
Submitted:
20 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
We developed a protein to rapidly and accurately diagnose Chagas disease, a life-threatening illness identified by the WHO as a critical worldwide public health risk. Complicating the current health situation are serological test limitations that contribute to most infected persons being unaware of their condition and therefore untreated. To improve diagnostic testing, we developed an immunological mimic of the etiological agent, Trypanosoma cruzi, by combining ten pathogen specific epitopes within the beta-barrel protein structure of Thermal Green Protein. The resulting multi-epitope protein, DxCruziV3, displayed high specificity and sensitivity as the antibody capture reagent in an ELISA platform with an analytical sensitivity that exceeds WHO recommendations. Within an immunochromatographic platform, DxCruziV3 showed excellent performance for the point of application diagnosis in a region endemic for multiple diseases, the municipality of Barcelos in the state of Amazonas, Brazil. In total, 167 individuals were rapidly tested using whole blood from a finger stick. As recommended by the Brazilian Ministry of Health, venous blood samples were laboratory tested by conventional assays for comparison. Test results suggest utilizing DxCruziV3 in different assay platforms can confidently diagnosis chronic infections by T. cruzi. Rapid and more accurate results will benefit everyone but will have the most noticeable impact in resource-limited rural areas where the disease is endemic.
Keywords:
Chagas disease
; Diagnostics
; ELISA
; Lateral Flow tests
; Synthetic protein
; T. cruzi
1. Introduction
Chagas disease, also known as American trypanosomiasis, is a potentially life-threatening illness caused by the protozoan parasite Trypanosoma cruzi [1]. Endemic to Latin America, this disease was primarily transmitted through the feces of triatomine bugs until the 1990s when programs were implemented to control Triatoma infestans, the species mostly responsible for infections inside homes [2,3].
Today, many people in tropical areas remain at risk from other species of triatomine in remote work areas or drinking T. cruzi contaminated fresh unpasteurized fruit juices [4,5]. Other modes of transmission include blood transfusions, organ transplants, and from mother to child during pregnancy, which can occur in non-endemic regions due to the migration of infected individuals [6,7,8]. Overall, up to 75 million people are at risk of infection globally [9].
Of the 6 to 7 million people worldwide that are estimated to be infected with T. cruzi, only around 10% are aware of their condition [9]. Chagas disease progresses through two phases: acute and chronic [10,11]. Because the acute phase is often asymptomatic or presents mild symptoms, the disease is frequently undiagnosed at this stage. Untreated, the disease progresses to the chronic phase, which can remain silent for decades. In about 30% of chronically infected individuals, the disease can cause cardiac, digestive, or neurological complications. The most severe form of Chagas disease involves cardiomyopathy, leading to heart failure, arrhythmias, and sudden death [12,13,14]. Gastrointestinal manifestations include megaesophagus and megacolon, which can cause severe digestive issues [15,16].
Although serological diagnosis plays an essential role in identifying persons with a chronic infection for managing Chagas disease, at least two different serological tests are recommended for a definitive diagnosis. Cross-reactivity with antibodies generated against other parasitic infections has been a major issue for specificity. Moreover, many commercial tests show variability in sensitivity according to the geographical origins of an individual [17,18]. Our solution focuses on epitopes specific to T. cruzi and grouping them together by their incorporation into the loops of the beta barrel structure from green fluorescent protein [19]. While two resulting recombinant proteins, DxCruziV1 & V2, were recovered in inclusion bodies, in-house ELISAs showed that each displayed nearly perfect specificity and excellent sensitivity using two overlapping sets of ten epitopes.
To improve accessibility, affordability and speed to screen populations in resource-limited areas, where the disease burden is highest, the Brazilian Unified Health System suggested that using a point-of-care diagnostic test could be a viable alternative for diagnosing chronic Chagas disease [20]. A rapid test would also be strongly favored for use during prenatal examinations and during delivery when Chagas disease is suspected in the pregnant woman [21].
To meet the government’s goals, we exchanged the beta-barrel to a fully synthetic construct, Thermal Green Protein [22], with the intention of improving the production of soluble protein for use in both ELISA and lateral flow platforms. Previously characterized patient sera and international biological standards for Chagas disease were used to evaluate the performance of the resulting protein, DxCruziV3, in an ELISA format. Its utility in a lateral flow platform was also evaluated by the comparison field results to laboratory-based assays. The results suggest that the newest version of our multiepitope protein, DxCruziV3, could rapidly deliver a high confidence diagnosis for Chagas disease at the point of application as well as confirmatory results in an ELISA format.
2. Materials and Methods
2.1. Ethical Approval
All procedures were performed in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki) for experiments involving humans and complied with relevant laws and institutional guidelines. Research protocols were pre-approved by the Research Ethics Committee of the Oswaldo Cruz Foundation involving Humans (CAAE: 52892216.8.0000.5248). In compliance with Resolution 466/12 of the National Health Council, participants were included after accepting and signing a Consent Form.
2.2. Serological Panels and Participant Profile
Serological samples were obtained from the Central Public Health Laboratories (LACENs) in the Brazilian states Ceará, Maranhão, Paraíba and Sergipe, as well as samples from the city of Barcelos in Amazonia, Brazil. Samples from the LACENs had been diagnosed for chronic Chagas disease by accepted protocols at the respective centers and reevaluated with Chagastest ELISA Rec. v3.0 (Wiener Lab, São Paulo, BR) and IFA-Chagas (Bio-Manguinhos, Rio de Janeiro, BR). The WHO International Biological Reference Standards - NIBSC 09/188 and NIBSC 09/186 that represent the TcI (Mexico) and TcII (Brazil and Chile) T. cruzi DTUs, respectively, were used to determine analytical sensitivity by a dilution series from 1:2 to 1:256 performed in triplicate [23].
In the municipality of Barcelos, AM, BR, 167 participants were recruited. They either lived in riverside communities or in boats temporarily anchored on the banks of the Negro River. Inclusion of indigenous persons was graciously permitted by the Special Health District for Indians (DSEI Alto Rio Negro). In total, the cohort included 134 people with a history of frequenting plantations with piassava palm trees, a region reported to support the presence of triatomines [24]. Two participants did not respond to this question and 31 said they did not frequent the piassava extraction area.
At the time of sampling, only five participants were suspected or known to have contracted the disease. One participant displayed symptoms of an acute infection that was confirmed by a thick drop slide examination performed by the Health Surveillance Foundation FVS of Barcelos. Four participants were undergoing Benznidazole treatment for an acute infection that was reported after drinking natural patauá juice [25].
Blood procurement varied by requirement. A digital puncture was used to obtain whole blood for application in the immunochromatographic rapid tests prototypes (two, 10 µL samples). A brachial puncture was also performed to collect 5 mL of whole blood into a Vacutainer® (BD, Franklin Lakes, New Jersey, US) tube containing clotting factor. Serum was separated by centrifugation (2,000 x g, 10 min @ 25 °C), frozen at -20 °C, shipped to Manus, AM, BR for transport to Rio de Janeiro, BR on dry ice.
2.3. Cloning, Expression, and Purification of DxCruziV3
The amino acid sequence of DxCruziV3 was back translated and optimized for expression in Escherichia coli (Figure S1). The 3’ was extended to include a ribosome binding site and a 6xHis tag was added to the carboxy terminus. A gene fragment flanked by XbaI and XhoI was synthesized (Integrated DNA Technologies, Coralville, IA, US) and inserted into the corresponding sites of pET28a. The final clone was confirmed by sequencing. Procedures to produce recombinant protein were pre-appoved by the Internal Biosecurity Commissions of the IOC (No°5/5342017) and Bio-Manguinhos (N° 006/2022) as a Class I biological risk.
For inclusion body-derived DxCruziV3 (V3ib), a confirmed clone of transformed E. coli BL21 (DE3) bacteria was cultured to log phase in 100 ml of Luria Broth (LB) with kanamycin (35 µg/ml) overnight at 37 °C with shaking (200 rpm). Next, 2 ml was inoculated into 100 mL of LB and cultured to an OD of 0.6 before induction with isoproypl-ß-D-thiogalactoside (IPTG; 1 mM). After 3h, bacteria were collected by centrifugation (3,500 x g, 20 min @ 25 °C), resuspended in lysis buffer A (50 mM Na2HPO4, 500 mM NaCl; pH 7.4), and sonicated (amplitude 20 in pulses of 30 sec with a 60 sec pause for 2 min). Inclusion bodies were collected and washed twice with lysis buffer plus detergent (1% Triton X-100) by centrifugation (16,000 x g, 20 min @ 4 °C). The final pellet was solubilized with 4M urea in lysis buffer A, clarified (16,000 x g, 20 min @ 4 °C), and applied to a pre-equilibrated 1 mL HisTrap® HP column (Cytiva Latin America, São Paulo, BR) attached to an Äkta 10 chromatograph system (Cytiva Latin America). The column was washed with 10 column volumes (CV) of lysis buffer A with 4M urea followed by 3 CV of lysis buffer A with 4M urea and 22 mM imidazole. Bound V3ib was eluted by a gradient of imidazole up to 500 mM. Elution fractions were analyzed by SDS-PAGE and western blot (anti-6xHis antibody, ThermoFisher, São Paulo, BR) to identify fractions to combine for dialysis to reduce the molarity of urea to 1M with PBS. The final concentration of V3ib was determined by the Bradford method [26].
To produce soluble DxCruziV3 (sV3), a colony of the confirmed clone in E. coli BL21 (DE3) was cultured in 100 mL of LB with kanamycin (35 µg/ml) in a 37 °C incubator shaker at 220 rpm. After 16h, the preculture was used to inoculate 2L of LB with kanamycin in a bioreactor (Biostat® B, Sartorius, Göttingen, DE) maintained at 37 °C with an rpm of 500, an aeration rate of 1VVM, and a pH of 6.8. At an OD600 of 0.6, expression was induced with IPTG (1 mM). After an additional 3h, bacteria from 750 mL of culture were harvested by centrifugation and resuspended in 200 mL of lysis buffer B (50 mM sodium phosphate, 500 mM NaCl, 5% glycerol, and 20 mM imidazole; pH 7.5) containing cOmplete®, EDTA-free protease inhibitor cocktail (Roche, Basil, CH). Cells were disrupted by pressure (PandaPlus1000, GEA Niro Soavi, Parma, IT) at 1000 bar and cycled for 30 min at 4 °C. The extract was centrifuged (16,000 x g, 30 min @ 4 °C) and the supernatant containing sV3 was filtered (0.22 µm). A 5 mL HisTrap® HP column (Cytiva Latin America) attached to an Äkta Pure chromatography system (Cytiva Latin America) was pre-equilibrated with 10 CV of lysis buffer B before loading sample. It was washed with 10 CV of lysis buffer B followed by 5 CV of lysis buffer B with 50 mM imidazole and then 5 CV of lysis buffer containing 100 mM imidazole. Protein was eluted in a linear gradient range of imidazole in lysis buffer B from 100 to 500 mM. Fractions were analyzed by SDS-PAGE (12%) to pool those with sV3. Buffer exchange and sample concentration were performed using the Amicon® Stirred system (Merck, São Paulo, BR) with a cellulose regenerated membrane (cutoff 5 kDa). The concentration of sV3 was calculated from its absorbance at 280 nm and calculated molar absorption coefficient (30370 M-1cm-1). Purity was estimated by densitometry (GS-900, Bio-Rad, Hercules, CA, US) of samples resolved by SDS-PAGE (12%).
2.4. Enzyme-Linked Immunosorbent Assays
Two commercial ELISA kits, Biolisa Chagas Recombinant (Bioclin, Belo Horizonte, MG, BR) and Chagatest recombinant v3.0 (Wiener Labs, Rosario, AR), were executed as directed by the manufacturer. In-house ELISA assays consisted of 96-well plates (Nunc™ MicroWell™, Rochester, NY, US) sensitized overnight at 4 °C with 500 ng/well of V3ib diluted into sodium carbonate-bicarbonate buffer (0.05 M, pH 9.6). Next, plates were washed 3x with PBS and blocked at 37 °C with 1X PBS-T (pH 7.4 with 0.05% Tween® 20) with 5% powdered non-fat milk. After 1h, buffer was removed and serological samples diluted as indicated in 100 µl of PBS-T with 1% powdered non-fat milk or the dilution series of the WHO International Biological Reference Standards was added followed by another 1h incubation at 37 °C. After three washes with PBS-T, goat anti-human IgG Fc specific antibody conjugated to HRP (Cat # A0170, Sigma-Aldrich, St. Louis, MO, US) was added at a dilution of 1:60,000 followed by another 1h incubation at 37 °C. After three final washes with PBS-T, One Step – TMB Ultra (Scienco, Santa Catarina, BR) was added followed by an incubation at room temperature for 15 minutes in the dark. Enzymatic activity was stopped by the addition 0.5M H2SO4 and the absorbance was measured at 450 nm in an automated plate reader (Hidex Sense, Hidex, Turku, FI).
2.5. Lateral Flow Immunochromatographic Assay Preparation
Lateral flow assays prepared with V3ib, (V3ib-LFA) began with the placement of a strip of nitrocellulose membrane to the center segment of a 3-segment, adhesive card (30 cm x 5 cm) followed by the simultaneous application of lines of V3ib (1 mg/mL at 1.5 µl/cm) and Protein A (1 mg/mL at 0.75 µl/cm) with an XYZ Platform Dispenser (HM3030, Shanghai Kinbio Tech, Shanghai, CH). A glass fiber membrane previously treated in buffer (0.05 M Tris, 0.5% Casein, 1.0% PVP, 0.05% Tween®20, 0.02% sodium azide, pH 8.0) was impregnated with a spray of colloidal gold labeled rabbit anti-human IgG antibody (ThermoFisher). After drying at 37 °C for 1hr, it was applied to the first segment of the adhesive card. A thick absorption pad was placed in the third segment. Strips (4 mm) were prepared on a programable strip cutter (ZQ2002, Shanghai Kinbio Tech) that were assembled in cassettes.
Lateral flow tests prepared with sV3 were manufactured by Bio-Manguinhos (FIOCRUZ, Rio de Janeiro, BR) under GMP conditions. The test strip consisted of a segment of nitrocellulose membrane immobilized on adhesive card. Solutions of sV3 (500 ug/mL, 0.85 µl/cm) and Protein A (50 mg/mL, 1 µl/cm) were applied as parallel lines by a lateral flow dispenser (Isoflow Dispenser, Imagene Technology Inc., Lebanon, NH, US). Membranes were placed in an oven at 37 °C for 30 min to fix the proteins. A conjugation of a colloidal gold nanoparticle with protein A was deposited on a glass fiber, dried for 1h in a 37 °C oven, and added to the adhesive card. After including an absorbent pad manually with the aid of a template, strips (5 mm) were obtained with a guillotine shear cutter (Matrix 2360, Kinematic, Sonora, CA, US) that were fitted into plastic cassettes and closed with a compressor roller (AR3000, BioDot, Irvine, CA, US).
2.6. Indirect Immunofluorescence
The indirect immunofluorescence assays (IFI Chagas, Bio-Manguinhos) were performed according to the included instructions.
2.7. Statistical Analysis
Students t-test analyses, ANOVA and receiver operating characteristic (ROC) calculations were performed using Prism software (V8.0.2, GraphPad, Boston, MA, US). Statistical significance was defined as a p value less than 0.05. The ROC analysis was used to define the cutoff value for reactive and non-reactive samples as well as to calculate the reactivity index (RI) as the division of an absorbance value divided by the cutoff (O.D./cutoff). Results within 10% of an RI of 1 were considered inconclusive.
3. Results
3.1. Design and Production of DxCruziV3
The third version of DxCruzi maintained the epitopes contained in the second version with an extension of epitope 6 by four amino acids on the amino terminus (Table 1). The major difference among versions was a switch in the beta-barrel sequences to those of Thermal Green Protein (TGP). As a fully synthetic construct, the potential presence of pre-existing antibodies in patient serum would be further reduced compared to eGFP. In addition, TGP contains specific amino acid changes in the beta sheets of the barrel structure that diminish aggregation and improve stability, all desirable characteristics for an immunological mimic of a pathogen for serodiagnosis. As a first prediction for the potential to successfully produce protein, the projected primary sequence was submitted to the I-TASSER server [27,28].
A projection of three views of the computationally determined tertiary structure
1Based on the sequence in FPbase (https://www.fpbase.org/protein/thermostable-green-protein).2Colorization of the sequence in the model presented in Figure 1. displayed a fully intact beta barrel structure that suggested that the insertion of the epitope sequence did not disrupt its formation (Figure 1). This was confirmed by the robust expression of DxCruziV3 using the pET28a plasmid based on T7 RNA polymerase [39], which distributed to both the soluble fraction (~15%) and inclusion bodies (~85%). Due to the greater yield from inclusion bodies, insoluble DxCruziV3 (V3ib) was initially purified by IMAC after solubilization in 4M urea (Figure S2). To reduce the urea concentration, samples were dialyzed against PBS to a final concentration of 1M.
3.2. Performance of DxCruziV3 in ELISAs
In-house ELISAs were generated using 200 ng of purified V3ib maintained in 1M urea to sensitize each well followed by an incubation with patient serum diluted 1:200. As can be seen in Figure 2A, the detection of human IgG showed excellent performance for differentiating reactive, infected from non-reactive, not infected individuals. A ROC analysis calculated that an area under the curve of 0.9892 with a standard error of 0.0096 and a P value <0.0001. With a cutoff of 0.153 OD450, the ELISA presented a sensitivity of 96.0% and a specificity of 100% (Table 2). Using the cutoff, we determined the reactivity index for the detection of cross-reactivity and non-specific reactivity (Figure 2B). Using patient serum infected with dengue or Leishmania spp., only one of 57 samples showed a reactivity above 1, which was not previously diagnosed for Chagas disease. Importantly, none of the healthy samples showed reactivity.
3.3. Analytical and Geographical Sensitivity of DxCruziV3
The analytical sensitivity of the in-house ELISA assay was assessed by the application of the two WHO International Biological Standards for Chagas disease that represent patient sera from areas predominantly occupied by T. cruzi DTUs TcI (NIBSC 09/188; Mexico) and TcII (NIBSC 09/186; Brazil) over a 2-fold serial dilution. After converting absorbance values to a reactivity index (RI), the use of V3ib as the antibody capture reagent resulted in an assay that exceeds the minimum recommended dilution value by a factor of 4 (Figure 3A). One aspect of the two standards is to be able to evaluate the potential for differences in sensitivity related to the geographical origin of the patient samples that could reflect the distribution of different subtypes of T. cruzi. To extend this to our panel of patient serum that represent populations across northern Brazil, the RI was calculated and plotted separately based on geography (Figure 3B). Following an INOVA statistical analysis with a Kruskal-Willis test, the performance of the ELISA was significantly different in samples from the state of Maranhão, BR to the three more eastern states, but not Amazonas, BR.
3.3. Lateral Flow Immunochromatographic Prototypes
The high level of sensitivity observed with V3ib encouraged us to develop a lateral flow assay (LFA), which in general are considered to present lower sensitivities than an ELISA platform. A single lot of 300 LFAs was produced with DxCruziV3 purified from inclusion bodies. Two different flows were utilized to apply the control line of Protein A (0.5 µl/cm) and test lines of V3ib (1.5 µl/cm) to achieve 200 ng and 450 ng per strip, respectively. Over the course of evaluating buffer compositions, the primary target performance profile was to eliminate the appearance of any coloration of the test line when non-reactive sera was applied. After finalizing the conditions for the V3ib-LFA, 100% sensitivity was confirmed using a pool of low titer Chagastic patient sera and another with a high titer (Figure S3). Specificity was evaluated with serum pools with serum from patients with cutaneous leishimaniose, visceral leishimaniose and dengue as well as healthy individuals and persons diagnosed as negative for chronic Chagas disease.
In parallel, a single lot of LFAs was independently developed at Bio-Manguinhos using sV3 (Figure S4). Internal quality control assays determined a sensitivity of 96% and a specificity of 100% (Table S1). A more extensive comparison was made with a panel of patient serum collected in the states of Minas Gerais and Piauí in 2022 and 1999, respectfully. As can be seen in Figure 4, samples reactive in the sV3-LFA (blue) primarily displayed a higher reactivity index than non-reactive samples (red). Although some of the samples non-reactive by the LFA showed reactivity by ELISA, in no instance was there reactivity in the LFA and no reactivity by ELISA suggesting that the LFA had the same high specificity but with a lower sensitivity than the ELISA platform. After the standardization phase, production lots of the two independent prototypes were evaluated by an application in the field followed by an in-depth laboratory analysis of the patient samples.
3.4. Performance of the LFA Prototypes
The municipality of Barcelos, AM, BR was chosen as the geographical site for testing the LFAs due to its long standing history as the site for studies on Chagas disease in the Amazon rainforest. A total of 16 communities within the municipality were represented by the recruited individuals. Overall, 43 of 167 participants (25.7%) were reactive by both LFAs (Table 3). Two were reactive by V3ib-LFA and non-reactive by sV3-LFA along with two reactive by sV3-LFA and non-reactive by V3ib-LFA. The Nova Jerusalém community had the highest prevalence of reactive LFAs (19 of 52 reactive), followed by Mariuá (12 of 34 reactive) (Table S2). Men showed a higher prevalence than woman (32 vs. 11) and the vast majority were over the age of 40 years old (Table S3).
In the lab, each correlate serum sample was subjected to a series of assays (Table 3). Initially, the two LFAs were repeated on field reactive samples along with a random subset of non-reactive due to a limitation in the number of tests produced in the first lot. In the place of 10 µl of whole blood, 5 µl of serum was applied under the controlled environment of the lab. One of the samples that was reactive in the field by V3ib-LFA was non-reactive in the lab while three non-reactive samples showed a weak reactivity in the lab. For sV3-LFA, three samples reactive and one non-reactive in the field presented the opposite result in the lab.
The next assays were an indirect immunofluorescence (IFI) assay and the commercial ELISA kit from Bioclin in duplicate. In addition, the commercial ELISA kit from Wiener and in-house ELISAs utilizing V3ib were employed. The results were defined as reactive (1) or non-reactive (0) and summed to group the results based on consistency between platforms. Of the 167 samples, 150 presented with a high level of agreement between the results from the assays. Within this group, both LFAs were calculated to have 100% sensitivity and specificity (Table 4). The ranking by decreasing sensitivity was the IF assay (93.8%) and then the ELISAs by Bioclin (90.6%), with V3ib (87.5%), and by Wiener (71.9%). For specificity, the V3ib and Wiener ELISAs showed very high performance followed by the IF assays and the Bioclin ELISA. The remaining 17 samples were interrogated further to make a best evidence final call on the status of the patient through up to 18 different tests. The IFI, Bioclin and Wiener assays were executed as instructed. A serial dilution of sera (1:50, 1:100 and 1:200) was applied to the V3ib-ELISA that showed a bifurcation of the samples between reactive and non-reactive (Figure S5). Ultimately, the calculations decreased for each assay although the effect was greater on sensitivity than specificity.
4. Discussion
While the connection between T. cruzi and Chagas disease was made by Carlos Chagas over 100 years ago [40], this parasite has been plaguing South America for thousands of years as evident by the detection of T. cruzi kinetoplast in the DNA of human mummies [41,42]. Today, it continues to be a predominantly silent life-threatening disease [43]. A troubling trend is the spread of Chagas beyond endemic regions with human migration and changes in climate [44]. With a goal of ending transmission, the 2030 WHO roadmap for neglected tropical diseases seeks to disrupt transmission routes and greatly increase treatment for afflicted individuals [45]. These goals cannot be met without an effective, and economical, diagnostic test.
It is well known that current serological tests for Chagas disease do not meet the minimal target performance profile [46]. Nearly all display cross-reactivity with other diseases, especially Leishmania spp. [47,48], and variances in sensitivity related to geography [18]. Both issues contribute to the need for two independent tests that can extend the time from sample collection to final diagnosis by several months. Recently, we reported on two multi-epitope proteins, DxCruziV1 & DxCruziV2, that could overcome these issues [19]. Built on the beta-barrel structure of eGFP, each displayed excellent qualities to serve as the antibody capture reagent in ELISAs.
Here, the beta barrel was changed to that of Thermal Green Protein (TGP) to yield DxCruziV3. The rapid folding and high stability of TGP was predicted to improve solubility of the DxCruziV3 compared to the earlier versions. As a fully synthetic construct, the move to TFP was also anticipated to minimize background from non-specific antibody interactions. As expected, computer modeling showed a retention of the barrel structure with the epitopes exposed on surface (Figure 1). To decisively demonstrate its potential to deliver high confidence diagnoses, DxCruziV3 was incorporated into ELISAs and rapid test platforms for interrogation with challenging patient samples and conditions. The resulting rapid test performances were extensively compared to three commercial tests.
As with the previous versions, DxCruziV3 proved to be easily expressed to high levels (>200 µg/ml) in bacteria with ~15% distributed in the soluble fraction. However, considering the yield and initial purity of protein in inclusion bodies, we decided to generate in-house ELISAs with V3ib. Using 200 ng/well, this could translate to nearly a million assays per liter of culture. To calculate specificity and sensitivity, we screened a panel of patient serum obtained from Brazilian state reference laboratories (LACENs) that had been diagnosed by the recommended protocol of two different assays followed by a third assay for divergent results. From 75 positive samples and 104 controls (Figure 2a), a specificity of 100% and a sensitivity of 96.0% were calculated by a ROC analysis (Table 1). Only a single sample from a panel of sera of patients diagnosed with either cutaneous Leishmaniosis, visceral Leishmaniosis or dengue showed a reactivity index above one (Figure 2b). As the sample had not been previously screened for Chagas disease and these diseases are co-endemic, there is a good probability it could represent a dual infection rather than cross-reactivity. This potential highlights a difficulty related to reagent development in which most control patient serum samples are only defined for a single condition/disease. When that sample also represents the target profile, its inclusion can skew calculations towards a poorer performance.
While all of the serum samples in our panel for the ROC analysis have an excellent provenance, they have been stored for years and subjected to an unknown number of freeze/thaws. For this reason, it was important to evaluate the V3ib-ELISA performance utilizing the two International Biological Standards for Chagas disease, which permits direct comparisons in analytical sensitivity between assays [49]. The initial flatness of the curve suggests that the available antibody binding sites were saturated up to a dilution of 1:16 (Figure 3a). Notably, a reactive index of greater than one was measurable through to a dilution of 1:256. This exceeds the minimum target dilution factor of 1:64 set by the WHO, which no commercial test in Brazil has achieved [50].
While no significant difference was observed between the two standards to suggest an influence of geographic origin on performance, significant differences were calculated between our samples from Maranhão compared to Ceará, Paraíba, and Sergipe, but not Amazonas (Figure 3b). This result could reflect a greater loss of antibody titer during cold storage for low titer samples, a small sample size, or could be restricted to the northern reaches of Brazil. It remains to be determined if the initial antibody titer influences retention during cold storage. Expanding the application of the assay to more regions of Brazil, other South American countries, and screening more recently obtained patient samples should conclusively demonstrate geographical differences. If it persists, and compromises performance, a new group of epitopes could be organized to build a new reagent. Since the design of DxCruziV3, a complete atlas of epitopes in T. cruzi has been described that examined differences across the Americas [51].
It is clear our protein can aid with the creation of a high performance ELISA that will aid in managing Chagas disease. Equally importantly, DxCruziV3 can be used in a lateral flow test format, which could rapidly deliver actionable results in remote endemic areas with an insufficient healthcare infrastructure to support laboratory-based tests but harbor a greater at risk population. We had an opportunity to generate LFAs that utilized either sV3 or V3ib completely independently of each other, which covered protein production, purification, and cassette assembly. Using pools of patient serum, the V3ib-LFA was assessed to be 100% specific and sensitive while the post-production quality control results for sV3-LFA returned a specificity of 100% and as sensitivity of 96%. For a comparison of sensitivity between the LFA and ELISA formats, sV3-LFA was investigated with two serum panels that had been analyzed by V3ib-ELISA. While one panel had been stored nearly 25 years, the results suggest that the LFA format is less sensitive but retains a high specificity (Figure 4).
The municipality of Barcelos, AM, BR was selected as the real world test site to evaluate the LFA prototypes. This area has the archetype characteristics for the most challenging demographics and geographics: 1) it is a location where conventional tests have the displayed low sensitivity; 2) it is endemic for multiple other diseases including malaria, dengue and leishmaniosis; 3) many members of the community spend months on plantations that house T. cruzi transmitting triatomines [52]; 4) the closest hospital is hours away by boat; 5) only two communities, Mariuá and Piloto, have an electrical grid while the others rely on diesel generators; 6) it is an archipelago that is difficult to access. To reach Barcelos, test kits were sent by plane from Rio de Janeiro, RJ, BR to Manaus, AM, BR, and then traveled by boat for two days. While the boxed kits contained prototype LFAs sealed into individual foil packs with a silica gel pouch, no special effort was made during transport to avoid sunlight or the higher temperatures of springtime in the Amazon in the middle of a river.
In practice, the results of the LFAs at the point of application were clearly non-reactive or reactive with no failures to mark the control line. Forty-three (43) individuals were suspected to be infected by T. cruzi and most reported working with piassava collection. To determine the validity of the results, venous blood samples were collected to obtain serum for laboratory analyses. These proved to provide less definitive results. In the end, six different assays were performed multiple times (Table 3). For 150 samples, there was a high level of agreement to define a sample as reactive or non-reactive, which suggested that the LFA results agreed to the highest possible extent with all the others. The remaining 17 samples required many other assays to reach a final call that still presented disagreements between assays.
Regardless, the final calculation showed a small decrease in specificity for the LFAs, V3ib-ELISA and Wiener ELISA that remained above 95%. This most likely reflects the inclusion of many of the same epitopes in the capture reagent of the Wiener ELISA as in DxCruziV3. With the IFI assay, the fixed parasites would contain natural proteins that present the highest number of possible antibody binding sites, but also a higher number of non-specific sites that would decrease specificity [53]. The greatest impact was on sensitivity for all assays except the V3ib-ELISA. From the V3ib-ELISA assays on divergent samples, serial dilutions of 1:50, 1:100, and 1:200 revealed that the highest dilution could not definitively detect all reactive samples. In comparison, both of the lower dilutions showed a good differentiation and the results from two assays at 1:100 were used for the calculations.
Overall, the results suggest that this subgroup of reactive samples from the Amazon had anti-T. cruzi antibody titers near the limit of detection for the LFAs, and below the limit of the commercial tests. As the ELISAs from Wiener and Bioclin as well as the IFI assay use 10 µl of patient serum, attempting to increase sensitivity with a greater volume of sample would most likely be impractical. The volume used in the LFAs is also at a maximum, but the calculated sensitivity suggests that it is only slightly less than the V3ib-ELISA at a dilution 1:100. This would be equivalent to a reactivity index approaching one, which converges with the analytical sensitivity of V3ib-ELISA at a dilution of 1:256 and suggests that both serodiagnostic platforms would satisfy WHO targets.
In the case of Chagas disease, we would argue that high specificity should take precedence over high sensitivity, more so if a low cost test is available. False positives from a lower specificity would necessitate a need for a confirmation of the diagnosis before a medical decision since a subset would not require intervention. For Chagas disease, the available drugs require a long treatment period and have many undesirable side effects [54]. In contrast, a false negative would represent an undetectable antibody titer, which is also an end point for successful treatments [55,56]. Low titer would suggest a less active immune response to a less aggressive infection. Changes in an infection, as reflected by this immune response, could be detected by a yearly test for persons originating from or who had worked in endemic regions. Ultimately, a false negative result would still allow all positive individuals to receive immediate medical advice since there would be no doubt a reactive person is infected. Combined with a confirmatory ELISA with higher sensitivity, persons under medical supervision could be better accommodated.
While the absence of a definitive diagnosis for the patient samples gathered in Barcelos excludes an absolute calculation of specificity and sensitivity for the LFA protypes, it is clear that they exceed those of commercially available ELISA kits. Increasing the number of individuals tested and expanding the geographical areas to other regions of Brazil and other countries will demonstrate the level confidence one can associate with the results. Chagas disease has been on ongoing challenge for developing an immunological reagent and we anticipate that the platform utilized to create DxCruzi will have applications for other diseases and infectious agents.
5. Conclusions
Our multiepitope protein DxCruziV3 represents a technology breakthrough for grouping a large number of epitopes into a single molecule. The experimental design and results suggest that as an immunological reagent it addresses current test limitations for the diagnosis of the chronic Chagas disease. The outcome is two-fold: a high-performance ELISA for improving the management of Chagas disease, and a qualified lateral flow assay for rapidly delivering actionable results. Their analytical sensitivities are greater than the recommendation by the WHO and their high specificity suggests that these assays would be refractory to co-infections. As a single recombinant protein produced in bacteria, its production on a larger scale could allow tests to be manufactured in greater quantities and at a more affordable cost. Together, the attributes of DxCruzi can facilitate diagnosis, seroepidemiology investigations, quality control, and improve studies in endemic areas.
6. Patents
A.M.D., S.G.D-S., and D.W.P. are listed as inventors on patents submitted in Brazil (BR10.2019.017792.6), USA (PCT/BR2020/05034 1), Europe (PCT: 26/06/2023), India (PCT: 26/06/2023), and China (PCT: 26/03/2023) filed by and under the administration of FIOCRUZ that may serve as a future source of funding.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprint.org, Table S1: Quality control of sV3-LFA produced by Bio-Manguinhos; Table S2: Test sites of the V3-LFA prototypes in the municipality of Barcelos, AM, BR.; Table S3: Sex, age, and rapid test results of participants with percent reactive per group; Figure S1: DxCruziV3. Amino acid sequence with the back translated DNA coding sequence along with the 5’ extension as well as the restriction sites of XbaI and XhoI (underlined) for insertion into pET28a; Figure S2: Preparation of DxCruziV3 from inclusion bodies. An SDS-PAGE (12%) separation of equal sample volumes (10 µl) from a representative purification of DxCruziV3 ( 36 kDa) with HisTrap™ column on an Äkta chromatograph system. Lane 1- Molecular weight standard (kDa); 2- Solubilized inclusion body fraction (load) 3- Flow through; 4- Elution Fraction 20; 5- Elution Fraction 24; 6- Elution Fraction 25. Figure S3: Reactivity of serum pools in a V3ib ELISA. The development of the V3ib lateral flow assay employed serum pools assembled from patients with chronic Chagas disease and a high (Sergipe) or low (Barcelos) antibody titer, cutaneous leishmaniasis (CL), visceral leishmaniasis (VL), dengue, or negative for Chagas disease from Barcelos (Barcelos (-)). Each pool combined 10 individual serological samples and applied to a V3ib-ELISA at a dilution of 1:200. CO = cutoff. Figure S4: Preparation of soluble DxCruziV3. (a) A chromatograph of the metal (Ni-NTA) affinity purification of DxCruziV3 with the measured absorbance units (280 nm; left y-axis) of the eluate and the percentage of 500 mM imidazole (right y-axis). (b) An SDS-PAGE (12%) analysis of the pool of fractions 16-41 at 1 µg and 5 µg total protein in comparison to molecular weight standards. (c) A densitometry of the lane with 5 µg of protein with the percentage of signal for each protein band. Figure S5: Serial dilution analysis of antibody titer in select patient samples with divergent serodiagnostic results. Serum dilutions of 1:200, 1:100, and 1:50 were screened by V3ib ELISA that either showed little difference in absorbance values (non-reactive, red) or increasing values (reactive, blue).
Author Contributions
Conceptualization, Carlos M. Morel, Salvatore G. De-Simone, Angela C. V. Junqueira and D. Provance, Jr; Data curation, Evandro R. Dias and Andressa M. Durans; Formal analysis, Evandro R. Dias, Andressa M. Durans, Guilherme C. Lechuga, Gabriela S. Esteves and D. Provance, Jr; Funding acquisition, Carlos M. Morel, Salvatore G. De-Simone and D. Provance, Jr; Investigation, Evandro R. Dias, Andressa M. Durans, Barbara B. Succar, Luiz André L. T. Pinto, Guilherme C. Lechuga, Mariana G. Miguez, Janaina Figueira-Mansur, Ana P. C. Argondizzo, Aline R. Bernardo and Rafaela L. Diniz; Methodology, Evandro R. Dias, Andressa M. Durans, Barbara B. Succar, Luiz André L. T. Pinto, Guilherme C. Lechuga, Mariana G. Miguez, Janaina Figueira-Mansur, Ana P. C. Argondizzo, Aline R. Bernardo and Rafaela L. Diniz; Project administration, Carlos M. Morel, Salvatore G. De-Simone and D. Provance, Jr; Resources, Gabriela S. Esteves, Edimilson D. Silva, Carlos M. Morel, José Borges-Pereira, Salvatore G. De-Simone, Angela C. V. Junqueira and D. Provance, Jr; Supervision, Gabriela S. Esteves, Edimilson D. Silva, Salvatore G. De-Simone, Angela C. V. Junqueira and D. Provance, Jr; Validation, Evandro R. Dias, Andressa M. Durans, Edimilson D. Silva, José Borges-Pereira, Salvatore G. De-Simone and D. Provance, Jr; Visualization, Evandro R. Dias and Andressa M. Durans; Writing – original draft, Evandro R. Dias, Andressa M. Durans and D. Provance, Jr; Writing – review & editing, Evandro R. Dias, Andressa M. Durans, Carlos M. Morel, Salvatore G. De-Simone and D. Provance, Jr. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by: INOVA PROGRAM/FIOCRUZ to D.W.P. (VPPIS-004-FIO-18-52); Carlos Chagas Filho Foundation for Research Support of the State of Rio de Janeiro (FAPERJ) to S.G.D-S. (#110.198–13, #210.470–2021, #200.960/2022); National Council for Scientific and Technological Development (CNPq)/Coordination for the Improvement of Higher Education Personnel (CAPES)/FAPS-National Institutes of Science and Technology Program to C.M.M. (#465313/2014-0); CNPq Productivity Grant in Technological Development and Innovative Extension - DT to C.M.M. (04001/2022-8); Coordination of Superior Level Staff Improvement CAPES to E.R.D. (#88887.493091/2020-00); Brazilian Council for Scientific Research/CNPq to S.G.D-S. (#305.157/2020-5). The funding agencies had no role in study design, data collection, data analysis, publication decisions, or manuscript preparation.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the Oswaldo Cruz Institute (CAAE: 52892216.8.0000.5248; January 2017).
Informed Consent Statement
Informed, signed consent to obtain blood samples for this study was obtained from all subjects involved in the study and their identity has been maintained anonymous.
Data Availability Statement
Dataset available on request from the authors.
Acknowledgments
We greatly appreciate the excellent review of English by D.S. Provance. Patient recruit and sample collection were facilitated by the contributions of the Barcelos Municipal Health Department, Barcelos General Hospital, Basic Health Units, Municipal Health Surveillance Department (FVS), Municipal Education Department, Barcelos City Hall, Rio Negro FM radio channel and Pousada Água Negra.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Medina-Rincón, G.J.; Gallo-Bernal, S.; Jiménez, P.A.; Cruz-Saavedra, L.; Ramírez, J.D.; Rodríguez, M.J.; Medina-Mur, R.; Díaz-Nassif, G.; Valderrama-Achury, M.D.; Medina, H.M. Molecular and Clinical Aspects of Chronic Manifestations in Chagas Disease: A State-of-the-Art Review. Pathogens 2021, 10, 1493. [Google Scholar] [CrossRef] [PubMed]
- Massad, E. The Elimination of Chagas’ Disease from Brazil. Epidemiol Infect 2008, 136, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.L.; Silva, T.P. [Transmission Elimination of Chagas’ Disease by Triatoma Infestans in Brazil: An Historical Fact]. 2006.
- Coura, J.R. The Main Sceneries of Chagas Disease Transmission. The Vectors, Blood and Oral Transmissions--a Comprehensive Review. Mem Inst Oswaldo Cruz 2015, 110, 277–282. [Google Scholar] [CrossRef]
- Amanda Coutinho de Souza; José Rodrigues Coura; Catarina Macedo Lopes; Angela Cristina Verissimo Junqueira Eratyrus Mucronatus Stål, 1859 and Panstrongylus Rufotuberculatus (Champion, 1899) (Hemiptera, Reduviidae, Triatominae): First Records in a Riverside Community of Rio Negro, Amazonas State, Brazil. Check List 2021, 17, 905–909. [CrossRef]
- Coura, J.R.; Viñas, P.A.; Junqueira, A.C. Ecoepidemiology, Short History and Control of Chagas Disease in the Endemic Countries and the New Challenge for Non-Endemic Countries. Mem Inst Oswaldo Cruz 2014, 109, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R.; Vinas, P.A. Chagas Disease: A New Worldwide Challenge. Nature 2010, 465, S6–7. [Google Scholar] [CrossRef]
- 8Coura, J.R.; Dias, J.C. Epidemiology, Control and Surveillance of Chagas Disease: 100 Years after Its Discovery. Mem Inst Oswaldo Cruz 2009, 104 Suppl 1, 31–40. [Google Scholar] [CrossRef]
- WHO Chagas Disease (American Trypanosomiasis). 2023.
- Rassi, A.; Marcondes de Rezende, J. American Trypanosomiasis (Chagas Disease). Infect Dis Clin North Am 2012, 26, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.C.; Ramos, A.N.; Gontijo, E.D.; Luquetti, A.; Shikanai-Yasuda, M.A.; Coura, J.R.; Torres, R.M.; Melo, J.R.; Almeida, E.A.; Oliveira, W.; et al. [Brazilian Consensus on Chagas Disease, 2015]. Epidemiol Serv Saude 2016, 25, 7–86. [Google Scholar] [CrossRef]
- Benziger, C.P.; do Carmo, G.A.L.; Ribeiro, A.L.P. Chagas Cardiomyopathy: Clinical Presentation and Management in the Americas. Cardiol Clin 2017, 35, 31–47. [Google Scholar] [CrossRef]
- Nunes, M.C.P.; Beaton, A.; Acquatella, H.; Bern, C.; Bolger, A.F.; Echeverría, L.E.; Dutra, W.O.; Gascon, J.; Morillo, C.A.; Oliveira-Filho, J.; et al. Chagas Cardiomyopathy: An Update of Current Clinical Knowledge and Management: A Scientific Statement from the American Heart Association. Circulation 2018, 138, e169–e209. [Google Scholar] [CrossRef]
- Sabino, E.C.; Nunes, M.C.P.; Blum, J.; Molina, I.; Ribeiro, A.L.P. Cardiac Involvement in Chagas Disease and African Trypanosomiasis. Nat Rev Cardiol 2024. [Google Scholar] [CrossRef]
- Baldoni, N.R.; de Oliveira-da Silva, L.C.; Gonçalves, A.C.O.; Quintino, N.D.; Ferreira, A.M.; Bierrenbach, A.L.; Padilha da Silva, J.L.; Pereira Nunes, M.C.; Ribeiro, A.L.P.; Oliveira, C.D.L.; et al. Gastrointestinal Manifestations of Chagas Disease: A Systematic Review with Meta-Analysis. Am J Trop Med Hyg 2024, 110, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Dantas, R.O. Management of Esophageal Dysphagia in Chagas Disease. Dysphagia 2021, 36, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Camargo, C.L.; Albajar-Viñas, P.; Wilkins, P.P.; Nieto, J.; Leiby, D.A.; Paris, L.; Scollo, K.; Flórez, C.; Guzmán-Bracho, C.; Luquetti, A.O.; et al. Comparative Evaluation of 11 Commercialized Rapid Diagnostic Tests for Detecting Trypanosoma Cruzi Antibodies in Serum Banks in Areas of Endemicity and Nonendemicity. J Clin Microbiol 2014, 52, 2506–2512. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Alquezar, A.; Junqueira, A.C.V.; Durans, A.M.; Guimarães, A.V.; Corrêa, J.A.; Borges-Pereira, J.; Zauza, P.L.; Cabello, P.H.; Albajar-Vinãs, P.; Provance, Jr., DW; et al. Geographical Origin of Chronic Chagas Disease Patients in Brazil Impacts the Performance of Commercial Tests for Anti-T. Cruzi IgG. Mem Inst Oswaldo Cruz 2021, 116. [Google Scholar]
- Durans, A.M.; Napoleão-Pêgo, P.; Reis, F.C. G.; Dias, E.R.; Machado, L.E.S.F.; Lechuga, G.C.; Junqueira, A.C.V.; De-Simone, S.G.; Provance, Jr., D.W. Chagas Disease Diagnosis with Trypanosoma Cruzi Exclusive Epitopes in GFP. Vaccines (Basel) 2024.
- Ministry of Health PORTARIA No 57. 2018.
- Edwards, M.S.; Montgomery, S.P. Congenital Chagas Disease: Progress toward Implementation of Pregnancy-Based Screening. Curr Opin Infect Dis 2021, 34, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Close, D.W.; Paul, C.D.; Langan, P.S.; Wilce, M.C.; Traore, D.A.; Halfmann, R.; Rocha, R.C.; Waldo, G.S.; Payne, R.J.; Rucker, J.B.; et al. Thermal Green Protein, an Extremely Stable, Nonaggregating Fluorescent Protein Created by Structure-Guided Surface Engineering. Proteins 2015, 83, 1225–1237. [Google Scholar] [CrossRef]
- World Health Organization WHO Expert Committee on Biological Standardization. World Health Organ Tech Rep Ser 2011, 1–244.
- Coura, J.R.; Junqueira, A.C. Ecological Diversity of Trypanosoma Cruzi Transmission in the Amazon Basin. The Main Scenaries in the Brazilian Amazon. Acta Trop 2015, 151, 51–57. [Google Scholar] [CrossRef]
- A. M. Saravia, S.; F. Montero, I.; M. Linhares, B.; A. Santos, R.; A. F. Marcia, J. Mineralogical Composition and Bioactive Molecules in the Pulp and Seed of Patauá (Oenocarpus Bataua Mart.): A Palm from the Amazon. International Journal of Plant & Soil Science 2020, 1–7. [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal Biochem 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. I-TASSER Server for Protein 3D Structure Prediction. BMC Bioinformatics 2008, 9, 40. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A Unified Platform for Automated Protein Structure and Function Prediction. Nat Protoc 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Thomas, M.C.; Longobardo, M.V.; Carmelo, E.; Marañón, C.; Planelles, L.; Patarroyo, M.E.; Alonso, C.; López, M.C. Mapping of the Antigenic Determinants of the T. Cruzi Kinetoplastid Membrane Protein-11. Identification of a Linear Epitope Specifically Recognized by Human Chagasic Sera. Clin Exp Immunol 2001, 123, 465–471. [Google Scholar] [CrossRef]
- Buscaglia, C.A.; Alfonso, J.; Campetella, O.; Frasch, A.C. Tandem Amino Acid Repeats from Trypanosoma Cruzi Shed Antigens Increase the Half-Life of Proteins in Blood. Blood 1999, 93, 2025–2032. [Google Scholar] [PubMed]
- bañez, C.F.; Affranchino, J.L.; Macina, R.A.; Reyes, M.B.; Leguizamon, S.; Camargo, M.E.; Aslund, L.; Pettersson, U.; Frasch, A.C. Multiple Trypanosoma Cruzi Antigens Containing Tandemly Repeated Amino Acid Sequence Motifs. Mol Biochem Parasitol 1988, 30, 27–33. [Google Scholar] [CrossRef]
- Peralta, J.M.; Teixeira, M.G.; Shreffler, W.G.; Pereira, J.B.; Burns, J.M.; Sleath, P.R.; Reed, S.G. Serodiagnosis of Chagas’ Disease by Enzyme-Linked Immunosorbent Assay Using Two Synthetic Peptides as Antigens. J Clin Microbiol 1994, 32, 971–974. [Google Scholar] [CrossRef]
- Thomas, M.C.; Fernández-Villegas, A.; Carrilero, B.; Marañón, C.; Saura, D.; Noya, O.; Segovia, M.; Alarcón de Noya, B.; Alonso, C.; López, M.C. Characterization of an Immunodominant Antigenic Epitope from Trypanosoma Cruzi as a Biomarker of Chronic Chagas’ Disease Pathology. Clin Vaccine Immunol 2012, 19, 167–173. [Google Scholar] [CrossRef]
- Burns, J.M.; Shreffler, W.G.; Rosman, D.E.; Sleath, P.R.; March, C.J.; Reed, S.G. Identification and Synthesis of a Major Conserved Antigenic Epitope of Trypanosoma Cruzi. Proc Natl Acad Sci U S A 1992, 89, 1239–1243. [Google Scholar] [CrossRef] [PubMed]
- Houghton, R.L.; Benson, D.R.; Reynolds, L.D.; McNeill, P.D.; Sleath, P.R.; Lodes, M.J.; Skeiky, Y.A.; Leiby, D.A.; Badaro, R.; Reed, S.G. A Multi-Epitope Synthetic Peptide and Recombinant Protein for the Detection of Antibodies to Trypanosoma Cruzi in Radioimmunoprecipitation-Confirmed and Consensus-Positive Sera. J Infect Dis 1999, 179, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- 3Balouz, V.; Cámara, M.L.; Cánepa, G.E.; Carmona, S.J.; Volcovich, R.; Gonzalez, N.; Altcheh, J.; Agüero, F.; Buscaglia, C.A. Mapping Antigenic Motifs in the Trypomastigote Small Surface Antigen from Trypanosoma Cruzi. Clin Vaccine Immunol 2015, 22, 304–312. [Google Scholar] [CrossRef]
- Gruber, A.; Zingales, B. Trypanosoma Cruzi: Characterization of Two Recombinant Antigens with Potential Application in the Diagnosis of Chagas’ Disease. Exp Parasitol 1993, 76, 1–12. [Google Scholar] [CrossRef]
- Bottino, C.; Gomes, L.P.; Coura, J.B.; Provance, D.W.J.; De-Simone, S.G. Chagas Disease-Specific Antigens: Characterization of Epitopes in CRA/FRA by Synthetic Peptide Mapping and Evaluation by ELISA-Peptide Assay. BMC Infect Dis 2013, 13, 568. [Google Scholar] [CrossRef]
- Studier, F.W.; Moffatt, B.A. Use of Bacteriophage T7 RNA Polymerase to Direct Selective High-Level Expression of Cloned Genes. J Mol Biol 1986, 189, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C. Nova Tripanozomiaze Humana. Mem Inst Oswaldo Cruz 1909, 1, 159–219. [Google Scholar]
- Aufderheide, A.C.; Salo, W.; Madden, M.; Streitz, J.; Buikstra, J.; Guhl, F.; Arriaza, B.; Renier, C.; Wittmers, L.E.; Fornaciari, G.; et al. A 9,000-Year Record of Chagas’ Disease. Proc Natl Acad Sci U S A 2004, 101, 2034–2039. [Google Scholar] [CrossRef]
- Guhl, F.; Jaramillo, C.; Yockteng, R.; Vallejo, G.A.; Cárdenas-Arroyo, F. Trypanosoma Cruzi DNA in Human Mummies. 1997.
- Coura, J.R. The Discovery of Chagas Disease (1908-1909): Great Successes and Certain Misunderstandings and Challenges. Rev Soc Bras Med Trop 2013, 46, 389–390. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Castro, O.; Moo-Llanes, D.A.; Ramsey, J.M. Impact of Climate Change on Vector Transmission of Trypanosoma Cruzi (Chagas, 1909) in North America. Med Vet Entomol 2018, 32, 84–101. [Google Scholar] [CrossRef] [PubMed]
- WHO Ending the Neglect to Attain the Sustainable Development Goals: A Road Map for Neglected Tropical Diseases 2021–2030. 2020.
- Porrás, A.I.; Yadon, Z.E.; Altcheh, J.; Britto, C.; Chaves, G.C.; Flevaud, L.; Martins-Filho, O.A.; Ribeiro, I.; Schijman, A.G.; Shikanai-Yasuda, M.A.; et al. Target Product Profile (TPP) for Chagas Disease Point-of-Care Diagnosis and Assessment of Response to Treatment. PLoS Negl Trop Dis 2015, 9, e0003697. [Google Scholar] [CrossRef]
- Caballero, Z.C.; Sousa, O.E.; Marques, W.P.; Saez-Alquezar, A.; Umezawa, E.S. Evaluation of Serological Tests to Identify Trypanosoma Cruzi Infection in Humans and Determine Cross-Reactivity with Trypanosoma Rangeli and Leishmania Spp. Clin Vaccine Immunol 2007, 14, 1045–1049. [Google Scholar] [CrossRef]
- Daltro, R.T.; Leony, L.M.; Freitas, N.E.M.; Silva, Â.A.O.; Santos, E.F.; Del-Rei, R.P.; Brito, M.E.F.; Brandão-Filho, S.P.; Gomes, Y.M.; Silva, M.S.; et al. Cross-Reactivity Using Chimeric Trypanosoma Cruzi Antigens: Diagnostic Performance in Settings Where Chagas Disease and American Cutaneous or Visceral Leishmaniasis Are Coendemic. J Clin Microbiol 2019, 57, e00762-19. [Google Scholar] [CrossRef]
- Otani, M.; Hockley, J.; Bracho, C.G.; Rijpkema, S.; Luguetti, A.O.; Duncan, R.; Rigsby, P.; Albajar-Viñas, P.; Padilla, A. Evaluation of Two International Reference Standards for Antibodies to Trypanosoma Cruzi in a WHO Collaborative Study. WHO Technical Report 2011. [Google Scholar]
- Saez-Alquezar, A.; Junqueira, A.C.; Durans, A.M.; Guimarães, A.V.; Corrêa, J.A.; Provance, J., DW; Cabello, P.H.; Coura, J.R.; Viñas, P.A. Application of WHO International Biological Standards to Evaluate Commercial Serological Tests for Chronic Chagas Disease. Mem Inst Oswaldo Cruz 2020, 115. [Google Scholar]
- Ricci, A.D.; Bracco, L.; Salas-Sarduy, E.; Ramsey, J.M.; Nolan, M.S.; Lynn, M.K.; Altcheh, J.; Ballering, G.E.; Torrico, F.; Kesper, N.; et al. The Trypanosoma Cruzi Antigen and Epitope Atlas: Antibody Specificities in Chagas Disease Patients across the Americas. Nat Commun 2023, 14, 1850. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R.; Junqueira, A.C.; Ferreira, J.M.B. Surveillance of Seroepidemiology and Morbidity of Chagas Disease in the Negro River, Brazilian Amazon. Mem Inst Oswaldo Cruz 2018, 113, 17–23. [Google Scholar] [CrossRef] [PubMed]
- William Provance, D.; da Matta Durans, A.; Lechuga, G.C.; da Rocha Dias, E.; Morel, C.M.; De Simone, S.G. Surpassing the Natural Limits of Serological Diagnostic Tests. hLife 2024, 0. [Google Scholar] [CrossRef]
- Ramos, L.G.; de Souza, K.R.; Júnior, P.A.S.; Câmara, C.C.; Castelo-Branco, F.S.; Boechat, N.; Carvalho, S.A. Tackling the Challenges of Human Chagas Disease: A Comprehensive Review of Treatment Strategies in the Chronic Phase and Emerging Therapeutic Approaches. Acta Trop 2024, 256, 107264. [Google Scholar] [CrossRef]
- Alonso-Padilla, J.; López, M.C.; Esteva, M.; Zrein, M.; Casellas, A.; Gómez, I.; Granjon, E.; Méndez, S.; Benítez, C.; Ruiz, A.M.; et al. Serological Reactivity against T. Cruzi-Derived Antigens: Evaluation of Their Suitability for the Assessment of Response to Treatment in Chronic Chagas Disease. Acta Trop 2021, 221, 105990. [Google Scholar] [CrossRef]
- Alonso-Padilla, J.; Abril, M.; Alarcón de Noya, B.; Almeida, I.C.; Angheben, A.; Araujo Jorge, T.; Chatelain, E.; Esteva, M.; Gascón, J.; Grijalva, M.J.; et al. Target Product Profile for a Test for the Early Assessment of Treatment Efficacy in Chagas Disease Patients: An Expert Consensus. PLoS Negl Trop Dis 2020, 14, e0008035. [Google Scholar] [CrossRef]
Figure 1.
Computer model of DxCruziV3. Views at 0°, 120°, and 240°of the predicted structure. The retained beta barrel is displayed with a ribbon for the primary sequence and a transparent surface (green). Epitopes are shown with a solid surface in the colors listed in Table 1. Projections obtained with PyMol software (V3.0, Schrödinger).
Figure 1.
Computer model of DxCruziV3. Views at 0°, 120°, and 240°of the predicted structure. The retained beta barrel is displayed with a ribbon for the primary sequence and a transparent surface (green). Epitopes are shown with a solid surface in the colors listed in Table 1. Projections obtained with PyMol software (V3.0, Schrödinger).
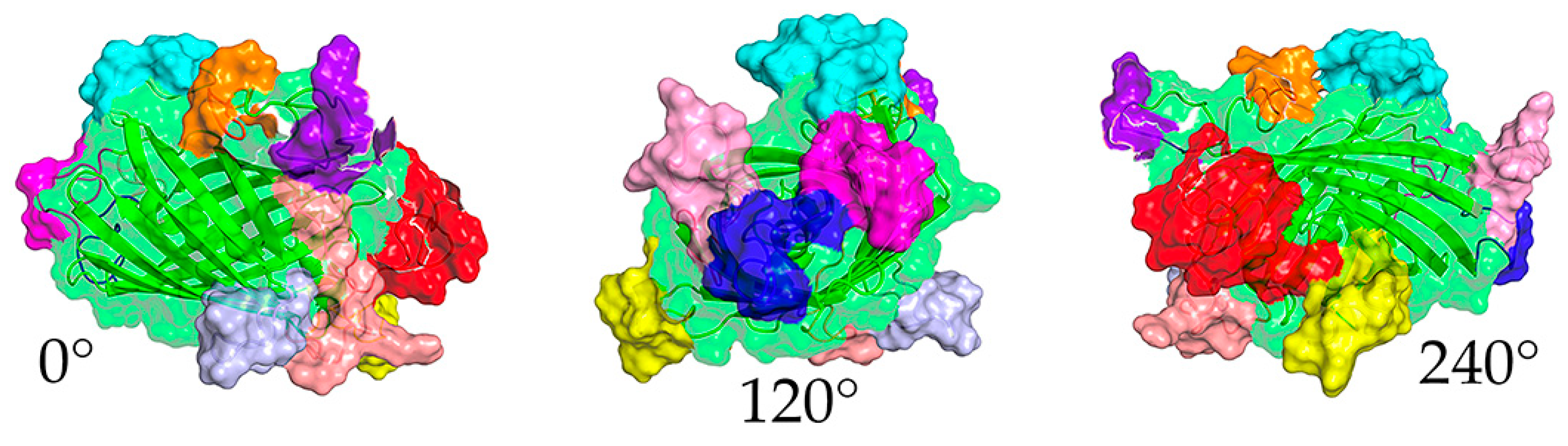
Figure 2.
Performance of DxCruziV3 in an ELISA format. (a) Performance of an in-house ELISA against a panel of sera reactive (T. cruzi infected; n = 75) or non-reactive (n=104) from patients diagnosed by LACENs in the Brazilian states of Ceará, Sergipe, Paraíba, and Maranhão. (b) Evaluation for cross-reactivity against serum from patients diagnosed with cutaneous Leishmaniosis, visceral Leishmaniosis, and dengue as well as non-specific reactivity with samples from healthy blood donors (HEMORIO).
Figure 2.
Performance of DxCruziV3 in an ELISA format. (a) Performance of an in-house ELISA against a panel of sera reactive (T. cruzi infected; n = 75) or non-reactive (n=104) from patients diagnosed by LACENs in the Brazilian states of Ceará, Sergipe, Paraíba, and Maranhão. (b) Evaluation for cross-reactivity against serum from patients diagnosed with cutaneous Leishmaniosis, visceral Leishmaniosis, and dengue as well as non-specific reactivity with samples from healthy blood donors (HEMORIO).
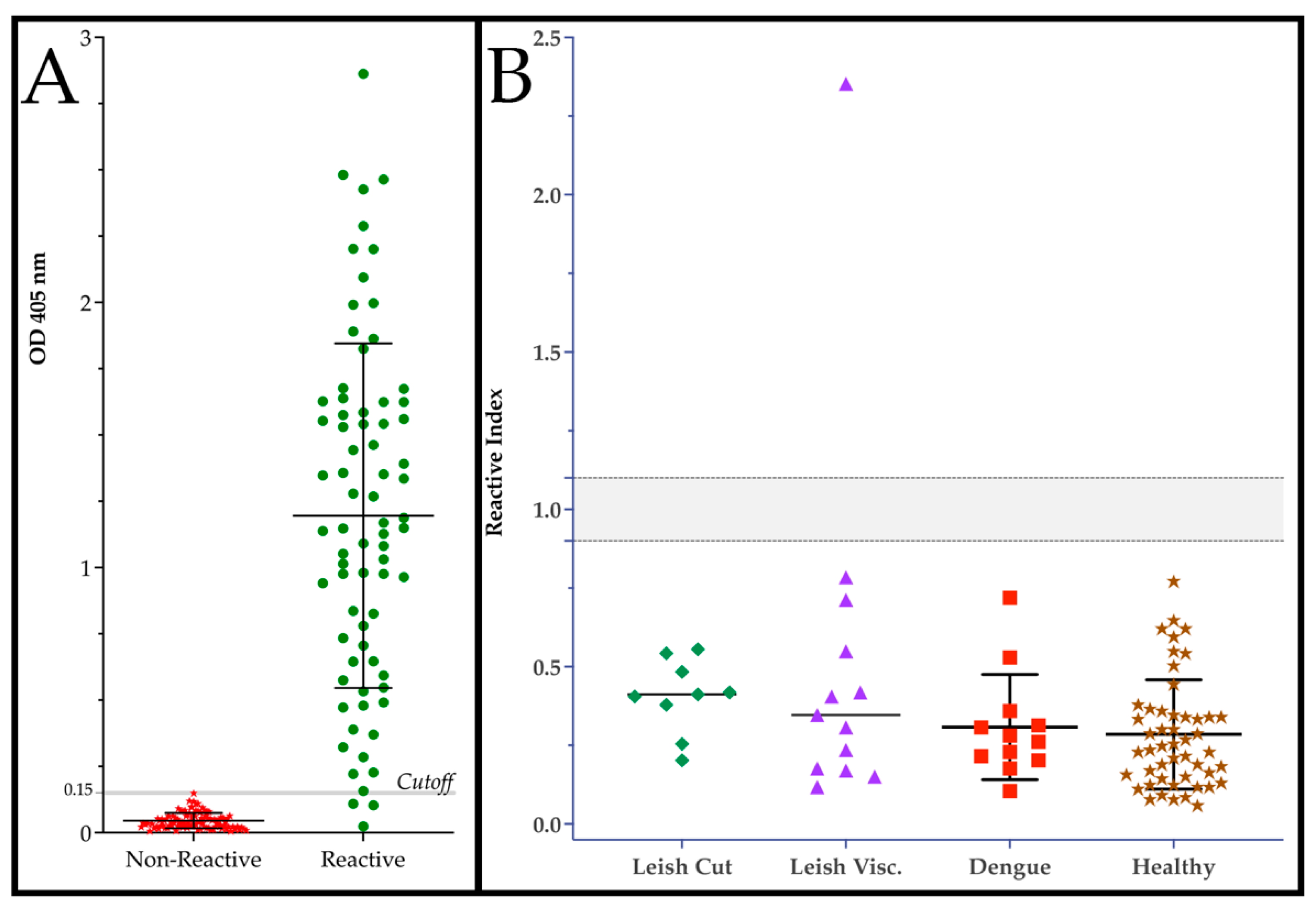
Figure 3.
Analytical sensitivity and influence of geographical origin of patient sample on ELISA performance. (a) In-house ELISAs generated with 200 ng of IB-V3 were exposed to a two-fold dilution series (1:2 to 1:256) of the two WHO International Biological Standards NIBSC 09/188 (TcI) and NIBSC 09/186 (TcII). Bound human antibodies were detected with a horseradish peroxidase labeled secondary. (b) Reactivity index calculated for patient samples originating from different states of Brazil displaying the P value from an ANOVA analysis with a Kruskal-Wallis test. AM-Amazonas; MA-Maranhão; CE-Ceará; PB-Paraíba; SE-Sergipe; Neg.-Negative controls. *WHO recommended minimum dilution.
Figure 3.
Analytical sensitivity and influence of geographical origin of patient sample on ELISA performance. (a) In-house ELISAs generated with 200 ng of IB-V3 were exposed to a two-fold dilution series (1:2 to 1:256) of the two WHO International Biological Standards NIBSC 09/188 (TcI) and NIBSC 09/186 (TcII). Bound human antibodies were detected with a horseradish peroxidase labeled secondary. (b) Reactivity index calculated for patient samples originating from different states of Brazil displaying the P value from an ANOVA analysis with a Kruskal-Wallis test. AM-Amazonas; MA-Maranhão; CE-Ceará; PB-Paraíba; SE-Sergipe; Neg.-Negative controls. *WHO recommended minimum dilution.
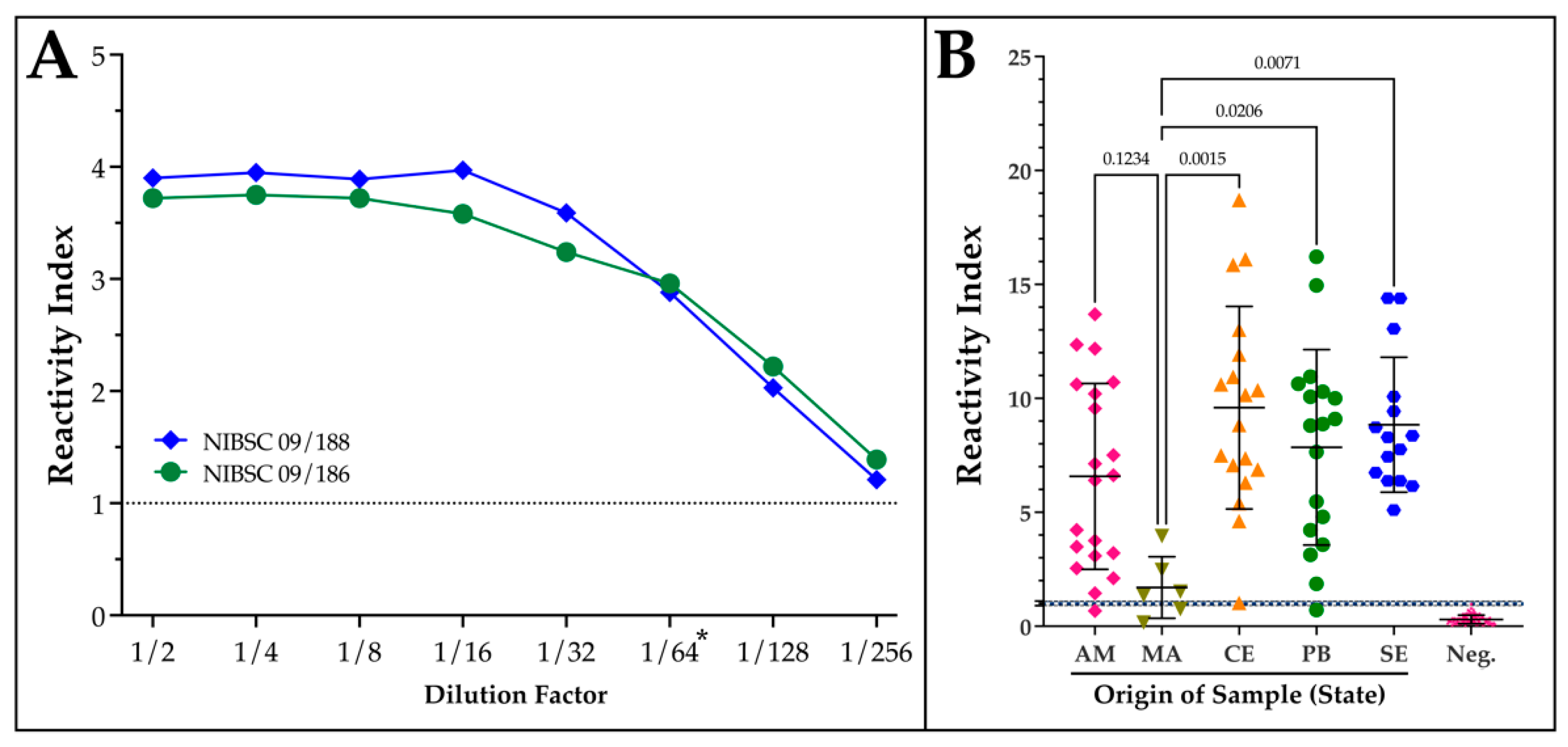
Figure 4.
Performance of sV3-LFA in relation to V3ib-ELISA. Serum samples collected in the states of Minas Gerais (MG, BR) in 2022 and Piauí (PI, BR) in 1999 were applied (5 µl) to the sV3-LFA and defined as reactive (blue) or non-reactive (red). The samples were also analyzed by the V3ib, in-house ELISA (1:200 dilution) and their reactivity index calculated.
Figure 4.
Performance of sV3-LFA in relation to V3ib-ELISA. Serum samples collected in the states of Minas Gerais (MG, BR) in 2022 and Piauí (PI, BR) in 1999 were applied (5 µl) to the sV3-LFA and defined as reactive (blue) or non-reactive (red). The samples were also analyzed by the V3ib, in-house ELISA (1:200 dilution) and their reactivity index calculated.
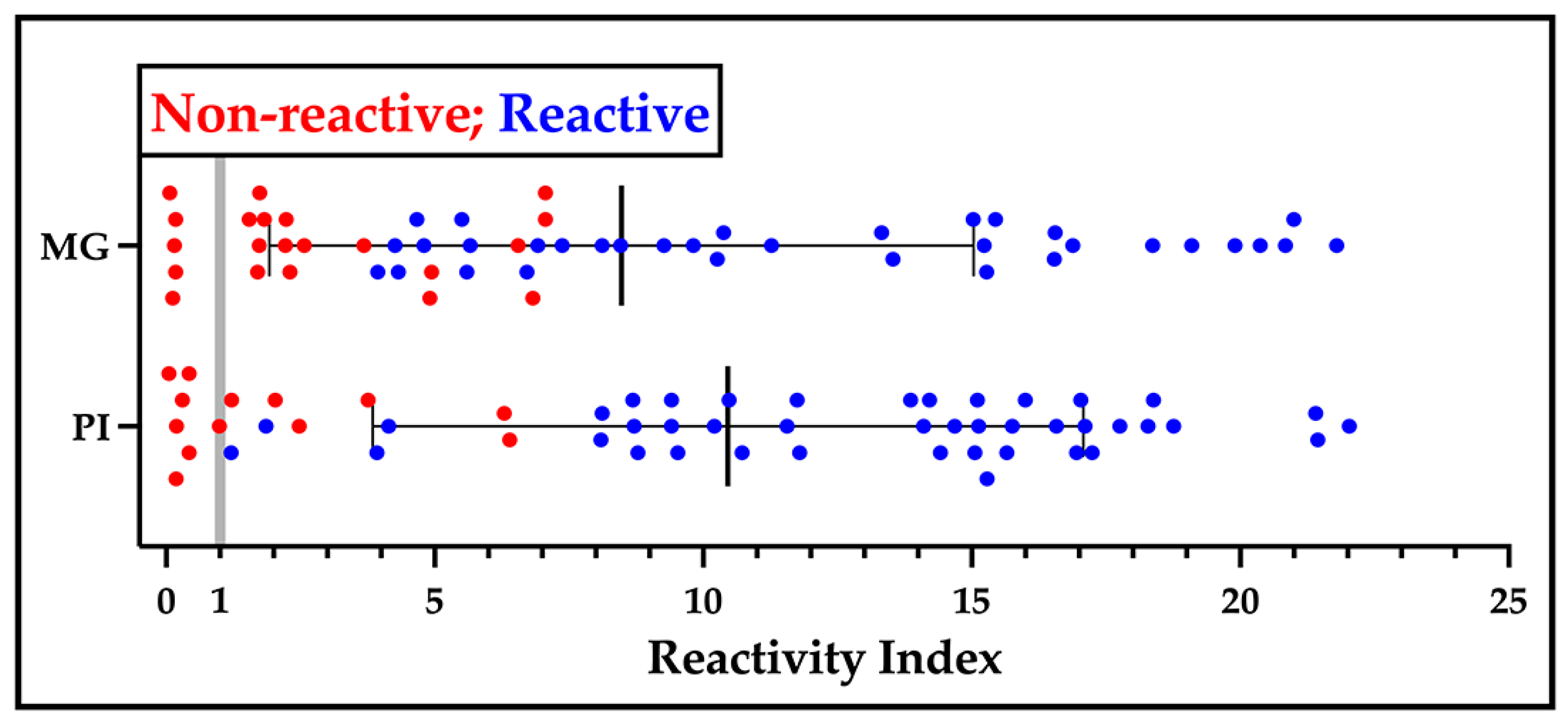

Table 1.
Epitopes contained in DxCruziV3 and their position within TGP.
| Epitope | Sequence | Insertion Site1 | Color2 | Protein Origin | Reference |
|---|---|---|---|---|---|
| 1 | KFAELLEQQKNAQFPGK | N-term | Red | KMP11 | [29] |
| 2 | DSSAHSTPSTPA | 50 | Blue | SAPA | [30] |
| 3 | GDKPSPFGQAAAADK | 114 | Yellow | PEP-2 | [31,32] |
| 4 | FGQAAAGDKPS | 127 | Magenta | TcCA-2 | [33] |
| 5 | AEPKPAEPKS | 153 | Lt. Blue | TcD-2 | [34,35] |
| 6 | TSSTPPSGTENKPAT | 167 | Cyan | TSSA | [36] |
| 7 | GTSEEGSRGGSSMPS | 183 | Salmon | TcLo1.2 | [35] |
| 8 | SPFGQAAAGDK | 244 | Pink | B13 | [32,37] |
| 9 | KAAIAPA | C-term | Purple | TcE | [35] |
| 10 | KQRAAETK | C-term | Orange | CRA | [38] |
Table 2.
Parameters of performance for V3ib ELISA.
| Sensitivity | Specificity | Likelihood Ratio | Pos. Likelihood Ratio | Neg. Likelihood Ratio |
|---|---|---|---|---|
| 96.0% | 100% | 98.6 | 98.8% | 95.3% |
Table 3.
Results obtained by different assays. 1- In cases of divergence, additional assays were performed to determine a final call. 2- Whole blood was applied. 3- The number of tests in the first lot were insufficient to repeat all samples. 4- Second lot of sV3-LFA produced by Bio-Manguinhos.
Table 3.
Results obtained by different assays. 1- In cases of divergence, additional assays were performed to determine a final call. 2- Whole blood was applied. 3- The number of tests in the first lot were insufficient to repeat all samples. 4- Second lot of sV3-LFA produced by Bio-Manguinhos.
| Assay | Repititions1 | Volume | Reactive | Non-reactive |
|---|---|---|---|---|
| V3ib-LFA Field2 | 1 | 10 µl | 43 | 124 |
| sV3-LFA Field2 | 1 | 10 µl | 43 | 124 |
| V3ib-LFA Lab3 | 0-1 | 5 µl | 45 | 25 |
| sV3-LFA Lab (Lot#1)3 | 0-1 | 5 µl | 44 | 123 |
| sV3-LFA Lab (Lot #2)4 | 1 | 5 µl | 36 | 131 |
| V3ib ELISA (in-house) | 3-8 | 0.5-2 µl | 39 | 128 |
| ELISA Wiener | 2-3 | 10 µl | 32 | 135 |
| ELISA Bioclin | 2 | 10 µl | 61 | 106 |
| Immunofluorescence Indirect | 1-2 | 5 µl | 48 | 119 |
Table 4.
Performance of diagnostic assays for reactivity to chronic T. cruzi infections.
| Assay | Non-Divergent (n=150) | Final Call (n=167) | ||
|---|---|---|---|---|
| Sensitivity | Specificity | Sensitivity | Specificity | |
| Field V3ib LFA | 100% | 100% | 95.2% | 97.6% |
| Field sV3 LFA | 100% | 100% | 90.5% | 96.0% |
| V3ib-ELISA | 87.5% | 98.3% | 87.5% | 97.6% |
| ELISA Wiener | 71.9% | 97.5% | 64.3% | 96.0% |
| ELISA Bioclin | 90.6% | 79.7% | 83.3% | 79.2% |
| Immunofluorescence | 93.8% | 88.1% | 76.2% | 87.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



