
Submitted:
21 July 2024
Posted:
22 July 2024
You are already at the latest version
Abstract
Melanoma is the most aggressive and deadly form of skin cancer, and its incidence has been steadily increasing over the past few decades, particularly in the Caucasian population [1]. Current treatment strategies, the combination of BRAF and MEK inhibitors (BRAF/MEKi) in BRAFV600 mutant melanoma, and immune checkpoint inhibitors (ICI), have changed the clinical evolution of this disease [2]. However, a significant percentage of patients do not benefit from these therapies. Therefore, to improve patient selection, it is imperative to look for novel biomarkers. Immune subsets, particularly the quantification of lymphoid populations, could contribute to the identification of ICI responders. The aim of this review is to thoroughly examine significant published data on this topic in melanoma patients and to evaluate the potential role of lymphoid subsets as prognostic and predictive response biomarkers.
Keywords:
Melanoma
; Immune checkpoint inhibitors
; lymphoid subsets
; biomarker
; predictive biomarker
; prognostic biomarker
1. Introduction
Since the approval of ICI that target Cytotoxic T-Lymphocyte Antigen 4 (CTLA-4) and Programmed cell Death 1 (PD-1), the course of treatment and prognosis for metastatic melanoma (MM) have undergone significant progress. Both belong to the CD28 family and function as inhibitory receptors that impede the activation and proliferation of T lymphocytes. However, they intervene at different phases T cell activation process [3]. At first stage of T cells activation, through a process that occurs in lymph nodes, CTLA-4 expressed by naïve T cells acts by binding to the costimulatory protein B7 secreted by antigen-presenting cells (APCs), in order that the costimulatory signal CD28 cannot attach, thus preventing the activation of naive T cells. On the other hand, PD-1 receptor, expressed by an already activated circulating T cell or present in an inflamed tissue, interacts with its ligand PD-L1, expressed by other cell types such as a tumor cell. The interaction of the PD-1 receptor with its ligand, PD-L1, causes the deactivation of T cells and prevents an effective immune response at the peripheral level [3]. PD-1 is moderately expressed on naive T, B and NK cells, upregulated in response to T and B cell receptor activation, and induced on activated monocytes and myeloid cells.
Phase III clinical trials have shown that ipilimumab, a human IgG1 monoclonal antibody that blocks CTLA-4, significantly improves progression-free survival (PFS) and overall survival (OS) in patients with advanced melanoma when compared to gp100 vaccination [4] or dacarbazine (DTIC) [5]. Subsequently, pembrolizumab and nivolumab, both anti-PD-1, where approved, as first-line anti-PD-1 monoclonal antibodies for the treatment of advanced and/or MM. The approval of nivolumab was based on the results of pivotal trials in which anti-PD-1 monotherapy achieved an objective response rate (ORR) of approximately 35 to 42% and a 7-year OS of 37.8%, demonstrating superiority over chemotherapy (ChT) and anti-CTLA-4 as first-line treatment in patients with advanced melanoma [6]. Several months earlier, pembrolizumab was approved based on the results from an early phase trial, due to promising outcomes, with a 5-year survival of 34% in all patients and 41% in immune naive patients [7].
Later on, data from the pivotal studies of the combination of anti-PD-1 and anti-CTLA-4 were reported, which have been shown to act synergistically, achieving ORR superior to monotherapy, of 50-70%, with also greater OS at 5 years than monotherapy, around 59-68% [8,9] varying between studies, even in patients with central nervous system (CNS) involvement. Mechanisms underlying the greater efficacy of combination therapy may include enhanced priming and activation of helper and cytotoxic T cells and regulatory T cell clearance mediated by CTLA-4 blockade, together with restored cytotoxic T cell killing activity induced by both, PD-1 and CTLA-4 inhibition [9].
The effectiveness of ICI in treating melanoma is due to their highly immunogenic nature [10]. The major reason of the considerable immunogenicity is generally attributed to a substantial number of tumor mutations, mostly resulting from UV-induced alterations in the genetic material. This leads to the production of neoantigens, which are identified as foreign by the immune cells of the host causing their activation [11]. Even so, suboptimal activation of tumor antigen-specific T lymphocytes, resulting from low or absent expression of these antigens or molecules involved in their presentation [12], may inhibit T cell homing to the tumor [13]. In the event of optimal T cell activation, however, problems may also arise from the lack of appropriate chemo attractants in the tumor area or from their receptors on T lymphocytes. According to literature, various immune populations can either enhance or hinder an effective immune response [10,11,12,13]. For instance, studies on melanoma patients have shown that a high density of activated T cells surrounding the tumor, or an elevated count of B lymphocytes are associated with improved outcomes [14]. Furthermore, because of the immunogenic nature of melanoma, analyzing the immune infiltrate components might serve as biomarkers to predict the effectiveness of treatment in patients receiving (ICI, ChT, or BRAF/MEKi). This is in addition to its prognostic usefulness.
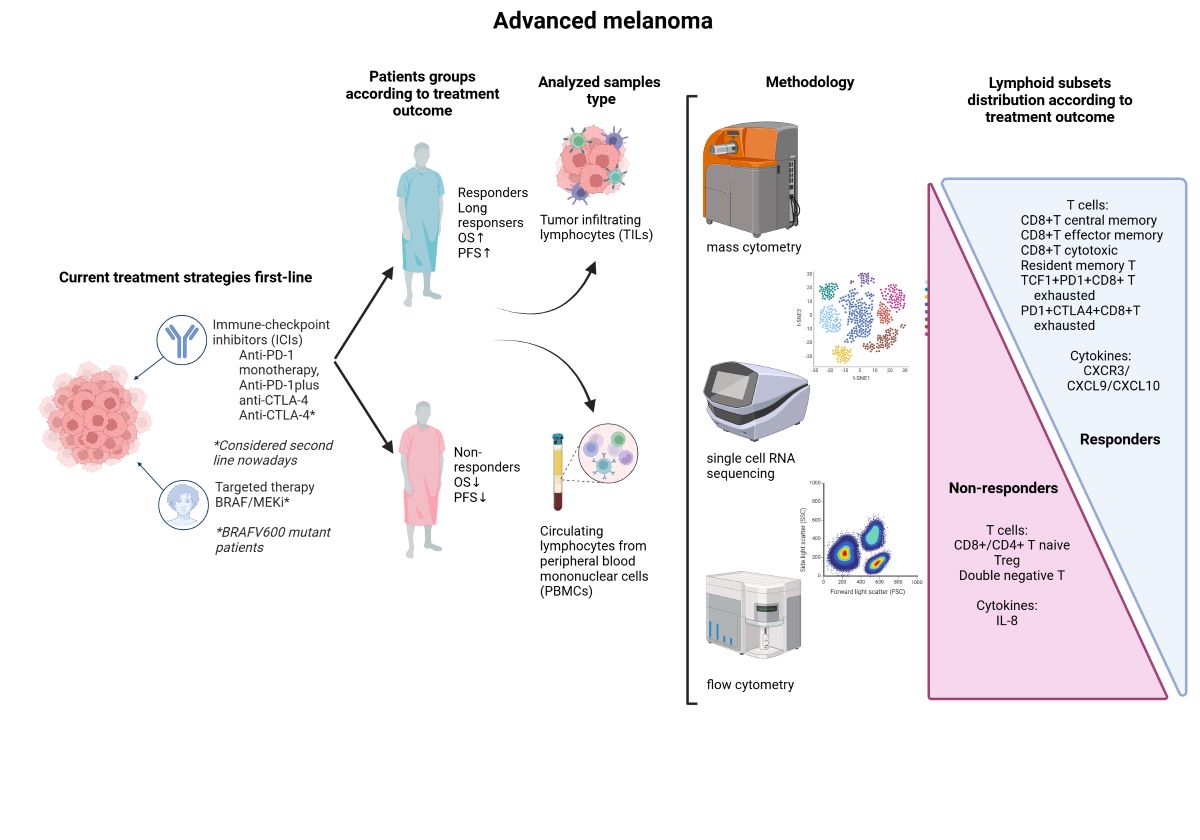
The present review focuses on preclinical and translational studies that explore the usefulness of identifying and monitoring several populations of peripheral lymphocyte subsets in melanoma patients undergoing treatment with ICI.
Particularly, we will focus on the T lymphocyte compartment within the total lymphoid lineage, as it is the most explored and has the most scientific evidence for its potential role as a prognostic and predictor of response to ICI including anti-CTLA-4, anti-PD-1 monotherapy and anti-PD-1 plus anti-CTLA-4 combination, in melanoma patients.
2. Lymphoid Populations and Outcomes of Melanoma Patients Treated with ICI
2.1. Lymphoid T Populations
Although the functional differences between T cell subpopulations are also reflected in gene expression, chromatin, metabolism, and ultimately at the functional level, in this review we will focus on T cell surface expressed antigens and intracellular proteins. Historically, multiparameter flow cytometry has been useful in identifying new subsets of T cells with a defined set of specialized markers and functions [15]. Nowadays, the development of innovative high-content technologies, such as single-cell RNA sequencing (scRNA-seq) or proteomics, is redefining the phenotypic classification and functional landscape of T cells [15]. Due to the diversity of the human immune system, there is currently a significant initiative called the Human Immunology Project. Its goal is to establish standardized protocols for all flow cytometry and also encompass more advanced methods like mass cytometry and already mentioned scRNA-seq [16]. T cells consist of significant subgroups, including CD4+ and CD8+. However, improved scientific knowledge has proven that certain cells within these subsets possess plasticity and can differentiate into various subpopulations.
In melanoma, some studies have reported that a prominent lymphocytic infiltrate is an independent parameter associated with a good prognosis [11], particularly when lymphocytes infiltrate the tumor in the vertical growth phase [17]. In particular, T lymphocyte infiltration, especially CD8+ and CD45RO+ subsets, has been shown to be a positive prognostic factor in several cancer types [18]. However, activated T cells in the tumor microenvironment are often functionally defective, with reduced proliferative capacity, cytokine production, or cytotoxicity [13]. Therefore, when assessing their role as biomarkers, it is important to characterize their functional state in addition to their quantitative determination in peripheral blood (PB) or tumor-infiltrating lymphocytes (TILs), for example by measuring ki67 expression.
2.1.1. Memory Compartment from Lymphoid T Cell Population
Naive (Tn), Central memory (Tcm), Effector memory cells (Tem), Terminally differentiated effector T cells (Temra) and Effector T cells (Teff)
Hematopoietic stem cells give rise to different types of blood cells, essentially, two main lineages: myeloid and lymphoid. T cells, B cells and NK cells arise from the lymphoid lineage (Figure 1). After maturation in the thymus, where they undergo somatic gene rearrangement resulting in the expression of a unique T cell receptor (TCR), T cells are fated to a particular CD8+ or CD4+ lineage. This occurs through a process called positive selection, which involves recognition of major histocompatibility complex class 1 (MHC1) or class 2 (MHC2) antigens corresponding to CD8+ or CD4+ T cell subsets, respectively [19] (Figure 1). In case of a self-antigen presentation, this process enables and ensures the elimination of autoreactive T cells [19]. Ultimately, remaining positively selected T cells enter the circulation, patrolling the blood and secondary lymphoid organs (SLOs), such as the spleen and lymph nodes, displaying a CD45RA+CCR7+ naïve T cell phenotype (Tn) [15,19]. Afterwards, during their journey, if they encounter APCs carrying their cognate antigen, the Tn cells bind to it via the TCR in the presence of a sufficient co-stimulatory signal. Once this co-stimulation process occurs, Tn cells are activated and differentiate into effector cells (Teff), which migrate to sites of inflammation and attack infected cells [15]. Once the pathogen or other threat, such as tumor cells, has been eliminated, 95% of the effector cells undergo apoptosis. Surviving T cells form a memory fraction that can recognize an antigen upon re-exposure, presenting a phenotype with loss of CD45RA and increase of CD45RO [15](Figure 2). We can identify CCR7+ central memory (Tcm), CCR7- effector memory (Tem), and CCR7- effector memory T cells that re-express CD45RA (Temra) [15]. In peripheral organs, Temra and Tem detect antigens, trigger a fast cytokine response, and release cytotoxic chemicals to kill antigen-expressing cells [15]. On the other hand, Tn and Tcm cells migrate through the SLOs, where they can be activated by mature dendritic cells, undergo expansion, and develop into Teff and Tem cells. These cells then circulate in the peripheral tissues through the bloodstream [15].
In the setting of advanced but curable melanoma, a report using multiparameter flow cytometric profiling showed that the proportion of naive CD45RA+CD4+ TILs in metastatic lymph nodes of 39 patients with stage III cutaneous melanoma were inversely correlated with the frequency of CD8+ T cells [20] and was significantly associated with reduced progression free survival (PFS). Related to this issue, another study, which analyzed PB T cell distribution in a cohort of 35 patients with locally advanced melanoma treated with ipilimumab in a neoadjuvant setting, showed an increased frequency of memory T cells expressing activating cytokines but no increase in Tn cells after 6 weeks of treatment (p<0.05) [21].
In the recurrent and advanced setting, similar observations have been made. For instance, the analysis of PB using flow cytometry from patients included in ipilimumab pivotal trials (CA184-004 and CA184-007), where effector memory and memory populations showed and increase after four weeks of treatment and Tn CD8+ and CD4+ declined [22].
Also, a study involving 77 patients with advanced melanoma treated with ipilimumab [23], an early increase in central and effector subsets was observed in PB samples from patients with disease control (DC). In addition, patients with DC had a slightly delayed but sustained increase in CD8+ Tem cells (p<0.05). The same study [23] assessed the dynamics between memory T cell subsets based on the expression of the Ki67 protein. Ipilimumab treatment resulted in a significant increase in CD4+ and CD8+ T cell proliferation (higher Ki67 expression) (p<0.05), involving the Tcm and Tem subsets, while remaining barely detectable in Tn.
Furthermore, in another study analyzing flow cytometry data with supervised clustering from peripheral blood mononuclear cells (PBMCs) in 90 advanced melanoma patients treated with anti-PD-1 and anti-CTLA-4 combination, an increase in circulating CD4+ Tem cells correlated positively with longer PFS [24].
In addition, high-dimensional single-cell mass cytometry was used for in-depth characterization of immune cell subsets in PB and matched tumor biopsies of 20 patients with MM before and after 12 weeks of anti-PD-1 treatment [25]. Patients classified as responders had higher numbers of CD4+ and CD8+ TILs, a significantly lower frequency of circulating CD4+ Tem cells, and a lower frequency of CD8+ Tn cells at baseline and after treatment initiation (p<0.05). In addition, the CD8+ T cell subpopulation of responders had a higher frequency of Tcm cells before and after the start of treatment than the CD8+ T cell subpopulation of non-responders [25].
Importantly, recent studies suggest that the level of CX3CR1, a chemokine receptor and a marker of T cell differentiation within memory subsets could behave as a predictor of response to ICI therapy. According to a study published in Nature [26], administering ICI to a tumor bearing mice increased the frequency and clonality of the T cell receptor belonging to the CX3CR1+CD8+ subset. The same study observed a higher frequency of the CX3CR1+ subset on circulating CD8+ T cells shortly after treatment initiation in patients with non-small-cell lung cancer (NSCLC) treated with anti-PD-1. ORR, PFS, and OS correlated positively with this frequency. Primarily, CX3CR1 is irreversibly expressed once T cells are fully differentiated; thus, in this study, its upregulation was maintained during treatment, but Ki67 expression on CD8+ T cells peaked at day 14 and returned to baseline by day 21 [26].
Moreover, Ribas and colleagues [27] reported an increase in memory T cells (CD8+CD45RO+) in tumoral samples from melanoma patients who presented a good outcome with pembrolizumab treatment. Also in another study, flow cytometry from PBMCs isolation from 31 melanoma patients treated with anti-PD-1 or anti-PD-1 plus anti-CTLA-4 combination revealed that responders showed an increase in the CD8+ Tem subset after three weeks of treatment [28].
Further evidence supporting that Tem and Tcm correlate with ICI response was observed in another study including 137 patients with advanced melanoma treated with ipilimumab. In this study those patients with 13% or higher percentage of Tcm CD8+ in PB were longer responders to treatment [29].
Finally, several studies have increased interest in the Tcm/Teff ratio as a predictor of ICI response. In this regard, in a study of patients with MM (n=43) and NSCLC (n=40) treated with nivolumab [30], tumors with increased inflammatory gene transcripts had increased ratios of CD4+ and CD8+ Tcm cells to Teff in the blood. In addition, the Tcm/Teff ratio correlated positively with PFS. The study [30] hypothesizes that a higher Tcm/Teff ratio does not reflect the immune response against the tumor, but rather the individual's ability to mount this response, which can be potentiated by ICI.
Resident memory T cells (Trm)
After migrating to different lymphoid and non-lymphoid tissues, some of the memory T cells generated from persistent activation against specific antigens eventually acquire tissue-resident properties characterized by up-regulation of CD69 and CD103, which are hallmarks of tissue-resident memory T cells (Trm). The Trm subpopulation persists long term in tissues without expanding into PB and acts as a sentinel in the event of antigen re-exposure [31] (Figure 2).
These cells have many intrinsic properties that imply they could regulate the growth of tumors: (i) they react far faster than circulating memory cells to re-exposure to cognate antigen; (ii) they express large amounts of cytotoxic molecules; and (iii) they are highly expressed in the tumor microenvironment. Trm are found in many human cancers, including melanoma, and they share all the features of memory CD8+ T cells (gated as CD45RO+CD62L-CD28-CD27-CCR7-) as well as an antitumor role in general [32]. They are the lymphocyte population with more immune checkpoint molecules expression [33] and preliminary data have shown that they may expand early during anti-PD-1 treatment.
A study examined the functional profiles of T cells within individual lymph metastases in 44 patients with stage III melanoma using single-cell mass cytometry. The outcomes were contrasted with circulating memory-phenotypic T cells, which were mainly CD8+ T cells [34]. The percentage of intratumoral T cells that expressed checkpoint point inhibitory proteins, PD-1 and TIM-3 was greater than that of circulating T cells. Nearly 60% of CD8+ T cells were found to be CD45RO+CD69+CCR7-, which is consistent with the Trm cell phenotype. Moreover, in the same study, a greater quantity of tumor-resident CD69+CD103+CD8+ T cells was linked to better melanoma-specific survival in stage III patients not receiving adjuvant ICI [35].
In line with these results, Wei et al. showed that T cell clones expanding in PB during anti-PD-1 treatment expressed high levels of CD69, PD-1, LAG-3 and CD45RO, corresponding to the phenotype of the CD8+ Trm population [36].
Interestingly, it is also worth mentioning that in a study including 103 resectable stage III melanoma patients, the presence of CD39+Trm in the surgical margin was associated with better relapse free survival (RFS)[37].
2.1.2. CD4+T Cells
For CD4+ T cells to identify an antigen, it must be delivered via MHCII, which is only expressed by specialized APCs such as dendritic cells (DCs), B cells, and macrophages [13]. When these APCs identify pathogens or other agents such as tumor cells that they recognize as danger signals, they become activated and migrate to the SLO. There, they are dedicated to display proteins and peptides derived from pathogens on their surface and presenting them to different types of cells, among them CD4+ T cells, which need to recognize MHCII, to cause their activation. At the time of activation of the CD4+ T cells, depending on which cytokines are present in the environment, a differentiation towards a specific subtype takes place, allowing a more efficient response according to the type of pathogen. In the presence of the cytokines interleukin (IL)-12 and interferon gamma (IFN-γ) they induce the expression of the transcription factor T-bet in T cells, resulting in T helper type 1 cells (Th1) producing IFN-γ which at the same time contributes to the death of the pathogen. In contrast, if IL-4 and IL-2 are present, IL-4 will induce Gata3 expression resulting in helper type 2 (Th2) cells that secrete IL-4, IL-5, and IL- 13, critical mediators of extracellular parasite clearance [13].
However, in recent years, the spectrum of CD4+ T cell subsets has been broadened, with the identification of T helper cells type 17 and 22 (Th17, Th22) and regulatory T cells (Treg). Th17 cells are generated by the expression of RORγt and secrete IL-17. IL-17, in turn, stimulates antimicrobial defense at the epithelial level and results in the recruitment and activation of neutrophils [13]. Th22 cells exert their effects on epithelial cells by releasing IL-22, which facilitates wound healing and provides tissue protection against injury [13]. A comprehensive examination of CD4+ T cell subsets has demonstrated distinct migratory capacities, as indicated by the presence of a specific group of chemokine receptors. The chemokine receptors CCR4, CCR6, CCR10, and CXCR3 have been identified as distinctive combinations for each CD4+ T cell subtype. These combinations are as follows: Th1 (CCR6-CCR4-CXCR3+), Th2 (CCR6-CCR4+CXCR3-), Th17 (CCR6+CCR4+CXCR3-CCR10-), and Th22 (CCR6+CCR4+CXCR3-CCR10+)[13].
Importantly, CXCR3 receptor interacts with CXCL9, CXCL10, and CXCL11 chemokines, which are released in the presence of IFN-γ and attract CXCR3-positive cells such as Th1 to sites of inflammation [13](Figure 3).
Conversely, Forkhead Box P3 (FoxP3)+ Treg cells do not aid in the defense against pathogens, but instead act to prevent autoimmune disorders by inhibiting undesired immune responses [13].
T helper (Th) subset
Although specific roles of different subsets of CD4+ T cells in various types of tumors are not well understood and require more investigation, it is widely acknowledged that Th1 cells enhance anti-tumor immune responses by producing significant quantities of IFN-γ. This not only promotes the function of cytotoxic CD8+ (Tc) T lymphocytes, but also attracts classically activated NK cells and M1 macrophages [38]. Nevertheless, within the framework of MM, there is a lack of definitive research that establishes the prognostic significance of evaluating CD4+ TIL by histopathology [12]. A positive association between higher densities of CD3+ and CD8+ TIL and OS has been observed, but not for CD4+ TIL [39]. In vivo murine models, CD4+ Th1 and Th2 cells have been shown to efficiently eliminate B16 melanomas [40].
One study found higher expression of Th1-associated genes, such as tumor necrosis factor alpha (TNF-α) and IL-2, in 20 primary melanoma tumors that spontaneously regressed, indicating activation of the host anti-tumor immune response, compared with non-regressing tumors [41]. In a phase II study [42], which included 75 patients with high-risk resected melanoma, HLA-A*0201 positive patients received a peptide vaccine in combination with ipilimumab. In this population, increased peripheral blood Th17 frequencies were correlated with RFS (p<0.05).
Additionally, a study evaluated whether PD-1 blockade influenced T helper polarization. To do this, whole blood and PBMCs from 9 patients with MM, 15 patients with prostate cancer and healthy controls were confronted with a superantigen (staphylococcus enterotoxin B) and re-exposure to the tetanus toxoid antigen to evaluate the reactivity of T cells [43]. Adding anti-PD-1 to the medium was observed to change the polarization of CD4+ T cells reacting to these antigens towards a more pro-inflammatory response of the Th1 and Th17 subtypes based on increased IFN-γ, IL-2, TNF-α, IL-6 and IL-17 production and the decrease in cytokines in the environment typically produced by Th2, IL-5 and IL-13.
T regulatory (Treg) subset
Treg are known for their potent immunosuppressive capabilities that regulates immunological balance [44]. Sakaguchi and his colleagues made the discovery that the transplantation of CD4+ T cells might effectively reverse autoimmune disorders in mice that had their thymus removed [45]. These findings suggested that the reduction of suppressive T cells might be a possible factor contributing to autoimmune diseases. These cells in question are often known as Treg and play a vital role in suppressing autoimmune reactions and preserving immunological balance. The predominant subset of Treg CD4+ cells has a high expression of the IL-2 receptor (CD25) alpha chain and the FoxP3 transcription factor. These factors are crucial for the production and maintenance of these cells, preventing any harmful effects [46]. Currently, the most extensively studied population of regulatory T cells is the CD4+CD25hiFoxP3+ Treg [47].
Current knowledge distinguishes three Treg subpopulations: naïve or resting Tregs (CD45RA+FoxP3loCD25lo), correspond to fraction 1 (Fr. I) of the total Tregs; effector Tregs (eTreg) (CD45RA-FOXP3hiCD25hi), fraction 2 (Fr. II) of the total set of Tregs, which come from naïve Treg cells after TCR stimulation and possess a high immunosuppressive capacity; and finally the so-called non-Tregs (CD45RA-FOXP3loCD25lo), fraction 3 (Fr. III) of the total number of Tregs, which do not possess immunosuppressive capacity but can produce pro-inflammatory cytokines. A significant number of eTreg cells, primarily belonging to the Fr. II subtype, are present in solid human malignancies.
While Treg cells account for just 2-5% of total CD4+ T cells in the peripheral blood of healthy individuals, they constitute 10-50% of CD4+ T cells in malignancies, with most of them being eTreg cells as already mentioned. In particular, the presence of Treg cells in the tumor microenvironment (TME) has been linked to decreased survival rates in melanoma patients [48]. Melanoma tumor cells secrete local immune-suppressing chemicals known as transforming growth factor beta (TGF-ß) and IL-10. They facilitate the proliferation of both endogenous Treg cells and the generation of novel induced Treg cells [49]. Additionally, the Teff/Treg ratio in the TME has consistently been linked to a favorable OS in melanoma patients [50].
Specifically, the presence of Treg cells in the melanoma has been linked to worse results in patients undergoing ICI treatment. According to findings from preclinical models, anti-CTLA-4 appears to specifically eliminate intratumoral Treg [51], which consistently expresses CTLA-4, while also activating Teff. The reduction of intratumoral Treg carried by anti-CTLA-4 is achieved by antibody-dependent cell-mediated cytotoxicity (ADCC) in the presence of macrophages expressing FCcRIIIA within the TME [52].
To provide more evidence that anti-CTLA-4 such as ipilimumab facilitates the depletion of Treg cells at the location of the tumor, metastatic biopsies obtained from 11 MM patients treated with ipilimumab showed a direct association between a high ratio of Teff to Treg and the occurrence of tumor necrosis [53]. Also, a comprehensive analysis of biomarkers indicating the likelihood of survival or response to ICI treatment in patients with advanced melanoma revealed a consistent negative correlation between Treg cells and response rates, PFS, and OS in most of the studies [54].
Moreover, in another research work [55], it was shown that lower levels of circulating Treg cells at the beginning of ipilimumab therapy in 31 MM patients were associated with improved RFS (p<0.05).
In addition, as most tumor antigens are typical self-antigens, melanoma has the potential to generate tumor-specific Treg cells, which can then inhibit the body's ability to mount an efficient anti-tumor response [56]. Comparative in vitro experiments demonstrated that Treg-mediated suppression is more prominent in T cells expressing self-antigenic tumor cells (Melan-A) than in those expressing neoantigens (cytomegalovirus), specifically targeting CD8+ T cells. This finding was supported by a study that reported a higher level of Treg-mediated suppression in self-antigen-specific CD8+ T cells compared to non-specific CD8+ T cells [57]. This might partially elucidate why tumors with a higher number of neoantigens are more vulnerable to anti-PD-1 therapy [58].
However, PD-1 inhibition can stimulate the growth of CD8+ T cells and the secretion of cytokines such as IFN-γ and IL-2. These cytokines decrease the amount of Treg cells by raising the ratio of Teff cells to Treg cells [58]. Simultaneously, IFN-γ can render Treg cells within tumors making them vulnerable, and causing them to lose their capacity to inhibit growth and produce additional IFN-γ, while still maintaining their Foxp3 expression and overall characteristics [59].
Administering anti-PD-1 in a mouse model of adenocarcinoma resulted in an increase in the quantity of IFN-γ+ Treg cells and a clinical response to anti-PD-1. This finding suggests that the treatment made these cells more susceptible to damage. When Treg cells lacked sensitivity to IFN-γ (using an Ifngr1L/LFoxp3Cre-YFP mouse), rats exhibited total resistance to PD-1 blocking, in contrast to a 40% response observed in WT mice [60]. This finding indicates that the fragility of Treg cells plays a role in determining their responsiveness to immunotherapy.
Experiments conducted on PBMCs derived from 14 individuals with advanced stage melanoma (stage III or IV) demonstrate that the inhibition of PD-1 with pembrolizumab can enhance the production of cytotoxic T lymphocytes that specifically target melanoma antigens and prevents Treg from limiting the function of these cytotoxic T lymphocytes [61].
In addition, a study using mice models found that Treg expressing PD-1 have a greater level of immunosuppressive activity compared to Treg without PD-1 [62]. Thus, PD-1-blocking antibodies have the potential to stimulate anti-tumor immunity and restore the functional activity of defective Teff cells by targeting Treg cells.
Finally, in a clinical study exploring this issue, an analysis of 32 individuals with MM treated either with nivolumab or pembrolizumab revealed a notable decrease in the presence of peripheral PD-1+ Treg cells (from 23% to 8.6%, p<0.05) following the initial cycle of immunotherapy. This reduction was observed specifically in responders to treatment [63] and correlated to PFS.
2.1.3. CD8+T Cells
CD8+ T cells, unlike CD4+ T cells, serve as cytotoxic T lymphocytes (Tc) and can directly eliminate tumoral and infected cells [31]. They possess the ability to identify antigens presented by MHCI, a molecule present on nearly all cells within the body. Subsequently, they can induce cell death by secreting chemicals that degrade cells, such as perforin and granzymes, or by initiating apoptosis via Fas.
Multiple investigations have demonstrated the essential role of CD8+ T cells in the immunological response to MM [64]. A decreased ratio Treg to Teff, as indicated before [50], has been linked to a higher ORR and OS in patients with MM who received ICI. Tumeh et al. [65] examined samples obtained from 46 individuals with advanced melanoma both before and after receiving pembrolizumab therapy. The researchers discovered that the quantity of CD8+ T lymphocytes located near the invasive margin and intratumoral was the most accurate predictor of response to treatment [65].
An alternative investigation examined PBMCs samples from 131 patients with MM who received treatment with either anti-PD-1 alone or in combination with anti-CTLA-4 [66]. On the 21st day, the patients who responded to treatment showed a higher number of big CD8+ T clones (those that make up more than 0.5%) in their PB compared to both healthy individuals and non-responsive patients. Flow cytometry analysis demonstrated a robust association between the abundance of big clones and the quantity of Tem CD8+ cells in PBMCs collected prior to and after treatment. In a separate group of MM patients who received ICI was studied within the same work, scRNA-seq was performed to investigate the extent to which transcriptional alterations occur in CD8+ T cells, based on their phenotypic subgroup and clonal size. The researchers discovered that the cytotoxic effector subset had the highest number of genes expressed differently and maintained a consistent clonal size following ICI treatment.
For instance, the activation of the IFN-γ pathway [67] demonstrates that the CD8+ T cell response to immune checkpoint inhibitors involves several components, such as cell division during mitosis and enhanced cytotoxicity.
Furthermore, a distinct investigation was carried out on a group of 31 individuals with advanced melanoma who had treatment with ipilimumab and stereotactic ablative radiation therapy (SBRT). The study revealed that patients who obtained therapeutic benefit, namely CR, PR, or SD ≥6 months, exhibited a higher prevalence of peripheral T cells CD8+. Furthermore, these individuals had an elevated CD8+/CD4+ cell ratio and a greater proportion of CD8+ cells expressing PD-1 [68], further described in section 2.1.4.
Eventually, it is crucial to consider the precise location of the metastases in the tissue samples that were examined in each research. To address this issue, research was conducted on patients with MM who were administered ipilimumab. The study examined immune subsets in lymph node tissue samples, as well as metastases located on the surface and beneath the skin [69]. The median densities of immune cells in lymph node metastases were consistently at twice as high as those in cutaneous or subcutaneous metastases for most of the indicators examined. Immunohistochemistry (IHC) analysis revealed that those who responded to treatment had a higher abundance of CD8+ T cells compared to those who did not react. This difference was observed specifically in lymph node metastases [69].
2.1.4. Dysfunctional Programs: From T Progenitor Exhausted to Terminally Exhausted and Reinvigoration
T cells go through different phases of maturation from Tn to Tcm, Tem and Teff, until reaching a final phase of exhaustion that takes place in the face of prolonged chronic activation and which are called exhausted T cells characterized by a deficient secretion of cytokines and expression of co-inhibitory signals such as PD-1 and TIM-3 [70]. These two markers that are immune checkpoints considered co-inhibitory signals make it possible to identify CD8+ T cells at opposite ends of the functional spectrum. It is considered that CD8+ TIM-3-PD-1- cells are highly functional with a great effector capacity of the immune response while CD8+ TIM-3+PD-1+ cells are highly dysfunctional, and each population has an own transcriptional profile [70]. Single-cell analysis of 25 immune infiltrates from melanoma tumors revealed a separation between bystander cytotoxic T cells and a population that shows a continuous progression from a transitional early effector to a dysfunctional T cell state [71]. This suggested differentiation pathway seems to be something that occurs to all CD8+ T cells in melanoma, according to several studies [71].
Within the exhausted CD8+ TILs, there are two distinct subpopulations: 'progenitor or stem cells' that preserve their ability to perform several functions and survive over a long period of time, and 'terminally exhausted cells' [72]. CD8+ Tex progenitor cells, show a middle level of PD-1 expression and are exposed to the CXCR5 chemokine receptor. Conversely, CD8+ Tex terminally exhausted have elevated expression of PD-1, TIM-3, and other co-inhibitory receptors [73].
There are notable functional distinctions between these two subpopulations. Researchers have discovered that the exhausted cells that expand after anti-PD-1 treatment display a progenitor phenotype [74]. In a study that analyzed CD8+ TILs from baseline tumor samples of 25 melanoma patients treated with nivolumab and ipilimumab, researchers identified TCF1+PD-1+CD8+ T cells consistent with the progenitor exhausted phenotype in almost all biopsies [74]. No significant differences in the frequency of TCF1+PD-1+CD8+ T cells were observed between responders and non-responders. However, in responders, on the other hand, the duration of the response was linked to the number of TCF1+PD-1+CD8+ T cells, and a higher number of these CD8+ T cells was significantly associated to longer PFS and OS (p<0.05) [74].
Another study of neoadjuvant anti-PD-1 in 20 locally advanced melanoma patients showed a rapid pathological response associated with the accumulation of exhausted CD8+ T cells in the tumor at 3 weeks and reinvigoration in the blood as early as 1 week, as determined by the expansion of Ki67+PD-1+ and Ki67+PD-1+CTLA-4+CD8+ T cells [75]. Most of the CD8+ T cells in the tumor were CD45RAloCD27hi, and expressed PD-1, TIM-3, CTLA-4, LAG-3, TIGIT, and CD39 [75]. Also, a lot of these CD8+ T cells had high Eomes levels but low T-bet levels, which is in line with the Tex phenotype [84] (further explained in section 2.1.5). In the same study, a high percentage of EomeshiT-betlo Tex was associated with clinical benefit, while Treg proliferation was associated with relapse and poor disease free-survival (DFS).
Consistently, in a systematic Medline search for biomarkers of survival or response to ICI in melanoma patients [54], PD-1+CD8+ T cells were the only biomarker significantly associated with OS in more than one study [54]. Consistent with these findings, in two studies of a cohort of 20 advanced MM patients each, samples from patients treated with pembrolizumab or nivolumab [76,77], PFS, and response were positively correlated with ≥20% higher proportions of CD8+ T cells in tumor biopsies characterized by high surface expression of CTLA-4 and PD-1 [76,77], regardless of the anatomical site of biopsy or prior therapy.
Further evidence was provided by another study [78] that used PBMCs of 29 MM patients before and after pembrolizumab treatment to identify pharmacodynamic changes in circulating exhausted CD8+ T cells. However, clinical failure in many patients was not due to a failure to induce immune reconstitution but to an imbalance between T cell reconstitution and tumor burden. This conclusion was reached by finding that a Ki67 to tumor burden ratio ≥1.94 at 6 weeks was associated with a better outcome based on ORR, PFS, and OS. An increase in Ki67 expression was observed in CD8+ T cells among melanoma patients (p<0.05) who received anti-PD-1 treatment, mostly in the PD-1+CD8+T subgroup (p<0.05), indicating the presence of an immune response before treatment [78]. The majority of the Ki67+CD8+T cell population from patients that responded had a phenotype CD45RAloCD27hi, as well as cells with elevated levels of CTLA-4 and PD-1. Furthermore, the upregulation of Ki67 expression was more prominent in PD-1+CD8+ cells compared to PD-1-CD8+ cells (p < 0.05). At baseline, about half of the PD-1+CTLA-4+ cells had Ki67 on them. After the therapy, that number rose to about 75%, which is in line with the recovery of Tex cells [78].
Also in a preclinical study with animal models, a study of CD8+ TIL from mice given PBS or an ICI control substance, revealed that, ICI-treated mice significantly increased the proportion of TILs displaying a reinvigorated Eomes+PD-1+Ki67+T-bet+ phenotype [79].
All these data show that in melanoma patients, blocking the inhibitory immune checkpoint triggers the regeneration of T cells within the tumor. This promotes the transformation of T cells into effector cells, which contribute to the anti-tumor response. Viral infections, as mentioned above, have described a hierarchy of CD8+ T cell dysfunction [77]. Partially exhausted CD8+ T cells have been defined as cells that can produce IFN-γ but lack the ability to produce TNF-α and IL-2, a phenotype consistent with tumor-infiltrating CTLA-4hiPD-1hiCD8+ T cells. In two cohorts of patients with advanced melanoma treated with ICI, there was a strong correlation between the proportions of PD-1hiCTLA-4hi cells within the tumor-infiltrating CD8+ T cell subset and the ORR and PFS [80]. The ORR was 0% (0 of 6) in the low CTLA-4hiPD-1hi T cell expression group, compared to 85.7% in the higher frequency CTLA-4hiPD-1hi T cell expression group [80].
Finally, researchers from this same work [80] sed a preclinical model to find out how many PD-1-CD8+ or PD-1+CD8+ TILs were present after each cycle of anti-TIM-3 and anti-PD-1 therapy [80]. After three cycles, the number of PD-1-CD8+ TILs increased, whereas the number of PD-1+CD8+ TILs did not change [80]. They hypothesize that PD-1-CD8+ TILs are in a less advanced state of differentiation than PD-1+CD8+ TILs. Therefore, they anticipated that PD-1-CD8+ TILs would have a greater capacity for revitalization and the ability to sustain long-term immunity. This study [80] classified T cells based on their memory state and the expression of three markers: CX3CR1, CD62L, and Slam7. They identified three subsets within the TILs PD-1-CD8+ group: the naive subset (CD62LhiSlamf7hiCX3CR1-PD-1-CD8+), the memory precursor phenotype subset (CD62L-Slamf7hiCX3CR1-PD1-CD8+), and the effector phenotype subgroup (CD62L-Slamf7hiCX3CR1+PD-1-CD8+). Interestingly, memory precursor phenotype and effector phenotype were the subpopulations that expanded more upon ICI treatment.
Therefore, as previously stated [26], CX3CR1 has been identified as an indicator of T cell differentiation and a prognostic factor for the effectiveness of ICI treatment. The data collected in this work [80] demonstrated that the transcription alterations caused by the inhibition of immune checkpoints corresponded with the development of memory and effector capabilities in CD8+ TILs. This phenomenon was observed to a greater extent in the PD-1-CD8+ subpopulation compared to the PD-1+CD8+ subpopulation.
Therefore, taking all these findings into account, T cells that are more prompt to experience reinvigoration driven by ICI treatment are the ones that are less differentiated; however, it can be a balance between the differentiation within exhaustion and memory states, which defines the phenotype that expands more frequently after ICI treatment.
2.1.5. Transcription Factors and Encoding Genes Regulators of Lymphoid T Cell Populations
Tcf7 and Tcf1/TCF1
Tcf7 is a gene that encodes for a transcription factor called Tcf1/TCF1. The primary function of this is to sustain the survival of CD8+ memory T cells and facilitate their transformation into effector T cells [81]. TCF1 is essential for the differentiation within the Th1/2/17/22 cell lineage. Additionally, it plays a crucial role in controlling the suppressive function of Treg on the immune system [82].
As previously mentioned, it has been shown that Tex-stem or progenitor TILs, which were developed following the anti-PD-1 breakthrough, had significant stem-like characteristics, including the capacity for self-renewal and the potential to transform into Tex–terminally exhausted TILs. TILs that express TIM-3, known as Tex-stem TILs, have lower levels of the co-inhibitory receptors PD-1 and LAG-3 compared to TILs that express both TIM-3 and PD-1 considered Tex-term. Significantly, Tex-stem TILs specifically correlate with the presence of TCF1 expression, whereas Tex-term TILs lack this expression [82].
In a preclinical research, a mouse model with a deletion of Tcf7 (E8i-Cre+Tcf7fl/fl) in mature CD8+ T cells was used [82]. The PD-1-CD8+ TIL exhibited a significant reduction in the number of memory precursor-like subsets in this animal. These findings indicate that Tcf7 is crucial for the formation and preservation of the memory precursor-like subgroup. Also, a study was conducted on 48 patients with advanced melanoma, 35 patients received anti-PD-1 monotherapy, 11 anti-PD-1 and anti-CTLA-4, and 2 anti-CTLA-4 monotherapy [83]. Among all patients, 31 responded to the treatment while 17 did not. Immune cells from tumor samples were chosen based on the CD45+ marker, using CellProfiler [84], an automated imaging system and scRNA-seq was performed. The scRNA-seq analysis revealed that responders had a >1 ratio of TCD8+TCF7+ cells to TCD8+TCF7- cells. Based on this discovery, the study determined that a higher proportion of progenitor cells compared to terminally exhausted cells is associated with a positive response to anti-PD-1-based therapy in patients with MM [83].
Siddiqui et al. conducted research where they discovered the presence of TCF1+PD1+CD8+ T cells within tumors in animal models following vaccination and checkpoint blockade treatment. These cells had characteristics like stem cells and played a crucial role in tumor control [85]. The previous understanding was that depletion impeded the formation of memory T cells. However, the identification of a specific group of virus-specific CD8+ T cells, which express both PD-1 and Tcf1, and are able to maintain the immune response during chronic infection, has revealed a link between T cell memory and exhaustion [85]. Therefore, Tcf1 is crucial in the development and functioning of CD8+ Tcm cells following the resolution of an acute infection [85].
Therefore, there is a link between exhaustion phase and differentiation memory phase, which defines the phenotype that expands more frequently after ICI treatment and TCF1 has a crucial role in determination in which differentiation state are found T cells within memory and exhaustion subsets.
Eomes and T-bet
Regulatory transcription factors, such as Tcf1 (encoded by Tcf7), already reviewed, Eomes, and T-bet, play a crucial role in governing the functional development of memory T cell subsets and their transformation into effector cells. Furthermore, T-bet and Eomes promote Th1 effector activity in cytotoxic T cells by increasing the expression of IFN-γ and cytotoxic granules, such as GzB. Afterwards, the absence of T-bet leads to impaired function of effector T cells [86].
T-bet and Eomes work together to sustain the antiviral CD8+ T cell population during long-lasting viral infection. T-bethi cells exhibit a low inherent rate of replacement, but they undergo cell division in the presence of persistent antigens, resulting in the production of Eomeshi terminal progenitor. Modified genetics elimination of either subset resulted in failure to control chronic infection, which suggests that an imbalance in differentiation and renewal could underlie the collapse of immunity in humans with chronic infections [80,81,87].
A preclinical investigation indicated that inhibition of the PD-1 pathway may result in T-bet collaborating with Eomes to bind to Pdcd1 gene or competing with it for binding. This competition leads to increased transcriptional repression. This interaction among T-box transcription factors is probable to happen in additional genes. Eomes can partially counteract the function of T-bet and presumably adjust the program of Tex by controlling its location inside the nucleus and potentially competing for DNA binding [88].
Elevated Eomes expression is necessary for the development of long-lasting memory cytotoxic T cells. If Eomes is absent, it leads to impaired maintenance of memory populations [88]. In addition, CTLA-4 decreases the transcriptional expression of Eomes, resulting in a reduced number of CD8+ T cells that generate IFN-γ and GzB [20]. Wei et al. [36] discovered a group of EomeshiT-bethiCD8+ T cells that were partly exhausted and showed considerable expansion when treated with anti-CTLA-4. In addition, Gide et al. demonstrated that in a group of 158 melanoma patients who received treatment with either anti-PD-1 alone or plus anti-CTLA-4, the presence of Eomes+CD69+CD45RO+ Tem cells in tumor samples was linked to a response and longer PFS (89).
Eomes ultimately facilitates the development of the terminal exhausted subpopulation [78], whereas T-bet and Tcf7 facilitate the development of the progenitor exhausted subpopulation [74]. As previously stated, following anti-PD-1 administration, the progenitor population undergoes expansion and originates from the subgroup of cells that are in a state of terminal exhaustion [74]. However, they must evolve to a terminal exhaustion state to promote a long-lasting response, and at these two points, T-bet and Eomes intervene.
Again, we can see the link between memory differentiation states from more precursor subsets to effector cells induced by Eomes expression that also facilitates the transition within exhausted population phases from progenitor to terminal exhausted cells.
2.1.6. Double Negative T Cells (DNTs)
Within T cells, defined by the expression of CD3+, apart from the two major groups, CD4+ and CD8+, a subset of cells is gaining interest that is characterized by the expression of TCR αβ or γδ but in the absence of surface T markers such as CD4, CD8, CD56. Based on this, these are called double negative T cells (DNTs) because they do not express any of the markers that classify T cells into the large groups [90]. They represent about 3% of total circulating T cells and have been shown to play an important role in preventing organ-versus-host disease [91] and in modulating the magnitude of the immune response [92]. In one study, including 68 patients with advanced melanoma treated with anti-CTLA-4 or anti-PD-1, the median level of circulating DNT decreased, while CD4+ and NK cells increased in patients who responded to treatment compared to those who did not [93].
So far, no further clinical data about DNTs potential predictive of response or prognostic role in melanoma patients treated with ICI have been published. According to these findings, DNTs could hinder immune response mediated by ICI but additional research is needed to confirm this assumption.
2.2. Chemokines and Outcomes of Melanoma Patients Treated with ICI
CXCR3
CXCR3 is a chemokine receptor that promotes the migration of Th1 cells and Teff CD8+ cells into inflamed tissues including tumors [94]. CXCR3 is mostly expressed in activated T cells and NK cells that release IFN-γ [94]. CXCL9 and CXCL10 act as ligands for CXCR3 and are generated in response to IFN-γ activation. Moreover, the IFN-γ-CXCL9/10-CXCR3 pathway serves a vital role in recruiting IFN-producing cells [94].
A study of 43 melanoma patients receiving pembrolizumab therapy found a significant increase in the percentage of CXCR3+ T cells in the blood following the first infusion [94]. However, the rate of expansion decreased following the second infusion in patients who responded well, whereas individuals with progressive disease continued to routinely show large percentages of these cells. The presence of CXCR3+ T cells in the circulation suggests a deficiency of IFN-γ-producing T cells stepping into tumors, leading to an insufficient response to anti-PD-1 treatment [94]. Consistently, individuals who did not respond exhibited a significant decrease in the levels of CXCL9/10. Furthermore, the study found that when anti-CXCR3 antibodies were used in preclinical experimental settings, there was a consistent acceleration of tumor growth in mice that had B16-F10 tumor cells [94]. In addition, the concurrent use of intramuscular CXCL9/10 and intraperitoneal anti-PD-1 administration successfully inhibited tumor growth. Therefore, this study hypothesized that the increase in CXCR3+ T cells in the blood of treatment-resistant patients might be caused by a decrease in the activation of CXCL9 and CXCL10 inside the tumor microenvironment.
Further in this setting, another study that performed flow cytomety analysis to investigate the presence of circulating or CD8+ TILs in 52 patients with MM found a direct relationship between the expression of CXCR3 in CD8+ TILs and prolonged OS [95]. Also, in another scientific work which analyzed plasma samples from 28 patients with advanced melanoma who were treated with pembrolizumab and nivolumab with ipilimumab, , responders (n=18) had a greater quantity of CXCL9 and CXCL10 compared to non-responders (n=10) [96].
Furthermore, separate research examined the immune profile and transcriptomics of 158 tumor samples obtained from advanced melanoma patients who received either anti-PD-1 monotherapy (n=63) or a combination of anti-PD-1 and anti-CTLA-4 (n=57) [97,98]. CXCL9 and CXCL10 were identified as two of the most prevalent and significantly increased chemokines following the administration of combined immune checkpoint blockade (p<0.005). Significantly, it was shown that the expression of CXCL9, CXCL10 was notably greater in tumors obtained from individuals who responded to treatment, both before and throughout therapy. Moreover, in these individuals, increased levels of CXCL9 and CXCL10 were linked to improved OS and a greater presence of CD8+ T cells within the tumor [97,98].
More recently published research has shown that the CXCL10/CXCR3 axis has a paracrine function in the anti-tumor activities of Th1 cells, Tc and NK cells. However, CXCL10/CXCR3 signaling promotes tumor cell proliferation, angiogenesis, and metastasis when mediated by an autocrine pathway in tumor cells. In general, the specific kinds of receptors and cells involved, appear to be the determining factors in the contradictory effects of CXCL10 on tumor growth. However, further research is needed to fully understand the underlying processes [99].
Preclinical evidence demonstrates that the injection of B16F10 tumor cells with decreased CXCR3 expression into C57BL/6 mice, greatly decreases the occurrence of metastasis in lymph nodes [100]. Thus, administering full Freund's adjuvant to the mice prior to therapy resulted in an increase in the levels of CXCL9 and CXCL10 in the draining lymph nodes. As a result, there was a significant 2.5–3.0-fold increase (p < 0.05) in the number of B16F10 cells that spread to lymph nodes, forming bigger clusters. Significantly, the growth of metastasis was greatly inhibited by lowering the expression of CXCR3 in B16F10 cells using reverse RNA or by dosing mice with antibodies against CXCL9 and CXCL10 [100].
Considering all published data to date, IFN-γ-CXCL9/10-CXCR3 pathway could have an antitumoral role when driven by paracrine signals and protumoral when driven by autocrine signals. This second occasion is caused by tumor cells.
IL-8
CXCL8, commonly referred to as IL-8, is a cytokine that exhibits several actions, including immunological effects like attracting neutrophils and promoting angiogenesis. The alpha and beta receptors, CXCR1 and CXCR2, are G-protein coupled receptors. Various cell types, such as monocytes, neutrophils, epithelial, fibroblast, endothelial, mesothelial, and cancer cells, release IL-8 when exposed to an inflammatory stimulation. It has a crucial function in the process of inflammation and the healing of wounds. Additionally, it can attract T cells and non-specific inflammatory cells to areas of inflammation by activating neutrophils. Patients diagnosed with various types of cancer have increased levels of IL-8, which seem to be associated with a more advanced tumor stage, grade, and disease severity [101].
Tumor-derived IL-8 has the capacity to have significant impacts on the tumor microenvironment [102]. It facilitates the maintenance of cancer cells in the epithelial-mesenchymal transition (EMT) state, enabling their mobility and invasiveness. Additionally, it promotes angiogenesis [103]. An instance of this is when cancer cells release IL-8, which can stimulate the growth and survival of cancer cells themselves through autocrine signaling pathways [102]. The attraction of myeloid-derived suppressor cells due to IL-8 secretion, in turn hinders the infiltration of T cells and weakens the expression of the IFN-γ. Quantifying the amounts of IL-8 in serum is a straightforward process that may be easily done using ordinary blood samples in regular clinical practice [101,104].
On this topic, a study was conducted with 1,344 patients who had advanced solid tumors treated with nivolumab and/or ipilimumab, everolimus, or docetaxel within clinical trials [101]. This study found that elevated levels of IL-8 in the bloodstream, specifically ≥23 pg/ml, were linked to poorer OS, independently of tumor type. Notably, the subgroup of patients with the strongest correlation between IL-8 levels and prognosis were MM patients treated nivolumab with ipilimumab within CheckMate 067 trial.
Moreover, Schalper et al. conducted an analysis of data from four clinical trials that included patients with melanoma among other solid tumors who were treated with ICI [105]. They found that greater baseline levels of IL-8 were consistently associated with poorer survival outcomes in patients who received anti-PD-1, anti-CTLA-4, or a combination of both treatments. Furthermore, patients with elevated IL-8 exhibited a decrease in ORR. The levels of IL-8 in circulation showed a favorable correlation with the expression of the CXCL8 gene inside the tumor and the number of neutrophils and monocytes in the peripheral blood.
To provide more evidence on this matter, we reference another study that examined the serum level of IL-8 in 29 MM patients who were treated with either nivolumab or pembrolizumab as a single therapy, or nivolumab in combination with ipilimumab. Levels were measured at the beginning of the treatment and again 2-4 weeks later [106]. Among patients who showed a response to treatment, the median levels of IL-8 in their blood decreased dramatically at the time when they had the best response, compared to the initial levels. Early changes in serum IL-8 levels were also strongly linked to responses to anti-PD-1 and anti-PD-1 plus anti-CTLA-4, but not baseline levels. Also, patients who were treated with anti-PD-1 and whose serum IL-8 levels dropped early had a significantly longer OS than patients whose levels rose early (p<0.05).
Finally, in a multitumor study, serum IL-8 levels correlated positively with tumor burden and negatively with OS and ORR in patients with various solid cancers, including MM patients (n=24) treated with BRAFi and ipilimumab [107].
However, all these studies were published several years ago, and the potential efficacy of anti-IL-8 is being evaluated within clinical trials in early development phases.
2.3. Distinct Effects of Anti-CTLA-4 and Anti-PD-1 on Lymphoid Populations
Immune checkpoint blockade with anti-CTLA-4, anti-PD-1 monotherapy or combined results in a distinct signature of gene expression changes in T cells in vivo as previously exemplified [36]. Both CD4+ and CD8+ T cells induced Ki67 in a flow cytometric analysis of pre- and post-therapy PBMC samples from 48 MM patients and mouse melanoma models treated with CTLA-4 and PD-1 blockade combination and monotherapy [108], with the Ki67+ cells exhibiting a CD45RO+ memory phenotype. However, combination therapy increased GzB, but neither anti-CTLA-4 nor anti-PD-1 monotherapy did. Anti-PD-1 alone led to the expansion of certain CD8+ T cell subsets that entered tumors and were worn out. In contrast, anti-CTLA-4 induces the expansion of an ICOS+T-bet+ Th1-like CD4+ effector population, as well as specific subsets of Tex CD8+ T cells in this study [108].
Another study, already mentioned, conducted by Wei et al. used a mass cytometry-based approach to analyze PBMCs from 48 patients treated with either ipilimumab monotherapy (n=13), pembrolizumab monotherapy (n=22), or ipilimumab+nivolumab combination therapy (n=13). Immune profiling analyzing T-cell responses revealed that terminally differentiated CD8+ effector T cells expanded only after the combination therapy as well as combination therapy increased Th1-like CD4 effector T cells greater than anti-CTLA-4 alone, but anti-PD-1 monotherapy failed to do so [36].
3. Discussion
Although the therapeutic landscape for advanced melanoma has radically changed in the last decade since the approval of ICI and BRAF/MEKi, a significant percentage of patients still do not benefit from these therapies [3,4,5,6,7,8,9,10,11,12,13]. The aim of this review to evaluate the potential role of lymphoid subsets as prognostic or predictive biomarkers for ICI treatment in melanoma patients by reviewing all meaningful published data on this topic. Some interesting and possibly useful results were found. It is still necessary to determine a cut-off value for the percentage distribution of lymphoid subsets in the PB, the appropriate timepoint for their determination (at baseline and/or longitudinally during treatment), and whether the PB distribution matches the TME distribution. .
First, considering the published data, we believe that memory T cells and exhausted T cells are crucial in determining the efficacy of ICI. Although, there are conflicting results regarding the role of CD4+ memory T cells, more solid evidence supports the association of higher frequency in PB and TILS of CD8+ Tcm and Tem cells with better response to anti-PD-1 therapy and longer duration of this, contributing to longer PFS and OS [21,22,23,24,25,26,27,28]. In comparison, higher levels of naive T cells (Tn) in both compartments, CD4+ and CD8+, generally correlate with poorer outcomes. We highlight the chemokine CX3CR1 as an important indicator of T cell differentiation within the memory subset, and its higher expression in CD8+ T cells may help predict a better response to ICI [26].
As a natural process following chronic activation of the immune response, there is a progressive maturation of T cells from Tn cells to Tcm and Tem to Teff cells and finally Tex, which show features of dysfunction such as impaired cytokine secretion and expression of markers of exhaustion such as PD-1 and TIM-3 [71,72,73]. Thus, analysing both markers expression enables to identify CD8+ T cells at opposite ends of exhaustion spectrum. It is considered that CD8+ TIM-3-PD-1- cells are highly functional with a great effector capacity of the immune response while CD8+ TIM-3+PD-1+ cells are highly dysfunctional [70].
Moreover, it is also relevant to mention the T exhaustion process from progenitor Tex cells to terminally exhausted Tex cells and the fact that anti-PD-1 based treatments have been shown to reinvigorate these cells, in particular CD8+ T cells expressing PD-1, making them differentiate, proliferate and become re-activated, being able to mediate an anti-tumor response in PB and intratumorally [75,76,77]. Higher presence of CTLA-4hiPD-1hiCD8+ T cells in peripheral blood or TILS has been correlated with good prognosis and ICI response in several studies [75,77,80].
Several transcription factors are involved in the regulation of this process. Eomes facilitates the development of terminally exhausted T cells, whereas T-bet and TCF7 are associated with progenitor exhausted T cells. Anti-PD-1 therapy primarily expands progenitor exhausted T cells [74], which are derived from cells in a state of terminal exhaustion. CD8+ T cells expressing TCF7 and its transcription factor Tcf1/TCF1 are more frequently expanded after ICI treatment in responders [82,83,85].
However, in a study [80] using a preclinical model, they observed that after three cycles of anti-TIM-3 and anti-PD-1 therapy, the number of PD-1-CD8+ TILs increased while the number of PD-1+CD8+ TILs did not change. They hypothesize that PD-1-CD8+ TILs are in a less advanced state of differentiation than PD-1+CD8+ TILs. In the same study, they identified three subsets within the PD-1-CD8+ group of TILs and found that memory precursor and effector phenotypes were the ones that expanded more after ICI treatment. One of the key markers to subclassify these subsets was CX3CR1.
Therefore, taking all these findings into account, T cells that are more responsive to ICI treatment are the ones that are less differentiated; however, it may be a balance between the exhaustion phase and the differentiation phase that defines the phenotype that expands more frequently after ICI treatment.
Also, CD8+ T cells, in general, but especially those with a cytotoxic phenotype, have been widely associated with a good prognosis in melanoma patients, and a high infiltration of CD8+ T cells in tumors has been considered a strong predictor of response to ICI [64,65,66,67]. Furthermore, the presence and expansion of large CD8+ T cell clones are critical for anti-PD-1 mediated responses. A higher CD8/CD4 ratio is also predictive of better outcomes with ICI treatment, presumably at the expense of the CD8+ cytotoxic subset, as these express IFN-γ, a key element for successful antitumor immune responses [68].
In contrast, the literature is inconclusive in clearly linking an increase in Th1, Th2, or Th17 cells after ICI treatment to improved responses. However, combined blockade with anti-PD-1 and anti-CTLA-4 may potentiate Th1 cells, which are characterized by their production of IFN-γ and have been more frequently associated with better responses to ICI than other T helper subtypes [36,108].
However, Treg, especially eTregs (with greater suppressive capacity), which have been extensively correlated with poor prognosis in several tumors including melanoma [55], have been shown to hinder response mediated by anti-PD-1 therapy and anti-CTLA-4 based therapies [51,52,53,54,55,56,57,58,59,60,61]. PD-1+ Tregs may be particularly affected by anti-PD-1 blockade, and their reduction correlates with better responses [63]. The reduction of intratumoral Treg by anti-CTLA-4 is achieved by ADCC in the presence of FCcRIIIA-expressing macrophages in the TME [52], whereas anti-PD-1 agents may have a more potent indirect mechanism by enhancing Teff, thereby increasing the Teff/Treg ratio. Thus, a high ratio of Tregs to conventional T cells has been associated with resistance to ICI.
Data on DNTs are limited for the scenario of this review, but evidence suggests that they may play a suppressive role in anti-tumor immunity. Further research is needed to understand their exact function and potential impact on immunotherapy outcomes [90,93].
Importantly, the representation of lymphocyte populations in peripheral blood may or may not reflect the TME, depending on the lymphocyte subset. Some subsets, such as Trm, may also be important for a successful immune response after ICI, but they are not represented in peripheral blood, which is less accessible for routine determination [35,36,37].
Finally, the measurement of some circulating cytokines may also be relevant. For instance, the IFN-γ-CXCL9/10-CXCR3 pathway can play both antitumor and protumor roles depending on the signaling context [94,95,96,97,98,99,100]. When driven by paracrine signals, this pathway can enhance the immune response against tumors by promoting the activity of Th1, Tc, and NK cells. Conversely, when driven by autocrine signals from tumor cells, this pathway can facilitate tumor growth and metastasis. Understanding these dual roles and analyzing which one is acting in each situation is crucial for carefully evaluating the potential use of CXCR3/CXCL9/CXCL10 plasma determination as a predictive biomarker for identifying ICI responders among MM patients.
Another protumoral cytokine that can be easily determined in peripheral blood is IL-8 [101,102,103,104,105,106,107]. Several data have shown its correlation with poor prognosis in a variety of solid tumors, including melanoma, and it could help identify non-responders to ICI [104,105,106,107]. Nevertheless, as mentioned above, most of the studies were published some years ago and, to our knowledge, clinical trials exploring the potential efficacy of anti-IL-8 are still in early development phases.

Table 1.
Lymphoid population subsets and findings in melanoma patients treated with ICI.
| Lymphoid cell population | Patients population | Observation | Ref* | ||
|---|---|---|---|---|---|
| T cell | Memory populations | Tn CD4+ | Stage III melanoma | ↑ in worse PFS | [20] |
| Locally advanced melanoma treated with neoadjuvant ipilimumab (IPI) | ↓ after treatment | [21] | |||
| Tn CD8+ | MM treated with Pembrolizumab (PEM) | ↓ in responders | [25] | ||
| Tcm CD8+ | ↑ in responders | ||||
| MM treated with IPI | ≥13% Tcm CD8+ in long responders | [29] | |||
| Tcm | MM treated with IPI | ↑ in patients (pts) with DC | [23] | ||
| Tem | |||||
| Tem CD8+ | ↑ in pts with DC | ||||
| MM treated with NIVO or PEM or NIVO+IPI | ↑ after 3 weeks in responders | [28] | |||
| Tem CD4+ | MM treated with nivolumab (NIVO) + IPI | ↑ in better PFS | [24] | ||
| MM treated with PEM | ↓ in responders | [25] | |||
| CX3CR1+CD8+ | Non-small cell lung cancer (NSCLC) treated with NIVO | ↑ better ORR, PFS and OS | [26] | ||
| Tcm/Teff | MM and NSCLC treated with NIVO | ↑ better PFS | [30] | ||
| Trm CD8+ | MC38-OVA and B16-OVA melanoma in murine model | ↓ tumor growth in conjuntion with anti-PD-1 | [32] | ||
| MM treated with anti-PD-1 | ↑ in responders | ||||
| MM treated with anti-PD-1 | ↑ better melanoma specific survival and ORR | [35] | |||
| Advanced resectable melanoma | ↑ better RFS | [37] | |||
| CD4+ | Th1 and Th2 | B16 melanoma in murine model | ↓ tumor growth | [40] | |
| Th1 | Spontaneously regressing tumours | ↑ Th1 genes expression | [41] | ||
| Th17 | MM treated with IPI +/- HLA-A*0201 | ↑ better RFS | [42] | ||
| Treg | MM treated with IPI | ↓ after treatment | [51] | ||
| MM treated with IPI | Lower Treg expression better RFS | [55] | |||
| PD-1+Treg | MM treated with PEM | Significant ↓ after 1rst cycle was predictor of PFS and Melanoma specific death | [63] | ||
| Teff/Treg | MM patients | ↑ associated with better OS | [50] | ||
| MM treated with IPI | ↑ was related to tumor necrosis | [53] | |||
| MM treated with PEM or NIVO | ↑ ratio | [61] | |||
| MM treated with combined anti-PD1+anti-CTLA-4 | ↑ within the tumor | [48] | |||
| CD8+ | Intratumoral CD8+ | MM treated with PEM | ↑ density at the invasive margin in responders | [65] | |
| CD8+ | MM treated with PEM or NIVO | ↑ PFS and ORR | [76] | ||
| CD8+Tc | MM treated with anti-PD-1 alone or with anti-CTLA-4 | ↑ORR | [66] | ||
| CD8+Teff | MM treated with IPI | ↑ associated with DC | [23] | ||
| DNTs | CD4-CD8-CD56- | MM treated with anti-PD-1 alone or anti-CTLA-4 | ↓ in responders | [90,110] | |
| Dysfunctional programs | TCF1+PD-1+CD8+ | MM treated with NIVO plus IPI | ↑ among responders and associated with longer response, OS and PFS | [74] | |
| Ki67+PD-1+CTLA-4+CD8+ | MMtreated with PEM | ↑ after treatment | [75] | ||
| CTLA-4+PD-1+CD8+ | MM treated with PEM or NIVO | ↑ associated with better ORR and PFS | [77] | ||
| Ki67+PD-1+CD8+ | MM treated with PEM | ↑ Ki67+PD-1+CD8+to tumor burden ratio associated with better OS, PFS and ORR | [78] | ||
| PD-1hiCTLA-4hiCD8+ | MM treated with ICI | ↑ better ORR, OS and PFS | [80] | ||
| Progenitor Tex/ Terminally exhausted Tex | MM treated with anti-PD-1 | ↑ ratio correlated to ORR | [74] | ||
| Transcription factors | TCF7+CD8+ | MM treated with anti-PD-1, anti-CTLA-4 or combined treatment | ↑ in responders | [83] | |
| TCF7+CD8+/TCF7-CD8+>1 | MM treated with anti-PD-1 | ↑ in responders | [83] | ||
| EomeshiT-BEThiCD8+ | MM treated with anti-CTLA-4 | ↑ after treatment | [36,88,89] | ||
| Eomes+CD69+CD45RO+Tem | MM treated with anti-PD-1, anti-CTLA-4 or combined treatment | ↑ better response and longer PFS | |||
| Ki67+Eomes+CD8+ | MM treated with IPI | ↓ baseline level associated with relapse | [88] | ||
| Cytokines | CXCR3+CD8+ | MM treated with PEM | Marked increase and decrease after second infusion in responders | [94] | |
| MM | ↑ better OS | [95] | |||
| CXCL9 and CXCL10 | MM treated with PEM or IPI+NIVO | ↑in responders | [96] | ||
| MM treated with anti-PD-1 alone or with anti-CTLA-4 | ↑intratumoral expression in responders and longer OS | [97] | |||
| IL-8 | MM patients treated with PEMBRO or NIVO alone or with IPI | ↓ levels in responders | [106] | ||
| Early ↓ associated with better OS | |||||
| MM treated with IPI or iBRAF | Positive correlation with tumor burden and negative with ORR and OS | [107] | |||
[*]Ref: Reference.
4. Conclusion
From an oncologist’s perspective, while significant progress has been made in the systemic treatment of advanced melanoma, ongoing research and clinical trials are essential to refine these strategies, overcome challenges, and improve patient outcomes. The integration of lymphocyte subsets profiling into clinical practice represents a promising avenue to personalize and enhance cancer immunotherapy. This is especially relevant when it comes to determining their presence in peripheral blood, since it is accessible and allows longitudinal determinations and may provide a more comprehensive understanding of the entire functioning of the immune system. Based on the existing research, Tcm, Tem, Tc, Tex from CD8+ T cells compartment, also PD-1, CTLA-4, CXC3R1 expression in CD8+ T cells among others, have shown potential as biomarkers for predicting the response to ICI. Nevertheless, the primary obstacle comes in integrating these biomarkers into everyday clinical practice, due to the diverse methodology employed in all studies, the majority of which are now limited to research applications.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J Clin. 2021 Jan;71(1):7-33. https://doi.org/10.3322/caac.21654. Epub 2021 Jan 12. Erratum in: CA Cancer J Clin. 2021 Jul;71(4):359. [CrossRef]
- Michielin O van Akkooi, A.C.J. Ascierto, P. A. Dummer, R., Keilholz, U. Cutaneous melanoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2019, 30, 1884–1901. [Google Scholar] [CrossRef]
- Teixidó, C.; González-Cao, M.; Karachaliou, N.; Rosell, R. Predictive factors for immunotherapy in melanoma. Ann Transl Med. 2015 Sep;3(15):208. [CrossRef]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved survival with ipilimumab in patients with metastatic melanoma, N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef]
- Robert, C.; Thomas, L.; Bondarenko, I.; O’Day, S.; Weber, J.; Garbe, C.; Lebbe, C.; Baurain, J.F.; Testori, A.; Grob, J.J.; et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma, N. Engl. J. Med. 2011, 364, 2517–2526. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Long, G.V.; Robert, C.; Brady, B.; Dutriaux, C.; Di Giacomo, A.M.; Mortier, L.; Hassel, J.C.; Rutkowski, P.; McNeil, C.; Kalinka-Warzocha, E.; Savage, K.J.; Hernberg, M.M.; Lebbé, C.; Charles, J.; Mihalcioiu, C.; Chiarion-Sileni, V.; Mauch, C.; Cognetti, F.; Ny, L.; Arance, A.; Svane, I.M.; Schadendorf, D.; Gogas, H.; Saci, A.; Jiang, J.; Rizzo, J.; Atkinson, V. Survival Outcomes in Patients With Previously Untreated BRAF Wild-Type Advanced Melanoma Treated With Nivolumab Therapy: Three-Year Follow-up of a Randomized Phase 3 Trial. JAMA Oncol. 2019 Feb 1;5(2):187-194. [CrossRef]
- Hamid, O.; Robert, C.; Daud, A.; Hodi, F.S.; Hwu, W.J.; Kefford, R.; Wolchok, J.D.; Hersey, P.; Joseph, R.; Weber, J.S. , et al. Five-year survival outcomes for patients with advanced melanoma treated with pembrolizumab in KEYNOTE-001. Ann. Oncol. 2019, 30, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Rutkowski, P.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Wagstaff, J.; Schadendorf, D.; Ferrucci, P.F.; et al. Overall Survival with Combined Nivolumab Ipilimumab in Advanced Melanoma, N. Engl. J. Med. 2017, 377, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; Ferrucci, P.F.; Smylie, M.; Butler, M.O.; Hill, A.; Márquez-Rodas, I.; Haanen, J.B.A.G.; Guidoboni, M.; Maio, M.; Schöffski, P.; Carlino, M.S.; Lebbé, C.; McArthur, G.; Ascierto, P.A.; Daniels, G.A.; Long, G.V.; Bas, T.; Ritchings, C.; Larkin, J.; Hodi, F.S. Long-Term Outcomes With Nivolumab Plus Ipilimumab or Nivolumab Alone Versus Ipilimumab in Patients With Advanced Melanoma. J Clin Oncol. 2022, Jan 10;40(2):127-13m7. [CrossRef]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in cancer immunotherapy. Science. 2015, 348:69–74 154. [CrossRef]
- Azimi, F.; Scolyer, R.A.; Rumcheva, P.; Moncrieff, M.; Murali, R.; McCarthy, S.W.; Saw, R.P.; Thompson, J.F. Tumor-infiltrating lymphocyte grade is an independent predictor of sentinel lymph node status and survival in patients with cutaneous melanoma. J Clin Oncol. 2012, 30:2678–2683. [CrossRef]
- Ladányi, A. Prognostic and predictive significance of immune cells infiltrating cutaneous melanoma. Pigment Cell Melanoma Res. 2015, Sep,28(5):490-500. [CrossRef]
- Anichini, A.; Vegetti, C.; Mortarini, R. The paradox of T cell–mediated antitumor immunity in spite of poor clinical outcome in human melanoma. Cancer Immunol. Immunother. 2004, 53, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Bogunovic, D.; O’Neill, D.W.; Belitskaya-Levy, I. Immune profile and mitotic index of metastatic melanoma lesions enhance clinical staging in predicting patient survival. Proc. Natl Acad. Sci. 2009, 106, 20429–20434. [Google Scholar] [CrossRef] [PubMed]
- Brummelman, J.; Pilipow, K.; Lugli, E. The Single-Cell Phenotypic Identity of Human CD8+ and CD4+ T Cells. Int Rev Cell Mol Biol. 2018, 341:63-124. [CrossRef]
- Maecker, H.T.; McCoy, J.P.; Nussenblatt, R. Standardizing immunophenotyping for the Human Immunology Project. Nat Rev Immunol. 2012, Feb 17,12(3):191-200. [CrossRef]
- Clark, W.H., Jr.; Elder, D.E.; Guerry, D.I.V.; Braitman, L.E.; Trock, B.J.; Schultz, D.; Synnestvedt, M.; Halpern, A.C. Model predicting survival in stage I melanoma based on tumor progression. J. Natl Cancer Inst. 1989, 81, 1893–1904. [Google Scholar] [CrossRef]
- Senovilla, L.; Vacchelli, E.; Galon, J.; et al. Prognostic and predictive value of the immune infiltrate in cancer. OncoImmunology. 2012, 1, 1323–1343. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, I. Views on helper/cytotoxic lineage choice from a bottom-up approach. Immunol Rev. 2016, 271:98–113. [CrossRef]
- Jacquelot, N.; Roberti, M.P.; Enot, D.P.; Rusakiewicz, S.; Semeraro, M.; Jégou, S.; et al. Immunophenotyping of stage III melanoma reveals parameters associated with patient prognosis. J Invest Dermatol. 2016, 136, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Tarhini, A.A.; Edington, H.; Butterfield, L.H.; Lin, Y.; Shuai, Y.; Tawbi, H.; Sander, C.; Yin, Y.; Holtzman, M.; Johnson, J.; Rao, U.N.; Kirkwood, J.M. Immune monitoring of the circulation and the tumor microenvironment in patients with regionally advanced melanoma receiving neoadjuvant ipilimumab. PLoS One. 2014, Feb 3,9(2):e87705. [CrossRef]
- Weber, J.S.; Hamid, O.; Chasalow, S.D.; Wu, D.Y.; Parker, S.M.; Galbraith, S.; Gnjatic, S.; Berman, D. Ipilimumab increases activated T cells and enhances humoral immunity in patients with advanced melanoma. J Immunother. 2012 Jan;35(1):89-97. [CrossRef]
- Felix, J.; Lambert, J.; Roelens, M.; Maubec, E.; Guermouche, H.; Pages, C.; Sidina, I.; Cordeiro, D.J.; Maki, G.; Chasset, F.; Porcher, R.; Bagot, M.; Caignard, A.; Toubert, A.; Lebbé, C.; Moins-Teisserenc, H. Ipilimumab reshapes T cell memory subsets in melanoma patients with clinical response. Oncoimmunology. 2016, Feb 18,5(7):1136045. [CrossRef]
- Kverneland, A.H.; Thorsen, S.U.; Granhøj, J.S.; Hansen, F.S.; Konge, M.; Ellebæk, E.; Donia, M.; Svane, I.M. Supervised clustering of peripheral immune cells associated with clinical response to checkpoint inhibitor therapy in patients with advanced melanoma. Immunooncol Technol. 2023 Aug 24;20:100396. [CrossRef]
- Krieg, C.; Nowicka, M.; Guglietta, S.; Schindler, S.; Hartmann, F.J.; Weber, L.M.; Dummer, R.; Robinson, M.D.; Levesque, M.P.; Becher, B. High-dimensional single-cell analysis predicts response to anti-PD-1 immunotherapy. Nat Med. 2018 Feb;24(2):144-153. Epub 2018 Jan 8. Erratum in: Nat Med. 2018 Nov;24(11):1773-1775. [CrossRef]
- Yamauchi, T.; Hoki, T.; Oba, T.; Jain, V.; Chen, H.; Attwood, K.; Battaglia, S.; George, S.; Chatta, G.; Puzanov, I.; Morrison, C.; Odunsi, K.; Segal, B.H.; Dy, G.K.; Ernstoff, M.S.; Ito, F. T cell CX3CR1 expression as a dynamic blood-based biomarker of response to immune checkpoint inhibitors. Nat Commun. 2021, Mar 3,12(1):1402. [CrossRef]
- Ribas, A.; Shin, D.S.; Zaretsky, J.; Frederiksen, J.; Cornish, A.; Avramis, E.; Seja, E.; Kivork, C.; Siebert, J.; Kaplan-Lefko, P.; Wang, X.; Chmielowski, B.; Glaspy, J.A.; Tumeh, P.C.; Chodon, T.; Pe'er, D.; Comin-Anduix, B. PD-1 Blockade Expands Intratumoral Memory T Cells. Cancer Immunol Res. 2016 Mar;4(3):194-203. [CrossRef]
- Valpione, S.; Galvani, E.; Tweedy, J.; Mundra, P.A.; Banyard, A.; Middlehurst, P.; Barry, J.; Mills, S.; Salih, Z.; Weightman, J.; Gupta, A.; Gremel, G.; Baenke, F.; Dhomen, N.; Lorigan, P.C.; Marais, R. Immune-awakening revealed by peripheral T cell dynamics after one cycle of immunotherapy. Nat Cancer. 2020 Feb;1(2):210-221. [CrossRef]
- Wistuba-Hamprecht, K.; Martens, A.; Heubach, F.; Romano, E.; Geukes Foppen, M.; Yuan, J.; Postow, M.; Wong, P.; Mallardo, D.; Schilling, B.; Di Giacomo, A.M.; Khammari, A.; Dreno, B.; Maio, M.; Schadendorf, D.; Ascierto, P.A.; Wolchok, J.D.; Blank, C.U.; Garbe, C.; Pawelec, G.; Weide, B. Peripheral CD8 effector-memory type 1 T cells correlate with outcome in ipilimumab-treated stage IV melanoma patients. Eur J Cancer. 2017 Mar;73:61-70. [CrossRef]
- Manjarrez-Orduño, N.; Menard, L.C.; Kansal, S.; Fischer, P.; Kakrecha, B.; Jiang, C.; Cunningham, M.; Greenawalt, D.; Patel, V.; Yang, M.; Golhar, R.; Carman, J.A.; Lezhnin, S.; Dai, H.; Kayne, P.S.; Suchard, S.J.; Bernstein, S.H.; Nadler, S.G. Circulating T Cell Subpopulations Correlate With Immune Responses at the Tumor Site and Clinical Response to PD1 Inhibition in Non-Small Cell Lung Cancer. Front Immunol. 2018, Aug 3,9:1613. [CrossRef]
- Sabat, R.; Wolk, K.; Loyal, L.; Döcke, W.D.; Ghoreschi, K. T cell pathology in skin inflammation. Semin Immunopathol. 2019 May,41(3):359-377. [CrossRef]
- Blanc, C.; Hans, S.; Tran, T.; Granier, C.; Saldman, A.; Anson, M.; Oudard, S.; Tartour, E. Targeting Resident Memory T Cells for Cancer Immunotherapy. Front Immunol. 2018, Jul 27,9:1722. [CrossRef]
- Boddupalli, C.S.; Bar, N.; Kadaveru, K.; Krauthammer, M.; Pornputtapong, N.; Mai, Z.; Ariyan, S.; Narayan, D.; Kluger, H.; Deng, Y.; Verma, R.; Das, R.; Bacchiocchi, A.; Halaban, R.; Sznol, M.; Dhodapkar, M.V.; Dhodapkar, K.M. Interlesional diversity of T cell receptors in melanoma with immune checkpoints enriched in tissue-resident memory T cells. JCI Insight. 2016 Dec 22;1(21):e88955. [CrossRef]
- Enamorado, M.; Iborra, S.; Priego, E.; Cueto, F.J.; Quintana, J.A.; Martinez-Cano, S.; et al. Enhanced anti-tumor immunity requires the interplay between resident and circulating memory CD8(+) T cells. Nat Commun (2017) 8:16073. [CrossRef]
- Edwards, J.; Wilmott, J.S.; Madore, J.; Gide, T.N.; Quek, C.; Tasker, A.; Ferguson, A.; Chen, J.; Hewavisenti, R.; Hersey, P.; Gebhardt, T.; Weninger, W.; Britton, W.J.; Saw, R.P.M.; Thompson, J.F.; Menzies, A.M.; Long, G.V.; Scolyer, R.A.; Palendira, U. CD103+ Tumor-Resident CD8+ T Cells Are Associated with Improved Survival in Immunotherapy-Naïve Melanoma Patients and Expand Significantly During Anti-PD-1 Treatment. Clin Cancer Res. 2018, Jul 1,24(13):3036-3045. [CrossRef]
- Wei, S.C.; Levine, J.H.; Cogdill, A.P.; Zhao, Y.; Anang, N.A.S.; Andrews, M.C., et al. Distinct cellular mechanisms underlie anti-CTLA-4 and anti-PD-1 checkpoint blockade. Cell. 2017, 170(6):1120–33.e17. [CrossRef]
- Attrill, G.H.; Owen, C.N.; Ahmed, T.; Vergara, I.A.; Colebatch, A.J.; Conway, J.W.; Nahar, K.J.; Thompson, J.F.; Pires da Silva, I.; Carlino, M.S.; Menzies, A.M.; Lo, S.; Palendira, U.; Scolyer, R.A.; Long, G.V.; Wilmott, J.S. Higher proportions of CD39+ tumor-resident cytotoxic T cells predict recurrence-free survival in patients with stage III melanoma treated with adjuvant immunotherapy. J Immunother Cancer. 2022 Jun;10(6):e004771. [CrossRef]
- Kim, H.-J.; Cantor, H. CD4 T cell subsets and tumor immunity: the helpful and the not-so-helpful. Cancer Immunol Res. 2014, 2:91–8. [CrossRef]
- Erdag, G.; Schaefer, J.T.; Smolkin, M.E.; Deacon, D.H.; Shea, S.M.; Dengel, L.T. Immunotype and immunohistologic characteristics of tumor infiltrating immune cells are associated with clinical outcome in metastatic melanoma. Cancer Res. 2012, 72:1070–80. [CrossRef]
- Eftimie, R.; Hamam, H. Modelling and investigation of the CD4+ T cells – macrophages paradox in melanoma immunotherapies. J Theor Biol. 2017, 420:82–104. [CrossRef]
- Lowes, M.A.; Bishop, G.A.; Crotty, K.; Barnetson, R.S.C.; Halliday, G.M. T helper 1 cytokine mRNA is increased in spontaneously regressing primary melanomas. J Invest Dermatol. 1997, 108:914–9. [CrossRef]
- Sarnaik, A.; Yu, B.; Yu, D.; Morelli, D.; Hall, M.; Bogle, D.; Yan, L.; Targan, S.; Solomon, J.; Nichol, G.; Yellin, M.; Weber, J.S. Extended dose ipilimumab with a peptide vaccine: immune correlates associated with clinical benefit in patients with resected high-risk stage IIIc/IV melanoma. Clin Cancer Res. 2011, Feb 15,17(4):896-906. [CrossRef]
- Dulos, J.; Carven, G.J.; van Boxtel, S.J.; Evers, S.; Driessen-Engels, L.J.; Hobo, W.; Gorecka, M.A.; de Haan, A.F.; Mulders, P.; Punt, C.J.; Jacobs, J.F.; Schalken, J.A.; Oosterwijk, E.; van Eenennaam, H.; Boots, A.M. PD-1 blockade augments Th1 and Th17 and suppresses Th2 responses in peripheral blood from patients with prostate and advanced melanoma cancer. J Immunother. 2012, Feb-Mar;35(2):169-78. [CrossRef]
- Josefowicz, S.Z.; Lu, L.F.; Rudensky, A.Y. Regulatory T cells: mechanisms of differentiation and function. Annual Rev Immunol. 2012, 30: 531–564. [CrossRef]
- Sakaguchi, S.; Takahashi, T.; Nishizuka, Y. Study on cellular events in postthymectomy autoimmune oophoritis in mice. I. Requirement of Lyt-1 effector cells for oocytes damage after adoptive transfer. J Exp Med 1982, 156, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Strauss, L.; Bergmann, C.; Szczepanski, M.; Gooding, W.; Johnson, J.T.; Whiteside, T.L. A unique subset of CD4+CD25highFoxp3+ T cells secreting interleukin-10 and transforming growth factor-beta1 mediates suppression in the tumor microenvironment. Clin Cancer Res. 2007, 13(15):4345–4354. [CrossRef]
- Callahan, M.K.; Postow, M.A.; Wolchok, J.D. CTLA-4 and PD-1 pathway blockade: combinations in the clinic. Front Oncol. 2014, 4: 385. [CrossRef]
- Tanaka, A.; Sakaguchi, S. Targeting Treg cells in cancer immunotherapy. Eur J Immunol. 2019 Aug;49(8):1140-1146. [CrossRef]
- Nizar, S.; Meyer, B.; Galustian, C.; Kumar, D.; Dalgleish, A. T regulatory cells, the evolution of targeted immunotherapy. Biochim Biophys Acta. 2010, 1806:7–17. [CrossRef]
- Quezada, S.A.; Peggs, K.S.; Curran, M.A.; Allison, J.P. CTLA4 blockade and GM-CSF combination immunotherapy alters the intratumor balance of effector and regulatory T cells. J Clin Invest. 2006, 116:1935–1945. [CrossRef]
- Selby, M.J.; Engelhardt, J.J.; Quigley, M.; Henning, K.A.; Chen, T.; Srinivasan, M.; Korman, A.J. Anti-CTLA-4 antibodies of IgG2a isotype enhance antitumor activity through reduction of intratumoral regulatory T cells. Cancer Immunol Res. 2013, Jul,1(1):32-42. [CrossRef]
- Simpson, T.R.; Li, F.; Montalvo-Ortiz, W., et al. Fc dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma. J Exp Med. 2013, 210: 1695–1710. [CrossRef]
- Hodi, F.S.; Butler, M.; Oble, D.A.; Seiden, M.V.; Haluska, F.G.; Kruse, A., et al. Immunologic and clinical effects of antibody blockade of cytotoxic T lymphocyte-associated antigen 4 in previously vaccinated cancer patients. Proc Natl Acad Sci USA. 2008,105:3005–10. [CrossRef]
- Ouwerkerk, W.; van den Berg, M.; van der Niet, S.; Limpens, J.; Luiten, R.M. Biomarkers, measured during therapy, for response of melanoma patients to immune checkpoint inhibitors: a systematic review. Melanoma Res. 2019, Oct, 29(5):453-464. [CrossRef]
- Retseck, J.; Nasr, A.; Lin, Y.; Lin, H.; Mendiratta, P.; Butterfield, L.H.; Tarhini, A.A. Long term impact of CTLA4 blockade immunotherapy on regulatory and effector immune responses in patients with melanoma. J Transl Med. 2018, Jul 4,16(1):184. [CrossRef]
- von Herrath, M.G.; Harrison, L.C. Antigen-induced regulatory T cells in autoimmunity. Nat Rev Immunol. 2003, 3:223–232. [CrossRef]
- Takeuchi, Y.; Nishikawa, H. Roles of regulatory T cells in cancer immunity. Int Immunol. 2016, Aug,28(8):401-9. [CrossRef]
- Charoentong, P.; Finotello, F.; Angelova, M.; Mayer, C.; Efremova, M.; Rieder, D.; Hackl, H.; Trajanoski, Z. Pan-cancer Immunogenomic Analyses Reveal Genotype-Immunophenotype Relationships and Predictors of Response to Checkpoint Blockade. Cell Rep. 2017, Jan 3,18(1):248-262. [CrossRef]
- Overacre-Delgoffe, A.E.; Vignali, D.A.A. Treg Fragility: A Prerequisite for Effective Antitumor Immunity? Cancer Immunol Res. 2018, Aug,6(8):882-887. [CrossRef]
- Overacre-Delgoffe, A.E.; Chikina, M.; Dadey, R.E.; Yano, H.; Brunazzi, E.A.; Shayan, G., et al. Interferon-g drives Treg fragility to promote anti-tumor immunity. Cell 2017,169:1130–41e11. [CrossRef]
- Wang, W.; Lau, R.; Yu, D.; Zhu, W.; Korman, A.; Weber, J. PD1 blockade reverses the suppression of melanoma antigen-specific CTL by CD4+ CD25(Hi) regulatory T cells. Int Immunol 2009, 21: 1065–1077. [CrossRef]
- Park, H.J.; Park, J.S.; Jeong, Y.H.; et al. PD-1 upregulated on regulatory T cells during chronic virus infection enhances the suppression of CD8+ T cell immune response via the interaction with PD-L1 expressed on CD8+ T cells. J Immunol. 2015, 194(12): p. 5801-11. [CrossRef]
- Gambichler, T.; Schröter, U.; Höxtermann, S.; Susok, L.; Stockfleth, E.; Becker, J.C. Decline of programmed death-1-positive circulating T regulatory cells predicts more favorable clinical outcome of patients with melanoma under immune checkpoint blockade. Br J Dermatol. 2020. May,182(5):1214-1220. [CrossRef]
- Fairfax, B.P.; Taylor, C.A.; Watson, R.A.; Nassiri, I.; Danielli, S.; Fang, H.; Mahé, E.A.; Cooper, R.; Woodcock, V.; Traill, Z.; Al-Mossawi, M.H.; Knight, J.C.; Klenerman, P.; Payne, M.; Middleton, M.R. Peripheral CD8+ T cell characteristics associated with durable responses to immune checkpoint blockade in patients with metastatic melanoma. Nat Med. 2020, Feb;26(2):193-199. [CrossRef]
- Tumeh, P.C.; Harview, C.L.; Yearley, J.H.; Shintaku, I.P.; Taylor, E.J.; Robert, L.; Chmielowski, B.; Spasic, M.; Henry, G.; Ciobanu, V.; West, A.N.; Carmona, M.; Kivork, C.; Seja, E.; Cherry, G.; Gutierrez, A.J.; Grogan, T.R.; Mateus, C.; Tomasic, G.; Glaspy, J.A.; Emerson, R.O.; Robins, H.; Pierce, R.H.; Elashoff, D.A.; Robert, C.; Ribas, A. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature. 2014 Nov 27,515(7528):568-71. [CrossRef]
- Watson, R.A.; Tong, O.; Cooper, R.; Taylor, C.A.; Sharma, P.K.; de Los Aires, A.V.; Mahé, E.A.; Ruffieux, H.; Nassiri, I.; Middleton, M.R.; Fairfax, B.P. Immune checkpoint blockade sensitivity and progression-free survival associates with baseline CD8+ T cell clone size and cytotoxicity. Sci Immunol. 2021 Oct;6(64):eabj8825. [CrossRef]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Albright, A.; Cheng, J.D.; Kang, S.P.; Shankaran, V.; Piha-Paul, S.A., et al. IFN-γ-related mRNA profile predicts clinical response to PD-1 blockade. J Clin Investig. 2017, 127: 2930–2940. [CrossRef]
- Tang, C.; Welsh, J.W.; de Groot, P.; Massarelli, E.; Chang, J.Y.; Hess, K.R.; Basu, S.; Curran, M.A.; Cabanillas, M.E.; Subbiah, V.; Fu, S.; Tsimberidou, A.M.; Karp, D.; Gomez, D.R.; Diab, A.; Komaki, R.; Heymach, J.V.; Sharma, P.; Naing, A.; Hong, D.S. Ipilimumab with Stereotactic Ablative Radiation Therapy: Phase I Results and Immunologic Correlates from Peripheral T Cells. Clin Cancer Res. 2017 Mar 15;23(6):1388-1396. [CrossRef]
- Balatoni, T.; Mohos, A.; Papp, E.; Sebestyén, T.; Liszkay, G.; Oláh, J.; Varga, A.; Lengyel, Z.; Emri, G.; Gaudi, I.; Ladányi, A. Tumor-infiltrating immune cells as potential biomarkers predicting response to treatment and survival in patients with metastatic melanoma receiving ipilimumab therapy. Cancer Immunol Immunother. 2018, Jan,67(1):141-151. [CrossRef]
- Fourcade, J.; Sun, Z.; Benallaoua, M.; Guillaume, P.; Luescher, I.F.; Sander, C.; Kirkwood, J.M.; Kuchroo, V.; Zarour, H.M. Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patients. J. Exp. Med. 2010, 207, 2175–2186. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Wang, C.; Cong, L.; Marjanovic, N.D.; Kowalczyk, M.S.; Zhang, H.; Nyman, J.; Sakuishi, K.; Kurtulus, S.; Gennert, D. , et al. A distinct gene module for dysfunction uncoupled from activation in tumor-infiltrating T cells. Cell. 2016, 166, 1500–1511e9. [Google Scholar] [CrossRef] [PubMed]
- Paley, M.A.; Kroy, D.C.; Odorizzi, P.M.; Johnnidis, J.B.; Dolfi, D.V.; Barnett, B.E.; Bikoff, E.K.; Robertson, E.J.; Lauer, G.M.; Reiner, S.L.; Wherry, E.J. Progenitor and terminal subsets of CD8+ T cells cooperate to contain chronic viral infection. Science. 2012 Nov 30;338(6111):1220-5. [CrossRef]
- Im, S.J.; et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature. 2016. 537, 417–421. [CrossRef]
- Miller, B.C.; Sen, D.R.; Al Abosy, R.; Bi, K.; Virkud, Y.V.; LaFleur, M.W.; Yates, K.B.; Lako, A.; Felt, K.; Naik, G.S.; Manos, M.; Gjini, E.; Kuchroo, J.R.; Ishizuka, J.J.; Collier, J.L.; Griffin, G.K.; Maleri, S.; Comstock, D.E.; Weiss, S.A.; Brown, F.D.; Panda, A.; Zimmer, M.D.; Manguso, R.T.; Hodi, F.S.; Rodig, S.J.; Sharpe, A.H.; Haining, W.N. Subsets of exhausted CD8+ T cells differentially mediate tumor control and respond to checkpoint blockade. Nat Immunol. 2019, Mar,20(3):326-336. [CrossRef]
- Huang, A.C.; Orlowski, R.J.; Xu, X.; Mick, R.; George, S.M.; Yan, P.K.; Manne, S.; Kraya, A.A.; Wubbenhorst, B.; Dorfman, L.; D'Andrea, K.; Wenz, B.M.; Liu, S.; Chilukuri, L.; Kozlov, A.; Carberry, M.; Giles, L.; Kier, M.W.; Quagliarello, F.; McGettigan, S.; Kreider, K.; Annamalai, L.; Zhao, Q.; Mogg, R.; Xu, W.; Blumenschein, W.M.; Yearley, J.H.; Linette, G.P.; Amaravadi, R.K.; Schuchter, L.M.; Herati, R.S.; Bengsch, B.; Nathanson, K.L.; Farwell, M.D.; Karakousis, G.C.; Wherry, E.J.; Mitchell, T.C. A single dose of neoadjuvant PD-1 blockade predicts clinical outcomes in resectable melanoma. Nat Med. 2019 Mar;25(3):454-461. [CrossRef]
- Maibach, F.; Sadozai, H.; Seyed Jafari, S.M.; Hunger, R.E.; Schenk, M. Tumor-Infiltrating Lymphocytes and Their Prognostic Value in Cutaneous Melanoma. Front Immunol. 2020. Sep 10,11:2105. [CrossRef]
- Daud, A.I.; Loo, K.; Pauli, M.L.; Sanchez-Rodriguez, R.; Sandoval, P.M.; Taravati, K., et al. Tumor immune profiling predicts response to anti-PD-1 therapy in human melanoma. J Clin Invest. 2016, 126:3447–52. [CrossRef]
- Huang, A.C.; Postow, M.A.; Orlowski, R.J.; Mick, R.; Bengsch, B.; Manne, S.; Xu, W.; Harmon, S.; Giles, J.R.; Wenz, B.; Adamow, M.; Kuk, D.; Panageas, K.S.; Carrera, C.; Wong, P.; Quagliarello, F.; Wubbenhorst, B.; D'Andrea, K.; Pauken, K.E.; Herati, R.S.; Staupe, R.P.; Schenkel, J.M.; McGettigan, S.; Kothari, S.; George, S.M.; Vonderheide, R.H.; Amaravadi, R.K.; Karakousis, G.C.; Schuchter, L.M.; Xu, X.; Nathanson, K.L.; Wolchok, J.D.; Gangadhar, T.C.; Wherry, E.J. T cell invigoration to tumor burden ratio associated with anti-PD-1 response. Nature. 2017, May 4,545(7652):60-65. [CrossRef]
- Kuehm, L.M.; Wolf, K.; Zahour, J.; DiPaolo, R.J.; Teague, R.M. Checkpoint blockade immunotherapy enhances the frequency and effector function of murine tumor-infiltrating T cells but does not alter TCRβ diversity. Cancer Immunol Immunother. 2019, Jul,68(7):1095-1106. [CrossRef]
- Kurtulus, S.; Madi, A.; Escobar, G.; Klapholz, M.; Nyman, J.; Christian, E.; Pawlak, M.; Dionne, D.; Xia, J.; Rozenblatt-Rosen, O.; Kuchroo, V.K.; Regev, A.; Anderson, A.C. Checkpoint Blockade Immunotherapy Induces Dynamic Changes in PD-1-CD8+ Tumor-Infiltrating T Cells. Immunity. 2019, Jan 15,50(1):181-194.e6. [CrossRef]
- Marshall, H.T.; Djamgoz, M.B.A. Immuno-oncology: emerging targets and combination therapies. Front Oncol. 2018, 8:315. [CrossRef]
- Zhao, X.; Shan, Q.; Xue, H.H. TCF1 in T cell immunity: a broadened frontier. Nat Rev Immunol 2022, 22, 147–157. [Google Scholar] [CrossRef]
- Sade-Feldman, M.; Yizhak, K.; Bjorgaard, S.L.; Ray, J.P.; de Boer, C.G.; Jenkins, R.W.; Lieb, D.J.; Chen, J.H.; Frederick, D.T.; Barzily-Rokni, M.; Freeman, S.S.; Reuben, A.; Hoover, P.J.; Villani, A.C.; Ivanova, E.; Portell, A.; Lizotte, P.H.; Aref, A.R.; Eliane, J.P.; Hammond, M.R.; Vitzthum, H.; Blackmon, S.M.; Li, B.; Gopalakrishnan, V.; Reddy, S.M.; Cooper, Z.A.; Paweletz, C.P.; Barbie, D.A.; Stemmer-Rachamimov, A.; Flaherty, K.T.; Wargo, J.A.; Boland, G.M.; Sullivan, R.J.; Getz, G.; Hacohen, N. Defining T Cell States Associated with Response to Checkpoint Immunotherapy in Melanoma. Cell. 2018 Nov 1;175(4):998-1013.e20. [CrossRef]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J. , et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef]
- Siddiqui, I.; Schaeuble, K.; Chennupati, V.; Fuertes Marraco, S.A.; Calderon-Copete, S.; Pais Ferreira, D.; Carmona, S.J.; Scarpellino, L.; Gfeller, D.; Pradervand, S.; Luther, S.A.; Speiser, D.E.; Held, W. Intratumoral Tcf1+PD-1+CD8+ T Cells with Stem-like Properties Promote Tumor Control in Response to Vaccination and Checkpoint Blockade Immunotherapy. Immunity. 2019, Jan 15,50(1):195-211.e10. [CrossRef]
- Jeannet, G.; Boudousquie, C.; Gardiol, N.; Kang, J.; Huelsken, J.; Held, W. Essential role of the Wnt pathway effector Tcf-1 for the establishment of functional CD8 T cell memory. Proc. Natl. Acad. Sci. 2010, USA 107, 9777–9782. [Google Scholar] [CrossRef]
- Kong, Y.C.; Wei, W.Z.; Tomer, Y. Opportunistic autoimmune disorders: from immunotherapy to immune dysregulation. Annals of the New York Academy of Sciences. 2010, 1183:222-36. [CrossRef]
- McLane, L.M.; Ngiow, S.F.; Chen, Z.; Attanasio, J.; Manne, S.; Ruthel, G.; Wu, J.E.; Staupe, R.P.; Xu, W.; Amaravadi, R.K.; Xu, X.; Karakousis, G.C.; Mitchell, T.C.; Schuchter, L.M.; Huang, A.C.; Freedman, B.D.; Betts, M.R.; Wherry, E.J. Role of nuclear localization in the regulation and function of T-bet and Eomes in exhausted CD8 T cells. Cell Rep. 2021 May 11;35(6):109120. [CrossRef]
- Gide, T.N.; Quek, C.; Menzies, A.M.; Tasker, A.T.; Shang, P.; Holst, J.; Madore, J.; Lim, S.Y.; Velickovic, R.; Wongchenko, M.; Yan, Y.; Lo, S.; Carlino, M.S.; Guminski, A.; Saw, R.P.M.; Pang, A.; McGuire, H.M.; Palendira, U.; Thompson, J.F.; Rizos, H.; Silva, I.P.D.; Batten, M.; Scolyer, R.A.; Long, G.V.; Wilmott, J.S. Distinct Immune Cell Populations Define Response to Anti-PD-1 Monotherapy and Anti-PD-1/Anti-CTLA-4 Combined Therapy. Cancer Cell. 2019, Feb 11,35(2):238-255.e6. [CrossRef]
- Strippoli, S.; Fanizzi, A.; Negri, A.; Quaresmini, D.; Nardone, A.; Armenio, A.; Sciacovelli, A.M.; Massafra, R.; De Risi, I.; De Tullio, G.; Albano, A.; Guida, M. Examining the Relationship between Circulating CD4- CD8- Double-Negative T Cells and Outcomes of Immuno-Checkpoint Inhibitor Therapy-Looking for Biomarkers and Therapeutic Targets in Metastatic Melanoma. Cells. 2021, Feb 16,10(2):406. [CrossRef]
- Zhang, Z.X.; Young, K. ; Zhang LCD3+CD4-CD8- alphabeta-TCR+ T cell as immune regulatory cell, J. Mol. Med. Berl. 2001, 79, 419–427. [Google Scholar] [CrossRef] [PubMed]
- D’Acquisto, F.; Crompton, T. CD3+CD4-CD8- (double negative) T cells: Saviours or villains of the immune response? Biochem. Pharmacol. 2011, 82, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Delyon, J.; Mateus, C.; Lefeuvre, D.; Lanoy, E.; Zitvogel, L.; Chaput, N., et al. Experience in daily practice with ipilimumab for the treatment of patients with metastatic melanoma: an early increase in lymphocyte and eosinophil counts is associated with improved survival. Annals of oncology : official journal of the European Society for Medical Oncology / ESMO 2013, 24:1697- 703. [CrossRef]
- Han, X.; Wang, Y.; Sun, J.; Tan, T.; Cai, X.; Lin, P.; Tan, Y.; Zheng, B.; Wang, B.; Wang, J.; Xu, L.; Yu, Z.; Xu, Q.; Wu, X.; Gu, Y. Role of CXCR3 signaling in response to anti-PD-1 therapy. EBioMedicine. 2019, Oct;48:169-177. [CrossRef]
- Mullins, I.M.; Slingluff, C.L.; Lee, J.K.; Garbee, C.F.; Shu, J.; Anderson, S.G.; Mayer, M.E.; Knaus, W.A.; Mullins, D.W. CXC chemokine receptor 3 expression by activated CD8+ T cells is associated with survival in melanoma patients with stage III disease. Cancer Res. 2004 Nov 1;64(21):7697-701. [CrossRef]
- Chow, M.T.; Ozga, A.J.; Servis, R.L.; Frederick, D.T.; Lo, J.A.; Fisher, D.E.; Freeman, G.J.; Boland, G.M.; Luster, A.D. Intratumoral Activity of the CXCR3 Chemokine System Is Required for the Efficacy of Anti-PD-1 Therapy. Immunity. 2019 Jun 18;50(6):1498-1512.e5. [CrossRef]
- House, I.G.; Savas, P.; Lai, J.; Chen, A.X.Y.; Oliver, A.J.; Teo, Z.L.; Todd, K.L.; Henderson, M.A.; Giuffrida, L.; Petley, E.V.; Sek, K.; Mardiana, S.; Gide, T.N.; Quek, C.; Scolyer, R.A.; Long, G.V.; Wilmott, J.S.; Loi, S.; Darcy, P.K.; Beavis, P.A. Macrophage-Derived CXCL9 and CXCL10 Are Required for Antitumor Immune Responses Following Immune Checkpoint Blockade. Clin Cancer Res. 2020 Jan 15;26(2):487-504. [CrossRef]
- XYang, Y., Chu, Y. Wang, R. Zhang, S. Xiong, Targeted in vivo expression of IFN-gamma-inducible protein 10 induces specific antitumor activity. J Leuk Biol 80, 2006, 1434–1444. [CrossRef]
- Bagheri, H.; Pourhanifeh, M.H.; Derakhshan, M.; Mahjoubin-Tehran, M.; Ghasemi, F.; Mousavi, S.; Rafiei, R.; Abbaszadeh-Goudarzi, K.; Mirzaei, H.R.; Mirzaei, H. CXCL-10: a new candidate for melanoma therapy? Cell Oncol (Dordr). 2020 Jun;43(3):353-365. [CrossRef]
- Kawada, K.; Sonoshita, M.; Sakashita, H.; Takabayashi, A.; Yamaoka, Y.; Manabe, T.; Inaba, K.; Minato, N.; Oshima, M.; Taketo, M.M. Pivotal role of CXCR3 in melanoma cell metastasis to lymph nodes. Cancer Res. 2004 Jun 1;64(11):4010-7. [CrossRef]
- Bakouny, Z.; Choueiri, T.K. IL-8 and cancer prognosis on immunotherapy. Nat Med. 2020, May;26(5):650-651. [CrossRef]
- Alfaro, C.; Sanmamed, M.F.; Rodríguez-Ruiz, M.E.; Teijeira, Á.; Oñate, C.; González, Á.; Ponz, M.; Schalper, K.A.; Pérez-Gracia, J.L.; Melero, I., et al. Interleukin-8 in cancer pathogenesis, treatment and follow-up. Cancer Treat Rev. 2017, 60:24–31. [CrossRef]
- Gonzalez-Aparicio, M.; Alfaro, C. Influence of Interleukin-8 and Neutrophil Extracellular Trap (NET) formation in the tumor microenvironment: is there a pathogenic role? J Immunol Res. 2019, 2019:7. [CrossRef]
- Salerno, E.P.; Olson, W.C.; McSkimming, C.; Shea, S.; Slingluff, C.L. T cells in the human metastatic melanoma microenvironment express site-specific homing receptors and retention integrins. Int J Cancer. 2014 Feb 1;134(3):563-74. [CrossRef]
- Schalper, K.A.; Carleton, M.; Zhou, M.; Chen, T.; Feng, Y.; Huang, S.P.; Walsh, A.M.; Baxi, V.; Pandya, D.; Baradet, T.; Locke, D.; Wu, Q.; Reilly, T.P.; Phillips, P.; Nagineni, V.; Gianino, N.; Gu, J.; Zhao, H.; Perez-Gracia, J.L.; Sanmamed, M.F.; Melero, I. Elevated serum interleukin-8 is associated with enhanced intratumor neutrophils and reduced clinical benefit of immune-checkpoint inhibitors. Nat Med. 2020, May;26(5):688-692. [CrossRef]
- Sanmamed, M.F.; Perez-Gracia, J.L.; Schalper, K.A.; Fusco, J.P.; Gonzalez, A.; Rodriguez-Ruiz, M.E.; Oñate, C.; Perez, G.; Alfaro, C.; Martín-Algarra, S.; Andueza, M.P.; Gurpide, A.; Morgado, M.; Wang, J.; Bacchiocchi, A.; Halaban, R.; Kluger, H.; Chen, L.; Sznol, M.; Melero, I. Changes in serum interleukin-8 (IL-8) levels reflect and predict response to anti-PD-1 treatment in melanoma and non-small-cell lung cancer patients. Ann Oncol. 2017 Aug 1;28(8):1988-1995. [CrossRef]
- Sanmamed, M.F.; Carranza-Rua, O.; Alfaro, C.; Oñate, C.; Martín-Algarra, S.; Perez, G.; Landazuri, S.F.; Gonzalez, A.; Gross, S.; Rodriguez, I.; Muñoz-Calleja, C.; Rodríguez-Ruiz, M.; Sangro, B.; López-Picazo, J.M.; Rizzo, M.; Mazzolini, G.; Pascual, J.I.; Andueza, M.P.; Perez-Gracia, J.L.; Melero, I. Serum interleukin-8 reflects tumor burden and treatment response across malignancies of multiple tissue origins. Clin Cancer Res. 2014, Nov 15,20(22):5697-707. [CrossRef]
- Wei, S.C.; Anang, N.A.S.; Sharma, R.; Andrews, M.C.; Reuben, A.; Levine, J.H.; Cogdill, A.P.; Mancuso, J.J.; Wargo, J.A.; Pe'er, D.; Allison, J.P. Combination anti-CTLA-4 plus anti-PD-1 checkpoint blockade utilizes cellular mechanisms partially distinct from monotherapies. Proc Natl Acad Sci U S A. 2019 Nov 5;116(45):22699-22709. [CrossRef]
Figure 1.
Lymphoid lineage cells. Hematopoietic stem cells (HSC) give rise to different types of blood cells, in lines called myeloid and lymphoid. Lymphoid cells include T cells, B cells, natural killer cells. The common lymphoid progenitor (CLP) is the precursor of the 3 lymphoid lineages and gives rise to pro-B cells, pro-T cells and pro-NK cells. T cells mature in the thymus and B cells in the bone marrow. On the other hand, pro-NK cells become NK cells on the periphery when a threat occurs.
Figure 1.
Lymphoid lineage cells. Hematopoietic stem cells (HSC) give rise to different types of blood cells, in lines called myeloid and lymphoid. Lymphoid cells include T cells, B cells, natural killer cells. The common lymphoid progenitor (CLP) is the precursor of the 3 lymphoid lineages and gives rise to pro-B cells, pro-T cells and pro-NK cells. T cells mature in the thymus and B cells in the bone marrow. On the other hand, pro-NK cells become NK cells on the periphery when a threat occurs.
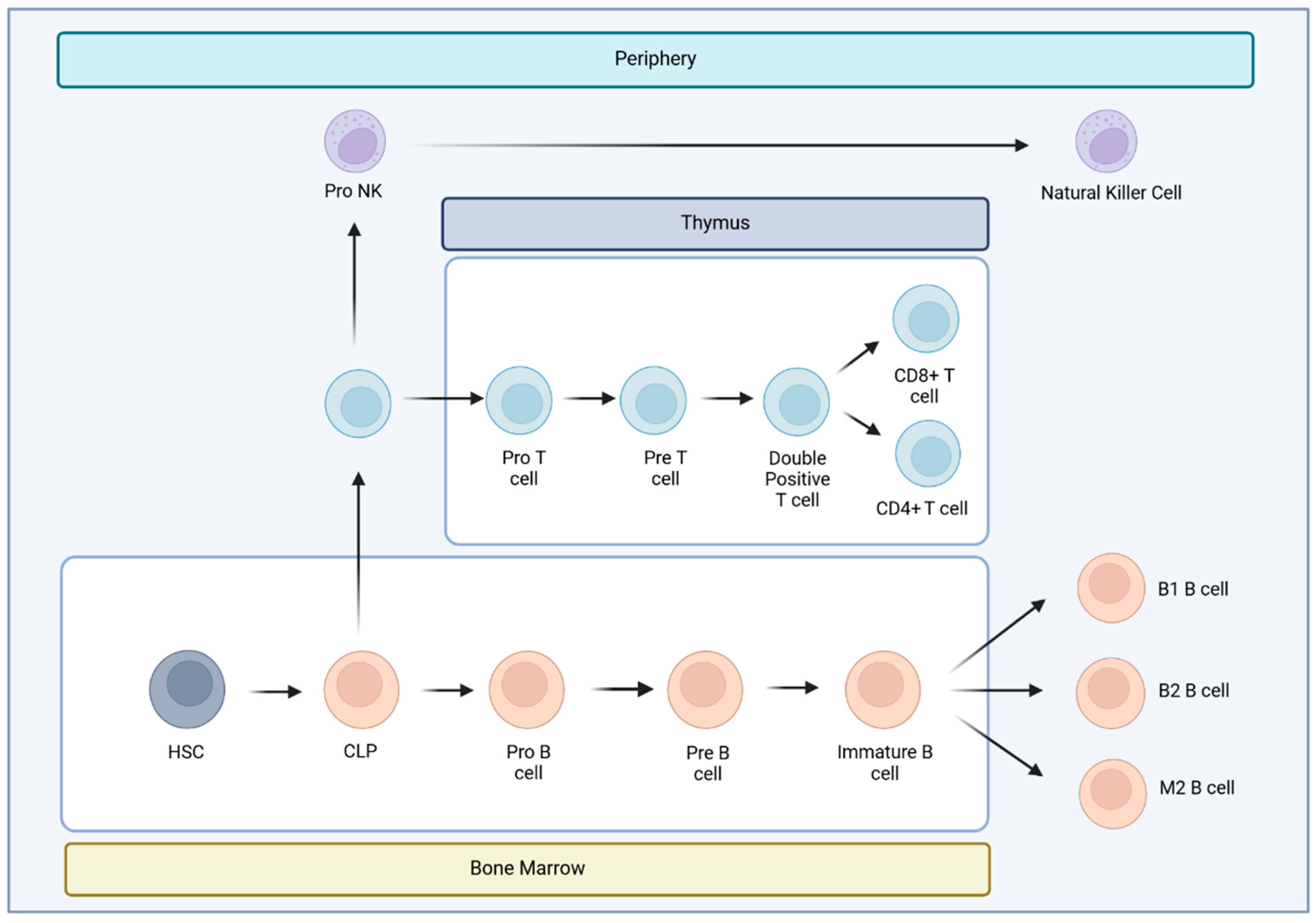
Figure 2.
Memory compartment cells activation against melanoma tumor cells threat. Once antigen-presenting cells, such Langherhans cell, detects a threat of melanoma tumor cells, they travel to lymph node to find Tn cells. Tn cells bind to APC cells it via the TCR, accompanied by a costimulatory signal. Through this process, they become activated and differentiate into effector cells (Teff) that migrate to inflamed sites and attack infected. After pathogen clearance, effector cells undergo apoptosis; the remaining T cells give rise to memory cells that have lost CD45RA expression and instead express CD45RO. Some Tn when they become activated, they differentiate into memory T cells with stem cell-like with an enhanced self-renewal capacity and multipotency to differentiate into all memory T cells. Also, a progressive maturation of T cells from Tn cells via Tcm and Tem to Teff cells that is associated with chronic activation and show features of dysfunction such as impaired cytokine secretion and the expression of markers of exhaustion, such as PD-1 (T exhausted CD45RO+CCR7-PD-1+). Some Tem differentiate into a population of memory CD8+ T cells, resident memory T cells (Trm), characterized for their persistence in tissue, non-recirculating in the blood.
Figure 2.
Memory compartment cells activation against melanoma tumor cells threat. Once antigen-presenting cells, such Langherhans cell, detects a threat of melanoma tumor cells, they travel to lymph node to find Tn cells. Tn cells bind to APC cells it via the TCR, accompanied by a costimulatory signal. Through this process, they become activated and differentiate into effector cells (Teff) that migrate to inflamed sites and attack infected. After pathogen clearance, effector cells undergo apoptosis; the remaining T cells give rise to memory cells that have lost CD45RA expression and instead express CD45RO. Some Tn when they become activated, they differentiate into memory T cells with stem cell-like with an enhanced self-renewal capacity and multipotency to differentiate into all memory T cells. Also, a progressive maturation of T cells from Tn cells via Tcm and Tem to Teff cells that is associated with chronic activation and show features of dysfunction such as impaired cytokine secretion and the expression of markers of exhaustion, such as PD-1 (T exhausted CD45RO+CCR7-PD-1+). Some Tem differentiate into a population of memory CD8+ T cells, resident memory T cells (Trm), characterized for their persistence in tissue, non-recirculating in the blood.
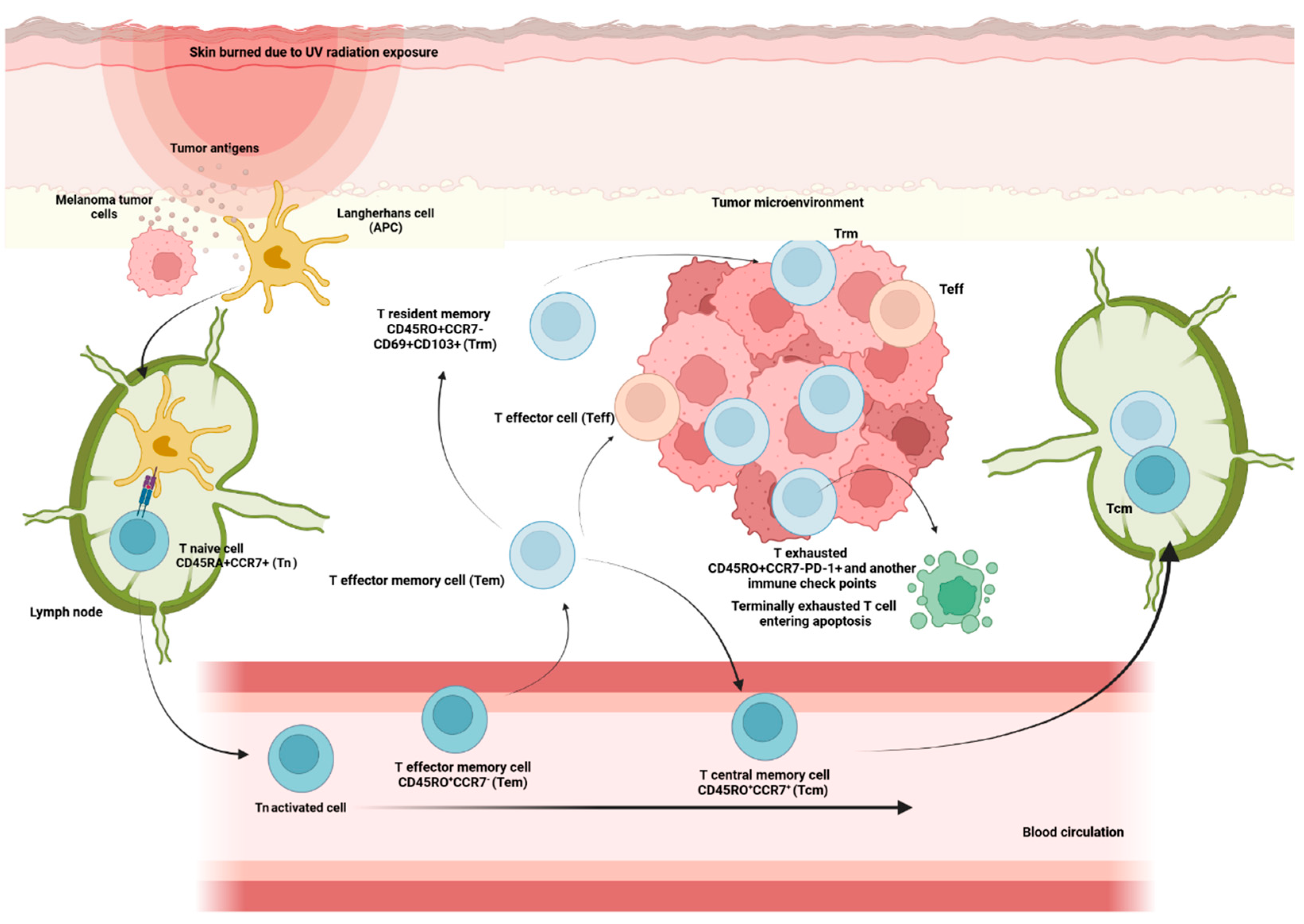
Figure 3.
Distinct process of immune system activation against melanoma tumor cells threat. Once antigen-presenting cells (APC), such Langherhans cell, detects a threat of melanoma tumor cells they travel to lymph node to find Tn cells. Tn cells bind to APC cells it via the TCR, accompanied by a costimulatory signal. Through this process, they become activated and differentiate into effector cells (Teff) that migrate to inflamed sites and attack infected. Tn CD4+ cells differentiate into Treg or Th1 depending on the transcription factor present on the original cell (FoxP3 or T-bet). Tn CD8+ cells differentiate into Tc CD8+ cells. CXCR3+ cells, can be Th1 cells and effector CD8+ T cells, migrate into inflamed tissues and tumors and producte IFN-γ. IFN-γ stimulates CXCL9 and CXCL10+ cells such as NK cells, Tc CD8+ and M1 macrophage recruitment. IL-8 secretion, which is increased in MM patients, causes neutrophil recruitment and stimulation of angiogenesis. At the tumor site, tumor cells produce negative immune check point such as PD-L1 causing Tc CD8+ inactivation.
Figure 3.
Distinct process of immune system activation against melanoma tumor cells threat. Once antigen-presenting cells (APC), such Langherhans cell, detects a threat of melanoma tumor cells they travel to lymph node to find Tn cells. Tn cells bind to APC cells it via the TCR, accompanied by a costimulatory signal. Through this process, they become activated and differentiate into effector cells (Teff) that migrate to inflamed sites and attack infected. Tn CD4+ cells differentiate into Treg or Th1 depending on the transcription factor present on the original cell (FoxP3 or T-bet). Tn CD8+ cells differentiate into Tc CD8+ cells. CXCR3+ cells, can be Th1 cells and effector CD8+ T cells, migrate into inflamed tissues and tumors and producte IFN-γ. IFN-γ stimulates CXCL9 and CXCL10+ cells such as NK cells, Tc CD8+ and M1 macrophage recruitment. IL-8 secretion, which is increased in MM patients, causes neutrophil recruitment and stimulation of angiogenesis. At the tumor site, tumor cells produce negative immune check point such as PD-L1 causing Tc CD8+ inactivation.
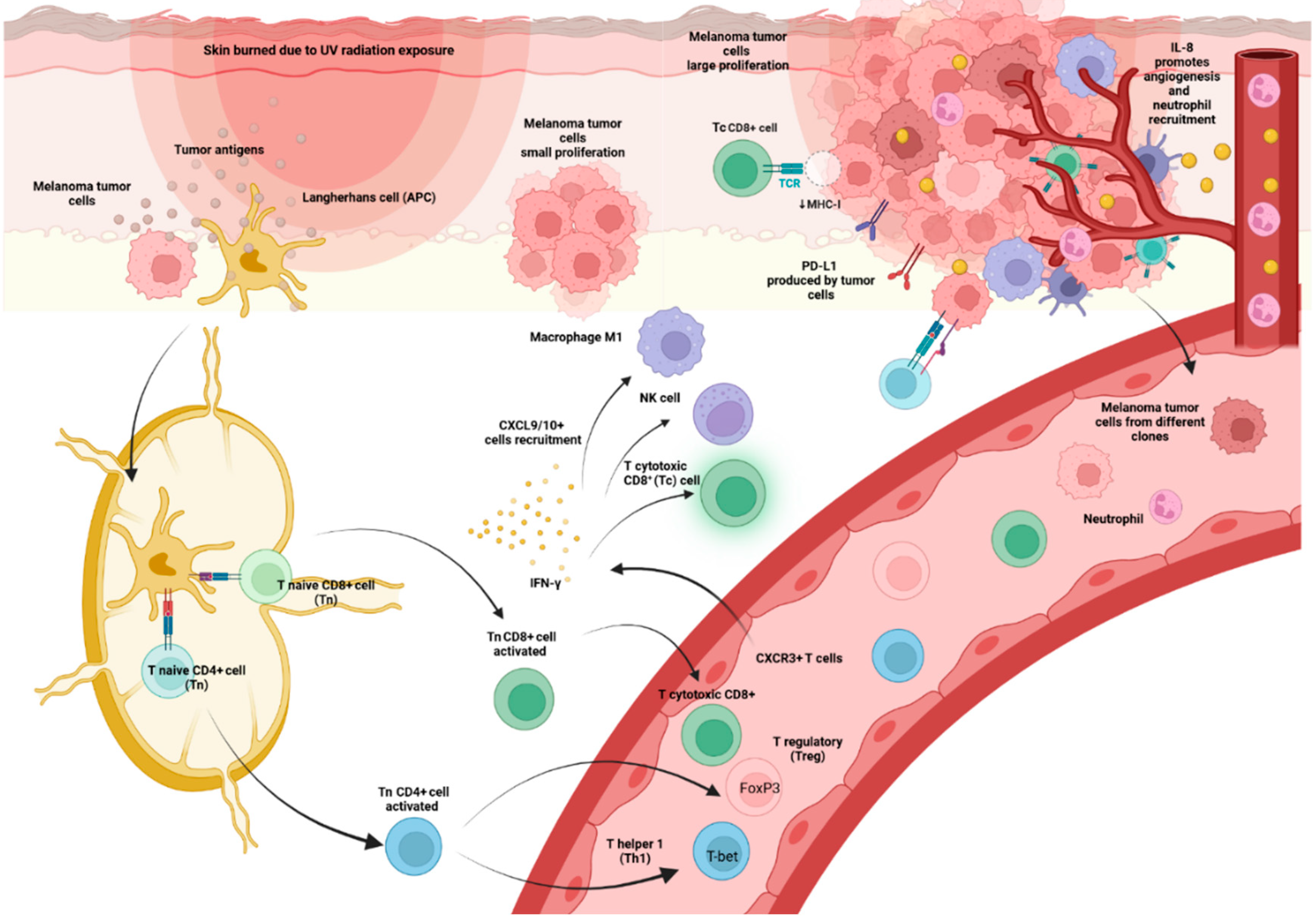
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



