflammatory agents, and biosurfactants [14,15,16]. Streptomyces, in particular, is known to produce a large number of specialized metabolites with different properties [13,17,18,19]. In this regard, several genera such as Streptomyces, Nocardia, Micromonospora, Thermomonospora, Actinoplanes, Microbispora, Streptosporangium, Actinomadura, Rhodococcus, and Kitasatospora [20] include microorganisms that produce a wide variety of pigments. The most reported pigment-producing genus is Streptomyces, including species such as S. griseus, S. griseoviridis, S. coelicolor [21,22], S. cyaneus [23], S. vietnamensis [24], S. echinoruber [25], S. shaanxiensis [26], and S. caeruleatus [27].
Microbial pigments are emerging as alternatives to synthetic pigments, offering several advantages such as abundant raw materials independent of season, high yields, higher concentrations with simpler purification processes, regulatory approval, high stability to environmental factors [6,28,29,30,31], and biodegradability [32]. Additionally, some microbial pigments possess antioxidant, anti-inflammatory, anticancer, and antimicrobial properties [30,33] with potential applications in food colorants, nutraceuticals, and cosmetics [32].
Actinobacteria are widely distributed in diverse natural ecosystems, including soil, plants, limestone, fresh and seawater, sponges, volcanic caves, deserts, air, and insects [16]. Studies of actinobacteria from unexplored or underexploited habitats as new sources of specialized metabolites have increased [13]. Colombia, known for its megadiversity in fauna and flora, also exhibits a rich diversity of microorganisms [34].
Given this background, this study evaluated the bioactive potential of pigmented extracts from Streptomyces isolated from the Guaviare and Arauca rivers of Colombia. Hence, pigmented ethanolic extracts were prepared and evaluated for their antioxidant capacity using 2,2-diphenyl-1-picrylhydrazyl (DPPH) and 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) assays, antibacterial activity by disc diffusion assay, and cytotoxic activity by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay against human dermal fibroblasts (HDFa), cervical-uterine cancer (HeLa), and MCF-7 (breast cancer) cell lines. Therefore, the aim of this study was to explore and characterize the bioactive properties of pigmented extracts of selected Streptomyces strains isolated from these unique Colombian habitats. By assessing their antioxidant, antibacterial, and cytotoxic activities, this study seeks to identify promising sources of natural pigments with potential applications in various industries, particularly cosmetics and pharmaceuticals. This study contributes to the growing field of natural product chemistry and highlights the significance of Actinobacteria as a valuable resource for biotechnological advancements.
2. Materials and Methods
2.1. Reagents and Culture Media
The components and culture media used were as follows: agar (DIBICO, Sta. Cruz Mexico), starch (Carlo Erba, Eure, France), glucose, CaCO3, K2HPO4, (NH4)2SO4, CuSO4.5H2O, FeSO4.7H2O, KNO3, MgSO4.7H2O, MnCl2.4H2O, ZnSO4.7H2O (PanReac, Barcelona Spain), yeast extract (OXOID, Hampshire UK), malt extract (CDH, New Delhi India), NaCl (Honeywell, Seelze Germany), KH2PO4 (DUKSAN, Kyungkido Korea), and TSA (Criterion, California USA). DPPH, ABTS, trolox, methanol, and ethanol were purchased from MilliporeSigma (St. Louis, MO, USA). MTT was acquired from Thermo Fisher Scientific Inc. (Waltham, MA USA).
2.2. Strains, Media and Culture Conditions
The Bioprospecting Research Group at Universidad de La Sabana has a collection including 380 isolates from sediments of the Guaviare riverbank and 780 isolates from sediments of the Arauca riverbank with Streptomyces-like morphological characteristics. These isolates have been cultured, isolated, and cryogenically preserved at -80°C in ISP2 medium. The colored isolates (20 isolates) were grown in four different culture media (see Table 1) at pH 7, in an incubator at 30°C for 7 days (Friocell, Querétaro, Mexico).
The pathogens employed were Klebsiella pneumoniae ATCC11486, Escherichia coli ATCCBAA2469, Pseudomonas aeruginosa ATCC27853, Bacillus subtilis ATCC 55033, Staphylococcus epidermidis ATCC14990, and Staphylococcus aureus ATCCBAA44. These pathogens were cultured on trypticase soy agar (TSA) at 37°C for 24 h.
2.3. Fermentation of Isolates
The colored isolates were activated on the isolation medium (ISP2) by seeding the spores and incubating for 7 days at 30°C. They were then seeded on the other three culture media (ISP4, modified ISP9, and starch nitrate). Media in which the isolate did not grow or did not produce pigment were discarded and re-evaluated in liquid culture. A 1-cm² section was cut from each medium and placed in 3 mL of the same medium in liquid form, and these cultures were incubated at 30°C for 7 days with agitation at 150 rpm (New Brunswick Scientific, Connecticut, USA). The fermentation volume of the cultures was then increased by maintaining a proportion of 10% of the liquid culture to 90% of new liquid medium until a fermentation volume of 100 mL was achieved.
2.4. Preparation of extracts from pigmented strains
The final fermentation volume (100 mL) of each strain was distributed in 50 mL Falcon tubes and centrifuged (Thermo Scientific, Massachusetts, USA) at 6000 xg for 10 minutes. The phase (supernatant and/or sediment) containing the pigment was lyophilized for 24 hours (Labconco, Kansas, USA). A double extraction of the lyophilizate from each Falcon tube was performed: first, 75% ethanol (10 mL) was added, vortexed, and agitated for 24 h at 150 rpm. The mixture was then centrifuged at 6000 xg for 5 min, and the supernatants were placed in amber vials. The procedure was repeated with an additional volume of 75% ethanol (5 mL) for the second extraction.
2.5. Biological Activity Tests
2.5.1. Antioxidant Capacity
Two methods were performed to determine the antioxidant capacity, i.e, DPPH and ABTS-related procedures, using the respective stable radicals for scavenging capacity. In both cases, the antioxidant capacity of the extracts was evaluated at concentrations of 5000, 500, 50, and 5 ppm in 96-well plates in triplicate, using trolox as a positive control.
For DPPH, the extract (100 µL) and 0.2 mM DPPH (100 µL) were added to a 200-µL well. The plate was incubated for 30 minutes in the dark, and absorbance was measured at 515 nm. For the ABTS method, aqueous solutions of 7 mM ABTS and 2.45 mM ammonium persulfate were separately prepared and subsequently mixed (1:1 ratio) and incubated in the dark for 16 h. The resulting ABTS+· radical was adjusted to an absorbance of 0.7. Finally, the extract (10 µL) and ABTS+· radical solution (190 µL) were added to a 200-µL well. It was incubated for 30 min in the dark and the absorbance was measured at 734 nm (Bio-Rad, California USA) [38,39].
2.5.2. Antibacterial Activity
The pigmented extracts were evaluated against the pathogens K. pneumoniae ATCC11486, E. coli ATCCBAA2469, P. aeruginosa ATCC27853, Bacillus subtilis ATCC 55033, S. epidermidis ATCC14990, S. aureus ATCCBAA44 using antibiograms by disk diffusion. Solutions of the extract (30 mg/mL) were prepared in 75% ethanol, and pathogen dispersions were prepared using the MacFarland scale 0.5% in 0.85% saline solution. Mass seeding of pathogenic bacteria was performed using a sterile swab in TSA plates. The discs were impregnated with 5, 15, and 30 µL of the pigment solutions to examine the variation of bioactivity depending on pigment amounts in the range of 150 µg to 900 µg. The tests were performed in triplicate. Finally, the TSA plates were incubated at 37°C for 24 h (Friocell, Querétaro Mexico) using the extraction solvent (75% ethanol) as a negative control and the respective antibiotic as a positive control. The antibiograms were photographed in the colony counter ProtoCOL3 (Microbiology International, Maryland USA) for the measurement of the inhibition halos using the ImageJ program [40].
2.5.3. Cytotoxic Activity
The cytotoxic activity of the pigmented extracts was evaluated against cancer lines such as HeLa (cervical-uterine cancer) and MCF-7 (breast cancer), and healthy cell lines like HDFa (human dermal fibroblasts), using the standard MTT assay. Cells were cultured in DMEM medium for HeLa and HDFa, and RPMI for MCF-7, with 10% fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin, and incubated at 37°C in a 5% CO2 incubator. After incubation, cells were treated with different concentrations of the pigmented extracts in triplicate (5-500 ppm) and incubated for 24 h. Following the 24-h treatment, MTT solution (100 µL) at 5 mg/mL was added and incubated for 4 h. MTT was removed, and dimethyl sulfoxide (DMSO) (100 µL) was added to each well. The assay was measured at an absorbance of 570 nm using a plate reader (Bio-Rad, California, USA), with DMSO as the control [41].
2.6. Molecular Identification
At the end of the seven-day incubation period, a spore solution of each isolate was prepared, and genomic DNA was extracted using a Quick-DNA Fungal/Bacterial Microprep kit (Zymo Research Corporation, Irvine, CA, USA) according to the manufacturer’s instructions. Molecular identification was carried out by sequencing a 1500 bp fragment of the 16S ribosomal RNA gene obtained by PCR amplification using the universal primers 1493R and 27F, under the following PCR cycling conditions: initial denaturation at 94°C for 3 minutes, followed by 30 cycles of 94°C for 1 minute, 50°C for 1 minute, and 72°C for 2 minutes, with a final extension of 72°C for 7 minutes (Bio-Rad, California, USA). The amplification products were verified by agarose electrophoresis (Bio-Rad, California, USA). The sequence obtained was compared in the GenBank nucleotide sequence database. A phylogenetic tree was constructed using the maximum likelihood method and Hasegawa-Kishino-Yano model [42] in MEGA X [43].
2.7. High-Performance Liquid Chromatography Coupled with Mass Spectrometry
The hydroalcoholic extracts were chemically characterized using a Shimadzu Corp LCMS-2020 system (Japan). This system featured a single quadrupole analyzer and a dual ion source (DUIS) that performs electrospray (ESI) and atmospheric pressure chemical (APCI) ionizations. The testing procedure employed a Phenomenex Synergy RP C18 column (150 × 4.4 mm, 2.6 µm). The column was heated to 30°C and maintained a flow rate of 0.7 mL/min. Two mobile phases, labeled A and B, consisted of 0.1% formic acid in water and 0.1% formic acid in acetonitrile, respectively. A gradient elution was conducted over 30 min, starting at 10% B and gradually increasing to 100% B until 25 min, and then returning to 10% B until 30 min. The DUIS was operated in both positive and negative ion modes, employing a scan range of 200–1000 m/z, a CDL temperature of 250°C, a detector voltage of 1.2 kV, a nebulizing gas flow rate of 1.3 L/min, and a drying gas flow rate of 8.0 L/min [44,45]. The characterization was supplemented by HRMS using a Shimadzu HPLC system coupled with a Bruker MicrOToF-Q II spectrometer equipped with a Quadrupole-Time of Flight (QToF) analyzer and ESI. Chromatographic parameters identical to those previously mentioned were employed, operating ESI in both positive and negative ion modes, scanning between 100 to 1000 m/z, and setting capillary voltage at 4.5 kV, desolvation line temperature at 400 °C, nebulizer gas (N2) at 4 Bar, drying gas at 8 L/min, and quadrupole and collision energies at 6 and 12 eV, respectively.
The raw data were pre-processed using MZmine 3 software [46], and the detected features were annotated (level 3) from the MS data using the StreptomeDB v3.0 database [47]. The possible metabolites responsible for the pigmentation or tested bioactivities were searched and additionally filtered using StreptomeDB and other databases such as PubChem [48], LOTUS [49] and the natural products atlas [50,51].
3. Results and Discussion
3.1. Pigmentation of the Strains in the Different Culture Media
Evaluating different culture media is crucial as the production of specialized metabolites, including pigments, is influenced by changes in the primary carbon and nitrogen sources. These changes can either increase the supply or limit access to key precursors [52,53,54], triggering the activation of biosynthetic enzymes or simultaneously affecting both processes [55,56]. Bacteria often prefer certain single carbon sources over others, preventing the synthesis of enzymes necessary for the catabolism of non-selected carbon sources through carbon catabolite repression (CCR) [57]. Similarly, nitrogen source preference (nitrogen repression) affects the expression of primary nitrogen metabolism and subsequently the biosynthesis of specialized metabolites, which require nitrogen compound precursors [56,58].
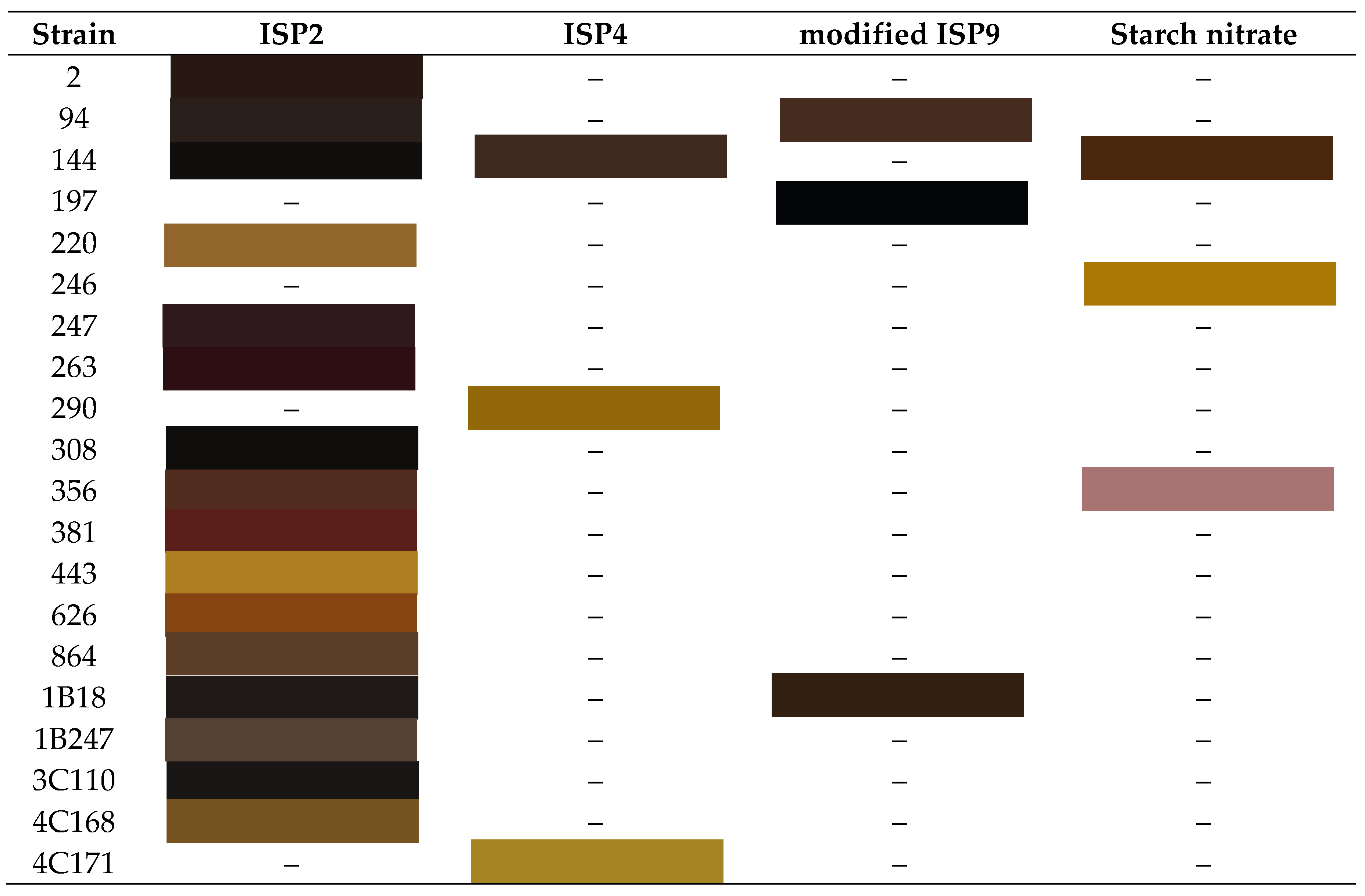
When evaluating pigment production of each strain in different culture media (see Table 1), it was observed that the highest number of pigments were produced in ISP2, which contains malt and yeast extract providing amino acids, peptides, vitamins, and minerals essential for bacterial growth [59,60,61,62], while glucose supplies the necessary energy for specialized metabolite synthesis like pigments [63,64]. This medium has previously been used to produce red pigments with anticancer activity from Streptomyces A 16-1 [65] and Streptomyces sp. PM4 [66], brown pigments with antibacterial and anti-HIV activities from Streptomyces sp. S45 [67], melanoid pigments like eumelanin with antiproliferative and antioxidant capacities from S. parvus BSB49[68], and pyomelanin from Streptomyces sp. MPPS2 [69].
The predominant colors produced were black and brown. These dark pigments, possibly melanin or melanoid, can be useful criteria for taxonomical studies [70]. However, varied colors such as yellow, pink, and purple were also observed in other culture media. The common nutrient for these particular pigments is starch as a carbon source, which can induce the expression of genes involved in specialized metabolite biosynthesis and ensure a sustained release of glucose for growth and metabolite production [71,72,73,74]. This carbon source has been used in starch casein nitrate medium to produce red pigment with antimicrobial and anticancer activities from Streptomyces sp. JAR6 [75], yellow pigments with antimicrobial activity from S. coeruleorubidus [76] and S. parvulus C5-5Y [77] and with anticancer activity from S. griseoaurantiacus JUACT 01 [41] and S. aurantiacus AAA5 [78].
It should be noted that pigment production in the starch nitrate culture medium is slow, but it has the advantage that pigment production is greater than the amount of biomass produced. This may be due to the fact that biosynthesis of specialized metabolites likely competes with biomass formation, slowing growth [56]. Additionally, some strains that produced pigment in agar lost their production in liquid medium, and when fermentation volumes were increased, a decrease in pigment production was observed, or the coloration became less visible compared to the culture medium color.

Table 2.
Pigmentation produced by the strains in the different culture media. ((–) = no pigmentation).
Table 2.
Pigmentation produced by the strains in the different culture media. ((–) = no pigmentation).
 |
3.2. Molecular Identification
The characterization of Streptomyces strains using 16S rRNA gene sequencing is a fundamental approach in microbial taxonomy and phylogenetics. This method provides a high level of accuracy in identifying and classifying bacterial species based on their genetic sequences [80,81]. In this study, a BLAST search of the GenBank database using the 16S rRNA gene sequences for each pigmented strain confirmed that all identified isolates belong to the genus Streptomyces. The 16S rRNA gene sequence similarities between the strains and type strains of the genus Streptomyces ranged from 98% to 100% (Table 4). This high similarity level indicated a close genetic relationship among the strains, aligning with previous findings in the literature where 16S rRNA gene sequencing has been effectively used to delineate species within the Streptomyces genus (Table 4).
A phylogenetic tree was constructed based on 16S rRNA gene sequences of Streptomyces species using the Maximum Likelihood method and Hasegawa-Kishino-Yano model [42] in MEGA X [43]. The bootstrap consensus tree, inferred from 1000 replicates, is presented in Figure 1, with the percentage of replicate trees supporting the clustered taxa shown next to the branches [79]. In the phylogenetic tree, some strains cluster closely together, such as strains S. lacticiproducens 864, S. albospinus 381, and S. humi 144, and strains S. murinus 4C168, S. humi 308, and S. mediolani 1B247. However, there is no clear pattern in strain relationships, with the most promising yellow pigment-producing strains (S. noursei 290, S. murinus 246 and 4C171) being relatively distant from each other.
Some strains with identical BLAST identifications are found in different branches of the phylogenetic tree and exhibit varying bioactivities. This suggests that more extensive morphological, biochemical, or sequencing analyses might be necessary for accurate identification. Additionally, the novel sources of isolation could indicate that some strains represent new identifications. The use of 16S rRNA gene sequencing is well-documented in the literature for its robustness in identifying bacterial strains [80]. This gene serves as a reliable molecular marker due to its presence in all bacteria and the slow rate of evolutionary change in its sequence, making it ideal for phylogenetic studies. Similarly, it has been previously demonstrated the utility of 16S rRNA gene sequencing in the classification of Streptomyces species, highlighting its effectiveness in distinguishing between closely related strains [81]. However, despite its strengths, 16S rRNA gene sequencing has limitations [80]. The resolution of this method may not be sufficient to differentiate between very closely related species or strains with highly similar sequences. Consequently, additional genetic markers or whole-genome sequencing might be required for more precise identification and classification. Hence, while 16S rRNA gene sequencing is a powerful tool for the identification and characterization of Streptomyces strains, combining this method with other genetic and phenotypic analyses enhances the accuracy and depth of microbial taxonomy studies [82]. This integrative approach is crucial for uncovering the full potential of Streptomyces species in producing bioactive compounds.
3.3. Biological Activity Assessment
3.3.1. Antioxidant Capacity
Antioxidants are crucial for preventing oxidative stress, which can cause damage to lipids, proteins, and DNA, leading to diseases such as cancer, neurodegeneration, and cardiovascular conditions [83,84]. Various in-vitro and in-vivo methods are utilized to evaluate antioxidant capacity, playing a significant role in understanding how antioxidants function [85]. Two commonly used methods for evaluating antioxidant capacity are the DPPH and ABTS assays (see Figure 2). Differences between results from these methods may arise because the ABTS assay is applicable to both hydrophilic and lipophilic antioxidant systems, while the DPPH assay is applicable only to hydrophobic systems [86,87,88]. Additionally, it is suggested that the ABTS assay better reflects antioxidant contents than the DPPH assay [89], being highly sensitive, practical, fast, very stable [90] and useful for high-moisture or highly pigmented nutrients [91].
In most pigmented extracts, a higher test concentration exhibits a higher antioxidant capacity, indicating a dose-dependent response [92]. Differences in antioxidant capacities determined by the two assays can be greater in highly pigmented sources [86]. According to the DPPH assay, extracts with the highest antioxidant potential (TEAC > 20 µM) are from strains S. humi 144 in ISP4, S. misionensis 197 in modified ISP9 (mISP9), S. hygroscopicus 356 in ISP2, and S. lactacystinicus 1B18 in mISP9. These extracts have dark brown or black colors, possibly indicating melanin pigments. Previously, Streptomyces strains have reported melanin with antioxidant capacity, such as Streptomyces girseorubiginosus D5 isolated from desert soil in India [93], Streptomyces glaucescens NEAE-H isolated from Gulbarga region in India [36], and Streptomyces parvus BSB49 from Bayburt province (Turkey) [68]. In the case of the ABTS assay, almost all extracts at maximum concentration exhibit good antioxidant capacities except for strains S. murinus 246 and S. humi 144 in starch nitrate medium, and strains S. noursei 290 and S. murinus 4C171 in ISP4, which resulted in TEAC < 20 µM. Notably, three of these are yellow pigments with low antioxidant capacity in both methods (see Table S1 and Table S2).
The use of different culture media affects the antioxidant capacities and specialized metabolite profiles [52,53,54], and explain the differences between the two methods, noting that a hydrophilic antioxidant may not be detected by the DPPH method, whereas the ABTS method could measure easier its scavenging capacity [86]. The strain S. tendae 94-derived extract shows slight changes in antioxidant capacity between ISP2 and mISP9 media in both methods. The strain S. humi 144-derived extract evaluated in three media shows significant differences: in ISP2 and ISP4, the DPPH-scavenging variation is slight (6.86 µM) and larger in ABTS (16.95 µM). However, in starch nitrate medium, the change was considered drastic with null DPPH and low ABTS-scavenging ability. Similarly, strain S. hygroscopicus 356-derived extracts show high DPPH capacity when cultured in ISP 2 and almost null in starch nitrate, while ABTS assay shows antioxidant capacity with a difference of 9.59 µM.
3.3.2. Antibacterial Activity
The discovery and development of new antibiotics are crucial due to the increasing threat of antimicrobial resistance (AMR). This problem is exacerbated by the overuse and misuse of antibiotics, inappropriate waste management, and environmental transmission [94,95]. It is estimated that at least 700,000 people worldwide die each year as a result of drug-resistant infections [94]. Additionally, the high numbers of secondary infections during COVID-19 pandemic [96,97] and the massive use of antibiotics as (co)treatment [98] worsened the situation, adding to the ongoing emergence of AMR [99], with likely unfavorable outcomes in clinical, economic, and societal aspects [94,100,101].
Among the six pathogenic microorganisms tested, only three—B. subtilis ATCC 55033, S. epidermidis ATCC 14990, and S. aureus ATCC BAA44—demonstrated sensitivity to the pigmented extracts (see Table 3). The last two are of clinical interest. In this regard, S. epidermidis is part of the human skin microbiome but can cause opportunistic infections, such as nosocomial infections, particularly in immunocompromised patients [102,103]. It may outcompete other bacteria in skin wounds and can acquire antibiotic resistance through horizontal gene transfer from other Staphylococcus species [104,105]. On the other hand, S. aureus ATCC BAA44 is an Iberian MRSA clone isolated from a hospital. It can lead to a range of infections, including those affecting the skin, subcutaneous tissues, and potentially invasive infections such as osteomyelitis, meningitis, pneumonia, lung abscess, empyema, and endocarditis [106,107,108].
Table 3.
Antibacterial activity of the pigmented extracts.
| Pigmented Extracts |
Tested bacterial strains | Pigmented Extracts |
Tested bacterial strains | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | ||
| 2-A | – | – | – | – | – | – | 356-A | – | – | – | – | ++ | – |
| 94-A | – | – | – | – | – | – | 356-D | – | – | – | – | – | – |
| 94-C | – | – | – | – | – | – | 381-A | – | – | – | – | – | – |
| 144-A | – | – | – | – | – | – | 443-A | – | – | – | – | – | – |
| 144-B | – | – | – | – | – | – | 626-A | – | – | – | – | – | – |
| 144-D | – | – | – | – | – | – | 864-A | – | – | – | – | – | – |
| 197-C | – | – | – | – | – | – | 1B18-A | – | – | – | – | – | – |
| 220-A | – | – | – | – | – | – | 1B18-C | – | – | – | – | – | – |
| 246-D | – | – | – | + | ++ | ++ | 1B247-A | – | – | – | – | ++ | +++ |
| 246-A | – | – | – | – | +++ | ++ | 3C110-A | – | – | – | – | ++ | – |
| 263-A | – | – | – | – | – | – | 4C168-A | – | – | – | – | – | – |
| 290-B | – | – | – | +++ | ++ | ++ | 4C171-B | – | – | – | ++ | +++ | +++ |
| 308-A | – | – | – | – | – | – | |||||||
Pathogenic microorganisms: (1) Klebsiella pneumoniae ATCC11486 (2) Escherichia coli ATCCBAA2469 (3) Pseudomonas aeruginosa ATCC27853 (4) Bacillus subtilis ATCC 55033 (5) Staphylococcus epidermidis ATCC14990 (6) Staphylococcus aureus ATCCBAA44. +++ (inhibition diameter >15 mm), ++ (10-15 mm), + (1-10mm), – (inhibition diameter <1 mm). Pigmented extracts listed according to a code comprising the codified strain (Table 4) hyphenated to the culture medium type, i.e., ISP2 (A) ISP4 (B) modified ISP9 (C) starch nitrate (D). For more information, all the antibiograms are presented in supplementary material (Figures S1 to S15).
Table 4.
Molecular identification of strains with pigmentation.
| Strain code | Identification | % Similarity |
|---|---|---|
| 2 | Streptomyces griseochromogenes | 99.93% |
| 94 | Streptomyces tendae | 99.86% |
| 144 | Streptomyces humi | 99.55% |
| 197 | Streptomyces misionensis | 99.72% |
| 220 | Streptomyces argillaceus | 100% |
| 246 | Streptomyces murinus | 99.93% |
| 290 | Streptomyces noursei | 99.24% |
| 308 | Streptomyces humi | 99.64% |
| 356 | Streptomyces hygroscopicus | 99.93% |
| 381 | Streptomyces albospinus | 99.18% |
| 443 | Streptomyces murinus | 98.96% |
| 626 | Streptomyces albospinus | 99.85% |
| 864 | Streptomyces lacticiproducens | 99.17% |
| 1B18 | Streptomyces lactacystinicus | 99.35% |
| 1B247 | Streptomyces mediolani | 99.64% |
| 3C110 | Streptomyces fodineus | 99.86% |
| 4C168 | Streptomyces murinus | 99.93% |
| 4C171 | Streptomyces murinus | 99.86% |
Pigmented extracts from strains S. murinus 246 grown in starch nitrate medium, S. noursei 290 and S. murinus 4C171 in ISP4 medium exhibited antimicrobial activity against these three bacteria and show promising potential as antibiotics. These strains are characterized by their yellow pigmentation and the use of starch as a carbon source. Notable, previously reported antibiotic yellow pigments include D-actinomycin from S. parvulus C5-5Y, isolated from the leather industry [77], a fraction from S. hygroscopicus subsp. ossamyceticus (D10), isolated from desert soil, which was active against vancomycin-resistant S. aureus [109], a crude pigment from Streptomyces sp. D25 with activity against S. aureus MTCC96 [110], grixazone A and B from S. griseus IFO13350, obtained from the Institute of Fermentation, Osaka, Japan (IFO), which was active against B. subtilis ATCC6633 and S. aureus 209P JC-1[111].
To a lesser extent, extracts from strains S. murinus 246 and S. mediolani 1B247 grown in ISP2 showed activity against S. epidermidis ATCC 14990 and S. aureus ATCC BAA44. Extracts from strains S. fodineus 3C110 and S. hygroscopicus 356 in ISP2 exhibited activity only against S. epidermidis ATCC 14990. In most cases, changing the culture medium did not affect antibacterial activity. However, strain S. hygroscopicus 356 inhibited the growth of S. epidermidis ATCC 14990 in ISP2 medium but not in starch nitrate medium, suggesting that the same strain may produce different specialized metabolites with varying bioactivities depending on the medium.
3.3.3. Cytotoxic Activity
Identifying anti-cancer compounds is crucial due to the limitations of current chemotherapies, such as multidrug resistance and secondary effects. Natural products, including those from microorganisms, have shown potential in inhibiting cell proliferation and promoting apoptosis [112]. Bacteria are highlighted as sources of specialized metabolites with anti-cancer properties, potentially leading to the development of more effective cancer therapies [113,114]. Streptomyces species, known for producing clinically significant drugs like doxorubicin and bleomycin [115,116], also produce pigmented compounds with anticancer activity. Notable examples include red pigmented crude extracts from Streptomyces sp. PM4 [66] and Streptomyces sp. A 16-1 [65], pure compounds such as undecylprodigiosin from Streptomyces sp. JAR6 [75] and prodigiosin from Streptomyces sp. WMA-LM31 [117]. Other pigments include a yellow pigment from Streptomyces griseoaurantiacus JUACT 01 [41] with activity against HeLa and liver cancer (HepG2), melanin from Streptomyces glaucescens NEAE-H [36], and eumelanin from Streptomyces parvus BSB49 [68] with activity against skin cancer (HFB4) and HeLa, respectively.
When evaluating the pigmented extracts against three cell lines—human dermal fibroblasts (HDFa), cervical cancer cell line (HeLa), and breast cancer cell line (MCF-7) (see Figure 3)—the MCF-7 cell line was the most resistant to the extracts. Extracts from strains S. murinus 246 in starch nitrate and S. murinus 4C171 in ISP4 medium showed promising anticancer activity against HeLa and MCF-7 without being toxic to the HDFa cell line. Extract Streptomyces noursei 290 in ISP4 medium demonstrated good anticancer activity against HeLa at all concentrations evaluated and against MCF-7 at maximum concentration, although it was toxic to HDFa. Strain S. murinus 443 in ISP 2 also showed promise at all concentrations without being toxic to HDFa.
In general, tested extracts exhibited a dose-dependent response (Figure 3). However, some extracts exhibited atypical behavior. For instance, strain S. tendae 94 in mISP9 showed higher cytotoxicity against MCF-7 at intermediate concentrations than at the highest concentration, while strain S. misionensis 197 in mISP 9 and strain S. tendae 94 in ISP2 showed higher activity at lower concentrations. Half-maximal inhibitory concentrations (IC50) were calculated for those extracts showing a dose-dependent response (see Figures S16-S20). Strain S. murinus 4C171 in starch nitrate had IC50 values of 146.4 ppm against HeLa and 242.6 ppm against MCF-7, which were relatively higher than those reported in literature, ranging from 0.04 to 48.07 ppm [118]. Strain S. murinus 246 in starch nitrate had an IC50 of 119.3 ppm against HeLa, which is lower than undecylprodigiosin from Streptomyces sp. JAR6 [75] but higher than a yellow pigment from Streptomyces griseoaurantiacus JUACT 01 [41]. For strains showing cytotoxicity, IC50 values were 152.6 ppm for S. mediolani 1B247 in ISP2 and 104.8 ppm for S. noursei 290 in ISP4. These values are higher than those reported for melanin from Streptomyces glaucescens NEAE-H, which were 37.05 ppm and 48.07 ppm against noncancer cell lines [37]. This suggests that higher concentrations are required for cytotoxicity, though these extracts are not entirely safe.
3.4. HPLC-MS-Based Annotation (Level 3) of Metabolites Possibly Responsible for the Observed Activities by the Different Streptomyces-Derived Extracts
The most promising pigmented extracts were analyzed by HPLC-MS to chemically characterize such extracts and annotate those detected compounds. The annotated metabolites were searched in literature and StreptomeDB database to determine a plausible role in pigmentation and bioactivity, as well as to find previous sources related to Streptomyces species that produce them. In this context, strain S. humi 144, cultured on ISP2 and ISP4 media, exhibited antioxidant capacity, as evidenced by DDPH and ABTS assays. HPLC-MS-based characterization suggested that this activity may be associated with chlorogenic acid [49], a phenylpropanoid previously identified in Streptomyces sp. PM9 [119], known for its antioxidant efficacy comparable to ʟ-ascorbic acid [120]. This effectiveness is often attributed to the hydroxyl groups in the aromatic ring structure [121]. Additionally, strain S. humi 144 in ISP2 medium demonstrated moderate cytotoxic activity against HeLa cells, possibly due to the presence of certain compounds, detected and annotated in this extract (Table S6), which previously reported to have cytotoxic activity. In contrast, strain S. noursei 290 in ISP4 showed antibacterial activity against B. subtilis ATCC 55033, S. epidermidis ATCC14990, and S. aureus ATCCBAA44, which may be attributed to the compounds listed in Table S7. This extract also had good anticancer activity against HeLa and MCF-7 cells, potentially due to the detected, annotated compounds listed in Table S8. Furthermore, HPLC-MS analysis suggested that the pigments responsible for coloration in this strain may include phenazine-1-carboxylate (also known as tubermycin B) with a greenish-yellow hue [122], or tetracenomycin D1, related to tetracenomycin C, a pale yellow antibiotic [123]. Other compounds with potential antimicrobial and cytotoxic activity included pentostatin, phenazine-1-carboxylate, yautomycetin, and tetracenomycin D1.
Strain S. murinus 246 in starch nitrate exhibited antibacterial activity against B. subtilis ATCC 55033, S. epidermidis ATCC14990, and S. aureus ATCCBAA44, possibly due to action of some of the annotated compounds (Table S9), which are related to antibiotics such as cephamicyn C, lankamycin, and sisomicin. It also showed promising cytotoxic activity against HeLa and MCF-7 cells without toxicity to the normal cell line (HDFa), potentially due to the presence of fumiformamide, maculosin, and salinomycin as annotated compounds with reported cytotoxic activity (Table S10). The pigments responsible might include a xanthone-related compound, such as xantholipin B. Xanthones, derived from the greek word “xanthos” meaning yellow, are polyketide-derived compounds, firstly isolated from Streptomyces but also found in plants, with a C6-C1-C6 carbon skeleton [124]. Xanthones have various properties, including antimicrobial, antioxidant, anti-inflammatory, and antitumor activities [121,122,123]. Recently, a new polycyclic xanthone, namely sattahipmycin, was identified from marine Streptomyces sp. GKU 257-1, and exhibited antimicrobial and cytotoxic activity [125]. Remarkably, other S. murinus strain (i.e., 4C171) in ISP4 medium showed antibacterial activity against B. subtilis ATCC 55033, S. epidermidis ATCC14990, and S. aureus ATCCBAA44, potentially due to a diverse metabolite classess listed in Table S11. The extract of this strain exhibited low antioxidant capacity, possibly related to the low action or low concentration of annotated compounds in this extract (Table S12). However, it demonstrated promising anticancer activity against HeLa and MCF-7 cells without toxicity to the healthy cell line (HDFa), likely due to the presence of active metabolites (Table S13). Potential pigments include carotenoids like beta-carotene, γ-carotene, neurosporene, 1,2,1’,2’-tetrahydrolycopene, and lycopene, which have been reported in Streptomyces species without photoinduction [126]. Other possible pigments include resistomycin, a natural antibiotic with cytotoxic activity [78,118,127]; flavonoids such as taxifolin or dihydroquercetin [118], phenazine-1-carboxylate which has a greenish-yellow coloration [122], tetracenomycin C which is pale yellow antibiotic [123], riboflavin, identified in species such as Streptomyces sp. TS-2-2 [128], and the red pigment roseoflavin from species like Streptomyces davaonensis [131,132].
On the other hand, strain S. fodineus 3C110 growth in ISP2 medium exhibited antibacterial activity against S. epidermidis ATCC14990, potentially due to those metabolites filtered as bioactives (Table S14). It also showed moderate antioxidant capacity, possibly related to salinomycin, the only antioxidant compound detected in this extract by HPLC-MS. This 3C110-derived extract was found to be active against HeLa cells at maximum dose, involving the presence of three cytotoxic compounds (Table S15). Additionally, strain S. hygroscopicus 356 in ISP2 medium showed antibacterial activity against S. epidermidis ATCC14990, moderate antioxidant capacity, and reasonable cytotoxic activity against HeLa cells at maximum concentration, whose bioactivity were rationalized by the reported properties of those annotated compounds for this extract (Tables S16-S18). Remarkably, this strain S. hygroscopicus 356 grown in starch nitrate showed no antibacterial activity against the pathogens tested and lower antioxidant capacity compared to ISP2 medium, exhibiting a different profile and most compounds were not produced and accumulated in such a medium (i.e., starch nitrate), excepting 2-acetylpyrrole, previously produced by Streptomyces sp. MUM273b [129], which was additionally detected. This extract also showed cytotoxic activity against HeLa cells at maximum concentration but to a lesser degree than in ISP2, involving cytotoxic compounds such as chartreusin, jadomycin A, and pentalenolactone (Table S19).
HPLC-MS-based characterization of S. hygroscopicus 356-derived extracts obtained from two culture media (ISP2 and starch nitrate) revealed the plausible production of annotated aromatic polyketide diphenyl ethers, i.e., Violaceol I and II, purple pigments known from co-cultures of Streptomyces rapamycinicus and Aspergillus nidulans [130], or endophytic fungi Aspergillus austroafricanus and Streptomyces lividans [131]. These pigments or related compounds might account for the pinkish-purple color of strain S. hygroscopicus 356 in starch nitrate and its darker shade in ISP2 medium. However, the coloration of this strain 356 in ISP2 differs slightly from purple, possibly due to the presence of other pigments like β-isorenieratene, luteothin, and xanthone-like metabolites.
Particularly, strain S. misionensis 197 in mISP9 medium shows no antibacterial activity against evaluated pathogens or cytotoxicity against cell lines but exhibits certain toxicity against HDFa. It also showed moderate antioxidant activity, potentially due to antioxidant compounds (Table S20). Similarly, strain S. murinus 443 in ISP2 medium showed no antibacterial activity against evaluated pathogens and low antioxidant capacity but it was active against HeLa and MCF-7 cells, possibly related to the presence of previously reported Streptomyces-derived compounds (Table S21-S22). Detected pigments in this extract included carotenoids such as β-carotene, γ-carotene, β-isorenieratene, phytoene, 1,2,1’,2’-tetrahydrolycopene, and lycopene [126]. Finally, strain S. lacticiproducens 864 in ISP2 medium exhibits no antibacterial activity against evaluated pathogens, moderate antioxidant capacity, and cytotoxic activity against HeLa and MCF-7 cells at the highest tested concentrations, whose properties could be explained by the detection of bioactive metabolites listed in Tables S23 and S24. In the case of strain S. humi 308 in ISP2 medium exhibits only antioxidant capacity, possibly due to the production of phenolic-like antioxidant metabolites [132,133,134].
4. Conclusions
The genus Streptomyces stands out as a remarkable source of diverse pigments with a spectrum of bioactivities, including antioxidant, antibacterial, and cytotoxic properties. This study underscores the significant potential of pigmented extracts from various Streptomyces strains to be active against pathogenic microorganisms and exhibit anticancer properties. Among the six pathogenic microorganisms tested, three (B. subtilis, S. epidermidis, and S. aureus) showed antibiotic sensitivity to certain pigmented extracts, while six Streptomyces-derived extracts exhibited cytotoxic activity (cell viability < 40%) against two cancer cell lines (HeLa and MCF-7), and various pigmented extracts demonstrated antioxidant capacity at different levels (TEAC > 20 µM) through DPPH and ABTS methods. Notably, strains S. noursei 290, S. murinus 246 and 4C171, which produce yellow pigments and utilize starch as a carbon source, exhibited promising antibacterial and anticancer activities. Extracts from strains S. murinus 4C171 and 246 also showed potent anticancer properties across various concentrations, exhibiting selectivity without compromising the viability of healthy HDFa cell lines. Molecular identification through 16S rRNA gene sequencing confirmed that all isolated strains belong to the Streptomyces genus, with similarities ranging from 98% to 100%. The HPLC-MS-based chemical characterization of active extracts identified several bioactive compounds responsible for the observed activities. Strain S. humi 144’s antioxidant capacity is likely linked to the presence of chlorogenic acid, while strain S. noursei 290’s broad-spectrum bioactivities might be attributed to compounds such as phenazine-1-carboxylate and tetracenomycin D1. Similarly, S. murinus strains’ activities (strains 246 and 4C17) are possibly due to carotenoids and xanthone-related compounds. This study also highlighted the importance of culture media in influencing the chemical composition of resulting extracts. Thus, the exploration of distinct culture mediums is crucial in characterizing the variations on the bioactivity associated with the specialized metabolite production, as their synthesis is intricately linked to primary carbon and nitrogen sources. For instance, strain S. hygroscopicus 356 exhibited different antibacterial and antioxidant capacities depending on the employed medium, suggesting that the same strain can produce different specialized metabolites with varying bioactivities. Notably, starch is a particularly favorable carbon source, driving the biosynthesis of yellow pigmented extracts that showcase the most pronounced potential as both antibacterial and anticancer agents. These findings underscore the multifaceted nature of Streptomyces-derived compounds (e.g., pigments) and extracts and their promising applications in the pharmaceutical and therapeutic industries. Hence, the pigmented extracts from these Streptomyces strains show reasonable promise as sources of novel antibiotics and anticancer agents. Further research, including the purification of these compounds and exploration of their mechanisms of action, is warranted to fully realize their therapeutic potential.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, L.D.; methodology, A.A.S.-T., S.J.P.-R., and J.G.-R.; software, A.A.S.-T. and E.C.-B.; validation, L.D. and E.C.-B.; formal analysis, A.A.S.-T., S.J.P.-R., and J.G.-R.; investigation, A.A.S.-T., S.J.P.-R., and J.G.-R.; resources, E.C.-B. and L.D.; data curation, E.C.-B. and L.D.; writing—original draft preparation, A.A.S.-T.; writing—review and editing, E.C.-B. and L.D.; supervision, L.D.; project administration, L.D.; funding acquisition, L.D.
Funding
This research was funded by Universidad de La Sabana (General Research Directorate, project ING-204-2018) and Universidad Militar Nueva Granada (project EXT-CIAS-3854).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors are grateful to the members of the groups ProNIUS and GIBP, Universidad de La Sabana, for substantially supporting the research with all the equipment, reagents, and academic support with helpful comments and encouraging remarks.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Marcano, D. Introducción a La Química de Los Colorantes; 2018. ISBN 9789806195592.
- Lyu, X.; Lyu, Y.; Yu, H.; Chen, W.; Ye, L.; Yang, R. Biotechnological Advances for Improving Natural Pigment Production: A State-of-the-Art Review. Bioresour. Bioprocess. 2022, 9, 8. [Google Scholar] [CrossRef]
- Ahmad, W.A.; Yusof, N.Z.; Nordin, N.; Zakaria, Z.A.; Rezali, M.F. Production and Characterization of Violacein by Locally Isolated Chromobacterium Violaceum Grown in Agricultural Wastes. Appl. Biochem. Biotechnol. 2012, 167, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- Sajda. S. Affat Classifications, Advantages, Disadvantages, Toxicity Effects of Natural and Synthetic Dyes: A Review. Univ. Thi-Qar J. Sci. 2021, 8, 130–135. [Google Scholar]
- Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Bacterial Pigments and Their Applications. Process Biochem. 2013, 48, 1065–1079. [Google Scholar] [CrossRef]
- Paul, T.; Bandyopadhyay, T.K.; Mondal, A.; Tiwari, O.N.; Muthuraj, M.; Bhunia, B. A Comprehensive Review on Recent Trends in Production, Purification, and Applications of Prodigiosin. Biomass Convers. Biorefinery 2020. [Google Scholar] [CrossRef]
- Husain, Q. Potential Applications of the Oxidoreductive Enzymes in the Decolorization and Detoxification of Textile and Other Synthetic Dyes from Polluted Water: A Review. Crit. Rev. Biotechnol. 2006, 26, 201–221. [Google Scholar] [CrossRef]
- Khattab, T.A.; Abdelrahman, M.S.; Rehan, M. Textile Dyeing Industry: Environmental Impacts and Remediation. Environ. Sci. Pollut. Res. 2020, 27, 3803–3818. [Google Scholar] [CrossRef]
- Sarayu, K.; Sandhya, S. Current Technologies for Biological Treatment of Textile Wastewater–A Review. Appl. Biochem. Biotechnol. 2012, 167, 645–661. [Google Scholar] [CrossRef]
- Lellis, B.; Fávaro-Polonio, C.Z.; Pamphile, J.A.; Polonio, J.C. Effects of Textile Dyes on Health and the Environment and Bioremediation Potential of Living Organisms. Biotechnol. Res. Innov. 2019, 3, 275–290. [Google Scholar] [CrossRef]
- Sanchez-Marroquín, A.; Zapata, M. Observations on the Pigment of Streptomyces Coelicolor. 1954, 2.
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.-K. Actinobacterial Melanins: Current Status and Perspective for the Future. World J. Microbiol. Biotechnol. 2013, 29, 1737–1750. [Google Scholar] [CrossRef] [PubMed]
- Dharmaraj, S. Marine Streptomyces as a Novel Source of Bioactive Substances. World J. Microbiol. Biotechnol. 2010, 26, 2123–2139. [Google Scholar] [CrossRef]
- Mann, J. Natural Products as Immunosuppressive Agents. Nat. Prod. Rep. 2001, 18, 417–430. [Google Scholar] [CrossRef]
- Olano, C.; Méndez, C.; Salas, J.A. Antitumor Compounds from Marine Actinomycetes. Mar. Drugs 2009, 7, 210–248. [Google Scholar] [CrossRef]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary Metabolites and Biodiversity of Actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19. [Google Scholar] [CrossRef]
- Pathom-aree, W.; Stach, J.E.M.; Ward, A.C.; Horikoshi, K.; Bull, A.T.; Goodfellow, M. Diversity of Actinomycetes Isolated from Challenger Deep Sediment (10,898 m) from the Mariana Trench. Extremophiles 2006, 10, 181–189. [Google Scholar] [CrossRef]
- Bull, A.T. Microbial Diversity and Bioprospecting; Bull, A.T., Ed.; ASM Press: Washington, DC, USA, 2003. ISBN 9781683672173.
- Berdi, J. Bioactive Microbial Metabolites. J. Antibiot. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Devi Salam, M. Antimicrobial Potential of Actinomycetes Isolated from Soil Samples of Punjab, India. J. Microbiol. Exp. 2014, 1. [Google Scholar] [CrossRef]
- Darshan, N.; Manonmani, H.K. Prodigiosin and Its Potential Applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.C.; Zhan, J.X.; Su, K.M.; Zhang, Y.X. A Kind of Potential Food Additive Produced by Streptomyces Coelicolor: Characteristics of Blue Pigment and Identification of a Novel Compound, Lambda-Actinorhodin. Food Chem. 2006, 95, 186–192. [Google Scholar] [CrossRef]
- Petinate, S.D.; Martins, R.M.; Coelho, R.R.; Meirelles, M.N.L.; Branquinha, M.H.; Vermelho, A.B. Influence of Growth Medium in Proteinase and Pigment Production by Streptomyces Cyaneus. Mem. Inst. Oswaldo Cruz 1999, 94, 173–177. [Google Scholar] [CrossRef]
- Zhu, H.-H.; Guo, J.; Yao, Q.; Yang, S.-Z.; Deng, M.-R.; Le Phuong, T.B.; Hanh, V.T.; Ryan, M.J. Streptomyces Vietnamensis Sp. Nov., a Streptomycete with Violet Blue Diffusible Pigment Isolated from Soil in Vietnam. Int. J. Syst. Evol. Microbiol. 2007, 57, 1770–1774. [Google Scholar] [CrossRef]
- Gupta, C.; Garg, A.P.; Prakash, D.; Goyal, S. Microbes as Potential Sources of Biocolours. Pharmacologyonline 2011, 1309–1318. [Google Scholar]
- Lin, Y.B.; Wang, X.Y.; Fang, H.; Ma, Y.N.; Tang, J.; Tang, M.; Wei, G.H. Streptomyces Shaanxiensis Sp. Nov., a Blue Pigment-Producing Streptomycete from Sewage Irrigation Soil. Int. J. Syst. Evol. Microbiol. 2012, 62, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-H.; Guo, J.; Yao, Q.; Yang, S.-Z.; Deng, M.-R.; Li, T.-H. Streptomyces Caeruleatus Sp. Nov., with Dark Blue Diffusible Pigment. Int. J. Syst. Evol. Microbiol. 2011, 61, 507–511. [Google Scholar] [CrossRef]
- Malik, K.; Tokkas, J.; Goyal, S. Microbial Pigments : A Review. Int. J. Microb. Resour. Technol. 2012, 361–365. [Google Scholar]
- Morales-Oyervides, L.; Oliveira, J.; Sousa-Gallagher, M.; Méndez-Zavala, A.; Montañez, J.C. Assessment of the Dyeing Properties of the Pigments Produced by Talaromyces Spp. J. Fungi 2017, 3. [Google Scholar] [CrossRef]
- Narsing Rao, M.P.; Xiao, M.; Li, W. Fungal and Bacterial Pigments : Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pombeiro-Sponchiado, S.R.; Sousa, G.S.; Andrade, J.C.R.; Lisboa, H.F.; Gonçalves, R.C.R. Production of Melanin Pigment by Fungi and Its Biotechnological Applications. Melanin 2017. [Google Scholar] [CrossRef]
- Pailliè-Jiménez, M.E.; Stincone, P.; Brandelli, A. Natural Pigments of Microbial Origin. Front. Sustain. Food Syst. 2020, 4, 1–8. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Multifaceted Applications of Microbial Pigments: Current Knowledge, Challenges and Future Directions for Public Health Implications; 2019; Vol. 7; ISBN 3326221754.
- Uribe Vélez, D. Metagenómica ¿Una Oportunidad Para El Estudio de La Diversidad Microbiana En Colombia? Rev. Colomb. Biotecnol. 2009, XI, 4–7. [Google Scholar]
- Shirling, E.B.; Gottlieb, D. Methods for Characterization of Streptomyces Species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; El-Ewasy, S.M. Bioproduction, Characterization, Anticancer and Antioxidant Activities of Extracellular Melanin Pigment Produced by Newly Isolated Microbial Cell Factories Streptomyces Glaucescens NEAE-H. Sci. Rep. 2017, 7, 42129. [Google Scholar] [CrossRef] [PubMed]
- Benouagueni, S.; Ranque, S.; Gacemi Kirane, D. A Non-Polyenic Antifungal Produced by a Streptomyces Yatensis Strain Isolated from Mellah Lake in El Kala, North-East of Algeria. J. Mycol. Med. 2015, 25, 2–10. [Google Scholar] [CrossRef]
- Siddharth, S.; Vittal, R.R. Evaluation of Antimicrobial, Enzyme Inhibitory, Antioxidant and Cytotoxic Activities of Partially Purified Volatile Metabolites of Marine Streptomyces Sp.S2A. Microorganisms 2018, 6, 72. [Google Scholar] [CrossRef]
- Francenia Santos-Sánchez, N.; Salas-Coronado, R.; Villanueva-Cañongo, C.; Hernández-Carlos, B. Antioxidant Compounds and Their Antioxidant Mechanism. In Antioxidants; IntechOpen, 2019; pp. 1–29.
- Taroco, R.; Seija, V.; Vignoli, R. Métodos de Estudio de La Sensibilidad Antibiótica. In Temas de Bacterología y Virología Médica; 2008; pp. 663–671.
- Prashanthi, K.; Suryan, S.; Varalakshmi, K.N. In Vitro Anticancer Property of Yellow Pigment from Streptomyces Griseoaurantiacus JUACT 01. Brazilian Arch. Biol. Technol. 2015, 58, 869–876. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the Human-Ape Splitting by a Molecular Clock of Mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ramirez-Rodriguez, L.; Stepanian-Martinez, B.; Morales-Gonzalez, M.; Diaz, L. Optimization of the Cytotoxic Activity of Three Streptomyces Strains Isolated from Guaviare River Sediments (Colombia, South America). Biomed Res. Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Bernal, F.A.; Orduz-Díaz, L.L.; Coy-Barrera, E. Application of Parafac and OPLS-DA Analyses on HPLC Fingerprints for the Characterization of Hibiscus Sabdariffa Calyxes. Quim. Nova 2016. [Google Scholar] [CrossRef]
- Schmid, R.; Heuckeroth, S.; Korf, A.; Smirnov, A.; Myers, O.; Dyrlund, T.S.; Bushuiev, R.; Murray, K.J.; Hoffmann, N.; Lu, M.; et al. Integrative Analysis of Multimodal Mass Spectrometry Data in MZmine 3. Nat. Biotechnol. 2023, 41, 447–449. [Google Scholar] [CrossRef]
- Moumbock, A.F.A.; Gao, M.; Qaseem, A.; Li, J.; Kirchner, P.A.; Ndingkokhar, B.; Bekono, B.D.; Simoben, C. V; Babiaka, S.B.; Malange, Y.I.; et al. StreptomeDB 3.0: An Updated Compendium of Streptomycetes Natural Products. Nucleic Acids Res. 2021, 49, D600–D604. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 Update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Rutz, A.; Sorokina, M.; Galgonek, J.; Mietchen, D.; Willighagen, E.; Gaudry, A.; Graham, J.G.; Stephan, R.; Page, R.; Vondrášek, J.; et al. The LOTUS Initiative for Open Knowledge Management in Natural Products Research. Elife 2022, 11. [Google Scholar] [CrossRef]
- van Santen, J.A.; Poynton, E.F.; Iskakova, D.; McMann, E.; Alsup, T.A.; Clark, T.N.; Fergusson, C.H.; Fewer, D.P.; Hughes, A.H.; McCadden, C.A.; et al. The Natural Products Atlas 2.0: A Database of Microbially-Derived Natural Products. Nucleic Acids Res. 2022, 50, D1317–D1323. [Google Scholar] [CrossRef]
- van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The Natural Products Atlas: An Open Access Knowledge Base for Microbial Natural Products Discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef]
- Wink, J.; Fatemeh, M.; Javad, H. Biology and Biotechnology of Actinobacteria; Wink, J. , Mohammadipanah, F., Hamedi, J., Eds.; Springer International Publishing: Cham, 2017; ISBN 978-3-319-60338-4. [Google Scholar]
- Anzai, K.; Ohno, M.; Nakashima, T.; Kuwahara, N.; Suzuki, R.; Tamura, T.; Komaki, H.; Miyadoh, S.; Harayama, S.; Ando, K. Taxonomic Distribution of Streptomyces Species Capable of Producing Bioactive Compounds among Strains Preserved at NITE/NBRC. Appl. Microbiol. Biotechnol. 2008, 80, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.-G.; Butler, M.J.; Chater, K.F.; Lee, K.J. Engineering of Primary Carbohydrate Metabolism for Increased Production of Actinorhodin in Streptomyces Coelicolor. Appl. Environ. Microbiol. 2006, 72, 7132–7139. [Google Scholar] [CrossRef]
- Demain, A.L.; Sanchez, S. Microbial Drug Discovery: 80 Years of Progress. J. Antibiot. (Tokyo). 2009, 62, 5–16. [Google Scholar] [CrossRef]
- Romero-Rodríguez, A.; Maldonado-Carmona, N.; Ruiz-Villafán, B.; Koirala, N.; Rocha, D.; Sánchez, S. Interplay between Carbon, Nitrogen and Phosphate Utilization in the Control of Secondary Metabolite Production in Streptomyces. Antonie Van Leeuwenhoek 2018, 111, 761–781. [Google Scholar] [CrossRef]
- Görke, B.; Stülke, J. Carbon Catabolite Repression in Bacteria: Many Ways to Make the Most out of Nutrients. Nat. Rev. Microbiol. 2008, 6, 613–624. [Google Scholar] [CrossRef]
- Omura, S.; Tanaka, Y.; Mamada, H.; Masuma, R. Effect of Ammonium Ion, Inorganic Phosphate and Amino Acids on the Biosynthesis of Protylonolide, a Precursor of Tylosin Aglycone. J. Antibiot. (Tokyo). 1984, 37, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Tomé, D. Yeast Extracts: Nutritional and Flavoring Food Ingredients. ACS Food Sci. Technol. 2021, 1, 487–494. [Google Scholar] [CrossRef]
- Narayana, K.J.P.; Kumar, K.G.; Vijayalakshmi, M. L-Asparaginase Production by Streptomyces Albidoflavus. Indian J. Microbiol. 2008, 48, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Khopade, A.; Ren, B.; Liu, X.-Y.; Mahadik, K.; Zhang, L.; Kokare, C. Production and Characterization of Biosurfactant from Marine Streptomyces Species B3. J. Colloid Interface Sci. 2012, 367, 311–318. [Google Scholar] [CrossRef]
- Langheinrich, W.; Ring, K. Regulation of Amino Acid Transport in Growing Cells of Streptomyces Hydrogenans. Arch. Microbiol. 1976, 109, 227–235. [Google Scholar] [CrossRef]
- Van Wezel, G.P.; Mahr, K.; König, M.; Traag, B.A.; Pimentel-Schmitt, E.F.; Willimek, A.; Titgemeyer, F. GlcP Constitutes the Major Glucose Uptake System of Streptomyces Coelicolor A3(2). Mol. Microbiol. 2004, 55, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Čihák, M.; Kameník, Z.; Šmídová, K.; Bergman, N.; Benada, O.; Kofroňová, O.; Petříčková, K.; Bobek, J. Secondary Metabolites Produced during the Germination of Streptomyces Coelicolor. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Saengkhae, C.; Srivibool, R.; Watanadilok, R.; Enomoto, K. Partially Purified Pigment Extract from Streptomyces A 16-1 Induces Apoptosis of Human Carcinoma of Nasopharynx Cell (KB Cells) via the Mitochondrial and Caspase-3 Pathway. Walailak J. Sci. Technol. 2017, 14, 51–63. [Google Scholar] [CrossRef]
- Karuppiah, V.; Aarthi, C.; Ramohan, *!!! REPLACE !!!*; Sivakumar, K.; Kannan, L. Statistical Optimization and Anticancer Activity of a Red Pigment Isolated from Streptomyces Sp. PM4. Asian Pac. J. Trop. Biomed. 2013, 3, 650–656. [Google Scholar] [CrossRef]
- Vaishnavi, M.; Manigundan, K.; Smalia, T.; N, *!!! REPLACE !!!*; Hini, S.U.; Gopikrishnan, V.; Kumar, A.; Hanna, L.E.; Radhakrishnan, M.; Aruni, W. Antibacterial and Anti-HIV Activity of Extracellular Pigment from Streptomyces Sp. S45 Isolated from Sabarimala Forest Soil, India. Indian J. Exp. Biol. 2020, 58, 861–868. [Google Scholar] [CrossRef]
- Bayram, S.; Dengiz, C.; Gerçek, Y.C.; Cetin, I.; Topcul, M.R. Bioproduction, Structure Elucidation and in Vitro Antiproliferative Effect of Eumelanin Pigment from Streptomyces Parvus BSB49. Arch. Microbiol. 2020, 202, 2401–2409. [Google Scholar] [CrossRef] [PubMed]
- Bayram, S. Production, Purification, and Characterization of Streptomyces Sp. Strain MPPS2 Extracellular Pyomelanin Pigment. Arch. Microbiol. 2021. [Google Scholar] [CrossRef]
- Dastager, S.G.; Wen-Jun, L.; Dayanand, A.; Shu-Kun, T.; Xin-Peng, T.; Xiao-Yang, Z.; Li-Hua, X.; Cheng-Lin, J. Seperation, Identification and Analysis of Pigment (Melanin) Production in Streptomyces. African J. Biotechnol. 2006, 5, 1131–1134. [Google Scholar]
- Tang-um, J.; Niamsup, H. Extracellular Amylase Activity from Endophytic Streptomyces Griseoflavus P4. Chiang Mai J. Sci. 2012, 39, 346–350. [Google Scholar]
- Singh, R.; Tiwari, M.; Singh, R.; Lee, J.-K. From Protein Engineering to Immobilization: Promising Strategies for the Upgrade of Industrial Enzymes. Int. J. Mol. Sci. 2013, 14, 1232–1277. [Google Scholar] [CrossRef] [PubMed]
- Elibol, M. Optimization of Medium Composition for Actinorhodin Production by Streptomyces Coelicolor A3(2) with Response Surface Methodology. Process Biochem. 2004, 39, 1057–1062. [Google Scholar] [CrossRef]
- Elyasi Far, B.; Ahmadi, Y.; Yari Khosroshahi, A.; Dilmaghani, A. Microbial Alpha-Amylase Production: Progress, Challenges and Perspectives. Adv. Pharm. Bull. 2020, 10, 350–358. [Google Scholar] [CrossRef]
- Abraham, J.; Chauhan, R. Profiling of Red Pigment Produced by Streptomyces Sp. JAR6 and Its Bioactivity. 3 Biotech 2018, 8, 22. [Google Scholar] [CrossRef]
- Azimi, S.; Baserisalehi, M.; Bahador, N. Evaluation of Antimicrobial Pigment Produced by Streptomyces Coeruleorubidus. Nat. Environ. Pollut. Technol. 2014, 13, 641–644. [Google Scholar]
- Soundari, A. Parimala Gnana Mani, V.M.; Bose, V.S.C.; Jabastin, J.; Priyadarisini, V.B. A Preliminary Assessment of Yellow Pigment from Streptomyces Parvulus C5-5Y. J. Pure Appl. Microbiol. 2017, 11, 197–203. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Bruheim, P.; Andreassen, T.; Raja, D.S.; Devi, P.B.; Sathyabama, S.; Priyadarisini, V.B. Assessment of Resistomycin, as an Anticancer Compound Isolated and Characterized from Streptomyces Aurantiacus AAA5. J. Microbiol. 2011, 49, 920–926. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution (N. Y). 1985, 39, 783. [Google Scholar] [CrossRef]
- Abellan-Schneyder, I.; Matchado, M.S.; Reitmeier, S.; Sommer, A.; Sewald, Z.; Baumbach, J.; List, M.; Neuhaus, K. Primer, Pipelines, Parameters: Issues in 16S RRNA Gene Sequencing. mSphere 2021, 6, 10–1128. [Google Scholar] [CrossRef]
- Labeda, D.P.; Goodfellow, M.; Brown, R.; Ward, A.C.; Lanoot, B.; Vanncanneyt, M.; Swings, J.; Kim, S.-B.; Liu, Z.; Chun, J.; et al. Phylogenetic Study of the Species within the Family Streptomycetaceae. Antonie Van Leeuwenhoek 2012, 101, 73–104. [Google Scholar] [CrossRef] [PubMed]
- Nicault, M.; Tidjani, A.-R.; Gauthier, A.; Dumarcay, S.; Gelhaye, E.; Bontemps, C.; Leblond, P. Mining the Biosynthetic Potential for Specialized Metabolism of a Streptomyces Soil Community. Antibiotics 2020, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant Activity of Citrus Fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The Role of Antioxidants in the Chemistry of Oxidative Stress: A Review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in Vivo and in Vitro Methods Evaluation of Antioxidant Activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-O.; Lee, K.W.; Lee, H.J.; Lee, C.Y. Vitamin C Equivalent Antioxidant Capacity (VCEAC) of Phenolic Phytochemicals. J. Agric. Food Chem. 2002, 50, 3713–3717. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The Hydrophilic and Lipophilic Contribution to Total Antioxidant Activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Hyun, H.B.; Shrestha, S.; Boo, K.H.; Cho, S.K. Evaluation of Antioxidant Potential of Ethyl Acetate Fraction of Rosmarinus Officinalis L. and Its Major Components. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 715–722. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH Assays to Measure Antioxidant Capacity in Popular Antioxidant-Rich US Foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Arnao, M.B. Some Methodological Problems in the Determination of Antioxidant Activity Using Chromogen Radicals: A Practical Case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Chun, O.K. Comparison of ABTS/DPPH Assays for the Detection of Antioxidant Capacity in Foods. FASEB J. 2010, 24, 535–9. [Google Scholar] [CrossRef]
- Kuskoski, E.M.; Asuero, A.G.; Troncoso, A.M.; Mancini-filho, J.; Fett, R. Aplicación de Diversos Métodos Químicos Para Determinar Actividad Antioxidante En Pulpa de Frutos. Ciência e Tecnol. Aliment. 2005, 25, 726–732. [Google Scholar] [CrossRef]
- Thirumalai, D.; Manikkam, R.; Ramasamy, B. Antioxidant Activity of Melanin Pigment from Streptomyces Species D5 Isolated from Desert Soil, Rajasthan, India. Drug Invent. Today 2010, 2, 12–13. [Google Scholar]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the Sustainable Discovery and Development of New Antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef]
- Byrne, M.K.; Miellet, S.; McGlinn, A.; Fish, J.; Meedya, S.; Reynolds, N.; van Oijen, A.M. The Drivers of Antibiotic Use and Misuse: The Development and Investigation of a Theory Driven Community Measure. BMC Public Health 2019, 19, 1425. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Liu, Z.; Chen, Y.; Xiao, Y.; Huang, X.; Fan, X.-G. Bacterial and Fungal Infections in COVID-19 Patients: A Matter of Concern. Infect. Control Hosp. Epidemiol. 2020, 41, 1124–1125. [Google Scholar] [CrossRef] [PubMed]
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Westwood, D.; MacFadden, D.R.; Soucy, J.-P.R.; Daneman, N. Bacterial Co-Infection and Secondary Infection in Patients with COVID-19: A Living Rapid Review and Meta-Analysis. Clin. Microbiol. Infect. 2020, 26, 1622–1629. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, V.M.; Gandhi, T.N.; Petty, L.A.; Patel, P.K.; Prescott, H.C.; Malani, A.N.; Ratz, D.; McLaughlin, E.; Chopra, V.; Flanders, S.A. Empiric Antibacterial Therapy and Community-Onset Bacterial Coinfection in Patients Hospitalized With Coronavirus Disease 2019 (COVID-19): A Multi-Hospital Cohort Study. Clin. Infect. Dis. 2021, 72, e533–e541. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J. How Covid-19 Is Accelerating the Threat of Antimicrobial Resistance. BMJ 2020, m1983. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Giacobbe, D.R. A Look at the Clinical, Economic, and Societal Impact of Antimicrobial Resistance in 2020. Expert Opin. Pharmacother. 2020, 21, 2067–2071. [Google Scholar] [CrossRef] [PubMed]
- Nieuwlaat, R.; Mbuagbaw, L.; Mertz, D.; Burrows, L.L.; Bowdish, D.M.E.; Moja, L.; Wright, G.D.; Schünemann, H.J. Coronavirus Disease 2019 and Antimicrobial Resistance: Parallel and Interacting Health Emergencies. Clin. Infect. Dis. 2021, 72, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Rupp, M.E.; Fey, P.D.; Heilmann, C.; Götz, F. Characterization of the Importance of Staphylococcus Epidermidis Autolysin and Polysaccharide Intercellular Adhesin in the Pathogenesis of Intravascular Catheter–Associated Infection in a Rat Model. J. Infect. Dis. 2001, 183, 1038–1042. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus Epidermidis — the “accidental” Pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Forbes, B.A.; Schaberg, D.R. Transfer of Resistance Plasmids from Staphylococcus Epidermidis to Staphylococcus Aureus: Evidence for Conjugative Exchange of Resistance. J. Bacteriol. 1983, 153, 627–634. [Google Scholar] [CrossRef]
- Loggenberg, S.R.; Twilley, D.; De Canha, M.N.; Lall, N. Chapter 4 - Medicinal Plants Used in South Africa as Antibacterial Agents for Wound Healing. In Medicinal Plants as Anti-Infectives; Chassagne, F.B.T., Ed.; Academic Press, 2022; pp. 139–182. ISBN 978-0-323-90999-0.
- Khan, T.M.; Kok, Y.L.; Bukhsh, A.; Lee, L.-H.; Chan, K.-G.; Goh, B.-H. Incidence of Methicillin Resistant Staphylococcus Aureus (MRSA) in Burn Intensive Care Unit: A Systematic Review. Germs 2018, 8, 113–125. [Google Scholar] [CrossRef]
- Clebak, K.T.; Malone, M.A. Skin Infections. Prim. Care Clin. Off. Pract. 2018, 45, 433–454. [Google Scholar] [CrossRef]
- Baddour, L.M.; Wilson, W.R.; Bayer, A.S.; Fowler, V.G.; Tleyjeh, I.M.; Rybak, M.J.; Barsic, B.; Lockhart, P.B.; Gewitz, M.H.; Levison, M.E.; et al. Infective Endocarditis in Adults: Diagnosis, Antimicrobial Therapy, and Management of Complications. Circulation 2015, 132, 1435–1486. [Google Scholar] [CrossRef]
- Selvameenal, L.; Radhakrishnan, M.; Balagurunathan, R. Antibiotic Pigment from Desert Soil Actinomycetes; Biological Activity, Purification and Chemical Screening. Indian J. Pharm. Sci. 2009, 71, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Manikkam, R.; Venugopal, G.; Ramasamy, B.; Kumar, V. Effect of Critical Medium Components and Culture Conditions on Antitubercular Pigment Production from Novel Streptomyces Sp D25 Isolated from Thar Desert, Rajasthan. J. Appl. Pharm. Sci. 2015, 5, 15–19. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Furusho, Y.; Higashi, T.; Chun, H.K.; Furihata, K.; Sakuda, S.; Horinouchi, S. Structures of Grixazone A and B, A-Factor-Dependent Yellow Pigments Produced under Phosphate Depletion by Streptomyces Griseus. J. Antibiot. (Tokyo). 2004, 57, 218–223. [Google Scholar] [CrossRef]
- Lu, J.-J.; Wang, Y.-T. Identification of Anti-Cancer Compounds from Natural Products. Chin. J. Nat. Med. 2020, 18, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Mohan, C.D.; Rangappa, S.; Nayak, S.C.; Jadimurthy, R.; Wang, L.; Sethi, G.; Garg, M.; Rangappa, K.S. Bacteria as a Treasure House of Secondary Metabolites with Anticancer Potential. Semin. Cancer Biol. 2022, 86, 998–1013. [Google Scholar] [CrossRef]
- Schabel, F.M.; Pittillo, R.F. Screening for and Biological Characterization of Antitumor Agents Using Microorganisms. In; 1961; pp. 223–256.
- Tan, L.T.H.; Chan, K.G.; Pusparajah, P.; Yin, W.F.; Khan, T.M.; Lee, L.H.; Goh, B.H. Mangrove Derived Streptomyces Sp. MUM265 as a Potential Source of Antioxidant and Anticolon-Cancer Agents. BMC Microbiol. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cui, J.; Wang, P.; Wang, X.; Wen, J. Enhancement of Bleomycin Production in Streptomyces Verticillus through Global Metabolic Regulation of N-Acetylglucosamine and Assisted Metabolic Profiling Analysis. Microb. Cell Fact. 2020, 19, 32. [Google Scholar] [CrossRef]
- Sajjad, W.; Ahmad, S.; Aziz, I.; Azam, S.S.; Hasan, F.; Shah, A.A. Antiproliferative, Antioxidant and Binding Mechanism Analysis of Prodigiosin from Newly Isolated Radio-Resistant Streptomyces Sp. Strain WMA-LM31. Mol. Biol. Rep. 2018, 45, 1787–1798. [Google Scholar] [CrossRef]
- Sarmiento-Tovar, A.A.; Silva, L.; Sánchez-Suárez, J.; Diaz, L. Streptomyces-Derived Bioactive Pigments: Ecofriendly Source of Bioactive Compounds. Coatings 2022, 12, 1858. [Google Scholar] [CrossRef]
- Salla, T.D.; Astarita, L. V.; Santarém, E.R. Defense Responses in Plants of Eucalyptus Elicited by Streptomyces and Challenged with Botrytis Cinerea. Planta 2016, 243, 1055–1070. [Google Scholar] [CrossRef]
- Kim, S.S.; Lee, C.K.; Kang, S.S.; Jung, H.A.; Choi, J.S. Chlorogenic Acid, an Antioxidant Principle from the Aerial Parts of Artemisia Iwayomogi That Acts on 1,1-Diphenyl-2-Picrylhydrazyl Radical. Arch. Pharm. Res. 1997, 20, 148–154. [Google Scholar] [CrossRef]
- Saqib, M.; Iqbal, S.; Mahmood, A.; Akram, R. Theoretical Investigation for Exploring the Antioxidant Potential of Chlorogenic Acid: A Density Functional Theory Study. Int. J. Food Prop. 2016, 19, 745–751. [Google Scholar] [CrossRef]
- Keudell, K.C.; Huang, J.K.; Wen, L.; Klopfenstein, W.E.; Bagby, M.O.; Lanser, A.C.; Plattner, R.D.; Peterson, R.E.; Weisleder, D. Fatty Acids Enhanced Tubermycin Production by Pseudomonas Strain 2HS. Microbios 2000, 102, 27–38. [Google Scholar] [PubMed]
- Weber, W.; Zähner, H.; Siebers, J.; Schröder, K.; Zeeck, A. [Metabolic products of microorganisms. 175. Tetracenomycin C (author’s transl)]. Arch. Microbiol. 1979, 121, 111–116. [Google Scholar] [CrossRef]
- Remali, J.; Sahidin, I.; Aizat, W.M. Xanthone Biosynthetic Pathway in Plants: A Review. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Leetanasaksakul, K.; Koomsiri, W.; Suga, T.; Matsuo, H.; Hokari, R.; Wattana-Amorn, P.; Takahashi, Y.; Shiomi, K.; Nakashima, T.; Inahashi, Y.; et al. Sattahipmycin, a Hexacyclic Xanthone Produced by a Marine-Derived Streptomyces. J. Nat. Prod. 2022, 85, 1211–1217. [Google Scholar] [CrossRef]
- Matselyukh, B.P.; Matselyukh, D.Y.; Golembiovska, S.L.; Polishchuk, L. V; Lavrinchuk, V.Y. Isolation of Streptomyces Globisporus and Blakeslea Trispora Mutants with Increased Carotenoid Content. Mikrobiol. Z. 2013, 75, 10–16. [Google Scholar]
- Han, Z.; Zhao, X.; Zhang, E.; Ma, J.; Zhang, H.; Li, J.; Xie, W.; Li, X. Resistomycin Induced Apoptosis and Cycle Arrest in Human Hepatocellular Carcinoma Cells by Activating P38 MAPK Pathway In Vitro and In Vivo. Pharmaceuticals 2021, 14. [Google Scholar] [CrossRef]
- Shaaban, K.A.; Singh, S.; Elshahawi, S.I.; Wang, X.; Ponomareva, L. V; Sunkara, M.; Copley, G.C.; Hower, J.C.; Morris, A.J.; Kharel, M.K.; et al. Venturicidin C, a New 20-Membered Macrolide Produced by Streptomyces Sp. TS-2-2. J. Antibiot. (Tokyo). 2014, 67, 223–230. [Google Scholar] [CrossRef]
- Tan, L.T.; Mahendra, C.K.; Yow, Y.; Chan, K.; Khan, T.M.; Lee, L.; Goh, B. Streptomyces Sp. MUM273b: A Mangrove-derived Potential Source for Antioxidant and UVB Radiation Protectants. Microbiologyopen 2019, 8. [Google Scholar] [CrossRef]
- Berger, H.; Bacher, M.; Labuda, R.; Eppel, I.M.; Bayer, F.; Sulyok, M.; Gasparotto, E.; Zehetbauer, F.; Doppler, M.; Gratzl, H.; et al. Polaramycin B, and Not Physical Interaction, Is the Signal That Rewires Fungal Metabolism in the Streptomyces–Aspergillus Interaction. Environ. Microbiol. 2022, 24, 4899–4914. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, W.; El-Neketi, M.; Lewald, L.-I.; Orfali, R.S.; Lin, W.; Rehberg, N.; Kalscheuer, R.; Daletos, G.; Proksch, P. Metabolites from the Fungal Endophyte Aspergillus Austroafricanus in Axenic Culture and in Fungal–Bacterial Mixed Cultures. J. Nat. Prod. 2016, 79, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Kemung, H.M.; Tan, L.T.-H.; Chan, K.-G.; Ser, H.-L.; Law, J.W.-F.; Lee, L.-H.; Goh, B.-H. Antioxidant Activities of Streptomyces Sp. Strain MUSC 14 from Mangrove Forest Soil in Malaysia. Biomed Res. Int. 2020, 2020, 6402607. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.M.A.; Pereira, P.S.; Silva, B.I.M. da; Ribeiro, N.E.; Borba, E.F. de O.; Oliveira Tintino, C.D. de M.; Randau, K.P.; Coutinho, H.D.M.; Lima-Gomes, G.M. de S.; Chávez-González, M.L.; et al. Antioxidant, Antimicrobial and Cytotoxic Activities of Secondary Metabolites from Streptomyces Sp. Isolated of the Amazon-Brazil Region. Res. Soc. Dev. 2021, 10, e366101018974. [Google Scholar] [CrossRef]
- de Moura, P.A.; de Albuquerque Lima, T.; Ferreira, M.R.A.; Soares, L.A.L.; de Souza Lima, G.M.; Napoleão, T.H.; da Silva, M.V.; de Oliveira, A.P.S.; Paiva, P.M.G. The Relevance of Actinobacteria as Sources of Antioxidant Compounds: Evaluation of Streptomyces Isolates from Rhizosphere Collected at Brazilian Caatinga. In Microbial and Natural Macromolecules - Synthesis and Applications; Das, S., Dash, H.R.B.T.-M. and N.M., Eds.; Academic Press: London UK, 2021; pp. 401–418. ISBN 978-0-12-820084-1. [Google Scholar]
Figure 1.
Phylogenetic tree of some the bioactive isolates.
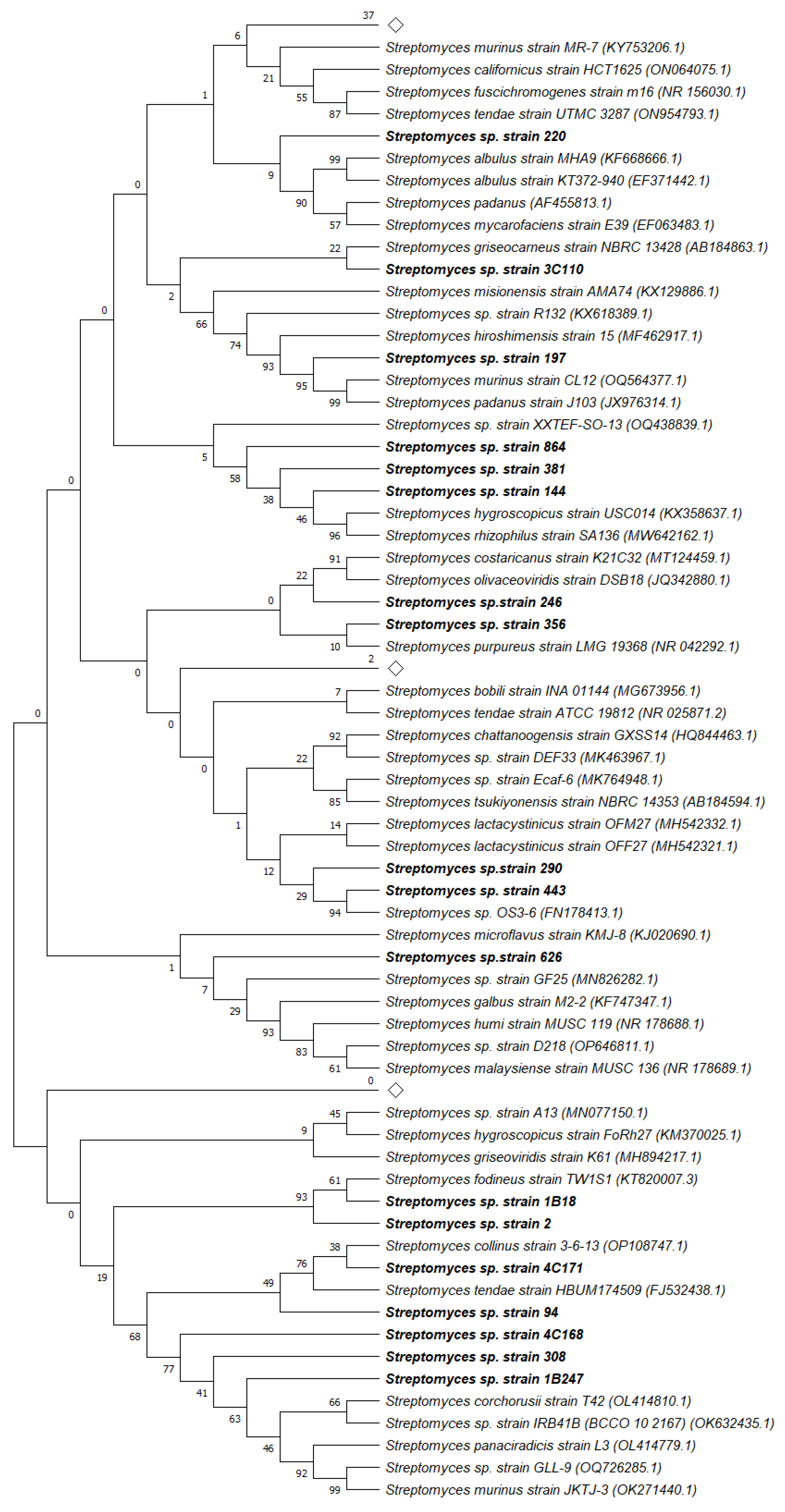
Figure 2.
Antioxidant capacity of the pigmented extracts. (a) DPPH Method (b) ABTS Method. Extracts listed according to a code comprising the codified strain (Table 4) hyphenated to the culture medium type, i.e., ISP2 (A) ISP4 (B) modified ISP9 (C) starch nitrate (D). For more information, the mean data with standard deviation (n = 3) are presented in supplementary material (Tables S1 and S2).
Figure 2.
Antioxidant capacity of the pigmented extracts. (a) DPPH Method (b) ABTS Method. Extracts listed according to a code comprising the codified strain (Table 4) hyphenated to the culture medium type, i.e., ISP2 (A) ISP4 (B) modified ISP9 (C) starch nitrate (D). For more information, the mean data with standard deviation (n = 3) are presented in supplementary material (Tables S1 and S2).
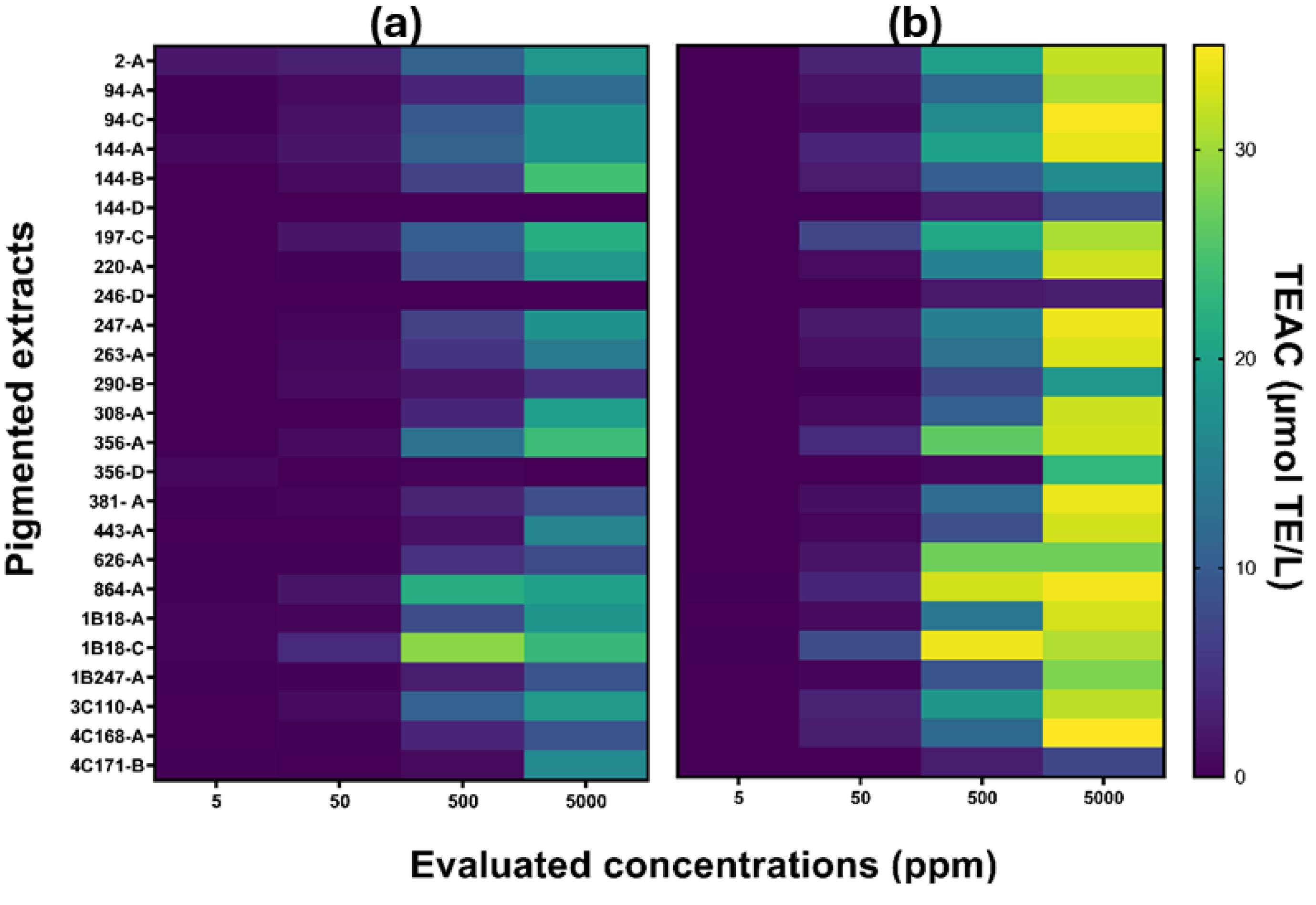
Figure 3.
Cytotoxicity test of the pigmented extracts. Human Dermal Fibroblasts (HDFa), Cervical cancer cell line (HeLa), Breast cancer cell line (MCF-7). Extracts listed according to a code comprising the codified strain hyphenated to the culture medium type, i.e., ISP2 (A) ISP4 (B) modified ISP9 (C) starch nitrate (D). For more information, the mean data with standard deviation (n = 3) are provided in supplementary material (Tables S4 to S6).
Figure 3.
Cytotoxicity test of the pigmented extracts. Human Dermal Fibroblasts (HDFa), Cervical cancer cell line (HeLa), Breast cancer cell line (MCF-7). Extracts listed according to a code comprising the codified strain hyphenated to the culture medium type, i.e., ISP2 (A) ISP4 (B) modified ISP9 (C) starch nitrate (D). For more information, the mean data with standard deviation (n = 3) are provided in supplementary material (Tables S4 to S6).
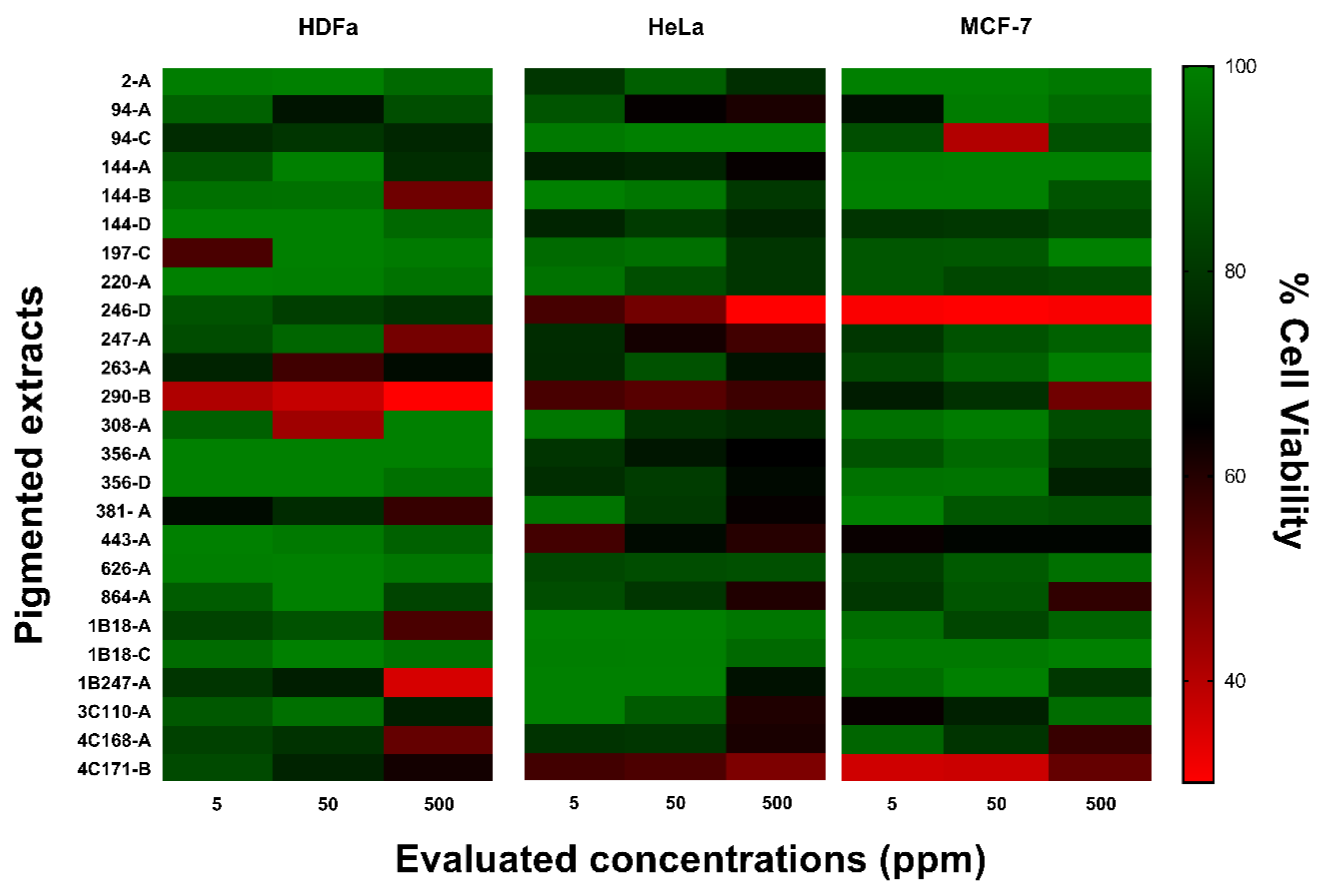
| Composition/Medium | ISP2 | ISP4 | modified ISP9 | Starch nitrate |
|---|---|---|---|---|
| agar | 15 | 15 | 15 | 15 |
| starch | 0 | 10 | 0 | 20 |
| CaCO3 | 0 | 2 | 0 | 3 |
| CuSO4·5H2O | 0 | 0 | 0.00064 | 0 |
| yeast extract | 4 | 0 | 0 | 0 |
| malt extract | 10 | 0 | 0 | 0 |
| FeSO4·7H2O | 0 | 0.01 | 0.00011 | 0.01 |
| glucose | 4 | 0 | 10 | 0 |
| K2HPO4 | 0 | 1 | 5.65 | 1 |
| KH2PO4 | 0 | 0 | 2.38 | 0 |
| KNO3 | 0 | 0 | 0 | 2 |
| MgSO4·7H2O | 0 | 1 | 1 | 0.5 |
| MnCl2·4H2O | 0 | 0.001 | 0.00079 | 0 |
| NaCl | 0 | 0.5 | 0 | 0 |
| (NH4)2SO4 | 0 | 2 | 2.64 | 0 |
| ZnSO4·7H2O | 0 | 0.001 | 0.00015 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



