
Submitted:
07 August 2024
Posted:
09 August 2024
You are already at the latest version
Abstract
The genus Chaetogaster belongs to the subfamily Naidinae (Naididae), includes mostly species of small size and is diverse and abundant in surface coarse sediments in streams. The aim of the present study is to initiate an inventory of lineages of Chaetogaster in Switzerland. We used 135 specimens collected at six sites in four streams of four cantons. We sequenced the cytochrome c oxidase (COI) gene from all specimens and ITS2 and rDNA 28S from all or several specimens of each lineage delimited using COI data and preserved a voucher of almost all sequenced specimens. We could delimit one lineage (=species) for Chaetogaster diaphanus (Gruithuisen, 1828), three within Chaetogaster diastrophus (Gruithuisen, 1828), one for Chaetogaster langi Brestcher, 1896, one for Chaetogaster setosus Svetlov, 1925 and three unidentified Chaetogaster spp. Two lineages of Chaetogaster spp. could correspond to a new morphological group, but this should be confirmed on more specimens. We described the three C. diastrophus lineages and two Chaetogaster spp. as new species for science. The prospects of the present work are to complete the data of the molecular diversity of this genus in Switzerland and to describe on a molecular/morphological basis the new Chaetogaster species that we will probably find.
Keywords:
aquatic oligochaetes
; Chaetogaster
; cryptic diversity
; molecular systematics
; genetic lineages
; new species
1. Introduction
The genus Chaetogaster von Baer, 1827 belongs to the subfamily of Naidinae (Naididae) and includes species of small size (0.5 – 4 mm long, 0.08 – 0.6 mm wide) that reproduce mainly asexually by paratomy (budding and fission), are predators of other invertebrates and/or feed on algae [1,2]. They are frequently found as part as biomonitoring surveys in streams in fine to coarse sediments [3]. Chaetogaster species (except C. parvus Pointner, 1914) are characteristic of surface habitats and their presence in the hyporheic zone indicates infiltration of surface water [4]. In addition, they are all considered as sensitive to moderately resistant to chemical pollution [4].
In total, eighteen species of Chaetogaster have been described based on morphological features [5]. In Switzerland, six species of Chaetogaster have been recorded in streams so far: C. diaphanus (Gruithuisen, 1828), C. diastrophus (Gruithuisen, 1828), C. langi Bretscher, 1896, C. parvus Pointner, 1914, C. limnaei (von Baer, 1827) and C. setosus Svetlov, 1925. While C. diaphanus, C. diastrophus and C. langi are frequent, C. parvus has been only found in some streams [3,6], C. setosus at two sites in one stream [7] and C. limnaei at one stream site [3].
Recently, Mack et al. [2] suggested that three commonly referenced species in North America and Europe, C. diaphanus, C. diastrophus and C. limnaei, are a complex of mostly 24 cryptic species. They sequenced mitochondrial and nuclear markers of 128 Chaetogaster specimens from ponds, lake shores and creeks at many different locations in North America (Maryland) and Europe (mostly Scandinavian countries) and found two cryptic species for C. diaphanus, 3 for C. limnaei and 19 for C. diastrophus. In other words, the overall species diversity of Chaetogaster has been largely underestimated on morphological grounds. For this reason, we initiated an inventory of lineages of Chaetogaster in Switzerland based on a DNA barcoding approach.
The aim of this study is not only to reveal the hidden diversity among Chaetogaster in Switzerland but also to genetically characterize the species C. langi and C. setosus and to attribute to them DNA barcode sequences. We selected 33 specimens of C. diaphanus, 67 of C. diastrophus, 30 of C. langi and 5 of C. setosus collected at six sites in four streams of four cantons. We sequenced COI from all specimens and ITS2 and 28S from all or several specimens of each lineage delimited using COI data. We preserved and analyzed morphologically the anterior part of almost all sequenced specimens. We also described five lineages we found within Chaetogaster as a new species for science on a DNA sequence basis.
2. Material and Methods
2.1. Specimen Collection
We used for present study 135 Chaetogaster specimens collected in surface coarse sediments at six different sites in four different streams in Canton of Vaud (Sorge River, one site), Canton of Bern (Suze River, on site), Canton of St-Gallen (Glatt River, 4 sites) and Canton Aargau (Bünz River, one site) as part of surveys for biomonitoring purposes (Table S1). The sediments were collected using a shovel and oligochaetes were fixed in the field with neutral buffered formalin (final concentration of formalin 10% (=formaldehyde of 4%) in the recipients). Formalin optimally fixes oligochaete specimens, and a study showed that fixation and storage of oligochaete specimens in 4% neutral buffered formalin for up to 30 days was suitable for subsequent genetic analyses [8].
In the laboratory, sediments were sieved through a column of 5 and 0.2 mm mesh size sieves. The material retained on the 0.2 mm mesh size sieve was transferred to a Tupperware box and absolute ethanol was added to obtain a concentration of ethanol of about 100%. These boxes were preserved at -20oC. We selected in these samples 33 specimens of C. diaphanus, 67 of C. diastrophus, 30 of C. langi and 5 of C. setosus using a stereomicroscope. C. diaphanus, C. langi and C. setosus are recognizable using a stereomicroscope, but it is impossible to distinguish C. diastrophus from C. limnaei and C. parvus using the magnification of a stereomicroscope. The “C. diastrophus” specimens were therefore supposed to belong to this morphospecies (morphospecies = group of morphologically identical species), as according to the morphological inventories it is much more frequent in Switzerland than C. limnaei and C. parvus.
Each specimen (except one specimen, Chaetogaster sp. MOTU 8) was cut into two pieces, the anterior part for morphological analysis and the posterior part for genetic analyses (preserved in absolute ethanol at -20oC in Eppendorf Tubes).
2.2. Morphological Analysis
The anterior parts were mounted on slides in a coating solution (containing lactic acid, glycerol and Mowiol) [9] and specimens were identified to species or morphospecies level using a compound microscope. The anterior parts of all specimens (vouchers), except isolates Glatt6, Nos 29, 30, 37, 38, 155, 204, 205, are deposited in the “Muséum cantonal des sciences naturelles” of Lausanne (Table S1). Almost all vouchers were photographed with an Olympus SC50 camera, 10x and 40x magnification (photos available upon request).
The genus Chaetogaster is characterized by a fusion of the first five segments forming a head containing a muscular pharynx, by the absence of ventral chaetae in segments III-V and by overall absence of dorsal chaetae [10]. C. diaphanus is different from the other species of this genus (found in the present study) by its much larger size (length and width; see below). C. diaphanus, C. langi and C. setosus have no or a vestigial prostomium, while C. diastrophus has a distinct prostomium. C. setosus has simple pointed, long and prominent ventral chaetae (compared to its body) while C. diaphanus, C. diastrophus and C. langi have bifid and no prominent ventral chaetae. Based on these characters, we have created the following key to identify C. diaphanus, C. diastrophus, C. langi and C. setosus. The mentioned sizes in the key correspond to the specimens of the present study (fixed material).
1 Body size: 1.5-4 mm long and 0.3-0.6 mm wide (at head); prostomium absent, buccal cavity large C. diaphanus
- Body size: 0.5-1.2 mm long, 0.08-0.16 mm wide (at head); prostomium present or absent; buccal cavity large or not large 2
2(1) Simple-pointed chaetae C. setosus
- Bifid chaetae 3
3(2) Body size: 0.6-1.2 mm long, 0.11-0.16 mm wide (at head); prostomium present; buccal cavity not large to large C. diastrophus
- Body size 0.5-0.7 mm long, 0.08-0.11 mm wide (at head); prostomium absent or presence of a vestigial prostomium (notch in its place at the edge of the mouth); buccal cavity generally large C. langi
2.3. Molecular Analyses
2.3.1. DNA Extraction and Sequencing
We sequenced a fragment of 658 base pairs of the mitochondrial COI gene from 127 specimens, a fragment of 313 base pairs of the mitochondrial COI gene from 7 specimens, a fragment of 558 base pairs of the nuclear ITS2 region from 112 specimens and a fragment of 796 base pairs of the 28S gene from 67 specimens. ITS2 and 28S were sequenced from all or several specimens per lineage delimited using COI sequences.
Total genomic DNA was extracted using guanidine thiocyanate as described by Tkach & Pawlowski [11]. We used the LCO 1490 and HCO 2198 primers [12] for amplifying the COI fragment of 658 bp, the “mlCOIintF” and “jgHCO2198” primers [13] for amplifying the COI fragment of 313 bp, the ITS2-1 and ITS2-2 primers [14] for amplifying the ITS2 rDNA region and the 28SC1 et 28SD2 primers [15] for amplifying the 28S region.
PCR amplifications were performed in a total volume of 20 μl containing 0.2 μl of Taq polymerase 5U/μl (Roche, Basel, Switzerland), 2 μl of the PCR buffer (10x concentrated) with MgCl2 (Roche), 0.5 μl of each primer (10 μM each), 0.4 μl of a mix containing 10 mM of each dNTP (Roche) and 1 μl of DNA template. The PCR comprised an initial denaturation step at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 40 s, annealing at 44°C for 45 s and elongation at 72°C for 1 min and a final elongation step at 72°C for 8 min.
COI, ITS2 and 28S products were then bi-directionally Sanger-sequenced on an Applied Biosystems 96-capillary 3730xl instrument by the company Fasteris (Switzerland) using the same primers (mentioned above) and following the manufacturer’s protocol. The raw sequence editing and the generation of contiguous sequences were accomplished using CodonCode Aligner (CodonCode Corporation). All ITS2 and 28S sequences were obtained by direct sequencing.
The COI, ITS2 and 28S sequences are provided as Supplemental Files S1-S3 and are accessible in GenBank: COI: PP996388-PP996521; ITS2: PQ013378-PQ013490; 28S: PQ013491-PQ013557.
2.3.2. Molecular Phylogeny
All sequences were aligned for each marker using the MUSCLE algorithm (default options) [16] implemented in Seaview v. 5.0.5 [17]. For each marker, a phylogenetic tree was inferred by maximum likelihood using IQ-TREE v. 2.2.0 for macOS [18,19], as well as optimization of its parameters, and data partitioned according to codon position when relevant (COI: TPM3+F+G4 (codon 1), TNe+G4 (codon 2), F81+F+I (codon 3); ITS2: TNe+I+G4; 28S: TIM3+F+I+G4). Branch support was obtained with the ultrafast bootstrap with 1000 replicates [20].
2.3.3. Distance Analysis
Uncorrected pairwise genetic distances in COI and ITS2 were calculated using MEGA 11 [21]. Genetic distances were calculated between sequences within consensus MOTUs (Molecular Operational Taxonomic Units) and between sequences of the different MOTUs as identified by the single-locus approaches (see below). To estimate the barcoding gap of each MOTU, we determined the maximal genetic distance within sequences of a same MOTU (=maximal intra-MOTU variation) and the minimal genetic distance between sequences of a MOTU and sequences of the other MOTUs (=minimal inter-MOTU variation). Genetic distances were also computed between the COI (Chaetogaster) sequences obtained by Mack et al. [2] and the COI sequences of our Chaetogaster MOTUs, to study if our lineages were new or already found by Mack et al. [2]. For this analysis, COI sequences diverging by less than 5% of genetic divergence were considered as belonging to the same lineage.
2.3.4. Single-Locus Species Delimitation
Species were delineated following two complementary approaches [22]: a distance-based method, ASAP [23], and a tree-based method, the “Poisson Tree Processes” (PTP) method [24]. Although very popular over the last two decades, the GMYC approach [25] has not been used here because the method has been shown on several occasions to have a strong tendency to oversplit species [26,27], in particular to delimit oligochaete species [28], being extremely parameter-sensitive [29].
ASAP was run using p-distances as well as both the Jukes-Cantor (JC69) and the Kimura 2-parameter (K80) substitution models to compute the distances, in order to investigate the possible impact of different distance models on the partitioning. Analyses were performed on the dedicated public web server (https://bioinfo.mnhn.fr/abi/public/asap/).
PTP analyses were performed using multi-rate PTP (mPTP), which, unlike PTP, considers differences in intraspecific variation, due to the evolutionary history or sampling of each species. mPTP is presented as an improvement on the single-rate model PTP [24], making possible to obtain more accurate estimates than the latter [30]. PTP analyses do not require an ultrametric input tree, which is a potentially error-prone process. Therefore, the phylogenetic tree produced by the I-QTREE analysis was used as the input tree for analyses based on the PTP model. The stand-alone version of mPTP was preferred to its web implementation because certain functionalities are not available in the web service (https://mptp.h-its.org/). The last release of the pre-compiled macOS binary (mPTP 0.2.4) was downloaded from GitHub (https://github.com/Pas-Kapli/mptp).
3. Results
3.1. Delimitation of Lineages
Species delimitation based on the COI gene leads to the identification of 10 MOTUs, either with ASAP or mPTP, which correspond to strongly supported singletons or clades in the ML tree (BV: 98-100) (Figure 1). The ASAP analyses consistently suggest the same partitioning into 10 different MOTUs, regardless of how the distances were estimated (p-distances, JC69, K80). While the grouping of specimens identified as C. langi and C. setosus remains consistent with their morphological identifications, the morphospecies C. diaphanus and C. diastrophus split into 2 (MOTU6, MOTU7) and 3 (MOTU3, MOTU9, MOTU10) groups, respectively. ASAP analyses also show a clear barcoding gap (p-distances: 4 - 9%; JC69: 5 - 9%; K2P: 4 -10%). The correspondence of each lineage found in the present study to the species (=lineages) described by Mack et al. [2] is indicated in Table 1 and the mean genetic variations between lineages (present study and Mack et al. [2]) are provided in Table S2.
Delineations based on ITS2 and 28S confirm these results, except for the morphospecies C. diaphanus which appears as a single lineage, so that only 9 MOTUs are delineated based on nuclear genes (Figures 2 and S1).
Figure 2.
Molecular phylogeny constructed using the maximum likelihood method and ITS2 gene fragment of Chaetogaster specimens. Partitions at the right side of the figure represent the results of the species delimitation analyses with single-locus methods (ASAP, mPTP). For comparison purposes, the MOTU numbers correspond to those used for the MOTUs identified from the COI. Numbers at nodes are ultrafast bootstrap values (BV). Nodes were considered as supported if BVs were higher or equal to 90 [20]. For the sake of clarity, BVs are not shown within delimited MOTUs.
Figure 2.
Molecular phylogeny constructed using the maximum likelihood method and ITS2 gene fragment of Chaetogaster specimens. Partitions at the right side of the figure represent the results of the species delimitation analyses with single-locus methods (ASAP, mPTP). For comparison purposes, the MOTU numbers correspond to those used for the MOTUs identified from the COI. Numbers at nodes are ultrafast bootstrap values (BV). Nodes were considered as supported if BVs were higher or equal to 90 [20]. For the sake of clarity, BVs are not shown within delimited MOTUs.
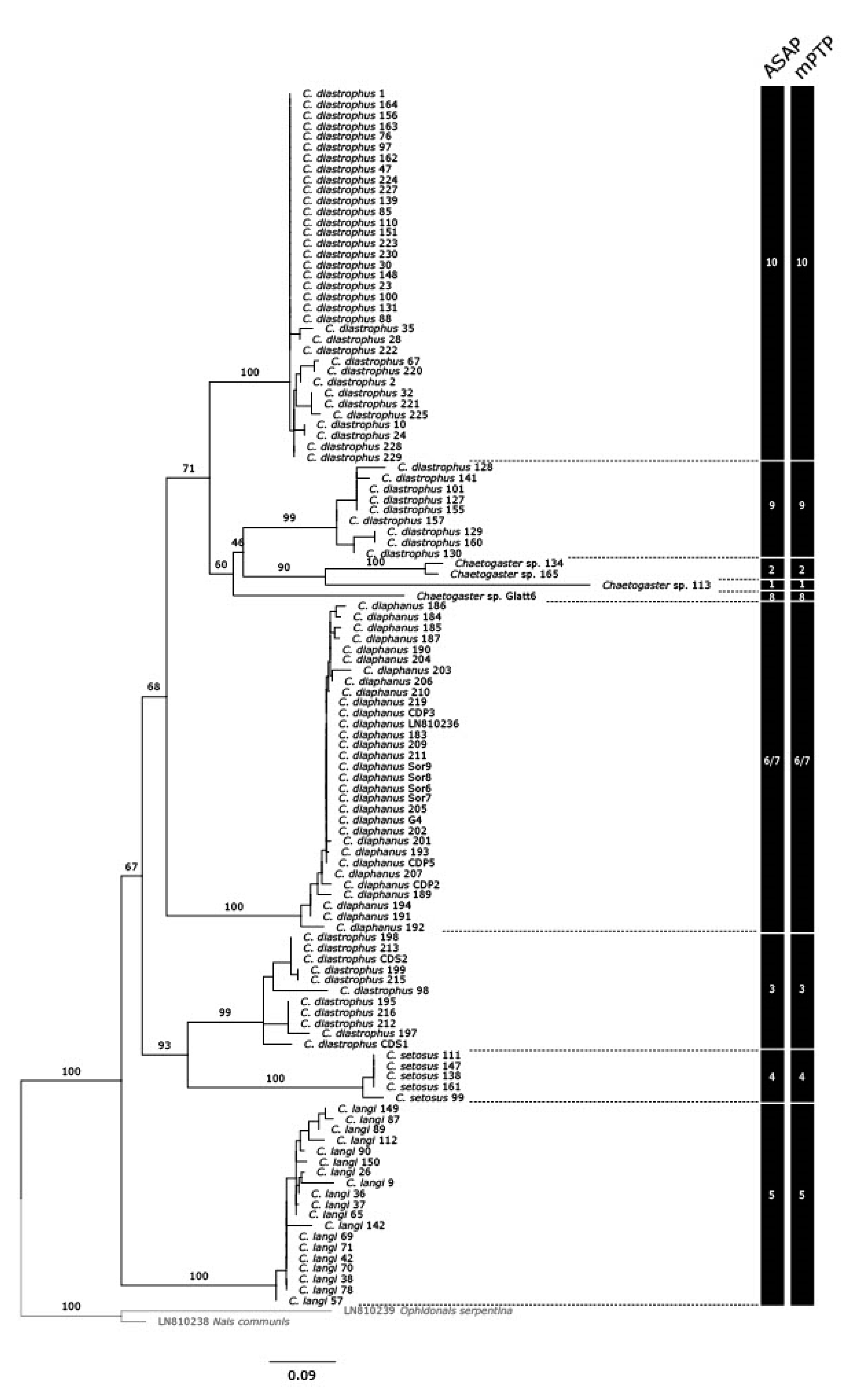
ITS2 produces identical delineations with both ASAP and mPTP. These delimitations also correspond to strongly supported singletons or clades in the ML tree (BV: 99-100). A barcoding gap remains, albeit narrower than that observed on the basis of COI (p-distances, JC69: 6-8%; K2P: 8-10%).
28S produced less consistent results between methods than ITS2. A discordance is observed in the ASAP analyses depending on whether the evolutionary model is p-distances and K2P (9 MOTUs identified) or JC69 (8 MOTUs by merging MOTU7 and MOTU8). However, it should be noted that, in the latter case, the second-best solution from the ASAP analysis delimits 9 MOTUs again. These MOTUs correspond to singletons or clades strongly supported in the ML tree (BV: 98-100), with the exception of M10 which forms a clade supported by a BV of 94. In contrast, the mPTP approach leads to inconsistent results, both in relation to the ASAP analyses or in relation to the clades identified by the ML analysis: the specimens identified as C. setosus (MOTU4) are split between two lineages, despite their genetic proximity, and, in addition to the lumping of MOTU6 and MOTU7 already observed with the ITS2, a new grouping occurs between MOTU5, MOTU8, MOTU9 and MOTU10, which appear as a single lineage; C. langi no longer appears as a distinct species. There is no visible barcoding gap.
3.2. Distance Analyses
In COI, the maximum intra-MOTU variations were low (max. 3.9%) and minimal inter-MOTU variations high (11.5%-14.9%) except for the two MOTUs of C. diaphanus (9.8%) (Table 1). In ITS2, the maximum intra-MOTU variations were between 2.7% and 6.2% and minimal inter-MOTU variations between 7.5% and 13.9%.
3.3. Morphological vs. Molecular Identifications
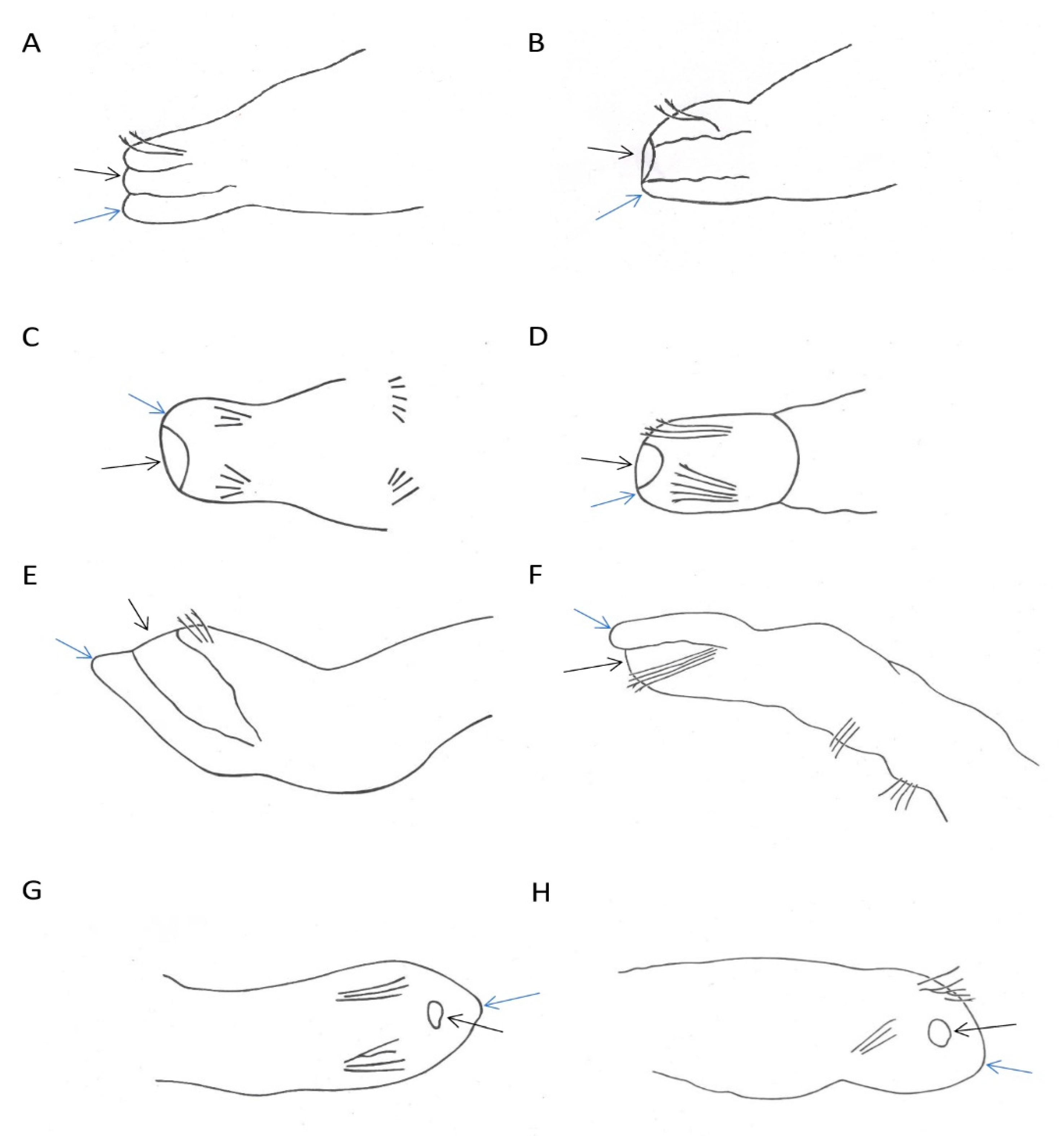
Out of the 135 Chaetogaster specimens collected at different periods (spring to autumn), none was sexually mature, a condition rarely reported in the literature. As we morphologically identified some specimens as C. langi while they belonged to a C. diastrophus lineage and inversely, we concluded that the morphological features attributed to C. langi were in practice not entirely reliable and that identification errors were possible. Indeed, fixation can cause a deformation or flattening of the prostomium and the position on slide of a C. langi specimen can falsely suggest that the specimen has a developed prostomium. We therefore suggest that doubtful specimens of C. langi/C. diastrophus should be identified as Chaetogaster sp. or genetic analysis carried out when possible. However, the morphological features specific to C. diastrophus and C. langi (Figure 3) could be verified on most specimens of each morphological group, so we consider that C. langi is significantly different from C. diastrophus and that a specific lineage can be attributed to this species.
Chaetogaster sp. MOTU1 and MOTU2 were characterized morphologically based on observations of only 3 specimens. They had a size close to the one of C. langi, so smaller than the one of C. diastrophus. However, we could not identify them as C. langi as they seemed to possess a developed prostomium. For Chaetogaster MOTU8, no voucher was available (entire specimen sequenced), so it could not be attributed to a morphospecies. We sequenced ulteriorly 11 additional specimens (No 220-230) from Site Dws4 in the Glatt River to obtain a voucher of this lineage but none of them belonged to it. In addition, we could not distinguish morphologically the lineages within the morphogroup C. diastrophus, which seemed therefore completely cryptic.
Figure 3.
Drawings of anterior parts of C. langi (A-D) and C. diastrophus (E-H) showing in C. langi an absence or a vestigial state of the prostomium and in C. diastrophus a well-developed prostomium (A=isolate No 150, B= isolate No 13, C= isolate No 87, D= isolate No 25, E= isolate No 85, F= isolate No 156, G= isolate No 164 and H= isolate No 23); lateral view in A, B, E, F, ventral view in C, D, G, H; the black arrows show the buccal cavity of the specimens; the blue arrows show the prostomium of the specimens or where it would be if it was present.
Figure 3.
Drawings of anterior parts of C. langi (A-D) and C. diastrophus (E-H) showing in C. langi an absence or a vestigial state of the prostomium and in C. diastrophus a well-developed prostomium (A=isolate No 150, B= isolate No 13, C= isolate No 87, D= isolate No 25, E= isolate No 85, F= isolate No 156, G= isolate No 164 and H= isolate No 23); lateral view in A, B, E, F, ventral view in C, D, G, H; the black arrows show the buccal cavity of the specimens; the blue arrows show the prostomium of the specimens or where it would be if it was present.
3.4. Taxonomy
Here, we describe the three C. diastrophus MOTUs 3, 9 and 10 and the Chaetogaster spp. MOTUs 1-2 as new species for science (cf. Discussion section) (Table 2). We also attribute a lineage to C. langi (MOTU 5) and C. setosus (MOTU 4) and describe material corresponding to C. diaphanus (MOTUs 6-7) and to the new lineage of Chaetogaster sp. MOTU8, for which no anterior part was preserved.
Morphological group Chaetogaster diastrophus (MOTUs 3, 9 and 10)
Length 0.6-1.2 mm, diameter 0.11-0.16 mm (head). Prostomium present. Fusion of the first five segments forming a head containing a musculous pharynx, buccal cavity not large to large. Ventral chaetae bifid, absent in segments III-V; dorsal chaetae completely lacking.
Chaetogaster communis sp. nov. Vivien, Lafont & Martin (MOTU10)
Holotype: Isolate No 1; Anterior part of the body mounted on a microscopic slide in a medium containing glycerol, lactic acid and Mowiol, specimen voucher: GBIFCH 1222926; DNA voucher stored in buffer at -20oC at the Ecotox Center in Lausanne
Type locality: Flawil (St. Gallen, Switzerland), 47.419157°N 9.195899°E
Paratypes: Isolates No 2, 10, 21, 23, 24, 28, 30, 32, 35, 47, 76, 85, 88, 97, 100, 110, 131, 139, 148, 151, 156, 162, 163, 164, 220-230 (voucher references in Table S1)
Etymology: Named “communis” (Latin for “common”), as seems frequent, according to the data of the present study
Distribution (previous studies, GenBank data): Reported from Switzerland (Geneva area), Sweden and Norway
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996388-PP996389, PP996393, PP996396-PP996398, PP996402, PP996404, PP996406, PP996408, PP996415, PP996421, PP996423, PP996426, PP996429, PP996432, PP996434, PP996442, PP996446, PP996451, PP996454, PP996456, PP996461, PP996511-PP996521; ITS2: PQ013378-PQ013379, PQ013381-PQ013383, PQ013385-PQ013388, PQ013393, PQ013396, PQ013400, PQ013402, PQ013404, PQ013407, PQ013410, PQ013412, PQ013420, PQ013423, PQ013427, PQ013430, PQ013432, PQ013436-PQ013438, PQ013481-PQ013490; 28S: PQ013492, PQ013497, PQ013499, PQ013506, PQ013510, PQ013515, PQ013518, PQ013521-PQ013523.
Chaetogaster fluvii sp. nov. Vivien, Lafont & Martin (MOTU3)
Holotype: Isolate CDS1; Anterior part of the body mounted on a microscopic slide in a medium containing glycerol, lactic acid and Mowiol, specimen voucher: GBIFCH 1223004; DNA voucher stored in buffer at -20oC at the Ecotox Center in Lausanne
Type locality: Muri (Aargau, Switzerland), 47.305217°N 8.327193°E
Paratypes: Isolates: CDS2, No 98, 195, 196, 197, 198, 199, 200, 212, 213, 214, 215, 216 (voucher references in Table S1)
Etymology: Named “fluvii” (Latin for “of river”), as this species was found in streams
Distribution (previous studies, GenBank data): Reported from Sweden, USA and Norway
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996430, PP996465-PP996466, PP996489-PP996494, PP996505-PP996509; ITS2: PQ013408, PQ013440-PQ013441, PQ013462-PQ013465, PQ013476-PQ013479; 28S: PQ013495, PQ013525-PQ013526, PQ013544-PQ013545, PQ013555-PQ013556.
Chaetogaster fluminis sp. nov. Vivien, Lafont & Martin (MOTU9)
Holotype: Isolate No 5; Anterior part of the body mounted on a microscopic slide in a medium containing glycerol, lactic acid and Mowiol, specimen voucher: GBIFCH 1222928; DNA voucher stored in buffer at -20oC at the Ecotox Center in Lausanne
Type locality: Flawil (St. Gallen, Switzerland), 47.419157°N 9.195899°E
Paratypes: Isolates No 34, 86, 101, 127, 128, 129, 130, 141, 155, 157, 159, 160 (voucher references in Table S1)
Etymology: Named “fluminis” (Latin for “of river”), as this species was found in streams
Distribution (previous studies, GenBank data): Reported from USA, Canada, Sweden, Norway, United Kingdom and Italy
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996390, PP996407, PP996424, PP996433, PP996438-PP996441, PP996448, PP996455, PP996457, PP996459; ITS2: PQ013411, PQ013416-PQ013419, PQ013424, PQ013431, PQ013433-PQ013434; 28S: PQ013498, PQ013503-PQ013505, PQ013512, PQ013519.
Morphological group Chaetogaster spp. (MOTUs 1-2)
Length 0.5-0.6 mm, diameter 0.07-0.1 mm (head). Prostomium present. Fusion of the first five segments forming a head containing a musculous pharynx, buccal cavity not large to large. Ventral chaetae bifid, absent in segments III-V; dorsal chaetae completely lacking.
Chaetogaster suzensis sp. nov. Vivien, Lafont & Martin (MOTU1)
Holotype: Isolate No 113; Anterior part of the body mounted on a microscopic slide in a medium containing glycerol, lactic acid and Mowiol, specimen voucher: GBIFCH 1222976; DNA voucher stored in buffer at -20oC at the Ecotox Center in Lausanne
Type locality: Villeret (Bern, Switzerland), 47,152502°N 7.014924°E
Paratypes: None
Etymology: Named after the stream (Suze River) in which the holotype was found
Distribution (previous studies): -
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession number for COI: PP996437; ITS2:PQ013415; 28S: PQ013502.
Chaetogaster sorgensis sp. nov. Vivien, Lafont & Martin (MOTU2)
Holotype: Isolate No 134; Anterior part of the body mounted on a microscopic slide in a medium containing glycerol, lactic acid and Mowiol, specimen voucher: GBIFCH 1222983; DNA voucher stored in buffer at -20oC at the Ecotox Center in Lausanne
Type locality: Ecublens (Vaud, Switzerland), geographical coordinates: 46.522661°N 6.573581°E
Paratypes: Isolate No 165 (voucher reference in Table S1)
Etymology: Named after the stream (Sorge River) in which the holotype was found
Distribution (previous studies, GenBank data): Reported from Sweden and Norway
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996444, PP996464; ITS2: PQ013421, PQ013439; 28S: PQ013508, PQ013524.
Chaetogaster langi (MOTU5)
Material examined: Isolate No 7, 9, 12, 13, 25, 26, 27, 29, 31, 36-40, 42, 57, 65, 69-71, 78, 87, 89, 90, 112, 132, 140, 142, 149, 150 (geographical coordinates in Table 2 and voucher references in Table S1)
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996391-PP996392, PP996394-PP996395, PP996399-PP996401, PP996403, PP996405, PP996409- PP996414, PP996416-PP996420, PP996422, PP996425, PP996427-PP996428, PP996436, PP996443, PP996447, PP996449, PP996452-PP996453; ITS2: PQ013380, PQ013384, PQ013389-PQ013392, PQ013394-PQ013395, PQ013397-PQ013399, PQ013401, PQ013403, PQ013405-PQ013406, PQ013414, PQ013425, PQ013428, PQ013429; 28S: PQ013491, PQ013493-PQ013494, PQ013501, PQ013507, PQ013511, PQ013513, PQ013516-PQ013517.
Chaetogaster setosus (MOTU4)
Material examined: Isolates No 99, 111, 138, 147, 161 (geographical coordinates in Table 2 and voucher references in Table S1)
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996431, PP996435, PP996445, PP996450, PP996460; ITS2: PQ013409, PQ013413, PQ013422, PQ013426, PQ013435; 28S: PQ013496, PQ013500, PQ013509, PQ013514, PQ013520.
Chaetogaster diaphanus (MOTUs 6-7)
Material examined : Isolates Sor6-9 ; G4; CDP1-5, No 183-187, 189-194, 201-207, 209-211, 219 (geographical coordinates in Table 2 and voucher references in Table S1)
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession numbers for COI: PP996467-PP996471, PP996473-PP996476, PP996478-PP996488, PP996495-PP996504, PP996510; ITS2: PQ013442-PQ013449, PQ013451-PQ013461, PQ013466-PQ013475; 28S: PQ013527-PQ013532, PQ013534-PQ013543, PQ013546- PQ013554, PQ013557.
Chaetogaster sp. with no morphological description (MOTU8)
Material examined: Isolate Glatt6 (geographical coordinates in Table 2)
Diagnostic molecular characters: The species can be distinguished from other Chaetogaster species on the basis of COI, ITS2 and 28S DNA sequences (Tab. S1). GenBank accession number for COI: PP996477; ITS2:PQ013450; 28S: PQ013533.

Table 2.
For each lineage, indication of the new species name (when applicable) and information on geographical coordinates, preservation of material (DNA and voucher); the habitat is the same for all specimens, surface coarse sediments in streams.
Table 2.
For each lineage, indication of the new species name (when applicable) and information on geographical coordinates, preservation of material (DNA and voucher); the habitat is the same for all specimens, surface coarse sediments in streams.
| Morphospecies/group | New species name | Geographical coordinates | Material preservation |
|---|---|---|---|
| Chaetogaster diaphanus MOTUs 6-7 | Isolates Sor6-9: 46.522661oN, 6.573581oE; Isolate G4: 47.419157oN, 9.195899oE; Isolates CDP1-5, No 201-207, 209-211, 219, No 183-187, 189-194: 47.305217oN, 8.327193oE |
DNA voucher of the 32 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of 31 specimens (all except No 204) preserved (mounted on slides) in the Muséum cantonal des sciences naturelles Lausanne | |
| Chaetogaster diastrophus MOTU10 |
Chaetogaster communis sp. nov. Vivien, Lafont & Martin |
Isolates No 1, 2, 10, 21: 47.419157oN, 9.195899oE; Isolates No 23, 24, 28, 30, 32, 35, 47, 76, 85: 47.414825oN, 9.198461oE; Isolates No 88, 97, 100, 110, 131, 139, 148, 151, 156, 162, 163, 164: 47,152502oN, 7.014924oE; Isolates No 220-230: 47.43167oN, 9.17485oE |
Holotype and paratypes: DNA voucher of the 36 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of 35 specimens (all except No 30) preserved (mounted on slides) in the Muséum cantonal des sciences naturelles of Lausanne |
| Chaetogaster diastrophus MOTU3 | Chaetogaster fluvii sp. nov. Vivien, Lafont & Martin | Isolates CDS1, CDS2, No 195, 196, 197, 198, 199, 200, 212, 213, 214, 215, 216: 47.305217oN, 8.327193oE; Isolate No 98: 47.152502oN, 7.014924oE |
Holotype and paratypes: DNA voucher of the 14 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of the 14 specimens preserved (mounted on slides) in the Muséum cantonal des sciences naturelles of Lausanne |
| Chaetogaster diastrophus MOTU9 |
Chaetogaster fluminis sp. nov. Vivien, Lafont & Martin |
Isolates No 5: 47.419157oN, 9.195899oE; Isolates No 34, 86: 47.414825oN, 9.198461oE; Isolate No 101: 47,152502oN, 7.014924oE Isolates No 127, 128, 129, 130 46.522661oN, 6.573581oE; Isolates No 141, 155, 157, 159, 160: 47,152502oN, 7.014924oE |
Holotype and paratypes: DNA voucher of the 13 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of 12 specimens (all except No 155) preserved (mounted on slides) in the Muséum cantonal des sciences naturelles of Lausanne |
| Chaetogaster sp. MOTU1 | Chaetogaster suzensis sp. nov. Vivien, Lafont & Martin | Isolate No 113: 47,152502oN, 7.014924oE | Holotype: DNA voucher of the specimen stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of the specimen (mounted on a slide) preserved in the Muséum cantonal des sciences naturelles of Lausanne |
| Chaetogaster sp. MOTU2 | Chaetogaster sorgensis sp. nov. Vivien, Lafont & Martin | Isolate No 134: 46.522661oN, 6.573581oE; Isolate No 165: 47,152502oN, 7.014924oE |
Holotype and paratype: DNA voucher of the 2 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of the 2 specimens preserved (mounted on slides) in the Muséum cantonal des sciences naturelles of Lausanne |
| Chaetogaster sp. MOTU8 | Isolate Glatt6: 47.43167oN, 9.17485oE | DNA voucher of the specimen (Glatt 6) stored in buffer at -20oC at the Ecotox Center in Lausanne | |
| Chaetogaster langi MOTU5 | Isolates No 7, 9, 12, 13: 47.419157oN, 9.195899 oE; Isolates No 25, 26, 27, 29, 31, 36-40, 42, 57, 65, 69-71, 78: 47.414825oN, 9.198461oE; Isolates No 87, 89, 90, 112: 47,152502oN, 7.014924oE; Isolate No 132: 46.522661oN, 6.573581oE; Isolates No 140, 142, 149, 150: 47,152502oN, 7.014924oE |
DNA voucher of the 30 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of 27 specimens (all except No 29, 37 and 38) preserved (mounted on slides) in the Muséum cantonal des sciences naturelles of Lausanne | |
| Chaetogaster setosus MOTU4 | Isolates No 99, 111, 138, 147, 161: 47,152502oN, 7.014924oE | DNA voucher of the 5 specimens stored in buffer at -20oC at the Ecotox Center in Lausanne; Anterior part of the 5 specimens preserved (mounted on slides) in the Muséum cantonal des sciences naturelles of Lausanne |
4. Discussion
4.1. Species Delimitations and Chaetogaster Diversity in Switzerland
It is generally accepted that, as a rule of thumb in clitellates, if two clusters delimited on the basis of the COI gene differ from each other by more than 10% uncorrected genetic distances, they are likely to belong to different species, and if they differ by less than 5%, they are likely to belong to one species [31] (but see Liu et al. [32] for exceptions within the Limnodrilus hoffmeisteri species complex). The different approaches and genes used yielded largely congruent results and confirmed the delineation obtained by ASAP with the COI gene, except for MOTUs 6 and 7, grouped together in all analyses based on nuclear genes, and the mPTP analysis of 28S sequences.
Incomplete lineage sorting (ILS) and/or introgression are the most common factors invoked to explain phylogenetic discordance between nuclear and mitochondrial markers [33,34]. Distinguishing between the two processes can be a challenging task [35], out of the scope of the present study. Although ILS can affect both nuclear and mitochondrial markers, the lack of recombination in mtDNA means that the genetic variation in mtDNA tends to coalesce more rapidly than in nuclear DNA, suggesting that MOTU6 and MOTU7 are possible recent separately evolving lineages, in other words, potential distinct species according to the de Queiroz's species concept [36].
By delimiting only 6 MOTUs, by merging MOTUs identified in the other analyses (MOTUs 6, 7; 5, 8, 9, 10) and despite the splitting of MOTU4 into two distinct lineages, the mPTP analysis of the 28S sequences clearly stands out from the other results. The splitting of MOTU4 into two lineages is clearly an artefact, given the genetic proximity between the 28S sequences that are split. Usually, mPTP is considered superior to its previous variants (PTP, bPTP) by producing more taxonomically congruent delineations [30]. However, it is possible that the lower variability of 28S compared to COI and ITS2 partially explains this result. Indeed, Dellicour and Flot [26] showed that PTP and bPTP tend to produce more overlumping when the mutation rate of the markers is low.
Chaetogaster sp. MOTUs 1-2, considered morphologically as possibly intermediate between C. diastrophus and C. langi, were, on the phylogenetic analysis of COI and 28S, early branching, which shows an important genetic divergence between these two lineages and the rest of the Chaetogaster lineages. Chaetogaster sp. MOTU2, corresponding to sp. 1 in Mack et al. [2], was also early branching on a concatenated tree based on COI, ITS2 and H3 markers built by these authors. The COI, ITS2 and 28S data suggest that Chaetogaster sp. MOTU8 is a cryptic species of C. diastrophus. But we cannot exclude that it belongs to the morphospecies C. limnaei or C. parvus (anterior part not preserved).
As ITS2 and 28S data suggested that all C. diaphanus MOTUs 6-7 sequences formed only one lineage, we decided to regroup all sequences of this morpho-group in a unique species, as a conservative measure. The genetic divergence in COI barely reached the threshold of 10% used to distinguish clitellate species, so the distinction based on COI data only was already doubtful. Mack et al. [2] considered these two MOTUs as significantly different based on the analysis of COI, the mitochondrial marker 16S rDNA and the nuclear region H3. The ITS2 sequences obtained by Mack et al. [2] also showed distinction between these MOTUs, which is in contradiction with our ITS2 data. This discordance is difficult to understand, and we cannot exclude that the ITS2 sequences provided by Mack et al. [2] for MOTU sp. 3 (= our MOTU7) correspond in fact to other specimens than those of this MOTU.
Chaetogaster langi was considered by some authors as synonym of C. diastrophus [2]. We could attribute a DNA barcode to the morphospecies C. langi and demonstrate that it was morphologically significantly different from C. diastrophus. Mack et al. [2] found this lineage but did not describe it as belonging to C. langi, probably because the small size of the Chaetogaster specimens required the extraction of DNA from whole specimens, making impossible to go back to the morphological study of the lineages.
In addition, Mack et al. [2] did not mention that one of the C. diastrophus lineages they found could belong to Chaetogaster parvus Poitner, 1914, probably because this species is considered by some authors as invalid or synonym of C. langi [10,37], despite the mention of its presence in France [38] and Switzerland and the fact that it can be easily differentiated from C. langi or C. diastrophus by the form of the crotchets [38,39]. Finally, Mack et al. [2] discussed the possibility that one of the two MOTUs of C. diaphanus could be confined to North America (sp. 3) and the other to Europe (sp. 4). But in the present study, the MOTU (MOTU7) supposed to be present only in North America was abundant in one stream, which suggests that it is also well represented in Europe.
In term of diversity, we observed a high number of Chaetogaster lineages in/at a low number of streams and sites. These results suggest that we only collected a part of the Chaetogaster lineages present in Switzerland and that a higher number of Chaetogaster species could be found in this small territory. The phylogenetic and morphological analyses suggest that the lineages Chaetogaster sp. MOTUs 1-2 could correspond to a new morphological group (morphospecies). However, this observation should be confirmed based on the morphological analysis of more specimens belonging to these lineages.
4.2. Description of New Chaetogaster Species for Science
It is widely acknowledged that cryptic species within morphospecies are frequent in metazoans and that they should be considered as distinct species [40]. It is now accepted that in many cases, speciation in metazoans can occur without any (visible) morphological modifications [40]. But so far, no consensus exists on the conditions required to attribute a status of nominal species to lineages indistinguishable morphologically. Many taxonomists agree that taxonomic descriptions should be integrative, i.e. consider all available data in literature (morphology, molecular sequences, behavior, ecology, reproductive isolation, etc.) for establishing if the different lineages within a same morphospecies are or not distinct species [41,42]. The taxonomists should however select some characteristics that could be pertinent to distinguish cryptic species in a particular group [43,44]. For example, experiments on behavioral characters and reproductive isolation could be tested on terrestrial oligochaetes such as lumbricids [45,46,47], but certainly not on rare aquatic oligochaete species that could not or hardly be isolated and cultivated. Such experiments could for example not be performed on cryptic species of Chaetogaster. Indeed, to attribute a lineage to a specimen of this genus, it is necessary to collect at least half of its body for genetic analyses and Chaetogaster specimens have not the property to regenerate their posterior segments. Given the current state of scientific knowledge and technological capabilities, it is probable that for the most cryptic metazoan species (e.g. worms, insects, etc.) on Earth, only DNA sequences could be used as a tool for distinguishing them. Molecular analyses that have proven to be effective for the identification of species in many invertebrate groups, including annelids e.g. [48,49,50], could be sufficient tools to delimitate and describe species.
The fact that cryptic species remain almost always considered as MOTUs, OTUs or putative species and named differently in the different studies lead to a denial of their existence as real species and so in their absence in the species inventories and conservation programs [43]. Once their validity established (based on DNA data and if possible other observations), these cryptic species should consequently be described as nominal species and considered by the scientific community as integrant part of biodiversity [31,50]. There are no official requirements of the ICZN that the description of species should be based only on morphological grounds [43].
We agree with Zamani et al. [51] and Martinsson & Erséus [50] that description of species using only DNA data should be avoided and that it should be accompanied with morphological observations. We preserved a voucher for most sequenced specimens and tried to find distinctive characters between the lineages of a same morphospecies and specific features to the lineages identified as Chaetogaster spp.
Currently, the newly described species of Chaetogaster could be recognized only using genetic analyses as we could not find any morphological characters such as the specimen size and the form of prostomium and crotchets specific to each of them. We cannot expect in the short and medium term characterizing each one using the morphology of their reproductive organs as sexually mature Chaetogaster specimens are very rarely found and in addition, it is necessary to cut specimens in two for sequencing them and so the segments bearing the genital features can be removed or damaged. Therefore, only an examination of live sexually mature specimens and photo acquisitions before sequencing could allow to differentiate and describe them based on these characters. This procedure would not allow to preserve a voucher containing the distinctive morphological features and of course it is not sure at all that the species of a same morphospecies would show observable differences in the genital apparatus.
The nominal species C. diastrophus [52] could potentially correspond to one of the three described lineages of this morphospecies. But its type has probably been lost or cannot be sequenced. In the 19th century, specimens were generally fixed using low-pH formalin and preserved in this medium or in 70% ethanol. In addition, the type could be preserved mounted on slide in a conservative medium, which could also hamper subsequent genetic analyses. When type material is lost or cannot be used for genetic analyses, assigning a genetic lineage to a nominal species remains still possible as long as new biological material is collected as close as possible to the original type locality [50]. Neotypes can thus be designated in compliance with the conditions required by the International Code of Zoological Nomenclature (Art. 75.3), in particular the provision of Article 75.3.6 relating to the original type locality, and genetically characterized. Sampling of C. diastrophus at the type locality would however not allow to know with certitude the lineage of the type as more than one lineage corresponding to this morphospecies can be present at a same site. For example, in the present study, the three lineages of C. diastrophus were present at site 9 in the Suze River.
As the C. diastrophus specimens of the present study were certainly not sampled at or close to the type locality (which is not mentioned in Gruithuisen [52]), we described each lineage corresponding to this morphospecies as a new species for science. If in the future it could be demonstrated that one of these new lineages corresponded to the nominal species, it would be possible to consider the new lineage as synonym to the nominal species. The nominal species C. diastrophus should currently be considered as species inquirendae. We recommend that any future studies based solely on a morphological approach should refer to it as species "sensu lato".
As it has not been demonstrated that the nominal species C. diaphanus, C. setosus and C. langi contained cryptic species, the lineages obtained in the present study for each of these species can be considered as belonging to the nominal species. If in the future cryptic species were detected within these species, we would recommend describing the cryptic lineages as new species for science and to consider the nominal species as species inquirendae. The new lineage MOTU8 could not be described as a new species for science as we could not preserve any voucher for this lineage. Even if the phylogenetic analyses suggest that it is an additional cryptic species of C. diastrophus, it is not impossible that it belongs to the species C. limnaei or C. parvus.
A high number of cryptic species of aquatic oligochaetes have been reported so far but are still not considered as integrant part of biodiversity, as no one was described as a new species for science. Several cryptic species (lineages) were reported for example within Tubifex tubifex [53], Limnodrilus hoffmeisteri [32,53], Globulidrilus riparius [54], Nais communis [55] and Haplotaxis gordioides [56]. We believe that these Chaetogaster cryptic species validated based on mitochondrial and nuclear data should be fully considered as individual species by the scientific community, especially because it cannot be expected to characterize each of them using morphological features. The genus Chaetogaster represents therefore an excellent candidate for initiating descriptions of cryptic species within aquatic oligochaetes.
4.3. Perspectives
The prospects of the present work are to complete the data of the molecular diversity of this genus in Switzerland and to describe on a molecular/morphological basis the future new Chaetogaster cryptic species that we will probably find. Special effort will be made to barcode C. parvus, well represented in Switzerland, to confirm its existence as a separate species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Details for each Chaetogaster specimen: sampling, morphological identification, Genbank accession number and voucher identifier (Place of deposit: Muséum cantonal des sciences naturelles of Lausanne); Table S2: Mean genetic variations (p-distances) between the MOTUs of the present study (M01-M10) and the lineages found by Mack et al. (2023) (Sp01-Sp24); Figure S1: Molecular phylogeny constructed using the maximum likelihood method and 28S gene fragment of Chaetogaster specimens. Partitions at the right side of the figure represent the results of the species delimitation analyses with single-locus methods (ASAP, mPTP). For comparison purposes, the MOTU numbers correspond to those used for the MOTUs identified from the COI. Numbers at nodes are ultrafast bootstrap values (BV). Nodes were considered as supported if BVs were higher or equal to 90 [20]. For the sake of clarity, BVs are not shown within delimited MOTUs. File S1: Aligned COI sequences of Chaetogaster obtained in the present study; File S2: Aligned ITS2 sequences of Chaetogaster obtained in the present study; File S3: Aligned 28S sequences of Chaetogaster obtained in the present study
Author Contributions
Conceptualization: R.V., M.L., C.I., B.J.D.F., P.M.; Methodology: R.V., P.M.; Choice of sites and sampling: R.V., B.J.D.F.; Investigation: R.V., M.L., C.I., P.M.; Formal Analysis: R.V., P.M.; Prepared figures and tables: R.V., P.M.; Writing – Original Draft Preparation: R.V., P.M..; Writing – Review & Editing: R.V., B.J.D.F., P.M. All authors have read and agree to the published version of the manuscript.
Funding
This research received no specific external funding.
Data Availability statement
The COI, ITS2 and 28S sequences are provided as Supplemental Files S1-S3 and are accessible in GenBank: COI: PP996388-PP996521; ITS2: PQ013378-PQ013490; 28S: PQ013491-PQ013557. The anterior parts of all specimens (vouchers), except isolates Glatt6, Nos 29, 30, 37, 38, 155, 204, 205, are deposited in the “Muséum cantonal des sciences naturelles” of Lausanne (Table S1).
Acknowledgments
We are grateful to Emmanuelle Rohrbach and Alessandro Scapuso for their help with PCR amplifications and sequencing.
Conflicts of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Timm T., Martin, P. J. Chapter 21 - Clitellata: Oligochaeta. In J. H. Thorp & D. C. Rogers (Eds) Thorp and Covich's Freshwater Invertebrates (Fourth Edition) pp. 529-549). Boston: Academic Press, 2015.
- Mack J.M., Klinth M., Martinsson S., Lu R., Stormer H., Hanington P, et al. Cryptic carnivores: Intercontinental sampling reveals extensive novel diversity in a genus of freshwater annelids. Mol. Phylogenet. Evol. 2023, 182, 107748. [CrossRef]
- Vivien, R., Lafont, M., Werner, I., Laluc, M., Ferrari, B.J.D. Assessment of the effects of wastewater treatment plant effluents on receiving streams using oligochaete communities of the porous matrix. Knowl. Manag. Aquat. Ec. 2019, 420, 18. [CrossRef]
- Lafont M., Jézéquel C., Vivier A., Breil P., Schmitt L., Bernoud S. Refinement of biomonitoring of urban watercourses by combining descriptive and ecohydrological approaches. Ecohydrol. Hydrobiol. 2010, 10, 3-11. [CrossRef]
- Martin P., Reynolds J., van Haaren, T. World List of Marine Oligochaeta. Chaetogaster von Baer, 1827. 2024. Accessed through: World Register of Marine Species at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=137356 on 2024-05-17.
- Vivien, R., Lafont, M. Note faunistique sur les oligochètes aquatiques de la région genevoise et de Suisse. Rev. Suisse Zool. 2015, 122, 207-212. [CrossRef]
- Vivien R., Ferrari, B.J.D. Assessment of the biological quality and functioning of the Suze River upstream and downstream of the wastewater treatment plant in Villeret (canton of Bern) using oligochaete communities in the porous matrix, Centre Ecotox, Eawag-EPFL, 2023.
- Vivien, R., Werner, I., Ferrari, B.J.D. Simultaneous preservation of the DNA quality, the community composition and the density of aquatic oligochaetes for the development of genetically based biological indices. PeerJ 2018, 6, e6050. [CrossRef]
- Reymond O. Préparations microscopiques permanentes d’oligochètes : une méthode simple. Bull. Soc. Vaud. Sci. 1994, 83, 1-3. [CrossRef]
- Timm T. A guide to the freshwater Oligochaeta and Polychaeta of Northern and Central Europe. Lauterbornia 2009, 66, 1–235.
- Tkach V., Pawlowski J. A new method of DNA extraction from the ethanol-fixed parasitic worms. Acta Parasitol. 1999, 44, 147-148.
- Folmer O., Black M., Hoeh W., Lutz R., Vrigenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294-299.
- Leray M., Yang J.Y., Meyer C.P., Mills S.C., Agudelo N., Ranwez V. et al. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity – application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [CrossRef]
- Navajas M., Lagnel J., Gutierrez J., Boursot P. Species wide homogeneity of nuclear ribosomal ITS2 sequences in the spider mite Tetranychus urticae contrasts with extensive mitochondrial COI polymorphism. Heredity 1998, 80, 742–752. pmid:9675873. [CrossRef]
- Jamieson B.G.M., Tillier S., Tillier A., Justine J.-L., Ling E., James S., et al. Phylogeny of the Megascolecidae and Crassiclitellata (Annelida, Oligochaeta): combined versus partitioned analysis using nuclear (28S) and mitochondrial (12S, 16S) rDNA. Zoosystema 2002, 24, 707-734.
- Edgar R.C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics 2004, 5, 113. [CrossRef]
- Gouy M., Guindon S., Gascuel O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221-224. [CrossRef]
- Kalyaanamoorthy S., Minh B.Q., Wong T. K. F., von Haeseler, A., Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587-589. [CrossRef]
- Nguyen L.T., Schmidt H.A., von Haeseler A., Minh B.Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268-274. [CrossRef]
- Hoang D.T., Chernomor O., von Haeseler A., Minh B.Q., Vinh L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518-522. [CrossRef]
- Tamura K., Peterson D., Peterson N., Stecher G., Nei M., Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731-2739. [CrossRef]
- Dellicour S., Flot J.-F. Delimiting species-poor data sets using single molecular markers: A study of barcode gaps, haplowebs and GMYC. Syst. Biol. 2015, 64, 900-908. [CrossRef]
- Puillandre N., Brouillet S., Achaz, G. ASAP: assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609-620. [CrossRef]
- Zhang, J., Kapli, P., Pavlidis, P. & Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869-2876. [CrossRef]
- Pons J., Barraclough T.G., Gomez-Zurita J., Cardoso A., Duran D.P., Hazell, S., et al. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 1-15. [CrossRef]
- Dellicour S., Flot J.-F. The hitchhiker's guide to single-locus species delimitation. Mol. Ecol. Resour. 2018, 18, 1234-1246. [CrossRef]
- Luo A., Ling C., Ho S.Y.W., Zhu C.D. Comparison of methods for molecular species delimitation across a range of speciation scenarios. Syst. Biol. 2018, 67, 830-846. [CrossRef]
- Goulpeau, A., Penel, B., Maggia, M.-E., Marchán, D.F., Steinke, D., Hedde, M. & Decaëns, T. OTU delimitation with earthworm DNA barcodes: A comparison of methods. Diversity 2022, 14, 10. [CrossRef]
- Phillips J.D., Gillis D.J., Hanner R.H. Lack of statistical rigor in DNA barcoding likely invalidates the presence of a true species' barcode gap. Front. Ecol. Evol. 2022, 10. [CrossRef]
- Kapli P., Lutteropp S., Zhang J., Kobert K., Pavlidis P., Stamatakis A., et al. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630-1638. [CrossRef]
- Schmelz R.M., Beylich A., Boros G., Dózsa-Farkas K., Graefe U., Hong Y., et al. How to deal with cryptic species in Enchytraeidae, with recommendations on taxonomical descriptions. Opusc. Zool. 2017, 48, 45–51. [CrossRef]
- Liu Y., Fend S.V., Martinsson S., Erséus C. Extensive cryptic diversity in the cosmopolitan sludge worm Limnodrilus hoffmeisteri (Clitellata, Naididae). Org. Divers. Evol. 2017, 17, 477–495.
- Dong X., Zhang H., Zhu X., Wang K., Xue H., Ye Z., et al. Mitochondrial introgression and mito-nuclear discordance obscured the closely related species boundaries in Cletus Stål from China (Heteroptera: Coreidae). Mol. Phylogenet. Evol. 2023, 184, 107802. [CrossRef]
- Toews, D. P. L. & Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907-3930. [CrossRef]
- Galtier N. An approximate likelihood method reveals ancient gene flow between human, chimpanzee and gorilla. Peer Community J. 2024, 4, e3. [CrossRef]
- de Queiroz K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879-886. [CrossRef]
- Sperber C. A taxonomical study of the Naididae. Zool. Bidr. Upps. 1950, 29, 45-81.
- Lafont M. Redescription de Chaetogaster parvus Poitner, 1914 (Oligochaeta, Naididae) avec quelques remarques sur la validité de cette espèce et sa répartition dans les eaux douces françaises. Ann. Limnol. 1981, 17, 211-217.
- Juget J. Quelques données Nouvelles sur les oligochètes du Léman: composition et origine du peuplement. Ann. Limnol. 1967, 3, 217-229.
- Bickford D, Lohman D.J., Sodhi N.S., Ng P.K.L., Meier R., Winker K., et al. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148-155. PMID: 17129636. [CrossRef]
- Dayrat B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [CrossRef]
- Padial J.M., Miralles A., De la Riva I., Vences M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [CrossRef]
- Jörger K.M., Schrödl M. How to describe a cryptic species? Practical challenges of molecular taxonomy. Front. Zool. 2013, 10, 59. [CrossRef]
- Marchán D.F., Díaz Cosín D.J., Novo M. Why are we blind to cryptic species? Lessons from the eyeless, Eur. J. Soil Biol. 2018, 86, 49-51. [CrossRef]
- Jones G.L., Wills A., Morgan A.J., Thomas R.J., Kille P., Novo M. The worm has turned: Behavioural drivers of reproductive isolation between cryptic lineages, Soil Biol. Biochem. 2016, 98, 11-17. [CrossRef]
- Dupont L, Audusseau H, Porco D, Butt KR. Mitonuclear discordance and patterns of reproductive isolation in a complex of simultaneously hermaphroditic species, the Allolobophora chlorotica case study. J. Evol. Biol. 2022, 35, 831-843. [CrossRef]
- Lowe C.N., Butt K.R. Culture techniques for soil dwelling earthworms: A review. Pedobiologia 2005, 49, 401-413.
- Knutson V.L., Gosliner T.M. The first phylogenetic and species delimitation study of the nudibranch genus Gymnodoris reveals high species diversity (Gastropoda: Nudibranchia). Mol. Phylogenet. Evol. 2022, 171, 107470. [CrossRef]
- Lawley J.W., Gamero-Mora E., Maronna M.M., Chiaverano L.M., Stampar S.N., Hopcroft R.R., et al. The importance of molecular characters when morphological variability hinders diagnosability: systematics of the moon jellyfish genus Aurelia (Cnidaria: Scyphozoa). PeerJ 2021, 9, e11954. [CrossRef]
- Martinsson S., Erséus C. Cryptic Clitellata: Molecular species delimitation of clitellate worms (Annelida): An overview. Diversity 2021, 13, 36. [CrossRef]
- Zamani, A; Faltynek Fric, Z.; Gante, H.F; Hopkins, T.; Orfinger, A.B; Scherz, M. D; Suchacko Bartonova, A.; Dal Pos, D. DNA barcodes on their own are not enough to describe a species. Systematic Entomology 2022, 47, 385-389. [CrossRef]
- Gruithuisen F.V.P. Über die Nais diaphana und Nais diastropha mit dem Nerven-und Blutsystem derselben. Nova Acta Physico-Medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum. 1828, 14, 407-420 + Plate I.
- Vivien, R., Holzmann, M., Werner, I., Pawlowski, J., Lafont, M., Ferrari, B.J.D. Cytochrome c oxidase barcodes for aquatic oligochaete identification: development of a Swiss reference database. PeerJ 2017, 5, e4122. [CrossRef]
- Martinsson S., Erséus C. Cryptic diversity in supposedly species-poor genera of Enchytraeidae (Annelida: Clitellata). Zool. J. Linn. Soc. 2018, 183, 749–762. [CrossRef]
- Envall I, Gustavsson L.M., Erséus C. Genetic and chaetal variation in Nais worms (Annelida, Clitellata, Naididae). Zool. J. Linn. Soc. 2012, 165, 495–520. [CrossRef]
- Martin P., Knüsel M., Alther R., Altermatt F., Ferrari B., Vivien, R. Haplotaxis gordioides (Hartmann in Oken, 1819) (Annelida, Clitellata) as a sub-cosmopolitan species: a commonly held view challenged by DNA barcoding. Zoosymposia 2023, 23, 78-93. [CrossRef]
Figure 1.
Molecular phylogeny constructed using the maximum likelihood method and COI gene fragment of Chaetogaster specimens. Partitions at the right side of the figure represent the results of the species delimitation analyses with single-locus methods (ASAP, mPTP). Numbers at nodes are ultrafast bootstrap values (BV). Nodes were considered as supported if BVs were higher or equal to 90 [20]. For the sake of clarity, BVs are not shown within delimited MOTUs.
Figure 1.
Molecular phylogeny constructed using the maximum likelihood method and COI gene fragment of Chaetogaster specimens. Partitions at the right side of the figure represent the results of the species delimitation analyses with single-locus methods (ASAP, mPTP). Numbers at nodes are ultrafast bootstrap values (BV). Nodes were considered as supported if BVs were higher or equal to 90 [20]. For the sake of clarity, BVs are not shown within delimited MOTUs.
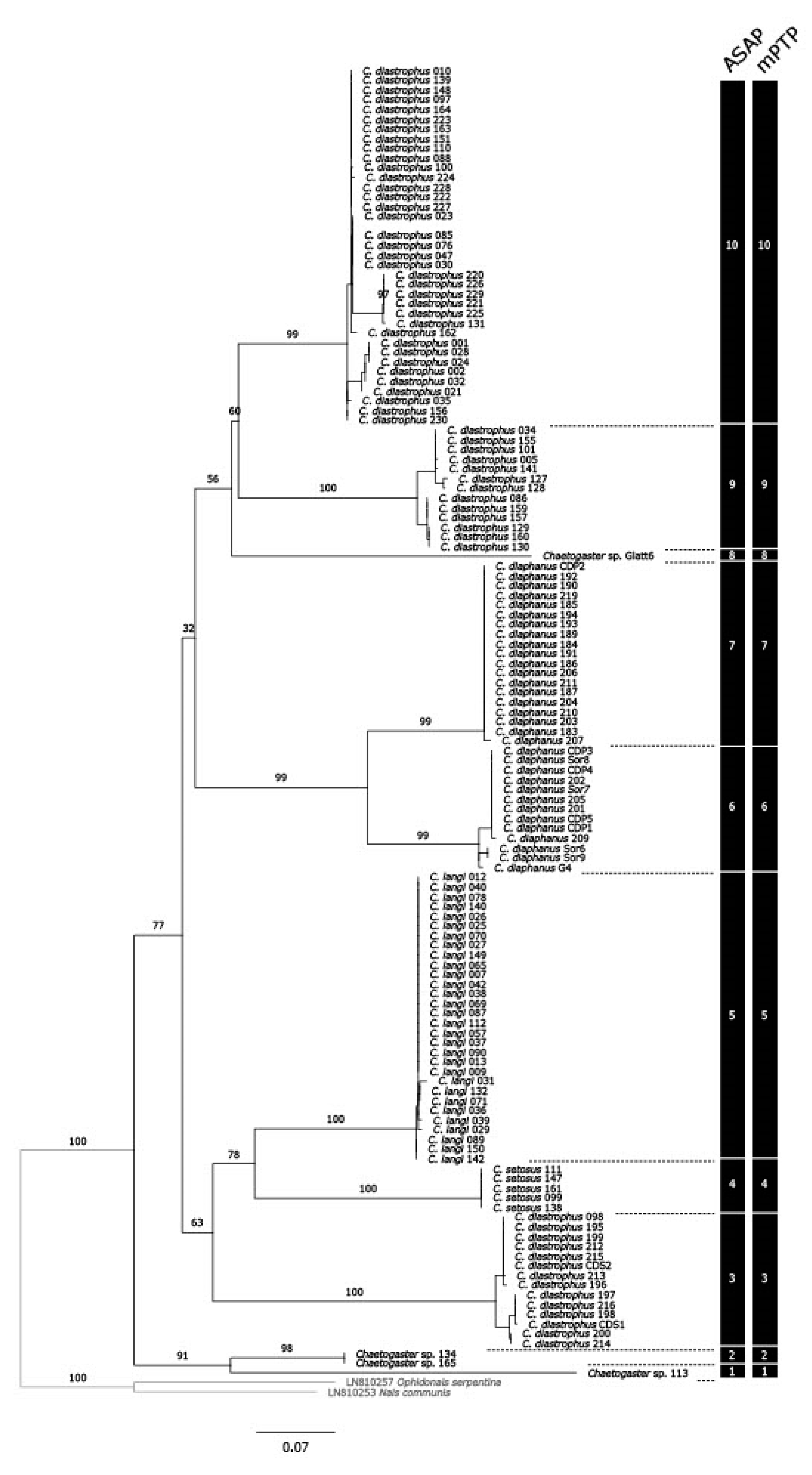
Table 1.
Percentages of maximal intra-MOTU and minimal inter-MOTU variations (p-distances) in COI and ITS2 of each MOTU of Chaetogaster found in the present study, with indication of the correspondence of each MOTU to the species (=lineages) described by Mack et al. [2].
Table 1.
Percentages of maximal intra-MOTU and minimal inter-MOTU variations (p-distances) in COI and ITS2 of each MOTU of Chaetogaster found in the present study, with indication of the correspondence of each MOTU to the species (=lineages) described by Mack et al. [2].
| No of species found by Mack et al. (2023) | Maximal intra- MOTU variation (%) in COI | Minimal inter- MOTU variation (%) in COI | Maximal intra- MOTU variation (%) in ITS2 | Minimal inter- MOTU variation (%) in ITS2 | |
| C. diaphanus MOTU6 | “C. diaphanus” sp.4 | 2.12 | 9.57 | 4.97 | 7.46 |
| C. diaphanus MOTU7 | “C. diaphanus” sp. 3 | 0.05 | 9.57 | ||
| C. diastrophus MOTU10 | “C. diastrophus” sp. 11 | 3.87 | 11.45 | 4.48 | 9.60 |
| C. diastrophus MOTU3 | “C. diastrophus” sp. 8 | 2.74 | 14.35 | 6.21 | 7.46 |
| C. diastrophus MOTU9 | “C. diastrophus” sp. 12 | 2.89 | 11.45 | 5.58 | 11.59 |
| C. langi MOTU5 | “C. diastrophus” sp. 19 | 0.91 | 13.67 | 5.68 | 10.34 |
| C. setosus MOTU4 | 0.00 | 12.61 | 2.73 | 12.50 | |
| Chaetogaster sp. MOTU1 | NC | 14.89 | NC | 13.86 | |
| Chaetogaster sp. MOTU2 | “C. diastrophus” sp. 1 | 0 | 12.00 | 2.78 | 12.29 |
| Chaetogaster sp. MOTU8 | “C. diastrophus” sp. 18 | NC | 11.89 | NC | 9.84 |
NC = not calculable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



