Submitted:
16 August 2024
Posted:
16 August 2024
You are already at the latest version
Abstract
Astragalus plants belong to the legume family and have obvious rhizosphere effect. The northwestern area of Yunnan China is the main distribution region of Astragalus plants with high application value, but low recognition and resource reserves. The aim of this research is to explore the species diversity of cultivable rhizosphere fungi derived from A. acaulis, A. forrestii, and A. ernestii growing in special high-cold environment of northwest Yunnan, and discover anti-infective components from these fungi. A total of 93 fungal strains belonging to 38 species in 18 genera were isolated and identified. Antibacterial and antimalarial screening yielded 10 target strains. Among them, the ethyl acetate crude extract of the fungus Aspergillus calidoustus AA12 from A. acaulis fermentation substance showed a broad antibacterial spectrum and the best antimalarial activity. Further chemical investigation led to the first discovery of seven compounds from the species of A. calidoustus, including a sesterterpine 6-epi-ophiobolin G (1), two sesquiterpenes penicisochroman A (2), pergillin (3) and 7-methyl-2-(1-methylethylethlidene)-furo[3,2-H]isoquinoline-3-one (4), and three polyketides trypacidin (5), 1,2-seco-trypacidin (6) and questin (7). Compound 1 exhibited moderate to strong antibacterial activity against six Gram-positive pathogens with the minimum inhibitory concentration (MIC) ranging from 25 to 6.25 μg/mL, and prominent inhibitory effect on the biofilm of Streptococcus agalactiae at MIC value of 3.125 μg/mL. This compound also displayed potent antimalarial activity against Plasmodium falciparum strains 3D7 and chloroquine-resistant Dd2 at the half maximal inhibitory concentration (IC50) values of 3.319 and 4.340 µmol/L at 72 h, respectively. This study contributed to our understanding of the cultivable rhizosphere fungi Astragalus-derived in special high-cold environment and further increased the library of fungi available for natural anti-infectious products screening.
Keywords:
Astragalus
; rhizosphere fungi
; species diversity
; secondary metabolites;antibacterial activity
; antimalarial activity
; special eco-environment
1. Introduction
The rhizosphere refers to the narrow region of soil, and is usually considered as a special eco-environment containing up to 1011 microbial cells [1,2]. These rhizosphere microbial communities are diverse, and is of great significance to the process of growth regulation, defence and stress tolerance, as well as quality of plants [3]. The application of high-throughput sequencing, such as metagenomics, enables researchers to comprehensively recognize the population distribution patterns of microbes in rhizosphere, and enhances the knowledge of environmental and ecological interactions [4]. However, to further explore the functional potential of some microbes, it is necessary to obtain pure cultured strains using traditional culture-dependent method.
Some pathogenic bacteria and Plasmodium parasites, especially drug-resistant strains, are posing a serious threat to human public health now [5]. The global problem of antibiotic resistance is fast becoming one of the major scientific issues of modern times [6]. Moreover, it does not seem to be a global problem, but in fact, malaria is impacting a considerable proportion of the population. Even now the Plasmodium parasites are present in eighty five countries spread across five WHO regions [7]. Worse yet, the P. falciparum has developed resistance to a variety of antimalarial medications, including chloroquine and artemisinin derivatives. Therefore, the development and exploitation of effectively antibacterial and antiplasmodial resources and lead compounds remains an urgent need for anti-infective drugs industry.
Fungi, particularly inhabiting in special eco-environment, have been proved as promising and prolific sources of bioactive secondary metabolites with potent anti-infectious applications [8]. In recent years, more and more research articles have reported works on the discovery of antibacterial or antimalarial natural products from marine-derived fungi [9,10,11]. However, there are few reports on the study of anti-infective medicinal sources of microorganisms in other special eco-environments. Hence, broadening the research visibility into antibacterial and antimalarial microbes in other special inhabits, such as polar, alpine, hyperhaline, plant rhizosphere, animal internal and so on, will be meaningful for the more comprehensive utilization of anti-infective fungal resources in special eco-environment.
The legume Astragalus is an important medicinal herb with noticeable rhizosphere effect in traditional Chinese medicine and thrive in high latitudes [12]. Although located in the low latitude, some northwestern regions of Yunnan province of China, have the similarly climatic characteristics of high latitudes because of the effect of high altitude (> 3000 m). During our latest field investigation of characteristic plant resources distribution of Yunnan, a total of around 30 species of Astragalus plants were found to grow endemically in northwest Yunnan with altitudes ranging from 3,040 to 4,479 m. Besides our previous study on the diversity of rhizosphere microbial communities associated with three species of Astragalus using metagenomics [13] and the new species of rhizofungus Varicosporellopsis shangrilaensis isolated from the plant A. polycladus [14], there is still large amount unknown about these Astragalus-derived microorganisms. But should not be ignored is that the microbes in rhizosphere of these characteristic Astragalus may be abundant, and could produce secondary metabolites with excellent bioactivity being due to the dual action of rhizosphere and high-cold conditions. Herein, it is reasonably suspected that the rhizofungi derived from Astragalus plants in alpine environment of northwestern Yunnan should have promising potential for the development and exploitation of antibacterial and antimalarial medical resources.
In this study, we isolated and identified rhizofungi from three species of Astragalus plants, including A. acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae) in special high-cold environment of Yunnan at different altitudes ranging from 3,225 to 4,353 m, analyzed the diversity and structure of the culturable fungal communities comparing with the previously reported metagenomic results [13], conducted anti-infective (antibacterial activity against seven Gram-positive pathogenic bacteria and antimalarial effect on two strains of Plasmodium falciparum) screening of the ethyl acetate (EA) crude extracts of the cultivable fungi, and performed further isolation, structure elucidation, and anti-infective evaluation for the obtained pure compounds from Aspergillus calidoustus AA12. The information of cultivated fungal diversity and distribution in the three species of Astragalus is very scarce and important, which provides a reference for the overall awareness of correlation between the host plants, ecological environment and community composition of fungal microbes between different Astragalus plants growing in the northwestern Yunnan. The results of anti-infectious evaluation and chemical investigation of these rhizofungi, meanwhile, suggested that ten strains in five species, such as Aspergillus calidoustus, A. tabacinus, Penicillium brevicompactum, Lecanicillium aphanocladii and Umbelopsis nana, should be considered as potent resources of anti-infective natural products, which is being instructive on exploring these distinctive fungi as bioactive microbial resource in special eco-environment.
2. Materials and Methods
2.1. Rhizosphere Soil Samples Collection
Soil samples were collected from the rhizosphere sediments (at depths of 4.0–5.0 cm) of three species of Astragalus plants in Dêqên county, Shangri-La city, Dêqên Tibetan Autonomous Prefecture, Yunnan Province, China. The plants of Astragalus acaulis (Aa) thriving in alpine grassland and A. ernestii (Ae) surviving in the steep slopes of flowstone beaches distribute in Baima Snow Mountain at respective 4353 and 4048 m of altitude (99°2′ E, 28°21′ N). The plant A. forrestii (Af) surrounded by more pine trees and deciduous foliage inhabits in Xiaozhongdian town of Shangri-La at an elevation of 3, 225 m (99°49ʹ E, 27°26ʹ N). The detailed sampling sites, plant appearance and growing environment of these three Astragalus are shown in Figure S1 and S2 [13]. These three healthy plants and the soil closely attached to the roots (within 2.5 mm) [15] were randomly collected in each plot. These samples packed in aseptic bags were immediately low-temperature transported to the laboratory, and stored at 4 °C until processing.
2.2. Isolation and Identification of Fungi
Fungal strains were isolated from the three Astragalus rhizosphere soil using a standard dilution-plating technique. About 1.5 g soil sample was weighed and suspended in 15 mL of sterile distilled water, and then diluted to the final concentration of 10-3 g/mL using gradient dilution method. About 200 μL of soil suspension was evenly spread on the surface of potato dextrose agar (PDA) plates with 100 mg/L of streptomycin sulfate and penicillin [14]. The plates were incubated at 28 °C for 3-5 days until several individual colonies formed, and were then observed with a stereomicroscope. The fungal mycelia were transferred to fresh PDA plates using a sterile needle. This step was repeated until the pure culture was obtained.
Cultivable fungal genomic DNA was extracted using a Rapid Genomic DNA Isolation Kit (Sangon Biotech Co., Ltd., Shanghai, China) according to the manufacturer’s instructions. Polymerase Chain Reaction (PCR) amplification was performed using primers ITS1(5′-TCCGTAGGTGAACCTGCGG3′) and ITS4(5′-TCCTCCGCTTATTGATATGC-3′) for the extracted fungal genome. The PCR amplification was performed in a 25 μL reaction system containing 2 μL of DNA template, 1 μL of each primer, 8.5 μL of ddH2 O, and 12.5 μL of 2 × Taq PCR MasterMix (Sangon, Biotech Company, Limited, Shanghai, China). PCR amplifications were performed using the following program: after pre-denaturation at 94°C for 10 min, amplification was performed with 30 cycles, denaturation at 94°C for 30 s, annealing at 55°C for 30 s, extension at 72°C for 90 s, and extension at 72°C for 5 min. The PCR products were verified by agarose gel electrophoresis. All sequencing results were analyzed for shearing using SeqMan software, and then the isolates were identified by the blast program in the NCBI database. The identified strains were preserved in the Yunnan Key Laboratory of Screening and Research on Anti-pathogenic Plant Resources from Western Yunnan, Dali, Yunnan, China.
2.3. Fermentation and Extraction
All fungal isolates were cultivated in rice medium (100 mL double-distilled water, 100 g rice, 0.6 g peptone) in 1 L Erlenmeyer flask at 27 °C for 4 weeks. The ethyl acetate (EA) was used to extract repeatedly (3 × 300 mL) the fermented substrate. Then the extractive EA was concentrated by vacuum evaporation to afford a residue. Finally, the EA crude extract of the fungal fermentation was obtained and dissolved in indimethyl sulfoxide (DMSO) to afford 100 mg/mL stock solution for testing.
2.4. Antibacterial Bioassay
A total of seven strains of Gram-positive pathogenic bacteria, including methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300, S. aureus ATCC 25923, S. epidermidis ATCC 12228, Streptococcus agalactiae ATCC 13813, vancomycin-resistant Enterococcus faecalis ATCC 51299, Enterococcus faecalis ATCC 29212 and E. faecium ATCC 35667, were used for antibacterial bioassay. The method of antibacterial evaluation (represented by MIC value) was in accordance with described by Song et al. [16]. These pathogens were incubated in Lysogeny broth (LB) at 28°C and collected at OD595 value in a range of 0.1 to 0.2. Then the culture collection was diluted by LB medium to obtain the final bacterial suspension of 5×10−5 CFU/mL, 200 µL of which was transferred into 96-well microplate (Corning, Corning, NY, United States). The desired stock solution of samples were added to each well containing 100 µL microbial suspension, and further diluted using a serial twofold dilution to obtain the tested solutions with five concentrations. There were five replicate wells per concentration. The wells containing DMSO–suspension (v/v 1:100) and different concentrations of vancomycin were served as the negative and positive control, respectively. These tested microplates was then incubated at 37°C and the OD595 value was measured after 24 h for the determination of MIC value. All the experiments were repeated three times.
2.5. Antimalarial Bioassay (In Vitro)
Two strains Plasmodium falciparum of 3D7 and chloroquine-resistant Dd2 were used as model protozoa, which were provided by Shanghai Institute of Immunity and Infection, Chinese Academy of Sciences. As the modified protocol stated by Gudla et al. [17] with little differences, Plasmodium strains were maintained in RPMI-1640 medium at 2–5% hematocrit supplemented with 0.2% w/v glucose, 0.5% w/v AlbuMaxII, 0.22% w/v NaHCO3, 50 µg/mL gentamycin and 50 µg/mL hypoxanthine, and incubated at 37°C for 72 h under micro-aerophilic atmosphere (4 % CO2, 3 % O2 and 93 % N2). The 5% w/v D-sorbitol method was used for parasite synchronization. The stock solution of the tested samples were serially diluted two-fold with culture medium. A total of 10 μL diluted substance were added to 190 μL parasite inoculum (1% parasitaemia, 2% haematocrit) of 96-well tissue culture plate. After 72 h of incubation at 37°C, each well was further processed by 100 µL of RBCs lytic buffer (5 mM EDTA, 20 mM Tris pH 7.5, 0.12% v/v Triton X-100, and 0.012% w/v Saponin) containing the SYBR green 1X final concentration. Then plates were incubated at room temperature for 2 h in the dark, and conducted under a fluorescence reader at an excitation of 485 nm and an emission of 535 nm. The chloroquine-treated group as the positive control. The inhibition rate for the antimalarial activity of the tested samples was determined based on the DNA content of the parasites. The IC50 value for Plasmodium was calculated by GraphPad software. Five biological replicates and three biological replicates were carried out in the antimalarial bioassay.
2.6. Isolation and Structural Elucidation of Secondary Metabolites from Aspergillus calidoustus AA12
The fungus A. calidoustus AA12 was finally chosen as the target strain because of its excellent antibacterial and antimalarial activities. A total of fifty Erlenmeyer flasks of the fungi AA12 were cultivated. The conditions of fermentation and extraction were the same as previously mentioned in the Section 2.3. About 37.2 g of EA crude extract was obtained, and then subjected to the general experimental procedures of compounds isolation. In this study, Silica gel GF254 plates (Qingdao Marine Chemical Ltd., Qingdao, People’s Republic of China) were used for thin-layer chromatography (TLC) analysis. Silica gel (Qingdao Marine Chemical Ltd., Qingdao, People’s Republic of China) and Sephadex LH-20 (Amersham Biosciences, Uppsala, Sweden) were applied for column chromatography (CC). High performance liquid hromatography (HPLC) analysis and separation was performed on Angilent 1260 system coupled with Angilent G1315D photodiode array detector using analytical (Agilent ZORBAX, 5 μm, 9.4 × 250 mm) and semipreparative (Agilent Eclipse XDB, 5 μm, 4.6 × 250 mm) C18 columns, respectively. The EA crude extract of fermented substances of strain AA12 was primarily subjected to vacuum liquid chromatography (VLC) on silica gel CC using step gradient elution with EA–petroleum ether (PE) (0–100%) and then with methanol (MeOH)–EA (0–100%) to yield eight fractions (Fr. 1–Fr. 8). Fr. 2 was isolated by silica gel CC (PE–EA, v/v 10:1–7:3), then applied to silica gel CC eluted by chloroform (CHCl3) –MeOH (v/v 80:1–50:1) and repeated Sephadex LH-20 CC eluting with MeOH, and further purified on HPLC (85% MeOH–H2O) to obtain compounds 1 (267 mg) and 2 (238 mg). Fr. 3 was primarily fractionated on silica gel CC (PE–EA, v:v 9:1–1:1) to afford four subfractions (Fr. 3-2–Fr. 3-4). Fr. 3-2 was subjected to silica gel CC (PE–acetone, v:v 4:1–1:1) and repeated Sephadex LH-20 CC (CHCl3-MeOH, v/v 1:1), and further purified on HPLC (87% MeOH–H2O) to afford compounds 3 (18 mg) and 4 (31 mg). Fr. 3-3 was isolated by silica gel CC (CHCl3–MeOH, v:v 50:1–40:1) to obtain compound 5 (13 mg), and then purified on HPLC using step gradient elution with MeOH–H2O (55–70%) to give compounds 6 (15 mg) and 7 (4.9 mg). The structure of compounds were elucidated by NMR spectra, which were recorded on a Bruker Avance III-400 instrument (Bruker, Faellanden, Switzerland) with TMS as an internal standard.
2.7. Scanning Electron Microscopic (SEM) Observation
According to the reported method with few modifications [16,18], a total of 400 μL logarithmic phase Streptococcus agalactiae ATCC 13813 cell suspension and 600 μL different concentrations of tested samples were added into each well of 24-well polystyrene plate (Nunc, Naperville, CA, USA). After incubation at 37°C for 10 h, the biofilms of each well was rinsed with PBS, and then fixed with glutaraldehyde for 10 h at 4°C, followed by dehydration with an increasing gradient of 30%, 50%, 70%, 90%, and 100% ethanol at 4°C for 10 min of each concentration. Finally, each processed sample was transferred to the circular slide, air dried and sputtered by coating with gold for 50 s. The SEM imaging was performed on JSM-7500F scanning electron microscope (JEOL, Tokyo, Japan).
2.8. Nucleotide Sequence Accession Number
The ITS sequences of all 93 fungal isolates obtained in this experiment were registered in GenBank. The accession numbers are PP380353–PP380390.
3. Results
3.1. Diversity of Cultivable Fungi in the Rhizosphere Communities of Three Species of Astragalus Plants
The diversity of culturable fungi in the rhizosphere communities of Astragalus acaulis (Aa), Astragalus forrestii (Af), and Astragalus ernestii (Ae) was determined at the genus and species level. A total of 93 fungal strains were isolated from the rhizosphere soil of Aa (34 isolates), Af (40 isolates) and Ae (19 isolates) based on the size, color, and other morphological observations of fungi, and identified according to the BLASTn results of ITS sequences, including 15 species of 12 genera for Aa, 19 species of 6 genera for Af, and 12 species of 9 genera for Ae (Table 1). In general, these 93 isolates belong to 38 differen species in 18 genera sharing the high similarity (98–100%) of ITS-rDNA sequences to their closest NCBI relatives (Table 1), which showed a great diversity of fungal communities in the rhizosphere of the plants Aa, Af and Ae. Furthermore, the most common trend of a decrease in biodiversity with increasing altitude observed in community ecology does not seem to occur in the rhizosphere fungal communities of Aa, Af and Ae with the elevation of 4,353, 3,225 and 4,048 m, respectively. This phenomena was also observed in our previous studyof the microbial diversity of Astragalus plants based on metagenomics [13]. As Ding et al. [13], soil physicochemical properties, plant species, the period of plant growth, and some other factors have much aggregate effect on the community structure of rhizosphere microbes.
There are four genera (Figure 1A, including Penicillium, Aspergillus, Mortierella and Trichoderma) and two species (Figure 1B, including A. fumigatus and M. alpine) commonly to the 18 genera and 38 species identified. Among them, the top two dominant genera are Penicillium and Aspergillus, which was aligned with the general idea that both genera tend to be common in soil worldwide [19,20]. Some members of these two genera are known as biocontrol fungal microbes and for their ability to produce large amounts of functional secondary metabolites with potent bioactive applications [21,22], hence justifying their highest abundance in the rhizosphere fungal communities of these three Astragalus plants growing in harsh environment. The genus Mortierella was the third most abundant genera, of which the species M. alpine, as the most dominant population, was commonly distributed in each Mortierella rhizosphere community of the three types of Astragalus. This finding was consistent with the previous research which reported the dominant distribution of the Mortierella in the soil samples of Tibet [23] and Greater Khingan Mountains [24], China. Therefore, it is reasonable to presume that the species of M. alpine may prefer to inhabit in the special high-cold environment, and herein this species can be considered as the dominant cultivable rhizofungi with distinctly eco-environmental characteristics for these three Astragalus. Meanwhile, a total of 7, 2 and 4 genera (Figure 2A and Table S1) and 10, 15, 7 species (Figure 2B and Table S2) were uniquely distributed in the rhizosphere fungi of Aa, Af and Ae. At the genus level, the rhizosphere fungi derived from Aa and Ae has better species uniqueness than those of Af. For instance, the Pseudogymnoascus roseus originally discovered in Antarctica [25] and the Samsoniella hepiali mainly from Ophiocordyceps sinensis distributing in the the plateau meadow of northwest Tibet, Qinghai, Sichuan, and Yunnan at 3,000−8,000 m of altitude [26] was only found in the rhizosphere of Aa group. This may be explained by the fact that the host plant Aa survive in the high altitude and low heat environment at an elevation of 4,353 all year round. In addition, Aporospora terricola, as the unique fungal species, was present in the Ae group, and had ever been repeatedly isolated from the the extremely arid desert nature reserve in Anxi, Gansu province of China [27]. As shown in Figure S2 [13], compared with the fertile vegetation-covered habitats of the plants Aa and Af, the plant Ae thrived in the steep slopes of flowstone beaches of 4,048 m that is a typical arid and cold mountain ecosystem, which has some similarities to that of the desert ecosystem. But in the end, there are something can not be neglected, that is, many dominant and unique fungi referred in the previously metagenomic study on the microbial diversity of rhizosphere soil of these three Astragalus plants [13] were not obtained and identified in this investigation, indicating that the selection of culture medium and conditions for these fungal microbes should be more diverse and specific.
3.2. Anti-Infectious Potential of Isolated Fungi
All 93 fungal isolates were fermented on a small scale in solid rice medium and the EA crude extracts of their fermented substances were evaluated for antibacterial and antimalarial activities. In total, ten extracts showed weak to strong antibacterial activity against seven Gram-positive pathogenic bacteria with MIC values ranging from 100 to 6.25 µg/mL (Table 2), and among which three extracts could inhibit the growth of 80-99% Plasmodium falciparum 3D7 at the concentration of 50 µg/mL (Table 3). Of these, ten bioactive fungal strains, about 70% species are belonged to the genera Aspergillus and Penicillium that is the dominant flora in the cultivated fungal communities of Aa, Af and Ae rhizosphere. Although many species of these two genera exist extensively in the nature and have been well explored as potential medicinal resources for producing novel and bioactive natural products [21,22], there are few reports on the detailed research of natural products of A. calidoustus, A. tabacinus, P. glabrum, and P. vasconiae. In addition, to our best knowledge, the study on the secondary metabolites of the other three anti-infective species of Cordyceps farinose, Lecanicillium aphanocladii and Umbelopsis nana has so far not been reported. These results of the primary anti-infective screening of fungi and the related literature investigation suggested that some fungal strains affiliating to the genera of Aspergillus, Penicillium, Cordyceps, Lecanicillium and Umbelopsis derived from these three Astragalus rhizosphere might be a potent source of anti-infective natural products.
Most notably, the EA crude extract of Aspergillus calidoustus AA12 fermented substance isolated from the plant Aa rhizosphere showed both excellent antibacterial and antimalarial activities. Specifically, the extract of strain AA12 revealed the strongest antibacterial against methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300, S. aureus ATCC 25923, and Enterococcus faecalis ATCC 29212 with the MIC values of 6.25 µg/mL, and showed significant inhibition for S. epidermidis ATCC 12228, vancomycin-resistant Enterococcus faecalis (VREF) ATCC 51299 and E. faecium ATCC 35667 with the MIC values of 12.5 µg/mL, and Streptococcus agalactiae ATCC 13813 with the MIC value of 25 µg/mL. Also, this extract showed the best antimalarial activity against Plasmodium falciparum 3D7 with the inhibitory rate reaching up to 99% under the concentration of 50 μg/mL at 72 h. The species of A. calidoustus was firstly proposed as a new species in Aspergillus section Usti by Varga et al. [28]. Valiante et al. [29] suggested that austinoid gene clusters involved in the biosynthesis of fungus A. calidoustus should lead to the production of various austinoid-type meroterpenoids. Mo et al. [30] and Zhang et al. [31] found that the isolates of A. calidoustus from the wetland soil of Dianchi Lake in Kunming City and East Lake in Wuhan City could be able to produce meroterpenoids and drimane sesquiterpenoids, respectively, which are a prominent group of natural products and have good biological and chemical diversity [32]. In this study, the strain A. calidoustus AA12 was eventually selected as the target fungus for further exploitation of anti-bacterial and anti-malarial natural products according to its prominent anti-infectious activity and chemical abundance (the HPLC spectra of the EA crude extracts of strain AA12 and other nine anti-infective fungal strains as shown in Figure S3).
3.3. Structural Elucidation and Anti-Infective Activity of the Isolated Natural Products
In total, seven compounds were isolated from Aspergillus calidoustus AA12, including 6-epi-ophiobolin G (1) [33], penicisochroman A (2) [34], pergillin (3) [35], 7-methyl-2-(1-methylethylethlidene)-furo[3,2-H]isoquinoline-3-one (4) [36], trypacidin (5) [37], 1,2-seco-trypacidin (6) [38] and questin (7) [39] (Figure 2), respectively, by comparing their spectroscopic data with the available literature (the 1H and 13C NMR spectra and data of these seven compounds as shown in Figures S4–S10 and Table S3–S5, respectively). As shown in Figure 2, the chemical structure types of these compounds comprises sesterterpene (1), sesquiterpenoids (2-4) and polyketides (5-7). In this study, all these seven compounds were firstly discovered from the species of A. calidoustus, which indicated that the fungus A. calidoustus AA12 might have a huge biosynthetic capacity to produce structural diverse natural products besides the previous-mentioned austinoid-type meroterpenoids [29,30] and drimane sesquiterpenoids [31]. Moreover, herein terpenoids 1-4 obtained from A. calidoustus of the plant Astragalus acaulis rhizosphere surviving in snow mountai at high elevations up to 4,353 m was previously reported as the main secondary metabolites of marine or plant rhizosphere-derived fungi [33,34,36], suggested that these sesterterpenes and sesquiterpenoids should be more likely to be biosynthesized by those fungal microorganisms in special eco-environment.
The antibacterial activity of compounds 1–7 were all evaluated. As shown in Table 4, all compounds displayed weak to strong antibacterial activity against seven strains of Gram-positive pathogens at MIC values in the range of 6.25 to 100 µg/mL except the 1,2-seco-trypacidin (6). Notably, the sesterterpene 6-epi-ophiobolin G (1) exhibited a broad-spectrum antibacterial activity against all pathogens tested with MIC values ranging from 6.25 to 50 µg/mL, and showed the most obvious inhibitory effect on the growth of Streptococcus agalactiae ATCC 13813 at the concentration of 6.25 µg/mL. As the SEM imaging observed (Figure 3), after treated with 1/2×MIC (3.125 µg/mL) of compound 1, a collapse of the S. agalactiae biofilm architecture performed that small cell clusters and short chains were loosely attached to the surface of glass slides with little debris. The further drastic damages on the S. agalactiae biofilm was obviously observed as the phenomenon that the total S. agalactiae biofilm biomass was significantly reduced when treated with compound 1 at the MIC value of 6.25 μg/mL. Bacterial biofilm is well known as one of contributing factors of favoring the growth of more bacteria resistant to antibiotic [9,16,18]. The pathogenic S. agalactiae poses a serious threat to high-risk groups such as pregnant women, newborns, and the elderly, causing severe clinical illnesses and increasing morbidity and mortality rates, part of which have become increasingly resistant to the second-line antibiotics like erythromycin and clindamycin [40]. Nowadays, the clinical treatment of S. agalactiae is more challenging with the emergence of penicillin-resistant strains [41]. Thus the remarkable disintegration efficacy of 6-epi-ophiobolin G (1) against S. agalactiae biofilm indicated that this compound might have potentially valuable for the utilization of controlling the drug-resistant properties of S. agalactiae.
Although antimalarial activity of compounds 1-7 against strains 3D7 and chloroquine-resistant Dd2 of Plasmodium falciparum was all detected, only the sesterterpene 6-epi-ophiobolin G (1) showed good antimalarial activity against strains 3D7 and Dd2 with IC50 values of 3.319 and 4.340 µmol/L at 72 h, respectively (Figure 4). In addition, as shown in Figure 5, compound 1 could effectively inhibit the period of Plasmodium trophozoites in order to achieve its excellent antiplasmodial effect.
In this study, 6-epi-ophiobolin G (1), as the most potent anti-infective sesterterpene compound isolated from Aspergillus calidoustus AA12, belongs to ophiobolins, which are a prominent group of natural products produced by the fungi derived from pathogenic plant [42,43], mangrove [44], and marine sediments [45]. This compound was successively isolated from the fungus Emericella variecolor GF10 derived from marine sediment collected at 70 m depth in the Gokasyo Gulf, Mie Prefecture, Japan [45], and the deep-sea derived fungus Aspergillus sp. WHU0154 at 3197 m depth, South China Sea [33], and only evaluated for its anti-inflammatory effect [33]. Therefore, it is the first report that the ophiobolin-type was isolated from the fungal microbe in high-cold terrestrial environment of snow mountain, and proved to have potent anti-infective activity against the biofilm of Streptococcus agalactiae ATCC 13813, Plasmodium falciparum 3D7 and chloroquine-resistant P. falciparum Dd2. These results will certainly do some inspiring action to the development and exploitation of ophiobolin-type sesterterpenoids with antibacterial and antimalarial activities against those drug-resistant strains.
4. Conclusions
This is the first report on the species diversity cultivable rhizofungi derived from Astragalus plants in special high-cold environment and their antibacterial and antimalarial potential. The fungal communities in the rhizosphere of A. acaulis, A. forrestii, and A. ernestii had a rich diversity and uniqueness, the microbial distribution and structure of which was influenced by a complex mixture of some factors such as the altitude, soil physicochemical property, plant species, and so on. Some fungal strains of Aspergillus and Penicillium displayed good anti-infective activity and chemical diversity, which not only might play an important role in helping their host Astragalus to grow healthy and enhance their resilience and adaptability in extremely harsh environment of high altitudes, but also provide a rich resource for our further research of bioactive natural products from Astragalus rhizofungi. Interestingly enough, all compounds of sesterterpine (1), sesquiterpenes (2-4) and polyketides (5-7) were firstly discovered in the species of Aspergillus calidoustus indicating the potent production of diverse chemical structural secondary metabolites of A. calidoustus AA12. Meanwhile, herein the broad-spectrum antibacterial activity and excellent antimalarial activity of 6-epi-ophiobolin G (1) was firstly proposed, which may be valuable, to some extent, for further exploring and utilizing anti-infectious natural products from characteristic microbes in special high-cold environment.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Map of sampling sites in the northwestern area of Yunnan province, China (A). In the picture B, the green dot is sampling site for Astragalus acaulis (Aa) growing in high altitude grassland of 4,353 m; as shown in the yellow dot, A. ernestii (AE) grows in the steep slopes of flowstone beaches of 4,048 m; The blue dot shows the A. forrestii (Af) survival environment, surrounded by more pine trees and deciduous foliage of 3,225 m; Figure S2: The plants of Astragalus acaulis (Aa) grows in alpine grassland (A), A. forrestii (Af) is surrounded by more pine trees and deciduous foliage (B), A. ernestii (Ae) grows in the steep slopes of flowstone beaches (C); Figure S3: HPLC spectra of the crude ethyl acetate (EA) extracts of fermented substrates of Aspergillus calidoustus AA12 (A③) and other nine target fungal strains of A. fumigatus AA15 (A②), A. tabacinus AF16 (B), Penicillium polonicum AA17 (C(②), Penicillium vasconiae AF5 (D), P. glabrum AF3-1 (E), P. brevicompactum AE7 (F), Cordyceps farinosa AA8 (G), Lecanicillium aphanocladii AF15 (H), and Umbelopsis nana AF31 (I); Figures S4–S10 The NMR spectrum of compounds 1–7; Table S1: A total of 7, 2 and 4 unique fungal genera identified in the rhizosphere fungal community of Astragalus acaulis (Aa), A. forrestii (Af) and A. ernestii (Ae), respectively; Table S2: A total of 10, 15 and 7 unique fungal species identified in the rhizosphere fungal community of Astragalus acaulis (Aa), A. forrestii (Af) and A. ernestii (Ae), respectively; Tables S3–S5: 1H and 13C NMR data of compounds 1–7 (δ in ppm).
Author Contributions
Conceptualization, G-J. Z. and K-L. W.; Data curation, G-J. Z., W-J. X., W.X. Z-R. D. and B-C. L.; Formal analysis, G-J. Z., W-J. X. and Z-R. D.; Funding acquisition, H-F. Li., Y-Z. Z. and K-L. W.; Investigation, G-J. Z., Z-R. D., Y-Z. Z. and B. J.; Methodology, W-J. X., W. X., Z-R. D., B-C. L., X-L. L. and H. D.; Resources, K-L. W.; Writing – original draft, G-J. Z.; Writing – review & editing, H-F. L., B. J. and K-L. W.
Funding
This research was supported by the National Natural Science Foundations of China (grant nos. 32060032, 82360708 and 32300181), the Natural Science Foundation of Yunnan Province of China (grant no. 202301AT070157) and the Expert Workstation of Jiang Yong Yunnan Province (grant No. 202305AF150048).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data of cultivable fungal strains in this study were uploaded to the NCBI database, and the accession number was PP380353-PP380390.
Acknowledgments
The authors acknowledge that Prof. Cai-Juan Zheng and Guo-Lei Huang in the Key Laboratory of Tropical Medicinal Resource Chemistry of Ministry of Education, Hainan Normal University, Haikou, China, provided great support for SEM imaging experiment.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: the microbial ecology of the rhizosphere. Nat Rev Microbiol 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Chen, D.; Zeng, J.; Wan, X.; Wang, Y.; Lan, S.; Zou, S.; Qian, X. Variation in community structure of the root-associated fungi of Cinnamomum camphora forest. J Fungi 2022, 8, 1210. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Yang, W.; Wang, R.; Zhang, H.; Meng, J.; Jing, H.; Zhang, G.; Wang, J.; Guo, L.; Gao, W. ZnO-S.cerevisiae: an effective growth promoter of Astragalus memeranaceus and nano-antifungal agent against Fusarium oxysporum. Chem Eng J 2024, 486, 149958. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, F.X.; Zeng, Z.; Xu, M.; Sun, F.; Yang, L.; Bi, X.; Lin, Y.; Gao, Y.; Hao, H.X.; Yi, W.; Li, M.; Xie, Y. Advances in metagenomics and its application in environmental microorganisms. Front Microbiol 2021, 12, 766364. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, A.R. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar]
- Centers for Disease Control and Prevention. Antibiotic resistance threats in the United States, 2019; Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; Ouellette, M.; Outterson, K.; Patel, J.; Cavaleri, M.; Cox, E.M.; Houchens, C.R.; Grayson, M.L.; Hansen, P.; Singh, N.; Theuretzbacher, U.; Magrini, N. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.; Zeng, Y. Novel bioactive natural products from marine-derived Penicillium fungi: A Review (2021-2023). Mar Drugs 2024, 22, 191. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.; Dou, Z.R.; Gong, G.F; Li, H.F.; Jiang, B.; Xu, Y. Anti-larval and anti-algal natural products from marine microorganisms as sources of anti-biofilm agents. Mar Drugs 2022, 20, 90. [Google Scholar] [CrossRef]
- Wang, X.; Ji, G.; Cun, J.; Xu, P.; Wang, X.; Ren, G.; Li, W. Screening of insecticidal and antifungal activities of the culturable fungi isolated from the intertidal zones of Qingdao, China. J Fungi 2022, 8, 1240. [Google Scholar] [CrossRef]
- Yang, X.; Yu, H.; Ren, J.; Cai, L.; Xu, L.; Liu, L. Sulfoxide-containing bisabolane sesquiterpenoids with antimicrobial and nematicidal activities from the marine-derived fungus Aspergillus sydowii LW09. J Fungi 2023, 9, 347. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, L.; Gao, C.; Chen, W.; Vong, C.; Yao, P.; Yang, Y.; Li, X.; Tang, X.; Wang, S.; Wang, Y. Astragali Radix (Huangqi): a promising edible immunomodulatory herbal medicine. J Ethnopharmacol 2020, 258, 112895. [Google Scholar] [CrossRef]
- Ding, J.J.; Zhou, G.J.; Chen, X.J.; Xu, W.; Gao, X.M.; Zhang, Y.Z.; Jiang, B.; Li, H.F.; Wang, K.L. Analysis of microbial diversity and community structure of rhizosphere soil of three Astragalus species grown in special high-cold environment of northwestern Yunnan, China. Microorganisms 2024, 12, 539. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.R.; Zhang, F.; Zhou, G.J.; Gong, G.F.; Ding, J.J.; Fan, X.H.; Jiang, B.; Wang, K.L. Varicosporellopsis shangrilaensis sp. nov. (Nectriaceae, Hypocreales), a new terricolous species isolated from the rhizosphere soil of Astragalus polycladus in northwestern Yunnan. Phytotaxa, 2023; 600, 230–238. [Google Scholar]
- Liu, A.; Li, Y.; Wang, Q.; Zhang, X.; Xiong, J.; Li, Y.; Lei, Y.; Sun, Y. Analysis of microbial diversity and community structure of rhizosphere soil of Cistanche salsa from different host plants. Front Microbiol 2022, 13, 971228. [Google Scholar] [CrossRef]
- Song, Z.M.; Zhang, J.L.; Zhou, K.; Yue, L.M; Zhang, Y.; Wang, C.Y.; Wang, K.L.; Xu, Y. Anthraquinones as potential antibiofilm agents against methicillin-resistant Staphylococcus aureus. Front Microbiol 2021, 12, 709826. [Google Scholar] [CrossRef]
- Gudla, C.S.; Selvam, V.; Selvaraj, S.S.; Tripathi, R.; Joshi, P.; Shaham, S.H.; Singh, M.; Shandil, R.K.; Habib, S.; Narayanan, S. Novel baicalein-derived inhibitors of Plasmodium falciparum. Pathogens 2023, 12, 1242. [Google Scholar] [CrossRef]
- Yin, Q.; Liang, J.; Zhang, W.; Zhang, L.; Hu, Z.-L.; Zhang, Y.; Xu, Y. Butenolide, a marine derived broad-spectrum antibiofilm agent against both Gram-positive and Gram-negative pathogenic bacteria. Mar Biotechnol 2019, 21: 88–98.
- Krijgsheld, P.; Bleichrodt, R.; Veluw, G.J.; Wang, F.; Müller, W.H.; Dijksterhuis, J.; Wösten, H.A.B. Development in Aspergillus. Stud Mycol 2012, 74, 1–29. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozaloa, B.; Quero, J.L.; García-Gómez, M.; Gallardof, A.; Ulrichg, W.; Bowkerh, M.A.; Arredondoi, T.; Barraza-Zepedaj, C.; Brank, D.; Florentinol, A.; Gaitánm, J; Gutiérrezj, J.R.; Huber-Sannwaldi, E.; Jankjup, M.; Mauq, R.L.; Miritir, M.; Naserip, K.; Ospinal, A.; Stavis, I.; Wangt, D.; Woodsr, N.N.; Yuant, X.; Zaadyu, E.; Brajesh K. Singhb, BK. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc Natl Acad Sci USA 2015, 112, 15684–15689.
- Zhao, D.L.; Han, X.B.; Wang, M.; Zeng, Y.T.; Li, Y.Q.; Ma, G.Y.; Liu, J.; Zheng, C.J.; Wen, M.X.; Zhang, Z.F; Zhang, P.; Zhang, C.S. Herbicidal and antifungal xanthone derivatives from the alga-derived fungus Aspergillus versicolor D5. J Agric Food Chem 2020, 68, 11207–11214. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, J.; Mei, J.; Jiang, R.; Tu, S.; Deng, H.; Liu, J.; Yang, S.; Li, J. Origins, structures, and bioactivities of secondary metabolites from marine-derived Penicillium fungi. Mini Rev Med Chem 2021, 21, 2000–2019. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Pu-BU, R.C.; Lv, M.L.; Wang, M.; Liu, X.Y. Species diversity of Zygomycotan fungi in the Tibet Autonomous Region. Microbiology China 2018, 45, 1250–1261. [Google Scholar]
- Lv, M.L.; Liu, Z.; Song, Z.; Wang, Y.N.; Liu, X.Y. Diversity and distribution of culturable Mucoromycota fungi in the Greater Khinggan Mountains, China. Biodiversity Science 2019, 27, 821–837. [Google Scholar]
- Minnis, A.M.; Lindner, D.L. Phylogenetic evaluation of geomyces and allies reveals no close relatives of Pseudogymnoascus destructans, comb. nov., in bat hibernacula of eastern North America. Fungal Biol 2013, 117, 638–649.
- Meng, Y,; Tang, H. Y.; Shi, J.D.; Wang, D. Three fungal strains isolated from stroma of Ophiocordyceps sinensis and their culture conditions. Mycosystema 2021, 40, 1991–2007.
- Zhang, L.L. Species diversity and drought resistance of dark septate endophytes in the rhizospheres of different plants in Anxi extreme arid desert. Master’s dissertation, Hebei university, Hebei, Baoding, June 9, 2018.
- Varga, J.; Houbraken, J.; Van-Der-Lee, H.A.; Verweij, P.E.; Samson, R.A. Aspergillus calidoustus sp. nov., causative agent of human infections previously assigned to Aspergillus ustus. Eukaryot Cell 2008, 7, 630–638.
- Valiante, V.; Mattern, D.J.; Schüffler, A.; Horn, F.; Walther, G.; Scherlach, K.; Petzke, L.; Dickhaut, J.; Guthke, R.; Hertweck, C.; Nett, M.; Thines, E.; Brakhage, A.A. Discovery of an extended austinoid biosynthetic pathway in Aspergillus calidoustus. ACS Chem Biol 2017, 12, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Yin, J.; Ye, Z.; Li, F.; Lin, S.; Zhang, S.; Yang, B.; Yao, J.; Wang, J.; Hu, Z.; Zhang, Y. Asperanstinoids A-E: undescribed 3,5-dimethylorsellinic acid-based meroterpenoids from Aspergillus calidoustus. Phytochemistry 2021, 190, 112892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Mo, S.; Li, F.; Zhang, Y.; Wang, J.; Hu, Z.; Zhang, Y. Drimane sesquiterpenoids from a wetland soil-derived fungus Aspergillus calidoustus TJ403-EL05. Nat Prod Bioprospect 2022, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- He, J.L.; Chen, C.J.; Liu, Y.H.; Gao, C.H.; Wang, R.P.; Zhang, W.F.; Bai, M. Austin-Type Meroterpenoids from fungi reported in the last five decades: a review. J Fungi 2024, 10, 162. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Uvarani, C.; Wang, F.; Xue, Y.; Wu, N.; He, L.; Tian, D.; Chen, M.; Zhang, Y.; Hong. K.; Tang, J. New Ophiobolins from the deep-sea derived fungus Aspergillus sp. WHU0154 and their anti-inflammatory effects. Mar Drugs 2020, 18, 575.
- Trisuwan, K, Rukachaisiriku, V.; Sukpondma, Y.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Furo[3,2-h]isochroman, furo[3,2-h]isoquinoline, isochroman, phenol, pyranone,and pyrone derivatives from the sea fan-derived fungus Penicillium sp. PSU-F40. Tetrahedron 2010, 4484–4489.
- Kuramochi, K.; Tsubaki, K. Synthesis and structural characterization of natural benzofuranoids. J Nat Prod 2015, 78, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, A.; Murakami, C.; Kamisuki, S.; Kuriyama, I.; Yoshida, H.; Sugawara, F.; Mizushina, Y. Pseudodeflectusin, a novel isochroman derivative from Aspergillus pseudodeflectus a parasite of the sea weed, Sargassum fusiform, as a selective human cancer cytotoxin. Bioorg Med Chem Lett 2004, 14, 3539–3543. [Google Scholar] [CrossRef]
- Xu, X.; Guo, S.; Chen, H.; Zhang, Z.; Li, X.; Wang, W.; Guo, L. Bioassay-guided isolation and characterization of antibacterial compound from Aspergillus fumigatus HX-1 associated with Clam. 3 Biotech 2021, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Newaz, A.W.; Yong, K.; Yi, W.; Wu, B.; Zhang, Z. Antimicrobial metabolites from the indonesian mangrove sediment-derived fungus Penicillium chrysogenum sp. ZZ1151. Nat Prod Res 2023, 37, 1702–1708. [Google Scholar] [CrossRef]
- Guo, L.; Wang, L.; Li, X.; Xu, X.; Guo, J.; Wang, X.; Yang, W.; Xu, F.; Li, F. Enhanced production of questin by marine-derived Aspergillus flavipes HN4-13. 3 Biotech 2020, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Paveenkittiporn, W.; Ungcharoen, R.; Kerdsin, A. Streptococcus agalactiae infections and clinical relevance in adults, Thailand. Diagn Microbiol Infect Dis 2020, 97, 15005. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Moreno, M.; Trampuz, A.; Di-Luca, M. Synergistic antibiotic activity against planktonic and biofilm-embedded Streptococcus agalactiae, Streptococcus pyogenes and Streptococcus oralis. J Antimicrob Chemother 2017, 72, 3085–3092. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Miao, F.P.; Ming-Feng Qiao, M.F.; Robert, H.; Ji, N.Y. Terretonin, ophiobolin, and drimane terpenes with absolute configurations from an algiconus Aspergillus ustus. RSC Advances 2013, 3, 588–595. [Google Scholar] [CrossRef]
- Arai, M.; Niikawa, H.; Kobayashi, M. Marine-derived fungal sesterterpenes, ophiobolins, inhibit biofilm formation of Mycobacterium species. J Nat Med 2013, 67, 271–275. [Google Scholar] [CrossRef]
- Cai, R.; Jiang, H.; Mo, Y.; Guo, H.; Li, C.; Long, Y.; Zang, Z.; She, Z. Ophiobolin-type sesterterpenoids from the mangrove endophytic fungus Aspergillus sp. ZJ-68. J Nat Prod. 2019, 82, 2268–2278.
- Wei, H.; Itoh, T.; Kinoshita, M.; Nakai, Y.; Kurotaki, M.; Kobayashi, M. Cytotoxic sesterterpenes, 6-epi-ophiobolin G and 6-epi-ophiobolin N, from marine derived fungus Emericella variecolor GF10. Tetrahedron 2004, 60, 6015–6019. [Google Scholar] [CrossRef]
Figure 1.
Venn diagram showing the number of fungi at genus (A) and species (B) level in the rhizosphere soil samples of Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae). Each circle, with a different color in the diagram, represents the number of genera and species specific to the corresponding subgroup. Middle core numbers represent the number of genera and species commonly to all groups.
Figure 1.
Venn diagram showing the number of fungi at genus (A) and species (B) level in the rhizosphere soil samples of Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae). Each circle, with a different color in the diagram, represents the number of genera and species specific to the corresponding subgroup. Middle core numbers represent the number of genera and species commonly to all groups.
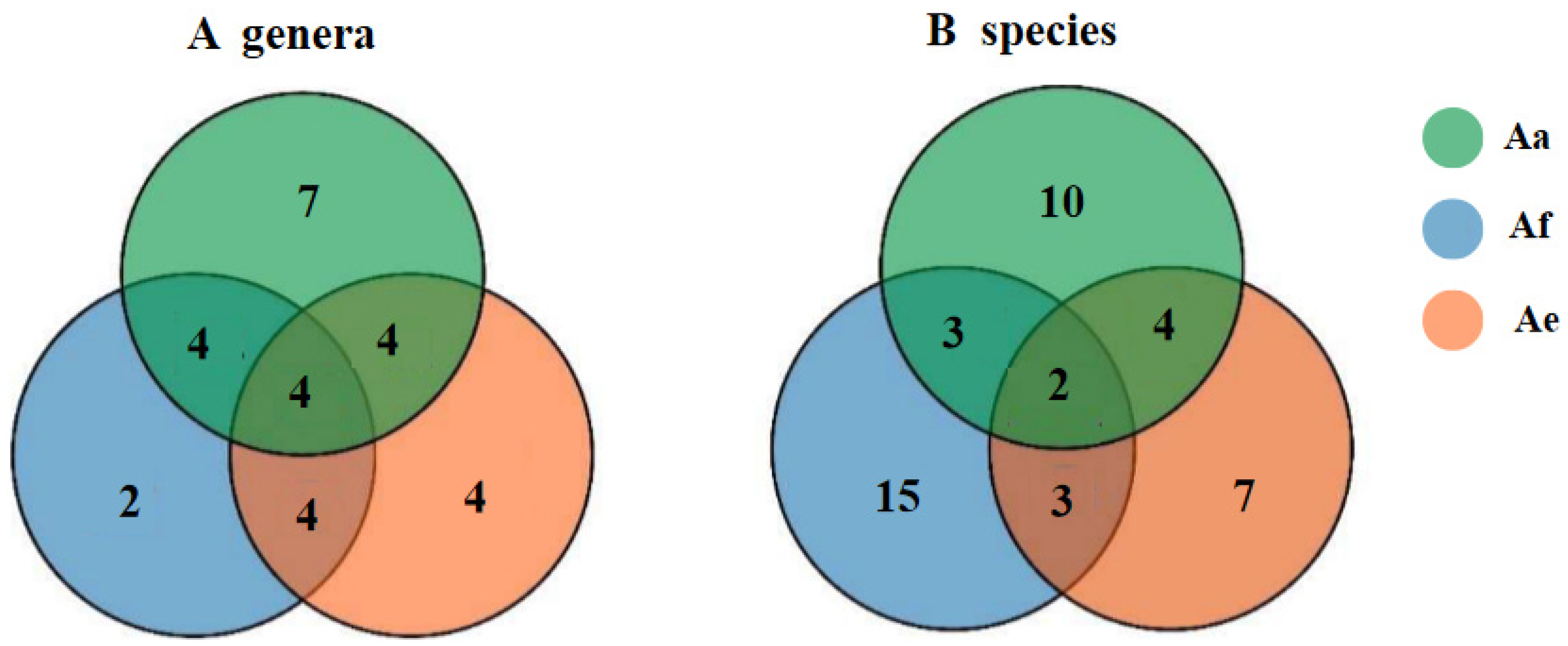
Figure 2.
Chemical structures of compounds 1–7.
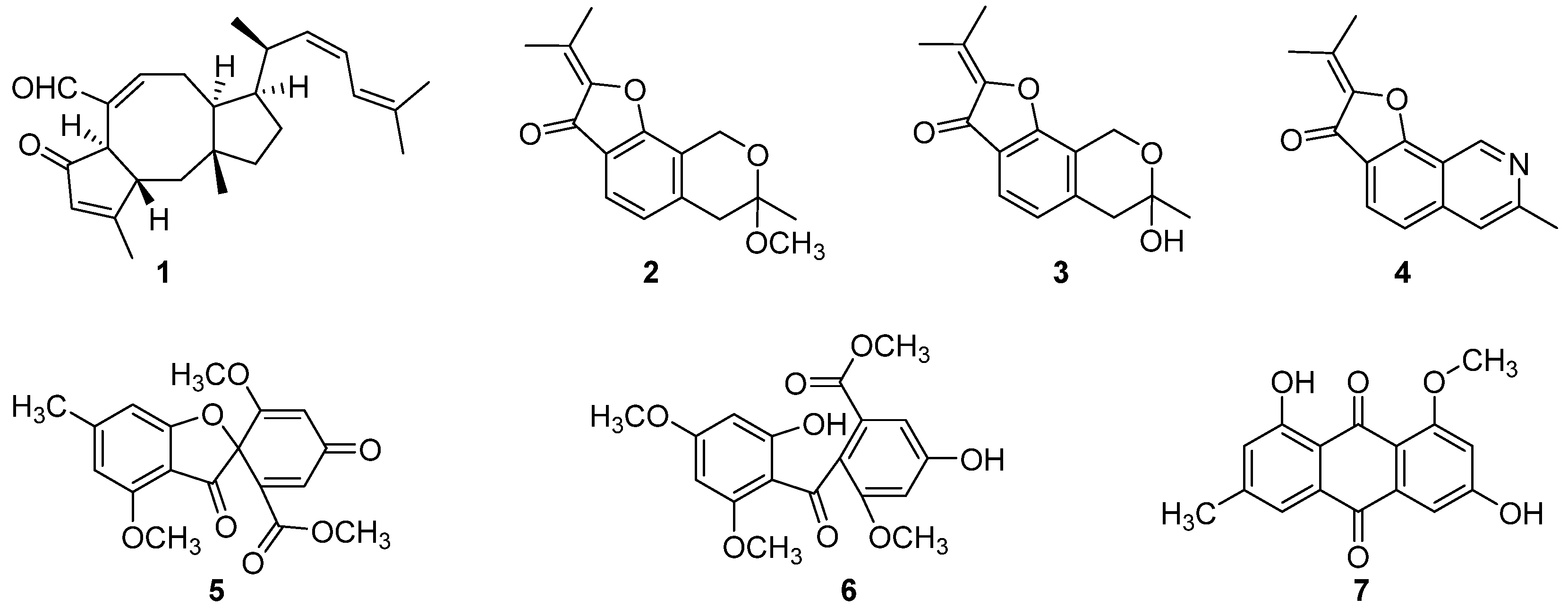
Figure 3.
Scanning electron micrographs (SEM) of 24 h preformed Streptococcus agalactiae ATCC 13813 biofilms treated with different concentrations of 6-epi-ophiobolin G (1). (A1–C2) Different treatment groups (A1, A2 DMSO control; B1, B2 treated with 1/2 × MIC =3.125 μg/mL of compound 1; C1, C2, treated with MIC = 6.25 μg/mL of compound 1). Series 1 and 2 were different magnifications with ×7,500 and ×15,000, respectively.
Figure 3.
Scanning electron micrographs (SEM) of 24 h preformed Streptococcus agalactiae ATCC 13813 biofilms treated with different concentrations of 6-epi-ophiobolin G (1). (A1–C2) Different treatment groups (A1, A2 DMSO control; B1, B2 treated with 1/2 × MIC =3.125 μg/mL of compound 1; C1, C2, treated with MIC = 6.25 μg/mL of compound 1). Series 1 and 2 were different magnifications with ×7,500 and ×15,000, respectively.

Figure 4.
Determination of antimalarial activity of 6-epi-ophiobolin G (1) at 72 h against Plasmodium falciparum 3D7 (A) and chloroquine-resistant P. falciparum Dd2 (B).
Figure 4.
Determination of antimalarial activity of 6-epi-ophiobolin G (1) at 72 h against Plasmodium falciparum 3D7 (A) and chloroquine-resistant P. falciparum Dd2 (B).
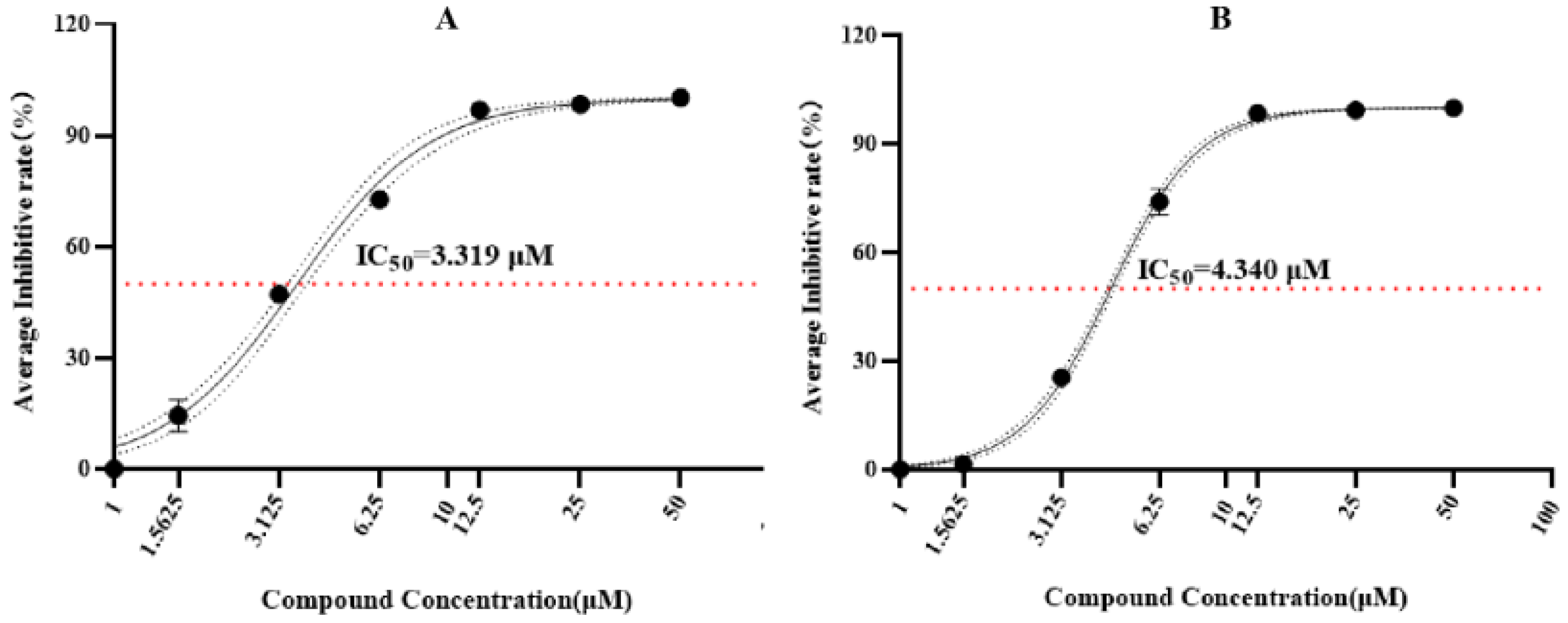
Figure 5.
Blood smear of Plasmodium falciparum 3D7 and chloroquine-resistant P. falciparum Dd2 treated by different concentrations of 6-epi-ophiobolin G (1). The pictures of A1-A3 belong to the P. falciparum 3D7 tested group including DMSO control (A1), and treated with 50 µmol/L (A2) and 25 µmol/L (A3) of compound 1. The pictures of B1-B3 belong to the chloroquine-resistant P. falciparum Dd2 tested group including DMSO control (B1), and treated with 50 µmol/L (B2) and 25 µmol/L (B3) of compound 1. I and II represent the periods of trophozoite and schizont of Plasmodium parasites, respectively.
Figure 5.
Blood smear of Plasmodium falciparum 3D7 and chloroquine-resistant P. falciparum Dd2 treated by different concentrations of 6-epi-ophiobolin G (1). The pictures of A1-A3 belong to the P. falciparum 3D7 tested group including DMSO control (A1), and treated with 50 µmol/L (A2) and 25 µmol/L (A3) of compound 1. The pictures of B1-B3 belong to the chloroquine-resistant P. falciparum Dd2 tested group including DMSO control (B1), and treated with 50 µmol/L (B2) and 25 µmol/L (B3) of compound 1. I and II represent the periods of trophozoite and schizont of Plasmodium parasites, respectively.
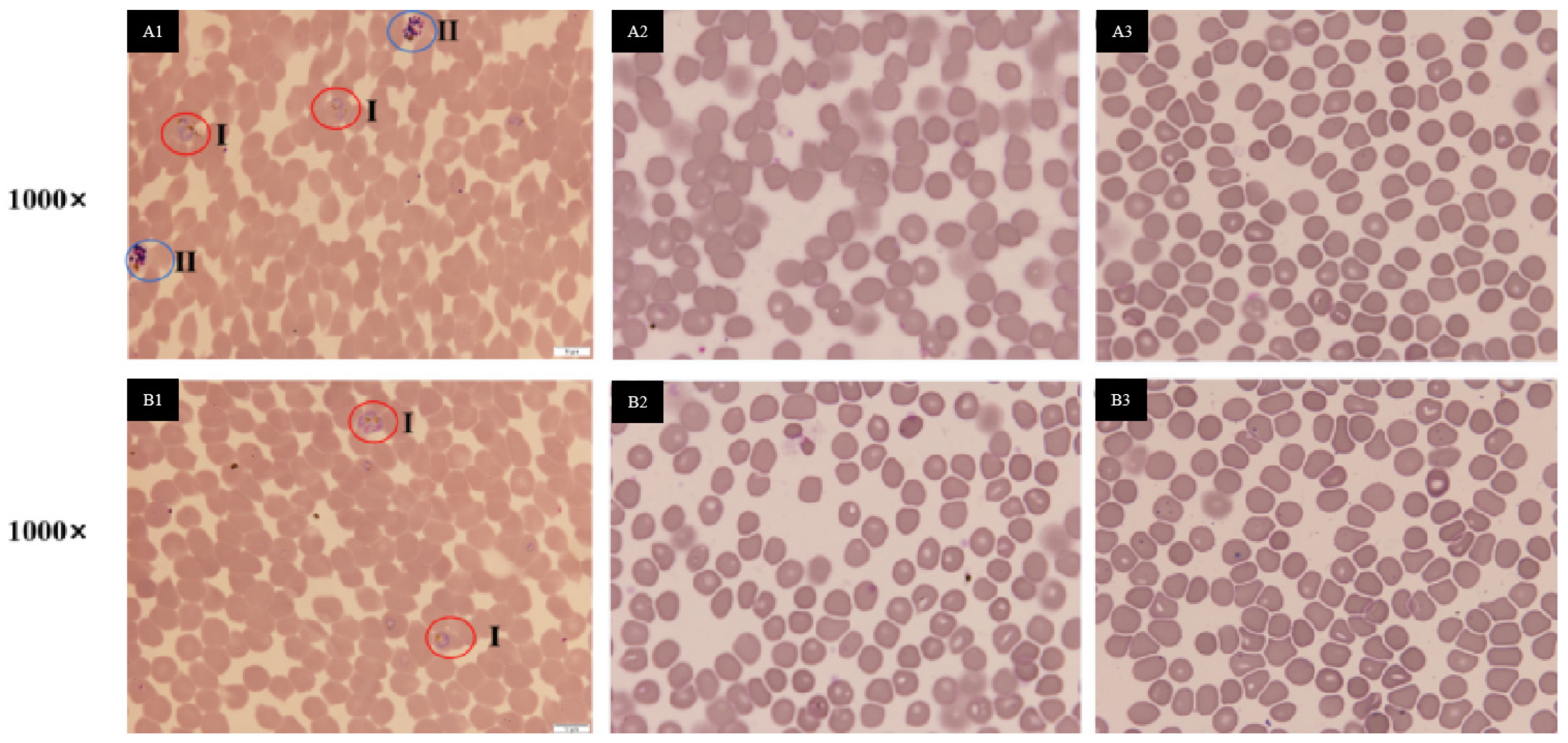

Table 1.
Diversity of the cultivable rhizosphere fungi from Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae). A total of 93 fungal strains belonging to 38 species in 18 genera were identified by comparison their internal transcribed spacer (ITS) sequences of the isolates with their reference strains in the GeneBank of NCBI. All these isolates have high identity percentage values with 98–100%.
Table 1.
Diversity of the cultivable rhizosphere fungi from Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae). A total of 93 fungal strains belonging to 38 species in 18 genera were identified by comparison their internal transcribed spacer (ITS) sequences of the isolates with their reference strains in the GeneBank of NCBI. All these isolates have high identity percentage values with 98–100%.
| Species | Representative Isolates (Accession Number in GenBank |
Similarity | Number of Fungal Isolates | ||
|---|---|---|---|---|---|
| Aa | Af | Ae | |||
| Penicillium wellingtonense | AA3(JN617713) | 100.00% | 2 | ||
| P. polonicum | AA17(OK510248) | 99.19% | 1 | 2 | |
| P. glabrum | AE3-1(OP179016) | 99.49% | 1 | 2 | |
| brevicompactum | AE7(MT558924) | 100.00% | 1 | ||
| P. vasconiae | AF5(OQ870819) | 98.86% | 8 | ||
| P. suaveolens | AF6(MH864249) | 99.48% | 1 | ||
| P. thomii | AF19(OR574374) | 99.58% | 1 | ||
| Aspergillus calidoustus | AA12(MZ267040) | 100.00% | 3 | ||
| A. fumigatus | AA15(MG991598) | 100.00% | 2 | 9 | 2 |
| A. niger | AE10(OM841707) | 99.74% | 1 | ||
| A. versicolor | AF2-1(MT582751) | 100.00% | 1 | ||
| A. tabacinus | AF16(OK011797) | 98.68% | 2 | ||
| Mortierella alpina | AA10(MT366045) | 100.00% | 8 | 2 | 3 |
| M.clonocystis | AA2(LC515184) | 100.00% | 3 | ||
| M. minutissima | AF9(MT072066) | 98.93% | 1 | ||
| M. verticillata | AF20(MT028128) | 99.50% | 1 | ||
| Truncatella angustata | AA1-1(MT514384) | 100.00% | 3 | ||
| Cordyceps farinosa | AA8(MF494607) | 99.82% | 3 | ||
| Exophiala tremulae | AA21(NR159874) | 98.84% | 1 | ||
| Beauveria pseudobassiana | AA4-1(KT368174) | 100/00% | 1 | ||
| Samsoniella hepiali | AA16(OL684609) | 98.91% | 2 | ||
| Cladosporium cladosporioides | AA2-1(ON970159) | 100.00% | 1 | 2 | |
| Neonectria radicicola | AA1(AJ875331) | 100.00% | 1 | ||
| Trichoderma atroviride | AA14(KX379158) | 99.63% | 1 | 3 | |
| T. paraviridescens | AF8(MN900599) | 100.00% | 1 | ||
| T. koningiopsis | AF23(KU645324) | 99.83% | 2 | ||
| T. viride | AF29(KU202222) | 98.71% | 1 | ||
| T. longipile | AF30(AY737763) | 100.00% | 2 | ||
| longibrachiatum | AE8(KU945846) | 99.71% | 1 | ||
| Pseudogymnoascus roseus | AA4(KJ755524) | 100.00% | 2 | ||
| Leptosphaeria sclerotioides | AE3(OR782803) | 99.76% | 2 | ||
| Fusarium solani | AE4-1(KT192216) | 98.98% | 1 | ||
| Nemania diffusa | AE4(MK336457) | 99.21% | 1 | ||
| Aporospora terricola | AE1(KC292841) | 98.47% | 1 | ||
| Lecanicillium aphanocladii | AF15(OR752316) | 99.32% | 1 | ||
| Umbelopsis vinacea | AF14(KT354998) | 99.34% | 1 | ||
| U. ramanniana | AF26 (GU934569) | 98.21% | 1 | ||
| U. nana | AF31(OR724064) | 100.00% | 1 | ||
Table 2.
Antibacterial screening of rhizosphere fungi associated with Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae) against seven strains of Gram-positive pathogenic bacteria, including methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300, S. aureus ATCC 25923, S. epidermidis ATCC 12228, vancomycin-resistant Enterococcus faecalis (VREF) ATCC 51299, E. faecalis ATCC 29212, E. faecium ATCC 35667 and Streptococcus agalactiae ATCC 13813 (µg/mL).
Table 2.
Antibacterial screening of rhizosphere fungi associated with Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae) against seven strains of Gram-positive pathogenic bacteria, including methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300, S. aureus ATCC 25923, S. epidermidis ATCC 12228, vancomycin-resistant Enterococcus faecalis (VREF) ATCC 51299, E. faecalis ATCC 29212, E. faecium ATCC 35667 and Streptococcus agalactiae ATCC 13813 (µg/mL).
| Fungal Strain | ATCC 43300 | ATCC 25923 | ATCC 12228 | ATCC 51299 | ATCC 29212 | ATCC 35667 | ATCC 13813 |
|---|---|---|---|---|---|---|---|
| Aspergillus calidoustus AA12 | 6.25 | 6.25 | 12.5 | 12.5 | 6.25 | 12.5 | 25 |
| A. fumigatus AA15 | 25 | 25 | 25 | 50 | 50 | 50 | 25 |
| A. tabacinus AF16 | 50 | 25 | 25 | - | - | - | 50 |
| Penicillium glabrum AE3-1 | 50 | 50 | 12.5 | - | - | - | 50 |
| P. polonicum AA17 | 50 | 50 | 50 | - | - | - | 50 |
| P. brevicompactum AE7 | 10 | 50 | 25 | - | - | - | 50 |
| P. vasconiae AF5 | - | - | - | - | - | - | 50 |
| Cordyceps farinosa AA8 | 100 | 100 | 100 | - | - | - | 50 |
| Lecanicillium aphanocladii AF15 | 25 | 50 | 50 | 50 | 50 | - | 50 |
| Umbelopsis nana AF31 | 50 | 50 | 25 | 50 | 50 | - | 50 |
| Positive control: vancomycin | 2 | 2 | 2 | - | 2 | 2 | 2 |
“-”: MIC > 100 µg/mL.
Table 3.
Antimalarial screening of rhizosphere fungi associated with Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae) against Plasmodium falciparum 3D7.
Table 3.
Antimalarial screening of rhizosphere fungi associated with Astragalus acaulis (Aa), A. forrestii (Af), and A. ernestii (Ae) against Plasmodium falciparum 3D7.
| Fungal strain | Test concentration (µg/mL) | Inhibitory rates (%) |
|---|---|---|
| Aspergillus calidoustus AA12 | 100 | 99.78 |
| 50 | 99.60 | |
| A. fumigatus AA15 | 100 | 81.81 |
| 50 | 14.31 | |
| A. tabacinus AF16 | 100 | 95.60 |
| 50 | 91.19 | |
| Penicillium glabrum AE3-1 | 100 | 29.57 |
| 50 | 0.93 | |
| P. polonicum AA17 | 100 | -51.88 |
| 50 | -32.53 | |
| P. brevicompactum AE7 | 100 | 87/95 |
| 50 | 81.91 | |
| Cordyceps farinosa AA8 | 100 | -45.30 |
| 50 | -15.46 | |
| Positive control: chloroquine | 0.16 | 99.88 |
Table 4.
Antibacterial activity of compounds 1-7 against seven strains of Gram-positive pathogenic bacteria, including methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300, S. aureus ATCC 25923, S. epidermidis ATCC 12228, Vancomycin-resistant Enterococcus faecalis (VREF) ATCC 51299, E. faecalis ATCC 29212, E. faecium ATCC 35667 and Streptococcus agalactiae ATCC 13813 (µg/mL).
Table 4.
Antibacterial activity of compounds 1-7 against seven strains of Gram-positive pathogenic bacteria, including methicillin-resistant Staphylococcus aureus (MRSA) ATCC 43300, S. aureus ATCC 25923, S. epidermidis ATCC 12228, Vancomycin-resistant Enterococcus faecalis (VREF) ATCC 51299, E. faecalis ATCC 29212, E. faecium ATCC 35667 and Streptococcus agalactiae ATCC 13813 (µg/mL).
| Compounds | ATCC 43300 | ATCC 25923 | ATCC 12228 | ATCC 51299 | ATCC 29212 | ATCC 35667 | ATCC 13813 |
|---|---|---|---|---|---|---|---|
| 1 | 25 | 12.25 | 12.5 | 50 | 12.5 | 25 | 6.25 |
| 2 | - | - | - | - | - | - | 80 |
| 3 | - | - | - | - | - | - | 80 |
| 4 | - | - | - | - | - | - | 50 |
| 5 | - | - | - | - | - | - | 100 |
| 6 | - | - | - | - | - | - | - |
| 7 | 100 | - | - | - | - | - | - |
| Vancomycin | 2 | 2 | 2 | - | 2 | 2 | 2 |
“-”: MIC > 100 µg/mL.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



