
Submitted:
07 August 2024
Posted:
09 August 2024
You are already at the latest version
Abstract
The development of a rapid, sensitive, specific method for detecting foodborne pathogens is paramount in supplying safe food to enhance public health safety. Despite the significant improvement in pathogen detection methods, key issues are still associated with rapid methods, such as distinguishing living cells from dead, pathogenic potential or health risk of the analyte at the time of consumption, detection limit, and sample-to-result. The mammalian cell-based assays analyze pathogens' interaction with the host cells and are responsive to only live pathogens or active toxins. In this study, a human embryonic kidney (HEK293) cell line expressing Toll-Like Receptor 5 (TLR-5) and chromogenic reporter system (HEK dual hTLR5) was used for the detection of viable Salmonella in a 96-well tissue culture plate. This cell line responds to low concentrations of TLR5 agonist flagellin. Stimulation of TLR5 ligand activates nuclear factor-kB (NF-κB) - linked alkaline phosphatase (AP-1) signaling cascade inducing the production of secreted embryonic alkaline phosphatase (SEAP). With the addition of a ρ-nitrophenyl phosphate as a substrate, a colored end product representing a positive signal is quantified. The assay's specificity was validated with the top 20 Salmonella enterica serovars and 19 non-Salmonella spp. The performance of the assay was also validated with spiked food samples. The total detection time (sample-to-result), including shortened pre-enrichment (4 h) and selective enrichment (4 h) steps with artificially inoculated outbreak-implicated food samples (chicken, peanut kernel, peanut butter, black pepper, mayonnaise, and peach) was 15 h when inoculated at 1–100 CFU/25 g sample. These results show the potential of HEK-DualTM hTLR5 cell-based functional biosensors for rapid screening of Salmonella.
Keywords:
Salmonella
; Cell-based sensor
; HEK-dual hTLR 5
; Detection
; Flagella
; Immunomagnetic Separation
; Food Safety
1. Introduction
Salmonella is a member of the Enterobacteriaceae family and is a Gram-negative, non-spore-forming motile bacillus with peritrichous flagella [1]. Salmonellae are widespread in the environment and found in various foods and ingredients, posing serious problems to the food industry [2,3,4,5,6]. Non-typhoidal Salmonella (NTS) infection is the second most common zoonotic disease after campylobacteriosis in the European Union member states, which reported over 91,000 cases in 2018. According to the Centers for Disease Control and Prevention (CDC), Salmonella causes about 1.35 million infections, 26,500 hospitalizations, and 420 deaths in the United States every year [7,8]. Salmonella enterica outbreaks have been associated with a variety of foods, including spices, nuts, flours, bakery products, fresh produce, and poultry, due to their robust physiological adaptability in harsh conditions [9,10,11].
Noteworthy outbreaks are reported for Salmonella enterica serovar Oranienburg (S. Oranienburg) involving black pepper [12] and onion [13,14]; S. Montevideo in black and red peppers [15]; S, Mbandaka, S. Stanley and S. Newport in peanut and peanut butter [16,17]; S. Agona, S. Anatum, S. Braenderup, S. Dessau, S. Hartford, S. Meleagridis, S. Muenchen, S. Rodepoort, S. Tennessee, and S. Tornow in shelled peanuts [18]; S. Enteritidis in fresh peaches [19], S. Heidelberg, S. Infantis, and S. Tennessee in chicken [20,21,22,23] and S. Typhimurium definitive type 49 in eggs and mayonnaise prepared with eggs [24], and S. Indiana associated with egg mayonnaise sandwiches [24,25]. Moreover, in Brazil, potato salad made with homemade mayonnaise was associated with S. Enteritidis [26], and buffet dishes containing mayonnaise were associated with S. Enteritidis outbreak in Germany [27].
Numerous rapid methods, including miniaturized biochemical tests, physicochemical methods that measure bacterial metabolites, nucleic acid-based tests, bacteriophage-based sensors, and antibody-based methods, have been developed [28,29,30]. Conventional methods for detecting Salmonella include pre-enrichment, selective enrichment, selective and differential plating, biochemical phenotyping of suspect colonies, and serotyping [28]. These procedures are time-consuming, laborious, and may take 5 to 7 days [31,32,33,34,35]. For consumer protection, risk-based rapid methods with high specificity and sensitivity are necessary to provide confirmed results in a day, and the quality of these results should at least be as reliable as the reference methods [36].
There is an increasing interest in the applications of biosensors that incorporate mammalian cells to detect foodborne pathogens to differentiate viable from non-living cells or toxins, which is a critical determinant for the food industry since nonviable pathogens are not considered a threat [35,37,38,39]. Cell-based biosensors (CBBs) have been applied to several detection systems and continue to serve as a reliable method to probe for the presence of pathogens in clinical, environmental, or food samples [40]. Mammalian CBBs exploit host-pathogen interactions, including pathogen adhesion, activation of host cell signaling events, cell-cycle arrest, apoptosis, and/or cytotoxicity [35,39,41]. The ability to detect host-pathogen interaction makes CBB a functional test, thus setting it apart from conventional methods. Most CBBs measure the optical properties of cellular metabolites or intracellular enzymes released after the pathogens' interaction with mammalian cells. Cytotoxicity assays also have been developed independently to detect Salmonella spp., using the Caco-2 cell line [42], RAW264.7 [43], and J774 macrophage cell lines [44]. However, due to the complex nature of food matrices, their specificity or sensitivity cannot be guaranteed only based on cytotoxicity analysis. Alternatively, an analyte-specific amplified cell signaling cascade approach could be adopted to improve the specificity and sensitivity of CBB.
In this study, our goal was to use an engineered human embryonic kidney (HEK293) cell line expressing Toll-Like Receptor 5 (TLR5) and chromogenic reporter system (HEK dual hTLR5) as the sensor platform. This cell line responds to low concentrations of TLR5 agonist flagellin [45,46]. Stimulation of TLR5 ligand activates canonical nuclear factor-kB (NF-κB) - linked alkaline phosphatase (AP-1) signaling cascade inducing the production of SEAP (secreted embryonic alkaline phosphatase) (Figure 1a). With the addition of ρ-nitrophenyl phosphate as a substrate, a colored end product representing a positive signal can be detected by the naked eye or quantified spectrophotometrically. Not all bacterial flagellin, but Salmonella flagellin is strongly recognized by TLR5 (Andersen-Nissen et al. 2005). In addition, employing an immunomagnetic separation (IMS) system such as anti-Salmonella Dynabeads (Invitrogen) can also improve assay specificity and sensitivity by capturing and concentrating target pathogens from complex food matrices before exposure to HEK dual hTLR5 cells [47,48,49,50].
Overall, the data showed that the HEK dual hTLR5 cell-based sensor could detect viable ~50-100 S. Enteritidis cells both in spiked food samples (black pepper, chicken, mayonnaise, peanut kernel, peanut butter, and peach) in 15 h (including enrichment steps) with high accuracy and specificity. Our results suggest the potential application of HEK-DualTM hTLR5 cell-based functional biosensors for rapid detection of Salmonella.
2. Results
2.1. Optimization of HEK Dual hTLR5 Sensor for Detection of Salmonella
HEK-Blue-hTLR5 cells (InvivoGen, CA, USA) were constructed with the human TLR5 gene and an inducible secreted embryonic alkaline phosphatase (SEAP) reporter gene (Figure 1a). The SEAP gene is placed under the control of the IFNβ minimal promoter fused to five NF-κB and AP-1-binding sites. Stimulation with a TLR5 ligand activates NF-κB and AP-1, which induce the production of SEAP. The amount of SEAP production can be determined qualitatively with the naked eye or quantified with the HEK-Blue Detection system (Quanti Blue substrate) by measuring absorbance at 620-655 nm (Figure 1b). Initial experiments were conducted to determine if HEK-Blue-hTLR5 cells are responsive to Salmonella. At multiplicity of infection (MOI) 10, HEK cells exposed to Salmonella enterica ser Enteritidis (PT21) for 6 h showed a significantly higher response (~6-fold increase; p < 0.0001) than Escherichia coli O157:H7, Listeria monocytogenes and Staphylococcus aureus (Figure 1c). These data indicate that HEK-Blue-hTLR5 can be used to detect Salmonella in a cell-based sensor.
Bacterial motility assay results on agar plates at different concentrations (0.3%, “swim plates”; 0.5%, 0.7%, and 1%, “swarm plates”) [51,52] confirmed S. Enteritidis to be highly motile and the most evident colony diameter differences were observed at 0.3% agar (swim plates) than the other bacteria tested (Figure S1). Significantly higher flagellar expression in Salmonella compared to other bacterial genera tested may contribute to the observed high signal with the HEK dual hTLR5 sensor than the other pathogens (Figure 1c).
Next, we determined the optimal Salmonella cells to HEK cell ratio (multiplicity of infection, MOI) needed to show a positive response after 4 or 6 h of incubation. At 4 h (Figure 1d), MOI 1 and 10 showed 6.7 and 8.7-fold higher signals than the uninfected controls, while at 6 h (Figure 1d), MOI 0.1, 1, and 10 showed 14.2, 15.3, and 10.3-fold higher signal, respectively. E. coli O157¯493-89, a non-motile strain, was used as a negative control. These data indicate that an MOI of 1 exhibits a positive signal after 4 h of exposure, while an MOI of 0.1 is sufficient after 6 h of exposure. However, TLR-5 response to MOI 0.1 compared to MOI 1 at 6 h did not significantly differ. Therefore, we used MOI 1 (4 h and 6 h) for further experiments.
Figure 1.
Cell-based assay for detection of Salmonella using HEK dual TLR5 sensor. (a-b) Schematic Representation: This panel illustrates the flow diagram of the cell-based assay. The process begins with preparing the Quantiblue solution to track the TLR5 response. HEK cells engineered to express TLR5 response are then exposed to varying concentrations of Salmonella bacteria, leading to an observable color change. The schematic details the steps in the assay, including cell culture, interaction with the bacteria, and the TLR5 response mechanism triggered by the bacterial flagella. Image created using Biorender.com. (c) This panel represents the analysis of the HEK dual TLR5 sensor's response to various foodborne bacterial pathogens tested at MOI of 1 for h and 6 h. (d) These panels present the analysis of the signal response from the HEK dual TLR5 sensor cells at different multiplicities of infection (MOI), examining the ratio of HEK cells to bacteria. The MOI ratios analyzed include various levels to determine the sensitivity and dynamic range of the assay. Left panel shows the signal response after 4 hours of incubation, while right panel depicts the response after 6 h. Data in c-d represent the average results from three independent experiments presented as mean ± SEM.
Figure 1.
Cell-based assay for detection of Salmonella using HEK dual TLR5 sensor. (a-b) Schematic Representation: This panel illustrates the flow diagram of the cell-based assay. The process begins with preparing the Quantiblue solution to track the TLR5 response. HEK cells engineered to express TLR5 response are then exposed to varying concentrations of Salmonella bacteria, leading to an observable color change. The schematic details the steps in the assay, including cell culture, interaction with the bacteria, and the TLR5 response mechanism triggered by the bacterial flagella. Image created using Biorender.com. (c) This panel represents the analysis of the HEK dual TLR5 sensor's response to various foodborne bacterial pathogens tested at MOI of 1 for h and 6 h. (d) These panels present the analysis of the signal response from the HEK dual TLR5 sensor cells at different multiplicities of infection (MOI), examining the ratio of HEK cells to bacteria. The MOI ratios analyzed include various levels to determine the sensitivity and dynamic range of the assay. Left panel shows the signal response after 4 hours of incubation, while right panel depicts the response after 6 h. Data in c-d represent the average results from three independent experiments presented as mean ± SEM.
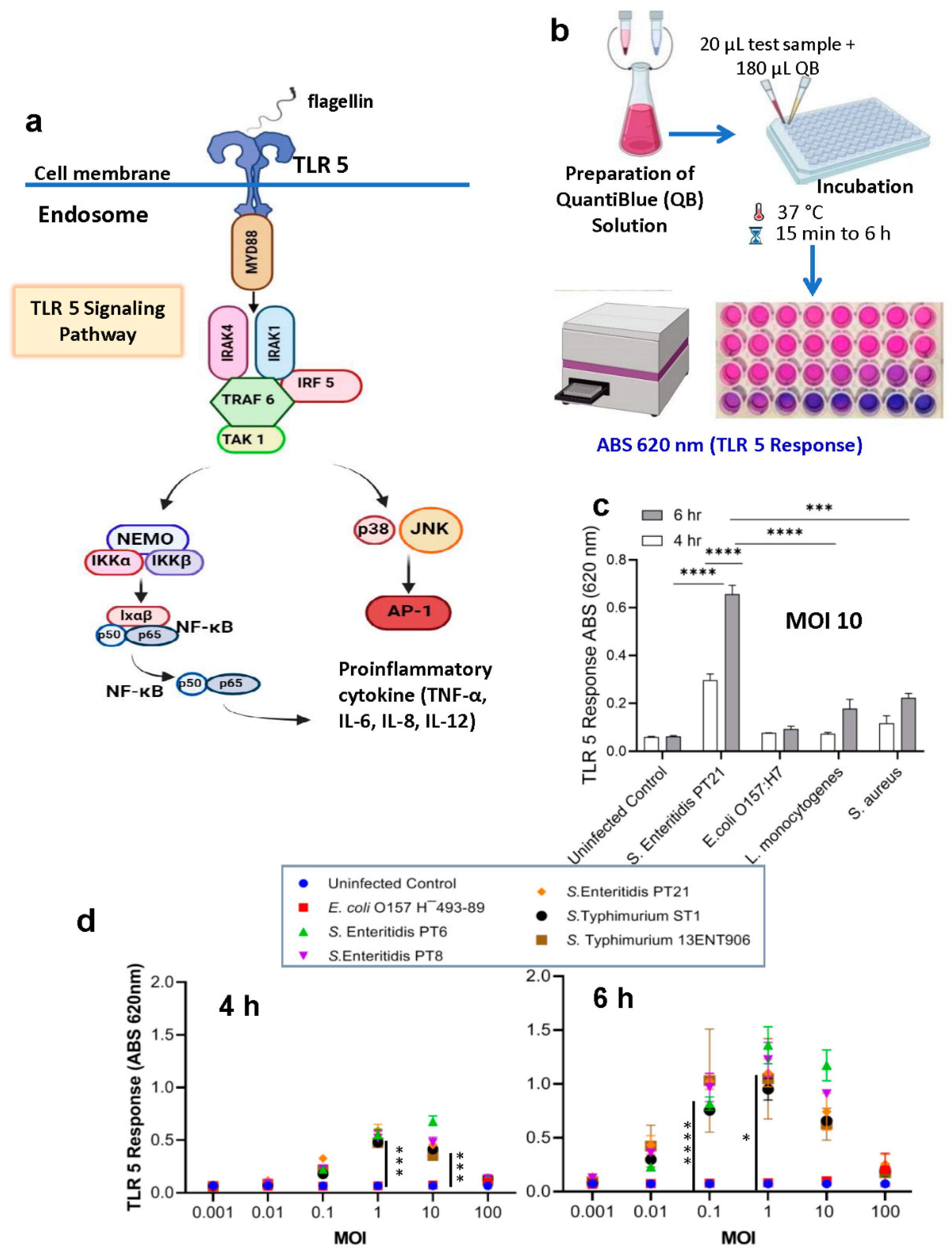
2.2. Specificity of HEK dual TLR5 sensor for detection of Salmonella serovars
The specificity of the HEK dual hTLR5 sensor for S. enterica was determined by testing the top 20 S. enterica serovars and non-Salmonella bacteria [53]. An MOI 1 and the bacterial exposure time of 4 and 6 h was used, and the cell supernatants were tested for AP activity. All 20 tested Salmonella strains showed positive signals and values were ~8 and 16-fold higher than the negative controls at 4 h and 6 h, respectively (Figure 2a). E. coli O157¯493-89, a non-motile strain, was used as a negative control and baseline to ensure that observed TLR5 responses were motility-dependent (Figure 2a).
Compared to Salmonella Enteritidis (PT21), most non-Salmonella bacteria, including Hafnia alvei, Serratia marcescens, Proteus mirabilis, Proteus vulgaris, Klebsiella pneumoniae, and few E. coli O157:H7, O26 and O103 strains showed a 2~3-fold decrease in TLR-5 response. In contrast, 2 of 3 Citrobacter freundii and E. coli O157:H7 strains showed higher responses (Figure 2b). The cytotoxic effect of the test strains on HEK dual hTLR5 cells was analyzed by monitoring the release of lactate dehydrogenase (LDH; Figure 2c). In general, cytotoxicity values were below 20% and significantly lower than the positive control (1% Triton X-100 or L. monocytogenes), suggesting that the flagella-mediated signal response obtained was largely independent of cellular damage. These results indicate that the HEK dual TLR5 sensor can be used to detect Salmonella serovars due to TLR-5 recognizable flagellin expression [54]. Any non-Salmonella bacteria that produce a TLR-5 response, could be eliminated during selective enrichment and/or IMS separation.
Figure 2.
Specificity analysis of the HEK dual TLR5 sensor's response to various Salmonella serovars. (a) Specificity analysis of the HEK dual TLR5 sensor's response to various Salmonella serovars. The assay was conducted at a multiplicity of infection (MOI) of 1, with 4 and 6 hours exposure times. The data illustrate the sensor's ability to specifically detect different Salmonella serovars. E. coli O157¯493-89, a non-motile strain, was used as a negative control. (b) Specificity analysis of the HEK dual TLR5 sensor's response against non-Salmonella bacteria: This panel shows the analysis of the HEK dual TLR5 sensor's response to a range of non-Salmonella bacteria. Like panel (a), the assay was performed at an MOI of 1, with signal responses measured after 4 and 6 hours of exposure. Salmonella PT21 was used as a positive control. (c) Cytotoxicity was determined by using an LDH release assay. The assay was conducted at a multiplicity of infection (MOI) of 1, with 6-hour exposure times. A 1% Triton-X100 was used as a positive control (100% cytotoxicity). Data in a-c represent the average results from three independent experiments presented as mean ± SEM.
Figure 2.
Specificity analysis of the HEK dual TLR5 sensor's response to various Salmonella serovars. (a) Specificity analysis of the HEK dual TLR5 sensor's response to various Salmonella serovars. The assay was conducted at a multiplicity of infection (MOI) of 1, with 4 and 6 hours exposure times. The data illustrate the sensor's ability to specifically detect different Salmonella serovars. E. coli O157¯493-89, a non-motile strain, was used as a negative control. (b) Specificity analysis of the HEK dual TLR5 sensor's response against non-Salmonella bacteria: This panel shows the analysis of the HEK dual TLR5 sensor's response to a range of non-Salmonella bacteria. Like panel (a), the assay was performed at an MOI of 1, with signal responses measured after 4 and 6 hours of exposure. Salmonella PT21 was used as a positive control. (c) Cytotoxicity was determined by using an LDH release assay. The assay was conducted at a multiplicity of infection (MOI) of 1, with 6-hour exposure times. A 1% Triton-X100 was used as a positive control (100% cytotoxicity). Data in a-c represent the average results from three independent experiments presented as mean ± SEM.
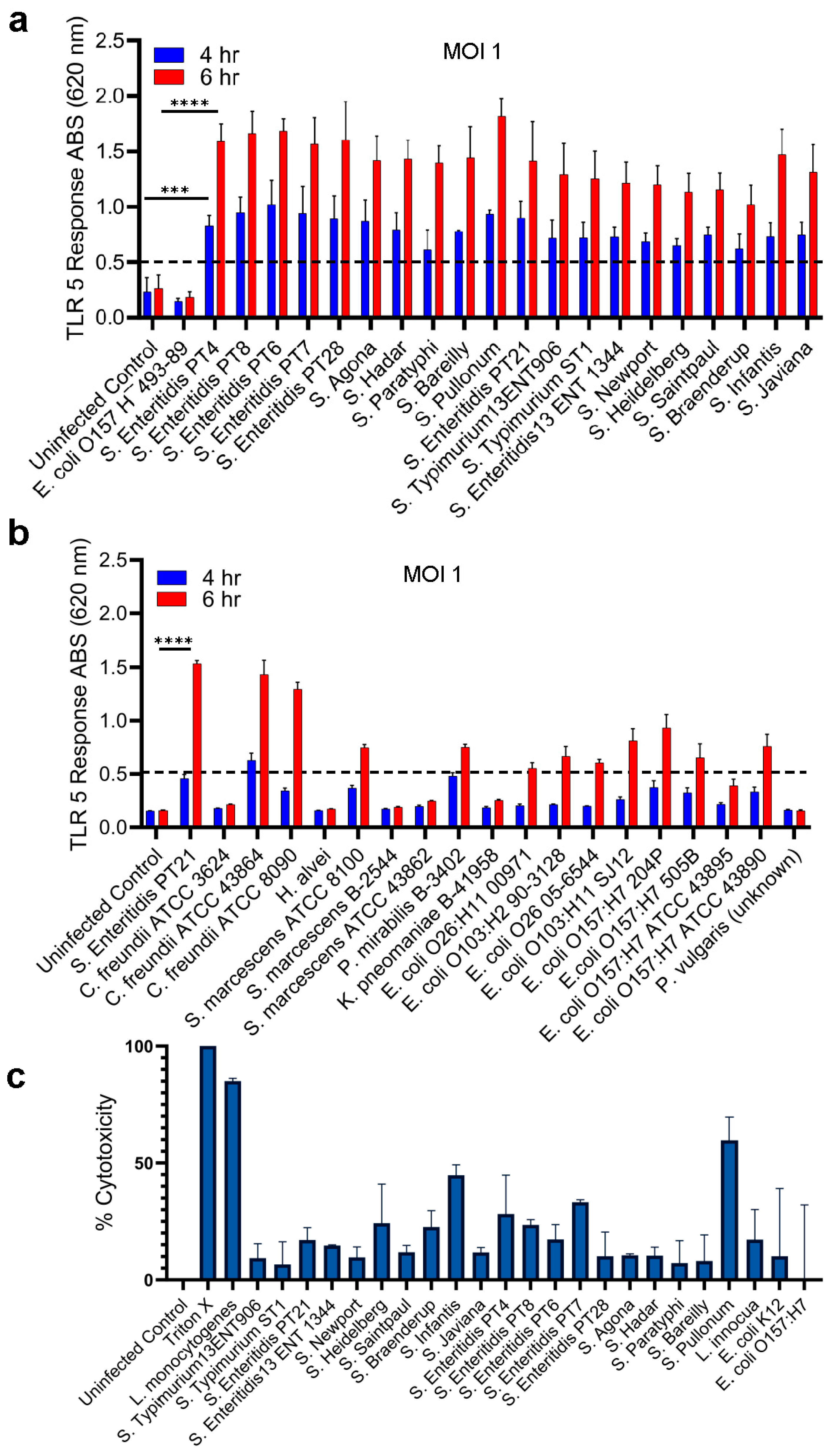
Figure 3.
Analysis of the HEK Dual TLR5 Sensor's Response in the presence of stressors. (a) Response to Live and Dead Bacteria: This panel illustrates the HEK dual TLR5 sensor's response to both live and dead bacteria after 6 hours of exposure at a multiplicity of infection (MOI) of 1. Panels on the top right depict representative color changes of the HEK dual TLR5 sensors between live (purple) and dead (pink) tested samples. (b, c) Effects of Stressors on TLR5 Response: These panels investigate how different stressors affect the TLR5 response of Salmonella PT21. Panel (b) shows the TLR5 response after 4 hours of exposure, while panel (c) presents the response after 6 h of exposure, both tested at an MOI of 1. Data in a-c represent the average results from three independent experiments presented as mean ± SEM.
Figure 3.
Analysis of the HEK Dual TLR5 Sensor's Response in the presence of stressors. (a) Response to Live and Dead Bacteria: This panel illustrates the HEK dual TLR5 sensor's response to both live and dead bacteria after 6 hours of exposure at a multiplicity of infection (MOI) of 1. Panels on the top right depict representative color changes of the HEK dual TLR5 sensors between live (purple) and dead (pink) tested samples. (b, c) Effects of Stressors on TLR5 Response: These panels investigate how different stressors affect the TLR5 response of Salmonella PT21. Panel (b) shows the TLR5 response after 4 hours of exposure, while panel (c) presents the response after 6 h of exposure, both tested at an MOI of 1. Data in a-c represent the average results from three independent experiments presented as mean ± SEM.
2.3. Detection of Live and Stress-Exposed Salmonella Using HEK dual TLR5 Sensor
We next tested if the HEK dual hTLR5 sensor could differentiate live from dead cells since processed food products may carry dead cells. To determine the sensor's response to live and dead cells, overnight-grown fresh cultures of different Salmonella serovars were prepared, and MOI was adjusted to 1. Salmonellae were treated with heat (80°C for 10 min) and plated on TSA to ensure cells were inactivated (non-viable). AP levels were monitored after 6 h exposure of HEK dual hTLR5 sensor to Salmonella. Only the live cells of S. enterica serovars showed positive signals (~10-fold increase), while heat-inactivated dead cells did not yield a signal (Figure 3a)
Next, we tested the effects of various stressors on detecting S. Enteritidis PT21 strain that was subjected to multiple stressors, including cold (4ºC), heat (45ºC), acidity (pH 5.5), ethanol (5%, v/v), NaCl (5.5%), and oxidative stress (H2O2, 15 mM) for 3 h followed by 3 h resuscitation in TSBYE [37,55]. Bacterial cell suspensions were plated to assess cell viability and cell counts (Table 1) to ensure the viability of Salmonella following stress induction and to confirm the desirable MOI before exposure to the HEK dual hTLR5 sensor. All stressors showed positive results in the tested Salmonella PT21 strain at both 4 and 6 h, respectively (Figs. 3b and 3c); however, there was a slight but negligible reduction in signal response (Figs. 3b & 3c). These results suggest that the HEK dual hTLR5 sensor can detect viable or stress-exposed Salmonella cells.
2.4. Detection of Salmonella from Spiked Food Samples Using HEK dual TLR5 Sensor
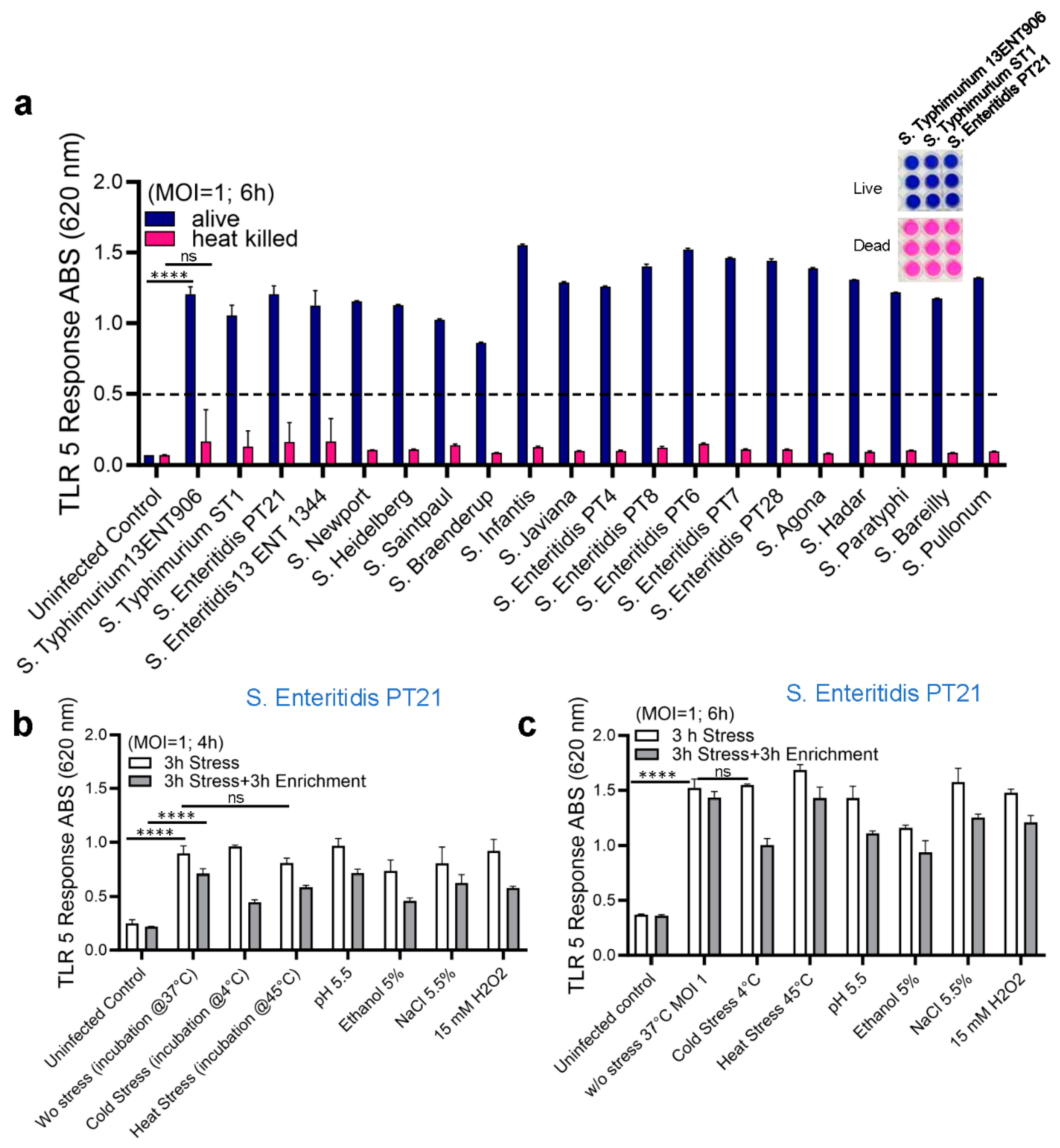
The performance of the HEK dual hTLR5 sensor was validated using artificially spiked food samples. Initially, the limit of detection (LOD) for the sensor was determined from spiked chicken and peanut butter samples. Background flora on tested chicken thigh meat samples was 1.92 CFU/cm2 but tested negative for Salmonella. Chicken thigh meat samples were artificially contaminated with different levels of S. Enteritidis PT21 cells (7.2 × 105 CFU/cm2, 7.2 × 104 CFU/cm2, 7.2 × 103 CFU/cm2, 7.2 × 102 CFU/cm2, 7.2 × 101 CFU/cm2) and incubated overnight in the cold room (4ºC). Skin surfaces were swabbed using Dacron swabs, transferred to tubes containing 1 ml of PBS (pH 7.2), serially diluted, and 100 µl samples added to HEK dual hTLR5 cells growing in a 96-well cell culture plate and incubated for 4- 6 h. Bacterial counts in the inoculums were enumerated to be 4.3 ×106, 3.6 × 105, 2.7 × 104, 3.2 × 103, and 1.0 × 103 CFU/mL, respectively. Relative to uncontaminated meat, S. Enteritidis cells from spiked samples at all inoculation levels showed positive signals at 4 h and 6 h. The values were 2.5-6-fold higher (p < 0.05) than the uninoculated sample (Figure 4a). Visual examination of color changes in the wells (6 h) corroborates with absorbance readings (Figure 4a, bottom panel). These data indicate that the TLR5 sensor could detect as few as 100 Salmonella cells from the chicken samples.
The LOD of the sensor was also determined using peanut butter. No background flora was found in tested peanut butter samples, which were also negative for Salmonella (Table 2). Peanut butter (25 g each portion) was spiked with S. Enteritidis PT21 to obtain an initial inoculum of about 5, 50, 500, and 5000 CFU /25 g, and enrichment steps followed a shortened USDA-FSIS procedure as described before and confirmed via plate CFU counts (Table 2) (Singh et al. 2014; USDA-FSIS). The initial inoculum containing 50 – 5 × 103 CFU/25 g showed positive signals, and the incremental values were proportional to the initial inoculum levels. Furthermore, these values significantly differed from the signals obtained from uninoculated or peanut butter inoculated with 5 CFU/25g (Figure 4b). These data indicate that the sensor can detect about 50 Salmonella cells (CFU/25g) (as an initial inoculum) without interference from peanut butter samples.
2.5. Validation of Sensor with Spiked Food Samples
Next, we tested the HEK dual hTLR5 sensor with additional spiked food samples (100 CFU/25g; chicken thigh meats, black pepper, peanut kernel, peanut butter, mayonnaise, peach) where sample preparation and enrichment steps followed a shortened USDA-FSIS procedure [53,56] (Table 3). One hundred microliters of enriched samples were tested for their response to HEK dual hTLR5 cells. Each experiment was repeated five times for all food samples on different days.
All S. Enteritidis-inoculated samples, irrespective of food types, produced ~5-10-fold higher signals (p < 0.001) than the uninoculated food samples (Figure 5a). Interestingly, the signal from inoculated mayonnaise was lowest among all the food samples tested. Furthermore, Microscopic observation of HEK dual hTLR5 cells after exposure to various food samples also did not affect the cell monolayer integrity or cellular morphology (Figure 5b). Parallelly, the same set of samples was tested with plate counting (on XLD) methods and PCR for verification. Compared to the TLR5 sensor, PCR could only detect Salmonella in peanut kernel, while other spiked food samples post-enrichment tested negative (Table 3). Negative PCR results from inoculated food samples are attributed to the presence of PCR inhibitors. These data suggest that the TLR5 cell sensor can detect as low as 100 CFU/25g of pre-enriched food samples with complex food matrix.
Next, we employed immunomagnetic separation (IMS) to concentrate Salmonella from enriched food samples before testing with the HEK dual hTLR5 sensor to determine if the signal could be augmented [47]. Before conducting food analysis using the IMS method, we investigated the immunomagnetic bead (IMB) based capture rate of freshly grown diluted (8.99 to 1.99 log CFU/mL bacteria) cultures of S. Enteritidis by plating. The capture rate was calculated to be >90% for all concentrations, regardless of the bacterial load (Table S1). The application of IMS significantly improved the signal except for the inoculated black pepper (Figure 5a). The sample-to-result time was estimated to be 15 h.
3. Discussion
The risk-based rapid and sensitive detection of Salmonella species is highly desirable to protect consumers from food-associated illnesses and promote food safety and biosecurity. Although various detection platforms have been developed in recent years, the traditional culture-based detection methods require 4–7 days to yield results [37,56,57,58], whereas rapid methods still take at least 24–48 h [28,38,59,60,61]. This delay poses a significant inconvenience to the food industry, especially for products with short shelf lives. Additionally, the requirement to store products until microbiological safety is confirmed increases storage costs. Therefore, products often enter the supply chain before test results are available, which is costly and results in hundreds of recalls each year and millions of pounds of food wasted, which is unsustainable to meet the future food demand [62,63]. This situation creates a high demand for quick, accurate, easy-to-use pathogen detection tools to reduce recalls, minimize food waste and financial losses, and prevent foodborne illnesses. Mammalian cell-based assays are highly promising for functional screening because they can detect viable pathogens in real-time [39,64]. These assays monitor host-pathogen interactions [40,41], ensuring that non-pathogenic, non-hazardous, dead, or non-toxic agents do not produce false results.
This study uses a human embryonic kidney (HEK293) cell line expressing Toll-Like Receptor 5 (TLR-5) and a chromogenic reporter system to detect viable Salmonella in a 96-well tissue culture plate. Flagellin is a bacterial protein that polymerizes into the flagellar filament and allows bacteria to be motile. During the bacterial invasion of the host cell, flagellin is recognized by TLR5 and activates innate immune signal transduction cascades [54,65,66]. TLR5 is an innate immune receptor located on the cell surface and consists of an extracellular leucine-rich repeat (LRR), transmembrane, and intracellular domains [67]. TLR5 uses the extracellular domain to recognize flagellin as a pathogen-associated molecular pattern (PAMP). It activates the MyD88-dependent signaling pathway and NF-κB-mediated production of proinflammatory cytokines, and this response can be measured using the chromogenic detection system (Figure 1) [45].
To test our hypothesis that the HEK-TLR-5 sensor can detect viable Salmonella selectively, we first determined the motility of different types of common foodborne bacterial pathogens (Salmonella Enteritidis PT21, E. coli O157:H7, and Listeria monocytogenes). The motility test (plating results performed on agar surface) and the signals obtained from the HEK dual hTLR5 sensor correlated as Salmonella was found to be the most motile bacteria with the highest swimming ability, and the sensor with a flagella recognition feature showed the highest response when tested with Salmonella Enteritidis PT21.
Furthermore, flagellar expression is critical for obtaining positive signals. We verified relative flagellar expression in test strains by measuring their motility on agar plates containing variable amounts of agar (0.3 – 1.0%; w/v) [52]. Swimming motility is the movement in liquid or low-viscosity conditions (up to 0.3% agar concentration). Unlike swarming motility, swimming motility requires a functional flagellum, but neither quorum sensing systems nor biosurfactants [51,52]. Plates with up to 0.3% agar concentration are referred to as ‘swim plates’ because highly motile flagellum-dependent bacteria can swim through the porous medium, leading to a large area of colonization [51]. Plates with agar concentration higher than 0.3% are known as ‘swarm plates’ because the reduced pore size prevents swimming through the agar, and extensive colonization of the plate occurs only through bacterial migration across the surface.
Specificity testing of the HEK-hTLR5 Reporter Cell-Based Sensor showed a high response (6-fold increase; Figure 2b) with all the top 20 Salmonella serovars tested. Testing with the non-Salmonella organisms suggested minimal signals with several common foodborne bacterial pathogens such as Enterohemorrhagic Escherichia coli (EHEC) strains, H. alvei, S. marcescens, K. pneumoniae, and Proteus vulgaris, suggesting their flagellin molecules may be poorly recognized by TLR5 [54]. However, 2 of 3 Citrobacter strains showed cross-reactivity, which may be attributed to high TLR-5 recognizable flagellin expression. More importantly, when tested with food samples, the sensor was specific as it showed no response to natural microflora or several uninoculated food samples. Black pepper, chicken, mayonnaise, peanut kernels, peanut butter, and peaches were chosen because these products are commonly linked with Salmonella outbreaks, and some are consumed without heat treatment. In addition, they have high protein, fat, and carbohydrate contents, which may cause interferences. Our data demonstrate that the tested uninoculated food samples had minimal or no interference with sensor response, but Salmonella inoculated samples showed a strong response. Surprisingly, PCR assay with inoculated food samples gave negative PCR results (except peanut kernel) (Table 3) and the negative PCR results are attributed to the presence of PCR inhibitors in these samples [68].
The sensitivity of the cell-based sensor in pure culture suggested that an MOI of 0.1 in the 105 seeded HEK-hTLR5 reporter cells on a 96-well plate elicited a positive signal (Figure 1d). These data indicate that the LOD in pure culture is ~ 105 CFU/mL (104 CFU/well in 100 µL). Our results are comparable with the LOD of the recently developed Next-Generation Sequencing Assay for direct detection of Salmonella [69]. Further testing of LOD in spiked food samples suggested the LOD is as low as 50-100 CFU/25g of food samples, which has the potential to outperform other molecular detection methods such as ELISA or PCR [61]. Our results are comparable with previous research showing an IMS-PCR procedure designed to isolate and detect S. enterica in vegetable salad, egg, and pork meat with a detection sensitivity of 102 CFU/mL [70]. However, one significant advantage of the HEK-hTLR5 cell-based sensor is its ability to selectively detect viable Salmonella and not respond to dead cells (Figure 3a). Additionally, although the sensitivity of the sensor may be high, accurate results may not be obtained due to the low level of pathogen survival in stressed food conditions. The HEK-hTLR5 reporter cell-based sensor was further validated and tested positive for the detection of Salmonella in stressed conditions such as cold (4◦C), heat (45◦C), acid (pH 5.5), ethanol (5%), NaCl (5.5%), and oxidative stress (15 mmol H2O2). A brief sample enrichment step allowed the resuscitation of stressed or injured cells before detection.
Our approach of combining enrichment steps with IMS improved the TLR-5 sensor sensitivity in certain food products, such as mayonnaise and yielded comparable results with other studies that used IMS tandem fluorescent probes based on quantum dots-antibody (QDs-Ab) for Salmonella testing [71]. However, the sensitivity of this assay was lower than our cell-based sensor (500 CFU/mL). Similarly, a protein chip immunosensor has been exploited to detect Listeria spp. by lowering the detection limit to 2.5 CFU/mL with the pre-enrichment culture of IMS [72]. While IMS can be evaluated as a potential pretreatment tool for the rapid isolation and enrichment of many foodborne pathogens in food, the bacterial surface antigens from dead cells could still bind the capture antibody. Therefore, our TLR-5 cell-based sensor combined with the IMS method is significantly more advantageous when viable pathogens are sought. Thus, developing cell-based assays is critical for functional screening and eliminating false-negative results from dead, non-hazardous, or non-pathogenic bacteria.
In conclusion, our two-step method (enrichment and TLR-5 sensor), with a significantly lower detection time of 15 h from sample to result, was found to be highly specific for the detection of live salmonellae in complex food matrixes in the presence of stressors with a sensitivity of 50 CFU/25 gram of food. Therefore, the TLR-5 sensor could be used as a risk-based screening tool for viable Salmonella.
4. Materials and Methods
4.1. Bacterial Cultures and Motility Testing
Bacterial strains (Table 4) were stored as 10% glycerol stocks at -80oC [53]. To recover from frozen stock, each strain was streaked onto tryptic soy agar (TSA) (Thermo Fisher Scientific, Rochester, NY, USA) plate and incubated at 37oC for 18 h to obtain pure colonies. A single colony of each strain was inoculated and propagated in tryptic soy broth (TSB) with 0.5% yeast (Thermo Fisher Scientific, Rochester, NY, USA) at 37oC for 18 h with shaking at 120 rpm.
For bacterial motility (swimming vs swarming) assay, a previous method [52] was followed. Briefly, bacteria were spot inoculated onto TSA plate containing either 0.3% w/v (for swimming motility) or 1.0% w/v (swarming motility) agar, incubated at 37ºC for 18-24 h and recorded colony diameter.
4.2. Cell Culture
HEK-Dual hTLR 5 (NF-KB-IL8) cells were purchased from InVivoGen (San Diego, CA, USA) and grown in Dulbecco's Modified Eagle Medium (DMEM) containing heat-inactivated 10% fetal bovine serum, antibiotics (Penicillin-Streptomycin, Normocin, Hygromycin B Gold and Zeocin). Cells were incubated at 37°C with 5% CO2 in a humidified incubator. Cells were seeded in tissue culture flasks, and medium was refreshed twice a week until cells reached the desired confluency. HEK 293 hTLR 5 cell monolayer was maintained and harvested using trypsin, and a hemocytometer with Trypan blue staining determined viable cell concentrations. A cell concentration of ~5 x 105/ml were added to each well of a 96-well cell culture plate and incubated overnight at 37°C under 5% CO2 in a humidified incubator for 24 h. Then, HEK293 cells were treated with bacteria to obtain the desired multiplicity of infection (MOI) value.
4.3. Cell-Based Detection of Bacterial Culture
HEK cell monolayers were prepared and maintained as described above in a 96-well plate. Overnight bacterial cultures were obtained and diluted to the desired concentration using PBS, which was then transferred to DMEM media. Heat-killed bacteria suspensions were prepared with heat treatment at 80°C for 10 min. Lethality of all bacteria spp. was confirmed by plating on tryptic soy agar (TSA) (Thermo Fisher Scientific, Rochester, NY, USA) plates. A 100 µL aliquot of culture suspension in DMEM was added to each well. After 4 and 6 h of incubation at 37oC, a 20 µL aliquot of the supernatant was collected from each well and transferred into another empty 96-well plate and mixed with 180 µL of the Quanti Blue Solution (InVivoGen, San Diego, CA, USA) used as a detection reagent to quantify Secreted Embryonic Alkaline Phosphatase (SEAP) activity. After one hour of incubation at room temperature in the dark, absorbance at 620 nm was measured using a microplate spectrophotometer (BioTech Epoch). SEAP is a truncated form of placental AP and is widely used as a reporter gene. SEAP is secreted into cell culture supernatant and offers many advantages over intracellular reporters. It allows the determination of reporter activity without disturbing the cells, does not require the preparation of cell lysates, and can be used for kinetic studies.
4.4. Specificity/Selectivity of Cell-Based Sensor
HEK Dual hTLR 5 (NF/IL8) reporter cells were generated from HEK-Dual Null cells by stable transfection of the human TLR5 (hTLR5) gene (InVivoGen, San Diego, CA). They respond to low concentrations of TLR5 agonist flagellin. To test the specificity and selectivity of this cell line, a non-motile E. coli O157¯493-89 and several non-Slamonella cultures were used (Table 5).
4.5. Limit of Detection of Cell-Based Sensor
Hek Dual hTLR 5 (NF/IL8) reporter cell line was used to detect bacterial flagellin. NTS salmonellae express flagellin and are motile. To determine the limit of detection of this sensor, we prepared Salmonella cultures with different dilution levels. The MOI was adjusted to 100, 10, 1, 0.1, 0.01, and 0.001 (HEK cell count was seeded to ~105 bacteria/well). After treating HEK Cells with specified bacteria levels for 4 h and 6 h, the SEAP level was assayed for each well by measuring the absorbance values at 620 nm. The lowest concentration of bacteria, which caused a significant TLR 5 response change, was considered the limit of detection for the sensor. Depending on the experimental design, in most cases, an absorbance (620 nm) cut-off value was set to 0.5.
4.6. Salmonella Enteritidis Analysis in Spiked Food Samples
Twenty-five grams of each food sample (except peach, which was sampled at 100 ± 5 g) was artificially contaminated with 500 µL of S. Enteritidis PT21 suspension and placed in sterile sampling bags in a biosafety cabinet. The contamination level was set to approximately 1 × 102 CFU/g. All samples were mixed with 225 ml (peach - 150 mL) of buffered peptone water and incubated at 37°C for 4 h. Then 1 ml of pre-enriched sample was transferred to 10 mL of RV Broth for selective enrichment and incubated at 42°C for 4 h. Finally, 1 mL of selective enriched sample mixed with 20 µL of Anti-Salmonella Dynabeads (Dynabeads™ anti-Salmonella. Catalog number: 71002; ThermoFisher) for immunomagnetic separation (IMS). Plating was performed after each step to determine the microbial load. After bead separation, bacteria were suspended in DMEM for cell-based analysis. A 100 µL of sample/well were transferred to the HEK cell monolayer in 96-well plates. TLR 5 response analyses were performed following 4 and 6 h of bacteria-cell interaction.
4.7. Detection of Stressed Cells
S. Enteritidis PT21 cells were freshly prepared in TSB and then exposed to cold (4◦C), heat (45◦C), acid (pH 5.5), ethanol (5%), NaCl (5.5%), and oxidative stress (15 mmol H2O2) for 3 h [55]. Bacterial cells were washed with PBS, resuspended in DMEM media, added to the HEK cell monolayer, and assayed as above.
4.8. Cytotoxicity Assays
To determine HEK cells' viability during pathogen exposure, supernatants from the wells were assayed for lactate dehydrogenase release (Cayman). Two controls were included for calculating percent cytotoxicity (LDH release). The low control consisted of supernatant from untreated cells with no exposure to bacteria. The high control was from cells treated with 0.1% Triton X-100 for one minute. For plate-based bacterial motility assay, a previous method [52] was followed.
4.9. Polymerase Chain Reaction (PCR) confirmation
Black pepper, Chicken thighs, mayonnaise, peanut kernel, peanut butter, and peach samples were inoculated with approximately 102 CFU/25 g of S. Enteritidis PT21 and were placed in 37oC for 4 h in pre-enrichment media (Buffered Peptone Water). After 4 h of pre-enrichment, samples were enriched selectively in Rappaport-Vassiliadis (RV) broth at 42oC for 4 h. Then, samples were subjected to testing using the TLR cell-based sensor platform. Parallelly, the same set of samples was tested with plate-counting methods. PCR was performed to verify the accuracy of both detection methods. DNA was extracted from the enriched samples using boiling methods.
A PCR reaction was used to amplify the gene targeting the Salmonella Invasion protein A (invA) with an amplicon size of 796 bp [37]. The forward primer sequence used was 5’CGGTGGTTTTAAGCGTACTCTT3’, and the reverse primer sequence used was 5’CGAATATGCTCCACAAGGTTA3’. The reaction mixture (25 μL) contained 1 μg of DNA template, 0.2 μM of each primer, 2.5 mM MgCl2, 200 μM of dNTP, 1 x GoTaq Flexi buffer and 1 U of GoTaq Flexi DNA polymerase (Promega). The reaction took place in the Profelx PCR system with an initial denaturation at 94oC for 3 min, 35 amplification cycles consisting of 1 min of denaturation at 94oC, 1.5 min of annealing at 50, and 1.5 min of elongation at 72oC. DNA amplicons (invA 796 bp) were detected in agarose gel (1.5%, wt/vol) containing 1 μg of ethidium bromide/mL [37].
4.10. Statistical Analysis
All data were analyzed through GraphPad Prism software. An unpaired t-test was used when comparing two datasets. ANOVA with Tukey’s multiple comparison test was used to compare more than two datasets. At least 3 independent experiments were performed for all data presented with mean ± standard error of the mean (SEM).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Analysis of Bacterial Motility on Agar Plates; Table S1: Performance of Anti-Salmonella Dynabeads.
Author Contributions
Conceptualization (AKB, EE, RD); Methodology and Data Analysis (EE, RD, VAF); Funding acquisition, Project administration, Resources, Supervision, and Review and editing (AKB, RD).
Funding
This research was funded by Scientific and Technological Research Council of Turkey (TUBITAK)–BIDEB under the 2219/ International Postdoctoral Research Fellowship Programme with Application Number 1059B191800917. The research was supported by funds from the National Institute of Food and Agriculture of USDA (Hatch grant # 1016249; A.K.B.), the start-up funds at ODU (R.D.), and USDA-NIFA (Award # 2023-67017-40051; R.D.).
Institutional Review Board Statement
“Not applicable” since this study does not involve humans or animals.
Data Availability Statement
All data are presented in the manuscript and supplementary files.
Acknowledgment
Esma Eser thanks the Scientific and Technological Research Council of Turkey (TUBITAK)–BIDEB under the 2219/ International Postdoctoral Research Fellowship Programme. The authors thank Vignesh Nathan for help with the cell culture experiments.
Conflict of Interest
The authors declare no conflict of interest. “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Bhunia, A. K., Salmonella enterica. In Foodborne Microbial Pathogens: Mechanisms and Pathogenesis, Bhunia, A. K., Ed. Springer New York: New York, NY, 2018; pp 271-287. 10.1007/978-1-4939-7349-1_15.
- Teklemariam, A. D.; Al-Hindi, R. R.; Albiheyri, R. S.; Alharbi, M. G.; Alghamdi, M. A.; Filimban, A. A.; Al Mutiri, A. S.; Al-Alyani, A. M.; Alseghayer, M. S.; Almaneea, A. M.; Albar, A. H.; Khormi, M. A.; Bhunia, A. K., Human Salmonellosis: A Continuous Global Threat in the Farm-to-Fork Food Safety Continuum. Foods 2023, 12, (9), 1756. [CrossRef]
- Popa, G. L.; Papa, M. I., Salmonella spp. infection-A continuous threat worldwide. Germs 2021, 11, (1), 88.
- Guard, J., Through the Looking Glass: Genome, Phenome, and Interactome of Salmonella enterica. Pathogens 2022, 11, (5), 581. 10.3390/pathogens11050581. [CrossRef]
- O'Bryan, C. A.; Ricke, S. C.; Marcy, J. A., Public health impact of Salmonella spp. on raw poultry: Current concepts and future prospects in the United States. Food Control 2022, 132, 108539. [CrossRef]
- Kowalska, B., Fresh vegetables and fruit as a source of Salmonella bacteria. Ann Agric Environ Med 2023, 30, (1), 9-14. [CrossRef]
- Peifer, M., beta-Catenin as Oncogene--The Smoking Gun. Science 1997, 275, (5307), 1752-0.
- CDC, Salmonella. In CDC; https://www.cdc.gov/salmonella/index.html: Atlanta, GA, 2020; Vol. https://www.cdc.gov/salmonella/index.html.
- Guillén, S.; Nadal, L.; Álvarez, I.; Mañas, P.; Cebrián, G., Impact of the Resistance Responses to Stress Conditions Encountered in Food and Food Processing Environments on the Virulence and Growth Fitness of Non-Typhoidal Salmonellae. Foods 2021, 10, (3), 617. [CrossRef]
- Horn, N.; Bhunia, A. K., Food-Associated Stress Primes Foodborne Pathogens for the Gastrointestinal Phase of Infection. Front. Microbiol. 2018, 9, (August), 1962. [CrossRef]
- Gruzdev, N.; Pinto, R.; Sela, S., Effect of desiccation on tolerance of Salmonella enterica to multiple stresses. Appl Environ Microbiol 2011, 77, (5), 1667-1673. [CrossRef]
- Gustavsen, S.; Breen, O., Investigation of an outbreak of Salmonella Oranienburg infections in Norway, caused by contaminated black pepper. Am. J. Epidemiol. 1984, 119, (5), 806-12. 10.1093/oxfordjournals.aje.a113801. [CrossRef]
- McCormic, Z. D.; Patel, K.; Higa, J.; Bancroft, J.; Donovan, D.; Edwards, L.; Cheng, J.; Adcock, B.; Bond, C.; Pereira, E., Bi-national outbreak of Salmonella Newport infections linked to onions: the United States experience. Epidemiol Infect 2022, 150, e199. [CrossRef]
- Mitchell Jr, M. R.; Kirchner, M.; Schneider, B.; McClure, M.; Neil, K. P.; Madad, A.; Jemaneh, T.; Tijerina, M.; Nolte, K.; Wellman, A., Multistate outbreak of Salmonella Oranienburg infections linked to bulb onions imported from Mexico–United States, 2021. Food Control 2024, 160, 110325.
- CDC, Multistate Outbreak of Human Salmonella Montevideo Infections https://www.cdc.gov/salmonella/2010/montevideo-5-4-2010.html.
- Kirk, M. D.; Little, C. L.; Lem, M.; Fyfe, M.; Genobile, D.; Tan, A.; Threlfall, J.; Paccagnella, A.; Lightfoot, D.; Lyi, H.; McIntyre, L.; Ward, L.; Brown, D. J.; Surnam, S.; Fisher, I. S., An outbreak due to peanuts in their shell caused by Salmonella enterica serotypes Stanley and Newport--sharing molecular information to solve international outbreaks. Epidemiol. Infect. 2004, 132, (4), 571-7. 10.1017/s095026880400216x. [CrossRef]
- Scheil, W.; Cameron, S.; Dalton, C.; Murray, C.; Wilson, D., A South Australian Salmonella Mbandaka outbreak investigation using a database to select controls. Aust N Z J Public Health 1998, 22, (5), 536-9. 10.1111/j.1467-842x.1998.tb01434.x. [CrossRef]
- Calhoun, S.; Post, L.; Warren, B.; Thompson, S.; Bontempo, A. R., Prevalence and concentration of Salmonella on raw shelled peanuts in the United States. J. Food Prot. 2013, 76, (4), 575-579.
- CDC, Outbreak of Salmonella Enteritidis Infections Linked to Peaches.
- CDC, Multistate Outbreak of Multidrug-Resistant Salmonella Heidelberg Infections Linked to Foster Farms Brand Chicken https://www.cdc.gov/salmonella/heidelberg-10-13/index.html.
- CDC, Outbreak of Salmonella Heidelberg Infections Linked to Tyson Brand Mechanically Separated Chicken at a Correctional Facility https://www.cdc.gov/salmonella/heidelberg-01-14/index.html.
- CDC, Outbreak of Salmonella Infections Linked to Chicken https://www.cdc.gov/salmonella/chicken-08-18/index.html.
- CDC, Multistate Outbreak of Multidrug-Resistant Salmonella Heidelberg Infections Linked to Foster Farms Brand Chicken https://www.cdc.gov/salmonella/heidelberg-10-13/index.html.
- Mitchell, E.; O'Mahony, M.; Lynch, D.; Ward, L. R.; Rowe, B.; Uttley, A.; Rogers, T.; Cunningham, D. G.; Watson, R., Large outbreak of food poisoning caused by Salmonella typhimurium definitive type 49 in mayonnaise. Brit. Med. J. 1989, 298, (6666), 99-101. 10.1136/bmj.298.6666.99. [CrossRef]
- Mason, B. W.; Williams, N.; Salmon, R. L.; Lewis, A.; Price, J.; Johnston, K. M.; Trott, R. M., Outbreak of Salmonella Indiana associated with egg mayonnaise sandwiches at an acute NHS hospital. Commun Dis Public Health 2001, 4, (4), 300-4.
- Carneiro, M. R.; Cabello, P. H.; Albuquerque-Junior, R. L.; Jain, S.; Candido, A. L., Characterization of a foodborne outbreak caused by Salmonella Enteritidis in Aracaju, State of Sergipe, Brazil. Revista da Sociedade Brasileira de Medicina Tropical 2015, 48, (3), 334-7. 10.1590/0037-8682-0260-2014. [CrossRef]
- von Wissmann, B.; Klinc, C.; Schulze, R.; Wolf, A.; Schreiner, H.; Rabsch, W.; Prager, R.; Hautmann, W., Outbreak of salmonellosis after a wedding party, Bavaria, Germany, summer 2010: the importance of implementing food safety concepts. Eurosurveillance 2012, 17, (6), 20076. doi:. [CrossRef]
- Bhunia, A. K., One day to one hour: how quickly can foodborne pathogens be detected? Fut. Microbiol. 2014, 9, (8), 935-946.
- Al-Hindi, R. R.; Teklemariam, A. D.; Alharbi, M. G.; Alotibi, I.; Azhari, S. A.; Qadri, I.; Alamri, T.; Harakeh, S.; Applegate, B. M.; Bhunia, A. K., Bacteriophage-Based Biosensors: A Platform for Detection of Foodborne Bacterial Pathogens from Food and Environment. Biosensors 2022, 12, (10), 905. 10.3390/bios12100905. [CrossRef]
- Foddai, A. C. G.; Grant, I. R., Methods for detection of viable foodborne pathogens: Current state-of-art and future prospects. Appl. Microbiol. Biotechnol. 2020, 104, 4281-4288. [CrossRef]
- Daquigan, N.; Grim, C. J.; White, J. R.; Hanes, D. E.; Jarvis, K. G., Early Recovery of Salmonella from Food Using a 6-Hour Non-selective Pre-enrichment and Reformulation of Tetrathionate Broth. Front. Microbiol. 2016, 7, (2103). 10.3389/fmicb.2016.02103. [CrossRef]
- Koyuncu, S.; Andersson, M. G.; Häggblom, P., Accuracy and Sensitivity of Commercial PCR-Based Methods for Detection of Salmonella enterica in Feed. Appl Environ Microbiol 2010, 76, (9), 2815. 10.1128/AEM.02714-09. [CrossRef]
- Moreirinha, C.; Trindade, J.; Saraiva, J. A.; Almeida, A.; Delgadillo, I., MIR spectroscopy as alternative method for further confirmation of foodborne pathogens Salmonella spp. and Listeria monocytogenes. J Food Sci Technol 2018, 55, (10), 3971-3978. 10.1007/s13197-018-3322-8. [CrossRef]
- Villamizar-Rodríguez, G.; Fernández, J.; Marín, L.; Muñiz, J.; González, I.; Lombó, F., Multiplex detection of nine food-borne pathogens by mPCR and capillary electrophoresis after using a universal pre-enrichment medium. Front. Microbiol. 2015, 6, (1194). 10.3389/fmicb.2015.01194. [CrossRef]
- Xu, L.; Bai, X.; Bhunia, A. K., Current State of Biosensors Development and their Application in Foodborne Pathogen Detection. J. Food Prot. 2021, 84, (7), 1213-1227. 10.4315/JFP-20-464. [CrossRef]
- Eijkelkamp, J. M.; Aarts, H. J. M.; van der Fels-Klerx, H. J., Suitability of Rapid Detection Methods for Salmonella in Poultry Slaughterhouses. Food Anal. Methods 2009, 2, (1), 1-13. 10.1007/s12161-008-9040-5. [CrossRef]
- Xu, L.; Bai, X.; Tenguria, S.; Liu, Y.; Drolia, R.; Bhunia, A. K., Mammalian cell-based immunoassay for detection of viable bacterial pathogens. Front. Microbiol. 2020, 11, 575615. 10.3389/fmicb.2020.575615. [CrossRef]
- Bhunia, A. K.; Singh, A. K.; Parker, K.; Applegate, B. M., Petri-plate, bacteria, and laser optical scattering sensor. Front. Cell. Infect. Microbiol. 2022, 12, 1087074. 10.3389/fcimb.2022.1087074. [CrossRef]
- To, C.; Banerjee, P.; Bhunia, A. K., Cell-Based Biosensor for Rapid Screening of Pathogens and Toxins. In Handbook of Cell Biosensors, Thouand, G., Ed. Springer International Publishing: Cham, 2020; pp 1-16. 10.1007/978-3-319-47405-2_102-1. [CrossRef]
- Lu, X.; Ye, Y.; Zhang, Y.; Sun, X., Current research progress of mammalian cell-based biosensors on the detection of foodborne pathogens and toxins. Crit. Rev. Food Sci. Nutr. 2021, 61, (22), 3819-3835. [CrossRef]
- Banerjee, P.; Bhunia, A. K., Mammalian cell-based biosensors for pathogens and toxins. Trends Biotechnol. 2009, 27, (3), 179-188. [CrossRef]
- Burkholder, K.; Bhunia, A., Salmonella enterica serovar Typhimurium adhesion and cytotoxicity during epithelial cell stress is reduced by Lactobacillus rhamnosus GG. Gut Pathog. 2009, 1, (1), 14. [CrossRef]
- Birhanu, B. T.; Park, N.-H.; Lee, S.-J.; Hossain, M. A.; Park, S.-C., Inhibition of Salmonella Typhimurium adhesion, invasion, and intracellular survival via treatment with methyl gallate alone and in combination with marbofloxacin. Vet. Res. 2018, 49, (1), 101. 10.1186/s13567-018-0597-8. [CrossRef]
- Chen, L. M.; Kaniga, K.; Galan, J. E., Salmonella spp. are cytotoxic for cultured macrophages. Mol. Microbiol. 1996, 21, (5), 1101-1115. [CrossRef]
- Yang, J.; Yan, H., TLR5: beyond the recognition of flagellin. Cell. Mol. Immunol. 2017, 14, (12), 1017-1019. [CrossRef]
- Song, W. S.; Jeon, Y. J.; Namgung, B.; Hong, M.; Yoon, S. I., A conserved TLR5 binding and activation hot spot on flagellin. Sci Rep 7: 40878. In 2017. [CrossRef]
- Gray, K. M.; Bhunia, A. K., Specific detection of cytopathogenic Listeria monocytogenes using a two-step method of immunoseparation and cytotoxicity analysis. J. Microbiol. Methods 2005, 60, (2), 259-268. [CrossRef]
- Mendonca, M.; Conrad, N.; Conceicao, F.; Moreira, A.; da Silva, W.; Aleixo, J.; Bhunia, A., Highly specific fiber optic immunosensor coupled with immunomagnetic separation for detection of low levels of Listeria monocytogenes and L. ivanovii. BMC Microbiol. 2012, 12, (1), 275. [CrossRef]
- de Cássia Dos Santos da Conceição, R.; Moreira, A. N.; Ramos, R. J.; Goularte, F. L.; Carvalhal, J. B.; Aleixo, J. A. G., Detection of Salmonella sp in chicken cuts using immunomagnetic separation. Braz J Microbiol 2008, 39, (1), 173-177. 10.1590/S1517-838220080001000034. [CrossRef]
- Mansfield, L. P.; Forsythe, S. J., The detection of Salmonella using a combined immunomagnetic separation and ELISA end-detection procedure. Lett Appl Microbiol 2000, 31, (4), 279-283. 10.1046/j.1472-765x.2000.00811.x. [CrossRef]
- Ha, D.-G.; Kuchma, S. L.; O’Toole, G. A., Plate-Based Assay for Swimming Motility in Pseudomonas aeruginosa. In Pseudomonas Methods and Protocols, Filloux, A.; Ramos, J.-L., Eds. Springer New York: New York, NY, 2014; pp 59-65. 10.1007/978-1-4939-0473-0_7. [CrossRef]
- Kearns, D. B.; Losick, R., Swarming motility in undomesticated Bacillus subtilis. Mol. Microbiol. 2003, 49, (3), 581-90. 10.1046/j.1365-2958.2003.03584.x. [CrossRef]
- Singh, A. K.; Bettasso, A. M.; Bae, E.; Rajwa, B.; Dundar, M. M.; Forster, M. D.; Liu, L.; Barrett, B.; Lovchik, J.; Robinson, J. P.; Hirleman, E. D.; Bhunia, A. K., Laser optical sensor, a label-free on-plate Salmonella enterica colony detection tool. mBio 2014, 5 (1), (1), e01019-13. 10.1128/mBio.01019-13. [CrossRef]
- Andersen-Nissen, E.; Smith, K. D.; Strobe, K. L.; Barrett, S. L. R.; Cookson, B. T.; Logan, S. M.; Aderem, A., Evasion of Toll-like receptor 5 by flagellated bacteria. Proc. Nat. Acad. Sci. USA 2005, 102, (26), 9247-9252. [CrossRef]
- Hahm, B. K.; Bhunia, A. K., Effect of environmental stresses on antibody-based detection of Escherichia coli O157:H7, Salmonella enterica serotype Enteritidis and Listeria monocytogenes. J. Appl. Microbiol. 2006, 100, (5), 1017-1027. [CrossRef]
- USDA-FSIS, Isolation and Identification of Salmonella from Meat, Poultry, Pasteurized Egg and Catfish Products. Method number MLG 4.06. In http://www.fsis.usda.gov/wps/wcm/connect/700c05fe-06a2-492a-a6e1-3357f7701f52/MLG-4.pdf?MOD=AJPERES: 2013.
- Bell, R. L.; Jarvis, K. G.; Ottesen, A. R.; McFarland, M. A.; Brown, E. W., Recent and emerging innovations in Salmonella detection: a food and environmental perspective. Microb.Biotechnol. 2016, 9, (3), 279-292. [CrossRef]
- FDA, Bacteriological Analytical Manual Online, 8th ed In AOAC International, Arlington, VA. [http://www.fda.gov/Food/ScienceResearch/LaboratoryMethods/BacteriologicalAnalyticalManualBAM/default.htm]: 2001. http://www.cfsan.fda.gov/~ebam/bam-toc.html.
- Rajapaksha, P.; Elbourne, A.; Gangadoo, S.; Brown, R.; Cozzolino, D.; Chapman, J., A review of methods for the detection of pathogenic microorganisms. Analyst 2019, 144, (2), 396-411. [CrossRef]
- Ricke, S. C.; Kim, S. A.; Shi, Z.; Park, S. H., Molecular-based identification and detection of Salmonella in food production systems: current perspectives. J. Appl. Microbiol. 2018, 125, (2), 313-327. 10.1111/jam.13888. [CrossRef]
- Lee, K.-M.; Runyon, M.; Herrman, T. J.; Phillips, R.; Hsieh, J., Review of Salmonella detection and identification methods: Aspects of rapid emergency response and food safety. Food Control 2015, 47, 264-276. [CrossRef]
- Buzby, J. C.; Farah-Wells, H.; Hyman, J., The estimated amount, value, and calories of postharvest food losses at the retail and consumer levels in the United States. USDA-ERS Economic Information Bulletin Number 121 2014, (121).
- Elkhishin, M. T.; Gooneratne, R.; Hussain, M. A., Microbial safety of foods in the supply chain and food security. Adv Food Technol Nutr Sci Open J 2017, 3, (1), 22-32. [CrossRef]
- Bhunia, A. K., Rapid pathogen screening tools for food safety. Food Technol. 2011, 65, (2), 38-43.
- Hayashi, F.; Smith, K. D.; Ozinsky, A.; Hawn, T. R.; Yi, E. C.; Goodlett, D. R.; Eng, J. K.; Akira, S.; Underhill, D. M.; Aderem, A., The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, (6832), 1099-1103. [CrossRef]
- Smith, K. D.; Andersen-Nissen, E.; Hayashi, F.; Strobe, K.; Bergman, M. A.; Barrett, S. L. R.; Cookson, B. T.; Aderem, A., Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat. Immunol. 2003, 4, (12), 1247-1253.
- Yoon, S.-i.; Kurnasov, O.; Natarajan, V.; Hong, M.; Gudkov, A. V.; Osterman, A. L.; Wilson, I. A., Structural basis of TLR5-flagellin recognition and signaling. Science 2012, 335, (6070), 859-864. [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R., PCR inhibitors–occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, (5), 1014-1026.
- Lin, A.; Singh, A.; Allred, A.; Allard, M.; Waltman, D.; Imanian, B.; Ng, J. H. J.; Sanahmadi, Y.; Khaksar, R., Targeted Next Generation Sequencing Assay for Direct Detection and Serotyping of Salmonella from Enrichment. J. Food Prot. 2024, 100256. [CrossRef]
- Vinayaka, A. C.; Ngo, T. A.; Kant, K.; Engelsmann, P.; Dave, V. P.; Shahbazi, M. A.; Wolff, A.; Bang, D. D., Rapid detection of Salmonella enterica in food samples by a novel approach with combination of sample concentration and direct PCR. Biosens Bioelectron 2019, 129, 224-230. 10.1016/j.bios.2018.09.078. [CrossRef]
- Kuang, H.; Cui, G.; Chen, X.; Yin, H.; Yong, Q.; Xu, L.; Peng, C.; Wang, L.; Xu, C., A one-step homogeneous sandwich immunosensor for Salmonella detection based on magnetic nanoparticles (MNPs) and quantum Dots (QDs). Int J Mol Sci 2013, 14, (4), 8603-8610. 10.3390/ijms14048603. [CrossRef]
- Cimaglia, F.; De Lorenzis, E.; Mezzolla, V.; Rossi, F.; Poltronieri, P., Detection of L. monocytogenes in Enrichment Cultures by Immunoseparation and Immunosensors. IEEE Sensors J. 2016, 16, (19), 7045-7052. 10.1109/JSEN.2016.2598700. [CrossRef]
Figure 4.
Limit of detection analysis of HEK dual TLR5 sensors to detect Salmonella from spiked chicken skin and peanut butter samples. (a) TLR 5 sensors response after 4 and 6 h incubation of spiked chicken samples. Plate image representative showing color development at 6 h (below). (b) TLR 5 sensors response after 4 and 6 h incubation of spiked peanut butter samples. Data in a-b represent the average results from three independent experiments presented as mean ± SEM.
Figure 4.
Limit of detection analysis of HEK dual TLR5 sensors to detect Salmonella from spiked chicken skin and peanut butter samples. (a) TLR 5 sensors response after 4 and 6 h incubation of spiked chicken samples. Plate image representative showing color development at 6 h (below). (b) TLR 5 sensors response after 4 and 6 h incubation of spiked peanut butter samples. Data in a-b represent the average results from three independent experiments presented as mean ± SEM.
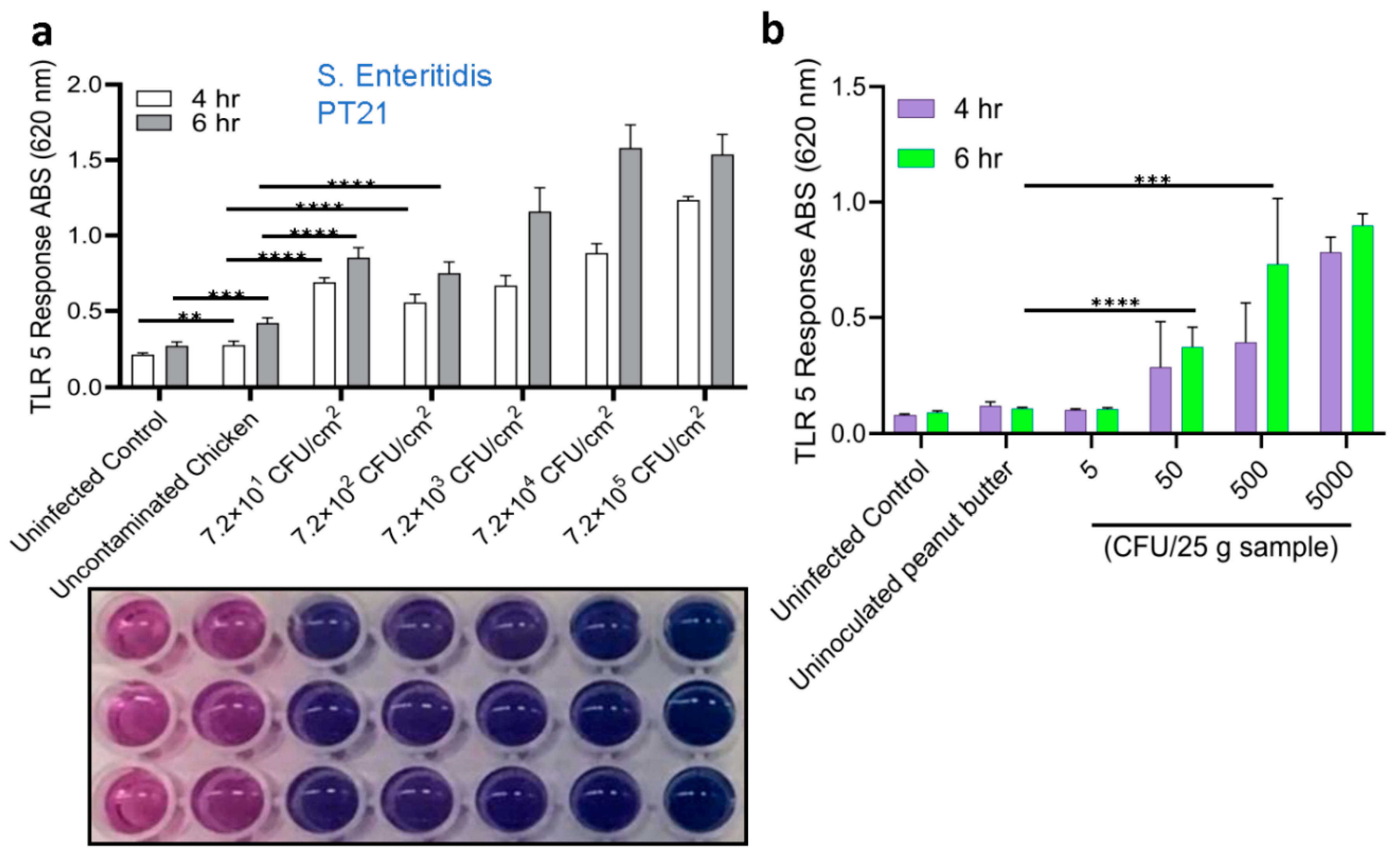
Figure 5.
Detection of Salmonella using HEKdualTLR 5 sensor from spiked food samples. (a) TLR 5 sensors response after 6 h incubation of spiked food samples Spiked food samples were subjected to 4-h pre-enrichment and 4-h selective enrichment (Initial contamination level was ~100 CFU /25 g food sample). Salmonella was captured and concentrated using immunomagnetic beads (IMB) in tests with IMS. Data represent the average results from three independent experiments presented as mean ± SEM. (b) Representative microscopic images show an intact cell monolayer treated with bacterial suspension concentrated from all food samples.
Figure 5.
Detection of Salmonella using HEKdualTLR 5 sensor from spiked food samples. (a) TLR 5 sensors response after 6 h incubation of spiked food samples Spiked food samples were subjected to 4-h pre-enrichment and 4-h selective enrichment (Initial contamination level was ~100 CFU /25 g food sample). Salmonella was captured and concentrated using immunomagnetic beads (IMB) in tests with IMS. Data represent the average results from three independent experiments presented as mean ± SEM. (b) Representative microscopic images show an intact cell monolayer treated with bacterial suspension concentrated from all food samples.
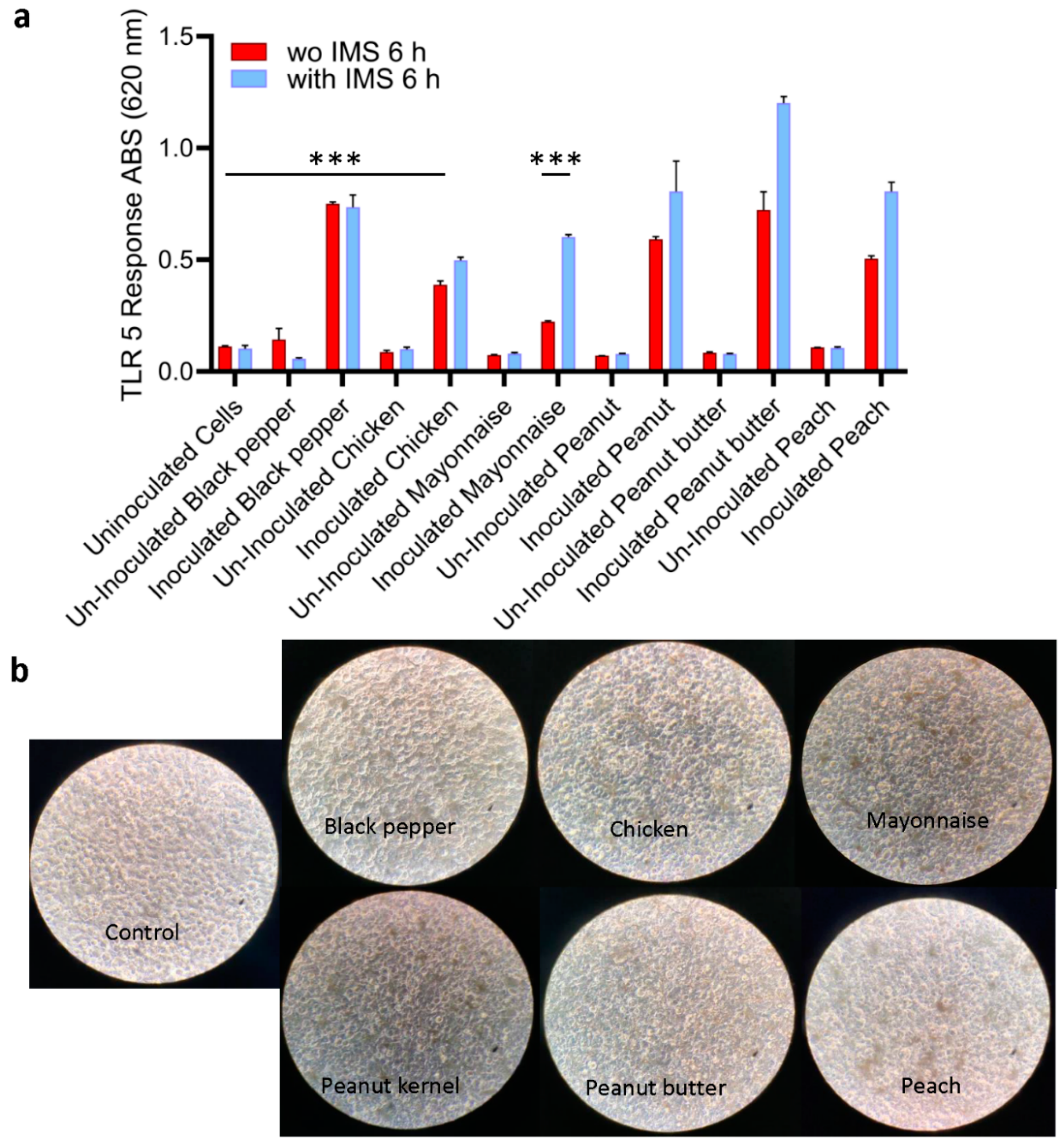

Table 1.
Effect of 3 h environmental stress on Salmonella Enteritidis PT21 viability and growth.
| Log 10 (CFU/mL) ± SEM (N=3) | |||||||
| No stress | Cold Stress | Heat Stress | Low pH | Ethanol | NaCL | H2O2 | |
| 3 h stress | 9.86±0.19 | 9.38±0.21 | 9.35±0.11 | 9.67±0.14 | 9.60±0.22 | 9.24±0.28 | 9.33±0.21 |
| 3 h stress+3 h enrichment | 10.49±0.22 | 10.44±0.24 | 10.01±0.21 | 10.21±0.24 | 9.69±0.29 | 10.33±0.19 | 10.65±0.24 |
Table 2.
Recovery of Salmonella Enteritidis PT21 from contaminated peanut butter at each step of sample enrichment.
Table 2.
Recovery of Salmonella Enteritidis PT21 from contaminated peanut butter at each step of sample enrichment.
| Log 10 (CFU/ml) ± SEM (N=3) | ||||
| Inoculation Level (CFU/25g sample) | Pre-Enriched | Enriched | ||
| TSA | XLD | TSA | XLD | |
| 0 | No growth | No growth | No growth | No growth |
| ~5 | No growth | No growth | 2.86±0.21 | 3.09±0.18 |
| ~50 | 2.68±0.17 | 2.51±0.11 | 4.06±0.14 | 4.18±0.19 |
| ~500 | 3.94±0.21 | 3.85±0.19 | 6.32±0.22 | 6.39±0.24 |
| ~500 | 4.35±0.13 | 4.44±0.18 | 6.60±0.23 | 6.42±0.14 |
TSA, Tryptic soy agar; XLD, Xylose lysine deoxycholate.
Table 3.
Recovery of Salmonella Enteritidis PT21 from spiked (~100 CFU/25 g, except peach (~100 CFU/100 g) food samples at each step of sample enrichment and concentration.
Table 3.
Recovery of Salmonella Enteritidis PT21 from spiked (~100 CFU/25 g, except peach (~100 CFU/100 g) food samples at each step of sample enrichment and concentration.
| Log 10 (CFU/ml)± SEM (N=3) | ||||||||
| Food Sample | After Selective Enrichment | Without IMS |
IMS |
Counts (CFU/100 µL) |
PCR (invA 796 bp) |
|||
| TSA | XLD | TSA | XLD | TSA | XLD | XLD | ||
| Black pepper (U) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Black pepper (I) | 3.91±0.18 | 4.04±0.11 | 3.87±0.14 | 4.05±0.21 | 3.98±0.08 | 4.07±0.14 | 3.07±0.11 | - |
| Chicken (U) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Chicken (I) | 3.85±0.15 | 3.92±0.08 | 3.94±0.19 | 4.10±0.21 | 3.91±0.15 | 4.10±0.11 | 3.1±0.09 | - |
| Mayonnaise (U) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Mayonnaise (I) | 3.94±0.22 | 4.02±0.14 | 3.72±0.18 | 3.94±0.12 | 3.72±0.21 | 3.92±0.18 | 2.92±0.13 | - |
| Peanut kernel(U) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Peanut kernel (I) | 3.92±0.26 | 4.09±0.19 | 4.26±0.22 | 4.29±0.18 | 4.07±0.14 | 4.13±0.19 | 3.13±0.18 | + |
| Peanut butter (U) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Peanut butter (I) | 5.53±0.24 | 5.18±0.18 | 5.36±0.25 | 5.46±0.28 | 5.39±0.22 | 5.47±0.24 | 4.47±0.16 | - |
| Peach (U) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Peach (I) | 3.71±0.14 | 3.88±0.16 | 3.72±0.12 | 4.08±0.19 | 3.91±0.16 | 4.33±0.14 | 3.33±0.11 | - |
U, uninoculated; I, inoculated; IMS, Immunomagnetic separation.
Table 4.
List of bacterial strains used in the study from our culture collection.
| Salmonella enterica serovars | Non-Salmonella cultures |
| Enteritidis PT4 | Escherichia coli O157 H¯493-89 (flagella Negative control) |
| Enteritidis PT8 | E. coli O26:H11 00971 |
| Enteritidis PT6 | E. coli O103:H2 90-3128 |
| Enteritidis PT7 | E. coli O26 05-6544 |
| Enteritidis PT28 | E. coli O103:H11 SJ12 |
| Enteritidis PT21 | E. coli O157:H7 204P |
| Enteritidis 13ENT1344 | E. coli O157:H7 505B |
| Typhimurium 13ENT906 | E. coli O157:H7 ATCC 43895 |
| Typhimurium ST1 | E. coli O157:H7 ATCC 43890 |
| Agona 12ENT1356 | Citrobacter freundii ATCC 3624 |
| Hadar 13ENT979 | Citrobacter freundii ATCC43864 |
| Paratyphi 11J85 | Citrobacter freundii ATCC8090 |
| Bareilly 12ENT1164 | Hafnia alvei |
| Pullorum DUP-PVUII 1006 | Proteus vulgaris DUP-10086 |
| Newport 13ENT1060 | Proteus mirabilis B-3402 |
| Heidelberg 18ENT1418 | Serratia marcescens ATCC 8100 |
| Saintpaul 13ENT1045 | S. marcescens B-2544 |
| Bradenderup 12ENT1138 | S. marcescens ATCC 43862 |
| Infantis 13ENT866 | Klebsiella pneumoniae B-41958 |
| Javiana 13ENT86F | Listeria monocytogenes 104033S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



