
Submitted:
19 August 2024
Posted:
19 August 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The current globally dominant SARS-CoV-2 variants are showing immune escape and reduced susceptibility to antiviral drugs. Therefore, agencies responsible for drug evaluation and regulation such as the FDA and EMA are revising their emergency authorization use of several COVID-19 neutralizing antibodies. These MAbs proved to be unlikely effective against new variants especially Omicron descendants and several pharmaceutical companies are pursuing the development of more potent neutralizing antibodies. To address this issue, we used In Silico method we previously developed to assess 10 anti-SARS-CoV-2 antibodies propensity to neutralize the new Omicron’s subvariants EG.5, BA.2.86 and JN.1, based on comparative binding affinity of 3D generated models and previous experimental and clinical observations. Nine of these MAbs were once granted emergency use authorization, and one is currently under clinical investigation. The results showed that one antibody showed a marked increase of the binding energy for EG.5 compared to two antibodies that showed a significant increase with Pirola (BA.2.86) and JN.1. This data indicates that the new SARS-CoV-2 variant escapes neutralization of most of the available therapeutic NAbs. Furthermore, the data showed new potential therapeutic MAbs combination that could be effective for the treatment countermeasure of the new Omicron’s descendants or potential novel variants.
Keywords:
In Silico
; Monoclonal
; Antibodies
; COVID-19
; EG.5
; BA.2.86
; JN.1
; Neutralization power
; Anti-SARS-CoV-2
; Variants
1. Introduction
Monoclonal antibodies (mAbs) are instrumental in combating viral infections [1,2]. Indeed, specific mAbs endowed with high affinity can efficiently neutralize and clear viruses through their effector functions [3,4]. In addition, current structural biology approaches combined with molecular engineering techniques allow the development of mAbs with high neutralizing power and the production in vitro of recombinant mAbs with improved effector functions and pharmacokinetics. Following the emergence of SARS-CoV-2 virus causing the COVID-19 pandemic in late 2019, several risk reduction strategies have been applied to prevent, overcome and/or mitigate its health impact. These measures diverse from personal safety measures to avoid contracting the virus to immunity boost via vaccination and treatment using selected neutralizing antibodies (NAbs). Anti-SARS-CoV-2 NAbs have first proven to be efficient in the clinical treatment of SARS-CoV-2 infection particularly the mild and moderate forms of the disease. Indeed, as of early 2021, nearly 18 months into pandemic, the U.S. Food and Drug Administration (FDA) and the European Medicine Agency (EMA) have granted emergency use authorizations (EUA) for several NAbs for the treatment of moderate to severe symptoms and as preventive protection from COVID-19. However, as new variants of SARS-CoV-2 emerged and due to increased frequency of resistant variants especially Omicron and its’ subvariants, the American National Institute of Health (NIH) COVID-19 Treatment Guidelines Panel started recommending against the use of anti-SARS-CoV-2 NAbs for the treatment or prevention of COVID-19. Based on marked reduction of in vitro susceptibility [5,6,7] and neutralizing activity studies [8,9,10] the EUAs have been revoked for many NAbs. Meanwhile, several pharmaceutical companies and organizations continue to invest in developing new Nabs. Several Nabs are now in early clinical development for example, REGN15160 and REGN14256 that are investigated by Regeneron, CPT63, and CTP59 developed by Celltrion and JS026 by Shangai Junshi, which was previously allied with Eli Lilly for the development of Etesevimab [11].Currently, several Omicron subvariants are monitored closely. Among which are EG.5 (Eris), BA.2.86 (Pirola) and JN.1. As of December 2023, theses variants are classified by the World Health Organization (WHO) as “variant of interest” (VOI) [12]. EG.5 is a descendant of Omicron XBB.1.9.2, it was first reported on February 2023 and it was related to increased prevalence with no solid evidence of disease severity [13]. BA.2.86 is a descendant of Omicron BA.2. and it was first reported on July 2023 in Denmark [14]. Up to date little is known about the severity or transmissibility of BA.2.86 variant [15]. According to Stanford University SARS-CoV-2 Variants database [6], EG.5 showed three different additional mutations to its parent subvariant (XBB), G252V, F456L and F486P. One particular mutation appears to be especially important, F456L, it is located at the interface between the cell Angiotensin Converting Enzyme-2 and the virus Receptor Binding Domain (ACE2-RBD) within the epitope of several Class I NAbs [16]. Another mutation F490S, is predicted to be resistant to class II antibody (Bamlanivimab). Recent study showed that this mutation is resistant to neutralization and causes antibody evasion [17]. On the other hand, BA.2.86 showed 31 differences in comparison to its parent subvariant (BA.2). It has parent mutations Q493R and R346T besides its own unique mutation L452W and all are linked to antibody evasion. In addition, both variants share four mutations (F486P, E484A/K, K417N, and N460K) that are also predicted to evade neutralization by class I and II NAbs [17,18]. Moreover, studies using BA.2.86 spike-pseudotyped virus model showed that this variant is resistant to several NAbs including Tixagevimab, Cilgavimab (Evusheld), Bebtelovimab (LY-CoV1404) and Sotrovimab (S309) [19,20,21]. The latest variant JN.1 which is a descendant of BA.2.86 was first collected on August 2023 and generated global attention as it’s represents 27.1% of the globally available sequences of the epidemiological week 48 and it was reported by 41 countries including France, USA, Singapore, Canada, United Kingdom and Sweden [22]. JN.1 variant has just one additional mutation (L455S) on the receptor binding domain compared to its precursor BA.2.86 [23]. This critical mutation added a new advantage to this new variant making it more transmissible with enhance immune evasion ability [24]. Overall, all three variants EG.5 and BA.2.86 and JN.1 contains three or more of especially important mutations previously reported as NAbs resistant mutations, including residues R346 and P337, G339, S371, N440, F486, V445, and G446 in addition to the flip mutation F456L of EG.5 [13,19,25,26,27].
In this study, we applied a computational method we previously described to evaluate SARS-CoV-2 antibodies neutralization power and to assess the effectiveness of the available SARS-CoV-2 neutralizing antibodies on the newly emerged variants Eris, Pirola and JN.1. The method is a simple computational approach that predicts the affinity of existing anti SARS-CoV-2 therapeutic antibody and evaluate their potential use with new emergent variants [28]. Therefore, we modeled nine different neutralizing antibodies previously approved by FDA and EMA for the treatment of mild to moderate COVID-19 symptoms and measured their binding affinities to the new variants in comparison to the original Wuhan strain for which they were granted emergency authorization use. These NAbs include Sotrovimab (S309) [29], Bamlanivimab (LY-CoV555) [30], Etesevimab (LY-CoV016, CB6, JS016) [31,32], Evusheld (Tixagevimab -AZD8895 and Cilgavimab-AZD1061) [33], Regdanvimab (CT-P59) [34], Casirivimab (REGN10933) [35], Imdevimab (REGN10987) [35] and Bebtelovimab (LY-CoV1404) [36]. Additionally, was also evaluated one anti-SARS-CoV-2 antibody (JS026) [37] that is currently in early clinical trials and shows excellent results with increased neutralizing efficacy in combination with Etesevimab [11,37]. The main modes of action for the these selected NAbs is to prevent virus entry to the host cell either by site competition or via steric hindrance [38]. Site competing NAbs binds to an epitope of the spike protein receptor binding domain which overlaps the ACE2 interacting site and effectively inhibits the interaction required for cell entry. These types of neutralizing antibodies bind the spike protein in its activated (open) state only where it has access to the specific ACE2 receptor epitope. On the other hand, neutralizing antibodies by steric hindrance binds distinct epitope and therefore can bind the spike protein in both states up and down.
2. Methods
2.1. Selection of the SARS-CoV-2 Neutralizing Antibodies
The selection of the antibodies we studied is based on their granting of the emergency use authorization (EUA). All the antibodies that were granted EUA by the FDA and/or EMA for the first SARS-CoV-2 strain (Wuhan) were retrieved, examined, and evaluated in addition to one that is not yet authorized but showed promising clinical results with resistant variants.
2.2. SARS-CoV-2 BA.2.86 (Pirola), JN.1 and EG.5 (Eris) Variants Sequence Retrieval, Modifications, and Modeling
The amino acid sequences of the extracellular domain of SARS-CoV-2 spike proteins were acquired from the National Center for Biological Information (NCBI) protein ID: YP_009724390.1 (amino acid 1-1261). SARS-CoV-2 variants-specific mutations were introduced to the collected sequence to generate the different variant sequences based on published mutations in the Stanford University SARS-CoV-2 Variants database (https://covdb.stanford.edu/variants/omicron_ba_1_3/) [39]. The spikes 3D monomeric structures were modeled in an open state form as described in our previous report [28]. using the SWISS-MODEL server - User Template Mode [40] (https://swissmodel.expasy.org/interactive#structure) and extracted cleaned open state monomer of Omicron’s template model (7XCO) [28].
2.3. NAb/SARS-CoV-2 RBD Reference Models’ Selection and Modification
Models representing the interaction of the selected neutralizing antibodies with receptor binding domain of SARS-CoV-2 (Wuhan variant) were extracted from RCSB Protein Data Bank (RCSB PDB) (https://www.rcsb.org). Table 1 lists the PDB IDs for the used 3D models representing each antibody and as some models show more than one NAb/RBD complex, used chain IDs for each model were indicated in the table. Each model was cleaned from any heteroatoms, modified so that only one unit of NAb/RBD is present, and only NAbs’ variable domain (Fv) is represented in the model. The modified models were used as reference models to generate complexes with BA.2.86, JN.1 and EG.5 variants by RBD replacement.
2.4. RBD/S309 Complexes Construction
RBDs of the BA.2.86, JN.1 and EG.5 SARS-CoV-2 variants were extracted from the generated models and complexes with the ten neutralizing antibodies were constructed by molecular replacement of the RBD domain. All constructed models were energy minimized using Swiss-pdb Viewer 4.1.0 (http://www.expasy.org/spdbv/) [41]. The energy minimization step was done in vacuo, without reaction field using GROMOS 43B1 force field [42].
2.5. Complex Binding Affinity Analysis
The stability and affinity were assisted based on thermodynamic measure of the formed complex energy, Gibbs free energy, (ΔG). This was performed using an antibody-antigen binding affinity online tool, CSM-AB (https://biosig.lab.uq.edu.au/csm_ab/prediction) [43]. Binding affinity percentage was calculated in reference to NAb/Wuhan complex’s binding affinity to which we attributed a 100% value. The interactions of some selected NAbs with the RBD of the new SARS-CoV-2 variants were analyzed based on polar and hydrophobic interaction using the LigPlot+ software [44].
2.6. RBD 3D Model Evaluation of SARS-CoV-2 BA.2.86 (Pirola), JN.1 and EG.5 (Eris) Variants
Generated 3D RBD models of SARS-CoV-2 BA.2.86 (Pirola), JN.1 and EG.5 (Eris) variants were evaluated by pairwise comparison alignment to the Wuhan RBD (PDB ID: 7R6W, resolution 1.83 Å, Chain R) as the target model. This was done using protein structure comparison service PDBeFold at European Bioinformatics Institute (http://www.ebi.ac.uk/msd-srv/ssm) [45]. Visualization and alignment of the resulted models was done using PyMol software [46].
3. Results
3.1. Antibodies Selection
Nine different SARS-CoV-2 neutralizing antibodies were selected based on the emergency use authorization granted by FDA and/or EMA. MAb Regdanvimab (CT-P59) was granted authorization in the European Union only and MAb Bebtelovimab (LY-CoV1404) was granted authorization by FDA but not EMA. The authorization was for the pre-exposure prophylaxis and/or treatment of COVID-19 symptoms caused by the original strain of SARS-CoV-2 (Wuhan). Currently, and due to the increase of resistant variants frequency, especially Omicron and its descendants, FDA revoked the EUA for all the neutralizing antibodies. However, EMA emergency use authorization is yet effective (Table 2). Meanwhile, several clinical trials are conducted to develop new antibodies that are capable of neutralizing the newly emerged subvariants. JS026 is one of the promising NAbs that showed to have increased neutralization effect against wild SARS-CoV-2 and its Alpha, Beta, Gamma, and Delta variants when combined with Etesevimab [37]. Therefore, although it is not an approved NAb yet, we add JS026 to the selected NAbs on this paper to analyze its neutralizing effect with the new variants BA.2.86, JN.1 and EG.5.
3.2. Models’ Generation
To analyze the binding affinity of the selected NAbs with the new variants BA.2.86 (Pirola), JN.1 and EG.5 (Eris), 30 models were generated representing the interaction between the NAb’s Fv domain and the RBD of SARS-CoV-2 newly emerged variants. In addition to 10 reference models, cleaned and energy minimized, representing the interaction of the selected NAbs with the original Wuhan strain RBD domain (Table 1). Each of the examined NAbs showed to have different binding site on the RBD domain. Moreover, as previously described [28], NAbs are categorized based on binding to a specific RBD epitope or based on whether it is competing with the angiotensin-converting enzyme 2 (ACE2). The selected NAbs were from class I, II and III as shown in Figure 1. Table 3 describes the previously reported neutralizing effect of the selected NAbs on SARS-CoV-2 and its variants.
3.3. Binding Affinity Analysis
The generated (RBD/NAb-Fv) interaction models were energy minimized and the binding energy of the 3D models were calculated by computational prediction of Gibbs free energy (ΔG). The percentage of the binding energy for each antibody was calculated in comparison to that of the same NAb with Wuhan strain (Figure 2). The results showed that the binding energy of most of the evaluated NAbs showed moderate to marked reduction with the newly emerged variants indicating that the new SARS-CoV-2 variant escapes neutralization of most of the available therapeutic antibodies. However, two antibody class I Evusheld (Cilgavimab-AZD1061) and class III Bebtelovimab (LY-CoV1404) retain nearly the same neutralization effect of Wuhan strain with minor increase of 1.1- and 1.2-fold respectively toward BA.286 variant and its 86 descendant JN.1. For EG.5 only class I Evusheld (Cilgavimab-AZD1061) antibody retain the same binding affinity of Wuhan strain (Figure 2) and Table A1.
Interestingly, one NAb JS026 that is currently under clinical trials, showed a significant increase in the binding affinity for all of the three new variants BA.2.86, JN.1 and EG.5 with 3-, 3.1- and 2.9-folds increase respectively. Additionally, NAbs Imdevimab (REGN10987) showed a 2.8-fold and 3-fold increase of affinity binding with BA.2.86 and JN.1 only. Although JS026 is to date under clinical evaluation, Imdevimab is still under emergency use authorization by EMA under the name Ronapreve and as a cocktail with Casirivimab for the treatment of COVID-19 symptoms in adults while its EUA was revoked by FDA on 2022 (Table 2).
3.4. Analysis of Imdevimab and JS026 Molecular Interactions with the RBD Domain of the New Variants EG.5 and BA.2.86
The new SARS-CoV-2 variants BA.2.86 and JN.1 that are a BA.2 descendants showed a significant increase in the binding affinity with Imdevimab and JS026 while EG.5 which is an XBB decedent showed an increase with JS026 only. Both antibodies are class III antibodies and share similar binding epitopes. Analyses of the interaction pattern of Imdevimad with BA.2.86 and JN.1 using LigPlot+ software, revealed one polar and 33 hydrophobic interaction in addition to one salt bridge. The Interaction involves the same residues of Wuhan in addition to four new interacting residues, Asp450, Gln498, and Pro499 (Table 4). However, interaction of Imdevimab with EG.5 includes 5 out of 6 residues with an additional four new ones with 1 polar, 28 hydrophobic and 1 salt bridge.
Conversely, JS026 binding affinity increases with all three variant BA.2.86, JN.1 and EG.5. Interaction analysis revealed that interacting residues for all variants are the same with fourteen residues as that of Wuhan except for Asn343 and an additional salt bridge for JN.1. Table 3 lists the interaction residues, number and type of interaction and mutated residues for each variant.
3.5. Variants’ 3D RBD Model Alignment and Evaluation
The alignment of the RBD of BA.2 descendant SARS-CoV-2 variants, BA.2.86, and JN.1 with the original Wuhan strain (PDB ID: 7R6W- chain R) showed an 86% sequence identity in 189 aligned amino acids and an RMSD of 0.86 and 0.85 respectively. EG.5, which is an XBB successor with a smaller number of mutations showed 88% sequence identity in 190 aligned amino acids and an RMSD of 0.83. Structural 3D alignment of theses variants displayed unmatched residues that interfere with the secondary structure spanning residues Tyr369, Asn370, Ser371 and Ala372 and involve a significant mutation Ser371Phe. This mutation causes an alteration of the loop as shown in Figure 3. Additionally, isolated mismatches including residue 386 and 387 for all the analyzed variants and residue 482 for EG.5 only were detected Table 5.
4. Discussion
Four years into pandemic and SARS-CoV-2 virus is still evolving. New variants are rising every day and so far, all lineages currently circulating are classified as Omicron variant sub lineages. As of December 2023, WHO listed five variants as variants of interest (VOIs); namely XBB.1.5, XBB.1.16, EG.5, BA.2.86 and JN.1 [12,53]. The most recent variants are EG.5 (Eris) BA.2.86 (Pirola) and JN.1. They were denoted as the most prevalent VOI as it has been reported by 39, 49 and 41 countries respectively with the EG.5 remaining to be the most reported VOI accounting for 36.3% of sequences shared on GISAID in epidemiological week 48 (27 November to 3 December 2023) [12].
Several recent studies [54,55,56,57] experimentally evaluated the immune evasion of neutralizing antibodies by the new evolving SARS-CoV-2 variants showing that Omicron’s new subvariants have high capacity of immune evasion and accordingly emergency use authorization revoked for almost all the currently available NAbs. In this work, we describe a handy rapid method that uses free online computational tools to assess existing SARS-CoV-2 variant/mutations or as a predictive method to anticipate the effect of possible new mutations. Molecular replacement is a straightforward approach to estimate the binding affinity of antibodies considering that an antibody may lose up to 40% of its affinity and retain therapeutic power and remain clinically useful. We evaluated the neutralization effect on the new SARS-CoV-2 variants (EG.5, BA.2.86, and JN.1) of ten of different classes of neutralizing antibodies, nine of them are granted or previously granted emergency use authorization and one (JS026) is under clinical trials. The neutralization effect was calculated in the form of ΔG percentage in comparison to Wuhan original strain. Our in-silico method showed similar results for the neutralization resistance of the new variants as seen in Figure 2. It is clearly showing that almost all of the available neutralizing antibodies are ineffective against the new SARS-CoV-2 variants EG.5, BA.2.86 and JN.1.
Moreover, our data is consistent with recently published clinical data where the neutralization evasion of SARS-Cov-2 pseudo-virus was less pronounced for BA.2.86 than XBB descendant EG.5 as per the neutralization titer measured in human sera samples with distinct vaccine and infection histories. These studies demonstrated that XBB descendant particularly EG.5 and EG.5.1 evades neutralizing antibodies with increased efficiency than BA.2 descendant BA.2.86 [17,58]. This enhanced neutralization of SARS-CoV-2 variant EG.5 over BA.2.86 could be the result of its unique spike protein mutations of confirmed reduced sensitivity to neutralizing antibody that are not acquired by BA.2.86 including F456L and F490S mutations [9,59,60]. F456L mutation is one of two mutations (L455S and F456L) known as the “Flip” mutations. Both mutations are known to enhance ACE2 binding and hence virus infectivity and they are well known to reduce the neutralization efficacy by evading Class 1 NAbs. Furthermore, the combination of these mutations is synergic [61,62]. This has been proven to be true for BA.2.86 infectivity in comparison to EG.5. However, BA.2.86 gain fitness in its descendants JN.1 with the L455S mutation [23]. Additionally, the 3D structural alignment pointed out a loop distortion caused by the point mutation Ser371Phe at the region between Tyr369 and Ala372. This structural alteration was described before as broadly affecting the binding of class 3 and class 4 RBD directed neutralizing antibodies [63,64].
Although, most of the NAbs responses (ΔG percentage) were higher for BA.2.86 and JN.1 than that for EG.5, they all fell below the neutralization of the Wuhan strain except for Evusheld (AZD1061), Imdevimab (REGN10987), Bebtelovimab (LY-CoV1404) and the new JS026 antibody. Both Evusheld (AZD1061) and Bebtelovimab (LY-CoV1404) showed a very slight increase for the binding affinity of BA.2.86 and JN.1 but not EG.5. The increase above the Wuhan threshold is almost negligible with less than 1.2-fold. Nevertheless, Imdevimab (REGN10987) which is one of the REGEN-COV (Ronapreve) cocktail showed a 2.8-fold increase in the neutralization effect of BA.2.86 and 3-fold increase for its descendant JN.1. Additionally the new potential therapeutic antibody JS026 [11] which is currently under clinical trials showed promising results with neutralization effect of variants EG.5, BA.2.86 and JN that display 3, 2.9 and 3.1 folds increase respectively. Clinically the potential effect of JS026 antibody was discussed in newly published research investigating the use of new therapeutic cocktails to countermeasure residual changes on the spike protein of new SARS-CoV-2 variants. It showed that using combinations of NAbs from different classes, in this case JS026 (class III) with Etesevimab (class I) can increase neutralizing efficacy [37]. Class I and class III were shown to be the best choice for NAbs cocktails as they have distinct epitopes on the RBD domain unlike class II that may overlap with the ACE2-binding site of class I causing steric clashes. Considering that currently clinical data is the ultimate basis for estimating the therapeutic usefulness of a given antibody to treat new variants; this method can be further validated with the availability of more clinical data with new or existing mAbs.
5. Conclusions
This work supports In silico method [28] to evaluate available SARS-CoV-2 antibodies neutralizing power with SARS-CoV-2 emerging variants as single therapy or in Mabs cocktails. It is the first paper to discuss the neutralizing efficiency of all available emergency authorized neutralizing antibodies with the new SARS-CoV-variants. Moreover, NAb cocktail Ronapreve also known as REGEN-COV (Casirivimab / imdevimab) may be still effective against BA.2.86 and JN.1 variants but not EG.5. It is highly probable that combination of Mab Etesevimab in with Mab JS026 is effective against new variants [37]. Furthermore, we propose to look into different NAbs cocktails’ mix that increases neutralizing efficacy against the new variants such as Regdanvimab (class I) with either of JS026 or imdevimab (class III) for strains BA.2.86 and JN.1 and Regdanvimab with JS026 for EG.5.
Author Contributions
In silico analysis, methodology, Illustrations and figures, data curation and analysis, writing, and editing Dana N. Ashoor.
Data availability statement: All data generated or analysis during this study are included in this published article.
Acknowledgments
We thank Dr. Maryam Marzouq for her support and insight in the preparation of this manuscript. We thank Mr. Nasser Abotaleb for continuous technical assistance during this research project.
Conflicts of Interest
No conflicts interests.
Appendix A

Table A1.
Calculated Gibbs free energy (ΔG) of the analyzed neutralizing antibodies with the reference Wuhan strain and the evaluated SARS-CoV-2 variants ; EG.5, BA.2.86 and JN.1.
Table A1.
Calculated Gibbs free energy (ΔG) of the analyzed neutralizing antibodies with the reference Wuhan strain and the evaluated SARS-CoV-2 variants ; EG.5, BA.2.86 and JN.1.
| Antibody | Wuhan ΔG | EG.5 ΔG | BA.2.86 (Pirola) ΔG |
|---|---|---|---|
| Sotrovimab (S309) |
-7.05 | -3.58 | -2.93 |
| Bamlanivimab (LY-CoV555) |
-16.15 | -8.14 | -13.15 |
| Evusheld (AZD8895) |
-9.58 | -2.71 | -3.93 |
| Evusheld (AZD1061) |
-10.84 | -11.32 | -11.91 |
| Regdanvimab (CT-P59) |
-15.05 | -12.62 | -13.22 |
| Etesevimab (LY-CoV016) |
-15.9 | -11.67 | -13 |
| Casirivimab (REGN10933) |
-14.31 | -9.11 | -11.03 |
| Imdevimab (REGN10987) |
-4.51 | -2.95 | -12.69 |
| Bebtelovimab (LY-CoV1404) |
-9.87 | -7.84 | -12.2 |
| JS026 | -2.64 | -8.27 | -7.58 |
References
- Casadevall, A.; Dadachova, E.; Pirofski, L.-a. Passive antibody therapy for infectious diseases. Nature Reviews Microbiology 2004, 2, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.C.; Adams, A.C.; Hufford, M.M.; De La Torre, I.; Winthrop, K.; Gottlieb, R.L. Neutralizing monoclonal antibodies for treatment of COVID-19. Nature Reviews Immunology 2021, 21, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Jiang, J.; He, M.; Zheng, Z.; Yu, H.; Li, T.; Xue, W.; Tang, Z.; Ying, D.; Li, Z. Viral neutralization by antibody-imposed physical disruption. Proceedings of the National Academy of Sciences 2019, 116, 26933–26940. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Muecksch, F.; Lorenzi, J.C.C.; Leist, S.R.; Cipolla, M.; Bournazos, S.; Schmidt, F.; Maison, R.M.; Gazumyan, A.; Martinez, D.R. Antibody potency, effector function, and combinations in protection and therapy for SARS-CoV-2 infection in vivo. Journal of Experimental Medicine 2020, 218, e20201993. [Google Scholar] [CrossRef] [PubMed]
- FDA. Fact sheet for healthcare providers: Emergency Use Authorization for bebtelovimab. Available online: https://www.fda.gov/media/156152/download (accessed on.
- FDA. Fact sheet for healthcare providers: Emergency Use Authorization (EUA) of sotrovimab. Available online: https://www.fda.gov/media/149534/download (accessed on.
- FDA. Fact sheet for healthcare providers: Emergency Use Authorization for Evusheld (tixagevimab co-packaged with cilgavimab). Available online: https://www.fda.gov/media/154701/download (accessed on.
- Imai, M.; Ito, M.; Kiso, M.; Yamayoshi, S.; Uraki, R.; Fukushi, S.; Watanabe, S.; Suzuki, T.; Maeda, K.; Sakai-Tagawa, Y. Efficacy of Antiviral Agents against Omicron Subvariants BQ. 1.1 and XBB. New England Journal of Medicine 2023, 388, 89–91. [Google Scholar] [CrossRef]
- Wang, Q.; Iketani, S.; Li, Z.; Liu, L.; Guo, Y.; Huang, Y.; Bowen, A.D.; Liu, M.; Wang, M.; Yu, J. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 2023, 186, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Yamayoshi, S.; Halfmann, P.; Wilson, N.; Ries, H.; Richardson, A.; Bobholz, M.; Vuyk, W.; Maddox, R.; Baker, D.A. In vitro efficacy of antiviral agents against òmicron subvariant BA. 4.6. New England Journal of Medicine 2022, 387, 2094–2097. [Google Scholar] [CrossRef]
- de Almeida Oliveira, A.; Praia Borges Freire, D.; Rodrigues de Andrade, A.; de Miranda Marques, A.; da Silva Madeira, L.; Moreno Senna, J.P.; Freitas Brasileiro da Silveira, I.A.; de Castro Fialho, B. The Landscape of Neutralizing Monoclonal Antibodies (nAbs) for Treatment and Prevention of COVID-19. Journal of Pharmaceutical Innovation 2023, 1–19. [Google Scholar]
- WHO. COVID-19 Epidemiological Update, Edition 162 published 22 December 2023. Available online: https://www.who.int/publications/m/item/covid-19-epidemiological-update---22-december-2023 (accessed on 2024).
- Dyer, O. Covid-19: Infections climb globally as EG. 5 variant gains ground. BMJ 2023, p1900. [Google Scholar] [CrossRef]
- Rasmussen, M.; Møller, F.T.; Gunalan, V.; Baig, S.; Bennedbæk, M.; Christiansen, L.E.; Cohen, A.S.; Ellegaard, K.; Fomsgaard, A.; Franck, K.T. First cases of SARS-CoV-2 BA. 2.86 in Denmark, 2023. Eurosurveillance 2023, 28, 2300460. [Google Scholar]
- Looi, M.-K. Covid-19: Scientists sound alarm over new BA. 2.86 “Pirola” variant. BMJ: British Medical Journal (Online) 2023, 382, p1964. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, Y.; Zhang, R.M.; Ho, J.; Mohri, H.; Valdez, R.; Manthei, D.M.; Gordon, A.; Liu, L.; Ho, D.D. Antibody neutralization of emerging SARS-CoV-2: EG. 5.1 and XBC. 1.6. bioRxiv 2023, 2023–2008. [Google Scholar]
- Lasrado, N.; Ai-ris, Y.C.; Hachmann, N.P.; Miller, J.; Rowe, M.; Schonberg, E.D.; Rodrigues, S.L.; LaPiana, A.; Patio, R.C.; Anand, T. Neutralization escape by SARS-CoV-2 Omicron subvariant BA. 2.86. Vaccine 2023. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Chatterjee, S.; Lee, S.-S.; Dhama, K.; Chakraborty, C. Antibody evasion associated with the RBD significant mutations in several emerging SARS-CoV-2 variants and its subvariants. Drug Resistance Updates 2023, 101008. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Z.; Guo, Y.; Guo, H.; Jian, L.; Xiao, J.; Yao, X.; Yu, H.; Cheng, T.; Zhang, Y.; et al. Evolving spike mutations in SARS-CoV-2 Omicron variants facilitate evasion from breakthrough infection-acquired antibodies. Cell Discovery 2023, 9, 86. [Google Scholar] [CrossRef]
- Sheward, D.J.; Yang, Y.; Westerberg, M.; Öling, S.; Muschiol, S.; Sato, K.; Peacock, T.P.; Hedestam, G.B.K.; Albert, J.; Murrell, B. Sensitivity of the SARS-CoV-2 BA. 2.86 variant to prevailing neutralising antibody responses. The Lancet Infectious Diseases 2023, 23, e462–e463. [Google Scholar] [CrossRef] [PubMed]
- Uriu, K.; Ito, J.; Kosugi, Y.; Tanaka, Y.L.; Mugita, Y.; Guo, Z.; Hinay, A.A.; Putri, O.; Kim, Y.; Shimizu, R. Transmissibility, infectivity, and immune evasion of the SARS-CoV-2 BA. 2.86 variant. The Lancet Infectious Diseases 2023, 23, e460–e461. [Google Scholar] [CrossRef]
- WHO. Initial Risk Evaluation of JN.1, 19 December 2023. 2023.
- Yang, S.; Yu, Y.; Xu, Y.; Jian, F.; Song, W.; Yisimayi, A.; Wang, P.; Wang, J.; Liu, J.; Yu, L. Fast evolution of SARS-CoV-2 BA. 2· 86 to JN. 1 under heavy immune pressure. The Lancet Infectious Diseases 2023.
- Kaku, Y.; Okumura, K.; Padilla-Blanco, M.; Kosugi, Y.; Uriu, K.; Hinay, A.A.; Chen, L.; Plianchaisuk, A.; Kobiyama, K.; Ishii, K.J. Virological characteristics of the SARS-CoV-2 JN. 1 variant. The Lancet Infectious Diseases 2023. [Google Scholar] [CrossRef]
- Magnus, C.L.; Hiergeist, A.; Schuster, P.; Rohrhofer, A.; Medenbach, J.; Gessner, A.; Peterhoff, D.; Schmidt, B. Targeted escape of SARS-CoV-2 in vitro from monoclonal antibody S309, the precursor of sotrovimab. Frontiers in immunology 2022, 13, 966236. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2022, 602, 657–663. [Google Scholar] [CrossRef]
- Abbasi, J. What to Know About EG.5, the Latest SARS-CoV-2 "Variant of Interest". Jama 2023, 330, 900–901. [Google Scholar] [CrossRef] [PubMed]
- Ashoor, D.; Marzouq, M.; Fathallah, M.D. In silico evaluation of anti SARS-CoV-2 antibodies neutralization power: A blueprint with monoclonal antibody Sotrovimab (preprint). 2023. [CrossRef]
- Starr, T.N.; Czudnochowski, N.; Liu, Z.; Zatta, F.; Park, Y.-J.; Addetia, A.; Pinto, D.; Beltramello, M.; Hernandez, P.; Greaney, A.J.; et al. SARS-CoV-2 RBD antibodies that maximize breadth and resistance to escape. Nature 2021, 597, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Brown-Augsburger, P.L.; Corbett, K.S.; Westendorf, K.; Davies, J.; Cujec, T.P.; Wiethoff, C.M.; Blackbourne, J.L.; Heinz, B.A.; Foster, D.; et al. The neutralizing antibody, LY-CoV555, protects against SARS-CoV-2 infection in nonhuman primates. 2021, 13, eabf1906. [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Laurini, E.; Marson, D.; Aulic, S.; Fermeglia, A.; Pricl, S. Molecular rationale for SARS-CoV-2 spike circulating mutations able to escape bamlanivimab and etesevimab monoclonal antibodies. Scientific Reports 2021, 11, 20274. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zost, S.J.; Greaney, A.J.; Starr, T.N.; Dingens, A.S.; Chen, E.C.; Chen, R.E.; Case, J.B.; Sutton, R.E.; Gilchuk, P.; et al. Genetic and structural basis for SARS-CoV-2 variant neutralization by a two-antibody cocktail. Nature Microbiology 2021, 6, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Ryu, D.-K.; Lee, J.; Kim, Y.-I.; Seo, J.-M.; Kim, Y.-G.; Jeong, J.-H.; Kim, M.; Kim, J.-I.; Kim, P.; et al. A therapeutic neutralizing antibody targeting receptor binding domain of SARS-CoV-2 spike protein. Nature Communications 2021, 12, 288. [Google Scholar] [CrossRef]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. 2020, 369, 1010-1014. [CrossRef]
- Westendorf, K.; Žentelis, S.; Wang, L.; Foster, D.; Vaillancourt, P.; Wiggin, M.; Lovett, E.; van der Lee, R.; Hendle, J.; Pustilnik, A.; et al. LY-CoV1404 (bebtelovimab) potently neutralizes SARS-CoV-2 variants. Cell reports 2022, 39, 110812. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, L.; Dou, Y.; Shi, R.; Duan, X.; Liu, H.; Zhang, J.; Liu, D.; Wu, J.; He, Y.; et al. Etesevimab in combination with JS026 neutralizing SARS-CoV-2 and its variants. Emerging Microbes & Infections 2022, 11, 548–551. [Google Scholar] [CrossRef]
- Huang, Q.; Han, X.; Yan, J. Structure-based neutralizing mechanisms for SARS-CoV-2 antibodies. Emerging Microbes & Infections 2022, 11, 2412–2422. [Google Scholar]
- Stanford-University. SARS-CoV-2 Variants. Available online: https://covdb.stanford.edu/variants/omicron_ba_1_3/ (accessed on 2023).
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic acids research 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: an environment for comparative protein modeling. electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- van Gunsteren, W.F.; Billeter, S.R.; Eising, A.A.; Hünenberger, P.H.; Krüger, P.; Mark, A.E.; Scott, W.R.P.; Tironi, I.G. Biomolecular simulation: the GROMOS96 manual and user guide. Vdf Hochschulverlag AG an der ETH Zürich, Zürich 1996, 86, 1–1044. [Google Scholar]
- Myung, Y.; Pires, D.E.V.; Ascher, D.B. CSM-AB: graph-based antibody–antigen binding affinity prediction and docking scoring function. Bioinformatics 2022, 38, 1141–1143. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: multiple ligand-protein interaction diagrams for drug discovery. Journal of chemical information and modeling 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Secondary-structure matching (SSM), a new tool for fast protein structure alignment in three dimensions. Acta Crystallographica Section D: Biological Crystallography 2004, 60, 2256–2268. [Google Scholar] [CrossRef]
- DeLano, W. The PyMOL Molecular Graphics System, Version 2.4. 0 Schrödinger, LLC. 2002.
- FDA. Emergency Use Authorization. Available online: https://www.fda.gov/emergency-preparedness-and-response/mcm-legal-regulatory-and-policy-framework/emergency-use-authorization#coviddrugs (accessed on 2023).
- EMA. COVID-19 treatments. Available online: https://www.ema.europa.eu/en/human-regulatory/overview/public-health-threats/coronavirus-disease-covid-19/treatments-vaccines/covid-19-treatments (accessed on 2023).
- Chen, Y.; Zhao, X.; Zhou, H.; Zhu, H.; Jiang, S.; Wang, P. Broadly neutralizing antibodies to SARS-CoV-2 and other human coronaviruses. Nature reviews Immunology 2023, 23, 189–199. [Google Scholar] [CrossRef]
- Touret, F.; Baronti, C.; Bouzidi, H.S.; de Lamballerie, X. In vitro evaluation of therapeutic antibodies against a SARS-CoV-2 Omicron B.1.1.529 isolate. Scientific Reports 2022, 12, 4683. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Casadevall, A. A critical analysis of the use of cilgavimab plus tixagevimab monoclonal antibody cocktail (Evusheld™) for COVID-19 prophylaxis and treatment. Viruses 2022, 14, 1999. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, X.; Zhang, Z.; Liu, X.; Wang, P.; Cao, J.; Liang, Q.; Qu, J.; Zhou, M. Serum neutralization of SARS-CoV-2 Omicron BA.2, BA.2.75, BA.2.76, BA.5, BF.7, BQ.1.1 and XBB.1.5 in individuals receiving Evusheld. Journal of medical virology 2023, 95, e28932. [Google Scholar] [CrossRef] [PubMed]
- WHO. Tracking SARS-CoV-2 variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 2024).
- Wang, L.; Møhlenberg, M.; Wang, P.; Zhou, H. Immune evasion of neutralizing antibodies by SARS-CoV-2 Omicron. Cytokine & growth factor reviews 2023.
- Miller, J.; Hachmann, N.P.; Collier, A.-r.Y.; Lasrado, N.; Mazurek, C.R.; Patio, R.C.; Powers, O.; Surve, N.; Theiler, J.; Korber, B. Substantial neutralization escape by SARS-CoV-2 Omicron variants BQ. 1.1 and XBB. 1. New England Journal of Medicine 2023, 388, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Faraone, J.N.; Qu, P.; Goodarzi, N.; Zheng, Y.-M.; Carlin, C.; Saif, L.J.; Oltz, E.M.; Xu, K.; Jones, D.; Gumina, R.J. Immune evasion and membrane fusion of SARS-CoV-2 XBB subvariants EG. 5.1 and XBB. 2.3. Emerging Microbes & Infections 2023, 12, 2270069. [Google Scholar]
- Qu, P.; Evans, J.P.; Faraone, J.N.; Zheng, Y.-M.; Carlin, C.; Anghelina, M.; Stevens, P.; Fernandez, S.; Jones, D.; Lozanski, G. Enhanced neutralization resistance of SARS-CoV-2 omicron subvariants BQ. 1, BQ. 1.1, BA. 4.6, BF. 7, and BA. 2.75. 2. Cell host & microbe 2023, 31, 9–17. [Google Scholar]
- Hu, Y.; Zou, J.; Kurhade, C.; Deng, X.; Chang, H.C.; Kim, D.K.; Shi, P.-Y.; Ren, P.; Xie, X. Less neutralization evasion of SARS-CoV-2 BA. 2.86 than XBB sublineages and CH. 1.1. Emerging Microbes & Infections 2023, 12, 2271089. [Google Scholar]
- Abbasi, J. What to know about EG. 5, the latest SARS-CoV-2 “Variant of Interest”. Jama 2023.
- Parums, D.V. a rapid global increase in COVID-19 is due to the emergence of the EG. 5 (Eris) subvariant of omicron SARS-CoV-2. Medical Science Monitor: International Medical Journal of Experimental and Clinical Research 2023, 29, e942244–942241. [Google Scholar] [CrossRef]
- Jian, F.; Feng, L.; Yang, S.; Yu, Y.; Wang, L.; Song, W.; Yisimayi, A.; Chen, X.; Xu, Y.; Wang, P. Convergent evolution of SARS-CoV-2 XBB lineages on receptor-binding domain 455–456 synergistically enhances antibody evasion and ACE2 binding. PLoS pathogens 2023, 19, e1011868. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Spezia, P.G.; Gueli, F.; Maggi, F. The Era of the FLips: How Spike Mutations L455F and F456L (and A475V) Are Shaping SARS-CoV-2 Evolution. 2023, 16, 3.
- Iketani, S.; Liu, L.; Guo, Y.; Liu, L.; Chan, J.F.W.; Huang, Y.; Wang, M.; Luo, Y.; Yu, J.; Chu, H. Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef]
- Liu, L.; Iketani, S.; Guo, Y.; Chan, J.F.W.; Wang, M.; Liu, L.; Luo, Y.; Chu, H.; Huang, Y.; Nair, M.S. Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature 2022, 602, 676–681. [Google Scholar] [CrossRef]
Figure 1.
Neutralizing antibodies’ binding position on SARS-CoV-2 RBD domain (magenta). Antibody (Fv domain) heavy chain (green) and light chain (cyan).
Figure 1.
Neutralizing antibodies’ binding position on SARS-CoV-2 RBD domain (magenta). Antibody (Fv domain) heavy chain (green) and light chain (cyan).
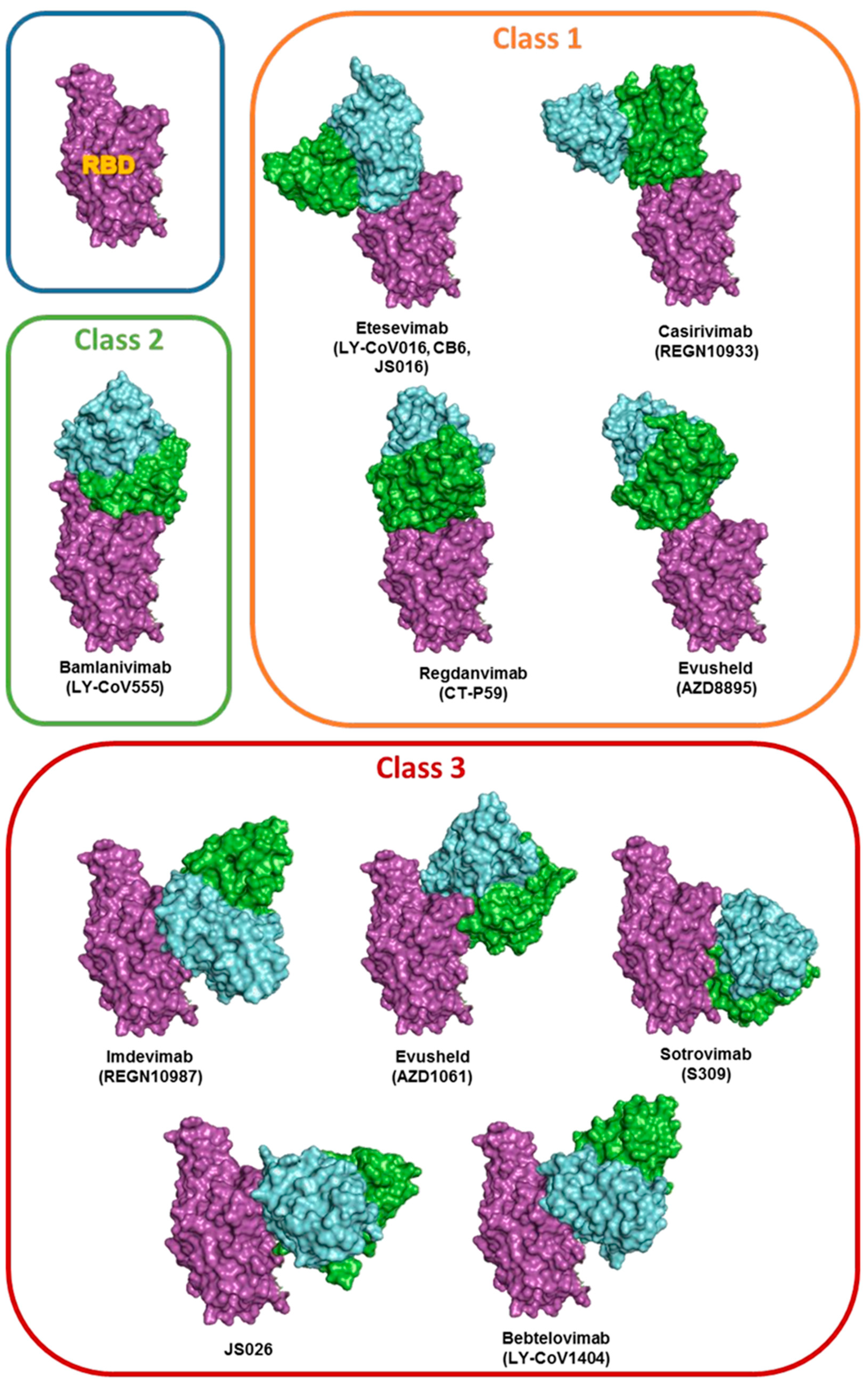
Figure 2.
Neutralization efficiency of nine different antibodies against the new SARS-CoV-2 variants EG.5, BA2.86 and JN1. based on increased/decreased binding affinity (ΔG) percentages are compared to the 100% affinity value attributed to the reference Wuhan strain.
Figure 2.
Neutralization efficiency of nine different antibodies against the new SARS-CoV-2 variants EG.5, BA2.86 and JN1. based on increased/decreased binding affinity (ΔG) percentages are compared to the 100% affinity value attributed to the reference Wuhan strain.
Figure 3.
3D alignment of RBD of SARS-CoV-2 BA.2.286 and JN.1 (Cyan) with Wuhan RBD (Green), showing the effect of Ser371Phe mutation. The location of the mismatch on the RBD domain is encircled in black.
Figure 3.
3D alignment of RBD of SARS-CoV-2 BA.2.286 and JN.1 (Cyan) with Wuhan RBD (Green), showing the effect of Ser371Phe mutation. The location of the mismatch on the RBD domain is encircled in black.
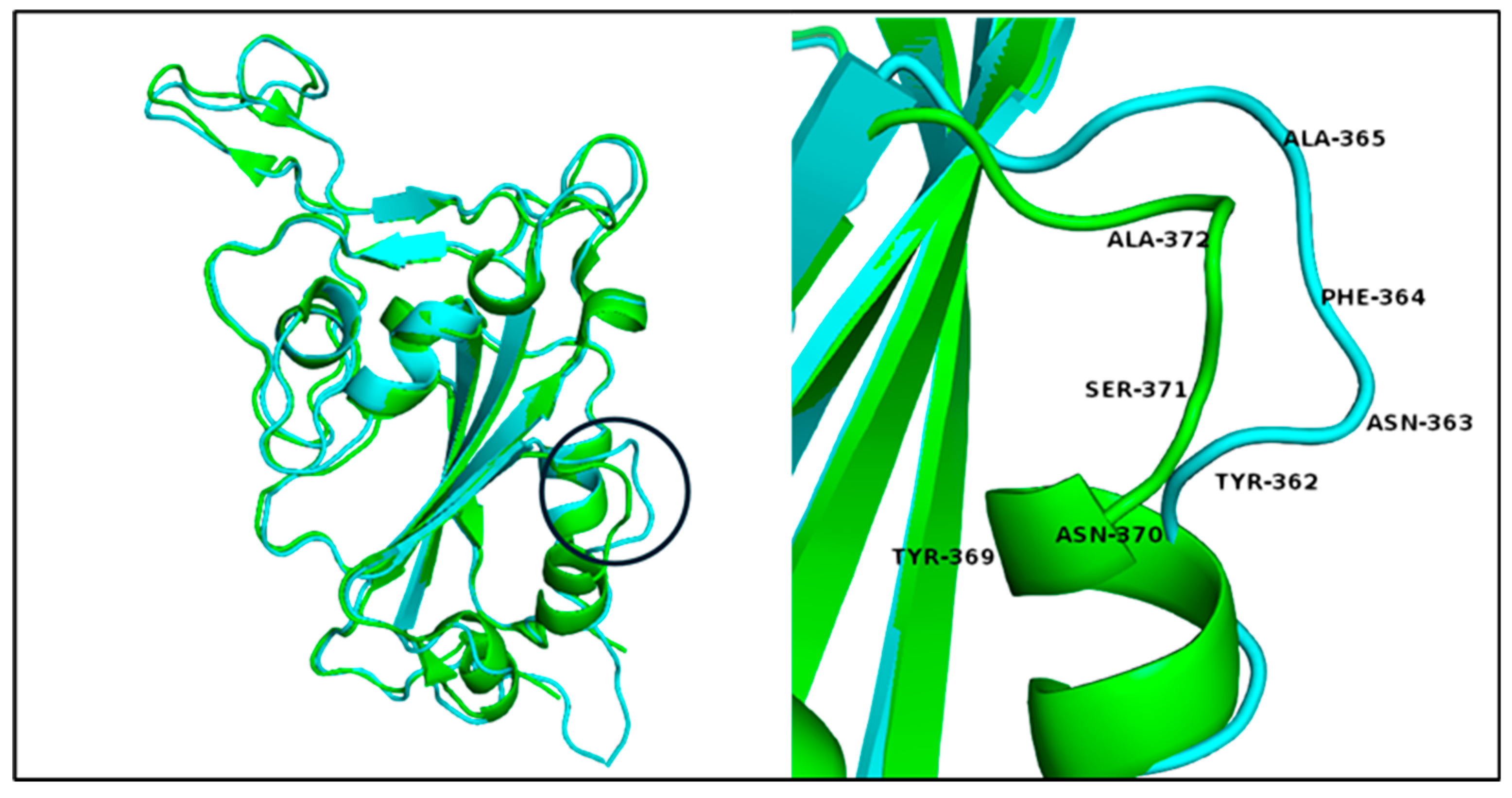
Table 1.
list of PDB template models and used chains IDs. All the PDB selected models represent the interaction of the neutralizing antibodies with Wuhan strain.
Table 1.
list of PDB template models and used chains IDs. All the PDB selected models represent the interaction of the neutralizing antibodies with Wuhan strain.
| # | Model | Resolution Å |
Antibody | Heavy chain ID |
Light chain ID |
RBD chain ID |
Reference |
|---|---|---|---|---|---|---|---|
| 1 | 7R6W | 1.83 | Sotrovimab (S309) | A | B | R | [29] |
| 2 | 7KMG | 2.16 | Bamlanivimab (LY-CoV555) | A | B | C | [30] |
| 3 | 7L7E | 3.00 | Evusheld (AZD8895) | A | B | G | [33] |
| 4 | 7L7E | 3.00 | Evusheld (AZD1061) | E | F | G | [33] |
| 5 | 7CM4 | 2.71 | Regdanvimab (CT-P59) | H | L | A | [34] |
| 6 | 6XDG | 3.90 | Casirivimab (REGN10933) | B | D | E | [35] |
| 7 | 6XDG | 3.90 | Imdevimab (REGN10987) | C | A | E | [35] |
| 8 | 7MMO | 2.43 | Bebtelovimab (LY-CoV1404) | A | B | C | [36] |
| 9 | 7C01 | 2.88 | Etesevimab (LY-CoV016) CB6, JS016, LY3832479 |
H | L | A | [31] [32] |
| 10 | 7F7E | 2.49 | JS026 | C | L | E | [37] |
Table 2.
Summary of SARS-CoV-2 neutralizing antibodies emergency authorization use by U.S. Food and Drug Administration (FDA) and European Medicine Agency (EMA).
Table 2.
Summary of SARS-CoV-2 neutralizing antibodies emergency authorization use by U.S. Food and Drug Administration (FDA) and European Medicine Agency (EMA).
| Antibody | FDA[47] | EMA[48] |
|---|---|---|
| Sotrovimab (S309) | Authorized May 2021 Revoked April 2022 * |
Authorized December 2021 up to date Under the name Xevudy |
| Bamlanivimab (LY-CoV555) |
Authorized September 2021 Administered together as a combination (Bamlanivimab/Etesevimab) Revoked January 2022* |
October 2021 Bamlanivimab and Etesevimab EMA ended the rolling review due to withdrawing from the process by the company (Eli Lilly Netherlands BV) |
| Etesevimab (LY-CoV016, CB6, JS016) | ||
| Evusheld (Tixagevimab -AZD8895) | Authorized August 2021 Revoked January 2023 Evusheld (tixagevimab co-packaged with cilgavimab) cocktail No longer authorized* |
Authorized March 2022 up to date Evusheld (tixagevimab co-packaged with cilgavimab) cocktail |
| Evusheld (Cilgavimab-AZD1061) | ||
| Regdanvimab (CT-P59) | No authorization | Authorized November 2021 up to date Under the name Regkirona |
| Casirivimab (REGN10933) | Authorized November 2020 Revoked January 2022 REGEN-COV (Casirivimab / Imdevimab) cocktail No longer authorized* |
Authorized November 2021 up to date Under the name Ronapreve (Casirivimab / imdevimab) cocktail |
| Imdevimab (REGN10987) | ||
| Bebtelovimab (LY-CoV1404) |
Authorized February 2022 Revoked November 2022 No longer authorized* |
No authorization |
| JS026 | Ongoing clinical trials [11] | |
* Due to increases frequency of resistant variants.
Table 3.
ACE2 competition and neutalization effect of the emergency authorized SARS-CoV-2 neutralization antibodies.
Table 3.
ACE2 competition and neutalization effect of the emergency authorized SARS-CoV-2 neutralization antibodies.
| Antibody | NAb’s class | RBD access |
ACE2 competing |
Viruses neutralized | Reference |
|---|---|---|---|---|---|
| Sotrovimab (S309) | Class 3 | Up/Down | No | SARS-CoV-2; Alpha, Beta, Gamma, Delta, BA.1, BA.2, BA.3, BA.4/5, and BA.2.75 |
[49] |
| Bamlanivimab (LY-CoV555) |
Class 2 | Up/Down | Yes | SARS-CoV-2; Alpha | [32,49,50] |
| Etesevimab (LY-CoV016, CB6, JS016 |
Class 1 | UP | Yes | SARS-CoV-2; Alpha and Delta | |
| Evusheld (Tixagevimab -AZD8895) | Class 1 | Up | Yes | SARS-CoV-2,; Alpha, Beta, Gamma, Delta | [50,51,52] |
| Evusheld (Cilgavimab-AZD1061) | Class 3 | Up/Down | No | SARS-CoV-2,; Alpha, Beta, Gamma, Delta, BA.2, BA.2.75, and BA.5 | |
| Regdanvimab (CT-P59) | Class1 | Up | Yes | SARS-CoV-2; Alpha, Beta, Gamma and Delta | [49] |
| Casirivimab (REGN10933) | Class 1 | UP | Yes | SARS-CoV-2; Alpha and Delta, BA.2.75 | [49] |
| Imdevimab (REGN10987) | Class 3 | Up/Down | No | SARS-CoV-2; Alpha, Beta, Gamma, Delta, BA.1, BA.2, and BA.4/5 | |
| Bebtelovimab (LY-CoV1404) |
Class 3 | Up/Down | No | SARS-CoV-2; Alpha, Beta, Gamma, Delta, BA.1, BA.2, BA.3, BA.4/5, and BA.2.75 | [49] |
| JS026 | Class 3 | Up/Down | No | In combination with Etesevimab. SARS-CoV-2; Alpha, Beta, Gamma and Delta | [37] |
Table 4.
The interaction of Imdevimab and JS026 with epitope residues of the spike RBD of the new variants in comparison to Wuhan strain. The Number and position of polar interactions are indicated by an asterisk (*). Mutated residues are in italic. Residues and numbering in-between brackets correspond to the Wuhan epitope. Salt bridges are underlined.
Table 4.
The interaction of Imdevimab and JS026 with epitope residues of the spike RBD of the new variants in comparison to Wuhan strain. The Number and position of polar interactions are indicated by an asterisk (*). Mutated residues are in italic. Residues and numbering in-between brackets correspond to the Wuhan epitope. Salt bridges are underlined.
| SARS-CoV-2 variant | Wuhan interaction epitope | EG.5 interaction epitope | BA.2.86 interaction epitope | JN.1 interaction epitope |
|---|---|---|---|---|
| Imdevimab | Arg346 | Arg339 (Arg346) | Arg339 (Arg346) | |
| Asn440* | Lys436 (Asn440) | Lys433 (Asn440) | Lys433 (Asn440) | |
| Leu441 | Leu437 (Leu441) | Leu434 (Leu441) | Leu434 (Leu441) | |
| Lys444 | Lys440 (Lys444) | Lys437 (Lys444) | Lys437 (Lys444) | |
| Val445 | Pro441* (Val445) | His438* (Val445) | His438* (Val445) | |
| Gly446 | Ser442 (Gly446) | Ser439 (Gly446) | Ser439 (Gly446) | |
| Gly443 (Gly447) | ||||
| Asn444 (Asn448) | ||||
| Gly440 (Gly447) | Gly440 (Gly447) | |||
| Asp443 (Asp450) | Asp443 (Asp450) | |||
| Arg494 (Gln498) | Arg490 (Gln498) | Arg490 (Gln498) | ||
| Pro495 (Pro499) | Pro491 (Pro499) | Pro491 (Pro499) | ||
| Bonds | 1 polar 14 hydrophobic |
1 polar 28 hydrophobic 1 salt bridge |
1 polar 33 hydrophobic 1 salt bridge |
1 polar 33 hydrophobic 1 salt bridge |
| JS026 | Asn343* | |||
| Thr345* | Thr341 (Thr345) | Thr338 (Thr345) | Thr338 (Thr345) | |
| Arg346 | Thr342 (Arg346) | Arg339 (Arg346) | Arg339 (Arg346) | |
| Asn439 | Asn435 (Asn439) | Asn432 (Asn439) | Asn432 (Asn439) | |
| Asn440** | Lys436* (Asn440) | Lys433* (Asn440) | Lys433* (Asn440) | |
| Leu441* | Leu437 (Leu441) | Leu434 (Leu441) | Leu434 (Leu441) | |
| Asp442* | Asp438* (Asp442) | Asp435* (Asp442) | Asp435* (Asp442) | |
| Ser443* | Ser439* (Ser443) | Ser436* (Ser443) | Ser436* (Ser443) | |
| Lys444 | Lys440* (Lys444) | Lys437* (Lys444) | Lys437* (Lys444) | |
| Val445 | Pro441 (Val455) | His438 (Val445) | His438 (Val445) | |
| Asn448 | Asn444* (Asn448) | Asn441* (Asn448) | Asn441* (Asn448) | |
| Tyr451 | Tyr447 (Tyr451) | Tyr444 (Tyr451) | Tyr444 (Tyr451) | |
| Pro499 | Pro495 (Pro499) | Pro491 (Pro499) | Pro491 (Pro499) | |
| Thr500 | Thr496 (Thr500) | Thr492 (Thr500) | Thr492 (Thr500) | |
| Arg509 | Arg505 (Arg509) | Arg501 (Arg509) | Arg501 (Arg509) | |
| Bonds | polar 54 hydrophobic |
4 polar 66 hydrophobic |
6 polar 72 hydrophobic |
75 polar 75 hydrophobic salt bridge |
Table 5.
3D structural alignment of the RBDs of SARS-CoV-2 variants and Wuhan strain.
| Wuhan PBD ID: 7R6W- Chain R |
BA.2.86 and JN.1 | EG.5 |
|---|---|---|
| Tyr 369 Asn 370 Ser 371 Ala 372 |
Tyr 362 Asn 363 Phe 364 Ala 365 |
Tyr 365 Asn 366 Phe 367 Ala 368 |
| - Lys 386 Leu 387 |
Lys 379 Leu 380 - |
Lys 382 Leu 383 - |
| Gly 482 | - | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



