
Submitted:
30 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
Hematological malignancies refer to a heterogeneous group of neoplastic conditions of lymphoid and hematopoietic tissues classified in leukemias, Hodgkin and non-Hodgkin lymphomas and multiple myeloma, according to their presumed cell of origin, genetic abnormalities, and clinical features.
Metabolic adaptation and immune escape, which influence various cellular functions including proliferation and survival of hematological malignant tumor cells, are major aspects of these malignancies that lead to therapeutic drug resistance. Targeting specific metabolic pathways is emerging as novel therapeutic strategy in hematopoietic neoplasms, particularly in acute myeloid leukemia and multiple myeloma. In this context, CD147, also known as Extracellular Matrix Metalloproteinase Inducer (EMMPRIN) or Basigin, is one target-candidate involved in reprogramming metabolism in different cancer cells, including hematological malignant tumor cells. CD147 overexpression significantly contributes to the metabolic transformation of these cancer cells, by mediating signaling pathway, growth, metastasis and metabolic reprogramming, through its interaction, direct or not, with various membrane proteins related to metabolic, regulation including monocarboxylated transporters, integrins, P-glycoprotein and Glucose Transporter 1.
This review explores the metabolic functions of CD147 and its impact on the tumor microenvironment, influencing the progression and neoplastic transformation in leukemias, myeloma and lymphomas. Furthermore, we highlight new opportunities for the development of targeted therapies against CD147, potentially improving the treatment of hematologic malignancies.
Keywords:
Hematological Malignancies
; Metabolism
; CD147
1. Introduction
Hematologic malignancies (HM) represent a wide range of malignant tumours of the blood and lymphatic system which share pathological alterations capable of inducing radical transformations in cells, characterized by genetic, histological and metabolic alterations associated with uncontrolled cell proliferation and high resistance to cell death [1,2]. Aberrant metabolism can significantly affect the immune microenvironment and thus modulate the immune cellular response of the hematological malignant tumor cells, contributing to disease progression [3].
Increasing progress has been made for the treatment of HM, involving drugs classified as chemotherapy agents, whose activity is due to disruption of the mitotic and/or DNA replication pathways and targeted agents, which inhibit the molecular targets involved in cancer cell growth and spread. New targeted therapies with small molecule inhibitors, but also monoclonal antibodies, bispecific T cell engagers, antibody-drug conjugates, recombinant immunotoxins, and, more recently, Chimeric Antigen Receptor T (CAR-T) cells have improved the clinical outcomes for blood cancers, especially for older patients and/or patients with relapsed or refractory hematological malignancies who do not respond to standard treatments [4,5,6].
Clinical and preclinical studies have shown that targeting metabolic features of hematologic malignancies is an appealing therapeutic approach [6,7,8].
As tumor cells utilize glucose aerobically as an energy source and an intermediate for other metabolic pathways, the most common metabolic change found in HM is an increased glucose consumption of the tumor cells through aerobic glycolysis ("Warburg effect"), which leads to increased production and accumulation of lactate [9]. Lactate, derived from glucose fermentation, is excreted from the cell. Inhibition of lactate production or transport of lactate out of the cell are considered two potential strategies to directly target the Warburg effect in hematological malignancies [7,9].
CD147, an Extracellular Matrix Metalloproteinase Inducer (EMMPRIN) also called Basigin [10,11], is a multifunctional glycoprotein which overexpression in various cancers is associated with poor prognosis, including in HM [10,11,12,13,14]. CD147 that interacts with membrane proteins related to metabolic regulation including the monocarboxylated transporters (MCT1 and MCT4), integrins such as CD98, P-glycoprotein (P-gp) and Glucose Transporter 1 (GLUT1), significantly contributes to the metabolic transformation of neoplastic cells [11,12,13,14]. CD147 is involved in the regulation of the main metabolic pathways such as glycolysis, oxidative phosphorylation and lipolysis [10,11,15]. Furthermore, immunomodulatory function of CD147 has emerged, particularly relevant in cancer [16]. Then, CD147 which overexpression can be used as a biomarker in HM [11,12,13,14,15,17], and its protein partners represent a potential therapeutic target in the treatment of hematologic malignancies for the development of new therapies aimed at identifying and manipulating specific metabolic vulnerabilities of the different main groups, leukemia, lymphoma and multiple myeloma, of HM that support neoplastic cells proliferation.
In this review we will examine recent advances in the characterization of metabolic pathways influenced by CD147 and its partners, and in the development of new therapies in hematological malignancies. The review will also highlight the role of CD147 in tumor glucose metabolism reprogramming and its interplays with the tumor microenvironment (TME).
2. Hematologic Malignancies
In the 5th edition of the WHO classification of hematolymphoid tumors (August 2022), the stage of maturation, phenotypic character, histomorphological characteristics, clinical information and cytogenetic/molecular genetic findings are taken into account for the classification criteria with the aim of increasing diagnostic accuracy [18,19,20]. From 3 main groups including leukemias, lymphomas and multiple myeloma, HM can be categorized into different subgroups, such as acute leukemias, chronic leukemias, myelodysplastic syndromes (MDS), myeloproliferative neoplasms (MPNs), non-Hodgkin lymphomas, and classic Hodgkin lymphomas [2,18,19,20].
2.1. Acute Myeloid Leukemia
Acute Myeloid Leukemia (AML) is a clonal hematopoietic disorder affecting hematopoietic stem and progenitor cells, which results in the blockage of myeloid differentiation and the suppression of hematopoietic functions. AML is the most commonly occurring acute leukemia in adults and its incidence increases with age. As a result of genetical mutations in hematopoietic stem/progenitor cells, AML is a highly heterogeneous disease [21]. An increase in glycolysis has been observed in AML cell lines and in primary human AML blasts. Leukemia cells to ensure high glycolytic levels use amino acids as fuel, in particular glutamine, glutamate and proline. Amino acids are converted into metabolic intermediates to support the anaplerosis of the TCA cycle. In addition to amino acid metabolism, AML cells can compensate metabolically by increasing fatty acid metabolism to promote their survival and quiescence [22].
2.2. Acute Lymphoblastic Leukemia
Acute lymphoblastic leukemia (ALL) originates from the transformation and uncontrolled proliferation of B cell precursor (B-ALL) or T lymphoid progenitors (T-ALL) [23]. ALL is the most prevalent acute common childhood malignancy, manifested by an expansion of immature B or T cells. Glycolytic pathway is a critical step in the malignant transformation of B-cells and B-lymphoid repression of glucose uptake and energy supply represents a previously unrecognized metabolic barrier against malignant pre-B-cell transformation, while B-lymphoid transcription factors function as metabolic gatekeepers by limiting the amount of cellular ATP to levels that are insufficient for malignant transformation [24].
2.3. Lymphomas
Lymphomas are part of a complex and highly heterogeneous group of lymphoid tissue neoplasms, which are divided into two main categories: Hodgkin lymphoma (HL) [25], and non-Hodgkin lymphoma (NHL) based on the presence of Reed-Sternberg cells, which is the hallmark of NHL [26]. Non-Hodgkin lymphoma is more common than Hodgkin lymphoma and include diffuse large B-cell lymphoma (DLBCL), follicular lymphoma (FL), mantle cell lymphoma, and Burkitt's lymphoma [26,27].
2.4. Myelodysplastic Syndrome/ Myeloproliferative Neoplasms
Myelodysplastic syndrome/ myeloproliferative neoplasms (MDS/MPNs) are a group of chronic onco-hematological diseases characterized by the coexistence of alterations in cell maturation, typically seen in MDS, with an increase in cell proliferation in bone marrow and peripheral blood, also seen in chronic MPNs. MDS and MPNs are the most common in elderly patients [28,29].
2.5. Multiple Myeloma
1Multiple myeloma (MM) is a cancer that affects a particular type of cell, plasma cells in the bone marrow. The abnormal and uncontrolled multiplication of a plasma cell gives rise to a population (a clone) of cancer cells [30,31].
Although increasing progress has been made with development of new chemotherapeutics, targeted therapies, and radiotherapy, significantly improving survival rates for cancer patients, many of these patients unfortunately experience treatment failure due to relapse and resistance. This clearly entails the need to continue research on new anticancer agents and new approaches to block neoplastic growth.
3. CD147
3.1. CD147 Structure and Expression
CD147 is a transmembrane glycoprotein which belongs to the superfamily of immunoglobulins and is present in four 4 isoforms (CD147/Basigin-1, -2, -3, -4) among these, isoform BSG-2 with two immunoglobulin domains, is the most abundant and most studied form, ubiquitously expressed in human by many types of cells, including hematopoietic, epithelial and endothelial cells [10,11].
CD147 is a potent inducer of extracellular matrix metalloproteinase (EMMPRIN) that plays a key role in the regulation of the extracellular matrix (ECM) remodeling during physiological and pathological processes such as wound healing, fibrotic and inflammatory-related diseases [10,32,33] and cancer, including haematological neoplasia such as AML, multiple myeloma, in myelodysplastic syndrome with 5q deletion, where CD147 expression levels have a prognostic value [10,11,13,14,15,16,17]. But CD147 is also a multifunctional protein that exerts pleiotropic functions by interacting with various binding partners, such as monocarboxylate transporters MCT1-4 involved in metabolic pathways and T-lymphocyte activation, cyclophilin A involved in adhesion, viral invasion and inflammation, E-selectin, integrins and caveolin-1 involved in adhesion [10,11]. CD147, expressed on the cell surface of leucocytes, plays a crucial role for the recruitment of innate or adaptative immune cells required to fight the virus infection and consequent inflammation [11,34,35].
The glycoprotein CD147 has a molecular weight between 28 and 60 KDa, depending on the level of glycosylation at the N-terminal end which consists of three N-bound glycosylation sites at Asn (Asn44, Asn 152 and Asn186). CD147 N-glycans determine the different molecular weights, respectively low LG-CD147 and high level of N-glycans in HG-CD147 form. The level of glycosylation reached by CD147 and the ratio between the two glycosylated forms LG-CD147 and HG-CD147, are two characteristics that play a fundamental role in affecting its biological functions; the N-glycosylation of CD147 is necessary for stabilizing CD147 glycoprotein, transporting CD147 to the cell membrane, modulating the secretion of MMPs, and finally initiating extracellular matrix remodeling. Furthermore, the different N-glycosylation on CD147 influences its association with protein partner [36,37,38]. N-glycosylation on CD147 is essential for its association with E-selectin during leukocyte infiltration in renal inflammation [39] and during the inflammatory response observed in CD147 overexpression-associated diseases [39,40] ; conversely, glycosylation can also inhibit the interaction between proteins as Tang et al. have shown by the increased interaction of the deglycosylated form of CD147 with caveolin-1, thus indicating that glycolysation on CD147 interferes with its interaction with caveolin-1 [40,41]. The possible role of N-glycosylation of CD147 in its interaction with other proteins, such as MCTs, integrins, and cyclophilins, has yet to be investigated with further studies. Importantly, the association of altered glycosylation of CD147 and multidrug resistance in human leukemia [42], the profound influence of the ratio of the two glycosylated forms HG-CD147 and LG-CD147 of CD147 on neoplastic diseases, and the correlation of the HG-CD147 form found in acute lymphoblastic leukemia (ALL) and its recurrence [43], have been demonstrated. Then, CD147 glycosylation that may be used as a marker to predict clinical prognosis of cancer, represents a potential therapeutic target in those cancers overexpressing the HG-CD147 form and the development of new target-specific inhibitors for CD147 glycosylation could significantly improve the prognosis of cancer patients [38].
CD147 is expressed in both membrane-bound and soluble forms [10,11,44]. The soluble form is considered to act in a paracrine fashion by binding to the membrane-bound form (i.e., dimerization), while CD147 dimerization is essential for promoting tumor invasion through its involvement in the production/activation of matrix metalloproteinases (MMPs) [45]. The soluble form of CD147 (sCD147) can be secreted from cells and released into the microenvironment, also as a full-length protein released via microvesicle [46]. sCD147 detection in the serum or plasma obtained from several types of tumors, associated with tumor growth, metastasis formation and chemoresistance, confers to CD147 its role of clinical biomarker in several cancers, including hematological malignancies [47,48,49,50,51]. Furthermore, sCD147 influences tumor cell proliferation through its interaction with the membrane-bound form of CD147. Internalization of sCD147 has also been shown to stimulate sCD147 production via positive feedback [47,48,49]. The peculiar structural characteristics of CD147 influence its ability to interact with different extracellular, intracellular and membrane proteins, ensuring its involvement in multiple biological process including cellular transport, adhesion, cell proliferation, angiogenesis, metastasis and apoptosis [10,11,12,13]. Although there are several/numerous studies on the role played by CD147 in various diseases and solid tumors [13,15], only recently in-depth research has clarified the biological role of this glycoprotein in normal hematopoiesis and in the development of hematological malignancies [10,11,12,35].
In a first study, expression of CD147 has been shown to be induced in activated leukocytes such as granulocytes, lymphocytes, and macrophages [52]. In our recent work we find that CD147 is expressed in normal CD34+ hematopoietic progenitor cells (HPCs) and down-regulated during monocytic and granulocyte differentiation of HPCs in "in vitro" cultures [12]. In blasts related to different subtypes of AML, CD147 is overexpressed compared to its normal counterpart. Specific inhibition of its activity with AC-73, inhibits proliferation and induces autophagy, demonstrating a critical role in the proliferation of leukemia cells [12]. In a subsequent study, to further characterize the downstream CD147 signaling pathway/network, we also extended our analysis on the expression and function of two partner molecules MCT1 and MCT4 in hematopoietic progenitor cells, in different leukemia cell lines and in primary leukemic blasts from AML patients [12,53]. We found an inverse correlation between MCT1 and MCT4 expression levels in leukemia cells and showed that MCT4 overexpression is associated with a poor prognosis in AML patients [53]. Interestingly, CD147 is expressed at the level of CD34/CD371 AML cells, previously described for their leukemia-initiating (LIC) properties [12]. Another study demonstrates that CD147 knockdown in AML cells suppressed growth and proliferation, reduced protein levels of NF-κB (p65), Bcl-2, Bcl-xL, and lowered anti- and pro-apoptotic protein ratios of Bcl-2/Bax and Bcl-xL/Bax, clarifying the role of CD147 in the signaling pathways involved in the development of leukemia [54]. Schmidt's study reports an overexpression of CD147 also in several lymphoma and leukemia cell lines, as well as in normal CD3+ T cells [55]. Protein analysis showed an increase in the high glycosylation form (HG-CD147) in ALK+ anaplastic large cell lymphoma (ALCL) cell lines, ALK− cell line and several B-cell lymphoma cell lines. The weakest expression of the HG-CD147 form has been detected in mantle cell lymphoma and T-cell acute lymphoblastic lymphoma cell lines. The low glycosylated form (LG-CD147) was strongly expressed in Mac-1 and HeLa, is weakly expressed in several cell lines of human diffuse large B-cell lymphoma, Raji (Burkitt lymphoma) e Granta B cell leukemia. In normal CD3+ T cell samples, however, no CD147 expression was detected [55]. Furthermore, CD147 is overexpressed in myelodysplastic syndrome (MDS) erythroid cells with 5q deletion and in multiple myeloma (MM), where CD147 expression levels have prognostic value and are necessary for the proliferation of MM cells [14,16,56].
3.2. CD147 Functions: Its Interaction with MCTs and Other Partners
CD147 glycoprotein was initially known as an matrix metalloproteinases (MMPs) inducer, capable of stimulating fibroblasts and endothelial cells to facilitate tumor invasion, metastasis, and angiogenesis [57,58]. Several studies have highlighted that CD147 also plays a role in several other functions and can interact with different partner proteins to regulate multiple signaling pathways [10,11] (Figure 1). Furthermore, CD147 is involved in angiogenesis through stimulation of vascular endothelial growth factor (VEGF) production [17,58]. Connected to cellular metabolic regulation, one of the prevalent biological functions carried out by CD147 concerns the regulation of the expression and the activity of monocarboxylate transporters-1 (MCT-1) and -4 (MCT-4) to form complexes on the membrane to transport the lactic acid produced from glycolysis [59]. This function is particularly critical in the context of neoplastic development, as CD147 supports glycolysis in the presence of oxygen in tumor cells [60]. MCTs catalyze the transport of lactate, pyruvate, and ketone bodies across the plasma membrane. There are four isoforms (MCT1–MCT4) with different modes of expression and distinct substrate affinities [61]. Mechanistically, CD147 serves as a chaperone required for plasma membrane translocation of MCTs [62]. The close association between CD147 and MCTs is consistent with the high level of CD147 expression in metabolically active cells, such as tumor cells.
Another partner of CD147 involved in the regulation of metabolism is the transmembrane protein GLUT1 that participates in the internalization of glucose. Several reports have demonstrated the interaction of CD147 with GLUT1, and in addition, it has also been reported that increased expression of GLUT1 and CD147 facilitates the entry of glucose into cancer cells to promote tumor glycolysis. GLUT1 overexpression has been correlated with an adverse prognosis in various cancer types as it leads to progression, invasion, and metastasis [63,64]. Overexpression of GLUT1 has been frequently associated with upregulation of CD147 in several types of tumors such as papillary renal cell carcinoma (pRCC) [63] or cervical squamous cell carcinoma [65], or melanoma, where it affects aerobic glycolysis favoring metabolic reprogramming and tumor growth [66]. In hematologic malignancies, GLUT1 disruption through genetic and pharmacological treatments, affects leukemic stem cells (LSCs) activity by disrupting energy metabolism, accompanied by increased apoptosis, differentiation, and autophagic activity. In human AML cell lines and in most samples tested from patients with AML, inhibition of GLUT1 and oxidative phosphorylation (OXPHOS), act synergistically impaired leukemia cell survival [67]. Although GLUT1 expression has been frequently found in a wide range of different expressions among the various Hodgkin lymphoma subtypes [68,69], it was not associated with clinical outcomes [68]. In multiple myeloma, an interesting study observed that some myeloma cell lines strongly express GLUT1 but found that the glucose uptake of myeloma cells is largely dependent on GLUT4 [70]. Since the suppression of GLUT1, through RNA interference, induces inhibition of cell growth and/or death, the possibility arises that different subtypes of GLUT are responsible for glucose uptake in different individual myeloma cells [71].
Interactions with integrins are particularly important when considering CD147 signaling. α 3 β 1 and α 6 β 1 integrins were colocalized with CD147 on the cell surface [72]. At least four molecules can mediate interactions between CD147 and integrins: CD98, CD43, MCT4 and galectin-3. CD147 binds directly to CD98, which in turn binds covalently to amino acid transporters and integrins [73]. CD98 is overexpressed in several hematological malignancies; in AML, CD98 promotes the propagation and lethality of AML by promoting the interaction of leukemic cells with their microenvironment and maintaining LSCs [74] and contributing to the growth of AML and the survival of LSCs [74]. In AML CD98 is highly expressed on proliferating cells and functions as a chaperone for I-type amino acid transporters such as LAT1 and LAT2. Furthermore, RNAi depletion of CD147 or CD98hc reduced cell surface expression of both molecules and reduced cell proliferation [75]. Aberrant regulation of CD98 expression has also been found in myeloma [76] and in lymphoma [77]. CD98 could also have a role in MDS, since it is known that the bone marrow microenvironment is altered in these pathologies and microenvironmental support is particularly important for the survival of MDS cells, so it is likely that leukemia-niche interactions that are important for the growth of AML play a role in the progression of MDS.
Among the molecules that bind to CD147 extracellularly and transmit their signals, cyclophilin A has been examined in detail [78]. Cyclophilin A is present intracellularly and is also secreted in response to immunological stimuli. Secreted cyclophilin A exhibits chemotactic activity for neutrophils, eosinophils, and T cells. CD147 has been found to be the major signaling receptor for cyclophilin A and its related molecule, cyclophilin B. Cyclophilin A typically binds to heparan sulfate, a heparin-like polysaccharide, and therefore to CD147 [78,79]. B-cell malignancies such as multiple myeloma frequently colonize the bone marrow. This colonization is mediated by cyclophilin A and CD147. Thus, cyclophilin A secreted by endothelial cells in bone marrow blood vessels attracts myeloma cells, which strongly express BSG, the major receptor for cyclophilin A.
The interaction between CD147 and P-glycoprotein may be important for chemoresistance [80]. Other proteins also use CD147 as receptor, such as cyclophilins, Platelet glycoprotein VI (GPVI) which intervenes in platelet adhesion and S100A9, a component of the heterodimeric protein calprotectin, which is released during tissue damage and is implicated in inflammation and metastasis [78,79,80,81].
4. Regulation of Energy Metabolism in Hematological malignancies
A distinctive feature of cancer cells is their ability to sustain increased energy needs by adopting metabolic reprogramming that allows for a high rate of growth and proliferation [7]. Metabolic dysregulation that is associated to Hematologic Malignancies, such as leukemias, lymphomas, myelomas and MDS, involves various mechanisms that activate signaling pathways and regulate the expression of metabolism-related genes. Taking into account the heterogeneity between these HM, identify their metabolic signatures and key pathways as potential therapeutic targets, may offer specific therapeutic opportunities for the treatment of specific subgroups of HM [7,82,83,84,85]
Metabolic rewiring is characterized by a faster production of ATP and metabolic intermediates for the biosynthesis of proteins, lipids, nucleotides and NADPH useful for neoplastic growth, accompanied by the activation of different antioxidant systems to maintain redox homeostasis [7].
Tumour metabolism is characterized by an enhanced aerobic glycolysis ("Warburg effect") to the rapid generation of ATP, instead of producing ATP through oxidative phosphorylation (OXPHOS), and by an increased production of lactic acid, to meet the pressing demands of intensively proliferating tumour cells [86,87]. The increase in glycolysis in cancer cells, is not due to a defective mitochondrial respiratory chain, as cancer cells have functional mitochondria, although they have reduced activity. This characteristic metabolic bipotentiality, allows cancer cells to change their metabolism from a glycolytic phenotype to an oxidative phenotype and vice versa, allowing rapid adaptation to the change in the microenvironment and to acidosis and decreased availability of nutrients and oxygen, which occur during neoplastic development [88]. These metabolic adaptations likely differ between AML subtypes [89] and even within the same patient's own leukemia cells [90].
CD147 promotes metabolic rewiring by favouring the adaptation of tumour cells, which leads to an advantage of selective growth to some cell subclones and allows cell proliferation. The metabolic regulation of CD147 occurs through its direct or indirect interaction with specific protein partners related to glycolysis, OXPHOS, hypoxia and lipolysis, which are dysregulated metabolic pathways in cancer including hematologic malignancies [91,92].
4.1. Acute Myeloid Leukemia
In Acute myeloid leukemia (AML), CD147 has already been reported to be overexpressed in AML cells [12], the knockdown of CD147 in AML cells suppressed growth and proliferation suggesting the role of CD147 in signaling pathways that contribute to disease pathophysiology [54]. The aberrant expression of CD147 directly affects the various tumour metabolic activities including glycolysis, due to its participation in lactate transport with MCT1 and MCT4 transporters. Through the inhibition of the two MCTs transporters respectively AR-C and SYRO, glycolysis was reduced, resulting in the accumulation of intracellular lactate, in ATP depletion, decreased cell pH, and arrest of cell proliferation in AML cells. Although AR-C and SYRO have a different impact on the rate of lactate import and export in leukemia cells, SYRO appears to be more efficient in blocking the lactate flow capacity of leukemia cells by predominantly inhibiting the export of lactate via MCT4 targeting. The CD147-MCT1/4 axis was therefore crucial to support lactate and H+ trafficking and pH balance in AML. In another study in vitro, were compared antileukemic effects of seven metabolic inhibitors on patient-derived AML cells, targeting various metabolic pathways including glycolysis (among these the selective MCT1 inhibitor AZD3965), pentose phosphate pathway, glutaminolysis, and fatty acid oxidation. The metabolic inhibitors that had the greatest antiproliferative and proapoptotic effects on leukemia cells were found to be the glycolysis inhibitors that exerted a stronger effect for most patients [93].
Song et al. have found that AML cell chemoresistance was associated with increased glycolytic activity and low oxidative phosphorylation efficiency. Through expression analyses in HL-60 adriamycin-resistant cells and HL-60 sensitive cells, the authors showed that the increase in chemoresistance was linked to the aberrant expression of glycolysis-related molecules, such as hypoxia-inducible factor (HIF)-1α, glucose transporter (GLUT)1, hexokinase-II (HK-II), aldehyde dehydrogenase (ALDH) and CD147, in chemo-resistant cells. In addition, the authors demonstrated that inhibition of glycolysis with 2-DG or 3 BrPA can reverse the chemoresistance of the adriamycin-resistant HL-60 AML cell line [94,95].
Several studies have reported the physical association of the CD98 protein with CD147 to form a complex on the cell surface of different solid tumors and leukemias, highlighting the role of the CD147/CD98 complex and its contribution in both adhesive signaling and modulation of metabolic function [73,96]. From the study of this complex in different tumor cell lines, it has been shown that the loss of CD98hc inhibits the absorption of amino acids and glucose, suppresses glycolysis, interfering with the pentose phosphate (PPP) pathway, oxidative stress and cell cycle arrest [74,97]. CD98 is overexpressed in AML and associated with poor prognosis, demonstrating its crucial role in drives malignant transformation and tumour progression, as also shown for CD147 in AML [12,17,74]. In addition, CD147 is involved in the engraftment and propagation of primary leukemia cells in their microenvironment favouring tumour progression [74,96]. To further elucidate the role of CD98 in leukemogenesis, Bajaj and colleagues have deleted CD98 in mouse models of AML, increasing the survival of AML mice, while the proliferative and self-renewal abilities of normal hematopoietic stem cells (HSCs) were reduced in non-tumor mice; In addition, CD98 knockdown resulted in a decrease in the frequency of leukemic stem cells (LSCs). Furthermore, authors found that CD98-integrin-mediated signalling and CD98-mediated amino acid transport contribute to AML growth and LSCs survival [74].
4.2. Chronic Myeloid Leukemia
Biochemical analysis of a panel of imatinib-resistant Chronic Myeloid Leukemia (CML) cell lines and mononuclear cells isolated from CML patients who had relapsed after imatinib treatment, showed increased glucose uptake and increased glycolysis. Metformin treatment of these cells exerted antiproliferative effects by inhibiting the CD147 co-binding proteins, MCT1 and MCT4 transporters, leading to inhibition of lactate export. Metformin-mediated inhibition occurs through regulation of mTORC1 and HIF-1α. In addition, glucose uptake and ATP production were also inhibited following metformin treatment due to AMPK-dependent inhibition of GLUT1 and HK-II [98].
Although a direct relationship between CD147 and metabolic regulation in CML is still to be explored, a recent study "in vivo" investigated the antitumor efficacy of a novel CD147-targeting antibody (h4 # 147D) in three xenograft mouse model that housed chemo-resistant human cell lines from different cancers. In mouse xenograft model inoculated with KU812 CML cells (with low sensitivity to imatinib), treatment with anti-CD147 antibody exerts a 99% antitumor effect compared to the imatinib group (TGI 48%), resulting in complete tumor regression. In addition, analysis of xenograft tumor tissues after treatment with the CD147 antibody highlighted the reductions in the levels of CD147 and its binding proteins (CD44, integrin α 3, integrin α 6 and MCT1) [99].
4.3. Lymphoma
Several studies have shown that CD147 is overexpressed in different lymphoma and leukemia cell lines but not in normal CD3+ T cells [55,100,101].
The involvement of CD147 in the regulation of glycolysis has been demonstrated in anaplastic large cell lymphoma (ALCL), a T-cell lymphoma [55]. A recent study showed that CD147 resulted differentially expressed in systemic ALK (Anaplastic lymphoma kinase)-positive ALCL (ALK+ ALCL) versus ALK-negative ALCL (ALK− ALCL) and normal T cells as CD147 is a direct target of miR-146. Interestingly, the authors observed that CD147 contributes to the survival and proliferation of ALK+ ALCL cells in vitro and to the engraftment and tumour growth in vivo, in an ALK+ALCL-xenotransplant mouse model. The direct involvement of CD147 with glycolysis was demonstrated through CD147 knockdown in ALK+ALCL cells which led to loss of MCT1 expression, reduction of glucose consumption, and retardation of tumour growth [102].
In another study, the role of CD147 in the progression of T lymphoma was determined by a CD147-specific siRNA in the T cell line Jurkat CD147-expressing. Silencing CD147 expression, causes decreased proliferation and migration of Jurkat cells and reduced Jurkat cell adhesion to extracellular matrix fibronectin in vitro [103].
The possibility of CD147 interaction with other protein partners has favoured the exploration of possible therapeutic targets. The interaction of CD147 and CypA is involved in cutaneous T-cell lymphoma (CTCL), a rare T-cell lymphoma. CD147 and CypA are both overexpressed in CTCL cells, blocking these proteins with an anti-CD147 antibody and/or anti-CypA antibody, suppresses the proliferation of CTCL cell lines, both in vitro and in vivo, via downregulation of phosphorylated extracellular-regulated kinase 1/2 and Akt [104].
4.4. Chronic Lymphocytic Leukemia
Alteration of glycolysis characterizes Chronic Lymphocytic Leukemia (CLL) malignant cells [105]. CLL-B cell interactions with the supportive tissue microenvironment play a critical role in disease pathogenesis. CD147 has been described in CLL-B cells, and its expression functionally linked to leukemic clone activation during disease progression [106], as well as with CLL-B cell migration and BM colonization [14]. Expression of CD147 is heterogeneous and was detected along with increased MMP9/MMP2 activity in plasma samples from CLL patients compared to healthy subjects. In this context, it was shown that CD147 is associated with MMPs production by tumor and stromal cells, highlighting the role of CD147 and MMPs in the pathophysiology of CLL [107].
4.5. Multiple Myeloma and Myelodysplastic Syndrome
Myeloma metabolism is characterized by increased glycolysis and the use of fatty acid oxidation that promote tumour survival and proliferation. CD147 is involved in the modulation of aerobic glycolysis of Multiple Myeloma (MM) through the regulation of MCT1 expression and lactate exportation [108]. Studies in MM have shown that CD147 expression is increased, and this increase accompanies disease progression [109]. The transporters MCT1 and MCT4, partners of CD147 in lactate transport, were also overexpressed in MM, but only MCT1 appears to have a role in myeloma cell proliferation and to promote tumour growth within an acidic microenvironment [108]. In addition, single nucleotide polymorphisms (SNPs) in the gene encoding for CD147 (BSG SNPs)) and MCT1 (SLC16A1 SNPs) have been shown to affect MM survival [110]. The development and progression of myeloma are also linked to its ability to modify the behaviour of surrounding cells so that they make the tumour microenvironment more favourable to its growth and survival. From this point of view, MM is a type of cancer that more than others need the support of its own microenvironment that provides growth signals and survival factors. In addition, interactions with bone marrow stromal cells and alterations in oxygen metabolism contribute to MM cells' resistance to chemotherapy treatments.
Several studies have highlighted the involvement of CD147 in the response to chemotherapy treatment of MM [111], among these, the study by Eichner et al. found that immunomodulatory drugs (IMiDs), such as thalidomide and its derivatives lenalidomide and pomalidomide, commonly used for the treatment of MM and myelodysplastic syndrome (MDS) with isolated chromosome 5q deletion (del(5q) MDS), which constitutes the only MDS subtype clearly defined by a cytogenetic alteration, bind Cereblon (CRBN), a substrate receptor of the ubiquitin ligase complex CRL4, mediating antitumor and teratogenic effects [16]. From the analysis of primary MM and CD34+ MDS cells treated with IMiD, it was found that CRBN functions via an ubiquitin-independent chaperone-like mechanism to mediate the folding and maturation of the CD147 and MCT1 proteins, thereby allowing activation of the CD147–MCT1 transmembrane complex. IMiDs treatment abrogate this mechanism in a competitive manner to mediate their antitumor and teratogenic activities. As a result, IMiD-sensitive MM cells lose CD147 and MCT1 expression after being exposed to IMiDs, while IMiD-resistant cells retain their expression. In addition, del(5q) MDS cells have elevated CD147 expression, which attenuates after IMiD treatment. This study showed that the elimination of CD147 in zebrafish restores the teratogenic effects of thalidomide exposure [16].
5. CD147 in Leukemic Microenvironment
The impact of metabolism on the leukemic microenvironment is crucial for the proliferation of hematopoietic neoplasms. The regulation of the leukemic microenvironment is complex, the so-called "niches" within the bone marrow (BM), lymph nodes and secondary lymphoid organs, are composed of different non-tumour cell types, which can interact with each other and with tumour cells, providing regulatory factors and growth factors to proximal malignant cells supporting their growth [112,113,114]. Several studies have also revealed that the microenvironment is characterized by a hypoxic environment and that the intercellular interaction of leukemia cells with stromal cells can influence cellular metabolism and consequently play an important role in the growth and survival of leukemia cells [115,116,117,118]. In this context, CD147 is at the centre of a crossroads, directly or through signalling to downstream molecules, and favours the hypoxic and metabolic adaptation of leukemia stem cells [114].
Analysis of the immunophenotypic expression of mesenchymal stromal cells derived from bone marrow and umbilical cord blood, showed that the CD147 protein is expressed by tissue-specific stem cells [119]. In addition, CD147 has been previously shown to regulate the expression of hypoxia-inducible factor 1 α (HIF1-α) and HIF-2α, factors involved in angiogenesis, hypoxic and metabolic reprogramming of different types of neoplasms [120,121,122].
Interestingly, Bougatef et al observed that HUVEC endothelial cells express endogenous CD147 and showed that CD147 expression is responsible, through the transcription factor HIF-2α, for the increased production of soluble VEGF isoforms and its main receptor VEGFR-2, both in vitro and in experimental tumour models in vivo [123]. The authors also found increase in VEGFR-2 “in vivo”, in a mouse model of xenograft tumours that had an altered level of CD147 expression.
In the BM microenvironment, leukemia cells can be favoured by the increase in VEGF, as this factor regulates the expression of the lactate transporter MCT1, allowing the intracellular transport of lactate to be used for production of energy and nutrients useful for tumour growth [124]. Dependence on this factor also drives the localization of leukemia cells within the marrow according to the VEGF gradient. Through the analysis of BM samples from patients with M0-M5 AML, which express high levels of MCT1, it was observed that leukemia cells prefer to localize near the bones (paratrabecular region) of the BM where VEGF production is active [124].
Regarding metabolites in the leukemic microenvironment, it is known that lactate in addition to being present inside cells to be used for energy production, can also be used as a signaling molecule through the lactate receptor G-protein-coupled receptor 81 (GPR81) under physiological conditions [125]. Lactate-GPR81 signaling had previously been observed to drive the growth and repopulation of breast cancer cells [126].
A recent study, through a metabolomics analysis, identified metabolites in extracellular fluid derived from BM biopsies of AML patients. The result of these investigations showed that the levels of extracellular metabolites were elevated in the BM of AML patients, including lactate. The increase in lactate is a critical factor in the dysfunction of BM microenvironment induced by AML and is responsible for the progression of leukemia. The metabolic characteristics make the leukemic bone marrow microenvironment (BMME) unique [127].
The expression of CD147 on leukemia cells surface, is also important for preferential homing of the BM. In an interesting work, using multiple myeloma (MM) as a model of a B-cell malignancy that frequently colonize the BM, it was shown that bone marrow endothelial cells (BMECs) secrete cyclophilin A (eCyPA), which promotes the migration, proliferation and colonization of leukemic cells through binding to its receptor CD147, on MM cells surface. The study revealed an altered level of eCyPA in BM serum compared to peripheral blood (PB) in people with MM and blocking eCyPA-CD147 suppresses BM homing and tumor growth in a mouse model of MM xenotransplantation. eCyPA also promoted the migration of chronic lymphocytic leukemia (CLL) and lymphoplasmacytic lymphoma (LPL) cells, two other malignant B-cell neoplasms that colonize BM and express CD147 [14].
It should also be considered that the niche, with its favorable microenvironment, constitutes a refuge for leukemia stem cells (LSCs) responsible for relapse, which in this environment are able to survive and proliferate after treatment with chemotherapy [128,129]. Gastel et al reported an analysis of metabolomics in AML LSC cells, revealing a distinct metabolic profile in the pre-chemo versus post-chemo relapse phase [130].
Another study observed that CML LSCs use the hypoxic niche to escape treatment with imatinib. In detail, the hypoxic environment promoted the upregulation of hypoxia-inducible factor 1 α (HIF1-α) to allow survival independent of BCR-ABL. Along with HIF1-α, other pro-survival genes are also stimulated that are preferentially upregulated by hypoxia in CML compared to normal CD34+ blood progenitors [131].
6. Therapeutic Targeting of CD147
Numerous studies have ascertained the crucial role played by the aberrant expression of CD147 in cancer progression, demonstrating the potential application of CD147 not only as a potential diagnostic marker of the disease [50], but also as an effective therapeutic target for the development of new anticancer therapies (Table 1).
Several preclinic studies have reported decreased expression of the CD147 protein through the treatment with small compounds of synthetic or natural origin, and with RNAi [12,54,132], in addition, new anti-CD147 antibodies [99,133,134] or antigenic peptide vaccines [99,133,135] have been designed in order to block CD147 function.
We have reported previously the study on the small molecule AC-73 able to block the dimerization of CD147 which led to the inhibition of the proliferation of leukemia cells and the induction of autophagy. In addition to an antiproliferative action, AC-73 was able to improve sensitivity to chemotherapy treatment and allowed a decrease in the dosage of chemotherapy (arabinosylcytosine and arsenic trioxide) [12].
Recently, the study by Zou et al showed that pseudolaric acid B (PAB), a bioactive compound of plant origin, was able to inhibit CD147 in several AML leukemia lines, confirming the role of CD147 in AML proliferation [54].
Although significant clinical progress has been made in the development of monoclonal antibodies CD147-directed for the treatment of different types of neoplasms including hepatocellular carcinoma (HCC) [136,137], Cervical Cancer [138], lung cancer [139] and other solid tumours [13], few studies have explored the application of anti-CD147 antibodies in the field of hematological malignancies.
Fukuchi and colleagues generated a humanized anti-CD147 antibody, which showed a potent antitumor effect in three mouse xenograft models including the CML model imatinib insensitive. Particularly, the authors found complete tumour regression in the group of xenograft mice treated with the antibody compared to the group of mice treated with the drug imatinib [99].
In an interesting study, K562/ADR cells resistant to the common chemotherapy adriamycin were found to have significantly higher levels of P-glycoprotein (P-gp) and CD147 than drug-free K562/ADR cells. Using monoclonal antibodies against CD147, mRNA and protein levels of P-gp and MDR1 were significantly reduced. Activation of CD147 by anti-CD147 antibody also reduced CD147 expression [134].
As CD147 had elevated expression levels in tumour T-cells from patients with ALL T-cell and T-cell lymphoma, it was considered a potential antigenic target of T-CAR cell therapy for T-ALL. CD147-CAR T cells, which contain a humanized single-chain variable fragment targeting CD147, showed potent antitumor activity against the human T-ALL cell line and T-ALL blasts, releasing high levels of cytokines in the process. However, CD147-CAR T cells have shown a potential safety concern against normal human cells and CD147-deficient cells, which must be considered for future therapy [140]. These results highlight the therapeutic potential of CD147-CAR-modified immune cells for patients with ALL.
Several studies suggested that CD147 glycoprotein plays a crucial role in resistance to anticancer drugs. Different tumours, including hematologic malignancies shown a positive correlation between the level of CD147 expression and resistance to different chemotherapy drugs. In lymphoid neoplasms, CD147 has been observed to increase resistance to chemotherapy drugs through the formation of a complex consisting of CD147, the endothelial lymphatic vessel hyaluronic acid receptor-1 (LYVE-1) and a drug transporter known as breast cancer resistance protein/ABCG2 (BCRP), located on the cell surface of a primary effusive lymphoma tumour cell line (PEL). Targeting of every single component of CD147/LYVE-1/BCRP complex with RNAi, showed the interdependent roles of the three molecules in chemotherapy resistance [141].
7. Conclusion and Future Directions
CD147 performs multiple metabolic functions that profoundly influence cellular metabolism and tumour microenvironment in hematologic malignancies. The high expression of this glycoprotein in haematological malignancies represents a metabolic vulnerability that can be exploited therapeutically. Its interaction with monocarboxylated transporters or other partners such as CD98 or Glut1 and the regulation of energy metabolism are crucial for the proliferation and survival of leukemia cells. Understanding these mechanisms offers new opportunities for the development of CD147-targeted therapies, potentially improving the treatment of hematologic malignancies. Future research should focus on exploring these therapeutic approaches, with the aim of translating this knowledge into clinical benefits for patients
Author Contributions
I.S., C.L., E.S. gathered the literature. E.S. wrote the first draft. I.S., C.L., and E.S. revised the final draft. E.S. finalized the submission. All authors have read and agreed to the published version of the manuscript.
Funding
E.S. acknowledges Istituto Superiore di Sanità (Rome, Italy) for supporting her research for the identification of new therapeutic targets in AML.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ramdass, B.; Chowdhary, A.; Koka, P.S. Hematological malignancies: disease pathophysiology of leukemic stem cells. J Stem Cells. 2013, 8, 151–187. [Google Scholar]
- Taylor, J.; Xiao, W.; Abdel-Wahab, O. Diagnosis and classification of hematologic malignancies on the basis of genetics. Blood. 2017, 130, 410–423. [Google Scholar] [CrossRef]
- Traba, J.; Sack, M.N.; Waldmann, T.A.; Anton, OM. Immunometabolism at the Nexus of Cancer Therapeutic Efficacy and Resistance. Front Immunol, 2012; 12, 657293. [Google Scholar]
- Cordoba, R.; Eyre, T.A.; Klepin, H.D.; Wildes, T.M.; Goede, V. A comprehensive approach to therapy of haematological malignancies in older patients. Lancet Haematol. 2021, 8, e840–e852. [Google Scholar] [CrossRef]
- Tang, L.; Huang, Z.; Mei, H.; Hu, Y. Immunotherapy in hematologic malignancies: achievements, challenges and future prospects. Signal Transduct Target Ther. 2023, 8, 306. [Google Scholar] [CrossRef]
- Sochacka-Cwikła, A.; Maczynski, M.; Regiec, A. FDA-Approved Drugs for Hematological Malignancies-The Last Decade Review. Cancers 2022, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhou, X.; Wang, X. Metabolic Reprogramming in Hematologic Malignancies: Advances and Clinical Perspectives. Cancer Res. 2022, 82, 2955–2963. [Google Scholar] [CrossRef]
- Stine, Z.E.; Schug, Z.T.; Salvino, J.M.; Dang, C.V. Targeting cancer metabolism in the era of precision oncology. Nat Rev Drug Discov. 2022, 21, 141–162. [Google Scholar] [CrossRef] [PubMed]
- San-Millán, I.; Brooks, G.A. Reexamining cancer metabolism: lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis 2017, 38, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Iacono, K.T.; Brown, A.L.; Greene, M.I.; Saouaf, S.J. CD147 immunoglobulin superfamily receptor function and role in pathology. Exp Mol Pathol. 2007, 83, 283–295. [Google Scholar] [CrossRef]
- Muramatsu, T. Basigin (CD147), a multifunctional transmembrane glycoprotein with various binding partners. J Biochem. 2016, 159, 481–490. [Google Scholar] [CrossRef]
- Spinello, I.; Saulle, E.; Quaranta, M.T.; Pasquini, L.; Pelosi, E.; Castelli, G.; Ottone, T.; Voso, M.T.; Testa, U.; Labbaye, C. The small-molecule compound AC-73 targeting CD147 inhibits leukemic cell proliferation, induces autophagy and increases the chemotherapeutic sensitivity of acute myeloid leukemia cells. Haematologica. 2019, 104, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Nyalali, A.M.K.; Leonard, A.U.; Xu, Y.; Li, H.; Zhou, J.; Zhang, X. Rugambwa, T.K., Shi, X. L,i F. CD147: an integral and potential molecule to abrogate hallmarks of cancer. In Front Oncol; 2023; Volume 13, p. 1238051. [Google Scholar]
- Zhu, D.; Wang, Z.; Zhao, J.J. The Cyclophilin A-CD147 complex promotes the proliferation and homing of multiple myeloma cells. Nat Med. 2015, 21, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Edwards, C.K. 3rd.; Zhou, L. The biological function and clinical utilization of CD147 in human diseases: a review of the current scientific literature. Int J Mol Sci. 2014, 15, 17411–17441. [Google Scholar] [CrossRef] [PubMed]
- Eichner, R.; Heider, M.; Fernández-Sáiz, V. Immunomodulatory drugs disrupt the cereblon-CD147-MCT1 axis to exert anti-tumor activity and teratogenicity. Nat Med. 2016, 22, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Landras, A.; Reger de Moura, C.; Jouenne, F.; Lebbe, C.; Menashi, S.; Mourah, S. CD147 Is a Promising Target of Tumor Progression and a Prognostic Biomarker. Cancers (Basel). 2019, 11, 1803. [Google Scholar] [CrossRef] [PubMed]
- Arber, D.A.; Orazi, A.; Hasserjian, R.P.; Borowitz, M.J.; Calvo, K.R.; Kvasnicka, H.M.; Wang, S.A.; Bagg, A.; Barbui, T.; Branford, S.; et al. International Consensus Classification of Myeloid Neoplasms and Acute Leukemias: integrating morphologic, clinical, and genomic data. Blood. 2022, 140, 1200–1228. [Google Scholar] [CrossRef] [PubMed]
- Khoury, J.D.; Solary, E.; Abla, O.; Akkari, Y.; Alaggio, R.; Apperley, J.F.; Bejar, R.; Berti, E.; Busque, L.; Chan, J.K.C.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Myeloid and Histiocytic/Dendritic Neoplasms. Leukemia. 2022, 36, 1703–1719. [Google Scholar] [CrossRef] [PubMed]
- Alaggio, R.; Amador, C.; Anagnostopoulos, I.; Attygalle, A.D.; Araujo, I.B.O.; Berti, E.; Bhagat, G.; Borges, A.M.; Boyer, D.; Calaminici, M.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748, Erratum in: Leukemia 2023, 37, 1944–1951. [Google Scholar] [CrossRef]
- DiNardo, C.D.; Erba, H.P.; Freeman, S.D.; Wei, A.H. Acute myeloid leukaemia. Lancet. 2023, 401, 2073–2086. [Google Scholar] [CrossRef]
- Mishra, S.K.; Millman, S.E.; Zhang, L. Metabolism in acute myeloid leukemia: mechanistic insights and therapeutic targets. Blood. 2023, 141, 1119–1135. [Google Scholar] [CrossRef]
- Pagliaro, L.; Chen, S.J.; Herranz, D.; Mecucci, C.; Harrison, C.J.; Mullighan, C.G.; Zhang, M.; Chen, Z.; Boissel, N.; Winter, S.S.; et al. Acute lymphoblastic leukaemia. Nat Rev Dis Primers. 2024, 10, 41. [Google Scholar] [CrossRef]
- Chan, L.N.; Chen, Z.; Braas, D.; Lee, J.W.; Xiao, G.; Geng, H.; Cosgun, K.N.; Hurtz, C.; Shojaee, S.; Cazzaniga, V.; et al. Metabolic gatekeeper function of B-lymphoid transcription factors. Nature. 2017, 542, 479–483. [Google Scholar] [CrossRef]
- Momotow, J.; Borchmann, S.; Eichenauer, D.A.; Engert, A.; Sasse, S. Hodgkin Lymphoma-Review on Pathogenesis, Diagnosis, Current and Future Treatment Approaches for Adult Patients. J Clin Med. 2021, 10, 1125. [Google Scholar] [CrossRef]
- Bowzyk Al-Naeeb, A.; Ajithkumar, T.; Behan, S.; Hodson, D.J. Non-Hodgkin lymphoma. BMJ, 2018; 362, 3204. [Google Scholar]
- Kluckova, K.; D'Avola, A.; Riches, J.C. Advances in Understanding of Metabolism of B-Cell Lymphoma: Implications for Therapy. Cancers (Basel). 2022, 14, 5552. [Google Scholar] [CrossRef] [PubMed]
- Sekeres, M.A.; Taylor, J. Diagnosis and Treatment of Myelodysplastic Syndromes: A Review. JAMA 2022, 328, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Gerke, M.B.; Christodoulou, I.; Karantanos, T. Definitions, Biology, and Current Therapeutic Landscape of Myelodysplastic/Myeloproliferative Neoplasms. Cancers (Basel). 2023, 15, 3815. [Google Scholar] [CrossRef] [PubMed]
- Gulla, A.; Anderson, K.C. Multiple myeloma: the (r)evolution of current therapy and a glance into future. Haematologica 2020, 105, 2358–2367. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Nandakumar, B.; Rajkumar, S.V.; Kapoor, P.; Buadi, F.K.; Dingli, D.; Lacy, M.Q.; Gertz, M.A.; Hayman, S.R.; Leung, N.; et al. Mortality trends in multiple myeloma after the introduction of novel therapies in the United States. Leukemia 2022, 36, 801–808. [Google Scholar] [CrossRef]
- Butera, A.; Quaranta, M.T.; Crippa, L.; Spinello, I.; Saulle, E.; Di Carlo, N.; Campanile, D.; Boirivant, M.; Labbaye, C. CD147 Targeting by AC-73 Induces Autophagy and Reduces Intestinal Fibrosis Associated with TNBS Chronic Colitis. J Crohns Colitis. 2022, 16, 1751–1761. [Google Scholar] [CrossRef]
- Chuliá-Peris, L.; Carreres-Rey, C.; Gabasa, M.; Alcaraz, J.; Carretero, J.; Pereda, J. Matrix Metalloproteinases and Their Inhibitors in Pulmonary Fibrosis: EMMPRIN/CD147 Comes into Play. Int. J. Mol. Sci. 2022, 23, 6894. [Google Scholar] [CrossRef]
- Pushkarsky, T.; Zybarth, G.; Dubrovsky, L.; Yurchenko, V.; Tang, H.; Guo, H.; Toole, B.; Sherry, B.; Bukrinsky, M. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A. Proc Natl Acad Sci U S A. 2001, 98, 6360–6365. [Google Scholar] [PubMed]
- Spinello, I.; Saulle, E.; Quaranta, M.T.; Pelosi, E.; Castelli, G.; Cerio, A.; Pasquini, L.; Morsilli, O.; Dupuis, M.L.; Labbaye, C. AC-73 and Syrosingopine Inhibit SARS-CoV-2 Entry into Megakaryocytes by Targeting CD147 and MCT4. Viruses, 2024, 16, 82. [Google Scholar] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: mechanisms and clinical implications. Nat Rev Cancer. 2015, 15, 540–555. [Google Scholar] [PubMed]
- Cui, J.; Huang, W.; Wu, B.; Jin, J.; Jing, L.; Shi, W.P.; Liu, Z.Y.; Yuan, L.; Luo, D.; Li. L.; et al. N-glycosylation by N-acetylglucosaminyltransferase V enhances the interaction of CD147/basigin with integrin β1 and promotes HCC metastasis. J Pathol. 2018, 245, 41–52. [Google Scholar]
- Li, W.; Wang, D.; Ge, Y.; Zhang, L.; Wu, J.; Liu, D. Discovery and Biological Evaluation of CD147 N-Glycan Inhibitors: A New Direction in the Treatment of Tumor Metastasis. Molecules. 2020, 26, 33. [Google Scholar]
- Kato, N.; Yuzawa, Y.; Kosugi, T.; Hobo, A.; Sato, W.; Miwa, Y.; Sakamoto, K.; Matsuo, S.; Kadomatsu, K. The E-selectin ligand basigin/CD147 is responsible for neutrophil recruitment in renal ischemia/reperfusion. J Am Soc Nephrol. 2009, 20, 1565–1576. [Google Scholar] [CrossRef]
- Bai, Y.; Huang, W.; Ma, L.T.; Jiang, J.; Chen, Z.N. Importance of N-glycosylation on CD147 for its biological functions. Int J Mol Sci. 2014, 15, 6356–6377. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Chang, S.B.; Hemler, M.E. Links between CD147 function, glycosylation, and caveolin-1. Mol Biol Cell. 2004, 15, 4043–4050. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, Y.; Jiang, L.; Miao, X.; Zhou, H.; Jia, L. Glycomic alterations are associated with multidrug resistance in human leukemia. Int J Biochem Cell Biol. 2012, 44, 1244–1253. [Google Scholar] [CrossRef]
- Beesley, A.H.; Weller, R.E.; Kees, U.R. The role of BSG (CD147) in acute lymphoblastic leukaemia and relapse. Br J Haematol. 2008, 142, 1000–1002. [Google Scholar] [CrossRef]
- Belton, R.J. Jr.; Chen, L.; Mesquita, F.S.; Nowak, R.A. Basigin-2 is a cell surface receptor for soluble basigin ligand. J. Biol. Chem. 2008, 283, 17805–17814. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.Y.; Guo, T.; Wang, S.J.; Zhao, P.; Dong, Z.S.; Zhang, Y.; Jiang, J.L.; Chen, Z.N.; Yu, XL. Dimerization is essential for HAb18G/CD147 promoting tumor invasion via MAPK pathway. Biochem Biophys Res Commun. 2012, 419, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, S.S.; Mengistab, A.T.; Tauscher, A.N.; LaVail, J.; Basbaum, C. The microvesicle as a vehicle for EMMPRIN in tumor-stromal interactions. Oncogene 2004, 23, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Rode, A.; Nicoll, A.; Maczurek, A.E.; Lim, L.; Lim, S.; Angus, P.; Kronborg, I.; Arachchi, N.; Gorelik, A.; et al. Circulating CD147 predicts mortality in advanced hepatocellular carcinoma. J Gastroenterol Hepatol. 2016, 31, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.H.; Liu, Y.J.; Tang, L.L.; Wang, S.M.; Yan, G.J.; Liao, L.Q. Plasma soluble cluster of differentiation 147 levels are increased in breast cancer patients and associated with lymph node metastasis and chemoresistance. Hong Kong Med J. 2018, 24, 252–260. [Google Scholar] [CrossRef]
- Rurali, E.; Perrucci, G.L.; Gaetano, R.; Pini, A.; Moschetta, D.; Gentilini, D.; Nigro, P.; Pompilio, G. Soluble EMMPRIN levels discriminate aortic ectasia in Marfan syndrome patients. Theranostics. 2019, 9, 2224–2234. [Google Scholar] [CrossRef] [PubMed]
- Łacina, P.; Butrym, A.; Turlej, E.; Stachowicz-Suhs, M.; Wietrzyk, J.; Mazur, G.; Bogunia-Kubik, K. BSG (CD147) Serum Level and Genetic Variants Are Associated with Overall Survival in Acute Myeloid Leukaemia. J Clin Med. 2022, 11, 332. [Google Scholar] [CrossRef]
- Łacina, P.; Butrym, A.; Frontkiewicz, D.; Mazur, G.; Bogunia-Kubik, K. Soluble CD147 (BSG) as a Prognostic Marker in Multiple Myeloma. Curr Issues Mol Biol. 2022, 44, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Kasinrerk, W.; Fiebiger, E.; Stefanová, I.; Baumruker, T.; Knapp, W.; Stockinger, H. Human leukocyte activation antigen M6, a member of the Ig superfamily, is the species homologue of rat OX-47, mouse basigin, and chicken HT7 molecule. J Immunol. 1992, 149, 847–854. [Google Scholar] [CrossRef]
- Saulle, E. , Spinello, I., Quaranta, MT., Pasquini, L., Pelosi, E., Iorio, E., Castelli, G., Chirico, M., Pisanu, M.E., Ottone, T., Voso, M.T., Testa, U., Labbaye, C. Targeting Lactate Metabolism by Inhibiting MCT1 or MCT4 Impairs Leukemic Cell Proliferation, Induces Two Different Related Death-Pathways and Increases Chemotherapeutic Sensitivity of Acute Myeloid Leukemia Cells. Front Oncol. 2021, 1, 621458. [Google Scholar]
- Zou, S.; Parfenova, E.; Vrdoljak, N.; Minden, M.D.; Spagnuolo, P.A. Pseudolaric Acid B Targets CD147 to Selectively Kill Acute Myeloid Leukemia Cells. Int J Mol Sci. 2024, 25, 6517. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Bonzheim, I.; Steinhilber, J.; Montes-Mojarro, I.A.; Ortiz-Hidalgo, C.; Klapper, W.; Fend, F.; Quintanilla-Martínez, L. EMMPRIN (CD147) is induced by C/EBPβ and is differentially expressed in ALK+ and ALK- anaplastic large-cell lymphoma. Lab Invest. 2017, 97, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Panchabhai, S.; Schlam, I.; Sebastian, S.; Fonseca, R. PKM2 and other key regulators of Warburg effect positively correlate with CD147 (EMMPRIN) gene expression and predict survival in multiple myeloma. Leukemia 2017, 31, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Caudroy, S.; Polette, M.; Nawrocki-Raby, B.; Cao, J.; Toole, B.P.; Zucker, S.; Birembaut, P. EMMPRIN-mediated MMP regulation in tumor and endothelial cells. Clin Exp Metastasis. 2002, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Ghandour, F.; Kassem, S.; Simanovich, E.; Rahat, M.A. Glucose Promotes EMMPRIN/CD147 and the Secretion of Pro-Angiogenic Factors in a Co-Culture System of Endothelial Cells and Monocytes. Biomedicines 2024, 12, 706. [Google Scholar] [CrossRef] [PubMed]
- Kanekura, T. CD147/Basigin Is Involved in the Development of Malignant Tumors and T-Cell-Mediated Immunological Disorders via Regulation of Glycolysis. Int. J. Mol. Sci. 2023, 24, 17344. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, W. CD147-mediated reprogrammed glycolytic metabolism potentially induces immune escape in the tumor microenvironment (Review). Oncol Rep. 2019, 41, 2945–2956. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.; Wilson, M.C.; Heddle, C.; Brown, M.H.; Barclay, A.N.; Halestrap, A.P. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J. 2000, 19, 3896–3904. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P. The monocarboxylate transporter family--Structure and functional characterization. IUBMB Life. 2012, 64, 1–9. [Google Scholar] [CrossRef]
- Aït-Ali, N.; Fridlich, R.; Millet-Puel, G.; Clérin, E.; Delalande, F.; Jaillard, C.; Blond, F.; Perrocheau, L.; Reichman, S.; Byrne, L.C.; et al. Rod-derived cone viability factor promotes cone survival by stimulating aerobic glycolysis. Cell. 2015, 161, 817–832. [Google Scholar] [CrossRef]
- Almeida, L.M.C.A.; Silva, R.; Cavadas, B.; Lima, J.; Pereira, L.; Soares, P.; Sobrinho-Simões, M.; Lopes, J.M.; Máximo, V. GLUT1, MCT1/4 and CD147 overexpression supports the metabolic reprogramming in papillary renal cell carcinoma. Histol Histopathol. 2017, 32, 1029–1040. [Google Scholar] [PubMed]
- Huang, X.Q.; Chen, X.; Xie, X.X.; Zhou, Q.; Li, K.; Li, S.; Shen, L.F.; Su, J. Co-expression of CD147 and GLUT-1 indicates radiation resistance and poor prognosis in cervical squamous cell carcinoma. Int J Clin Exp Pathol. 2014, 7, 1651–1666. [Google Scholar] [PubMed]
- Su, J.; Gao, T.; Jiang, M.; Wu, L.; Zeng, W.; Zhao, S.; Peng, C.; Chen, X. CD147 silencing inhibits tumor growth by suppressing glucose transport in melanoma. Oncotarget. 2016, 7, 64778–64784. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Zabala, M.; Ramakrishnan, R.; Reinbach. ; Ghosh, S.; Oburoglu, L.; Falqués-Costa, A.; Bellamkonda, K.; Ehinger, M.; Peña-Martínez, P.; Puente-Moncada, N.; et al. Combined GLUT1 and OXPHOS inhibition eliminates acute myeloid leukemia cells by restraining their metabolic plasticity. Blood Adv. 2023, 7, 5382–5395. [Google Scholar] [CrossRef]
- Hartmann, S.; Agostinelli, C.; Diener, J.; Döring, C.; Fanti, S.; Zinzani, P.L.; Gallamini, A.; Bergmann, L.; Pileri, S.; Hansmann, M.L. GLUT1 expression patterns in different Hodgkin lymphoma subtypes and progressively transformed germinal centers. BMC Cancer. 2012, 12, 586. [Google Scholar] [CrossRef]
- Shim, H.K.; Lee, W.W.; Park, S.Y.; Kim, H.; Kim, S.E. Relationship between FDG uptake and expressions of glucose transporter type 1, type 3, and hexokinase-II in Reed-Sternberg cells of Hodgkin lymphoma. Oncol Res. 2009, 17, 331–337. [Google Scholar] [CrossRef]
- McBrayer, S.K.; Cheng, J.C.; Singhal, S.; Krett, N.L.; Rosen, S.T.; Shanmugam, M. Multiple myeloma exhibits novel dependence on GLUT4, GLUT8, and GLUT11: implications for glucose transporter-directed therapy. Blood. 2012, 119, 4686–4697. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Jimi, S.; Migita, K.; Takamatsu, Y.; Hara, S. Inhibition of glucose transporter 1 induces apoptosis and sensitizes multiple myeloma cells to conventional chemotherapeutic agents. Leuk Res. 2016, 41, 103–110. [Google Scholar] [CrossRef]
- Berditchevski, F.; Chang, S.; Bodorova, J.; Hemler, M.E. Generation of monoclonal antibodies to integrin-associated proteins. Evidence that alpha3beta1 complexes with EMMPRIN/basigin/OX47/M6. J Biol Chem. 1997, 272, 29174–29180. [Google Scholar] [CrossRef]
- Xu, D.; Hemler, M.E. Metabolic activation-related CD147-CD98 complex. Mol Cell Proteomics 2005, 4, 1061–1071. [Google Scholar] [CrossRef]
- Bajaj, J.; Konuma, T.; Lytle, N.K.; Kwon, H.Y.; Ablack, J.N.; Cantor, J.M.; Rizzieri, D.; Chuah, C.; Oehler, V.G.; Broome, E.H.; et al. CD98-Mediated Adhesive Signaling Enables the Establishment and Propagation of Acute Myelogenous Leukemia. Cancer Cell 2016, 30, 792–805. [Google Scholar] [CrossRef]
- Devés, R.; Boyd, C.A. Surface antigen CD98(4F2): not a single membrane protein, but a family of proteins with multiple functions. J Membr Biol. 2000, 173, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Ikeda, S.; Yaga, M.; Watanabe, K.; Urakawa, R.; Iehara, A.; Iwai, M.; Hashiguchi, S.; Morimoto, S.; Fujiki, F.; et al. Selective targeting of multiple myeloma cells with a monoclonal antibody recognizing the ubiquitous protein CD98 heavy chain. Sci Transl Med. 2022, 14, eaax7706. [Google Scholar] [CrossRef] [PubMed]
- Hayes, G.M.; Chinn, L.; Cantor, J.M.; Cairns, B.; Levashova, Z.; Tran, H.; Velilla, T.; Duey, D.; Lippincott, J.; Zachwieja, J.; et al. Antitumor activity of an anti-CD98 antibody. Int J Cancer 2015, 137, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, V.; Constant, S.; Eisenmesser, E.; Bukrinsky, M. Cyclophilin-CD147 interactions: a new target for anti-inflammatory therapeutics. Clin Exp Immunol. 2010, 160, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, V.; Zybarth, G.; O'Connor, M.; Dai, W.W.; Franchin, G.; Hao, T.; Guo, H.; Hung, H.C.; Toole, B.; Gallay, P.; et al. Active site residues of cyclophilin A are crucial for its signaling activity via CD147. J Biol Chem. 2002, 277, 22959–22965. [Google Scholar] [CrossRef]
- Wang, W.J.; Li, Q.Q.; Xu, J.D.; Cao, X.X.; Li, H.X.; Tang, F.; Chen, Q.; Yang, J.M.; Xu, Z.D.; Liu, X.P. Interaction between CD147 and P-glycoprotein and their regulation by ubiquitination in breast cancer cells. Chemotherapy, 2008; 54, 291–301. [Google Scholar]
- Seizer, P.; Borst, O.; Langer, H.F.; Bültmann, A.; Münch, G.; Herouy, Y.; Stellos, K.; Krämer, B.; Bigalke, B.; Büchele, B.; et al. EMMPRIN (CD147) is a novel receptor for platelet GPVI and mediates platelet rolling via GPVI-EMMPRIN interaction. Thromb Haemost. 2009, 101, 682–686. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.; Wang, J.H.; Zhao, A.H.; Xu, X.; Wang, Y.H.; Chen, T.L.; Li, J.M.; Mi, J.Q.; Zhu, Y.M.; Liu, Y.F.; et al. A distinct glucose metabolism signature of acute myeloid leukemia with prognostic value. Blood. 2014, 124, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, M.T.; Panagopoulou, P.; Paramera, E.; Pechlivanis, A.; Virgiliou, C.; Papakonstantinou, E.; Palabougiouki, M.; Ioannidou, M.; Vasileiou, E.; Tragiannidis, A.; et al. Metabolic Fingerprint in Childhood Acute Lymphoblastic Leukemia. Diagnostics (Basel). 2024, 14, 682. [Google Scholar]
- Maiso, P.; Huynh, D.; Moschetta, M.; Sacco, A.; Aljawai, Y.; Mishima, Y.; Asara, J.M.; Roccaro, A.M.; Kimmelman, A.C.; Ghobrial, I.M. Metabolic signature identifies novel targets for drug resistance in multiple myeloma. Cancer Res. 2015, 75, 2071–2082. [Google Scholar] [CrossRef]
- Balaian, E.; Wobus, M.; Bornhäuser, M.; Chavakis, T.; Sockel, K. Myelodysplastic Syndromes and Metabolism. Int. J. Mol. Sci. 2021, 22, 11250. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, M.; Szczudło, J.; Pietrzyk, A.; Shah, J.; Trojan, S.E.; Ostrowska, B.; Kocemba-Pilarczyk, K.A. The Warburg effect: a score for many instruments in the concert of cancer and cancer niche cells. Pharmacol Rep. 2023, 75, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does It Benefit Cancer Cells? Trends Biochem Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Pereira, O.; Teixeira, A.; Sampaio-Marques, B.; Castro, I.; Girao, H.; Ludovico, P. Signalling mechanisms that regulate metabolic profile and autophagy of acute myeloid leukaemia cells. J. Cell Mol. Med. 2018, 22, 4807–4817. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, N.; Poulain, L.; Birsen, R.; Tamburini, J.; Bouscary, D. Rationale for Targeting Deregulated Metabolic Pathways as a Therapeutic Strategy in Acute Myeloid Leukemia. Front Oncol. 2019, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz Concepción, B.; Bartolo-García, L.D.; Tizapa-Méndez, M.D.; Martínez-Vélez, M.; Valerio-Diego, J.J.; Illades-Aguiar, B.; Salmerón-Bárcenas, E.G.; Ortiz-Ortiz, J.; Torres-Rojas, F.I.; Mendoza-Catalán, M.Á.; et al. EMMPRIN is an emerging protein capable of regulating cancer hallmarks. Eur Rev Med Pharmacol Sci. 2022, 26, 6700–6724. [Google Scholar] [PubMed]
- Kendrick, A.A.; Schafer, J.; Dzieciatkowska, M.; Nemkov, T.; D'Alessandro, A.; Neelakantan, D.; Ford, H.L.; Pearson, C.G.; Weekes, C.D.; Hansen, K.C.; et al. CD147: a small molecule transporter ancillary protein at the crossroad of multiple hallmarks of cancer and metabolic reprogramming. Oncotarget 2017, 8, 6742–6762. [Google Scholar] [CrossRef] [PubMed]
- Grønningsæter, I.S.; Reikvam, H.; Aasebø, E.; Bartaula-Brevik, S.; Tvedt, T.H.; Bruserud, Ø.; Hatfield, K.J. Targeting Cellular Metabolism in Acute Myeloid Leukemia and The Role of Patient Heterogeneity. Cells 2020, 9, 1155. [Google Scholar] [CrossRef]
- Kui, S.; Xiaojun, X.; Li, X.; Xiao, Z.; Fuhua, Z.; Ke, Z.; Meiqing, W.; Ren, L.; Qifa, L. The Expression Profile of Glycolysis Associated Molecules and Chemoresistance in Acute Myeloid Leukemia. Blood 2012, 120, 4318. [Google Scholar]
- Song, K.; Li, M.; Xu, X.; Xuan, L.I.; Huang, G.; Liu, Q. Resistance to chemotherapy is associated with altered glucose metabolism in acute myeloid leukemia. Oncol Lett. 2016, 12, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Fox, D.A.; Horejsi, V.; Sagawa, K.; Skubitz, K.M.; Katz, D.R.; Chain, B. The functional interactions between CD98, beta1-integrins, and CD147 in the induction of U937 homotypic aggregation. Blood 2001, 98, 374–382. [Google Scholar] [CrossRef] [PubMed]
- de la Ballina, L.R.; Cano-Crespo, S.; González-Muñoz, E.; Bial, S.; Estrach, S.; Cailleteau, L.; Tissot, F.; Daniel, H.; Zorzano, A.; Ginsberg, M.H.; et al. Amino Acid Transport Associated to Cluster of Differentiation 98 Heavy Chain (CD98hc) Is at the Cross-road of Oxidative Stress and Amino Acid Availability. J Biol Chem. 2016, 291, 9700–9711. [Google Scholar] [CrossRef] [PubMed]
- Gayatri, M.B.; Kancha, R.K.; Patchva, D.; Velugonda, N.; Gundeti, S.; Reddy, A.B.M. Metformin exerts antileukemic effects by modulating lactate metabolism and overcomes imatinib resistance in chronic myelogenous leukemia. FEBS J. 2023, 290, 4480–4495. [Google Scholar] [CrossRef]
- Fukuchi, K.; Nanai, K.; Yuita, H.; Maru, C.; Tsukada, J.; Ishigami, M.; Nagai, Y.; Nakano, Y.; Yoshimura, C.; Yoneda, K.; et al. Novel Antibody Exerts Antitumor Effect through Downregulation of CD147 and Activation of Multiple Stress Signals. J Oncol. 2022, 2022, 3552793. [Google Scholar] [CrossRef] [PubMed]
- Thorns, C.; Feller, A.C.; Merz, H. EMMPRIN (CD 147) is expressed in Hodgkin's lymphoma and anaplastic large cell lymphoma. An immunohistochemical study of 60 cases. Anticancer Res. 2002, 22, 1983–1986. [Google Scholar] [PubMed]
- Nabeshima, K.; Suzumiya, J.; Nagano, M.; Ohshima, K.; Toole, B.P.; Tamura, K.; Iwasaki, H.; Kikuchi, M. Emmprin, a cell surface inducer of matrix metalloproteinases (MMPs), is expressed in T-cell lymphomas. J Pathol. 2004, 202, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Montes-Mojarro, I.A.; Steinhilber, J.; Griessinger, C.M.; Rau, A.; Gersmann, A.-K.; Kohlhofer, U.; Fallier-Becker, P.; Liang, H.-C.; Hofmann, U.; Haag, M.; et al. CD147 a direct target of miR-146a supports energy metabolism and promotes tumor growth in ALK+ ALCL. Leukemia 2022, 36, 2050–2063. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Su, J.; Chang, J.; Kanekura, T.; Li, J.; Kuang, Y.H.; Peng, S.; Yang, F.; Lu, H.; Zhang, J.L. Inhibition of CD147 gene expression via RNA interference reduces tumor cell proliferation, activation, adhesion, and migration activity in the human Jurkat T-lymphoma cell line. Cancer Investig. 2008, 26, 689–697. [Google Scholar] [CrossRef]
- Sakamoto, M.; Miyagaki, T.; Kamijo, H.; Oka, T.; Boki, H.; Takahashi-Shishido, N.; Suga, H.; Sugaya, M.; Sato, S. CD147-Cyclophilin a Interactions Promote Proliferation and Survival of Cutaneous T-Cell Lymphoma. Int. J. Mol. Sci. 2021, 22, 7889. [Google Scholar] [CrossRef]
- Donnelly, R.P; Finlay, D.K. Glucose, glycolysis and lymphocyte responses. Mol Immunol. 2015, 68 Pt C, 513–519. [Google Scholar] [CrossRef]
- Prieto, D.; Sotelo, N.; Seija, N.; Sernbo,S. ; Abreu, C.; Duran, R.; Gil, M.; Sicco, E.; Irigoin, V.; Oliver, C.; Landoni, A.I.; Gabus, R.; Dighiero, G.; Oppezzo, P. S100-A9 protein in exosomes from chronic lymphocytic leukemia cells promotes NF-kappaB activity during disease progression. Blood 2017, 130, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.M.; Gilardoni, M.B.; Remedi, M.M.; Sastre, D.; Heller, V.; Pellizas, C.G.; Donadio, A.C. Tumor-stroma interaction increases CD147 expression in neoplastic B lymphocytes in chronic lymphocytic leukemia. Blood Cells Mol Dis. 2020, 82, 102405. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.K.; Arendt, B.K.; Jelinek, D.F. CD147 regulates the expression of MCT1 and lactate export in multiple myeloma cells. Cell Cycle, 2013; 12, 3175–3183. [Google Scholar]
- Arendt, B.K.; Walters, D.K.; Wu, X.; Tschumper, R.C.; Huddleston, P.M.; Henderson, K.J.; Dispenzieri, A.; Jelinek, D.F. Increased expression of extracellular matrix metalloproteinase inducer (CD147) in multiple myeloma: Role in regulation of myeloma cell proliferation. Leukemia 2012, 26, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Łacina, P.; Butrym, A.; Mazur, G.; Bogunia-Kubik, K. BSG and MCT1 Genetic Variants Influence Survival in Multiple Myeloma Patients. Genes (Basel) 2018, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Bolomsky, A.; Hübl, W.; Spada, S.; Müldür, E.; Schlangen, K.; Heintel, D.; Rocci, A.; Weißmann, A.; Fritz, V.; Willheim, M.; et al. IKAROS expression in distinct bone marrow cell populations as a candidate biomarker for outcome with lenalidomide-dexamethasone therapy in multiple myeloma. Am J Hematol. 2017, 92, 269–278. [Google Scholar] [PubMed]
- Winkler, I.G.; Barbier, V.; Nowlan, B.; et al. The vascular niche E-selectin regulates hematopoietic stem cell dormancy, self-renewal and chemoresistance. Nat Med. 2012, 18, 1651–1657. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, A. ; Hsu, Y-M. S.; Day, R.B.; Schuettpelz, L.G.; Christopher, M.J.; Borgerding, J.N.; Nagasawa, T.; Link, D.C. CXCL12 in early mesenchymal progenitors is required for the maintenance of hematopoietic stem cells. Nature, 2013; 495, 227–230. [Google Scholar]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar]
- Wang, Y.H.; Israelsen, W.J.; Lee, D.; Yu, V.W.; Jeanson, N.T.; Clish, C.B.; et al. Specific metabolic dependence of cell status in hematopoiesis and leukemogenesis. Cell 2014, 158, 1309–1323. [Google Scholar] [CrossRef]
- Saito, Y.; Chapple, R.H.; Lin, A.; Kitano, A.; Nakada, D. AMPK protects the cells that initiate leukemia in myeloid leukemias from metabolic stress in the bone marrow. Cell Stem cell 2015, 17, 585–596. [Google Scholar]
- Zhang, W.; Trachootham, D.; Liu, J.; Chen, G.; Pelicano, H.; Garcia-Prieto, C.; et al. Stromal control of cystine metabolism promotes cancer cell survival in chronic lymphocytic leukemia. Nat Cell Biol. 2012, 14, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Rovida, E.; Peppicelli, S.; Bono, S.; Bianchini, F.; Tusa, I.; Cheloni, G.; et al. The metabolically modulated stem cell niche: a dynamic scenario that regulates cancer cell phenotype and resistance to therapy. Cell cycle 2014, 13, 3169–3175. [Google Scholar] [CrossRef] [PubMed]
- Amati, E.; Perbellini, O.; Rotta, G.; et al. High-throughput immunophenotypic characterization of bone marrow-derived mesenchymal stromal cells and umbilical cord blood reveals common and differentially expressed markers: identification of angiotensin-converting enzyme (CD143) as a differentially expressed marker between adult and perinatal tissue sources. Stem Cell Res Ther. 2018, 9, 10. [Google Scholar] [PubMed]
- Ke, X.; Chen, Y.; Wang, P.; Xing, J.; Chen, Z. Upregulation of CD147 protects hepatocellular carcinoma cells from apoptosis through glycolytic switch via HIF-1 and MCT-4 in hypoxia. Hepatol Int. 2014, 8, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, F.; Menashi, S.; Khayati, F.; Naïmi, B.; Porcher, R.; Podgorniak, MP.; et al. EMMPRIN promotes the malignant properties of melanoma cells through HIF-2alpha-mediated upregulation of the VEGF-2 receptor. PloS One 2010, 5, e12265. [Google Scholar] [CrossRef]
- Wang, C.H.; Yao, H.; Chen, L.N.; Jia, J.F.; Wang, L.; Dai, J.Y.; et al. CD147 induces angiogenesis through a vascular endothelial growth factor and a hypoxia-inducible transcription factor 1α-mediated pathway in rheumatoid arthritis. Arthritis Rheum. 2012, 64, 1818–1827. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, F. , Eougatef, F.; Quemener, C.; Kellouche, S.; Naimi, B.; Podgorniak, M.P.; Millot G.; Gabison, E.E.; Calvo, F.; Dosquet, C.; Lebbe, C.; et al. EMMPRIN promotes angiogenesis through hypoxia-inducible factor-2-mediated regulation of soluble VEGF isoforms and their VEGFR-2 receptor. Blood 2009, 114, 5547–5556. [Google Scholar]
- Lopes-Coelho, F.; Nunes, C.; Gouveia-Fernandes, S.; Rosas, R.; Silva, F.; Gameiro, P.; Carvalho, T.; Gomes da Silva, M.; Cabeçadas, J.; Dias, S.; Gonçalves, LG.; Serpa, J. Monocarboxylate transporter 1 (MCT1), a tool to stratify patients with acute myeloid leukemia (AML) and a vehicle to kill cancer cells. Oncotarget 2017, 8, 82803–82823. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, S.; Hata, K.; Hirose, K.; Okui, T.; Toyosawa, S.; Uzawa, N.; Nishimura, R.; Yoneda, T. The GPR81 lactate sensor regulates the glycolysis and tumor growth of breast cancer. Scientific reports, 2022; 12, 6261. [Google Scholar]
- Brown, TP.; Bhattacharjee, P.; Ramachandran, S.; et al. The lactate receptor GPR81 promotes breast cancer growth via a paracrine mechanism involving antigen-presenting cells in the tumor microenvironment. Oncogene 2020, 39, 3292–3304. [Google Scholar] [CrossRef]
- Soto, C. A.; Lesch, M.L.; Sharipol, A.; Khan, A.; Schafer, L. X.; Becker, M. W.; Munger, J. C.; Frisch B., J. Elevated Lactate in Acute Myeloid Leukemia Bone Marrow Microenvironment Dysfunction, with a Dual Role of GPR81 in Macrophage Polarization and Leukemia Cell Growth. BioRxiv 2023, 11, 566874. [Google Scholar]
- Ishikawa, F.; Yoshida, S.; Saito,Y; Hijikata A. ; Kitamura, H.; Tanaka, S.; et al. Chemotherapy-resistant human AML stem cells home to and engraft within the bone-marrow endosteal region. Nat Biotechnol. 2007, 25, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Nick van Gastel, N.; Spinelli, J.B.; Sharda, A.; Schajnovitz, A.; Baryawno, N.; Rhee, C.; Oki, T.; Grace, E.; Soled, H.J.; Milosevic, J.; Sykes, D.B.; Hsu, P.P.; Vander Heiden, M.G.; Vidoudez, C.; Trauger, S.A.; Haigis, M.C.; Scadden, D.T. Induction of a timed metabolic collapse to overcome cancer chemoresistance. Cell Metab. 2020, 32, 391–403. [Google Scholar] [CrossRef]
- Ng, K.P.; Manjeri, A.; Lee, K.L.; Huang, W.; Tan, S.Y.; Chuah, C.T.; Poellinger, L.; Ong, S.T. Physiologic hypoxia promotes maintenance of CML stem cells despite effective BCR-ABL1 inhibition. Blood 2014, 123, 3316–3326. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Jiang, Q.; Han, Y.; Peng, J.; Wang, C. shRNA-mediated EMMPRIN silencing inhibits human leukemic monocyte lymphoma U937 cell proliferation and increases chemosensitivity to adriamycin. Cell Biochem Biophys. 2015, 71, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.M.; Silva, C.; Wang, J.; Tong, J.P.; Yong, V.W. A novel anti-EMMPRIN function-blocking antibody reduces T cell proliferation and neurotoxicity: Relevance to multiple sclerosis. J Neuroinflamm. 2012, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Somno, A.; Anuchapreeda, S.; Chruewkamlow, N.M.; Pata, S.; Kasinrerk, W.; Chiampanichayakul, S. Involvement of CD147 on multidrug resistance through the regulation of P-glycoprotein expression in K562/ADR leukemic cell line. Leuk Res Rep. 2016, 6, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Simanovich, E.; Brod, V.; Rahat, M.M.; Drazdov, E.; Walter, M.; Shakya, J.; Rahat, M.A. Inhibition of tumor growth and metastasis by EMMPRIN multiple antigenic peptide (MAP) vaccination is mediated by immune modulation. Oncoimmunology 2016, 6, e1261778. [Google Scholar] [CrossRef]
- Wang, L.; Ku, X.M.; Li, Y.; Bian, H.J.; Zhang, S.H.; Ye, H.; Yao, X.Y.; Li, B.H.; Yang, X.M.; Liao, C.G.; Chen, Z.N. Regulation of matrix metalloproteinase production and tumor cell invasion by four monoclonal antibodies against different epitopes of HAb18G/CD147 extracellular domain. Hybridoma (Larchmt) 2006, 25, 60–67. [Google Scholar] [CrossRef]
- Chen, Z.N.; Mi, L.; Xu, J.; Song, F.; Zhang, Q.; Zhang, Z.; et al. Targeting radioimmunotherapy of hepatocellular carcinoma with iodine (131I) metuximab injection: clinical phase I/II trials. Int J Radiat Oncol Biol Phys. 2006, 65, 435–444. [Google Scholar] [CrossRef]
- Panich, T.; Tragoolpua, K.; Pata, S.; Tayapiwatana, C.; Intasai, N. Downregulation of extracellular matrix metalloproteinase inducer by scFv-M6-1B9 intrabody suppresses cervical cancer invasion through inhibition of urokinase-type plasminogen activator. Cancer Biother Radiopharm. 2017, 32, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Sun, Q.; Feng, F.; Huhe, M.; Mi, L.; Chen, Z. Preclinical pharmacokinetics, tolerability, and pharmacodynamics of metuzumab, a novel CD147 human-mouse chimeric and glycoengineered antibody. Mol Cancer Ther. 2015, 14, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.S.; Zhao, X.Y; Wei, D.; Miao, J.L; Liu, Z.K.; Yong, Y.L.; et al. CD147-specific chimeric antigen receptor T cells effectively inhibit T cell acute lymphoblastic leukemia. Cancer Lett. 2022, 542, 215762. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Dai, L.; Bratoeva, M.; Slomiany, M.G.; Toole, B.P.; Parsons, C. Cooperative roles for emmprin and LYVE-1 in the regulation of chemoresistance for primary effusion lymphoma. Leukemia 2011, 25, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Regulation of glycolytic metabolism by CD147-associated partners: a schematic overview to illustrate the regulation of glycolytic metabolism in Hematological malignancies.
Figure 1.
Regulation of glycolytic metabolism by CD147-associated partners: a schematic overview to illustrate the regulation of glycolytic metabolism in Hematological malignancies.
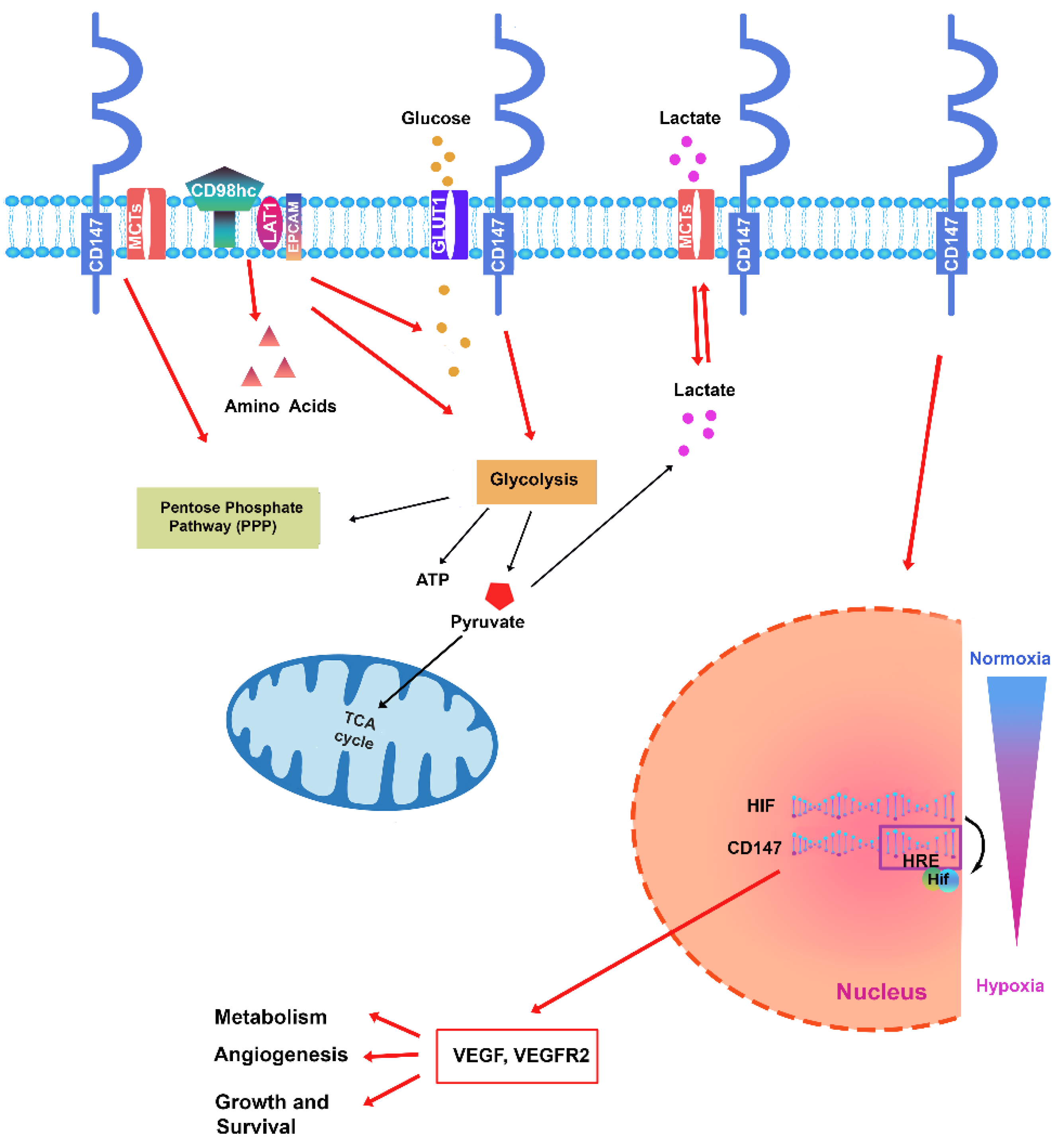

Table 1.
CD147-based therapies for Hematological Malignancies.
| Name |
Mechanism of Action |
Cell lines / tumor xenograft | Effects | References |
|---|---|---|---|---|
| siRNA | silencing | AML cell line | Inhibits leukemic cell proliferation, increases chemosensitivity to Adriamycin. | 132 |
| siRNA | silencing | Lymphoma Cell Line | increases chemosensitivity | 141 |
| AC-73 | Disrupts CD147 dimerization |
Cell lines and primary AML blasts |
Inhibits leukemic cell proliferation, increases chemosensitivity to ARA-C and ATO | 12 |
| PAB | CD147 inhibitor | Cell lines and primary AML blasts |
AML cell apoptosis | 54 |
| h4#147D | humanized anti-CD147 antibody | xenograft CML mouse models | strong antitumor effects with tumour regression | 99 |
| MEM-M6/6 | monoclonal CD147 antibodies (mAb) | CML cell line | Decreased expression of CD147 and caused downregulation of P-gp | 134 |
| CD147-CAR T cells | (CAR) T cell therapy | T-ALL cell line and T-ALL blasts | anti-tumor activity | 140 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



